
Suckling behaviour of Nelore, Gir and Caracu calves and their crosses
12
Suckling behaviour of Nelore, Gir and Caracu calves and their crosses Mateus Jose ´ Rodrigues Paranhos da Costa a,b,c, * , Lu ´cia Galva ˜o Albuquerque b,c , Joanir Pereira Eler c,d , Josineudson Augusto II de Vasconcelos Silva e a ETCO-Grupo de Estudos e Pesquisas em Etologia e Ecologia Animal, Brazil b Departamento de Zootecnia, Faculdade de Cie ˆncias Agra ´rias e Veterina ´rias, UNESP, 14884-900 Jaboticabal, SP, Brazil c Pesquisador CNPq, Brazil d Departamento de Cie ˆncias Ba ´sicas, Faculdade de Zootecnia e Engenharia de Alimentos, USP, Pirassununga, SP, Brazil e Estac ¸a ˜o Experimental de Zootecnia de Serta ˜ozinho, IZ-SAA-SP, Serta ˜ozinho, SP, Brazil Accepted 20 February 2006 Available online 18 April 2006 Abstract It is reasonable to assume that the knowledge of suckling behaviour contributes to optimal management and selection of beef cattle. However, there is little information about suckling behaviour of some beef cattle breeds. The aim of this study was to describe the suckling behaviour of two zebu (Bos indicus) and one criollo (Bos taurus) breeds, analysing the potential effects of breed and some environmental factors on suckling frequency and duration. Forty cows, 17 Nelore, 14 Gir (both zebu) and 9 Caracu (criollo) were bred in a diallelic crossing design. The cows and resulting calves were kept on pasture from birth to weaning. Their behaviour was recorded weekly during daylight. Three behavioural traits were considered: number of suckling meals (NSM), duration of each suckling meal (DSM) and total suckling duration (TSD). Allosuckling was not observed. The calves suckled at any time during the daylight and the overall means were: NSM = 2.57 0.05 meals/12 h (from back transformed data), DSM = 9.25 0.11 min/suckling meal and TSD = 23.76 0.47 min/12 h. There was an effect of dam’s breed on NSM and DSM; the calf’s genetic group within breed of cow influenced NSM and TSD when the dams were from www.elsevier.com/locate/applanim Applied Animal Behaviour Science 101 (2006) 276–287 * Corresponding author. Tel.: +55 16 32092678; fax: +55 16 32092684. E-mail address: [email protected] (M.J.R. Paranhos da Costa). 0168-1591/$ – see front matter # 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.applanim.2006.02.006
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Suckling behaviour of Nelore, Gir and Caracu calves and their crosses

Suckling behaviour of Nelore, Gir and
Caracu calves and their crosses
Mateus Jose Rodrigues Paranhos da Costa a,b,c,*,Lucia Galvao Albuquerque b,c, Joanir Pereira Eler c,d,
Josineudson Augusto II de Vasconcelos Silva e
a ETCO-Grupo de Estudos e Pesquisas em Etologia e Ecologia Animal, Brazilb Departamento de Zootecnia, Faculdade de Ciencias Agrarias e Veterinarias,
UNESP, 14884-900 Jaboticabal, SP, Brazilc Pesquisador CNPq, Brazil
d Departamento de Ciencias Basicas, Faculdade de Zootecnia e Engenharia de Alimentos,
USP, Pirassununga, SP, Brazile Estacao Experimental de Zootecnia de Sertaozinho, IZ-SAA-SP, Sertaozinho, SP, Brazil
Accepted 20 February 2006
Available online 18 April 2006
Abstract
It is reasonable to assume that the knowledge of suckling behaviour contributes to optimal
management and selection of beef cattle. However, there is little information about suckling
behaviour of some beef cattle breeds. The aim of this study was to describe the suckling behaviour
of two zebu (Bos indicus) and one criollo (Bos taurus) breeds, analysing the potential effects of
breed and some environmental factors on suckling frequency and duration. Forty cows, 17 Nelore,
14 Gir (both zebu) and 9 Caracu (criollo) were bred in a diallelic crossing design. The cows and
resulting calves were kept on pasture from birth to weaning. Their behaviour was recorded weekly
during daylight. Three behavioural traits were considered: number of suckling meals (NSM),
duration of each suckling meal (DSM) and total suckling duration (TSD). Allosuckling was not
observed. The calves suckled at any time during the daylight and the overall means were:
NSM = 2.57 � 0.05 meals/12 h (from back transformed data), DSM = 9.25 � 0.11 min/suckling
meal and TSD = 23.76 � 0.47 min/12 h. There was an effect of dam’s breed on NSM and DSM;
the calf’s genetic group within breed of cow influenced NSM and TSD when the dams were from
www.elsevier.com/locate/applanim
Applied Animal Behaviour Science 101 (2006) 276–287
* Corresponding author. Tel.: +55 16 32092678; fax: +55 16 32092684.
E-mail address: [email protected] (M.J.R. Paranhos da Costa).
0168-1591/$ – see front matter # 2006 Elsevier B.V. All rights reserved.
doi:10.1016/j.applanim.2006.02.006

the Nelore breed. The age of calf had significant effects on all traits. Males averaged higher NSM
and TSD (2.60 � 0.03 meals and 25.05 � 1.37 min/12 h, respectively) than females (2.12 � 0.04
meals and 21.51 � 1.55 min/12 h, respectively). The differences in suckling behaviour seem to be
produced by a complex combination of genetic and environmental factors, which result in a
particular behavioural relationship within mother–offspring pairs.
# 2006 Elsevier B.V. All rights reserved.
Keywords: Cattle; Cow–calf relationship; Milk transfer estimation; Breed differences; Suckling
1. Introduction
The importance of maternal influence on offspring’s phenotype has been recognised in
mammals since early animal husbandry. Milk production has been considered the main
cause of maternal effects in cattle (Eler et al., 1989). However, there is a large variation in
the correlation coefficient estimates between milk production of the dam and the
performance of the offspring from birth to weaning, ranging from 0.29 to 0.90 (Neville,
1962; Rutledge et al., 1971; Totusek et al., 1973; Robison et al., 1978; Chenette and Frahm,
1981; Clutter and Nielsen, 1987).
It is also recognised that knowledge of the nursing–suckling relationship has practical
significance, contributing to improve management and selection of cattle (Craig, 1981;
Paranhos da Costa and Cromberg, 1998). Although the obvious function of suckling is to
provide nutrients to the calf, it is also important for providing antibodies and improving
welfare through colostrum and maternal contact, respectively. Therefore, it is expected that
other important factors affect the performance of the calf besides the dam’s milk
production, and that the whole care and protection offered by the dam to offspring during
the suckling period should also be considered part of the maternal influence (Poindron and
Le Neindre, 1979; Toledo et al., 2003).
Important breed differences in suckling behaviour have been reported in several
studies (Nicol and Sharafeldin, 1975; Le Neindre, 1989; Alencar et al., 1995; Das et al.,
2000; Espasandin et al., 2001). Usually, these differences are associated with the
mother’s milk production. However, there is not a consensus on this last statement. The
breed differences in suckling behaviour could also result from non-nutritive suckling (de
Passile and Rushen, 1997), which, as described by Lidfors et al. (1994), represented a
high percentage (between 30.3 and 50.0%) of the total suckling bout duration of beef
cattle.
There are many papers addressing the behaviour of cows and calves from birth
to weaning, but most of them have addressed the issue in just a few breeds of Bos
taurus. The information about suckling behaviour of Bos indicus (zebu) and criollo
breeds is still scarce, especially for those reared in Brazil. The aim of this study was
to describe the suckling behaviour of Nelore, Gir and Caracu cattle and their
crosses, addressing its role in milk transfer estimation and analysing the poten-
tial effects of breed and some environmental factors on suckling frequency and
duration.
M.J.R. Paranhos da Costa et al. / Applied Animal Behaviour Science 101 (2006) 276–287 277

2. Animals and methods
The present study was carried out at the University of Sao Paulo campus of
Pirassununga, State of Sao Paulo, Brazil (218590S 478210W and 560 m of altitude). Forty
cows and their respective calves were observed with a variable frequency (ranging from 11
to 27 days).
The cows were bred in a ‘‘diallel crossing design’’, involving the three breeds mentioned
above. Therefore, all of the cows were pure-breds (being 14 Gir; 9 Caracu and 17 Nelore
breed) and the calves were either, pure (N = 22; being 8 Gir, 5 Caracu and 9 Nelore) or cross-
bred (N = 18; being 2 Gir-Caracu, 4 Gir-Nelore, 3 Caracu-Gir, 4 Caracu-Nelore, 3 Nelore-Gir
and 2 Nelore-Caracu). Nelore and Gir are zebu breeds (Bos indicus) first taken to Brazil in the
19th century and Caracu is a Bos taurus originated in Brazil through crossing of Iberian cattle
(Felius, 1985), which have been introduced into the country since the beginning of the 16th
century. The calving season was from November to March and the observations were carried
out from February to September. Each cow and its calf were identified with the same coloured
rubber collar. All the animals were kept together in pastures and the calves had free access to
suckling during the whole period of observation.
The behavioural data were collected by continuous, direct observations (Martin and
Bateson, 1993). We observed all occurrences of suckling and attempts to suckle, recording
the time of the beginning and ending of each occurrence. An attempt to suckle was
recorded when a calf tried to get a teat into its mouth without success, and suckling when a
calf succeeded in getting a teat into its mouth for any detectable period of time combined
with observable swallowing. The cow-calf pair identification (based on the colour of
collars) and the date of observation were also recorded.
Observations were performed by three people (previously trained) riding a horse with
the aid of binoculars. They were conducted during the daylight period (from 06:00 to
18:00 h) on 28 days. The date of birth of cows and calves, sex and genetic composition of
calves, were obtained from the farm files.
Milk intake was estimated monthly from the 30th to 180th days after calving. The
method of weighing the calf just before and after a suckling meal was used. The method
comprised the following steps: (1) cows and calves were separated at 11:00 h; (2) at
16:00 h the calves were weighed and immediately allowed to suckle their mothers during
approximately 30 min; (3) they were separated again overnight; (4) at 08:00 h the next day
the calves were weighed before suckling; (5) just after weighing they were allowed to
suckle; (6) the calves were reweighed just after suckling. It was assumed that the difference
between the calves’ weight (before and after the second suckling) estimated the normal
milk intake during the 16-h period. The milk production per lactation was estimated by
multiplying the milk intake on sampling days by the number of days between consecutive
milk intake records.
2.1. Data management and statistical analysis
The data file was composed by 774 daily records, characterizing the following variables:
number of suckling meals (NSM), average duration of each suckling meal (DSM) and total
suckling duration (TSD—total time spent suckling in 12 h).
M.J.R. Paranhos da Costa et al. / Applied Animal Behaviour Science 101 (2006) 276–287278

Repeated measures analysis of variance using the MIXED procedure of the SAS
software package (SAS, 2001) was applied and the model considered the fixed effects of
dam’s breed, calf’s genetic group within the dam’s breed; month of birth, sex of calf, age of
cow at calving and age of calf at the day of observation on NSM, DSM and TSD. Linear
effects of calf’s birth weight and dam’s milk production were also considered, as
covariates. The number of suckling meals was transformed into square root of (NSM + 0.5)
to meet the requirements of normal distribution.
3. Results
Two thousand one hundred ninety-one responses of nursing-suckling interactions from 40
cows and their calves were recorded. Ninety-three percent of them (2038 records) were
effective suckling and the remaining seven percent (153 records) referred to attempts of
suckling in which the calves, for various reasons, did not succeed. The lack of success in
suckling, in spite of the attempts, occurred mainly due to rejection by the cows (62.1%), or
when the calves stopped their attempts before suckling (8.5% of the cases) and when some
other animal disturbed those attempts (1.3%). In 28.1% of the cases the reason for terminating
the attempt could not be determined. All nursing–sucking interactions involved mother–
offspring interactions; allosuckling (a calf suckling from an alien cow) was not observed.
The calves suckled at any time during the daylight, but three peaks of suckling meals
were noted: from 6:00 to 7:00, from 10:00 to 12:00 and from 17:00 to 18:00 h. The overall
means were: NSM = 2.57 � 0.05 suckling meals/12 h (from back transformed data),
DSM = 9.25 � 0.11 min/suckling meal and TSD = 23.76 � 0.47 min/12 h.
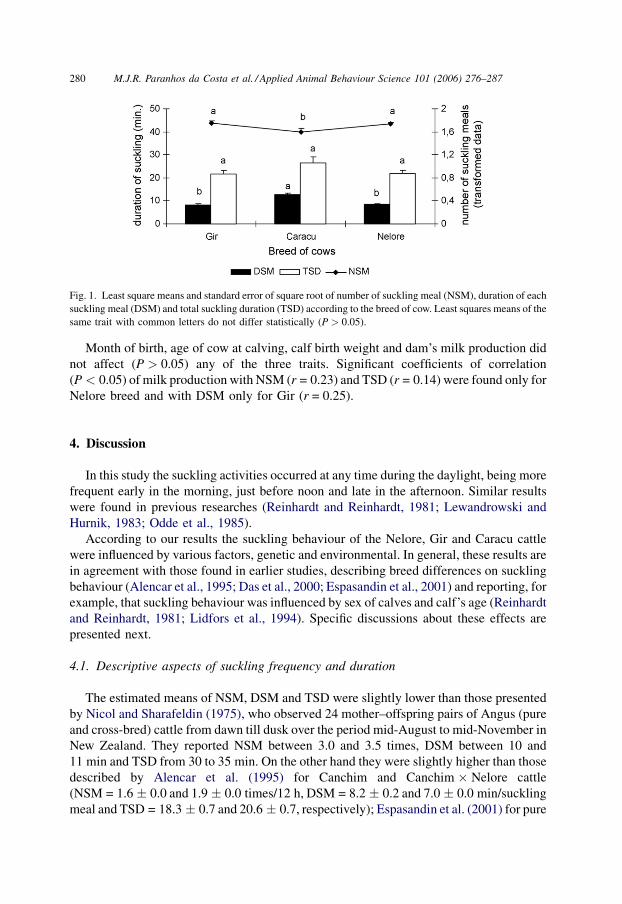
There was an effect of dam’s breed on NSM (P < 0.05) and DSM (P < 0.01), but not
TSD (P > 0.05). The NSM means (back-transformed data) of Gir and Nelore dams
(2.56 � 0.04 and 2.53 � 0.03 suckling meals/12 h, respectively) were higher than the
mean of Caracu dams (2.03 � 0.07 suckling meals/12 h). Regarding mean durations of
suckling meals (DSM) the results were the inverse, with Caracu dams presenting higher
mean than Nelore and Gir dams. The least square means of NSM (transformed data), DSM
and TSD, according to the dams breed, are presented in Fig. 1.
The calf’s genetic group within breed of cow influenced NSM and TSD traits only when
the dams were from Nelore breed. The adjusted means according to the calf’s genetic group
are presented in Fig. 2. The highest NSM was presented by pure-bred Nelore–Nelore
calves, which differed significantly (P < 0.05) from the Gir–Nelore and Caracu–Nelore
means. The same order was found for TSD means, and there were significant differences
between the three means (P < 0.05, Fig. 2C).
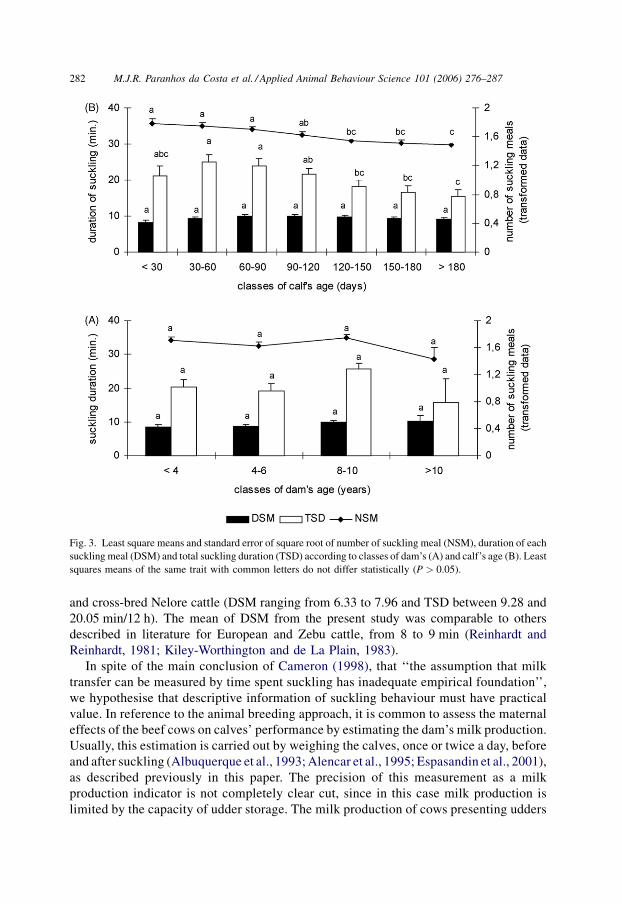
Age of calf, which corresponds to the stage of lactation, had significant effects
(P < 0.01) on NSM and TSD, but not DSM. NSM decreased continuously with age of calf.
On the other hand, means of TSD presented a light increase until the calf reached 60–90
days of age, decreasing continuously after that (Fig. 3B).
The sex of the calf affected NSM (P < 0.01) and TSD (P < 0.05), but not DSM. Males
presented higher NSM (2.60 � 0.03 suckling meals/12 h; back transformed data) and TSD
(25.05 � 1.37 min/12 h) than females (2.12 � 0.04 suckling meals/12 h and 21.51 � 1.55
min/12 h, respectively).
M.J.R. Paranhos da Costa et al. / Applied Animal Behaviour Science 101 (2006) 276–287 279
Month of birth, age of cow at calving, calf birth weight and dam’s milk production did
not affect (P > 0.05) any of the three traits. Significant coefficients of correlation
(P < 0.05) of milk production with NSM (r = 0.23) and TSD (r = 0.14) were found only for
Nelore breed and with DSM only for Gir (r = 0.25).
4. Discussion
In this study the suckling activities occurred at any time during the daylight, being more
frequent early in the morning, just before noon and late in the afternoon. Similar results
were found in previous researches (Reinhardt and Reinhardt, 1981; Lewandrowski and
Hurnik, 1983; Odde et al., 1985).
According to our results the suckling behaviour of the Nelore, Gir and Caracu cattle
were influenced by various factors, genetic and environmental. In general, these results are
in agreement with those found in earlier studies, describing breed differences on suckling
behaviour (Alencar et al., 1995; Das et al., 2000; Espasandin et al., 2001) and reporting, for
example, that suckling behaviour was influenced by sex of calves and calf’s age (Reinhardt
and Reinhardt, 1981; Lidfors et al., 1994). Specific discussions about these effects are
presented next.
4.1. Descriptive aspects of suckling frequency and duration
The estimated means of NSM, DSM and TSD were slightly lower than those presented
by Nicol and Sharafeldin (1975), who observed 24 mother–offspring pairs of Angus (pure
and cross-bred) cattle from dawn till dusk over the period mid-August to mid-November in
New Zealand. They reported NSM between 3.0 and 3.5 times, DSM between 10 and
11 min and TSD from 30 to 35 min. On the other hand they were slightly higher than those
described by Alencar et al. (1995) for Canchim and Canchim � Nelore cattle
(NSM = 1.6 � 0.0 and 1.9 � 0.0 times/12 h, DSM = 8.2 � 0.2 and 7.0 � 0.0 min/suckling
meal and TSD = 18.3 � 0.7 and 20.6 � 0.7, respectively); Espasandin et al. (2001) for pure
M.J.R. Paranhos da Costa et al. / Applied Animal Behaviour Science 101 (2006) 276–287280
Fig. 1. Least square means and standard error of square root of number of suckling meal (NSM), duration of each
suckling meal (DSM) and total suckling duration (TSD) according to the breed of cow. Least squares means of the
same trait with common letters do not differ statistically (P > 0.05).

M.J.R. Paranhos da Costa et al. / Applied Animal Behaviour Science 101 (2006) 276–287 281
Fig. 2. Least square means and standard error of square root of number of suckling meals (NSM), duration of each
suckling meal (DSM) and total suckling duration (TSD) for calves mothered by Gir (A), Caracu (B) and Nelore
(C) cows. Least squares means of the same trait with common letters do not differ statistically (P > 0.05).
and cross-bred Nelore cattle (DSM ranging from 6.33 to 7.96 and TSD between 9.28 and
20.05 min/12 h). The mean of DSM from the present study was comparable to others
described in literature for European and Zebu cattle, from 8 to 9 min (Reinhardt and
Reinhardt, 1981; Kiley-Worthington and de La Plain, 1983).
In spite of the main conclusion of Cameron (1998), that ‘‘the assumption that milk
transfer can be measured by time spent suckling has inadequate empirical foundation’’,
we hypothesise that descriptive information of suckling behaviour must have practical
value. In reference to the animal breeding approach, it is common to assess the maternal
effects of the beef cows on calves’ performance by estimating the dam’s milk production.
Usually, this estimation is carried out by weighing the calves, once or twice a day, before
and after suckling (Albuquerque et al., 1993; Alencar et al., 1995; Espasandin et al., 2001),
as described previously in this paper. The precision of this measurement as a milk
production indicator is not completely clear cut, since in this case milk production is
limited by the capacity of udder storage. The milk production of cows presenting udders
M.J.R. Paranhos da Costa et al. / Applied Animal Behaviour Science 101 (2006) 276–287282
Fig. 3. Least square means and standard error of square root of number of suckling meal (NSM), duration of each
suckling meal (DSM) and total suckling duration (TSD) according to classes of dam’s (A) and calf’s age (B). Least
squares means of the same trait with common letters do not differ statistically (P > 0.05).

with low storage capacity could be underestimated, just by weighing their calves only
once or twice a day.
This statement is based on empirical data. The NSM means from our data, recorded only
during the 12 daylight hours, were always higher than two. Similar results were found by
other authors. Nicol and Sharafeldin (1975), working during the daylight with crossed beef
cattle in New Zealand, reported that the calves presented a suckling frequency ranging
from 3.5 to 5.6 suckling meals/day at 120 and 7 days of age, respectively. Reinhardt and
Reinhardt (1981), studying zebu cattle in Kenya, found 4.8 as the average number of
suckling meals per 24 h; Odde et al. (1985), found an average of 5.0 suckling meals/24 h.
Besides, in our study Nelore and Gir herds presented higher NSM means and lower DSM
means than Caracu. This might explain the breed differences in milk production estimated
by Albuquerque et al. (1993) for Caracu (1077.76 � 134.43 kg), Gir (406.44 � 106.97 kg)
and Nelore breed (631.92 � 90.69 kg), considering 207 days of lactation. Although, some
of the coefficients of correlation of milk production with NSM and DSM were significantly
different from zero, they were always lower than 0.25, characterizing a weak association of
milk production (estimated by the weighing-suckling-weighing method) with suckling
frequency and duration. These results together led us to hypothesise that a calf could get the
same amount of milk from one cow with a low capacity of milk storage, by suckling more
frequently than a calf from a higher udder capacity cow.
We did not observe a single event of allosuckling. Previous studies described the
occurrence of allosuckling in cattle, ranging from 3% (Lewandrowski and Hurnik, 1983
and Das et al., 2000) to 19.02% of the total suckling events (Vıchova and Bartos, 2005).
There are strong evidences that, under restricted milk and space conditions, the probability
of allosuckling increases (Packer et al., 1992; Murphey et al., 1991, 1995). This is, in some
extension, supported by the comparison between the present results and those found by
Lewandrowski and Hurnik (1983), Das et al. (2000) and Vıchova and Bartos (2005). In our
study the animals were kept in free-range conditions, the cows were not milked and the
calves had free access to suckle; in their studies cows and calves had some restriction in
space and milk availability.
4.2. Effect of genetic groups on suckling frequency and duration
In relation to the effects of dams’ breed we have found that the highest NSM mean was
from the breed with the lowest DSM mean, and vice-versa. This could explain the lack of
significance of the dam’s breed effect on TSD, since this variable is a product of NSM and
DSM. This result disagrees with those presented by Das et al. (2000), who found higher
duration and number of suckling meals in zebu than in cross-bred cow–calf pairs, and
significant effects of the breed of cow-calf pairs on the total duration of suckling time.
As pointed out by Nicol and Sharafeldin (1975) breed differences in suckling behaviour
could be attributed to milk production. However, this was not the case in a study with Salers
and Friesian cows–calves pairs carried out by Le Neindre (1989), leading him to conclude
that the total suckling time should result from the need for mother–young physical contact
instead of from the milk production. Both conclusions might be correct, but certainly
incomplete. In our understanding the differences in suckling behaviour among breeds
could result from the combination of similarities and the differences in many traits that are
M.J.R. Paranhos da Costa et al. / Applied Animal Behaviour Science 101 (2006) 276–287 283

involved with physiological and psychological states, and are largely dependent on the
environmental circumstances.
As reported previously, significant differences among the calves’ genetic group were
found depending on the dams’ breed. The calves’ genetic group influenced all variables
when the dams were from Nelore breed. There were no differences among calves mothered
by Gir and Caracu cows. We did not find any logical explanation for such variation among
calves’ genetic group within dams’ breed.
The expectation of a change in suckling behaviour of cross-bred calves to sustain an
increase in nutritional demand promoted by a possible effect of individual heterosis was
not confirmed here, since their suckling frequency and duration were not higher than those
presented by the pure-bred calves. These results disagree with some of those presented
previously. For example, higher DSM means for crossbred than for pure-bred calves were
found by Nicol and Sharafeldin (1975), who proposed that the increase of DSM could
stimulate milk production and consequently proportionate a higher growth rate.
A comparison between pure and crossed Canchim and Canchim–Nelore calves
(Alencar et al., 1995) showed higher NSM for crossed than for pure calves (11.1%), and
lower DSM (14.6%). The difference between TSD means was non significant (18.3 � 0.7
and 20.6 � 0.7 min for pure and crossed calves, respectively). However, Espasandin et al.
(2001) did not find the same tendency when comparing the suckling behaviour of pure and
crossed Nelore calves (Canchim � Nelore, Angus � Nelore and Simental � Nelore) and
their dams’ milk production. The highest mean of DSM was from Simental � Nelore,
which did not differ significantly from the mean of pure Nelore in an intensive system. The
authors did not find significant effects of calves’ genetic group on NSM and TSD.
4.3. Environmental effects on suckling frequency and duration
Several researchers have reported the effects of the age of the calf (or stage of lactation) on
suckling behaviour (Walker, 1962; Nicol and Sharafeldin, 1975; Reinhardt and Reinhardt,
1981; Kiley-Worthington and de La Plain, 1983; Lidfors et al., 1994; Das et al., 2000). In the
present study DSM presented a slight increase between 30 and 60 days of age and remained
relatively constant until 120 days of age, decreasing slightly after this age. The TSD increased
initially, probably due to the increase in DSM, but after 90 days of age it followed the trend
observed for NSM, reducing with the age of calf. Similar results were described by Nicol and
Sharafeldin (1975), reporting an increase in the time spent in each suckling from the 1st to
35th day of life and a decrease in number of sucklings with increasing calf age. However,
other authors found different tendencies in suckling behaviour according to the calf’s age:
Hafez and Lineaver (1968) described an increase in the frequency of suckling from birth to
weaning, Lidfors et al. (1994) observed that the duration of suckling bout decreased with the
calf’s age and the number of suckling bouts increased between 7 and 65 days of age; Das et al.
(2000) reported a continuous decrease in frequency and duration of suckling from birth to 6
months postpartum and Espasandin et al. (2001) did not find any effect of calf’s age on
suckling frequency but the duration of each suckling meal decreased from 60 to 180 days of
age, following the same tendency of milk yield.
The sex of calf significantly affected NSM and TSD, but not DSM. This could be related to
the calves’ weight, since, on average, male calves were heavier than female. These results are
M.J.R. Paranhos da Costa et al. / Applied Animal Behaviour Science 101 (2006) 276–287284

different from those reported by Nicol and Sharafeldin (1975), Reinhardt and Reinhardt
(1981) and Espasandin et al. (2001). These authors did not find significant effects of sex on
suckling behaviour.
Our results lead us to hypothesise that male calves had a higher demand for milk than
the female calves, resulting in a significant increase in NSM, which led to an increase in
TSD and probably an increase in milk ingestion. This last statement is based upon the
well-known effect of increasing the milking frequency on milk production of dairy cows
(Klei et al., 1997). It is also in agreement with the hypothesis proposed by Trivers (1972),
assuming, for polygynous species like cattle, that natural selection favours parents that
invest more in males, since they present greater variance in reproductive success than
females. We did not find any result addressing the milk production of beef cattle which
could sustain this hypothesis. Instead, some studies with beef cattle (Albuquerque et al.,
1993 and Espasandin et al., 2001) did not find any effect of calf’s sex on milk production.
A similar result was found for water buffalo (Bubalus bubalis), with no difference in
milk production of cows mothering male and female calves (Paranhos da Costa et al.,
2000).
On the other hand, there are some papers showing the higher maternal investment in
male than in female offspring. Alley et al. (1995) studying feral goats (Capra hircus),
showed that single males suckled more frequently than single females, and they also
exhibited a higher growth rate. There are other results supporting the idea of differential
maternal investment based on the sex of offspring: bull calves were heavier at birth and
weaning time (Albuquerque et al., 1993), presented a higher weight gain (Paranhos da
Costa et al., 2000) and were naturally weaned later than heifer calves (Reinhardt and
Reinhardt, 1981).
Considering the results presented here and those reported previously we conclude that
the differences in suckling behaviour cannot be explained by a single isolated underlying
factor. It is more likely that differences in suckling behaviour are produced by a complex
combination of genetic and experiential factors, which would result in a particular
behavioural style for a mother–offspring pair. Nevertheless, under practical conditions, it
could be important to identify specific underlying factors to explain major effects of
suckling behaviour on calf survival or development.
Acknowledgements
We are grateful to Alan Bitu Alves, Andrei K.M. de Castro, Anselmo Bodenmuller,
Marcos V.B. da Silva, Ricardo B. Tonetto and Roberto Carlos de Souza for their help
during data collection. Financial supports were given by CNPq (Conselho Nacional de
Desenvolvimento Cientıfico e Tecnologico) and Fapesp (Fundacao de Amparo a Pesquisa
do Estado de Sao Paulo).
References
Albuquerque, L.G., Eler, J.P., Paranhos da Costa, M.J.R., Souza, R.C., 1993. Producao de leite e desempenho do
bezerro na fase de aleitamento em tres racas bovinas de corte. Rev. Soc. Braz. Zootec. 22, 745–754.
M.J.R. Paranhos da Costa et al. / Applied Animal Behaviour Science 101 (2006) 276–287 285

Alencar, M.M., Cruz, G.M., Tullio, R.R., Correa, L.A., 1995. Caracterısticas da amamentacao de bezerros da raca
Canchim e cruzados Canchim � Nelore. Rev. Braz. Zootec. 24, 706–714.
Alley, J.C., Fordham, R.A., Minot, E.O., 1995. Mother–offspring interactions in feral goats—a behavioural
perspective of maternal investment. N.Z. J. Zool. 22, 17–23.
Cameron, E.Z., 1998. Is suckling behaviour a useful predictor of milk intake? A review. Anim. Behav. 56, 521–
532.
Chenette, C.G., Frahm, R.R., 1981. Yield and composition of milk from various two-breed cross cows. J. Anim.
Sci. 52, 483–492.
Clutter, A.C., Nielsen, M.K., 1987. Effect of level of beef cow milk production on pre and post-weaning calf
growth. J. Anim. Sci. 64, 1313–1322.
Craig, J.V., 1981. Domestic Animal Behaviour. Prentice-Hall Inc., Englewood Cliffs, NJ.
Das, S.M., Redbo, I., Wiktorsson, H., 2000. Effect of age of calf on suckling behaviour and other behavioural
activities of Zebu and crossbred calves during restricted suckling periods. Appl. Anim. Behav. Sci. 67, 47–57.
de Passile, A.M., Rushen, J., 1997. Motivational and physiological analysis of the causes and consequences of
non-nutritive sucking by calves. Appl. Anim. Behav. Sci. 53, 15–31.
Eler, J.P., Lobo, R.B., Rosa, A.N., 1989. Influencia de fatores geneticos e de meio em pesos de bovinos da raca
nelore criados no Estado de Sao Paulo. Rev. Soc. Braz. Zootec. 18, 103–111.
Espasandin, A.C., Packer, I.U., Alencar, M.M., 2001. Producao de leite e comportamento de amamentacao em
cinco sistemas de producao de gado de corte. Rev. Braz. Zootec. 30, 702–708.
Felius, M., 1985. Genus Bos: Cattle Breeds of the World. MSD AGVET, Rahway, NJ, p. 234.
Hafez, E.S.E., Lineaver, J.A., 1968. Suckling behaviour in natural and artificially fed neonate calves. Z.
Tierpsychol. 25, 187–198.
Kiley-Worthington, M., de La Plain, S., 1983. The behaviour of beef suckler cattle (Bos taurus). Tierhaltung, vol.
14. Birkhauser, Basel, 195 pp.
Klei, L.R., Lynch, J.M., Barbano, D.M., Oltenaco, P.A., Lednor, A.J., Bandler, D.K., 1997. Influence of milking
three times a day on milk quality. J. Dairy Sci. 80, 427–436.
Le Neindre, P., 1989. Influence of rearing conditions and breed on social relationships of mother and young. Appl.
Anim. Behav. Sci. 23, 129–140.
Lewandrowski, N.M., Hurnik, J.F., 1983. Nursing and cross-nursing behaviour of beef cattle in confinement. Can.
J. Anim. Sci. 63, 849–853.
Lidfors, L.M., Jensen, P., Algers, B., 1994. Suckling in free-ranging beef cattle—temporal patterning of suckling
bouts and effects of age and sex. Ethology 98, 321–332.
Martin, P., Bateson, P., 1993. Measuring Behaviour: An Introductory Guide. Cambridge University Press,
Cambridge, 256 pp.
Murphey, R.M., Paranhos da Costa, M.J.R., Lima, L.O.S., Duarte, F.A.M., 1991. Communal suckling in water
buffalo (Bubalus bubalis). Appl. Anim. Behav. Sci. 28, 341–352.
Murphey, R.M., Paranhos da Costa, M.J.R., Silva, R.G., Souza, R.C., 1995. Allonursing in river buffalo, Bubalus
bubalis: nepotism, incompetence or thievery? Anim. Behav. 49, 1611–1616.
Neville Jr., W.E., 1962. Influence of dam’s milk production and other factors on 120 and 240 day weight in
Hereford calves. J. Anim. Sci. 21, 315–320.
Nicol, A.M., Sharafeldin, M.A., 1975. Observations on the behaviour of single-suckled calves from birth to 120
day. Proc. N.Z. Soc. Anim. Prod. 35, 221–230.
Odde, K.G., Kiracofe, G.H., Schalles, R.R., 1985. Suckling behaviour in range beef calves. J. Anim. Sci. 61, 307–
309.
Packer, C., Lewis, S., Pusey, A., 1992. A comparative analysis of non-offspring nursing. Anim. Behav. 43, 265–281.
Paranhos da Costa, M.J.R., Cromberg, V.U., 1998. Relacoes materno-filiais em bovinos de corte nas primeiras
horas apos o parto. In: Paranhos da Costa, M.J.R., Cromberg, V.U. (Eds.), Comportamento Materno em
Mamıferos. Sociedade Brasileira de Etologia, Sao Paulo, pp. 215–235.
Paranhos da Costa, M.J.R., Andriolo, A., de Oliveira, J.F.S., Schmidek, W.R., 2000. Suckling and allosuckling in
river buffalo calves and its relations with relations with weight gain. Appl. Anim. Behav. Sci. 66, 1–10.
Poindron, P., Le Neindre, P., 1979. Les relations mere-jeunez chez les ruminants domestiques et leurs
consequences en production animale. In: Action conjointe des effects direct et maternels des genes sur
les characteres de, production. Bulletin Technique du Department de Genetic Animale, pp. 29–30.
M.J.R. Paranhos da Costa et al. / Applied Animal Behaviour Science 101 (2006) 276–287286

Reinhardt, V., Reinhardt, A., 1981. Natural suckling performance and age of weaning in zebu cattle (Bos indicus).
J. Agric. Sci. 96, 309–312.
Robison, O.W., Youseff, M.K.M., Dillard, E.U., 1978. Milk production in Hereford cows. I. Means and
correlations. J. Anim. Sci. 47, 131–136.
Rutledge, J.J., Robison, O.W., Ahlschwede, W.T., Legates, J.E., 1971. Milk yield and its influence on 205-day
weight of beef calves. J. Anim. Sci. 33, 563–567.
SAS, 2001. Statistical Analysis System User’s Guide, version 8.2. SAS Institute, Cary, USA.
Toledo, L.M., Paranhos da Costa, M.J.R., Schmidek, A., 2003. The presence of black vultures has negative effects
on the cow-calf behaviour just after birth. Rev. Etol. 5, 214 (Abstract).
Totusek, R., Arnett, D.W., Holland, G.L., Whiteman, J.V., 1973. Relation of estimation method, sampling interval
and milk composition to milk yield of beef cows and calf gain. J. Anim. Sci. 37, 153–158.
Trivers, R.L., 1972. Parental investment and sexual selection. In: Campbell, B. (Ed.), Sexual Selection and the
Descent of Man 1871–1971. Aldine, Chicago, pp. 136–179.
Vıchova, J., Bartos, L., 2005. Allosuckling in cattle: gain or compensation? Appl. Anim. Behav. Sci. 94, 223–235.
Walker, D.E., 1962. Suckling and grazing behaviour of beef heifers and calves. N.Z. J. Agric. Res. 5, 331–338.
M.J.R. Paranhos da Costa et al. / Applied Animal Behaviour Science 101 (2006) 276–287 287



















