
Survival, male morphotypes, female and male proportion, female reproductive status and tag loss in...
12
Survival, male morphotypes, female and male proportion, female reproductive status and tag loss in crosses among three populations of freshwater prawn Macrobrachium rosenbergii (de Man) in India Bindu R Pillai 1 , Kanta D Mahapatra 1 , Raul W Ponzoni 2 , Lopamudra Sahoo 1,† , P L Lalrinsanga 1 , Wagdy Mekkawy 2,3 , Hooi Ling Khaw 2 , Nguyen H Nguyen 2, *, Swagathika Mohanty 1 , Sovan Sahu 1 & Gunamaya Patra 1 1 Central Institute of Freshwater Aquaculture, Bhubaneswar, Odisha, India 2 WorldFish, Bayan Lepas, Penang, Malaysia 3 Animal Production Department, Faculty of Agriculture, Ain Shams University, Cairo, Egypt Correspondence: B R Pillai, Principal Scientist, CIFA, Kausalyaganga, Bhubaneswar-751002, Odisha, India. E-mail: pillaibr@ hotmail.com *Present address: N H Nguyen, School of Science, Education and Engineering, University of the Sunshine Coast, Maroochydore, QLD 4558, Australia † Present address: L Sahoo, ICAR Research Complex for North East Region, Lembucherra, Tripura 799210, India Abstract The present study examined the variation in sur- vival, proportion of male morphotypes, female and male proportion, female reproductive status and tag losses in nine crosses from a complete (3 9 3) dial- lel mating of three populations of Macrobrachium rosenbergii in India. The populations originated from Gujarat (north-west), Kerala (south-west) and Odisha (east), representing different agro-ecological regions in India. Progeny from 60 families (4773 juveniles) were individually tagged and reared for 16–17 weeks in earthen ponds. Binary logistic regression was used to analyse the data. Survival rate was greatest (0.88) for Kerala 9 Odisha (KO; the first letter represents the sire), whereas it was lowest (0.75) for both Kerala 9 Kerala (KK) and Gujarat 9 Kerala (GK). Significantly greater pro- portions of large male morphotypes were observed in KK cross and in combinations involving Kerala population. The highest proportion of immature females, as well as of non-berried females, was observed in the Odisha 9 Odisha (OO) cross. Tag loss was greatest for KK (0.35) and lowest for GG (0.19). We found significant variations among crosses for all studied traits suggesting the possibil- ity of including them in genetic improvement pro- grammes of this species. Keywords: survival, male morphotype, diallel cross, tag loss, Macrobrachium rosenbergii Introduction The giant freshwater prawn Macrobrachium rosen- bergii (de Man) is an important species cultured globally, with a production of 215 029 t worth over a billion US dollars in 2010 (FAO 2012). Aquaculture production of this species in India, the third largest producer in 2005 with a produc- tion of 42 800 t, started declining in 2006 and by 2010 it reduced drastically to 13 525 t (FAO 2012). The genetic deterioration of brood stock, possibly due to unintentional inbreeding and nega- tive selection was perceived as a major contribut- ing factor to the decreased productivity. Similar decline in productivity has been reported from countries such as China, Taiwan and Thailand (De Bruyn, Wilson & Mather 2004; New, Valenty, Tidwell, D’Abramo & Kutty 2010). Genetic improvement of cultured stocks through selective breeding is an efficient way of increasing productivity in farmed terrestrial animals and plants, as well as in aquatic animal species (for recent review see, Gjedrem, Robinson & Rye 2012). Favourable response to selective breeding has been observed in a number of commercially © 2014 John Wiley & Sons Ltd 1 Aquaculture Research, 2014, 1–12 doi: 10.1111/are.12419
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Survival, male morphotypes, female and male proportion, female reproductive status and tag loss in...

Survival, male morphotypes, female and male
proportion, female reproductive status and tag loss in
crosses among three populations of freshwater
prawn Macrobrachium rosenbergii (de Man) in India
Bindu R Pillai1, Kanta D Mahapatra1, Raul W Ponzoni2, Lopamudra Sahoo1,†, P L Lalrinsanga1,
Wagdy Mekkawy2,3, Hooi Ling Khaw2, Nguyen H Nguyen2,*, Swagathika Mohanty1,
Sovan Sahu1 & Gunamaya Patra1
1Central Institute of Freshwater Aquaculture, Bhubaneswar, Odisha, India2WorldFish, Bayan Lepas, Penang, Malaysia3Animal Production Department, Faculty of Agriculture, Ain Shams University, Cairo, Egypt
Correspondence: B R Pillai, Principal Scientist, CIFA, Kausalyaganga, Bhubaneswar-751002, Odisha, India. E-mail: pillaibr@
hotmail.com
*Present address: N H Nguyen, School of Science, Education and Engineering, University of the Sunshine Coast, Maroochydore,
QLD 4558, Australia†Present address: L Sahoo, ICAR Research Complex for North East Region, Lembucherra, Tripura 799210, India
Abstract
The present study examined the variation in sur-
vival, proportion of male morphotypes, female and
male proportion, female reproductive status and tag
losses in nine crosses from a complete (3 9 3) dial-
lel mating of three populations of Macrobrachium
rosenbergii in India. The populations originated
from Gujarat (north-west), Kerala (south-west) and
Odisha (east), representing different agro-ecological
regions in India. Progeny from 60 families (4773
juveniles) were individually tagged and reared for
16–17 weeks in earthen ponds. Binary logistic
regression was used to analyse the data. Survival
rate was greatest (0.88) for Kerala 9 Odisha (KO;
the first letter represents the sire), whereas it was
lowest (0.75) for both Kerala 9 Kerala (KK) and
Gujarat 9 Kerala (GK). Significantly greater pro-
portions of large male morphotypes were observed
in KK cross and in combinations involving Kerala
population. The highest proportion of immature
females, as well as of non-berried females, was
observed in the Odisha 9 Odisha (OO) cross. Tag
loss was greatest for KK (0.35) and lowest for GG
(0.19). We found significant variations among
crosses for all studied traits suggesting the possibil-
ity of including them in genetic improvement pro-
grammes of this species.
Keywords: survival, male morphotype, diallel
cross, tag loss, Macrobrachium rosenbergii
Introduction
The giant freshwater prawn Macrobrachium rosen-
bergii (de Man) is an important species cultured
globally, with a production of 215 029 t worth
over a billion US dollars in 2010 (FAO 2012).
Aquaculture production of this species in India,
the third largest producer in 2005 with a produc-
tion of 42 800 t, started declining in 2006 and by
2010 it reduced drastically to 13 525 t (FAO
2012). The genetic deterioration of brood stock,
possibly due to unintentional inbreeding and nega-
tive selection was perceived as a major contribut-
ing factor to the decreased productivity. Similar
decline in productivity has been reported from
countries such as China, Taiwan and Thailand (De
Bruyn, Wilson & Mather 2004; New, Valenty,
Tidwell, D’Abramo & Kutty 2010).
Genetic improvement of cultured stocks through
selective breeding is an efficient way of increasing
productivity in farmed terrestrial animals and
plants, as well as in aquatic animal species (for
recent review see, Gjedrem, Robinson & Rye
2012). Favourable response to selective breeding
has been observed in a number of commercially
© 2014 John Wiley & Sons Ltd 1
Aquaculture Research, 2014, 1–12 doi:10.1111/are.12419

important finfish species such as salmon (Gjedrem
1997; Quinton, Mc Millan & Glebe 2005), tilapia
(Eknath, Bentsen, Ponzoni, Rye, Nguyen & Thode-
sen 2007; Ponzoni, Hamzah, Tan & Kamaruzz-
aman 2005) and rohu carp (Mahapatra, Jana,
Saha, Gjerde & Sarangi 2006). Selective breeding
programmes so far undertaken in commercially
important crustaceans including penaeids (Hetzel,
Crocos, Davis, Moore & Preston 2000; Argue,
Arce, Lotz & Moss 2002; Goyard, Patrois, Peignon,
Vanaa, Dufour, Viallon & Bedier 2002; Preston,
Crocos, Keys, Coman & Koeing 2004; de Donato,
Manrique, Ramirez, Mayer & Howell 2005; Gitte-
rle, Rye, Salte, Cock, Johansen, Lozano, Su�arez &
Gjerde 2005) and crayfish (Jones, Mc Phee & Rus-
coe 2000; Jerry, Purvis, Piper & Dennis 2005)
have also resulted in genetic gains in productivity.
Most aquaculture genetic improvement pro-
grammes have focused on growth traits. However,
there are other potentially important traits that
need to be considered as candidates for inclusion
in the breeding objective. Investigating the magni-
tude of genetic variation in such traits is impor-
tant in order to assess the feasibility of including
them in genetic improvement programmes. The
present investigation was therefore undertaken to
study the variation in important traits like sur-
vival, proportion of male morphotypes, female and
male proportion, female reproductive status as well
as tag loss among the nine crosses of three geo-
graphically distinct populations of M. rosenbergii
from India.
Material and methods
Sampling from different populations
A selective breeding programme aimed at increas-
ing the growth rate of giant freshwater prawn
(GFP) was initiated in 2007 in India, led by the
Central Institute of Freshwater Aquaculture (CIFA)
in collaboration with WorldFish, Malaysia. For the
establishment of the base population, different pop-
ulations of M. rosenbergii were sampled from three
locations in India, viz. Gujarat (north-west), Kerala
(south-west) and Odisha (East). These represent
different agro-ecological regions and are geograph-
ically distant from each other (Figure 1). Juvenile
Figure 1 Map of India showing
the collection sites for the popula-
tions of Macrobrachium rosenbergii
Gujarat, Kerala, Odisha.
© 2014 John Wiley & Sons Ltd, Aquaculture Research, 1–122
Selected traits in nine crosses of M. rosenbergii B R Pillai et al. Aquaculture Research, 2014, 1–12

prawns were collected from Narmada River, near
Bharuch, Gujarat (21º 420 N 72º 580 E) (2100
animals, average body weight (ABW): 0.30 g
(0.11–0.68 g), from the private hatcheries in Ker-
ala (9º 290 N 76º 190 E) [2000, ABW: 0.01 g
(0.008–0.02 g) and 200, ABW: 0.24 g (0.10–
0.55 g)], and from Brahmani River near Rajkani-
ka, Kendrapara, Odisha (20º 440 N 86º 420 E)
(900, ABW: 6.0 g (1.2–12.0 g). The juveniles
were transported to the institute farm and released
into indoor rearing facility separately. After
4 weeks, the three populations were tagged with
different coloured elastomer tags. Visible implant
elastomer tags (VIE) from Northwest Marine Tech-
nology, (NMT), USA, were used: green (Gujarat),
orange (Odisha) and red (Kerala). The tagged
prawns were then released into separate 0.04 ha
earthen ponds for grow-out. Prawns were fed com-
mercial freshwater prawn feed (Godrej Macro Gold,
30% crude protein, 5% lipid) twice daily at 10% of
biomass.
Family production and rearing
The experimental design consisted of a complete
3 9 3 diallel cross comprising three pure-bred mat-
ings (Kerala 9 Kerala (KK); Gujarat 9 Gujarat
(GG); Odisha 9 Odisha (OO)), and six crosses (Guja-
rat 9 Kerala (GK), the first letter representing the
sire; Gujarat 9 Odisha (GO); Kerala 9 Gujarat
(KG); Kerala 9 Odisha (KO); Odisha 9 Gujarat
(OG); Odisha 9 Kerala (OK)). The mating of the
brood stock was carried out in two batches. Table 1
shows the number of mating pairs for each popula-
tion and their combination. A total of 2233 juvenile
prawns (progeny of 27 sires and 30 dams) repre-
senting the three purebred and six crosses of the
three populations of GFP were tagged in the first
batch and another 2540 prawn juveniles (progeny
of 13 sires and 30 dams) representing six crosses
were tagged in the second batch.
For the production of healthy families, mature
female and male parents were carefully chosen to
maximize mating success. One male was stocked
in a mating tank (Ferrocement tanks
3 9 1.2 9 1.0 m) with four mature females (sex
ratio 1:4). Females carrying fertilized eggs (berried
females) were collected from the mating tank
within 24 h of spawning and maintained in sepa-
rate 500-L fibre glass tanks for 14–16 days until
the eggs had changed colour from bright orange
to grey. Grey-berried females were transferred to
the hatchery for egg hatching in 500-L circular
conical bottom fibreglass tanks. Larvae from indi-
vidual families from each population combination
were reared at a stocking density of 50/litre in
brackish water of 12 gL�1 salinity in the same
hatching containers. Clear water larval rearing
technique was employed that required water
exchange of 50% every second day. Larvae were
fed newly hatched brine shrimp nauplii for the first
10 days and a combination of brine shrimp nau-
plii and egg custard (egg, milk powder, mussel
flesh and cod liver oil and vitamin mineral mix).
One thousand postlarvae (PLs) were collected from
each family when approximately 90% of the lar-
vae metamorphosed to PL. They were reared at a
stocking density of 1 PL per litre for the first
20 days in the hatchery. Subsequently, PLs were
transferred to fine mesh hapas (2.5 9 191 m)
fixed in a 0.10 ha earthen pond for further
3 months rearing at a stocking density of 100/m2.
The PL were fed a 30% protein commercial prawn
pellet (Godrej Macro Gold) twice daily at 20% of
biomass.
Animal identification
Two tagging techniques were employed to identify
the animals either at the individual or at the
group level. Visible implant alpha numeric (VIA)
tags of standard size format (1.0 9 2.5 mm) from
Table 1 Numberofsires(s)anddams
(d)thatproducedprogenyforeachpop-
ulationcombinationinbatches1and2
(s1 9 d1)(s2 9 d2)a
Dams
Sires
Gujarat Kerala Odisha
Batch1
(s1 3 d1)
Batch2
(s2 3 d2)
Batch1
(s1 3 d1)
Batch2
(s2 3 d2)
Batch1
(s1 3 d1)
Batch2
(s2 3 d2)
Gujarat (5 9 5) (3 9 5) (3 9 3) (2 9 5) (2 9 2) (2 9 5)
Kerala (1 9 1) (2 9 5) (4 9 4) (2 9 5) (3 9 3) (0)*
Odisha (1 9 1) (2 9 5) (3 9 5) (0)* (5 9 6) (0)*
*No progeny produced.
© 2014 John Wiley & Sons Ltd, Aquaculture Research, 1–12 3
Aquaculture Research, 2014, 1–12 Selected traits in nine crosses of M. rosenbergii B R Pillai et al.

NMT, USA, with alpha numeric codes were used
for individual prawn identification. The VIE tags
mentioned earlier were used as means of identify-
ing prawns at a group level (nine codes, one for
each of the nine crosses using three colours and
three body locations). Purebreds were tagged on
both right and left sides with one colour whereas
crosses were tagged with one colour on one side
only. The VIA tags were inserted on the lateral
side of the second abdominal segment, whereas
the VIE tag was introduced into the ventral side of
the sixth abdominal segment. All prawns were
tagged with both types of tags.
Communal grow-out of tagged juveniles
The communal grow-out of tagged prawns was
carried out in earthen ponds of 0.04 ha
(400 m2) provided with continuous aeration
using an air blower. Two replicate ponds were
used for both batch 1 and batch 2. Stocking den-
sity was 2.8 and 3.2 individuals per square metre
of surface water area in batches 1 and 2 respec-
tively. About 130–140 days after hatching, the
prawns were tagged and stocked in the ponds.
Mean body weight at stocking for batch 1 was
1.3 g (0.46–6.3 g) whereas it was 2.1 g (0.30–
10.9 g) for batch 2. Prawns were fed twice daily
with a 30% protein commercial pellet (Godrej
Macrogold) at a feeding rate of 10% body weight.
Grow-out continued for 16–17 weeks for both
the batches. Water quality parameters (i.e. pH,
dissolved oxygen and total alkalinity) were mea-
sured weekly (once in the morning) whereas tem-
perature was measured twice daily. The ranges
for the measured water quality parameters during
the grow-out period were as follows: temperature
21.9 to 27.5°C; pH 7.5 to 8.4; dissolved oxygen
2.7 to 4.6 mg L�1; total alkalinity 85 to
121 mg L�1 as CaCO3.
Harvest and data recording
At harvest, surviving prawns were collected and
segregated sex-wise. The sex, body length and
bodyweight of each prawn were recorded. Only
those prawns that had a readable VIE tag could be
included in the analyses because otherwise the
effect of cross (the study of which was one of our
main aims) could not be fitted. The male prawns
were classified according to their morphological
appearance into Blue Claw (BC), Orange Claw (OC)
and Small (SM) as described by Kuris, Ra’anan,
Sagi and Cohen (1987). Large males without claws
were classified as no claw (NC) (Hulata, Karplus,
Wohlfarth & Halevy 1990). For the statistical
analysis BC, OC and NC were combined into a
large male category (LM) for comparison with SM.
Females were classified into four female maturity
stages: stage I (Immature, IM), stage II (early
maturity, EM), stage III (maturity, MA) and stage
IV (fully mature, FM) as per Rajyalakshmi (1961).
Ovaries in immature females show no visible sign
of ovarian maturation; the ovary is small and is
confined to the posterior region of the carapace. In
EM females, ovaries can be seen as a small orange
mass on the posterior region of the carapace. In
MA females, ovaries are larger and more intensely
coloured, occupying two-thirds of the carapace
cavity. In FM females, the ovary occupies the entire
carapace cavity. The number of females in EM and
MA categories was small. The reproductively active
females were combined with FM females and cate-
gorized as mature females. Females were further
classified into two categories: those carrying eggs
as berried (OB) females, and those not carrying
eggs as non-berried (NB) females. In this paper, the
maturity and berried status is generically referred
to as ‘female reproductive traits’. For the tag loss
analysis, two groups of surviving prawns at har-
vest were defined: a group that retained readable
VIA tags on their abdomen and another group that
either had lost the VIA tag or had an unreadable
VIA tag. The ‘tag loss’ trait referred to in this paper
is a combination of tag losses and unreadable tags.
Table 2 shows the total number of observations for
different traits and the proportion of the two cate-
gories defined for each trait.
Statistical analysis
A binary logistic regression model (LOGISTIC
REGRESSION procedure, SPSS (Statistical Package
for the Social Sciences) 2011) was fitted to the
data on survival to harvest, male morphotype,
female and male proportion, female reproductive
traits and tag loss.
The model fitted to survival, male morphotype,
female and male proportion, and female reproduc-
tive traits was:
logpijk
1� pijk
� �¼ lþ Ci þ BPj þ eijk
© 2014 John Wiley & Sons Ltd, Aquaculture Research, 1–124
Selected traits in nine crosses of M. rosenbergii B R Pillai et al. Aquaculture Research, 2014, 1–12
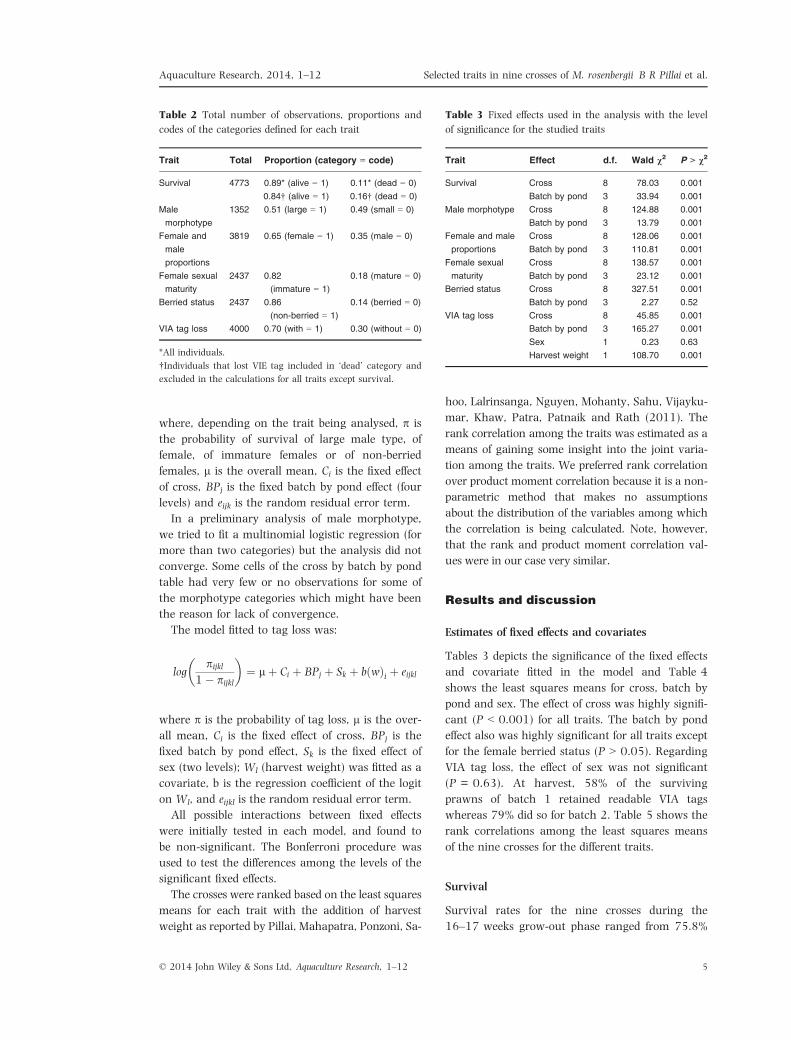
where, depending on the trait being analysed, p is
the probability of survival of large male type, of
female, of immature females or of non-berried
females, l is the overall mean, Ci is the fixed effect
of cross, BPj is the fixed batch by pond effect (four
levels) and eijk is the random residual error term.
In a preliminary analysis of male morphotype,
we tried to fit a multinomial logistic regression (for
more than two categories) but the analysis did not
converge. Some cells of the cross by batch by pond
table had very few or no observations for some of
the morphotype categories which might have been
the reason for lack of convergence.
The model fitted to tag loss was:
logpijkl
1� pijkl
� �¼ lþ Ci þ BPj þ Sk þ bðwÞi þ eijkl
where p is the probability of tag loss, l is the over-
all mean, Ci is the fixed effect of cross, BPj is the
fixed batch by pond effect, Sk is the fixed effect of
sex (two levels); Wl (harvest weight) was fitted as a
covariate, b is the regression coefficient of the logit
on Wl, and eijkl is the random residual error term.
All possible interactions between fixed effects
were initially tested in each model, and found to
be non-significant. The Bonferroni procedure was
used to test the differences among the levels of the
significant fixed effects.
The crosses were ranked based on the least squares
means for each trait with the addition of harvest
weight as reported by Pillai, Mahapatra, Ponzoni, Sa-
hoo, Lalrinsanga, Nguyen, Mohanty, Sahu, Vijayku-
mar, Khaw, Patra, Patnaik and Rath (2011). The
rank correlation among the traits was estimated as a
means of gaining some insight into the joint varia-
tion among the traits. We preferred rank correlation
over product moment correlation because it is a non-
parametric method that makes no assumptions
about the distribution of the variables among which
the correlation is being calculated. Note, however,
that the rank and product moment correlation val-
ues were in our case very similar.
Results and discussion
Estimates of fixed effects and covariates
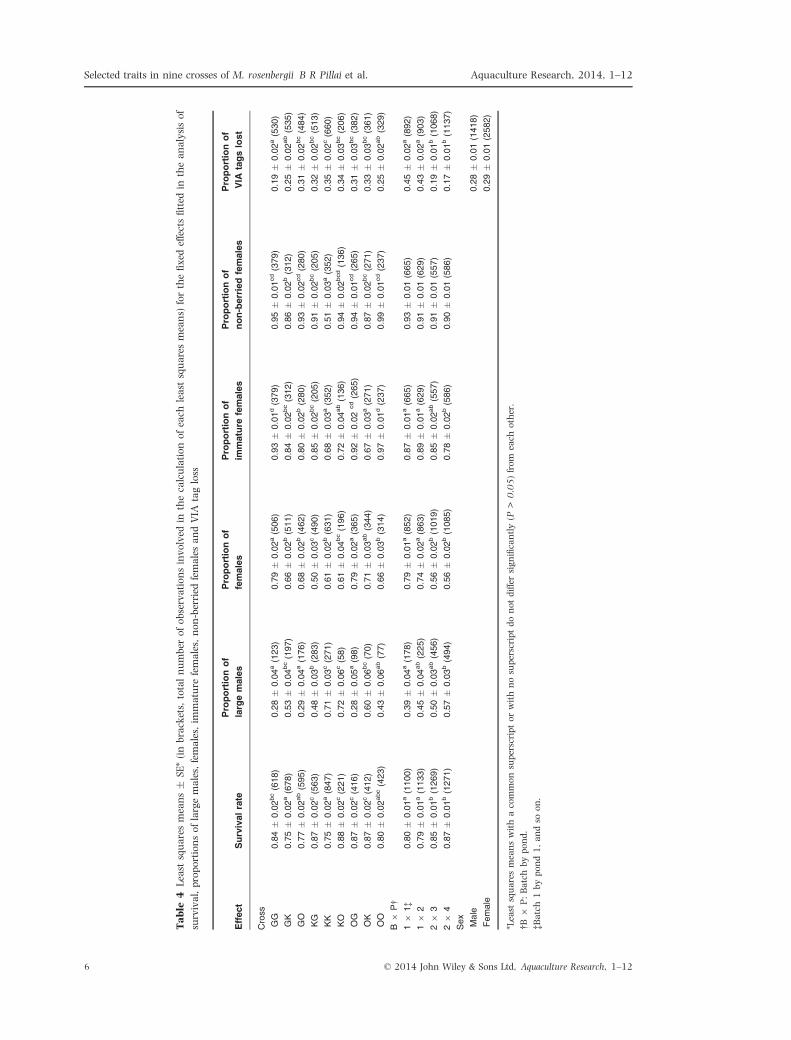
Tables 3 depicts the significance of the fixed effects
and covariate fitted in the model and Table 4
shows the least squares means for cross, batch by
pond and sex. The effect of cross was highly signifi-
cant (P < 0.001) for all traits. The batch by pond
effect also was highly significant for all traits except
for the female berried status (P > 0.05). Regarding
VIA tag loss, the effect of sex was not significant
(P = 0.63). At harvest, 58% of the surviving
prawns of batch 1 retained readable VIA tags
whereas 79% did so for batch 2. Table 5 shows the
rank correlations among the least squares means
of the nine crosses for the different traits.
Survival
Survival rates for the nine crosses during the
16–17 weeks grow-out phase ranged from 75.8%
Table 2 Total number of observations, proportions and
codes of the categories defined for each trait
Trait Total Proportion (category = code)
Survival 4773 0.89* (alive = 1)
0.84† (alive = 1)
0.11* (dead = 0)
0.16† (dead = 0)
Male
morphotype
1352 0.51 (large = 1) 0.49 (small = 0)
Female and
male
proportions
3819 0.65 (female = 1) 0.35 (male = 0)
Female sexual
maturity
2437 0.82
(immature = 1)
0.18 (mature = 0)
Berried status 2437 0.86
(non-berried = 1)
0.14 (berried = 0)
VIA tag loss 4000 0.70 (with = 1) 0.30 (without = 0)
*All individuals.
†Individuals that lost VIE tag included in ‘dead’ category and
excluded in the calculations for all traits except survival.
Table 3 Fixed effects used in the analysis with the level
of significance for the studied traits
Trait Effect d.f. Wald v2 P > v2
Survival Cross 8 78.03 0.001
Batch by pond 3 33.94 0.001
Male morphotype Cross 8 124.88 0.001
Batch by pond 3 13.79 0.001
Female and male
proportions
Cross 8 128.06 0.001
Batch by pond 3 110.81 0.001
Female sexual
maturity
Cross 8 138.57 0.001
Batch by pond 3 23.12 0.001
Berried status Cross 8 327.51 0.001
Batch by pond 3 2.27 0.52
VIA tag loss Cross 8 45.85 0.001
Batch by pond 3 165.27 0.001
Sex 1 0.23 0.63
Harvest weight 1 108.70 0.001
© 2014 John Wiley & Sons Ltd, Aquaculture Research, 1–12 5
Aquaculture Research, 2014, 1–12 Selected traits in nine crosses of M. rosenbergii B R Pillai et al.
Table
4Least
squaresmeans�
SE*(inbrackets,totalnumber
ofobservationsinvolved
inthecalculationofeach
least
squaresmeans)
forthefixed
effectsfitted
intheanalysisof
survival,proportionsoflargemales,females,im
mature
females,non-berried
femalesandVIA
tagloss
Effect
Survivalrate
Proportionof
largemales
Proportionof
females
Proportionof
immature
females
Proportionof
non-berriedfemales
Proportionof
VIA
tagslost
Cross
GG
0.84�
0.02bc(618)
0.28�
0.04a(123)
0.79�
0.02a(506)
0.93�
0.01d(379)
0.95�
0.01cd(379)
0.19�
0.02a(530)
GK
0.75�
0.02a(678)
0.53�
0.04bc(197)
0.66�
0.02b(511)
0.84�
0.02bc(312)
0.86�
0.02b(312)
0.25�
0.02ab(535)
GO
0.77�
0.02ab(595)
0.29�
0.04a(176)
0.68�
0.02b(462)
0.80�
0.02b(280)
0.93�
0.02cd(280)
0.31�
0.02bc(484)
KG
0.87�
0.02c(563)
0.48�
0.03b(283)
0.50�
0.03c(490)
0.85�
0.02bc(205)
0.91�
0.02bc(205)
0.32�
0.02bc(513)
KK
0.75�
0.02a(847)
0.71�
0.03c(271)
0.61�
0.02b(631)
0.68�
0.03a(352)
0.51�
0.03a(352)
0.35�
0.02c(660)
KO
0.88�
0.02c(221)
0.72�
0.06c(58)
0.61�
0.04bc(196)
0.72�
0.04ab(136)
0.94�
0.02bcd(136)
0.34�
0.03bc(206)
OG
0.87�
0.02c(416)
0.28�
0.05a(98)
0.79�
0.02a(365)
0.92�
0.02
cd(265)
0.94�
0.01cd(265)
0.31�
0.03bc(382)
OK
0.87�
0.02c(412)
0.60�
0.06bc(70)
0.71�
0.03ab(344)
0.67�
0.03a(271)
0.87�
0.02bc(271)
0.33�
0.03bc(361)
OO
0.80�
0.02abc(423)
0.43�
0.06ab(77)
0.66�
0.03b(314)
0.97�
0.01d(237)
0.99�
0.01cd(237)
0.25�
0.02ab(329)
B9
P†
19
1‡
0.80�
0.01a(1100)
0.39�
0.04a(178)
0.79�
0.01a(852)
0.87�
0.01a(665)
0.93�
0.01(665)
0.45�
0.02a(892)
19
20.79�
0.01a(1133)
0.45�
0.04ab(225)
0.74�
0.02a(863)
0.89�
0.01a(629)
0.91�
0.01(629)
0.43�
0.02a(903)
29
30.85�
0.01b(1269)
0.50�
0.03ab(456)
0.56�
0.02b(1019)
0.85�
0.02ab(557)
0.91�
0.01(557)
0.19�
0.01b(1068)
29
40.87�
0.01b(1271)
0.57�
0.03b(494)
0.56�
0.02b(1085)
0.78�
0.02b(586)
0.90�
0.01(586)
0.17�
0.01b(1137)
Sex
Male
0.28�
0.01(1418)
Female
0.29�
0.01(2582)
*Least
squaresmeanswithacommonsuperscriptorwithnosuperscriptdonotdiffersignificantly(P
>0.05)from
each
other.
†B9
P:Batchbypond.
‡Batch1bypond1,andso
on.
© 2014 John Wiley & Sons Ltd, Aquaculture Research, 1–126
Selected traits in nine crosses of M. rosenbergii B R Pillai et al. Aquaculture Research, 2014, 1–12
to 88.5%. These values are consistent with those
reported for this species (New et al. 2010). In our
data, because we only recorded as surviving those
prawns that had a VIE tag, the number of missing
VIE tags is confounded with the number of dead
prawns within cross. Accordingly, our estimate of
survival is underestimated. Note, however, that
losses of VIE tags were relatively small (6.2% in
batch 1 and 3.7% in batch 2). We have no evi-
dence of differential VIE tag losses among crosses
so the differences between crosses in survival is
most likely unbiased. Among the three popula-
tions, Kerala population showed the lowest sur-
vival whereas Gujarat population showed the
highest survival. The KK, GK, GO and OO had sig-
nificantly lower survival compared to other
crosses. After a grow-out period of 8 months in
earthen ponds, Aflalo, Raju, Bommi, Verghese,
Samraj, Hulata, Ovadia and Sagi (2012) reported
survival rates of approximately 20% to 60%
among nine different crosses of three populations
of M. rosenbergii from India indicating among
cross variation for this trait. By contrast, in Viet-
nam, Thanh, Ponzoni, Nguyen, Vu, Barnes and
Mather (2009) did not find significant differences
in survival among nine crosses of three popula-
tions of M. rosenbergii during a 15-week grow-out
period in hapas. They report survival rates rang-
ing from 73.3 to 77.4%. Survival rates in our
study were similar to those of Thanh et al.
(2009). Note that the latter authors recorded sur-
vival rates in a grow-out period of approximately
the same duration as ours. However, unlike
Thanh et al. (2009) we found significant among
cross variation in survival rate. The rank correla-
tions between survival rate of the different crosses
and the other traits were low and statistically not
significant (Table 5).
Proportion of male morphotypes
Adult M. rosenbergii populations from culture
ponds exhibit a wide range of sizes, especially
among males. This variability poses a challenge to
increased profitability because prawn prices are
size dependent. Karplus and Sagi (2010) observed
that in male GFP populations live weights are usu-
ally bi-modal or multi-modal and positively
skewed, comprising a large fraction of small,
unmarketable individuals and a smaller fraction of
large prawns. Earlier workers found that prawn
size variation is the consequence of complex intra-
population interactions that result in a distribution
comprising three distinct male classes viz. SM, OC
and BC. These classes differ in their morphology,
physiology, behaviour and transform from SM to
OC and then to BC (Karplus & Sagi 2010). The BC
males are characterized by very long, robust sec-
ond walking legs that are deep blue, and are the
largest of the three morphotypes. They are aggres-
sive, territorial and dominant over the other two
morphotypes (Karplus, Malecha & Sagi 2000). A
second type of male, SM, with small claws, was
first described in pond populations (Fujimura &
Okamoto 1972). The SM are the smallest of the
three morphotypes and are slow growing and sub-
ordinate to both BC and OC. Sandifer and Smith
(1977) described a faster growing third male mor-
photype, OC, that also grows to a large size and
has long, second walking legs and orange claws.
A greater proportion of BC and OC morphotypes is
preferred under commercial grow-out because the
yield and the market value are greater. Our study
revealed a significant difference in the proportion
of male morphotypes among the crosses. The Ker-
ala population showed the greatest proportion of
large male morphotypes, whereas it was lowest in
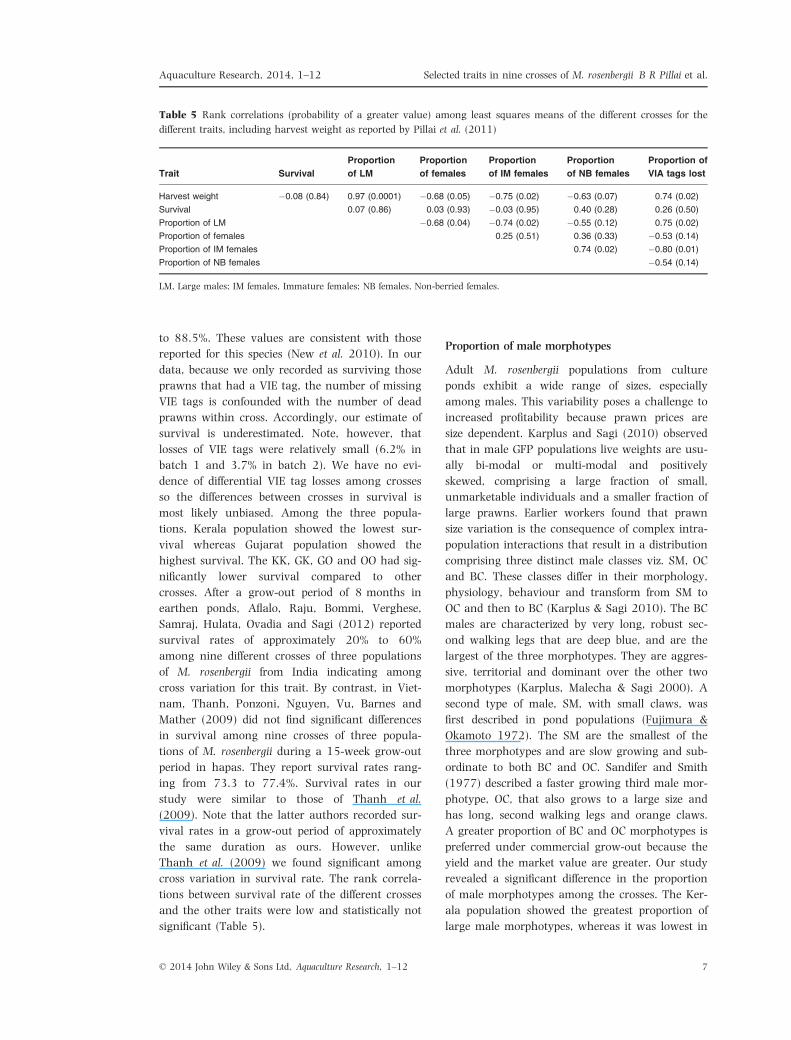
Table 5 Rank correlations (probability of a greater value) among least squares means of the different crosses for the
different traits, including harvest weight as reported by Pillai et al. (2011)
Trait Survival
Proportion
of LM
Proportion
of females
Proportion
of IM females
Proportion
of NB females
Proportion of
VIA tags lost
Harvest weight �0.08 (0.84) 0.97 (0.0001) �0.68 (0.05) �0.75 (0.02) �0.63 (0.07) 0.74 (0.02)
Survival 0.07 (0.86) 0.03 (0.93) �0.03 (0.95) 0.40 (0.28) 0.26 (0.50)
Proportion of LM �0.68 (0.04) �0.74 (0.02) �0.55 (0.12) 0.75 (0.02)
Proportion of females 0.25 (0.51) 0.36 (0.33) �0.53 (0.14)
Proportion of IM females 0.74 (0.02) �0.80 (0.01)
Proportion of NB females �0.54 (0.14)
LM, Large males; IM females, Immature females; NB females, Non-berried females.
© 2014 John Wiley & Sons Ltd, Aquaculture Research, 1–12 7
Aquaculture Research, 2014, 1–12 Selected traits in nine crosses of M. rosenbergii B R Pillai et al.

the Gujarat population and intermediate in the
Odisha population. Among the crosses, KO and OK
had a significantly greater proportion of large male
morphotypes, whereas it was significantly lower in
OG and GO. Greater proportions of large male
morphotypes in KK, KO and OK would result in
greater production compared to the other crosses.
Among OO, OG, GG and GO the frequency of the
SM morphotype was significantly greater (>60%)
indicating poor yield from these crosses.
Earlier studies of the distribution of male mor-
photypes during grow-out culture of M. rosenbergii
indicate a density-dependent trend, with a larger
proportion of SM morphotype (and lower propor-
tion of BC and OC) at greater stocking densities
(D’Abramo, Heinen, Robinette & Collins 1989;
Karplus, Hulata, Wohlfarth & Halevy 1986; Ti-
dwell, Webster, Goodgame-Tiu & D’Abramo 1994;
Tidwell, Coyle & Schulmeister 1998). Karplus et al.
(1986) suggest that the changes in morphotype
frequencies with density reflect increased stunting
of prawn populations with increasing densities. In
our study, prawns from different crosses exhibited
differences even when communally reared in a
pond. Hence, the observed differences in the fre-
quency of male morphotypes among crosses could
not be due to density, which suggests that there
may be genetic variation in the potential for
expression of the various morphotypes. Aflalo et al.
(2012) examined the male morphotype distribu-
tion in different populations of M. rosenbergii in a
diallel cross experiment involving three popula-
tions. They reported differences in the frequency of
male morphotypes among different crosses, with a
greater proportion of large morphotypes (BC and
OC) in a pure-bred population from West Bengal,
as well as in its cross with Kerala population.
The rank correlations between the cross least
squares means of proportion of large males and
other traits showed a significant (positive) associa-
tion with harvest weight and VIA tag losses. By
contrast, there was a negative association with the
proportion of females relative to males, and of
immature and non-berried females.
Female and male proportions
The proportion of females was greatest in the Guj-
arat population and lowest in the Kerala popula-
tion. Overall, there were a greater proportion of
females in all crosses except in KG. The GG and
OG had significantly more females (79%) than the
other crosses. Earlier studies report greater propor-
tions of female M. rosenbergii in cultured popula-
tions from different geographical areas (Willis &
Berrigan 1977; Smith, Sandifer & Smith 1978;
Sandifer, Smith, Stokes & Jenkins 1982; D’Abra-
mo, Robinette, Heinen, Ra’anan & Cohen 1986;
Karplus et al. 1986; Jose, Joseph, Shyama, Mohan
& Nair 2007). In a diallel cross experiment involv-
ing three populations of M. rosenbergii in Vietnam,
Thanh et al. (2009) also report sex ratios that
were significantly different from 1:1, in favour of
females in all populations. Malecha, Nevin, Ha,
Barck, La Madrid-Rose, Masuno and Hedgecock
(1992) found a greater proportion of females in
the progeny of five crosses between normal males
(not sex reversed) and females. So far, there have
been no reports on the effect of density on female
to male ratio. Several authors attribute the preva-
lence of females in prawn populations to a greater
male mortality. New et al. (2010) notes that
prawns are vulnerable to limb damage and canni-
balism during moulting even under conditions of
excess food. Hence, the selective male mortality
hypothesis seems plausible under crowded pond
culture conditions. The greater proportion of
females in the present study could partly be attrib-
uted to greater male mortality. It would be inter-
esting to investigate as to what phase of the
development of males and females the sex imbal-
ance sets in (e.g. zygote, larvae, juvenile).
The most obvious feature emerging from the
rank correlations among the least squares means
for the crosses involving male and female propor-
tions was that a greater proportion of females were
associated with lower harvest weight (Table 5).
Female reproductive status
There were marked variations in the proportions
of immature females among the different crosses.
The pure-bred Odisha population had the highest
proportion of immature females (97%), followed by
the Gujarat population (93%). By contrast, the
pure-bred Kerala population had about 32% of
females having ovaries with developing oocytes.
Among crosses, OG, KG, GK and GO had signifi-
cantly greater proportions of immature females
whereas it was lower in OK and KO. The propor-
tion of non-berried females exhibited a similar
trend as that of immature females.
Generally, female growth in decapod crustaceans
slows down after they reach maturity because part
© 2014 John Wiley & Sons Ltd, Aquaculture Research, 1–128
Selected traits in nine crosses of M. rosenbergii B R Pillai et al. Aquaculture Research, 2014, 1–12

of the energy intake is diverted to egg production
and incubation (Hartnoll 1982). Freshwater
prawns can reproduce either continuously or peri-
odically, depending on their geographical location
(New et al. 2010). In tropical regions where the
water temperature remains above 24°C for most of
the year, reproduction occurs continuously (Ling
1969). By contrast, in monsoon regions, reproduc-
tion is seasonal (New et al. 2010). Ra’anan, Sagi,
Wax, Karplus, Hulata and Kuris (1991) report that
the growth rate of immature female M. rosenbergii
is relatively high, initially exponential, but after
sexual maturation, this rate reduces considerably
as the reproductive processes require significant
energy mobilization. Aflalo et al. (2012) reported
differences in reproductive performance among
crosses of M. rosenbergii in a diallel experiment
involving three populations from India. There was
a greater proportion of females in progressive repro-
ductive stages (bearing developed gonads or ber-
ried) in the pure-bred West Bengal population and
its crosses when it was used as a male parent. This
indicates the existence of population specific differ-
ences in reproductive activity of female M. rosen-
bergii. Populations showing fast rate of maturation
are not preferred for culture as this would retard
the growth rate and the presence of greater number
of egg carrying females further reduces the price.
The observed differences in proportion of reproduc-
tively active females among different populations of
M. rosenbergii may affect the yield and returns from
farming. It may however be noted that the rank
correlations between least squares means of the
crosses for harvest weight with the female repro-
ductive traits (proportion of immature and of non-
berried females) were negative (Table 5). This
indicates that across populations, heavier weight
was associated with a lower proportion of imma-
ture and non-berried females, although female sex-
ual maturation itself may slow down growth rate.
VIA tag loss
Two types of tags were used in the present study;
VIE as a batch tag to identify the crosses and VIA
tags for individual prawn identification. The losses
among the former were considerably lower than
the latter. Furthermore, all the retained VIE tags
were readable. Earlier studies on decapod crusta-
ceans including freshwater prawn on retention
and readability of VIE tags (Woods & James 2003;
Ashley & Jean-Paul 2006; Pillai, Rath & Sahu
2007) compare well with the results from our
study. By contrast, studies on retention and read-
ability of VIA tags report higher retention and
readability than what we experienced. Isely and
Stockett (2001) report 100% readability of VIA
tags in juvenile red swamp crayfish Procabarus
clarkii 150 days posttagging. Jerry, Stewart, Purvis
and Piper (2001) reported 96% readability in juve-
nile yabby (Cherax destructor). The lower readabil-
ity of VIA tag observed in our study could partly
be attributed to the pigmented and thicker shell
(exoskeleton) of the adult M. rosenbergii. Melaniza-
tion of tissues surrounding the tag could also have
resulted in decreased readability of VIA tags.
Under laboratory conditions, Pillai, Sahoo, Ma-
hapatra, Ponzoni, Sahu, Mohanty, Vijaykumar
and Sahu (2009) did not find any significant effect
of VIA tags on growth and survival of M. rosen-
bergii. However, our results indicated that higher
body weight was associated with greater losses of
VIA tags. This finding is important because it may
affect the family based selective breeding pro-
gramme for improved harvest weight of M. rosen-
bergii that requires individual prawn identification.
Greater tag losses among the heaviest individuals
may lead to lower selection intensity and reduced
selection response.
Note that the differences between crosses were
non-significant (results are not presented) when
the male morphotype was fitted as a fixed effect in
the statistical model used to analyse tag loss. The
effect of harvest weight as a covariate remained
significant but with lower odds ratio (2%) com-
pared with the analysis that did not include male
morphotype (3%). The BC morphotype had the
greatest tag loss (0.61) compared to the other mor-
photypes which ranged from 0.36 to 0.41. We also
found a significant difference in tag loss between
berried (0.31) and non-berried females (0.25). The
differences between crosses remained significant
when female maturity stage was included in the
model as a fixed effect. Fitting harvest weight as a
covariate did not affect the significance of the other
fixed effects. Table 5 shows that tag losses were
greater in the heavier crosses, whereas they were
lower among those with a greater proportion of
females, especially immature ones.
Conclusions
Evidence of variation among populations and
among crosses in survival rate, proportion of male
© 2014 John Wiley & Sons Ltd, Aquaculture Research, 1–12 9
Aquaculture Research, 2014, 1–12 Selected traits in nine crosses of M. rosenbergii B R Pillai et al.

morphotypes, female and male proportions and
state of reproductive maturity in females were
observed. These are biological attributes of impor-
tance in the production system. The presence of
variation among populations and crosses in
prawns bred and reared in a common environ-
ment suggests that the observed variation could
have a genetic basis. This variation could be
exploited in a synthetic population by including all
populations and crosses and selection practised in
ushering in genetic improvement. Greater VIA tag
losses among heavier animals (and among the lar-
ger male morphotypes in particular) are a concern
since they are bound to affect selection intensity
and response. Freshwater prawn has unique fea-
tures which are absent in other species. The pres-
ent study indicates two avenues that appear as
potentially useful to achieving large increases in
productivity: (1) an increase in the proportion of
males and (2) an increase in the frequency of the
larger male morphotypes.
Acknowledgments
This study was carried out at the Central Insti-
tute of Freshwater Aquaculture, Bhubaneswar
under a bilateral collaborative research project
between Indian Council of Agricultural Research
(ICAR), New Delhi, India and WorldFish, Malay-
sia. The authors express sincere gratitude to Dr.
S. Ayyappan, Secretary, Department of Agricul-
tural Research and Education and Director Gen-
eral, ICAR and former Deputy Director General
(Fisheries), ICAR for his keen interest and support
to this work. We are thankful to the Director,
CIFA for encouragement and for providing facili-
ties. Help received from the state fisheries depart-
ments of Gujarat, Kerala and Odisha during the
collection of different populations of M. rosenbegii
is gratefully acknowledged. This work is a contri-
bution to the CGIAR Research Program on Live-
stock and Fish.
References
Aflalo E.D., Raju V.S.N.D., Bommi N.A., Verghese J.T.,
Samraj T.Y.C., Hulata G., Ovadia O. & Sagi A. (2012)
Toward a sustainable production of genetically
improved all-male prawn (Macrobrachium rosenbergii):
evaluation of production traits and obtaining neo-
females in three Indian strains. Aquaculture 338,
197–207.
Argue J.B., Arce S.M., Lotz M.J. & Moss S.M. (2002)
Selective breeding of Pacific white shrimp (Litopenaeus
vannamei) for growth and resistance to Taura
Syndrome Virus. Aquaculture 204, 447–460.
Ashley F. & Jean-Paul A.H. (2006) Long-term retention
of internal elastomer tags in a wild population of
painted crayfish (Panulirus versicolor [Latreille]) on the
great barrier reef. Journal of Experimental Marine
Biology and Ecology 339, 104–110.
D’Abramo L.R., Robinette H.R., Heinen J.M., Ra’anan Z.
& Cohen D. (1986) Polyculture of the freshwater
prawn (Macrobrachium rosenbergii) with a mixed size
population of channel catfish (Ictalurus punctatus).
Aquaculture 59, 71–80.
D’Abramo L.R., Heinen J.M., Robinette H.R. & Collins J.S.
(1989) Production of the freshwater prawn Macrob-
rachium rosenbergii stocked as juveniles at different den-
sities in temperature zone ponds. Journal of World
Aquaculture Society 20, 81–89.
De Bruyn M., Wilson J.A. & Mather P.B. (2004) Huxley’s
line demarcates extensive divergence between eastern
and western forms of the giant freshwater prawn,
Macrobrachium rosenbergii. Molecular Phylogenetics and
Evolution 30, 251–257.
de Donato M., Manrique R., Ramirez R., Mayer I. &
Howell C. (2005) Mass selection and inbreeding effects
on a cultivated strain of Penaeus (Litopenaeus) vannamei
in Venezuela. Aquaculture 247, 159–167.
Eknath A.E., Bentsen H.B., Ponzoni R.W., Rye M.,
Nguyen N.H. & Thodesen J. (2007) Genetic improve-
ment of farmed tilapias: composition and genetic
parameters of a synthetic base population of Oreochr-
omis niloticus for selective breeding. Aquaculture 273,
1–14.
FAO (2012) Statistics and Information Service of the Fisher-
ies and Aquaculture Department. FAO Yearbook, Fishery
and Aquaculture Statistics 2010, FAO, Rome.
Fujimura T. & Okamoto H. (1972) Notes on progress
made in developing a mass culturing technique for
Macrobrachium rosenbergii in Hawaii. In: Coastal Aqua-
culture in the Indo-Pacific Region (ed. by T.V.R. Pillay),
pp. 313–327. Fishing News Books, Blackwell Science,
Oxford.
Gitterle T., Rye M., Salte R., Cock J., Johansen H., Lozano
C., Su�arez J.A. & Gjerde B. (2005) Genetic (co)varia-
tion in harvest body weight and survival in Penaeus
(Litopenaeus) vannamei under standard commercial con-
ditions. Aquaculture 243, 83–92.
Gjedrem T. (1997) Selective breeding to improve aqua-
culture production. World Aquaculture 28, 33–45.
Gjedrem T., Robinson N. & Rye M. (2012) The impor-
tance of selective breeding in aquaculture to meet
future demands for animal protein. Aquaculture 350–
353, 117–129.
Goyard E., Patrois J., Peignon J.M., Vanaa V., Dufour R.,
Viallon J. & Bedier E. (2002) Selection for better
© 2014 John Wiley & Sons Ltd, Aquaculture Research, 1–1210
Selected traits in nine crosses of M. rosenbergii B R Pillai et al. Aquaculture Research, 2014, 1–12

growth of Penaeus stylirostris in Tahiti and New
Caledonia. Aquaculture 204, 461–468.
Hartnoll R.G. (1982) Growth. In: The Biology of Crustacea,
Vol. 2 (ed. by L.G. Abele), pp. 111–196, Embryology,
Morphology and Genetics. Academic Press, New York.
Hetzel D.J.S., Crocos P.J., Davis G.P., Moore S.S. & Pres-
ton N.C. (2000) Response to selection and heritability
for growth in the Kuruma prawn, Penaeus japonicus.
Aquaculture 181, 215–223.
Hulata G., Karplus I., Wohlfarth G.W. & Halevy A.
(1990) Effects of size and age of juvenile freshwater
prawn, Macrobrachium rosenbergii at stocking on popu-
lation structure and production in polyculture ponds.
Journal of World Aquaculture Society 21, 295–299.
Isely J.J. & Stockett P.E. (2001) Tag retention, growth
and survival of red swamp crayfish marked with a visi-
ble implant tag. North American Journal of Fisheries
Management 21, 422–424.
Jerry D.R., Stewart T., Purvis I.W. & Piper I.R. (2001)
Evaluation of visual implant elastomer and alphanu-
meric internal tag as a method to identify juveniles of
freshwater crayfish Cherax destructor. Aquaculture 193,
149–154.
Jerry D.R., Purvis I.W., Piper L.R. & Dennis C.A. (2005)
Selection for faster growth in the freshwater crayfish
Cherax destructor. Aquaculture 247, 169–176.
Jones C.M., Mc Phee C.P. & Ruscoe I.M. (2000) A review
of genetic improvement in growth rate in redclaw
crayfish Cherax quadricarinatus (von Martens) (Deca-
poda: Parastacidae). Aquaculture Research 31, 61–67.
Jose S., Joseph A., Shyama S., Mohan M.V. & Nair C.M.
(2007) Production trends of the giant freshwater
prawn, Macrobrachium rosenbergii (De Man) in two cul-
ture strategies. In: Proceedings of the International Sym-
posium on Freshwater prawns, 20–23 August 2003 (ed.
by C.M. Nair, D.D. Nambudiri, J. Susheela, T.M. Sanka-
ran, K.V. Jayachandran & K.R. Salin), pp. 404–409.
Kochi, India.
Karplus I. & Sagi A. (2010) The biology and manage-
ment of size variation. In: Freshwater Prawns Biology
and Farming (ed. by M.B. New, W.C. Valenty, J.C. Ti-
dwell, L. D Abramo & N. Kutty), pp. 316–347. Black-
well Science, Oxford.
Karplus I., Hulata G., Wohlfarth G.W. & Halevy A.
(1986) The effect of density of the Macrobrachium ro-
senbergii raised in earthen ponds on their population
structure and weight distribution. Aquaculture 52,
307–320.
Karplus I., Malecha S.R. & Sagi A. (2000) The biology
and management of size variation. In: Freshwater
Prawn Culture (ed. by M.B. New & W.C. Valenti), pp.
259–289. Blackwell Science, London.
Kuris A.M., Ra’anan Z., Sagi A. & Cohen D. (1987) Mor-
phometric differentiation of male Malaysian giant
prawns, Macrobrachium rosenbergii. Journal of Crustacean
Biology 7, 219–237.
Ling S.W. (1969) The general biology and development
of Macrobrachium rosenbergii (de Man), In: Proceedings
of World Scientific Conference on the Biology and Culture
of Shrimps and Prawns, Vol. 57, FAO, Rome, pp.
589–606.
Mahapatra K.D., Jana R.K., Saha J.N., Gjerde B. & Sarangi
N. (2006) Lessons from breeding program of rohu. In:
Development of Aquatic Animal Genetic Improvement and
Dissemination Programs: Current Status and Action Plans,
Vol. 73 (ed. by R.W. Ponzoni, B.O. Acosta & A.G. Ponn-
iah), pp. 34–40. WorldFish Center Conference Proceed-
ings. The WorldFish Center, Penang, Malaysia.
Malecha S.R., Nevin P.A., Ha P., Barck L.E., La Madrid-
Rose Y., Masuno S. & Hedgecock D. (1992) Sex-ratios
and sex determination in progeny from crosses of sur-
gically sex-reversed freshwater prawns, Macrobrachium
rosenbergii. Aquaculture 105, 201–218.
New M.B., Valenty W.C., Tidwell J.C., D’Abramo L. & Ku-
tty N. (2010) Freshwater Prawns Biology and Farming.
Wiley-Blackwell, Oxford.
Pillai B.R., Rath S.C. & Sahu S. (2007) Evaluation of a vis-
ible implant fluorescent elastomer internal tag in juve-
nile freshwater prawn Macrobrachium rosenbergii (de
Man). Indian Journal of Animal Science 77, 1054–1056.
Pillai B.R., Sahoo L., Mahapatra K.D., Ponzoni R.W.,
Sahu S., Mohanty S., Vijaykumar & Sahu S. (2009)
Evaluation of a new fluorescent internal tag (soft visi-
ble implant alphanumeric tag) in freshwater prawn
Macrobrachium rosenbergii. Israeli Journal of Aquaculture-
Bamidgeh 61, 345–350.
Pillai B.R., Mahapatra K.D., Ponzoni R.W., Sahoo L.,
Lalrinsanga P.L., Nguyen N.H., Mohanty S., Sahu S.,
Vijaykumar, Sahu S., Khaw H.L., Patra G., Patnaik S. &
Rath S.C. (2011) Genetic evaluation of a complete
diallel cross involving three populations of freshwater
prawn (Macrobrachium rosenbergii) from different geo-
graphical regions of India. Aquaculture 319, 347–354.
Ponzoni R.W., Hamzah A., Tan S. & Kamaruzzaman N.
(2005) Genetic parameters and response to selection
for live weight in the GIFT strain of Nile tilapia (Ore-
ochromis niloticus). Aquaculture 247, 203–210.
Preston N.P., Crocos P.J., Keys S.J., Coman G.J. & Koeing
R. (2004) Comparative growth of selected and non-
selected Kuruma shrimp Penaeus (Marsupenaeus) japoni-
cus in commercial farm ponds; implications for brood-
stock production. Aquaculture 231, 73–82.
Quinton C., Mc Millan I. & Glebe B.D. (2005) Develop-
ment of Atlantic salmon (Salmo salar) genetic improve-
ment program. Genetic parameters of harvest body
weight and carcass quality traits estimated with ani-
mal models. Aquaculture 247, 211–217.
Ra’anan Z., Sagi A., Wax Y., Karplus I., Hulata G. & Ku-
ris A.M. (1991) Growth, size rank, and maturation of
the freshwater prawn, Macrobrachium rosenbergii:
analysis of marked prawns in an experimental popula-
tion. Biological Bulletin 181, 379–386.
© 2014 John Wiley & Sons Ltd, Aquaculture Research, 1–12 11
Aquaculture Research, 2014, 1–12 Selected traits in nine crosses of M. rosenbergii B R Pillai et al.

Rajyalakshmi T. (1961) Studies on maturation and
breeding in some estuarine palaemonid prawns.
Proceedings of the National Institute of Science India 27,
179–188.
Sandifer P.A. & Smith T.I.J. (1977) Intensive rearing of
post larval Malaysian prawns (Macrobrachium rosen-
bergii) in a closed cycle nursery system. Proceedings of
World Mariculture Society 8, 225–235.
Sandifer P.A., Smith T.I.J., Stokes A.D. & Jenkins W.E.
(1982) Semi-intensive grow-out of prawns (Macrob-
rachium rosenbergii): preliminary results and prospects.
In: Giant Prawn Farming: Developments in Aquaculture
and Fisheries Science, Vol. 10 (ed. by M.B. New),
pp. 161–172. Elsevier, Amsterdam.
Smith T.I.J., Sandifer P.A. & Smith M.H. (1978) Popula-
tion structure of Malaysian prawns, Macrobrachium ro-
senbergii (De Man) reared in earthen ponds in South
Carolina 1974–1976. Proceedings of World Mariculture
Society 9, 21–38.
SPSS (Statistical Package for the Social Sciences) (2011).
IBM SPSS Statistics for Windows, Version 19.0. IBM,
Armonk, NY.
Thanh N.M., Ponzoni R.W., Nguyen N.H., Vu N.T.,
Barnes A. & Mather P.B. (2009) Evaluation of growth
performance in a diallel cross of three strains of giant
freshwater prawn (Macrobrachium rosenbergii) in Viet-
nam. Aquaculture 287, 75–83.
Tidwell J.H., Webster C.D., Goodgame-Tiu L. & D’Abramo
L.R. (1994) Population characteristics of Macrobrachi-
um rosenbergii fed diets containing different protein
sources under cool water conditions in earthen ponds.
Aquaculture 126, 271–281.
Tidwell J.H., Coyle S.D. & Schulmeister G. (1998) Effects
of added substrate on the production and population
characteristics of freshwater prawns Macrobrachium ro-
senbergii in ponds. Journal of World Aquaculture Society
29, 17–22.
Willis S.A. & Berrigan M.E. (1977) Effect of stocking size
and density on growth and survival of Macrobrachium
rosenbergii in ponds. Proceedings of World Mariculture
Society 8, 251–264.
Woods C.M.C. & James P.J. (2003) Evaluation of visible
implant fluorescent elastomer (VIE) as a tagging tech-
nique for spiny lobsters (Jasus Edwardsii). Marine and
Freshwater Research 54, 853–858.
© 2014 John Wiley & Sons Ltd, Aquaculture Research, 1–1212
Selected traits in nine crosses of M. rosenbergii B R Pillai et al. Aquaculture Research, 2014, 1–12




















