
The forage type (grazing versus hay pasture) fed to ewes and the lamb sex affect fatty acid profile...
15
E. Dervishi, M. Joy, J. Alvarez-Rodriguez, M. Serrano and J. H. Calvo acid profile and lipogenic gene expression in the longissimus muscle of suckling lambs The forage type (grazing versus hay pasture) fed to ewes and the lamb sex affect fatty doi: 10.2527/jas.2011-4057 originally published online August 19, 2011 2012, 90:54-66. J ANIM SCI http://jas.fass.org/content/90/1/54 the World Wide Web at: The online version of this article, along with updated information and services, is located on www.asas.org at Universitat de Lleida on January 4, 2012 jas.fass.org Downloaded from
Transcript of The forage type (grazing versus hay pasture) fed to ewes and the lamb sex affect fatty acid profile...

E. Dervishi, M. Joy, J. Alvarez-Rodriguez, M. Serrano and J. H. Calvoacid profile and lipogenic gene expression in the longissimus muscle of suckling lambsThe forage type (grazing versus hay pasture) fed to ewes and the lamb sex affect fatty
doi: 10.2527/jas.2011-4057 originally published online August 19, 20112012, 90:54-66.J ANIM SCI
http://jas.fass.org/content/90/1/54the World Wide Web at:
The online version of this article, along with updated information and services, is located on
www.asas.org
at Universitat de Lleida on January 4, 2012jas.fass.orgDownloaded from

The forage type (grazing versus hay pasture) fed to ewes and the lamb sex affect fatty acid profile and lipogenic gene
expression in the longissimus muscle of suckling lambs1
E. Dervishi,* M. Joy,* J. Alvarez-Rodriguez,*2 M. Serrano,† and J. H. Calvo*‡3
*Unidad de Tecnología en Producción Animal, Centro de Investigación y Tecnología Agroalimentaria (CITA), Avda Montañana 930, 50059 Zaragoza, Spain; †Departamento de Mejora Genética Animal, Instituto Nacional
de Investigación y Tecnología Agraria y Alimentaria (INIA), Ctra. La Coruña 7.5, 28040 Madrid, Spain; and ‡ARAID Fundacion Agencia Aragonesa para la Investigacion y Desarrollo, 50004 Zaragoza, Spain
ABSTRACT: Meat intramuscular fat (IMF) contrib-utes to meat quality and consumer acceptance. Molecu-lar events that occur during IMF deposition and the identification of genes that are differentially expressed during this process are important to the design of an optimal nutrition plan for animals. In the present study, we examined the effect of the forage type (grazing vs. hay pasture) fed to ewes and the effect of lamb sex on the LM fatty acid (FA) profile and gene expression of suckling lambs (10 to 12 kg of BW at slaughter); ewes received pasture hay (PH) or grazed pasture (GRE). Forage type had a significant effect on IMF FA pro-file. Ewes grazing green forage (GRE) promoted the formation and deposition of vaccenic acid (C18:1n-7), CLA, and PUFA n-3 in LM from their suckling lambs (P < 0.05). We found that forage type affected the expression of the sterol regulatory element binding transcription factor 1 (SREBF1) gene in females. How-
ever, in males, it modulated stearoyl CoA desaturase (SCD) gene expression (P < 0.05). Moreover, our re-sults showed that females, independent of the diet of the ewes (PH or GRE), are predisposed to develop fat and to upregulate the expression of key genes of tran-scriptional factors PPARA, CEBPB, SREBF1, and li-poprotein lipase (LPL) and SCD (P < 0.05). The data suggest that SREBF1, SCD, and most likely CEBPB gene expression in young suckling lambs is modulated by both lamb sex and forage type fed to ewes. Fat-ty acid indicators PUFA, n-6/n-3, CLA, and SFA are closely related to LPL, SCD, PPARA, and CEBPB gene expression depending on animal sex or the diet of ewes. This study suggests that grazing pasture affects FA composition promoting greater vaccenic, CLA, and total PUFA n-3 FA in female and male suckling lambs, and it is mediated through the regulation of lipogenic enzyme expression.
Key words: diet, ewe, fatty acid, gene expression, meat quality, suckling lamb
©2012 American Society of Animal Science. All rights reserved. J. Anim. Sci. 2012. 90:54–66 doi:10.2527/jas.2011-4057
INTRODUCTION
Accumulation of intramuscular fat (IMF) is influ-enced by animal intrinsic factors, such as genetics and age, and by extrinsic factors, such as the animal nutri-tion plan. Increasing demand for safe meat products is stimulating pasture-based production systems, and grazing systems can be a good alternative to achieve good animal performance (Alvarez-Rodriguez et al., 2007; Joy et al., 2008).
Previous studies have shown that feeding strategies, such as concentrate, pasture, or both, influence the fat-ty acid (FA) composition of meat IMF and milk fat in ruminants, but no studies have been performed about forage type. Moreover, forage type can alter the expres-sion of genes that are associated with fat metabolism (Wood and Enser, 1997; Scerra et al., 2007; Bernard et al., 2008; Graugnard et al., 2009; Dervishi et al.,
1 This study was funded by the Ministry of Science and Inno-vation of Spain and European Union Regional Development funds (INIA-RTA2008-0098; INIA-RTA2009-91). The authors thank the staff of La Garcipollera and of Centro de Investigacion y Tecnolo-gia Agroalimentaria (CITA) de Aragón (La Garcipollera Research Centre, Bescós de la Garcipollera, Huesca, Spain and CITA de Ara-gon, Unidad de Tecnologia en Produccion Animal, Zaragoza, Spain, respectively) for their assistance in sample collection and analysis. Special thanks go to A. Sanz, I. Blasco, R. Ripoll, and F. Molino (CITA de Aragon, Unidad de Tecnologia en Produccion Animal, Zaragoza, Spain) for their assistance and suggestions. E. Dervishi is supported by a doctoral grant from the Agencia Española de Coop-eracion Internacional (Madrid). The present study is dedicated to the memory of Teresa Fustero.
2 Present address: Departament de Producció Animal, Universitat de Lleida, Av. Rovira Roure 191, 25198 Lleida, Spain.
3 Corresponding author: [email protected] March 14, 2011.Accepted August 16, 2011.
54
at Universitat de Lleida on January 4, 2012jas.fass.orgDownloaded from

2010). Recent studies have explored the transcriptomic adaptation in skeletal muscle of cattle in response to different nutrition variables, such as high quality vs. low quality diet (Reverter et al., 2003), or during se-vere undernutrition (Byrne et al., 2005; Lehnert et al., 2006). Data from these studies have allowed investiga-tors to identify biochemical mechanisms that can be associated with key physiological processes in animals and to define specific markers to evaluate meat quality. Furthermore, Muhlhausler et al. (2008) also suggested that sex and birth weight determine the expression of adipogenic and lipogenic genes in perirenal adipose tis-sue in young adult sheep fed the same diet.
To further address these issues, we studied the effect of forage type (grazing vs. hay pasture) fed to ewes and lamb sex on intramuscular FA profile and the ex-pression of some lipogenic [lipoprotein lipase (LPL), acetyl-coenzyme A carboxylase α (ACACA), FA syn-thase (FASN), FA binding protein 4 (FABP4), acyl-CoA diacylglycerol acyltransferase1 (DGAT1), stea-royl CoA desaturase (SCD), AMP-activated protein kinase α2 subunit (PRKAA2)], and transcription fac-tor genes (SREBF1, PPARG, PPARA, and CEBPB) in LM from suckling lambs of Churra Tensina sheep breed.
MATERIALS AND METHODS
All experimental procedures were performed in ac-cordance with the guidelines of the European Union and Spanish regulations for the use and care of animals in research.
Experimental Site
The experiment was conducted in La Garcipollera Research Station, in the Pyrenees (north-eastern Spain, 42°37′ N, 0°30′ W, 945 m above sea level; a.s.l.), during autumn 2008. The average annual rainfall is bimodally distributed with peaks in spring and autumn, with dry summers and some precipitation in the form of snow in winter. The mean temperature during September, October, November, and December was 16.2, 11.6, 5.0, and 3.0°C, and precipitation in these months was 31.8, 120.6, 94.0, and 59.2 mm, respectively.
Animals and Diets
Forty-eight multiparous single-bearing ewes from the Churra Tensina breed were used in this study (46.5 ± 0.66 kg of BW, and BCS of 3.0 ± 0.03 of 5.0 at breeding). Ewes were mated through male effect during spring (May 5 to June 15) to produce suckling lambs for the autumn-lambing period. Ewes were kept on mountain pastures (920 to 1,500 m a.s.l.) during the mating period (43 to 49% NDF, 20 to 26% CP; Joy et al., 2008). In summer (early to mid pregnancy), all the ewes grazed on high mountain pastures (1,500 to 2,200
m a.s.l.; 68 to 75% NDF, 7 to 10% CP; Casasús et al., 2002). From September to lambing (average October 19 ± 8 d) ewes were grazing low mountain pastures (18.9 ± 4.7% DM, 20.1 ± 1.7% CP, 42.3 ± 4.0% NDF, and 21.6 ± 1.9% ADF), or fed alfalfa pellets occasionally (93.3 ± 0.02% DM, 15.7 ± 3.3% CP, 35.5 ± 0.9% NDF, and 23.8 ± 1.0% ADF) when the snow was limiting pasture availability. After lambing, one-half of the ewes grazed mountain pastures (GRE; described above) and the rest were allowed ad libitum pasture hay (PH; 87.5 ± 5.0% DM, 12.0 ± 1.1% CP, 50.4 ± 2.9% NDF, and 26.8 ± 1.9% ADF). Hay was made in late spring from the same pasture paddocks as those grazed in autumn. Outdoor grazing ewes had free access to a shelter area.
The pasture was composed of 22% legumes (mainly Trifolium repens), 68% grass (the main species were Festuca arundinacea, Festuca pratensis, and Dactylis glomerata), and 10% other species (mainly Rumex ace-tosa and Ranunculus bulbosus; Joy et al., 2008). Pasture biomass was estimated by measuring the sward weekly, taking 50 points evenly distributed in the paddock with the Hill Farm Research Organization stick (Barthram, 1986), and decreased steadily from 14.4 ± 4.2 cm to 5.1 ± 0.6 cm during the study. Herbage mass and forage quality were measured weekly by clipping with an elec-tric mower all plant material to ground level in ten 1 × 0.25 m2 quadrats per paddock. In the present study, the estimated dietary ME content according to Cannas et al. (2004) was 10.2 and 9.1 MJ of ME/kg of DM for hay and pasture, respectively. Also, the alfalfa pellets sup-plied 10.2 MJ of ME/kg of DM to grazing ewes when snow limited pasture availability.
The study was carried out during 5 wk of lactation, the time required to reach the target slaughter BW. The BW of the ewe, parity, lamb BW, and sex were taken into account to balance groups (PH group: fe-males, n = 12 and males, n = 12; GRE group: females, n = 12 and males, n = 12). The study finished when lambs reached 10 to 12 kg of BW to include them with-in the suckling lamb commercial category.
Milk production was recorded weekly by the oxyto-cin technique proposed by Doney et al. (1979) using machine milking with hand finishing (4 h interval). Ewes were injected with 5 IU of oxytocin in a jugular vein before machine milking at 0800 and 1200 h (with hand finishing). Ewes were returned to their paddock between the 2 milking while lambs were confined. The milk obtained in the second milking was weighed, and yield was extrapolated to the daily period (daily pro-duction = milk obtained × 6). Two individual milk samples (50 mL) were preserved by addition of potas-sium dichromate and stored at −20°C until chemical analysis.
When lambs reached 10 to 12 kg of BW, they were weighed at the farm and then transported to the exper-imental abattoir of the Research Institute in Zaragoza, which is located 180 km away from the farm. Imme-diately after arrival, lambs were slaughtered accord-
55The forage type fed affects gene expression
at Universitat de Lleida on January 4, 2012jas.fass.orgDownloaded from

ing to the guidelines of the Council Directive 86/609/EEC (European Communities, 1986) on the protection of animals used for experimental and other scientific purposes. Before exsanguination, lambs were stunned by a lightweight captive bolt pistol. After evisceration, samples of LM (12 thoracic vertebrae) were immediate-ly placed in liquid nitrogen for gene expression studies.
Carcasses were chilled at 4°C for 24 h. Thereafter, carcasses were split and LM from the left side were removed, and sampled from 6th to 10th thoracic ver-tebrae for chemical analysis and from 10th to 12th thoracic vertebrae for FA determination of IMF. All samples were vacuum-packed and frozen (−20°C) until analysis.
Chemical Analysis
Muscle samples were minced and freeze-dried to de-termine the chemical composition. Meat was weighed before and after freeze-drying to calculate DM content. Fat content was quantified using the Ankom procedure (AOAC, 2000) with an Ankom extractor (model XT10, Ankom Technology, Madrid, Spain). The FA of IMF were extracted (Bligh and Dyer, 1959), methylated, and analyzed with a gas chromatograph (Autosystem XL Agilent Technologies 7890 Net Work GC System, Perkin Elmer, Boston, MA) equipped with a flame ion-ization detector, a Hamilton injector, and an Omega-wax 320 capillary column (30 m × 0.32 mm with a film thickness of 0.25 μm; Supelco, Bellefonte, PA) with He as the carrier gas at 30 cm/s. The temperature of the inlet detector was 260°C, and the initial temperature of the oven was 190°C for 2 min, increasing to 205°C at a rate of 5°C/min for 3 min. Fatty acids were quantified using the internal standard (C21:0) after adjusting for the response as determined by Sigma-Aldrich (Madrid, Spain) standard measures.
Milk FA composition was determined by capillary gas chromatography of the FA methyl esters (FAME). These FAME were prepared by base-catalyzed metha-nolysis of the glycerides with KOH according to the UNE-EN ISO 5509:2000 methods (BOE, 2001). Fatty acid methyl esters were separated using a flame-ioniza-tion detector (gas chromatograph HP-6890), equipped with a capillary column (HP-88 100 m × 0.25 mm i.d. and 0.20-μm film thickness, Agilent Technologies, Waldbronn, Germany). The carrier gas was He, and the flow rate was 2 mL/min. The temperatures of the inlet and detector were maintained at 250 and 300°C, respec-tively. The temperature program was as follows: the initial temperature was held at 100°C after injection, then programmed to increase at 1.5°C/min to 170°C (held there for 15 min), then programmed to increase at 0.5°C/min to 180°C (held there for 2 min), and fi-nally to increase at 10°C/min to 215°C (held there for 20 min). Injection volume was 1.0 μL. The FAME iden-tification was based on retention times compared with those of the standard FAME mixture (Sigma-Aldrich).
The individual FA contents were expressed as weight percentages (g/100 g of total FA). Proportions of PUFA, MUFA, and SFA as well as n-6 and n-3 PUFA/SFA and n-6/n-3 ratios were obtained from individual FA percentages.
RNA Extraction and cDNA Synthesis
Total RNA was prepared from approximately 500 mg of LM using Tri Reagent (Sigma Life Science, Madrid, Spain), according to the manufacturer’s instructions. The concentration and quality of RNA were determined using nanophotometer (Implen, Madrid, Spain). To ex-clude possible amplification of contaminating genomic DNA, samples were treated with deoxyribonuclease. Single-stranded cDNA was synthesized from 1 μg of RNA using the SuperScript III Reverse Transcriptase kit (Invitrogen, Carlsbad, CA) following the manufac-turer’s instructions.
Real-Time PCR Analysis
Gene expression abundance was determined by real-time PCR in an ABI Prism 7500 platform (Applied Biosystems, Madrid, Spain). Primers for real-time PCR were designed using the Primer3 software (http:// frodo.wi.mit.edu/primer3/). The sequences of primers and the real-time reverse-transcription (RT) PCR con-ditions are shown in Table 1.
Before performing the real-time RT-PCR reactions, a conventional PCR was carried out for all genes to test the primers and verify the amplified products. The PCR products were sequenced to confirm gene identity using an ABI Prism3700 (Applied Biosystems) with standard protocols. Homology searches were performed with BLAST (National Center for Biotechnology Infor-mation: http://www.ncbi.nlm.nih.gov/BLAST/), con-firming the identity of the amplified fragments. When possible, the primers for real-time RT-PCR were de-signed across exon-exon junctions to prevent the am-plification of genomic DNA in the PCR reaction. The PCR reaction was carried out in a 10-μL PCR total reaction mixture containing SYBR Green PCR Master Mix (Applied Biosystems). Each reaction was run in triplicate, and the average was used to calculate the rel-ative amount of target genes. To normalize the results of the target genes, 8 candidate housekeeping genes were tested. In this study, analyzed housekeeping genes in-cluded ovine tyrosine 3-monooxigenase (YWHAZ), ri-bosomal protein L19 (RPL19), glyceraldehyde-3-phos-phate dehydrogenase (GAPDH), glucose-6-phosphate dehydrogenase (G6PDH), β actin (ACTB), ubiquitin C (UBC), β-2-microglobulin (B2M), and succinate dehydrogenase (SDHA; Table 1). Gene stability values were estimated using the NormFinder (Molecular Diag-nostic Laboratory, Aarhus, Denmark) software, which indentifies the optimal normalization gene among a set of candidates (Andersen et al., 2004). Standard curves for genes were generated with the aim of calculating
56 Dervishi et al.
at Universitat de Lleida on January 4, 2012jas.fass.orgDownloaded from

the amplification efficiency. The efficiency of PCR am-plification for each gene was calculated using the stan-dard curve method (E = 10−1/slope − 1). The standard curves for each gene were generated by 5-fold serial dilution of pooled cDNA. The amplification conditions included an initial step of 10 min at 95°C, followed by 40 cycles of 95°C for 15 s and 59 or 60°C for 30 s. The specificity of the amplification products and the lack of primer dimers were determined by performing melting curves. To quantify relative gene expression, the standard curve method was used according to the recommendation of Larionov et al. (2005). Normalized real-time PCR data were transformed into fold-change relative to the control group; therefore, data in the con-
trol group were transformed to obtain a perfect mean of 1. The PCR-normalized data are presented as relative to n-fold change.
Statistical Analyses
Statistical analyses were carried out using the SPSS statistical software package (SPSS Inc., Chicago, IL). The chemical composition and intramuscular FA con-tent of LM were assessed using the GLM procedure, where the diet of the ewes, lamb sex, and their interac-tions were included as fixed effects.
The relative differences in gene expression among the different groups were defined as the relative amounts
Table 1. Genes, forward (F) and reverse (R) primers, amplicon length, and GenBank accession numbers for ovine sequences1
Target gene2 Primers: F and R
Amplicon, bp Acc. number
PCR conditions
AT Nm R2 Slope
LPL F-5′aatgaagagatgaacggaacg-3′ 119 NM_001009394 59 900 0.994 −3.37R-5′gcactttccaaccaggatgt-3′ 900
ACACA F-5′tgaaggatgtggatgatgg-3′ 110 NM_001009256 59 300 0.991 −3.29R-5′cgcgttgttgacttttctga-3′ 300
FASN F-5′gtgtggtacagcccctcaag-3′ 110 GQ150557 59 300 0.989 −3.31R-5′acgcacctgaatgaccactt-3′ 300
FABP4 F-5′aagaagtgggtgtgggcttt-3′ 91 EU301804 60 900 0.998 −3.64R-5′atgttgaccacatccccatt-3′ 900
SCD F-5′cccagctgtcagagaaaagg-3′ 115 AJ001048 59 900 0.996 −3.35R-5′gatgaagcacaacagcagga-3′ 900
DGAT1 F-5′acgtcccggtaggaaaacag-3′ 100 EU178818 59 900 0.993 −3.38R-5′cgtggccttcctcctagagt-3′ 900
PRKAA2 F-5′accctcccatttgatgatga-3′ 97 NM_001112816 60 900 0.993 −3.56R-5′tggcaacagaacgattgaga-3′ 900
SREBF1 F-5′ctgctatgcaggcagcac-3′ 99 GU206528 59 900 0.980 −3.32R-5′ggttgatgggcagcttgt-3′ 900
PPARG F-5′cttgctgtggggatgtctc-3′ 121 AY137204 60 900 0.982 −3.35R-5′ggtcagcagactctgggttc-3′ 900
PPARA F-5′tgccaagatctgaaaaagca-3′ 101 AY369138 60 300 0.986 −3.38R-5′cctcttggccagagacttga-3 300
CEBPB F-5′gacaagcacagcgacgagt-3′ 102 AY371068 59 300 0.998 −3.38F-5′gtgctgcgtctccaggtt-3 300
ACTB F-5′ggacctgacggactacctcatg-3′ 136 U39357 60 100 0.989 −3.32R-5′ggccatctcctgctcgaagt-3′ 100
SDHA F-5′catccactacatgacggagca-3′ 90 AY970969 60 300 0.990 −3.36R-5′atcttgccatcttcagttctgcta-3′ 300
G6PDH F-5′tgacctatggcaaccgatacaa-3′ 76 DQ377364 60 900 0.994 −3.33R-5′ccgcaaaagacatccaggat-3′ 900
GAPDH F-5′atgcctcctgcaccacca-3′ 76 HM043737 60 200 0.991 −3.37R-5′agtccctccacgatgccaa-3′ 200
UBC F-5′cgtcttaggggtggctgtta-3′ 90 NM_001009202 59 100 0.981 −3.38R-5′aaattggggtaaatggctaga-3′ 100
B2M F-5′ggtgctgcttagaggtctcg-3′ 109 NM_001009284 59 200 0.987 −3.35R-5′acgctgagttcactcccaac-3′ 200
RPL19 F-5′caactcccgccagcagat-3′ 76 AY158223 60 200 0.994 −3.35R-5′ccgggaatggacatgcaca-3′ 200
YWHAZ F-5′tgtaggagcccgtaggtcatct-3′ 102 AY970970 59 200 0.996 −3.38R-5′ttctctctgtattctcgagccatct-3 200
1Real-time reverse-transcription PCR conditions: annealing temperature (AT), primer concentrations (Nm), correlation coefficient (R2), and slope of the standard curve.
2LPL = lipoprotein lipase; ACACA = acetyl-coenzyme A carboxylase α; FASN = fatty acid synthase; FABP4 = fatty acid binding protein 4; SCD = stearoyl CoA desaturase; DGAT1 = acyl-CoA diacylglycerol acyltransferase 1; PRKAA2 = AMP-activated protein kinase α2 subunit; SREBF1 = sterol regulatory element binding protein 1; ACTB = β-actin; SDHA = succinate dehydrogenase; G6PDH = glucose-6-phosphate de-hydrogenase; GAPDH = glyceraldehyde-3-phosphate dehydrogenase; UBC = ubiquitin C; B2M = β-2-microglobulin; RPL19 = ribosomal protein L19; YWHAZ = tyrosine 3-monooxygenase.
57The forage type fed affects gene expression
at Universitat de Lleida on January 4, 2012jas.fass.orgDownloaded from

after normalization. Relative differences between the groups were calculated and defined as the relative in-crease, setting the control means at 1. To evaluate the effect of lamb sex on gene expression, the analyses were performed separately for each treatment group. In this analysis, sex was included as a fixed effect, and birth weight, slaughter weight, and IMF were included as co-variates. To evaluate the effect of the diet on the ewe on gene expression, the analyses were performed sepa-rately for each sex, thus avoiding significant changes in gene expression due to animal sex. In this analysis, ewe diet was included as a fixed effect, and birth weight, slaughter weight, and IMF were included as covariates. The relationship between gene expression and sex, IMF, FA indicators, birth weight, and slaughter weight was determined using stepwise linear regression analysis.
The level of significance was set at P ≤ 0.05, whereas P-values >0.05 but ≤0.10 were considered trends.
RESULTS
Chemical Meat Composition and FA Profile
Forage type fed to ewes affected several milk FA cat-egories (Table 2). Significant differences were found in PUFA and CLA (C18:2 cis-9, trans-11; P < 0.001) and PUFA n-3 (P > 0.01) contents and in PUFA/SFA (P < 0.001) and PUFA n-6/n-3 (P < 0.05) ratios. Milk from ewes grazing pastures showed greater content of PUFA, CLA, PUFA n-3 than those fed hay. In contrast, the ratio of PUFA n-6/n-3 was less in the GRE group (P > 0.05).
Diet fed to ewes did not affect BW at birth or slaugh-ter BW (P > 0.05; Figure 1). In the GRE group, the average birth weight in males was 3.70 ± 0.14 kg, and in females was 3.55 ± 0.16 kg. In the PH group, birth weight was 3.87 ± 0.14 kg and 3.65 ± 0.16 kg for males and females, respectively. Slaughter weight, fixed be-tween 10 to 12 kg of BW, was 10.7 ± 0.35 and 10.5 ± 0.39 kg in the GRE group and 10.8 ± 0.35 and 10.5 ± 0.39 kg in the PH group for males and females, respec-tively. Regarding chemical composition of LM, no effect of forage type fed to dams or lamb sex on crude fat content was observed (P > 0.05; Figure 1). In the GRE group, fat content in males was 2.05 ± 0.21%, whereas in females, it was 2.06 ± 0.23%. In the PH group, fat content was 2.17 ± 0.21% and 2.05 ± 0.23% for male and females, respectively.
Table 2. Main fatty acid (FA) categories content (%) in milk of Churra Tensina ewes fed low mountain hay (PH) or grazing low mountain vegetation (GRE)
FA category PH GRE P-value
SFA 56.37 56.17 0.77MUFA 31.61 29.93 0.06PUFA 5.70b 6.19a <0.001CLA cis-9, trans-11 1.29b 1.63a <0.001PUFA/SFA 0.10b 0.11a <0.001PUFA n-6 2.53 2.48 0.47PUFA n-3 1.58b 1.74a 0.01n-6/n-3 1.64a 1.46b 0.02
a,bWithin a row, means without a common superscript differ (P < 0.05).
Figure 1. a) Birth weight in kilograms, b) slaughter weight in kilograms, and c) total intramuscular fat (IMF) in percentage in LM of male and female Churra Tensina suckling lambs, according to forage type fed to ewes. PH group: lambs whose mothers received low mountain hay; GRE group: lambs whose mothers grazed low mountain vegetation.
58 Dervishi et al.
at Universitat de Lleida on January 4, 2012jas.fass.orgDownloaded from
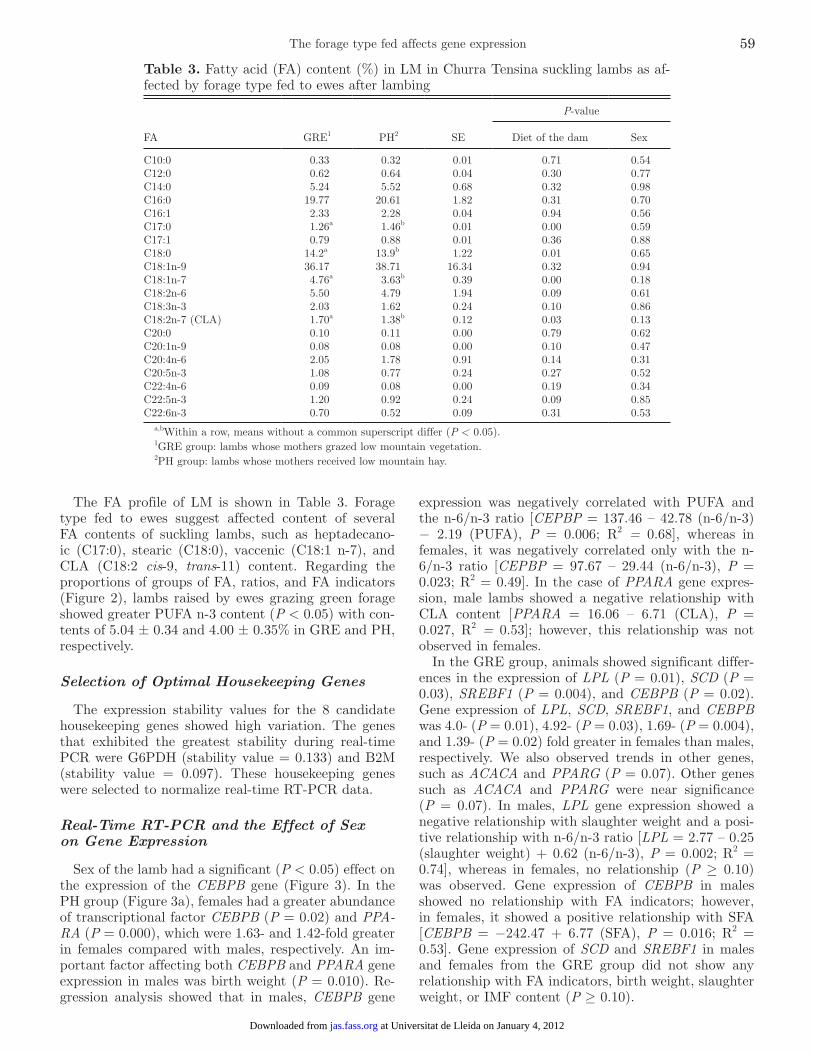
The FA profile of LM is shown in Table 3. Forage type fed to ewes suggest affected content of several FA contents of suckling lambs, such as heptadecano-ic (C17:0), stearic (C18:0), vaccenic (C18:1 n-7), and CLA (C18:2 cis-9, trans-11) content. Regarding the proportions of groups of FA, ratios, and FA indicators (Figure 2), lambs raised by ewes grazing green forage showed greater PUFA n-3 content (P < 0.05) with con-tents of 5.04 ± 0.34 and 4.00 ± 0.35% in GRE and PH, respectively.
Selection of Optimal Housekeeping Genes
The expression stability values for the 8 candidate housekeeping genes showed high variation. The genes that exhibited the greatest stability during real-time PCR were G6PDH (stability value = 0.133) and B2M (stability value = 0.097). These housekeeping genes were selected to normalize real-time RT-PCR data.
Real-Time RT-PCR and the Effect of Sex on Gene Expression
Sex of the lamb had a significant (P < 0.05) effect on the expression of the CEBPB gene (Figure 3). In the PH group (Figure 3a), females had a greater abundance of transcriptional factor CEBPB (P = 0.02) and PPA-RA (P = 0.000), which were 1.63- and 1.42-fold greater in females compared with males, respectively. An im-portant factor affecting both CEBPB and PPARA gene expression in males was birth weight (P = 0.010). Re-gression analysis showed that in males, CEBPB gene
expression was negatively correlated with PUFA and the n-6/n-3 ratio [CEPBP = 137.46 – 42.78 (n-6/n-3) − 2.19 (PUFA), P = 0.006; R2 = 0.68], whereas in females, it was negatively correlated only with the n-6/n-3 ratio [CEPBP = 97.67 – 29.44 (n-6/n-3), P = 0.023; R2 = 0.49]. In the case of PPARA gene expres-sion, male lambs showed a negative relationship with CLA content [PPARA = 16.06 – 6.71 (CLA), P = 0.027, R2 = 0.53]; however, this relationship was not observed in females.
In the GRE group, animals showed significant differ-ences in the expression of LPL (P = 0.01), SCD (P = 0.03), SREBF1 (P = 0.004), and CEBPB (P = 0.02). Gene expression of LPL, SCD, SREBF1, and CEBPB was 4.0- (P = 0.01), 4.92- (P = 0.03), 1.69- (P = 0.004), and 1.39- (P = 0.02) fold greater in females than males, respectively. We also observed trends in other genes, such as ACACA and PPARG (P = 0.07). Other genes such as ACACA and PPARG were near significance (P = 0.07). In males, LPL gene expression showed a negative relationship with slaughter weight and a posi-tive relationship with n-6/n-3 ratio [LPL = 2.77 – 0.25 (slaughter weight) + 0.62 (n-6/n-3), P = 0.002; R2 = 0.74], whereas in females, no relationship (P ≥ 0.10) was observed. Gene expression of CEBPB in males showed no relationship with FA indicators; however, in females, it showed a positive relationship with SFA [CEBPB = −242.47 + 6.77 (SFA), P = 0.016; R2 = 0.53]. Gene expression of SCD and SREBF1 in males and females from the GRE group did not show any relationship with FA indicators, birth weight, slaughter weight, or IMF content (P ≥ 0.10).
Table 3. Fatty acid (FA) content (%) in LM in Churra Tensina suckling lambs as af-fected by forage type fed to ewes after lambing
FA GRE1 PH2 SE
P-value
Diet of the dam Sex
C10:0 0.33 0.32 0.01 0.71 0.54C12:0 0.62 0.64 0.04 0.30 0.77C14:0 5.24 5.52 0.68 0.32 0.98C16:0 19.77 20.61 1.82 0.31 0.70C16:1 2.33 2.28 0.04 0.94 0.56C17:0 1.26a 1.46b 0.01 0.00 0.59C17:1 0.79 0.88 0.01 0.36 0.88C18:0 14.2a 13.9b 1.22 0.01 0.65C18:1n-9 36.17 38.71 16.34 0.32 0.94C18:1n-7 4.76a 3.63b 0.39 0.00 0.18C18:2n-6 5.50 4.79 1.94 0.09 0.61C18:3n-3 2.03 1.62 0.24 0.10 0.86C18:2n-7 (CLA) 1.70a 1.38b 0.12 0.03 0.13C20:0 0.10 0.11 0.00 0.79 0.62C20:1n-9 0.08 0.08 0.00 0.10 0.47C20:4n-6 2.05 1.78 0.91 0.14 0.31C20:5n-3 1.08 0.77 0.24 0.27 0.52C22:4n-6 0.09 0.08 0.00 0.19 0.34C22:5n-3 1.20 0.92 0.24 0.09 0.85C22:6n-3 0.70 0.52 0.09 0.31 0.53
a,bWithin a row, means without a common superscript differ (P < 0.05).1GRE group: lambs whose mothers grazed low mountain vegetation.2PH group: lambs whose mothers received low mountain hay.
59The forage type fed affects gene expression
at Universitat de Lleida on January 4, 2012jas.fass.orgDownloaded from

The Effect of the Diet of the Ewe on Gene Expression in Female Lambs
With the aim of studying the effect of forage type fed to ewes on gene expression in LM in suckling lambs, sta-tistical analyses were carried out independently in male and female lambs, thus avoiding significant changes in gene expression due to animal sex. The results showed that gene expression of SREBF1 (P = 0.03) in female lambs was significantly affected by the type of forage fed to their dams (Figure 4). Females in the GRE group had 1.47-fold greater expression of SREBF1 than ewe lambs in the PH group. Gene expression of PPARG and PPARA approached a significant (P = 0.08 and P = 0.06, respectively) difference due to feed treatment. Re-gression analysis did not show a relationship between SREBF1 and FA indicators (P ≥ 0.10).
The Effect of the Diet of the Ewe on Gene Expression in Male Lambs
In males, significant differences were found in SCD gene expression (P = 0.04), and CEBPB was at the limit of significance (P = 0.05; Figure 5). Relative gene expression of SCD was 0.78-fold less (P = 0.04) in males from the GRE group compared with the PH group males. However, CEBPB gene expression was 1.31-fold greater (P = 0.05) in the GRE group com-pared with the PH group. In the PH group, SCD gene expression showed a positive relationship with the PUFA n-6/n-3 ratio and with SFA [SCD = −7.25+
1.46 (n-6/n-3) + 1.46 (SFA), P = 0.008, R2 = 0.65]. When males were grazing, SCD gene expression showed a positive relationship with PPARG gene expression [SCD = −0.048 + 0.2 (PPARG), P = 0.002, R2 = 0.63] but not with other variables. Gene expression of CEB-PB had a negative relationship with n-6/n-3 in the PH group [CEBPB = 80.80 − 28.65 (n-6/n-3), P = 0.011, R2 = 0.49]. However, in the GRE group, CEBPB gene expression showed no relationship with the FA indica-tor, birth weight, or slaughter weight (P ≥ 0.10), but it showed a positive relationship with PPARA gene ex-pression [CEBPB = 9.86 + 2.29 (PPARA), P = 0.004, R2 = 0.57].
DISCUSSION
In the present study, the effect of the forage type fed to ewes and lamb sex on FA composition and gene expression in LM from Churra Tensina suckling lambs was investigated. No statistical differences were ob-served in total IMF infiltration between males and fe-males in LM, as already described in 4- to 5-wk-old animals (Miguélez et al., 2008). In 22-wk-old lambs, Muhlhausler et al. (2008) found that the percentage of body fat tended to be greater in females than in males. Female sheep showed a greater tendency to partition nutrients into fat, rather than lean tissue, in early postnatal life. In contrast to the above mention study, we analyzed very young animals, 5 wk old, and therefore we did not expect to find any differences in total IMF infiltration. Furthermore, the diets of the
Figure 2. Fatty acid (FA) indicators in LM of suckling lambs: a) FA groups according to the degree of saturation, b) the healthy point of view, and c) ratios related to human health. Different letters indicate a significant difference with at least P < 0.05. The ratio of SFA/PUFA was calculated as (C18:2 + C18:3 + C20:4 + C20:5 + C22:4 + C22:5 + C22:6)/(C10:0 + C12:0 + C14:0 + C16:0 + C18:0 + C20:0). The ratio of n-6/n-3 was calculated as (C18:2n-6 + C20:4n-6 + C22:4n-6)/(C18:3n-3 + C20:5n-3 + C22:5n-3 + C22:6n-3). PH group: lambs whose mothers received low mountain hay; GRE group: lambs whose mothers grazed low mountain vegetation.
60 Dervishi et al.
at Universitat de Lleida on January 4, 2012jas.fass.orgDownloaded from

ewes were similar in ME content, regardless of the type of forage fed to ewes. Although no difference in total IMF was observed, we found significant differences in some FA content. Stearic acid, vaccenic acid, CLA, and PUFA n-3 were greater in the GRE group compared with the PH group. Fatty acid content from suckling lambs could be modulated by dietary composition of their dams because milk FA composition also showed greater contents of CLA, PUFA, and PUFA n-3 and decreased PUFA n-6/n-3 ratio in grazing ewes. Further-more, in our study, significant differences in milk have been found in CLA (C18:2 cis-9, trans-11), PUFA, and PUFA n-3 content. Grazing pastures promotes CLA formation, and various studies reported the same re-
sults in the meat of ruminants (French et al., 2000; Aurousseau et al., 2004; Daniel et al., 2004; Kay et al., 2004; Dannenberger et al., 2005; Aurousseau et al., 2007; Dervishi et al., 2010) and their milk (Kelly et al., 1998). The effect of diet fed to ewes on intramuscular FA composition of suckling lambs has been studied by Scerra et al. (2007). They found that IMF from the pasture-fed lambs was richer in n-3 PUFA compared with the stall-fed lambs, and consequently, the n-6/n-3 ratio was less in the IMF from pasture-fed lambs. In the present study, we found that lambs whose dams grazed have greater content of PUFA n-3 and CLA compared with those whose dams were hay-fed. We did not ex-pect differences in FA profiles because the diets were
Figure 3. Effect of lamb sex on gene expression: a) gene expression in male and female lambs whose mothers received the low mountain hay (PH) treatment, and b) gene expression in male and female lambs whose mothers grazed the low mountain vegetation (GRE) treatment. Differ-ent letters (a,b) indicate a significant difference with at least P < 0.05. LPL = lipoprotein lipase; ACACA = acetyl-coenzyme A carboxylase α; FASN = fatty acid synthase; FABP4 = fatty acid binding protein 4; DGAT1 = acyl-CoA diacylglycerol acyltransferase 1; SCD = stearoyl CoA desaturase; SREBF1 = sterol regulatory element binding protein 1.
61The forage type fed affects gene expression
at Universitat de Lleida on January 4, 2012jas.fass.orgDownloaded from

not extremely different, such as in cases of starving vs. refeeding or grazing vs. concentrate. In the present study, both diets (GRE and PH group) were forages, which only differ in the way of conservation (directly grazed vs. dried). Moreover, suckling lambs from graz-ing ewes showed greater content of CLA and PUFA n-3, which is favorable in regard to current human dietary guidelines. From a human health perspective, a greater n-6/n-3 ratio has been found to promote the pathogen-esis of many chronic illnesses, including cardiovascular disease (Simopoulos, 2008). Similarly, CLA are a group
of positional and geometric isomers of octadecadienoic acids with conjugated double bonds and are receiving increasing attention because of their possible beneficial effects on human health. For example, they reduce the incidence of atherosclerosis, diabetes, and cancer in ani-mals (Park et al., 1997; Pariza et al., 1999; Kritchevsky et al., 2000).
The expression of genes related with fat metabolism was analyzed in LM of Churra Tensina male and fe-male suckling lambs. We tested 8 housekeeping genes, and the 2 most stable genes, G6PDH and B2M, were
Figure 4. Gene expression in LM of female suckling lambs. Different letters (a,b) indicate a significant difference with at least P < 0.05. PH group: female lambs whose mothers received low mountain hay; GRE group: female lambs whose mothers grazed low mountain vegetation. LPL = lipoprotein lipase; ACACA = acetyl-coenzyme A carboxylase α; FASN = fatty acid synthase; FABP4 = fatty acid binding protein 4; SCD = stearoyl CoA desaturase; DGAT1 = acyl-CoA diacylglycerol acyltransferase 1; SREBF1 = sterol regulatory element binding protein 1.
Figure 5. Gene expression in LM from male suckling lambs. Different letters (a,b) indicate a significant difference with at least P < 0.05. PH group: female lambs whose mothers received low mountain hay; GRE group: female lambs whose mothers grazed low mountain vegetation. LPL = lipoprotein lipase; ACACA = acetyl-coenzyme A carboxylase α; FASN = fatty acid synthase; FABP4 = fatty acid binding protein 4; DGAT1 = acyl-CoA diacylglycerol acyltransferase 1; SCD = stearoyl CoA desaturase; SREBF1 = sterol regulatory element binding protein 1.
62 Dervishi et al.
at Universitat de Lleida on January 4, 2012jas.fass.orgDownloaded from

selected to normalize the real-time RT-PCR data. The type of treatment and the type of cell or muscle type are sources of variance in housekeeping genes (Dheda et al., 2004; de Jonge et al., 2007). Therefore, house-keeping genes are highly specific for a particular ex-perimental model and more than 1 housekeeping gene is recommended to detect small expression differences accurately (Vandesompele et al., 2002). Recent studies explored the transcriptomic adaptation in skeletal mus-cle of cattle in response to a plane of nutrition such as high quality diet vs. low quality (Reverter et al., 2003), or during severe undernutrition (Byrne et al., 2005; Lehnert et al., 2006). Moreover, Wang et al. (2009) de-termined gene expression patterns during IMF devel-opment in 2 different crossbreed cattle using a micro-array approach. They reported changes in expression of adipose differentiation-associated genes, CEBPB and PPARG, and in genes involved with FA synthesis such as FABP4 and FASN. In the present study, GRE female lambs showed greater SREBF1 and tended to express more PPARG and PPARA compared with PH female lambs. The SREBF1 has been established as a lipid synthetic transcription factor for FA biosynthesis and cholesterol homeostasis (Felder et al., 2005) and has been associated with the concentration of stearic acid (C18:0), linoleic acid (C18:2), and PUFA in the Hanwoo cattle breed (Bhuiyan et al., 2009). In the pres-ent study, SREBF1 gene expression did not show a re-lationship with FA indicators in both the GRE and the PH group females. In the PH group males, SCD gene expression was greater compared with GRE males. The same tendency was found in female lambs; however, there were no significant differences. We also found that SCD gene expression of PH group males showed a positive relationship with n-6/n-3 ratio and SFA but not the GRE group, suggesting that FA indicators are important for the regulation of SCD gene expression in PH group suckling lambs but not in GRE group. Nota-bly, in GRE males, SCD gene expression showed a posi-tive relationship only with PPARG gene expression but not with other variables. The balance between n-6 and n-3 PUFA in the diet is important for the regulation of SCD through SREBP-1c in muscle of beef cattle, and n-3 PUFA enriched diet inhibits SREBP-1c in muscle tissue (Waters et al., 2009; Herdmann et al., 2010). These results are in agreement with previous reports, in which expression of the SCD gene is downregulated by PUFA (especially the n-6 and n-3 families), CLA, cho-lesterol, vitamin A, hormonal changes, and peroxisomal proliferators (Ntambi, 1995, 1999; Choi et al., 2000; Ntambi et al., 2002; Waters et al., 2009). In this sense, SCD gene expression showed a positive relationship with PPARG gene expression. Small-interference RNA-mediated knock-down of SCD1 in human adipocytes impaired adipogenesis and decreased PPARγ protein abundance (Christianson et al., 2008; Yao-Borengasser et al., 2008). Bovine studies reported that the expres-sion of the SCD gene is regulated by the transcrip-
tion factor PPARγ (Graugnard et al., 2009; Waters et al., 2009). Near significance was found for the CEBPB gene, which is involved in the preadipocyte differentia-tion process, and it is required for subsequent induction of PPARG expression in the preadipocyte differentia-tion process (Wu et al., 1995; Clarke et al., 1999). This finding may indicate that CEBPB expression was not regulated in response to the plane of nutrition.
In the PH group, CEBPB and PPARA expression was significantly affected by sex. Meanwhile, a signifi-cant effect on the expression of LPL, SCD, SREBF1, and CEBPB genes was observed in GRE lambs. Female lambs had greater gene expression compared with their male counterparts. Lamb sex affected CEBPB gene expression in the GRE and the PH group. Females of both groups showed greater CEBPB gene expres-sion compared with males. Muhlhausler et al. (2008) reported that sex and birth weight have an effect on the expression on adipogenic and lipogenic genes such as CEBPA, PPARG, LPL, and G3PDH in perirenal adipose tissue in young adult sheep fed the same diet. A sex-specific pattern of leptin mRNA expression was reported by the same authors in adipose tissue. The CEBPB gene, which is involved in the preadipocyte dif-ferentiation process, indicates the predisposition of the animal to develop fat (Wang et al., 2009). Muhlhausler et al. (2008) concluded that female sheep have a greater tendency to partition nutrients into fat, rather than lean tissue, in early postnatal life. Our results indicate that animal sex may be exerting its effect on transcriptional factors such as PPARA, CEBPB, SREBF1, and other genes that are considered to be adipogenic markers. It has been previously reported that the LPL, ACACA, and SCD genes are considered to be potential PPAR and CEBP target genes. The PPARG has a central role in controlling the transcription of genes that are known to regulate adipocyte differentiation and lipid filling in longissimus lumborum in cattle (Graugnard et al., 2009). These authors demonstrated that intramuscular adipocyte differentiation during the early growth phase involves greater expression of mature adipocyte gene markers such as PPARG, THRSP, and SCD. Moreover, terminal differentiation of adipocytes requires the up-regulation of SCD, FABP4, FASN, and ACACA genes (Graugnard et al., 2009; Hausman et al., 2009).
We hypothesize that these changes in gene expression between males and females fed the same diet during the early growth period may be due to hormonal factors. Sex steroid hormones, such as estrogens or androgens, are involved in the metabolism, accumulation, and dis-tribution of adipose tissues by binding to their recep-tors, and then the steroid-receptor complex regulates the transcription of given genes. Estrogen-mediated control of gene expression is a complex process that includes ligand–receptor interactions, receptor–DNA interactions and receptor–cofactor interactions (Gao and Dahlman-Wright, 2010). The LEP, LPL, ACC, SREBP-cleavage activating protein (SCAP), SCD,
63The forage type fed affects gene expression
at Universitat de Lleida on January 4, 2012jas.fass.orgDownloaded from

and PPARγ are key proteins in adipose tissue that are regulated by transcriptional control with sex ste-roid hormones (Machinal et al., 1999; Heemers et al., 2004; Mayes and Watson, 2004; Swinnen et al., 2004; Lundholm et al., 2008). Androgens and estrogens have a tremendous impact on the lipogenesis of adipose tis-sue (Lundholm et al., 2008) because androgens act as anti-adipogenic hormones and estrogens behave as pro-adipogenic hormones. Androgens, such as testosterone, regulate lineage determination in mesenchymal pluripo-tent cells by promoting their commitment to the myo-genic lineage and inhibiting their differentiation into adipogenic lineage through the androgen receptor-me-diated pathway (Singh et al., 2003). The opposite effect of androgens on adipogenesis in rat preadipocytes may involve the IGF-1 receptor and PPARG2 (Dieudonne et al., 2000). Taken together, the data suggest that SCD and SREBF1 gene expression in young suckling lambs is under the control of both sexes and the type of forag-ing. Our results allow us to speculate that females in both groups (PH and GRE) have the predisposition to develop fat by upregulating the expression of key genes of transcriptional factors and other genes that are con-sidered to be adipogenic markers.
To our knowledge, this is the first study that inves-tigates whether forage type fed to ewes and lamb sex affect intramuscular FA profile and the expression of genes related to fat metabolism.
Conclusions
Forage type fed to lactating ewes had a significant effect on the FA profile of milk and IMF of their lambs. Animals fed GRE were characterized by greater con-tent of vaccenic C18:1n-7, CLA, and n-3 FA compared with PH animals. Ewes grazing pasture promoted the formation and deposition of n-3 FA in LM of their suck-ling lambs. Lamb sex modulated expression of CEB-PB, PPARA, SREBF1, and SCD, with females having greater gene expression compared with males, inde-pendent from the forage type fed to ewes. The forage type affected milk composition and gene expression of SREBF1 and SCD in female and male suckling lambs, respectively.
The data suggest that SREBF1, SCD, and most like-ly CEBPB gene expression in young suckling lambs is modulated by both lamb sex and forage type. Fatty acid indicators PUFA, n-6/n-3, CLA, and SFA affect LPL, SCD, PPARA, and CEBPB gene expression ac-cording to animal sex and forage type fed to the ewe. This study suggests that the dietary manipulation of ewes affects FA composition in young female and male suckling lambs, and it is mediated through the regula-tion of lipogenic enzyme expression.
LITERATURE CITED
Alvarez-Rodriguez, J., A. Sanz, R. Delfa, R. Revilla, and M. Joy. 2007. Performance and grazing behaviour of Churra Tensina
sheep stocked under different management systems during lac-tation on Spanish mountain pastures. Livest. Sci. 107:152–161.
Andersen, C. L., J. L. Jensen, and T. F. Orntoft. 2004. Normaliza-tion of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data set. Cancer Res. 64:5245–5250.
AOAC. 2000. Official Methods of Analysis. Assoc. Off. Anal. Chem., Arlington, VA.
Aurousseau, B., D. Bauchart, E. Calichon, D. Micol, and A. Priolo. 2004. Effect of grass or concentrate feeding systems and rate of growth on triglyceride and phospholipid and their fatty acids in the M. longissimus thoracis of lambs. Meat Sci. 66:531–541.
Aurousseau, B., D. Bauchart, X. Faure, A. L. Galot, S. Prache, and D. Micol. 2007. Indoor fattening of lambs raised on pas-ture. Part 1: Influence of stall finishing duration on lipid classes and fatty acids in the longissimus thoracis muscle. Meat Sci. 72:241–252.
Barthram, G. T. 1986. Experimental techniques: The HFRO sward stick. Biennial report. 1984–1985. Hill Farming Research Or-ganisation, Edinburgh, UK.
Bernard, L., C. Leroux, and Y. Chilliard. 2008. Expression and nu-tritional regulation of lipogenic genes in the ruminant lactat-ing mammary gland. Chemistry and materials science bioactive components of milk. Adv. Exp. Med. Biol. 606:67–108.
Bhuiyan, M. S. A., S. L. Yu, J. T. Jeon, D. Yoon, Y. M. Cho, and E. W. Park. 2009. DNA polymorphisms in SREBF1 and FASN genes affect fatty acid composition in Korean cattle (Hanwoo). Asian-australas. J. Anim. Sci. 22:765–773.
Bligh, E. G., and W. J. Dyer. 1959. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 37:911–917.
BOE. 2001. Aceites y grasas de origen animal y vegetal. Preparación de ésteres metílicos de ácidos grasos. In Resolución de 7 de mayo de 2001, de la Dirección General de Política Tecnológica, por la que se publica la relación de normas UNE aprobadas por AENOR durante el mes de febrero de 2001, no. 136, de 7 de junio, 20223–20227.
Byrne, K. A., Y. H. Wang, S. A. Lehnert, G. S. Harper, S. M. Mc-William, H. L. Bruce, and A. Reverter. 2005. Gene expression profiling of muscle tissue in Brahman steers during nutritional restriction. J. Anim. Sci. 83:1–12.
Cannas, A., L. O. Tedeschi, D. G. Fox, A. N. Pell, and P. J. Van Soest. 2004. A mechanistic model for predicting the nutrient requirements and feed biological values for sheep. J. Anim. Sci. 82:149–169.
Casasús, I., A. Sanz, D. Villalba, R. Ferrer, and R. Revilla. 2002. Factors affecting animal performance during the grazing sea-son in a mountain cattle production system. J. Anim. Sci. 80:1638–1651.
Choi, Y., Y. Cheul Kim, Y. B. Han, Y. Park, M. W. Pariza, and J. M. Ntambi. 2000. The trans-10, cis-12 isomer of conjugated linoleic acid downregulates stearoyl-CoA desaturase 1 gene ex-pression in 3T3–L1 adipocytes. J. Nutr. 130:1920–1924.
Christianson, J. L., S. Nicoloro, J. Straubhaar, and M. P. Czech. 2008. Stearoyl CoA desaturase 2 is required for PPARγ expres-sion and adipogenesis in cultured 3T3-L1 cells. J. Biol. Chem. 283:2906–2916.
Clarke, S. D., P. Thuillier, R. A. Baillie, and X. Sha. 1999. Peroxi-some proliferator-activated receptors: A family of lipid-activat-ed transcription factors. Am. J. Clin. Nutr. 70:566–571.
Daniel, Z. C., R. J. Wynn, A. M. Salter, and P. J. Buttery. 2004. Differing effects of forage and concentrate diets on the oleic acid and conjugated linoleic acid content of sheep tissues: The role of stearoyl-CoA desaturase. J. Anim. Sci. 82:747–758.
Dannenberger, D., K. Nuernberg, G. Nuernberg, N. Scollan, H. Steinhart, and K. Ender. 2005. Effect of pasture vs. concentrate diet on CLA isomer distribution in different tissue lipids of beef cattle. Lipids 40:589–598.
de Jonge, H. J. M., R. S. N. Ferhmann, E. S. J. M. de Bont, R. M. W. Hofstra, F. Gerbens, W. A. Kamps, E. G. E. de Vries, A. G.
64 Dervishi et al.
at Universitat de Lleida on January 4, 2012jas.fass.orgDownloaded from

L. Van der Zee, G. J. Meerman, and A. ter Elst. 2007. Evidence based selection of housekeeping genes. PLoS ONE 2:e898.
Dervishi, E., C. Serrano, M. Joy, M. Serrano, C. Rodellar, and J. H. Calvo. 2010. Effect of the feedings system on the fatty acid composition, expression of the Δ9-desaturase, Peroxisome Pro-liferator-Activated Receptor Alpha, Gamma, and Sterol Regu-latory Element Binding Protein 1 genes in the semitendinous muscle of light lambs of the Rasa Aragonesa breed. BMC Vet. Res. 6:40.
Dheda, K., J. F. Huggett, S. A. Bustin, M. A. Johnson, G. Rook, and A. Zumla. 2004. Validation of housekeeping genes for nor-malizing expression in real-time PCR. Biotechniques 37:112–114.
Dieudonne, M. N., R. Pecquery, M. C. Leneveu, and Y. Giudicelli. 2000. Opposite effects of androgens and estrogens on adipo-genesis in rat preadipocytes: Evidence for sex and site-related specificities and possible involvement of insulin-like growth fac-tor 1 receptor and peroxisome proliferator-activated receptor 2. Endocrinology 141:649–656.
Doney, J. M., J. N. Peart, W. F. Smith, and F. Louda. 1979. Consid-eration of the techniques for estimation of milk yield by suckled sheep and a comparison of estimates obtained by two methods in relation to the effect of breed, level of production and stage of lactation. J. Agric. Sci. 92:123–132.
European Communities. 1986. Council Directive of 24 November 1986 on the approximation of laws, regulations and administra-tive provisions of the Member States regarding the protection of animals used for experimental and other scientific purposes. Off. J. Eur. Commun. Ser. L358:1–32.
Felder, T. K., K. Klein, W. Patsch, and H. Oberkofler. 2005. A novel SREBP-1 splice variant: Tissue abundance and transactivation potency. Biochim. Biophys. Acta 1731:41–47.
French, P., C. Stanton, F. Lawless, G. O’Riordan, F. J. Monahan, and P. J. Caffrey. 2000. Fatty acid composition, including con-jugated linoleic acid, of intramuscular fat from steers offered grazed grass, grass silage or concentrate-based diets. J. Anim. Sci. 78:2849–2855.
Gao, H., and K. Dahlman-Wright. 2010. The gene regulatory net-works controlled by estrogens. Mol. Cell. Endocrinol. 334:83–90.
Graugnard, D. E., P. Piantoni, M. Bionaz, L. L. Berger, D. B. Faulkner, and J. J. Loor. 2009. Adipogenic and energy me-tabolism gene networks in longissimus lumborum during rapid post-weaning growth in Angus and Angus × Simmental cattle fed high-starch or low-starch diets. BMC Genomics 10:142.
Hausman, G. J., M. V. Dodson, K. Ajuwon, M. Azain, K. M. Barnes, and L. L. Guan. 2009. BOARD-INVITED REVIEW: The biol-ogy and regulation of preadipocytes and adipocytes in meat animals. J. Anim. Sci. 87:1218–1246.
Heemers, H., G. Verrijdt, S. Organe, F. Claessens, W. H. G. Ver-hoeven, and J. V. Swinnen. 2004. Identification of an andro-gen response element in intron 8 of the sterol regulatory ele-ment-binding protein cleavage-activating protein gene allowing direct regulation by the androgen receptor. J. Biol. Chem. 279:30880–30887.
Herdmann, A., K. Nuernberg, J. Martin, G. Nuernberg, and O. Doran. 2010. Effect of dietary fatty acids on expression of lipo-genic enzymes and fatty acid profile in tissues of bulls. Animal 4:755–762.
Joy, M., J. Alvarez- Rodriguez, R. Revilla, R. Delfa, and G. Ripoll. 2008. Ewe metabolic performance and lamb carcass traits in pasture and concentrate-based production systems in Churra Tensina breed. Small Rumin. Res. 75:24–35.
Kay, J. K., T. R. Mackle, M. J. Auldist, N. A. Thomson, and D. E. Bauman. 2004. Endogenous synthesis of cis-9, trans-11 con-jugated linoleic acid in dairy cows fed fresh pasture. J. Dairy Sci. 87:369–378.
Kelly, M. L., J. R. Berry, D. A. Dwyer, J. M. Griinari, P. Y. Choui-nard, M. E. Van Amburgh, and D. E. Bauman. 1998. Dietary
fatty acid sources affect conjugated linoleic acid concentrations in milk from lactating dairy cows. J. Nutr. 128:881–885.
Kritchevsky, D., S. H. A. Tepper, S. Wright, P. Tso, and S. K. Czarnecki. 2000. Influence of conjugated linoleic acid (CLA) on establishment and progression of atherosclerosis in rabbits. J. Am. Coll. Nutr. 19:472S–477S.
Larionov, A., A. Krause, and W. Miller. 2005. A standard curve based method for relative real time PCR data processing. BMC Bioinformatics 6:62.
Lehnert, S. A., K. A. Byrne, A. Reverter, G. S. Nattrass, P. L. Greenwood, Y. H. Wang, N. J. Hudson, and G. S. Harper. 2006. Gene expression profiling of bovine skeletal muscle in response to and during recovery from chronic and severe undernutrition. J. Anim. Sci. 84:3239–3250.
Lundholm, L., H. Zang, A. Lind en Hirschberg, J. A. Gustafsson, P. Arner, and K. Dahlman-Wright. 2008. Key lipogenic gene ex-pression can be decreased by estrogen in human adipose tissue. Fertil. Steril. 90:44–48.
Machinal, F., M. N. Dieudonne, M. C. Leneveu, R. Pecquery, and Y. Giudicelli. 1999. In vivo and in vitro ob gene expression and leptin secretion in rat adipocytes: Evidence for a regional specific regulation by sex steroid hormones. Endocrinology 140:1567–1574.
Mayes, J. S., and G. H. Watson. 2004. Direct effects of sex steroid hormones on adipose tissues and obesity. Obes. Rev. 5:197–216.
Miguélez, E., J. M. Zumalacárregui, M. T. Osorio, A. C. Figueira, B. Fonseca, and J. Mateo. 2008. Quality traits of suckling- lamb meat covered by the protected geographical indication “Lechazo de Castilla y León” European quality label. Small Rumin. Res. 77:65–70.
Muhlhausler, B. S., V. Ritortoa, C. Schultz, B. E. Chatterton, J. A. Duffield, and I. C. McMillen. 2008. Birth weight and gender de-termine expression of adipogenic, lipogenic and adipokine genes in perirenal adipose tissue in the young adult sheep. Domest. Anim. Endocrinol. 35:46–57.
Ntambi, J. M. 1995. The regulation of stearoyl-CoA desaturase (SCD). Prog. Lipid Res. 34:139–150.
Ntambi, J. M. 1999. Regulation of stearoyl-CoA desaturase by poly-unsaturated fatty acids and cholesterol. J. Lipid Res. 40:1549–1558.
Ntambi, J. M., M. Miyazaki, J. P. Stoehr, H. Lan, C. M. Kendzior-ski, and B. S. Yandell. 2002. Loss of stearoyl-CoA desaturase-1 function protects mice against adiposity. Proc. Natl. Acad. Sci. USA 99:11482–11486.
Pariza, M. W., Y. Park, and M. E. Cook. 1999. Conjugated lin-oleic acid and the control of cancer and obesity. Toxicol. Sci. 52:107–110.
Park, Y., K. Albright, W. Liu, J. Storkson, M. Cook, and M. Pariza. 1997. Effect of conjugated linoleic acid on body composition in mice. Lipids 32:853–858.
Reverter, A., K. A. Byrne, H. L. Bruce, Y. H. Wang, B. P. Dalrym-ple, and S. A. Lehnert. 2003. A mixture model- based cluster analysis of DNA microarray gene expression data on Brahman and Brahman composite steers fed high-, medium-, and low-quality diets. J. Anim. Sci. 81:1900–1910.
Scerra, M., P. Caparra, F. Foti, V. Galofaro, M. C. Sinatra, and V. Scerra. 2007. Influence of ewe feeding systems on fatty acid composition of suckling lambs. Meat Sci. 76:390–394.
Simopoulos, A. P. 2008. The omega-6/omega-3 fatty acid ratio, ge-netic variation, and cardiovascular disease. Asia Pac. J. Clin. Nutr. 17:131–134.
Singh, R., J. N. Artaza, W. E. Taylor, N. F. Gonzalez-Cadavid, and S. H. Bhasin. 2003. Androgens stimulate myogenic differentia-tion and inhibit adipogenesis in C3H 10T1/2 pluripotent cells through an androgen receptor-mediated pathway. Endocrinol-ogy 144:5081–5088.
Swinnen, J. V., H. Heemers, T. van de Sande, E. de Schrijver, K. Brusselmans, W. Heyns, and G. Verhoeven. 2004. Androgens,
65The forage type fed affects gene expression
at Universitat de Lleida on January 4, 2012jas.fass.orgDownloaded from

lipogenesis and prostate cancer. J. Steroid Biochem. Mol. Biol. 92:273–279.
Vandesompele, J., K. De Preter, F. Pattyn, B. Poppe, N. Van Roy, A. De Paepe, and F. Speleman. 2002. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. Res. 3:34.
Wang, Y. H., N. I. Bower, A. Reverter, S. H. Tan, N. De Jager, R. Wang, S. M. McWilliam, L. M. Cafe, P. L. Greenwood, and S. A. Lehnert. 2009. Gene expression patterns during intramuscu-lar fat development in cattle. J. Anim. Sci. 87:119–130.
Waters, S. M., J. P. Kelly, P. O’Boyle, A. P. Moloney, and D. A. Kenny. 2009. Effect of level and duration of dietary n-3 poly-unsaturated fatty acid supplementation on the transcriptional regulation of delta 9-desaturase in muscle of beef cattle. J. Anim. Sci. 87:244–252.
Wood, D., and M. Enser. 1997. Factors influencing fatty acid in meat and the role of antioxidants in improving meat quality. Br. J. Nutr. 78:49–60.
Wu, Zh., Y. Xie, N. L. R. Bucher, and S. R. Farmer. 1995. Condi-tional ectopic expression of C/EBPP in NIH-3T3 cells induces PPARγ and stimulates adipogenesis. Genes Dev. 9:2350–2363.
Yao-Borengasser, A., N. Rassouli, V. Varma, A. M. Bodles, N. Ra-souli, R. Unal, B. Phanavanh, G. Ranganathan, R. McGehee Jr., and P. A. Kern. 2008. Stearoyl-coenzyme a desaturase 1 gene expression increases after pioglitazone treatment and is associated with peroxisomal proliferator-activated receptor re-sponsiveness. J. Clin. Endocrinol. Metab. 93:4431–4439.
66 Dervishi et al.
at Universitat de Lleida on January 4, 2012jas.fass.orgDownloaded from

Referenceshttp://jas.fass.org/content/90/1/54#BIBLThis article cites 54 articles, 25 of which you can access for free at:
at Universitat de Lleida on January 4, 2012jas.fass.orgDownloaded from




















