
Skull trauma in Egyptian and Hippocratic medicine - E-Periodica

498
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
© 2013 International Society of Zoological Sciences, Institute of Zoology/ Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd
Integrative Zoology 2014; 9: 498–516 doi: 10.1111/1749-4877.12062
ORIGINAL ARTICLE
Morphological integration and functional modularity in the crocodilian skull
Paolo PIRAS,1,2,3 Angela D. BUSCALIONI,4 Luciano TERESI,5 Pasquale RAIA,2,6 Gabriele SANSALONE,2,3 Tassos KOTSAKIS2,3 and Jorge CUBO7,8
1Pierre and Marie Curie University, Paris, France, 2Center for Evolutionary Ecology, Rome, Italy, 3Department of Sciences, ‘Roma Tre’ University, Rome, Italy, 4Biology Department, Universidad Autónoma de Madrid, Madrid, Spain, 5Department of Mathematics and Physics, ‘Roma Tre’ University, Rome, Italy, 6Earth Sciences Department, Federico II University, Naples, Italy, 7Sorbonne Universités, UPMC Univ Paris 06, UMR 7193, ISTeP, France and 8CNRS, UMR 7193, ISTeP, Paris, France
AbstractWe explored the morphological organization of the skull within Crocodylidae, analyzing functional and phy-logenetic interactions between its 2 constituent functional modules: the rostrum and the postrostrum. We used geometric morphometrics to identify localized shape changes, focusing on the differences between the major clades of the crown-group Crocodylia: Alligatoridae and Crocodylidae. We used published bite performance data to correlate rostral function with postrostral morphology. The skull modules appear more integrated with-in Alligatoridae than within Crocodyliade. Phylogenetic effects on shape variation are more evident in Alligato-ridae than in Crocodylidae, where functional parameters concerning the rostral morphology are proportionally more important than phylogeny. Long-snouted species are characterized by low structural performance, which is significantly associated with a reduction of the pterygoid-quadrate cranial nipper, suggesting that the nipper is important for the ingestion of large food items in generalist species. This functional association is coupled with a significant evolutionary allometry at the clade level, while Alligatoridae and Crocodylidae show different de-grees of evolutionary allometry for their entire shape and rostrum. The postrostrum is more conservative than the rostrum in terms of morphospace occupation, evolutionary allometry and phylogenetic signal.
Key words: crocodilian skull, functional performance, geometric morphometrics, modularity, morphological integration
Correspondence: Paolo Piras, Dipartimento di Scienze, Università Roma Tre, Largo San Leonardo Murialdo 1, 00146 Rome, Italy. Email: [email protected]
INTRODUCTIONThe crown-group Crocodylia is characterized by a
long evolutionary history. The most recent common an-
cestor of the Crocodylia (Gavialoidea + [Alligatoroi-dae + Crocodyloidea]) dates to 65 Ma (Brochu 2003). In this paper we analyze the evolution of the skull shape in Crocodylia by means of 3D geometric morphometrics (GM) and phylogenetic comparative methods.
The evolution of the skull shape in this group has been brilliantly studied in the past from the perspec-tive of the adaptive value of some structures within a phylogenetic framework (Iordansky 1964; Langston

499
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Morphological integration in crocodilian skull
© 2013 International Society of Zoological Sciences, Institute of Zoology/ Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
primary biological functions. Under strong functional modularity the number of functional interactions is high within a given module, but low between modules (Klin-genberg 2008). To test the modularity, we defined a pri-ori 2 morphofunctional units in the crocodilian skull: (i) the rostrum, as the portion of the skull anterior to the or-bits, primarily concerned with trophic functions (Busbey
Figure 1 Schema based on Desdevises et al. (2003) and Pi-ras et al. (2009) method showing the potential scenario for crocodylids and alligatorids when the rostral total variation is partitioned into purely ecological component (a), the phy-logenetically structured environmental variation (b), and the phylogenetically structured variation not correlated with en-vironment (c). The results of Piras et al. (2009) highlight that while factors ‘a’ and ‘b’ explained much of the rostral variation in crocodylids (27% and 19%), alligatorids variation was ex-plained mainly by factor ‘c’ (60.1%). According with this con-ceptual schema the rostral morphological evolution of crocody-lids will be traced mainly by local phenotypic responses, while alligatorid responses will be constrained by ancestors.
1965, 1973; Cleuren & de Vree, 1992; Brochu 2001). From the biomechanical perspective, Busbey (1995) pi-oneered the field, applying beam theory to understand the mechanical consequences of the acquisition of the platyrostral condition in modern crocodylians. There is now general agreement that differences among crocody-lian skulls are usually related to rostral shape and have adaptive significance. The early seminal contributions on crocodilian skull shape variation prompted empir-ical approaches testing the relationship between feed-ing performance and the occupation of ecological nich-es in both alligatorids and crocodylids (Erickson et al. 2003, 2005, 2012). Surprisingly, field experiments show that, pound per pound, maximum bite force is roughly constant across all extant crocodiles, suggesting that the evolution of adductor jaw muscles, and of the posterior portion of the crocodile skull, is very conservative (Er-ickson et al. 2012). Yet, new data on the relationship be-tween skull mechanical performance and morphology in extant crocodylians suggests that distantly-related spe-cies acquired similar snout morphologies as a response to similar ecological/functional constraints (McHen-ry et al. 2006; Pierce et al. 2008; Sadleir & Makovicky 2008; Piras et al. 2009). These authors found that dif-ferences in rostral length and width conform to different ecomorphological categories (blunt, general and slender snouts) and are correlated, in turn, to changes in oth-er skull regions (Pierce et al. 2008; Sadleir & Makov-icky 2008). One additional important notion is that the degree of skull shape variability in crocodiles is some-how limited by phylogenetic effects (Pierce et al. 2008; Piras et al. 2009). In fact, different evolutionary dynam-ics were found to apply to the evolution of the rostral region in Alligatoridae and Crococdylidae (Fig. 1) (Pi-ras et al. 2009). In Alligatoridae, shape variation in the rostrum does not reflect adaptation to current ecologi-cal conditions. Rather, it bears the signature of a strong phylogenetic conservatism. In stark contrast, within the Crococdylidae rostrum shape variation shows a strong ecological component. Therefore, the 2 major crocodyl-ian clades seem to have either history or function as the main determinant of current species skull shapes.
Here, we test the hypothesis that the crocodylian skull is differently integrated in Crococdylidae and Alli-gatoridae, as a result of different role of function con-trasted to evolutionary history of the 2 clades. To this aim, we explore patterns of morphospace occupation in the 2 clades, and the functional modularity in their skulls, here defined as the degree of connectivity within and among units/modules identified on the basis of their

500
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
P. Piras et al.
© 2013 International Society of Zoological Sciences, Institute of Zoology/ Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
1995), involving the anterior extension of the frontal, nasals, premaxillae and maxillae, as elements mainly as-sociated with prey capture and manipulation and (ii) the postrostrum, including elements of the secondary pal-ate and suspensorium that conform the skeletal support of jaw musculature, as well as those elements delimiting the central nervous (brain) and sensory (optic and audi-tory) systems. We then tested the degree of integration between these 2 modules. In addition, to test whether the rostrum is a major driving force in coordinating the changes of the entire skull, we also tested for covaria-tion between the rostrum and the entire shape, an analy-sis often called part–whole analysis (Marquez 2008) that allows assessing which, among the 2 modules we iden-tified, is the driving force in skull shape change. We ex-pect that crocodylids show less morphological integra-tion than alligatorids because of the greater emphasis on functional variation in their evolution (Piras et al. 2009). Finally, we analyzed evolutionary allometry in both the entire clade and the above mentioned subclades, taking phylogenetic relationships into account to control for co-variance between observations due to phylogeny.
MATERIALS AND METHODS
Materials
We refer to the Alligatoridae and Crocodylidae as the 2 main subclades of Crocodylia. Our dataset covers all extant crocodilian species over a wide spectrum of onto-genetic stages (juveniles to adults). Besides the true gh-arial Gavialis gangeticus Gmelin, 1789 (here represent-ed by 20 individuals), we collected shape information on skulls of Alligatoridae (157 individuals, 8 extant spe-cies) and Crocodylidae (223 individuals, 14 extant spe-cies) (see online Table S1). Three fossil Crocodylidae for which well-preserved 3D skulls were available were added to the dataset: Voay robustus (Grandidier and Vaillant, 1872) (Brochu 2007a) from the Holocene of Madagascar, Crocodylus ossifragus Dubois, 1908 from the Pleistocene of Java, which is considered by some au-thors to be a synonym of the extant Crocodylus siamen-sis Schneider, 1801 (Delfino & de Vos 2010), and Dol-losuchoides densmorei Brochu, 2007 (Brochu 2007b) from the Early Eocene of Belgium. Non-deformed 3D fossil Alligatoridae were not available for this study. De-tails about specimens are available online in Table S1.
Geometric morphometrics
A 3D configuration of 90 unilateral landmarks was digitized on the left side of the skull to capture its mor-
phology: landmark definitions, positions and their cor-responding configurations are shown in Figure 2. Data were collected with an Immersion Microscribe G2. Morphology has been approached by means of land-marks that were selected to approximate skull geome-try, taking into account: (i) extreme landmarks denot-ing relative dimensions (e.g. landmark 39-anterior tip of premaxilla); and (ii) bone relationships according to su-tural connections. Selected landmarks should capture the major shape differences between the 2 major clades of the Crocodylia crown-group (i.e. Crocodylidae and Alligatoridae plus Gavialis). To assess integration and modularity, landmarks were partitioned into 2 func-tional modules. Despite the fact that different modules might be defined from a developmental point of view, they are rarely recognized a priori (Marquez 2008). This happens because different mesenchymal ossifica-tions, which represent true developmental modules reg-ulated by different growth factors, can be barely recog-nized in a completely developed structure. This leads us to incorporate the 2 complexes, the rostrum and the pos-trostrum, as supermodules that include multiple regions descending from different mesenchymal ossifications. Thus, we consider 2 designated morphological modules (Fig. 2h,i) chosen on the basis of their functional role in the context of the general crocodylian skull morpholo-gy. In fact, the rostrum and postrostrum are easy to rec-ognize. Still, each module includes different morpho-logical components that provide quite distinct functional roles. The rostrum, as defined here, relates to prey cap-ture and manipulation. Different ecomorphs are recog-nized in crocodilians, from generalized (Alligator spp., Crocodylus niloticus Laurenti, 1768) to highly special-ized forms (Tomistoma schlegelii Müller, 1838; G. gan-geticus). The rostrum was here defined as the set of 21 landmarks including the bony elements with a prima-ry trophic function, such as the premaxilla, nares and maxilla (see Busbey 1995). The postrostrum includes a set of 41 landmarks delimiting the central sensory sys-tem encompassing the orbits, the braincase, the dermal bones of the skull table, the quadrate articulation for the lower jaw, and the posterior region of the palate.
Generalized Procrustes Analysis (GPA) was used to analyze shape (Bookstein 1991; Goodall 1991). GPA ro-tates, aligns and scales landmark configurations to cen-troid size (CS: the square root of the sum of squared distances of a set of landmarks from their centroid; Bookstein 1986). Rotation of the scaled and translat-ed landmark sets is achieved via comparison with a ref-erence configuration (the first specimen in the dataset).
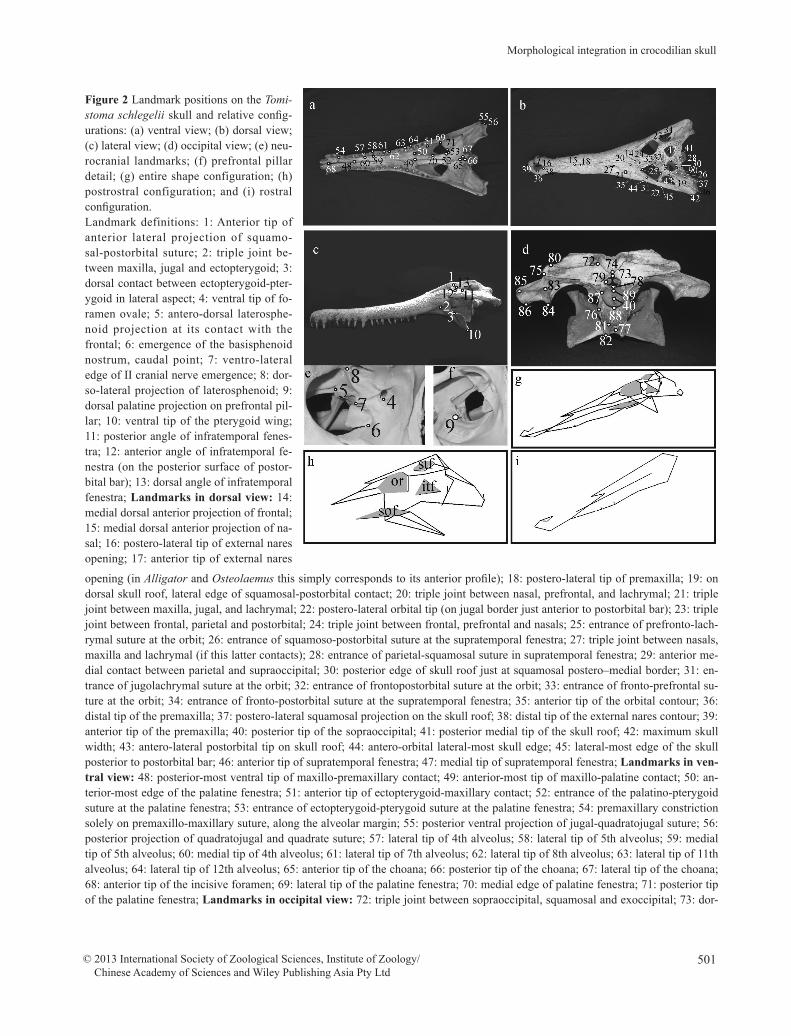
501
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Morphological integration in crocodilian skull
© 2013 International Society of Zoological Sciences, Institute of Zoology/ Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Figure 2 Landmark positions on the Tomi-stoma schlegelii skull and relative config-urations: (a) ventral view; (b) dorsal view; (c) lateral view; (d) occipital view; (e) neu-rocranial landmarks; (f) prefrontal pillar detail; (g) entire shape configuration; (h) postrostral configuration; and (i) rostral configuration. Landmark definitions: 1: Anterior tip of anterior lateral projection of squamo-sal-postorbital suture; 2: triple joint be-tween maxilla, jugal and ectopterygoid; 3: dorsal contact between ectopterygoid-pter-ygoid in lateral aspect; 4: ventral tip of fo-ramen ovale; 5: antero-dorsal laterosphe-noid projection at its contact with the frontal; 6: emergence of the basisphenoid nostrum, caudal point; 7: ventro-lateral edge of II cranial nerve emergence; 8: dor-so-lateral projection of laterosphenoid; 9: dorsal palatine projection on prefrontal pil-lar; 10: ventral tip of the pterygoid wing; 11: posterior angle of infratemporal fenes-tra; 12: anterior angle of infratemporal fe-nestra (on the posterior surface of postor-bital bar); 13: dorsal angle of infratemporal fenestra; Landmarks in dorsal view: 14: medial dorsal anterior projection of frontal; 15: medial dorsal anterior projection of na-sal; 16: postero-lateral tip of external nares opening; 17: anterior tip of external nares
opening (in Alligator and Osteolaemus this simply corresponds to its anterior profile); 18: postero-lateral tip of premaxilla; 19: on dorsal skull roof, lateral edge of squamosal-postorbital contact; 20: triple joint between nasal, prefrontal, and lachrymal; 21: triple joint between maxilla, jugal, and lachrymal; 22: postero-lateral orbital tip (on jugal border just anterior to postorbital bar); 23: triple joint between frontal, parietal and postorbital; 24: triple joint between frontal, prefrontal and nasals; 25: entrance of prefronto-lach-rymal suture at the orbit; 26: entrance of squamoso-postorbital suture at the supratemporal fenestra; 27: triple joint between nasals, maxilla and lachrymal (if this latter contacts); 28: entrance of parietal-squamosal suture in supratemporal fenestra; 29: anterior me-dial contact between parietal and supraoccipital; 30: posterior edge of skull roof just at squamosal postero–medial border; 31: en-trance of jugolachrymal suture at the orbit; 32: entrance of frontopostorbital suture at the orbit; 33: entrance of fronto-prefrontal su-ture at the orbit; 34: entrance of fronto-postorbital suture at the supratemporal fenestra; 35: anterior tip of the orbital contour; 36: distal tip of the premaxilla; 37: postero-lateral squamosal projection on the skull roof; 38: distal tip of the external nares contour; 39: anterior tip of the premaxilla; 40: posterior tip of the sopraoccipital; 41: posterior medial tip of the skull roof; 42: maximum skull width; 43: antero-lateral postorbital tip on skull roof; 44: antero-orbital lateral-most skull edge; 45: lateral-most edge of the skull posterior to postorbital bar; 46: anterior tip of supratemporal fenestra; 47: medial tip of supratemporal fenestra; Landmarks in ven-tral view: 48: posterior-most ventral tip of maxillo-premaxillary contact; 49: anterior-most tip of maxillo-palatine contact; 50: an-terior-most edge of the palatine fenestra; 51: anterior tip of ectopterygoid-maxillary contact; 52: entrance of the palatino-pterygoid suture at the palatine fenestra; 53: entrance of ectopterygoid-pterygoid suture at the palatine fenestra; 54: premaxillary constriction solely on premaxillo-maxillary suture, along the alveolar margin; 55: posterior ventral projection of jugal-quadratojugal suture; 56: posterior projection of quadratojugal and quadrate suture; 57: lateral tip of 4th alveolus; 58: lateral tip of 5th alveolus; 59: medial tip of 5th alveolus; 60: medial tip of 4th alveolus; 61: lateral tip of 7th alveolus; 62: lateral tip of 8th alveolus; 63: lateral tip of 11th alveolus; 64: lateral tip of 12th alveolus; 65: anterior tip of the choana; 66: posterior tip of the choana; 67: lateral tip of the choana; 68: anterior tip of the incisive foramen; 69: lateral tip of the palatine fenestra; 70: medial edge of palatine fenestra; 71: posterior tip of the palatine fenestra; Landmarks in occipital view: 72: triple joint between sopraoccipital, squamosal and exoccipital; 73: dor-
502
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
P. Piras et al.
© 2013 International Society of Zoological Sciences, Institute of Zoology/ Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Once that rotation has been completed, a mean shape is calculated and the rotation process is repeated using the mean shape as the reference configuration for the sam-ple (including the previous empirical reference-speci-men configuration). This mean-shape/rotation procedure is iterated to minimize rotation differences between sub-
sequent iterations. Residual differences, then, are caused only by shape differences. Principal component analy-sis (PCA) was performed on the Procrustes residuals to find the orthogonal axes of maximal variation (Table 1). This is a common procedure in geometric morphometric studies (Adams et al. 2004).
sal tip of the foramen magnum; 74: mid-joint between sopraoccipital and exoccipitals; 75: ventro-lateral projection of paraoccipital process; 76: lateral tip of basioccipital just at exoccipital-basioccipital suture; 77: ventro- medial contact between pterygoid and ba-sisphenoid; 78: ventral tip of the foramen magnum; 79: lateral tip of the foramen magnum; 80: dorso-lateral projection of the para-occipital process; 81: ventro-lateral tip of the basioccipital tubera; 82: postero-lateral tip of the pterygoid posterior process; 83: dor-so-lateral tip of medial quadrate hemicondyle; 84: ventro-lateral tip of medial quadrate hemicondyle; 85: dorso-lateral tip of distal quadrate hemicondyle; 86: ventro-lateral tip of distal quadrate hemicondyle; 87: lateral tip of the basioccipital condyle; 88: dorsal tip of the basioccipital condyle; 89: ventral tip of the basioccipital condyle; 90: posterior tip of the supratemporal fenestra. Note that the rostral configuration was obtained with the following landmarks: 14–18, 35–36, 38–39, 48–49, 54, 57–64, 68, while the postros-tral configuration was obtained with the following landmarks: 1, 10–14, 19–20, 22, 24–28, 31–34, 37, 40–47, 50, 69–71, 73, 75–79, 83, 85, 89–90.
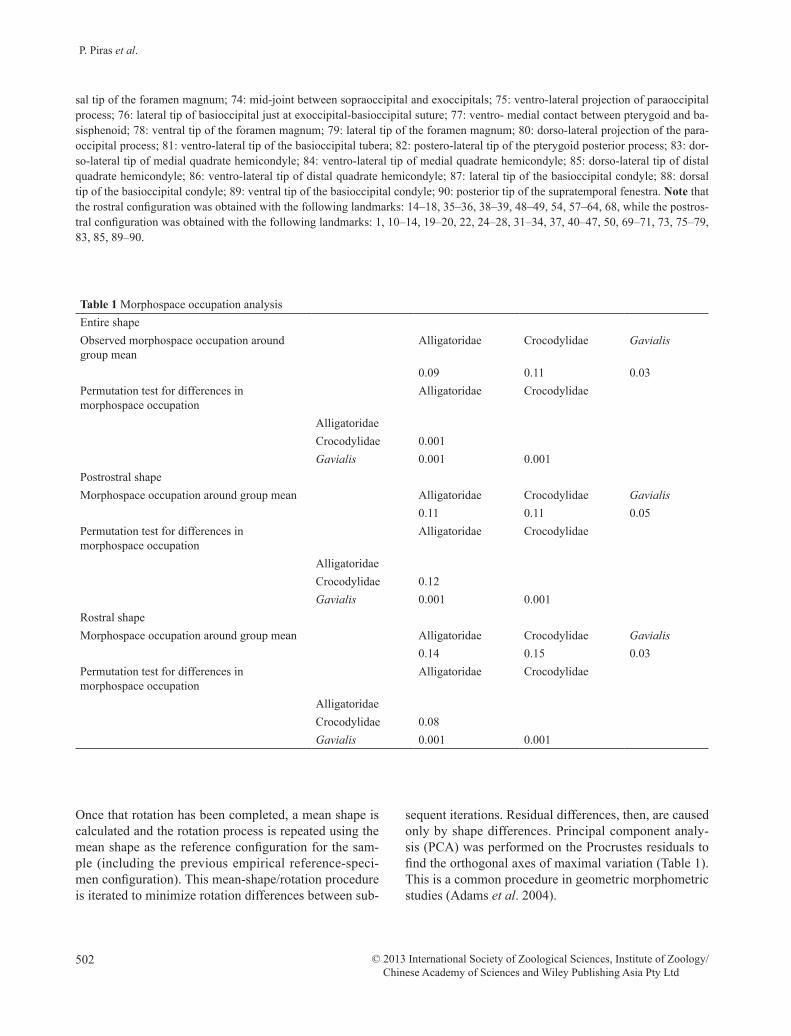
Table 1 Morphospace occupation analysisEntire shapeObserved morphospace occupation around group mean
Alligatoridae Crocodylidae Gavialis
0.09 0.11 0.03Permutation test for differences in morphospace occupation
Alligatoridae Crocodylidae
AlligatoridaeCrocodylidae 0.001Gavialis 0.001 0.001
Postrostral shapeMorphospace occupation around group mean Alligatoridae Crocodylidae Gavialis
0.11 0.11 0.05Permutation test for differences in morphospace occupation
Alligatoridae Crocodylidae
AlligatoridaeCrocodylidae 0.12Gavialis 0.001 0.001
Rostral shapeMorphospace occupation around group mean Alligatoridae Crocodylidae Gavialis
0.14 0.15 0.03Permutation test for differences in morphospace occupation
Alligatoridae Crocodylidae
AlligatoridaeCrocodylidae 0.08Gavialis 0.001 0.001

503
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Morphological integration in crocodilian skull
© 2013 International Society of Zoological Sciences, Institute of Zoology/ Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
As we were interested in studying interspecific shape differences and evolutionary allometry (if any), we re-moved the ontogenetic intraspecific variation by per-forming separate per-species multivariate regressions between shape and size. Then, for each species, the re-siduals were added to species specific shapes predict-ed at maximum species specific size values. This proce-dure ensures elimination of intraspecific allometry while maintaining the interspecific size-shape differences due to evolutionary allometry. In fact, species specific resid-uals alone have means of zero by definition thus elimi-nating any evolutionary allometric signal. This strategy is common in GM studies (Zeldtich et al. 2004, 2012) where it is often called (perhaps misleadingly) standard-ization at adult size. Data so arranged were then used in all successive analyses.
The 3 datasets representing entire shape, rostral and post-rostral configurations were analyzed separately in order to explore shape differences between species. To test for modularity and establish whether it differs be-tween the 2 major clades (Alligatoridae and Crocodyli-dae), 3 separate GPAs and successive PCAs procedures were performed on each configuration (entire shape, ros-trum and postrostrum). We successively tested morpho-logical integration for the pooled sample, and in Alliga-toridae and Crocodylidae samples separately.
Phenotypic disparity was studied by performing a Levene’s test on a performance variable (univariate [see below]) and morphospace occupation analysis on shape variables (Alligatoridae and Crocodylidae) using the betadisper() function of R package ‘vegan’ (Oksanen et al. 2008). Morphospace occupation was evaluated after a common GPA because it makes sense to calculate dis-parities and morphospace occupations only for groups that belong to a common empirical morphospace. We fi-nally assessed the differences in morphospace occupa-tion between rostral and postrostral configurations to look for a differential signal in conservatism of the 2 functional modules.
Beam theory
We used the data from Piras et al. (2009) as a proxy of structural performance of the rostrum. Piras et al. (2009) applied beam theory to geometries of rostra to quantify their mechanical behavior (Fig. 3a). Erickson et al. (2012) show that maximum bite force scales con-stantly in all extant Crocodylia, confirming their previ-ous findings (Erickson et al. 2003, 2005) upon which
Figure 3 (a) Geometric model for beam theory analysis start-ing from the actual skull morphology and analyzing 3 coronal sections: premaxillo-maxillary constriction, diameter at the 4th alveolus, and preorbital diameter and (b) loading conditions studied for the beam analysis. Schematic drawings show sec-tions of rostrum corresponding to the described anatomical lev-els (see text). Arrows indicate forces applied.
Piras et al. (2009) built their model. The authors found that, in all species (including brevirostral and longiros-tral forms), both absolute values and values scaled to al-lometric change follow the same trajectory. The perfor-mance variable used here (taken from Piras et al. 2009) represents sigma (Fig. 3b). Sigma is often used to pre-dict the failure points in brittle material, so we consid-er it to be a meaningful characteristic that assesses the overall mechanical performance of the rostrum.
To eliminate intraspecific ontogenetic allometry, we scaled this variable in the same way that we did for shape variables. Since Erickson et al. (2012) correlated their results with morphology using body mass and fur-nished a morphometric allometric equation using snout vent length and tail length (information unavailable for our study), we used postrostral centroid size as a proxy of body mass. We specifically avoided the use of cen-troid size of entire skull shape because in longirostral species the centroid size of the entire configuration is positively biased by their peculiar rostral morphology. Thus, we judge postrostral size to be a better proxy for body mass in comparison to the size of entire skull con-figuration.
Phylogeny
Crocodylian phylogeny is characterized by a conflict between molecular and morphological/paleontologi-cal studies (see Brochu 2003; McAliley et al. 2006; Jan-ke et al. 2006 with references for a review; Oaks 2011; Erickson et al. 2012). The bone of contention is the po-

504
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
P. Piras et al.
© 2013 International Society of Zoological Sciences, Institute of Zoology/ Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
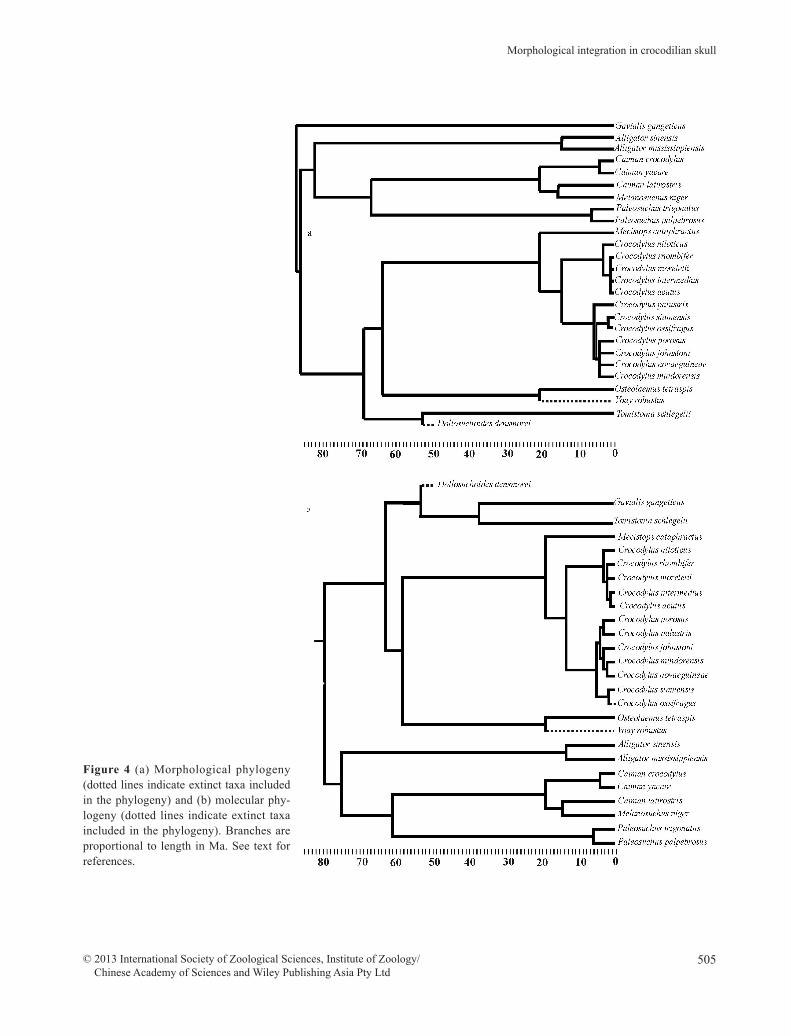
sition of Gavialis, which includes the sole extant spe-cies, G. gangeticus (the true gharial). In morphologi-cal/paleontological studies Gavialis is placed outside all other species (as the sister taxon of all the other mem-bers of the crown group Crocodylia) with a divergence time in the Late Cretaceous (Brochu 2003). In contrast, according to molecular studies, it is the sister taxon to Schlegel’s false gharial, T. schlegelii, and their separa-tion is estimated to have occurred in the Late Eocene, at the very earliest (Janke et al. 2006; Oaks 2011). Recent analyses focused on the ontogenetic allometry of Tomi-stoma and Gavialis as compared to Crocodylus support the morphological hypothesis (Piras et al. 2010). The re-maining extant species are unambiguously grouped into 2 clades: Alligatoridae and Crocodylidae. There remains some uncertain relationships within the genus Crocody-lus, but most nodes of the group are stable. We consid-ered the 2 phylogenies in order to test their differential effect in comparative analyses. The 2 topologies are pre-sented in Figure 4. The trees were based upon Brochu (2003), Gatesy et al. (2003), Janke et al. (2006), Wil-lis et al. (2007), Oaks (2011) and Erickson et al. (2012). Clearly, it is not possible to give a very precise estima-tion for all branches, but we doubt that small uncertain-ties would affect the results significantly. For example, for Paleosuchus spp. divergence time is not available and it was tentatively placed by Brochu (2003), and here accordingly, at the Early Pliocene, 5 Ma. Small changes in the topology or branch lengths, considering more re-cent divergences, should not change the results.
Morphological integration and functional modularity
We used the RV coefficient (Escoufier 1973) to test for integration between postrostrum and rostrum using the RV.rtest() function of the R package ‘ade4’ (Dray et al. 2007). We repeated this analysis in both the pooled sample and on Alligatoridae and Crocodylidae separate-ly to test their differential degree of integration (if any). Part–whole analyses were performed to assess the most influential module in entire skull shape change.
We used Partial Least Squares (PLS) analysis to vi-sualize shape changes associated with rostrum and pos-trostrum covariation (Rohlf & Corti 2000).
Linear models and comparative methods
We used per species maximum size values in linear models. This is consistent with the analyses performed here, which in some cases require standardization (of
shape) at adult size. In fact, we performed such stan-dardization by adding the shape predicted at maximum size recorded for each species to the residuals coming from per-species multivariate regression between shape and size.
We first assessed differences in size, structural per-formance and shape between major clades (coded as factor) by means of ANOVA (for size and performance) and MANOVA (for shape variables) using the adonis() function from the R package ‘vegan’ (Oksanen et al. 2008) that allows handling fatty data (i.e. with more variables than cases) by means of non-parametric test of significance based on permutations.
Then, we explored evolutionary allometry for both the pooled sample and for Alligatoridae and Crocody-lidae taken separately. We performed linear regression models between shape data (dependent) and size (inde-pendent).
In the case of significant slopes for subclades we per-formed the ontogenetic convergence test as described in Piras et al. (2010, 2011), to assess whether the tra-jectories are parallel, convergent or divergent. Just for the sake of visualization, we used canonical correlation analysis using the rcc() function of R package ‘CCA’ (Gonzalez & Dejean 2012). These analyses were per-formed separately on entire shape, postrostral shape and rostral shape.
Finally, we correlated the performance variable sig-ma with postrostral shape in order to assess the modifi-cations affecting the postrostrum that covary with per-formance. To do this we eliminated variation due to interspecific size differences in our model. In fact, the performance variable, averaged as explained above, still maintains the evolutionary allometric signal due to in-terspecific maximum size differences. Our strategy is suited to eliminate even the interspecific size depen-dence. Thus, we regressed the per-species averaged per-formance variable values (standardized as explained above) on per species maximum size of postrostrum (our best proxy of body mass) and retained the residuals. We then regressed, against these residuals, the residuals of postrostral shape derived from a multivariate regression between shape and size. In this way, we contrasted per-formance and shape data variation as not influenced by evolutionary allometry.
All these analyses were accompanied by their phylo-genetic versions using the phy.anova() and phy.mano-va() from R package ‘geiger’ (Harmon et al. 2009) and the R function genpgls() written by P. Piras is available
505
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Morphological integration in crocodilian skull
© 2013 International Society of Zoological Sciences, Institute of Zoology/ Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Figure 4 (a) Morphological phylogeny (dotted lines indicate extinct taxa included in the phylogeny) and (b) molecular phy-logeny (dotted lines indicate extinct taxa included in the phylogeny). Branches are proportional to length in Ma. See text for references.

506
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
P. Piras et al.
© 2013 International Society of Zoological Sciences, Institute of Zoology/ Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
in online Table S2; this function, when the dependent variable is univariate, automatically determines the best mode of Y table character evolution using fitContinuous() from the R package ‘geiger’ and, if the most support-ed mode is not Brownian, scales branch lengths accord-ing to the best model parameters before calculating the phylogenetic covariance matrix that transforms observa-tions during the Phylogenetic Generalized Least Squares (PGLS) procedure (Rohlf 2001, 2006). In the case of a multivariate Y table, Brownian motion is assumed.
Both molecular and morphological phylogenies shown in Figure 4 were used in these comparative anal-yses.
RESULTS
Morphospace occupation and skull shape variation
Figure 5 shows results of GM analyses. Figure S1 shows, in color, the same scatterplots with groups iden-tified at the specific level. For the entire shape (Fig. 5a), PC1 mainly captures the rostrum elongation: the longi-rostral G. gangeticus is located at PC1 positive extreme, while the alligatorids Caiman latirostris Daudin, 1802 and Melanosuchus niger Spix, 1825 occupy the area of brevi- and mesorostral skulls, on the opposite side of the axis. Shorter rostra are associated with broader orbits and reduced temporal fenestrae in alligatorids, while short-snouted crocodylids also bear large palatines (rel-ative to the orbits) and vertically-oriented infratemporal fenestrae. It must also be noted that species are segre-gated into 2 phylogenetically-consistent groups (Alliga-tor spp. versus caimans) in the alligatorid morphospace, while for crocodylids it is hard to recognize any phylo-genetic ordering. Postrostral shape (Fig. 5b) allows us to distinguish crocodylids and alligatorids. In fact, PC1 is related to the expansion of quadrates and to the relative elongation of lachrymal and prefrontal bones in both clades. Taxa with short quadrates and prefrontals are placed towards PC1 negative values, while those at pos-itive values have long lachrymals and laterally expanded quadrates. On PC2 the 2 clades are separated except for the dwarf crocodylid Osteolaemus tetraspis Cope, 1860, which falls within the alligatorid portion of the axis. This overlap is due to large orbits and small supratem-poral fenestrae, in relation to the skull size, of Osteolae-mus. In fact, PC2 captures the correlation between the relative sizes of the orbits, the supratemporal, infratem-poral and palatine fenestrae, and, an important anatomi-
cal aspect for discriminating between Crocodylidae and Alligatoridae, the configuration of the quadrate articu-lar area and of the paraoccipital process. In crocodylids the quadrate is posterolaterally extended while the para-occipital process is reduced. Palatine and infratemporal fenestrae follow a strikingly inverse pattern. At the neg-ative PC2 extreme the infratemporal fenestra becomes relatively larger in alligatorids, whereas the palatine fe-nestra appears smaller in comparison to crocodylids for negative values. PC2 separates Gavialis from Croco-dylia, which is shifted to the negative extremes of the axis. Gavialis morphology presents an increased dimen-sion of the supratemporal fenestrae relative to the orbit, a marked orbit telescopicity, an anterior extension of the frontal bone, a small lateral expansion of the quadrate, and a relative reduction in the verticalization of the pter-ygoid flange. The occupation of the postrostral space by Alligatoridae is shaped according to the relative dimen-sion of the supratemporal fenestra (Fig. 5b, PC1). At the negative extreme, Paleosuchus spp. has complete-ly closed supratemporal fenestrae, while Alligator spp. is at the opposite extreme. Other changes are related to the relative dimensions of the orbits (which are rela-tively larger towards negative values). The extreme po-sition of Paleosuchus spp. results from the correlation between the verticalization of the infratemporal fenes-tra and the occipital area, the reduction of the quadrate articular area, the relative shortening of the pterygoid flanges, and the enlargement of the palatine fenestra. PC2 is related to the length of the prefrontal, which is longer in Alligator (on negative values). The postrostral changes in crocodylids involve the posterolateral expan-sion of the quadrate for positive values of PC1 (where C. niloticus and Crocodylus porosus Schneider, 1801 are positioned). Negatives values are occupied by Croco-dylus mindorensis Schmidt, 1935, Crocodylus johnstoni Krefft, 1873 and O. tetraspis with shorter quadrates and visibly larger orbits. PC2 scores correlate to lachrymal lengthening and to the reduction of the supratemporal fenestra; the positive extreme is occupied by O. tetras-pis, and the negative extreme by Tomistoma, which has extremely long lachrymals and relatively wide fenes-trae.
The particular analysis of the rostral shape indicates a similar morphospace occupation for the entire skull shape (Fig. 5c). Rostral elongation is associated with maxillary and premaxillary (to a lesser extent) elon-gation and with relative nasal position. This implies a lengthening of nasals relative to the total length of ros-trum with a consequent contact between nasals and the

507
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Morphological integration in crocodilian skull
© 2013 International Society of Zoological Sciences, Institute of Zoology/ Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Figure 5 PC1/PC2 scatterplots after Gen-eralized Procrustes Analysis (GPA) per-formed for entire shape, rostrum and postrostrum, showing the deformations as-sociated to positive and negative value of these axes. Deformations refer to extreme negative or positive values of principal components. Online Figure S1 shows the same scatterplots in color with points iden-tification at specific level and abbreviations as in Figure 7.

508
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
P. Piras et al.
© 2013 International Society of Zoological Sciences, Institute of Zoology/ Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
external nares. Nonetheless, the relative position of the nasals is crucial in rostral shape analysis, because those of G. gangeticus do not contact the premaxilla or the ex-ternal nares. This discrete character (an autapomorphy of the genus Gavialis, see character 95 in Brochu 1999) was detected in the morphospace. Within the Alligato-ridae, the nasals of Alligator spp. bisect the external na-res (see PC2 in Fig. 5c: positive values divide Alligator spp. from caimans, which bear relatively shorter na-sals). Within Crocodylidae, Tomistoma and Osteolaemus have short nasals (see PC2 in Fig. 5c). The false ghari-al is morphologically extreme because of the posterior position of the nasals, and the premaxilla extending to the level of the fifth maxillary alveolus. Morphological variation along PC2 is extremely interesting because it explains, apparently, the same shape change of PC1 (i.e. rostrum elongation), but by means of opposite shape changes of nasal and premaxilla.
Morphospace occupation analysis (Table 1) revealed significant differences between Crocodylidae and Alli-gatoridae only for entire shape, while postrostrum and rostrum differences are not significant. Levene’s test on the performance variable is not significant when con-trasting Allgatoridae and Crocodylidae, although is close to the significance level (P = 0.08). Postrostrum occupation is significantly smaller than rostrum occupa-tion in both Alligatoridae (P = 0.001) and Crocodylidae (P = 0.001).
Morphological integration and functional modularity
The results of PLS analysis are shown in Figure 6. We show the covariation for the pooled sample, as well as for Alligatoridae and Crocodylidae separately.
Positive and negative extremes of the first pair of PLS axes reflect brevirostral and longirostral skull mor-photypes in all analyses. At the negative extremes (Fig. 6d–f) of the first singular vector, postrostral morphology is characterized by an increased relative size of the su-pratemporal fenestra and by the lengthening of the fron-tal spine. Another important change is the reduction of the pterygoidal flange, toward negative values, result-ing in a less open morphology of the skull profile (when viewed laterally) and a backward shift of the palatine fenestra. The reduction of the pterygoid flange, which implies a significant reduction of the cranial nipper in its vertical extent, could cause a reduction in maxi-mal mouth opening. These features correspond, along the first singular vector, to the longirostral morpholo-gy, such as those of T. schlegelii and G. gangeticus. At
the positive extremes (Fig. 6a–c) we found the opposite condition whereby more open cranial nippers were as-sociated with the broadest rostra. Morphological visual-ization shows that postrostrum varies differently in alli-gatorids and in crocodylids.
Results of the RV coefficient computed for morpho-logical integration are given in Table 2. As expected, all comparisons are statistically supported. Crocodylidae are less integrated than Alligatoridae.
Alligatoridae show larger RV for all comparisons, thus denoting a greater morphological integration than Crocodylidae. Higher values of rostral covariation and entire skull shape may indicate that this module is dom-inating the morphological variation even in the allig-atorid skull. Our hypothesis that rostrum drives shape changes of the entire skull configuration is strongly sup-ported by our results.
Figure 6 Morphological changes associated with the first sin-gular vector after the partial least squares analysis performed on rostrum–postrostrum comparison (rostrum in dorsal view, postrostrum in lateral view). Rostral and postrostral shapes are mutually contrasted in the figures and correspond to the 2 ex-tremes of the singular vectors. (a) Pooled sample, positive ex-treme of the first singular vector; (b) Alligatoridae, positive ex-treme of the first singular vector; (c) Crocodylidae, positive extreme of the first singular vector; (d) pooled sample, nega-tive extreme of the first singular vector; (e) Alligatoridae, nega-tive extreme of the first singular vector; (f) Crocodylidae, neg-ative extreme of the first singular vector.
509
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Morphological integration in crocodilian skull
© 2013 International Society of Zoological Sciences, Institute of Zoology/ Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Table 2 RV coefficients computed for integration on pooled sample, Alligatoridae and Crocodylidae
RV coefficient P after 10 000 permutationsPooled simple Entire shape–postrostrum 0.61 0.001
Entire shape–rostrum 0.93 0.001Postrostrum–rostrum 0.61 0.001
Alligatoridae Entire shape–postrostrum 0.81 0.001Entire shape–rostrum 0.86 0.001Postrostrum–rostrum 0.61 0.001
Crocodylidae Entire shape–postrostrum 0.56 0.001Entire shape–rostrum 0.95 0.001Postrostrum–rostrum 0.53 0.001
Table 3 P-values for ANOVA and MANOVA and their phylogenetic versions run on Crocodylidae and Alligatoridae treated as factor. Dashes are present when the corresponding test was not applicable
Maximum size recorded ShapeEntire shapeANOVA 0.048*Phyl.ANOVA molecular 0.068Phyl.ANOVA morphological 0.049*MANOVA — P < 0.001*Phyl.MANOVA molecular — P < 0.001*Phyl.MANOVA morphological — P < 0.001*Postrostral shapeANOVA 0.09Phyl.ANOVA molecular 0.11Phyl.ANOVA morphological 0.12MANOVA — P < 0.001*Phyl.MANOVA molecular — P < 0.001*Phyl.MANOVA morphological — P < 0.001*Rostral shapeANOVA 0.034*Phyl.ANOVA molecular 0.036*Phyl.ANOVA morphological 0.025*MANOVA — P < 0.001*Phyl.MANOVA molecular — P < 0.001*Phyl.MANOVA morphological — P < 0.001*
*Significant results. Phyl., phylogenetic.
Linear models and comparative methods
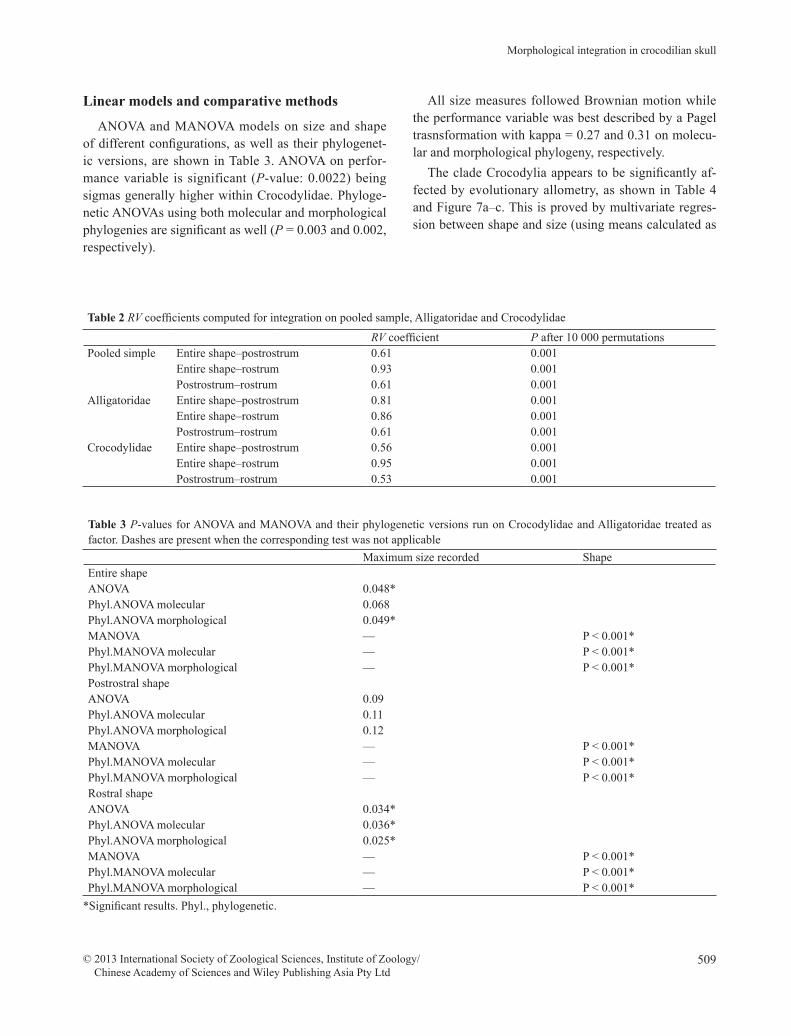
ANOVA and MANOVA models on size and shape of different configurations, as well as their phylogenet-ic versions, are shown in Table 3. ANOVA on perfor-mance variable is significant (P-value: 0.0022) being sigmas generally higher within Crocodylidae. Phyloge-netic ANOVAs using both molecular and morphological phylogenies are significant as well (P = 0.003 and 0.002, respectively).
All size measures followed Brownian motion while the performance variable was best described by a Pagel trasnsformation with kappa = 0.27 and 0.31 on molecu-lar and morphological phylogeny, respectively.
The clade Crocodylia appears to be significantly af-fected by evolutionary allometry, as shown in Table 4 and Figure 7a–c. This is proved by multivariate regres-sion between shape and size (using means calculated as

510
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
P. Piras et al.
© 2013 International Society of Zoological Sciences, Institute of Zoology/ Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Figure 7 Evolutionary allometries for en-tire shape (a), postrostrum (b), rostrum (c) and relationship between size free perfor-mance and postrostrum data (d). Species abbreviations: Am, Alligator mississippi-ensis; As, Alligator sinensis; Cc, Caiman crocodylus; Cl, Caiman latirostris; Cy, Caiman yacare; Mn, Melanosuchus niger; Pp, Paleosuchus palpebrosus; Pt, Paleo-suchus trigonatus; Ca, Crocodylus acutus; Mcat, Mecistops cataphractus; Cin, Croc-odylus intermedius; Cj, Crocodylus john-stoni; Cm, Crocodylus mindorensis; Cmor, Crocodylus moreletii; Cnil, Crocodylus ni-loticus; Cnov, Crocodylus novaeguineae; Cpal, Crocodylus palustris; Cpor, Croco-dylus porosus; Crho, Crocodylus rhombi-fer; Cs, Crocodylus siamensis; Ot, Osteo-laemus tetraspis; Ts, Tomistoma schlegelii; Crob, Voay robustus; Coss, Crocodylus os-sifragus; Dollo, Dollosuchoides densmorei; Gg, Gavialis gangeticus.
511
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Morphological integration in crocodilian skull
© 2013 International Society of Zoological Sciences, Institute of Zoology/ Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Table 4 P-values for evolutionary allometry analyses. Both OLS and PGLS are shown. PGLSmol: PGLS performed using molec-ular phylogeny; PGLSmor: PGLS performed using morphological phylogeny.
OLS PGLSmol PGLSmorphEntire shape dataPooled dataset 0.001* 5.30×10-7* 5.20×10-7*Alligatoridae 0.11 0.16 0.16Crocodylidae 0.03* 7.60×10-6* 7.50×10-6*Postrostrum shape dataPooled dataset 0.001* 8.40×10-7* 7.10×10-7*Alligatoridae 0.035* 0.11 0.11Crocodylidae 0.001* 1.14×10-7* 1.14×10-7*Rostrum shape dataPooled dataset 0.001* 4.50×10-5* 1.64×10-5*Alligatoridae 0.45 0.35 0.35Crocodylidae 0.001* 0.001* 0.001*PerformancePooled dataset 0.001* 0.0085* 0.0084*Alligatoridae 0.09 0.27 0.26Crocodylidae 0.055 0.03* 0.03*
*Significant results.
explained above). This holds true for the entire shape, the postrostral shape and the rostral shape. In pooled analysis smaller species have shorter rostra. The same signal is actually present within Crocodylidae for all configurations under OLS and PGLS using both topolo-gies. Alligatoridae do not show the same signal of Croc-odylidae for rostrum and entire shape data, while they do for postrostrum. An ontogenetic convergence test re-vealed that postrostrum trajectories of Alligatoridae and Crocodylidae have the same slope, but statistically dif-ferent elevation. However, under PGLS, even the alliga-torids postrostrum does not appear influenced by evolu-tionary allometry, thus appearing strongly influenced by the phylogeny.
The regression between the size-free data of perfor-mance and shape (Fig. 7d) is significant for the entire clade. The taxon with the best performance (i.e. the low-est sigma) is C. latirostris, followed by M. niger and Cayman yacare Daudin, 1802. For Alligatoridae this re-lationship is not significant under OLS and PGLS, while it is for Crocodilydae (P = 0.0019 and 0.0029, respec-tively).
DISCUSSIONComparative methods coupled with investigations on
morphological integration and modularity may offer a more complete perspective of crocodilian macroevolu-tionary dynamics than previous approaches did.
Crocodilian cranial variation has been considered as an example of convergent evolution (similar skull morphologies as a response to similar ecological/func-tional constraints) in distantly related species (Iordan-sky1964; Langston 1973; Busbey 1995; Brochu 2001; Pierce et al. 2008; Sadleir & Makovicky 2008; Piras et al. 2009). This convergent evolution mainly concerns the rostrum as the outcome of functional and ecologi-cal specializations, consistent with the fact that the ros-tral module explains much of the total skull shape varia-tion in crocodilians. Thus, we hypothesize that function (adaptation to current conditions, to past conditions, or both) is the driving force explaining patterns of skull shape evolution in crocodilians. The analyses of the en-tire skull shape variation show that the Crocodylidae oc-cupy a significantly larger portion of the morphospace than Alligatoridae (Fig. 5). Both broad-snouted and long-snouted combinations occur within the Crocody-

512
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
P. Piras et al.
© 2013 International Society of Zoological Sciences, Institute of Zoology/ Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
lidae, whereas the rostrum is invariably broad in Alli-gatoridae. When only rostral morphology is analyzed, the morphospace difference between Alligatoridae and Crocodylidae is only marginally significant (P = 0.08). This is due to local differences mainly associated with the shape of the nasals (which in the genus Alligator completely bisect the external nares). Thus, from a gen-eral point of view, shape variation in rostrum morphol-ogy, disregarding local morphological changes, is larger in Crocodylidae than in Alligatoridae. The rostrum is, in fact, the most plastic structure in the evolution of croco-dilians (Langston 1973; Busbey 1995; Brochu 2001).
In contrast, shared ancestry plays a comparatively more important role in explaining cranial variation of the postrostral shape, particularly in Alligatoridae. The strong influence of phylogeny in the Alligatoridae is herein proved by the results of the ontogenetic conver-gence test on evolutionary allometric trajectory of the postrostrum (which is significant under OLS but not un-der PGLS), and by the pattern of morphospace occupa-tion found between the species of this clade. In postros-tral morphology, the larger morphospace occupation of Crocodylidae reflects the peculiar morphology of O. tet-raspis. This species possesses particularly small supra-temporal fenestrae, and relatively large orbits and infra-temporal fenestrae, which morphologically converge on alligators.
The results also demonstrate that rostral shape affects a significant number of cranial postrostral characters, as pointed out in Sadleir and Makovicky (2008) using a concentrated change test. The effect of rostral variation (from long to short/blunt rostral extremes) on postros-tral cranial morphology is visualized on the first singu-lar vector in PLS analysis (Fig. 7). An important amount of postrostral shape change is linked to rostral variation in the crocodilian skull; specifically, the supratemporal fenestra region, the anterior frontal region, the pterygoid flange and the palatine fenestra relative positions. These should all be regarded as homoplasic features correlated to rostral variation in Crocodylia.
Despite the high degree of rostral variation in the crocodylian evolution, both Crocodylidae and Alliga-toridae express the same pattern of bite forces (Erick-son et al. 2012). The point here is to understand wheth-er the lack of variation in bite forces is the outcome of structural constraints (Seilacher 1970; Cubo et al. 2008) or the outcome of stabilizing selection. The structuralist hypothesis suggests that crocodilian skull architecture may limit maximal muscle length, and so maximum bite force, while the alternative hypothesis suggests that the
lack of variation in maximal bite force is the outcome of stabilizing selection because of the high adaptive value of this function.
Our results suggest that the correlation between the postrostrum and rostrum could be related to the constan-cy of the bite force among extant taxa. Rostral morpho-logical variation is linked to the maximum size of the food items ingested, so that the longirostral morphology (such as in G. gangeticus) accrues to piscivorous spe-cies, while short rostra (such as in Caiman latirostris) are associated with durophagy and with the handling of less agile but stout prey (see also Daniel & McHen-ry 2001; McHenry et al. 2006). This is confirmed in this study by the beam simulation that placed Gavialis and Caiman at the opposite extremes of the size free sig-ma distribution (Fig. 7), and by the differences found in the longirostrine and brevirostrine cranial nipper. In fact, Figure 7d illustrates the relationships between pos-trostrum shape and rostrum performance (once the sig-nificant evolutionary allometric signal evidenced in Fig-ure 7a–c is removed from both datasets), showing that the worst performances are associated with postrostral morphologies with narrow cranial nippers.
Modifications in the reduction of the cranial nipper associated with longirostral morphology suggest a close relationship between such shapes and the maximum di-mensions of food items (which is probably important in an exclusive fish diet). In fact, in opposition to longiros-tral species, generalized forms need to open their oral cavity wide in order to ingest larger food items. This means that different rostral conditions should require visible musculoskeletal reorganization related (in turn) with the pterygoid flange and the quadrate articulation. This, however, does not change the maximum bite force exerted by different species. Maximum prey size could not be the sole factor influencing mouth opening. Bite frequency must play an important role when contrasting 2 different rostral morphologies. In piscivorous species the mouth opens and closes rapidly to trap fish. Non-pi-scivorous species take prey in a variety of locations, fre-quently only against the resistant of air, so wider open-ing is also easier to accomplish. Similar conclusions are suggested by Walmsley et al. (2013), who perform finite element simulations and morphometrics assessments on different mandible morphologies.
Accordingly, the hypothesis suggesting a limit to the maximum muscle length and the maximum bite force is supported. Notice that the cranial musculature in croco-dilians is mediated by the presence of a cartilago tran-siliens. This has been recently characterized as a ses-

513
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Morphological integration in crocodilian skull
© 2013 International Society of Zoological Sciences, Institute of Zoology/ Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
amoid-like structure providing significant attachment sites for m. pseudotemporalis superficialis dorsally and m. intramandibularis ventrally, with some contribut-ing attachments from other surrounding muscles (Tsai & Holliday 2011). A sesamoid is able to increase a mus-cle’s mechanical advantage by lengthening the moment arm of the tendon, and should be considered in under-standing the pattern of bite forces in crocodylids and alligatorids.
Nonetheless, the lack of variation in maximal bite force as an outcome of stabilizing selection, because of the high adaptive value of this function, would also be sustained when the occupation of an empirical chro-no-morphospace is taken into account. Rostrum analysis suggests that the functional role of the crocodilian ros-trum is strongly constrained by a preferential pattern of variation involving either lengthening or shortening of the structure. Such a pronounced functional trait might be related to the transformation towards broad-rostral skulls operating throughout the evolutionary history of Crocodylia. Considering that, to date, no longirostral Alligatoridae have been found in the fossil record (Bro-chu 2001; Piras et al. 2009), adding new fossils should not change our results.
The use of modules is decisive in understanding how phylogenetic inertia might explain cranial variation in the Crocodylia. Piras et al. (2009) find substantial dif-ferences in the rostral morphological variation of Allig-atoridae and Crocodylidae. While crocodylids showed a stronger functional component of variation in their skulls, alligatorid variation was better explained as an outcome of phylogenetic inertia. Despite crocodylid dis-parity covering a larger portion of the total morphospace in PCA, alligatorids segregate into phylogenetically dis-crete groups, suggesting that skull shape is more conser-vative in alligators.
If, as noted in Piras et al. (2009), alligatorid rostral shape variation also has a strong phylogenetic compo-nent, there must be great covariation between rostral and postrostral modules, and, thus, great morphological integration in alligators, as we found here. Based on dif-ferent data, Sadleir and Makovicky (2008) reach a sim-ilar conclusion. They indicate that characters correlated with the blunt ecomorphotypes (i.e. in some alligato-rids) do not concentrate in the snout but are distributed throughout the skull (which means in the postrostrum as well).
This is confirmed here by RV coefficients; we found strong evidence that the rostrum is the main driving force in entire skull shape change and that it has the
smallest covariation with the postrostrum in Crocodyli-dae, suggesting higher modularity in this clade as com-pared to Alligatoridae. This suggests a stronger mor-phological decoupling between the 2 cranial modules in crocodylids.
In fact, the RV coefficient reveals higher covariation between the entire shape and the rostrum than between the 2 modules in Crocodylidae. Alligatoridae show, in-stead, a higher integration (Table 2). It is worth noting that in Alligatoridae the relationship between the en-tire shape and the rostrum is quite high and significant as well. This means that even in Alligatoridae the en-tire shape change is driven more by the rostrum than by the postrostrum. This singular condition indicates a gen-uine macroevolutionary difference. In placing the high modularity of the crocodylid skull in a macroevolu-tionary framework, it might be suggested that the pat-tern of rostrum variation at clade level is related to the outcome of a higher functional efficiency (Piras et al. 2009). Crocodylids would be characterized by a higher value of evolvability (defined as the capacity to generate heritable, selectable phenotypic variation [Eble 2005; Kirschner & Gerhart 2005]) than alligatorids.
Entire skull shape and rostrum evolutionary allome-tries show a similar pattern; the Crocodylia clade is sig-nificantly affected by evolutionary allometry as well as the Crocodylidae, while Alligatoridae does not present an evolutionary allometry for these configurations. Pos-trostra, in contrast, seem to maintain a similar evolu-tionary allometric trajectory between the 2 clades, dif-fering only in elevation (Fig. 7). This indicates that the postrostrum is more conservative. Therefore, evolution-ary allometry impacts the skull evolution of the entire configuration as well as that of its functional modules. As for the postrostrum, larger species are the generalists while larger rostra are confined to longirostral forms. In all configurations Paleosuchus spp., a very short snout-ed form, presents the smallest centroid size. In the light of these results we argue that size is, again, one of the most rapid ways to promote shape change (Meloro & Raia 2010) and it could represent a trajectory of least evolutionary resistance within crocodylians (Marroig & Cheverud 2005).
ACKNOWLEDGMENTSPaolo Piras had access to the collections of the Nat-
ural History Museum in London, the Institut Royal des Sciences Naturelles in Bruxelles and the Museum d’His-toire Naturelle in Paris thanks to the SYNTHESYS pro-gram, and to the collection of the Field Museum in Chi-

514
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
P. Piras et al.
© 2013 International Society of Zoological Sciences, Institute of Zoology/ Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
cago thanks to the Visiting Scholarship Program. Funds from the Geological Science Department of ‘Roma Tre’ University made possible the visit to the American Mu-seum of Natural History, New York. Part of this research was conducted during a postdoctoral visit by Paolo Pi-ras, supervised by Jorge Cubo, to the Université Pierre et Marie Curie. Funds were also provided by Project CGL-2009-11838 from the Spanish Ministerio de Cien-cia e Innovacion Cientifica, Spain.
REFERENCESAdams DC, Rohlf FJ, Slice DE (2004). Geometric mor-
phometrics: ten years of progress following the ‘rev-olution’. Italian Journal of Zoology 71, 5–16.
Bookstein FL (1986). Size and shape spaces for land-mark data in two dimensions. Statistical Science 1, 181–222.
Bookstein FL (1991). Morphometric Tools for Land-mark Data. Geometry and Biology. Cambridge Uni-versity Press, New York.
Brochu CA (1999). Phylogeny, systematics and histori-cal biogeography of Alligatoroidea. Society of Verte-brate Paleontolgy Memories 6, 9–100.
Brochu CA (2001). Crocodilian snouts in space and time: phylogenetic approaches toward adaptive radia-tion. American Zoologist 41, 564–85.
Brochu CA (2003). Phylogenetic approaches toward crocodilian history. Annual Review of Earth Plane-tary Science 31, 357–97.
Brochu CA (2007a). Morphology, relationships and bio-geographic significance of an extinct horned croco-dile (Crocodylia, Crocodylidae) from the Quaterna-ry of Madagascar. Zoological Journal of the Linnean Society 150, 835–63.
Brochu CA (2007b). Systematics and taxonomy of Eo-cene tomistomine crocodylians from Britain and Northern Europe. Palaeontology 50, 917–28.
Busbey AB (1995). The structural consequences of skull flattening in crocodilians. In: Thomason JJ, ed. Func-tional Morphology in Vertebrate Paleontology. Cam-bridge University Press, New York. pp. 173–92.
Cleuren J, de Vree F (1992). Kinematics of the jaw and hyolingual apparatus during feeding in Caiman croc-odilus. Journal of Morphology 212, 141–54.
Cubo J, Legendre P, de Ricqlès A et al. (2008). Phyloge-netic, functional, and structural components of vari-
ation in bone growth rate of amniotes. Evolution & Development 10, 217–27.
Daniel WJT, McHenry C (2001). Bite force to skull stress correlation: Modelling the skull of Alligator mississippiensis. In: Grigg C, Seebacher F, Franklin CE, eds. Crocodilian Biology and Evolution (G). Sur-rey Beatty & Sons, New South Wales, Australia, pp. 135–43.
Delfino M, de Vos J (2010). A revision of Dubois croc-odylians, Gavialis bengawanicus and Crocodylus os-sifragus, from the Pleistocene Homo erectus beds of Java. Journal of Vertebrate Paleontology 30, 427–41.
Desdevises Y, Legendre P, Azouzi L, Morand S (2003). Quantifying phylogenetically structured environmen-tal variation. Evolution 57, 2647–52.
Dray S, Dufour AB, Chessel D (2007). The ade4 pack-age-II: two-table and K-table methods. R News 7, 47–52.
Eble G (2005). Morphological modularity and mac-roevolution: conceptual and empirical aspects. In: Callebaut W, Rasskin-Guttman D, eds. Modulari-ty: Understanding the Development and Evolution of Natural Complex Systems. MIT Press, Cambridge and London, pp. 221–38.
Erickson GM, Lappin AK, Vliet K (2003). The ontog-eny of bite-force performance in American alligator (Alligator mississippiensis). Journal of Zoology 260, 317–27.
Erickson GM, Lappin AK, Vliet K, Brueggen J, Kled-zik D, Webb G (2005). Comparative bite-force per-formance of extant and fossil crocodilians: a window into evolutionary feeding biomechanics. Journal of Vertebrate Paleontology 26A, 46.
Erickson GM, Gignac PM, Steppan SJ et al. (2012). Insights into the ecology and evolutionary suc-cess of crocodilians revealed through bite-force and tooth-pressure experimentation. PLOS ONE 7, e31781.
Escoufier Y (1973). Le traitement des variables vectori-elles. Biometrics 29, 751–60.
Gatesy J, Amato G, Norell M, Desalle R, Hayashi C (2003). Combined support for wholesale taxic ata-vism in Gavialine Crocodilians. Systematic Biology 52, 403–22.
Goodall CR (1991). Procrustes methods in the statistical analysis of shape. Journal of the Royal Statistical So-ciety series B-statistical methodology 53, 285–339.

515
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Morphological integration in crocodilian skull
© 2013 International Society of Zoological Sciences, Institute of Zoology/ Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Gonzalez I, Dejean S (2012). CCA: Canonical correla-tion analisys. R package version 1.2. [Cited 13 Sep 2013.] Available from URL: http://CRAN.R-project.org/package=CCA
Harmon L, Weir J, Brock C, Glor R, Challenger W, Hunt G (2009). geiger: Analysis of evolutionary di-versification. R package version 1.99-3. [Cited 13 Sep 2013.] Available from URL: http://CRAN.R-project.org/package=geiger
Iordansky NN (1964). The jaw muscles of the croco-diles and some relative structures of the crocodilian skull. Anatomischer Anzeiger 155, 256–80.
Janke A, Gullberg A, Hughes S, Aggarwal RK, Arnason U (2006). Mitogenomic analyses place the gharial (Gavialis gangeticus) on the crocodile tree and pro-vide pre-K/T divergence times for most crocodilians. Journal of Molecular Evolution 61, 620–6.
Kirschner MW, Gerhart JC (2005). The Plausibility of Life. Resolving Darwin’s Dilemma. Yale University Press, New Heaven, London.
Klingenberg CP (2008). Morphological integration and developmental modularity. Annual Review of Ecolo-gy, Evolution and Systematics 39, 115–32.
Langston W (1965). Fossil crocodilians from Colombia and the Cenozoic history of the Crocodilia in South America. University of California Publications in Geological Sciences 53, 1–157.
Langston W (1973). The crocodilian skull in histori-cal perspective. In: Gans C, Parsons T, eds. Biology of the Reptilia, Vol. 4. Academic Press, London, pp. 263–84.
Marquez EJ (2008). A statistical framework for testing modularity in multidimensional data. Evolution 62, 2688–708.
Marroig G, Cheverud JM (2005). Size as a line of least evolutionary resistance: diet and adaptive morpho-logical radiation in new world monkeys. Evolution 59, 1128–42.
McAliley LR, Willis RE, Ray DA, Scott-White P, Bro-chu CA, Densmore III LD (2006). Are crocodiles re-ally monophyletic? Evidence for subdivisions from sequence and morphological data. Molecular Phylo-genetics and Evolution 39, 16–32.
McHenry CR, Clausen D, Daniel WJT, Meers MB, Pendharkar A (2006). Biomechanics of the rostrum in Crocodilians: a comparative analysis using finite-el-ement modelling. The Anatomical Record 288, 827–49.
Meloro C, Raia P (2010). Cats and dogs down the tree: the tempo and mode of evolution in the lower carnas-sial of fossil and living carnivora. Evolutionary Biol-ogy 37, 177–86.
Oaks JR (2011). A time-calibrated species tree of croco-dylia reveals a recent radiation of the true crocodiles. Evolution 65, 3285–97.
Oksanen J, Kindt R, Legendre P, O’Hara RB (2008). Vegan: Community Ecology Package. [Cited 25 Sep 2013.] Available from URL: http://cran.r-project.org/
Pierce SE, Angielczyk KD, Rayfield EJ (2008). Patterns of morphospace occupation and mechanical perfor-mance in extant crocodilian skulls: a combined geo-metric morphometric and finite element modelling approach. Journal of Morphology 269, 840–64.
Piras P, Teresi L, Buscalioni AD, Cubo J (2009). The shadow of forgotten ancestors differently constrains the fate of Alligatoroidea and Crocodyloidea. Global Ecology and Biogeography 18, 30–40.
Piras P, Colangelo P, Adams DC et al. (2010). The Gavi-alis–Tomistoma debate: the contribution of skull on-togenetic allometry and growth trajectories to the study of crocodylian relationships. Evolution and De-velopment 12, 568–79.
Piras P, Salvi D, Ferrara G et al. (2011). The role of post-natal ontogeny in the evolution of phenotypic diversity in Podarcis lizards. Journal of Evolutionary Biology 24, 2705–20.
Rohlf FJ (2001). Comparative methods for the analy-sis of continuous variables: geometric interpretations. Evolution 55, 2143–60.
Rohlf FJ, Corti M (2000). Use of two-block partial least-square to study covariation in shape. Systematic Biology 49, 740–53.
Rohlf FJ (2006). A comment on phylogenetic correc-tions. Evolution 60, 1509–15.
Sadleir RW, Makovicky PJ (2008). Cranial shape and correlated characters in crocodilian evolution. Jour-nal of Evolutionary Biology 21, 1578–96.
Seilacher A (1970). Arbeitskonzept zur konstruk-tions-morphologie. Lethaia 3, 393–6.
Tsai HP, Holliday CM (2011). Ontogeny of Alligator cartilago transiliens and its significance for sauropsid jaw muscle evolution. PLOS ONE 6, e24935.
Walmsley CW, Smits PD, Quayle MR et al. (2013). Why the long face? The mechanics of mandibular symphysis proportions in crocodiles. PLOS ONE 8, e53873.

516
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
P. Piras et al.
© 2013 International Society of Zoological Sciences, Institute of Zoology/ Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Willis RE, McAliley LR, Neeley ED, Densmore LD (2007). Evidence for placing the false gharial (Tomi-stoma schlegelii) into the family Gavialidae: infer-ences from nuclear gene sequences. Molecular Phy-logenetics and Evolution 43, 787–94.
Zelditch ML, Swiderski DL, Sheets HD, Fink WL (2004). Geometric Morphometrics for Biologists: A Primer. Elsevier, Academic Press. San Diego, CA.
Zelditch ML, Swiderski DL, Sheets HD (2012). Geo-metric Morphometrics for Biologists: A Primer. 2nd ed. Elsevier, Academic Press. San Diego, CA.
SUPPORTING INFORMATIONAdditional supporting information may be found in
the online version of this article at the publisher’s web-site.
Table S1 Material used for this studyTable S2 R function genpgls() written by P. Piras Figure S1 Scatterplots as the results of GM analyses
(Fig. 5) with groups identified at the specific level. Please note: Wiley is not responsible for the content
or functionality of any supporting information supplied by the authors. Any queries (other than missing content) should be directed to the corresponding author for the article.
Copyright © 2022 FDOKUMEN




















