
Molecular basis for the development of individual differences in the hypothalamic-pituitary-adrenal...
27
Cellular and Molecular Neurobiology, Vol. 13, No. 4, 1993 Molecular Basis for the Development of Individual Differences in the Hypothalamic- Pituitary-Adrenal Stress Response Michael J. Meaney, l'z Seema Bhatnagar, 1 Josie Diorio, 1 Sylvie Larocque, ~ Darlene Francis, ~ Dajan O'Donnell, ~ Nola Shanks, ~ Shakti Sharma, ~ James Smythe, 1 and Victor Viau ~ Received February 20, 1993; accepted May I0, 1993 KEY WORDS: hypothalamic-pituitary-adrenalresponse; stress; rats; handling. SUMMARY 1. Several years ago, investigators described the effects of infantile handling on the development of hypothalamic-pituitary-adrenal (HPA) responses to stress in the rat. Rat pups exposed to brief periods of innocuous handling early in life showed reduced HPA responses to a wide variety of stressors, and the effect persists throughout the life of the animal. These effects are robust and provide an excellent model for understanding how early environmental stimuli, which are external to the organism, alter neural differentiation and, thus, neuroendocrine responsivity to stress. 2. This paper reviews the endocrine mechanisms affected by early handling and our current understanding of the neural transduction of environmental events and their effects at the level of the target neurons (in the hippocampus and frontal cortex). 3. In brief, handling serves to increase glucocorticoid receptor gene trans- cription, increasing sensitivity to glucocorticoid negative feedback regulation and, thus, altering the activity within hypothalamic corticotropin-releasing factor/vasopressin neurons. Together these changes serve to determine neuroen- docrine responsivity to stress. 1Developmental Neuroendocrinology Laboratory, Douglas Hospital Research Centre, Departments of Psychiatry, and Neurology and Neurosurgery, McGill University, Montreal, Canada. ZTo whom correspondence should be addressed at Developmental Neuroendocrinology Laboratory, Douglas Hospital Research Centre, 6875 Boul. LaSalle, Montreal, Quebec H4H 1R3, Canada. 321 0272-4340/93/ll80(I-0321506.00/0 (~) 1993 Plenum Publishing Corporation
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Molecular basis for the development of individual differences in the hypothalamic-pituitary-adrenal...

Cellular and Molecular Neurobiology, Vol. 13, No. 4, 1993
Molecular Basis for the Development of Individual Differences in the Hypothalamic- Pituitary-Adrenal Stress Response
Michael J. Meaney, l'z Seema Bhatnagar, 1 Josie Diorio , 1 Sylvie Larocque, ~ Darlene Francis, ~ Dajan O'Donnel l , ~ Nola Shanks, ~ Shakti Sharma, ~ James Smythe, 1 and Victor Viau ~
Received February 20, 1993; accepted May I0, 1993
KEY WORDS: hypothalamic-pituitary-adrenal response; stress; rats; handling.
SUMMARY
1. Several years ago, investigators described the effects of infantile handling on the development of hypothalamic-pituitary-adrenal (HPA) responses to stress in the rat. Rat pups exposed to brief periods of innocuous handling early in life showed reduced HPA responses to a wide variety of stressors, and the effect persists throughout the life of the animal. These effects are robust and provide an excellent model for understanding how early environmental stimuli, which are external to the organism, alter neural differentiation and, thus, neuroendocrine responsivity to stress.
2. This paper reviews the endocrine mechanisms affected by early handling and our current understanding of the neural transduction of environmental events and their effects at the level of the target neurons (in the hippocampus and frontal cortex).
3. In brief, handling serves to increase glucocorticoid receptor gene trans- cription, increasing sensitivity to glucocorticoid negative feedback regulation and, thus, altering the activity within hypothalamic corticotropin-releasing factor/vasopressin neurons. Together these changes serve to determine neuroen- docrine responsivity to stress.
1Developmental Neuroendocrinology Laboratory, Douglas Hospital Research Centre, Departments of Psychiatry, and Neurology and Neurosurgery, McGill University, Montreal, Canada.
ZTo whom correspondence should be addressed at Developmental Neuroendocrinology Laboratory, Douglas Hospital Research Centre, 6875 Boul. LaSalle, Montreal, Quebec H4H 1R3, Canada.
321 0272-4340/93/ll80(I-0321506.00/0 (~) 1993 Plenum Publishing Corporation

322 Meaney et al.
INTRODUCTION
The adrenal glucocorticoids and catecholamines comprise a frontline of defense for mammalian species under conditions which threaten homeostasis (conditions commonly referred to as "stress," and the provocative stimuli as "stressors"). These hormones serve as major regulators of carbohydrate and lipid metabolism, cardiovascular tone, muscle function, immunocompetence, and behavior. The adrenal glucocorticoids represent the end product of the hypothalamic-pituitary- adrenal (HPA) axis. Under most conditions, this axis lies under the dominion of specific peptides secreted by neurons located in the paraventricular nuclei (PVN) of the hypothalamus. Most notable among these peptides are corticotropin- releasing factor (CRF) and arginine vasopressin (AVP). The neurons of the PVN are thus the major target for both the activational effects of the neural signals associated with stress and the inhibitory effects associated with glucocorticoid negative feedback. The nature of the HPA response to stress occurs as a function of the integration of these signals at the level of the CRF/AVP neurons in the hypothalamus.
The ability of the organism to mount an endocrine response to stress rapidly is imperative. Under stressful conditions, adrenalectomized animals are unable to sustain the necessary cardiovascular tone and blood glucose levels and, thus, perish. However, inappropriate or exaggerated HPA activity is also threatening to the health of the organism. Prolonged exposure to elevated glucocorticoid levels results in a general suppression of anabolic processes, muscle atrophy, decreased sensitivity to insulin and a risk of steroid-induced diabetes, hyperten- sion, hyperlipidemia, hypercholesterolemia, arterial disease, amenorrhea, and impotency, and the impairment of growth and tissue rapair, as well as immunosuppression (Brindley and Rolland, 1989; Munck et al . , 1984). In addition, elevated glucocorticoid levels are also associated with psychopathology, neuronal damage and impaired cognitive function (see Meaney et al . , 1993a; Sapolsky, 1990). Hence, an efficient response to stress is one that is rapidly immobilized in the presence of a threat and then effectively "turned off" once the threatening condition is no longer present.
The development of the HPA response is shaped by events occurring early in life. These effects are surprisingly robust and account, at least in part, for the individual differences that occur in neuroendocrine responses to stress. There is now considerable evidence for the idea that these early environmental factors ultimately influence activity at the level of the hypothalamic CRF/AVP neurons, either directly or indirectly via their effects on systems which mediate the inhibitory signals associated with glucocorticoid negative feedback. These changes persist throughout the life of the animal and are accompanied by altered endocrine responses to stress. Such developmental effects likely represent one way in which early life events can predispose an individual to pathology in later life. Thus, early environment can determine vulnerability to pathology in later life by determining the efficiency with which the animal responds to stress. At another level, these studies also provide a model for understanding the processes by which environmental stimuli can regulate development by altering gene

HPA Development 323
transcription. At this point, we can begin to understand what is meant by the " x " in the maxim "Genotype x Environment = Phenotype" under conditions where the relevant environmental stimuli are external to the organism and the phenotype refers to the animal's endocrine responses to stress.
THE ADRENOCORTICAL RESPONSE TO STRESS
The HPA axis, as described by Selye (1950), is highly responsive to stress. Neural signals associated with the stressor are transduced into endocrine responses via their effects at the level of the hypothalamus (see Fig. 1). Thus, the secretion of CRF and AVP from PVN neurons into the portal system of t h e anterior pituitary during stress causes an increase in the release of adrenocor- ticotropin (ACTH) from pituitary corticotropes into circulation (e.g., Antoni, 1984; Gibbs, 1986; Plotsky, 1987; Linton et al., 1985; Nakane et al., 1985; Rivier and Vale, 1983; Rivier et al., 1982). The elevated ACTH levels, in turn, stimulate an increase in the synthesis and release of adrenal glucocorticoids. The highly catabolic glucocorticoids produce lypolysis, increasing the level of free fatty acids, glycogenolysis, increasing blood glucose levels, and in protein catabolism, which increases amino acid availability as substrates for gluconeogenesis, further increasing blood glucose levels (Brindley & Rolland, 1989; Munck et al., 1984). Together, these actions assist the organism under stressful conditions, at least in part; by increasing the availability of energy substrates. The glucocorticoids also suppress immunological responses (Munck et al., 1984), protecting against the occurrence of inflammation at a time when mobility may be important to the animal.
However, for the reasons stated above, once the stressor is terminated, it is very clearly in the animal's best interest to terminate the HPA stress response. The efficacy of this process is determined by the ability of the glucocorticoids to inhibit subsequent ACTH release (i.e.glucocorticoid negative feedback; see Fig.
Fig. 1. A scheme describing the hypothalamic- pituitary-adrenal response to stress. Neural sig- nals associated with the stress or impinge upon hypothalamic (and extrahypothalamic sites) to stimulate the release of CRF and/or AVP stores from hypothalamic neurons into the hypophysial portal system of the anterior pituitary. These peptides stimulate the release of ACTH from corticotropes into circulation and ACTH, in turn, stimulates the release of adrenal glucocor- ticoids (primarily eortisol in primates and cor- cisoterone in rodents). Activity within the axis is regulated by a negative feedback loop whereby the end product of the axis, the adrenal gluco- corticoids, acts at various levels to inhibit CRF/AVP synthesis and release.
Extrahypotbalamic Sites ~ --- (_)
iHypotha'ousl (CRF/AVP)
ACTH (+)
/ - - Glucocorticoids - -
(-)
(9

324 Meaney et al.
1). The HPA axis is a classical closed-loop feedback system where the end product of the axis, the adrenal glucocorticoids, feedback onto higher structures in the axis to inhibit activity. Thus, circulating glucocorticoids feedback onto the pituitary and specific brain regions to inhibit the release of ACTH from the anterior pituitary cells (Dallman et al., 1987; Jones et al., 1982; Keller-Wood and Dallman, 1984; Plotsky and Vale, 1984; Plotsky et al. , 1987; Van Loon and DeSouza, 1987). The focus for glucocorticoid negative feedback inhibition is the population of CRF and CRF/AVP neurons in the parvocellular region of the PVN. Thus, glucocorticoids serve to decrease mRNA levels (e.g., Beyer et al. , 1988; Lightman and Young, 1987; Swanson and Simmons, 1989; Wolfson et al. , 1985; Young et al. , 1987), content (e.g., Kiss et al., 1984; Merchenthaler et al. , 1983; Swanson et al., 1983), and release (Plotsky and Sawchenko, 1987) of both CRF and AVP. These effects occur either directly on CRF and CRF/AVP neurons or via effects on other brain regions which, in turn, regulate activity within the PVN neurons. Thus, the pituitary and the hypothalamus are not the only relevant brain sites for glucocorticoid feedback effects.
There is now considerable evidence for the importance of extrahypothalamic regions in the inhibition of CRF synthesis and HPA activity. Most notable among these regions is the hippocampus (Jacobson and Sapolsky, 1991). In the rat, hippocampal lesions or ablations are associated with elevated corticosterone (B; the principal glucocorticoid in rodents) levels under basal, stress, and poststress conditions (e.g., Feldman and Conforti, 1976, 1980; Fischette et al. , 1980; Sapolsky et al. , 1984a; Wilson et al., 1980). Moreover, hippocampectomized animals show reduced suppression of ACTH following exogenous glucocorticoid administration (Feldman and Conforti, 1976, 1980) and increased CRF and AVP mRNA levels in the PVN of the hypothalamus (Herman et al. , 1989) and fornix lesions decrease glucocorticoid inhibition over CRF and AVP release in the portal system (Sapolsky et al. , 1990). These findings, together with the fact that the hippocampus is rich in corticosteroid receptors (McEwen et al. , 1986), suggest that this structure is involved in the inhibitory influence of glucocorticoids over adrenocortical activity [see Jacobson and Sapolsky (1991) and McEwen et al. (1986) for reviews of the issues and controversies on this topic].
C O R T I C O S T E R O I D R E C E P T O R S
The uptake of B in rat brain is associated with at least two distinct corticosteroid receptor subtypes (e.g., Beaumont and Fanestil, 1983; Emadian et al. , 1986; McEwen et al., 1986; Funder and Sheppard, 1987; Reul and De Kloet, 1985, 1986; Reul et al. , 1987; Sheppard and Funder, 1987). The mineralocorticoid (or type I) receptor binds in v i t ro to both B and the mineralocorticoids, aldosterone, spironolactone, and RU 26752, with a high affinity and binds the synthetic glucocorticoid, RU 28362, with a very low affinity. In the brain, mineralocorticoid receptor density is highest in the septal-hippocampal system. The glucocorticoid (or type II) receptor, which is far more diffusely distributed throughout the brain, binds B, dexamethasone, and RU 28362 with a high affinity

HPA Development 325
and RU 26752 and aldosterone with a lower affinity. Although both receptors bind B with a high affinity, the K~ of the mineralocorticoid receptor for B ( -0 .5 riM) is lower than that of the glucocorticoid receptor [ -2 .0-5 .0 nM (e.g., Reul and De Kloet, 1985)].
A physiological consequence of this difference in affinity for B is the fact that these receptors then show different rates of ligand occupancy under basal B levels. About 50-90% of the mineralocorticoid sites are occupied under basal B levels [-5/~g/dl (Reul and De Kloet, 1985; Reul et al. , 1987)]. This renders the hippocampal mineralocorticoid receptor relatively insensitive to dynamic varia- tions in B levels above those seen in the resting state. In contrast, the glucocorticoid receptor is highly responsive to dynamic changes in B titers, such as those occurring during the circadian peak or stress (Meaney et al., 1988a; Reul and De Kloet, 1985; Reul et al., 1987). Under conditions of basal circulating B, only about 10-15% of the glucocorticoid receptors are occupied. Stress results in a dramatic increase in the hormone-receptor signal, such that immediately following a 20-min period of restraint about 75% of glucocorticoid receptors are occupied. B injections that mimic the steroid levels seen during stress (-30/~g/d) also result in about 75% occupancy of glucocorticoid receptors. These findings, together with the known negative-feedback efficacy of the synthetic corticoids such as dexamethasone, once thought to bind selectively to the glucocorticoid receptor, suggested that it was this site and not the mineralocorticoid-like receptor that underlies the negative-feedback actions of glucocorticoids.
However, Dallman and her colleagues have recently provided evidence for the involvement of both hippocampal mineralocorticoid and glucocorticoid receptors in the regulation of ACTH levels in rats. In these studies hippocampal implants of both the glucocorticoid receptor antagonist, RU 38486, and the mineralocorticoid receptor antagonist, RU 26752, resulted in elevated levels of plasma ACTH (Bradbury and Dallman, 1989; also see Dallman et al. , 1989). Moreover, Ratka et al. (1989) have found that systemic injections of either antagonist resulted in elevated post-stress B levels in intact rats. These findings suggest that glucocorticoid negative feedback may involve both mineralocorticoid and glucocorticoid receptor sites. In addition, although there is presently considerable emphasis on the role of the hippocampus and hippocampal corticosteroid receptors in glucocorticoid negative-feedback regulation of HPA function, several other corticolimbic sites are likely to be relevant. Kovacs and Makara (1988) reported that glucocorticoid implants into the septum attenuated adrenalectomy-induced increases in plasma ACTH. Moreover, Diorio et al. (1993) have found that corticoid implants into the frontal cortex-cingulate gyrus dampen plasma ACTH responses to restraint stress. Interestingly, these regions contain both mineralocorticoid and glucocorticoid receptor sites (Diorio et al. , 1993; Meaney and Aitken, 1985a).
Probably the strongest support for the importance of the hippocampus in the regulation of HPA activity comes from studies on the role of hippocampal corticosteroid receptor systems. Evidence from a number of models (see Goldman et al., 1978; Jacobson and Sapolsky, 1991; McEwen et al. , 1986; Meaney et al., 1993; Sapolsky and Meaney, 1986) suggests that a decrease in

326 Meaney et al.
hippocampal corticosteroid receptor density is associated with a hypersecretion of B under basal conditions and/or following the termination of stress (i.e., less effective negative feedback). There are decreased levels of hippocampal cortico- steroid receptors binding in the aged rat, lactating rats, and immature rats and
these animals also hypersecrete B under basal and/or stressful conditions. Perhaps the most impressive evidence comes from studies with the vasopressin- deficient, Brattleboro rat (Sapolsky et al., 1984a). These animals show a deficit in corticosteroid receptors in the hippocampus and pituitary and hypersecrete B following stress. The hippocampal receptor deficit is reversed with vasopressin treatment, and so long as the treatment is continued, receptor levels remain elevated and the animals exhibit normal B secretion following stress (Sapolsky et al., 1984a).
THE EFFECT OF HANDLING ON THE HPA RESPONSE TO STRESS
Several years ago Levine, Denenberg, Ader, and others (Ader and Grota, 1969; Hess et al., 1969; Levine, 1957, 1962; Levine et al., 1967) described the effects of postnatal "handling" on the development of behavioral and endocrine responses to stress. As adults, handled rats exhibited attenuated fearfulness in novel environments and a less pronounced increase in the secretion of the adrenal glucocorticoids in response to a variety of stressors. These findings clearly demonstrated that the development of rudimentary, adaptive responses to stress could be modified by environmental events. In addition, the handling paradigm provides a marvelous opportunity to examine how subtle variations in the early environment alter the development of specific neurochemicai systems, leading to stable individual differences in biological responses to stimuli that threaten homeostasis.
The studies described below have been performed using laboratory rats. The following description of developmental landmarks is provided in order to give the reader an impression of the maturational process and its timing in this rodent species. Rat pups are born following a 21 to 22-week period of gestation. The animals remain highly dependent upon the mother for the first 2 weeks of life. During the third week of life the weaning process begins and the animals begin to regulate their own body temperature. Rat pups are weaned at 22 days of age, and puberty begins over the next few weeks of life; generally during days 35-40 in females and 40-50 in males, depending upon the measures used to indicate the onset of sexual maturity. By 70 days of life the animals have reached maturity. In each case, the postnatal manipulations described below are most effective during the first week of life.
The handling procedure usually involves removing rat pups from their cage, placing the animals together in small containers, and 15-20 min later, returning the animals to their cage and their mothers. The manipulation is generally performed daily for the first 21 days of life and the animals are tested as fully mature adults. In response to a wide variety of stimuli, handled (H) rats secrete less B and show a faster return to basal B levels following the termination of

HPA Development 327
stress than do nonhandled (NH) animals (Ader and Grota, 1969; Hess et al. , 1969; Levine, 1957, 1962; Levine et al. , 1967; Meaney et al. , 1988b, 1989). Another way to evaluate hormone data is to calculate the integrated response, which is a measure of hormone secretion over time (i.e., area under the curve/time). This procedure reflects the integrated hormonal signal and for certain hormones, such as ACTH, is physiologically very relevant. The integrated plasma B response to stress (calculated from the prestress to the 120 min poststress time points) is usually 75-100% higher in the NH rats (see Fig. 2). These endocrine differences are apparent as late as 24-26 months of age (Meaney et al., 1988b), indicating that the handling effect persists over the entire life of the animal. The differences in HPA function are not due to changes in adrenal sensitivity to ACTH or in pituitary sensitivity to CRF (Grota, 1975; Meaney et
" IF F- 0 <
1000
750
500
250
- - -o - - - H ~. NH
I I I I I
PRE 10 20 Post 20 Post 40
50
_~- 40
30 C O
I l l
20 O
O o
10
H
I I I I I
PRE 0 30 60 90 120
Fig. 2. Representative data from the authors' lab showing plasma ACTH and corticosterone levels in handled (H) and nonhandled (NH) animals prior to (PRE), during (10- and 20-min time points for ACTH and 0-time point for corticosterone), and at various times the following the termination of a 20-min period of restraint stress.

328 Meaney et al.
al., 1989). Moreover, there are no differences between H and NH animals in the metabolic clearance rate for ACTH or B (Grota, 1975; Meaney et al. , 1989). Rather, the difference lies in the fact that the NH animals show increased hormone secretion both during and following stress.
Young adult H and NH animals do not differ in levels of corticosteroid- binding globulin (CBG), the principal plasma binder for B (Meaney et al. , 1989, 1992) or in free B levels (Meaney et al. , 1992). This finding is of considerable importance since brain uptake of B appears to approximate the non-CBG-bound (free + albumin-bound) portion of the steroid (Partridge et al. , 1983). Thus, differences in total B are likely predictive of differences in brain uptake of the steroid. Interestingly, the handling effects on HPA function are specific to conditions of stress. Young adult NH and NH animals do not differ in basal B levels at any time point over the diurnal cycle (Meaney et al. , 1989, 1992). This finding also indicates that differences in HPA activity observed during stress cannot be accounted for by differences in prestress, basal glucocorticoid levels.
H and NH animals also differ in plasma ACTH responses to stress. As with B, plasma ACTH levels are higher both during and following stress in NH compared with H animals (Meaney et al. , 1989; Viau et al. , 1993). Generally, the integrated plasma ACTH response (prestress to 60 min poststress) is about 50-100% higher in the NH rats. These findings, along with an earlier report (Zarrow et al., 1972) on differences in CRF-like bioactivity between H and NH animals, suggest that the mechanism(s) for differences between N and NH animals is located above the level of the pituitary and may be related to differential sensitivity of CNS negative-feedback processes.
THE EFFECT OF HANDLING ON HPA NEGATIVE-FEEDBACK PROCESSES
Thus, we then examined whether the relative hypersecretion of ACTH and B by NH animals might be related to differences in negative-feedback sensitivity to circulating glucocorticoids between H and NH animals. We (Meaney et al. , 1989) used a classical negative-feedback paradigm based on the finding that high levels of circulating glucorticoids feedback onto the brain and/or pituitary to inhibit subsequent HPA activity (see Keller-Wood and Dallman, 1984; Jones, 1982). Such delayed, negative feedback persists for hours following exposure to elevated glucocorticoid levels. H and NH animals were injected with one of five doses of either B or dexamethasone 3 hr prior to a 20-min period of restraint. Both glucocorticoids were more effective in suppressing stress-induced HPA responses in the H animals (i.e., the IDs0 for both B and dexamethasone was 5-10 times lower in the H animals). These data suggest that H animals are indeed more sensitive to the negative-feedback effects of circulating glucocorticoids on HPA activity.
Since this delayed form of negative-feedback is mediated by the binding of B to soluble intracellular receptors, we measured both mineralocorticoid and glucocorticoid receptor sites in selected brain regions and pituitary of young adult

HPA Development 329
H and NH animals (Meaney and Aitken, 1985b; Meaney et al., 1985a, b, 1987, 1988b, 1989, 1992; Sarrieau et al., 1988). The results of these studies dem- onstrated significant and tissue-specific differences in glucocorticoid receptor binding capacity as a function of handling. H animals show increased gluco- corticoid receptor binding capacity in the hippocampus but not in the septum, amygdala, hypothalamus, or pituitary. The difference in the receptor binding capacity is clearly related to the number of receptor sites, and not to the affinity of the receptor for the 3H-radioligand, RU 28362. Moreover, the difference occurs in glucorticoid receptors but not the mineralocorticoid receptor sites. Finally, using in si tu hybridization with probes selective for either the glucocorti- coid or the mineralocorticoid receptor mRNA, we (Larocque et al., 1992) found that glucocorticoid receptor mRNA expression (grains/cell) was higher through- out the hippocampus of H compared with NH animals. There were no differences in mineralocorticoid receptor mRNA expression. These data indicate that postnatal handling permanently increases glucocorticoid receptor biosynthesis (perhaps by increasing basal transcription rates of the glucocorticoid receptor gene).
The difference in hippocampal glucocorticoid receptor density appears to be related to the more efficient suppression of poststress HPA activity in the H animals. Chronic administration of B results in a 30-45% down-regulation of hippocampal glucocorticoid receptor binding sites that persists for about 7 days following the cessation of treatment (Sapolsky et al., 1984b; Tornello et al. , 1982). The effect is highly specific to the hippocampus, such that receptor binding capacity in the hypothalamus and pituitary is unaffected. This somewhat peculiar effect provided us with an opportunity to examine the importance of differences in hippocampal glucocorticoid receptor density between the H and the NH for HPA responses to stress. Thus, in one experiment H animals were treated for 5 days with B and allowed 2 days for steroid clearance (Meaney et al. , 1989). Hippocampal glucocorticoid receptor density was down-regulated in the H + B animals to levels that were indistinguishable from those of NH animals and significantly less than that of the H + vehicle animals. There were no differences in glucocorticoid receptor density in the hypothalamus or pituitary. When the animals in these groups were exposed to a 20-min period of restraint, we found that the H + B animals, like the NH animals, hypersecreted B 60 and 120 min poststress in comparison to the H + vehicle, control animals. These data suggest that the differences in negative-feedback efficiency and in HPA responses to stress between H and NH are related to the differences in hippocampal glucocorticoid receptor density. Thus, the chronic B treatment reversed both the handling-induced increase in hippocampal glucorticoid receptor binding capacity and the difference in poststress HPA activity.
It appears that the increase in glucocorticoid receptor sites in the hippocam- pus is a critical feature for the handling effect on HPA function. The increase in receptor density appears to increase the sensitivity of the hippocampus to circulating glucocorticoids, enhancing the efficacy of negative-feedback inhibition over HPA activity and serving to reduce poststress secretion of ACTH and B in H animals.

330 Meaney et al.
The effects of postnatal handling on HPA negative feedback likely involves glucocorticoid receptor differences in at least one other region. Handling also increases glucocorticoid receptor density in the frontal cortex (Meaney et al., 1985b). We have recently provided evidence for the role of the prefrontal cortex in the regulation of stress-induced HPA activity (Diorio et al., 1993). Medial prefrontal cortex lesions produced increased levels of ACTH and B both during and following the termination of stress. B implants directly into this region produce a 40-50% decrease in stress-induced ACTH and B levels. Interestingly, these effects are apparent only with more moderate stimuli, in this case restraint. Neither the medial prefrontal cortex lesions nor B implants into this region had any effect on ACTH or B levels observed using ether stress; a more severe stressor associated with two to three times higher levels of ACTH. Moreover, these effects were observed only during or following stress; neither treatment altered basal ACTH or B levels at any point over the diurnal cycle. These findings suggest that the handling effect on HPA function may involve altered glucocorticoid receptor density in the frontal ocrtex.
THE NATURE OF THE GLUCOCORTICOID NEGATIVE FEEDBACK SIGNAL
In recent studies we have provided a more detailed description of plasma ACTH responses to ether and restraint stress in H and NH animals (Viau et al., 1993). The results of these studies have shown that very shortly following the onset of stress, plasma ACTH levels are higher in the NH rats. While ACTH level rise in both groups, this increase is greater in the NH animals. Thus, the integrated (hormone levels x time) plasma ACTH response in the NH rats is generally about twofold greater than in the H animals. At this point it is important to note that H and NH animals do not differ in glucocorticoid fast feedback sensitivity; the inhibitory signal associated with the rapidly increasing levels of plasma B that occur during the first few minutes of stress (Viau et al., 1993).
These data raise an interesting set of problems. First, assuming that differences in fast feedback are not relevant here, the other important negative feedback signal in determining the magnitude of the plasma ACTH response to stress is that associated with basal glucocorticoid levels. Moreover, while we have previously shown that basal B levels do not differ in H vs NH rats, there are differences in glucocorticoid receptor levels in brain regions known to regulate HPA activity. Since the increased receptor density confers a greater sensitivity to B, it remains possible that the basal glucocorticoid feedback signal might be stronger in the H rats despite the fact that basal hormone levels are comparable in H and NH rats.
The second problem concerns the differences in poststress levels of HPA activity. H and NH rats differ in poststress plasma levels of B; Nh animals show elevated B levels for a considerably longer period of time. Thus, the termination of the adrenocortical response to stress in the H animals occurs more efficiently

HPA Development 331
than in NH animals. We assumed that this difference is associated with the differences in negative feedback sensitivity between H and NH rats and that the relevant signal was the elevated B levels achieved during stress. Thus, we believed that the high levels of B served to provide a strong negative feedback signal which, in turn, inhibited HPA activity once the drive associated with the stressor was terminated. However, the adrenal is known to respond to the integrated ACTH signal. The integrated ACTH level determines not only the peak in B secretion, but also the duration of the response; higher levels of ACTH results not only in greater levels of B, but also in a longer period of elevated B. Therefore, the poststress differences in plasma B that are observed in the NH animals could occur in response to the amount of ACTH secreted during stress.
Thus, one very interesting question that emerges here concerns the nature of the relevant glucocorticoid feedback signal. There are at least two obvious signals: the basal B signal prior to stress and the signal associated with the elevated B levels occurring during stress. Stress-induced elevations in B could serve as the signal for the termination of the stress response, once the activational effect of the stress has been removed. Alternatively, the increase in plasma ACTH could be associated with differences in the tonic negative feedback signal associated with basal B levels.
In order to examine this question we (Viau et al. , 1993) compared ACTH responses to restraint in H and NH (1) adrenalectomized animals, which lack any glucocorticoid negative-feedback signal, (2) adrenalectomized animals provided the equivalent of a basal B signal (adrenalectomized +B) but lacking the negative-feedback signal associated with stress-induced in increases in B, and (3) intact, sham-operated animals which possess both basal and stress-induced glucocorticoid signals. B replacement was achieved by implanting fused pellets of B under the skin of the animal; a procedure which provides a constant level of plasma glucocorticoids (i.e., animals which possess a basal glucocorticoid signal but which are unable to mobilize a glucocorticoid stress response). These animals were studied 5 days following adrenalectomy. At this time the HPA axis has generally stabilized in adrenalectomized + B animals (see Dallman et al., 1987). The B pellets provided circulating hormone levels of about 5/,g/dl, equivalent to the integrated basal level of B over the diurnal cycle. Adrenalectomy results in a dramatic increase in pituitary ACTH release and in the synthesis of proopiomela- nocortin (POMC), which is the precursor for ACTH synthesis. Basal plasma ACTH and anterior pituitary POMC mRNA levels were substantially increased by adrenalectomized in both H and NH rats and these effects were effectively reversed with B pellet implants in both groups of animals. Thus, the B pellet was equally effective in reversing the effects of adrenalectomized on basal HPA activity in H and NH rats.
There were no differences in the plasma ACTH responses to restraint in adrenalectomized/NH vs. adrenalectomized/H animals. This was an important finding, for it suggests that in the absence of a glucocorticoid negative feedback signal, ACTH responses to stress are comparable in H and NH rats, thus confirming that the differences in HPA responses to stress observed in intact H and NH animals are associated with differences in glucocorticoid negative
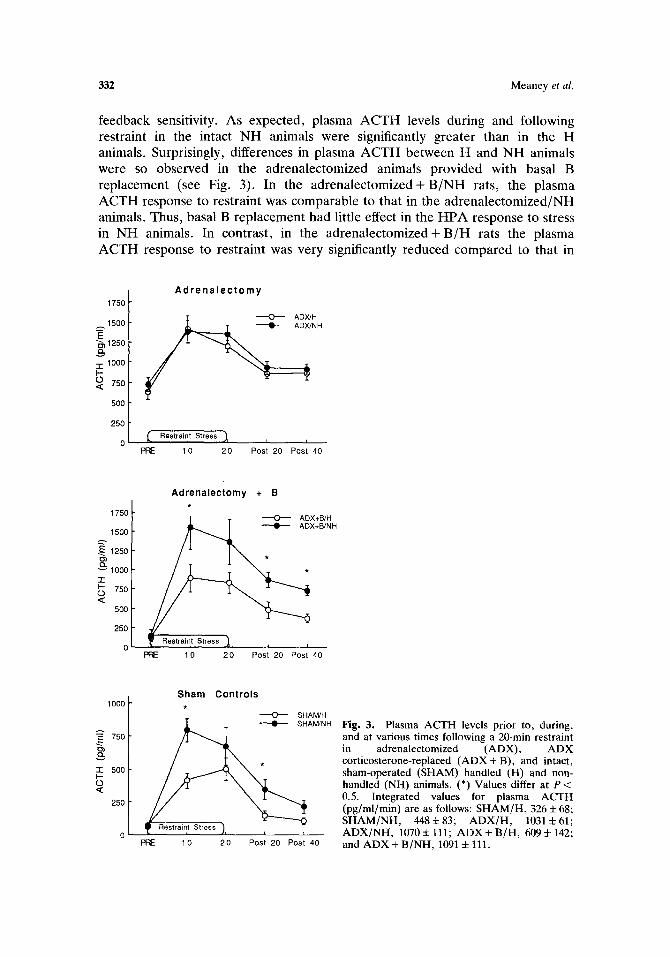
332 Meaney et al,
feedback sensitivity. As expected, plasma ACTH levels during and following restraint in the intact NH animals were significantly greater than in the H animals. Surprisingly, differences in plasma ACTH between H and NH animals were so observed in the adrenalectomized animals provided with basal B replacement (see Fig. 3). In the adrenalectomized+ B/NH rats, the plasma ACTH response to restraint was comparable to that in the adrenalectomized/NH animals. Thus, basal B replacement had little effect in the HPA response to stress in NH animals. In contrast, in the adrenalectomized+ B/H rats the plasma ACTH response to restraint was very significantly reduced compared to that in
1750
1 5 0 0
~ 1 2 5 0
"r- 1000 I-- O 750
500
250
A d r e n a l e c t o m y
Restraint Stress ~ i i
PRE 1 0 20 Post 20 Post 40
1750
1500
1250
8' v l000 "r
750 <
5O0
250
Adrenalectomy + B
' ~ O - - - - ADX+B/H * • ADX+B/NH
PIE 10 20 Post 20 Post 40
1000
. • 750
- r 500
0
25O
Sham Controls
SHAM/H AMJNH
PRE 10 20 Post 20 Post 40
Fig. 3. Plasma ACTH levels prior to, during, and at various times following a 20-min restraint in adrenalectomized (ADX), A D X corticosterone-replaced ( A D X + B), and intact, sham-operated (SHAM) handled (H) and non- handled (NH) animals. (*) Values differ at P < 0.5. Integrated values for plasma ACTH (pg/ml/min) are as follows: SHAM/H, 326 + 68; SHAM/NH, 448+83; A D X / H , 1031:t:61; A D X / N H , 1070 4- 111; A D X + B/H, 609 + 142; and A D X + B/NH, 1091 i 111.

HPA Development 333
adrenalectomized/H rats and significantly lower than that in adrenalectomized + B/NH animals. Thus, basal B replacement was sufficient to reinstate the differences between H and NH rats. These data reflect the importance of the increased sensitivity to the negative feedback effects of glucocorticoids.
It is also important to note the specificity of this effect. The B replacement regimen used in this study was sufficient to attenuate greatly the adrenalectomy- induced increase in pituitary POMC mRNA, basal ACTH, and plasma CBG in both H and NH animals but altered stress-induced ACTH secretion only in the H animals. A number of previous studies have shown that in laboratory rats B pellet replacement of this order of magnitude is sufficient to correct basal ACTH secretion (Akana et al., 1988; Jacobson et al., 1988) and CBG production (Levin et al., 1987) but not stress-induced increases in plasma ACTH; these findings are comparable to our data with adrenalectomized + B/NH rats. In contrast, basal B replacement did attenuate, the ACTH hypersection with stress in adrenalectomized + B/NH rats. The reduced ACTH secretion in response to stress in the adrenalectomized + B/H animals is consistent with previous data showing increased glucocorticoid negative-feedback sensitivity in H animals. Taken together, these data indicate that differences in HPA response to stress between H and NH animals are dependent upon the presence of glucocorticoids but are not dependent upon stress-induced elevations in glucocorticoid levels.
These findings also suggest that negative-feedback differences between H and NH animals can occur in response to basal B levels. These differences appear to be reflected in differences in median eminence content of various ACTH secretagogues. The median eminence contains the axon terminals of the CRF/AVP neurons of the PVN, and measures of peptide content in this region are assumed to reflect the size of the readily releasible hormone pool. Thus, we found that under resting-state conditions, median eminence levels of CRF and AVP (but not oxytocin) are significantly higher in NH compared with H rats (Viau et al., 1993). We have also recently found that hypothalamic CRF mRNA levels are about 2.5-fold higher in NH compared with H animals (Plotsky and Meaney, 1993). Thus, under resting conditions, hypothalamic CRF and AVP synthesis appears to be elevated in NH rats: a difference that occurs in the presence of basal glucocorticoid levels.
The differences in median eminence levels of CRF and AVP offer an important insight into understanding the nature of the differences in glucocortic- oid negative feedback between H and NH animals. Indeed, we feel that these findings represent a biological basis for individual differences in "responsivity" to stress. Since H and NH animals do not differ in basal levels of ACTH or B (Meaney et al., 1989, 1992), it seems likely that the differences in CRF and AVP represent differences in readily releasable storage pools of these peptides in axons terminals of PVN neurons located in the median eminence. The excitatory signal at the level of the PVN of the hypothalamus associated with stress likely results in greater CRF and AVP release in the animals. This, in turn, would result in a greater plasma ACTH signal. This idea is consistent with the finding that H and NH animals differ in plasma ACTH responses to a wide variety of stimuli. Indeed, the differences in the terminal pools of CRF and AVP suggest that H and

334 Meaney et al.
NH animals would differ in stressors mediated by either secretagogue. Note that pituitary ACTH responses to both restraint and ether stress appear to be mediated by dynamic variations in both CRF and AVP (e.g., Linton et al. , 1985; Nakane et al. , 1985). This hypothesis has been at least partially confirmed in one recent study. Plotsky and Meaney (1993) found that CRF release from the median eminence in response to restraint was significantly greater in NH compared with H rats.
It is also important to note that adrenalectomized + B animals do not differ in corticotrope sensitivity to CRF (Akana et al. , 1986) and that differences in ACTH release in response to stress most likely reflect differences in neural regulatory components of ACTH secretion. This idea is also consistent with available information on the role of the hippocampus in mediating glucocorticoid inhibition of HPA activity. Hippocampal lesions result in a prolonged elevation of B following stress (Sapolsky et al. , 1984a). Herman et al. (1989) found that hippocampal lesions resulted in increased CRF and AVP mRNA levels in the hypothalamus under basal B conditions. Moreover, Sapolsky et al. (1990) found
HANDLED NONHANDLED
I 1 I"P mpu 3 Grec ~ B ~ Grec (Basal)
i I
I Hypothalamus ; Q Hypothalamus'~ CRF/AVP I CRF/AVP~
~ t ++++ ~ S T R E S S = +++++++
Fig. 4. A summary of our current understanding of the mechan- isms underlying the differences in HPA responses to acute stress in handled and nonhandled animals. The increased glucocorticoid receptor levels confer greater sensitivity to corticosterone (B) in hippocampal tissue from handled animals. Therefore, under even under basal corticosterone levels (which do not differ in young, adult handled, and adult nonhandled rats), there is a greater tonic inhibitory signal ( - ) on hypothalamic corticotropin-releasing factor (CRF) and vasopressin (AVP) synthesis in handled rats. In response to the neural signals associated with stress and impinging upon the hypothalamus, there is a greater release of CRF/AVP into the portal circulation ( + ) in the nonhandled rats, resulting in a greater ACTH release.

HPA Development 335
that portal concentrations CRF and AVP were negatively correlated with hippocampal glucocorticoid receptor occupancy. Interestingly, hippocampal gluc- ocorticoid receptor occupancy was significantly correlated with resting (prestress) portal concentrations of both CRF and AVP. These findings suggest that an increased glucocorticoid receptor signal at the level of the hippocampus is associated with decreased levels of hypophysial CRF and AVP.
On the basis of these data, it seems reasonable to propose (see Fig. 4) that (1) H and NH animals differ in delayed negative-feedback (Meaney et al., 1989) and that this difference is reflected in differential rates of CRF and AVP synthesis in the PVN of the hypothalamus, (2) differences in negative-feedback regulation are apparent even in response to basal B signals and occur as a result of the increased glucocorticoid receptor density in the hippocampus (and perhaps the medial prefrontal cortex), (3) in response to stress there is a greater release of CRF and/or AVP in the NH animals (Plotsky and Meaney, 1993), (4) giving rise to a greater increase in plasma ACTH levels and (5) a greater increase in plasma B levels which persists for a longer period of time (i.e., higher poststress plasma B levels in the NH animals). Thus, differences between H and NH animals in HPA activity both during and following stress can occur independently of the stress-induced increase in plasma B. In our view, the central feature of the handling effect on HPA responsivity to stress involves the changes in hippocam- pal glucocorticoid receptor gene expression.
MECHANISM OF ACTION OF HANDLING ON GLUCOCORTICOID RECEPTOR DEVELOPMENT
These descriptive studies have served to characterize at least partially the individual differences in HPA responses to stress represented in H and NH animals. As mentioned above, we consider the difference in hippocampal glucocorticoid receptor levels to be a critical feature for the effect of handling on HPA responses to stress. In another series of studies we have begun to examine the way in which the stimuli associated with the handling procedure actually serve to increase hippocampal glucocorticoid receptor gene expression.
In our initial studies, animals were handled for the first 21 days of life. Based on the earlier work by Levine (1970), we wondered whether some portion of this 3-week period might represent a critical period for the handling effect on glucocorticoid receptors. In one study (Meaney and Aitken, 1985b), animals were handled daily and sacrificed on either postnatal Day 3, 7, 14, or 21 of life. We found that, in comparison to same-aged NH animals, H animals exhibited significantly increased hippocampal glucocorticoid receptor density as early as day 7 of life and that magnitude of the effect did not increase thereafter. We also found that handling between Day 1 and Day 7 of life was as effective in increasing hippocampal glucocorticoid receptor density as handling over the entire first 3 weeks. Handling over the second week of life (i.e., between Day 8 and Day 14) was somewhat less effective, whereas animals handled between Day 15 and Day 21 did not differ from NH animals in glucocorticoid receptor binding. Thus,

336 Meaney et al.
glucocorticoid receptor binding capacity appears to be especially sensitive to environmental regulation during the first week of life. This finding is consistent with the previous studies of Levine (1970), who found that handling during the first week of life was sufficient to decrease corticosteroid responses to stress.
This temporal pattern corresponds to the normal developmental changes in glucocorticoid receptor density occurring over the early postnatal life (see De Kloet et al., 19 ; Meaney et al., 1993; Sarrieau et al., 1988). Glucocorticoid receptor density is low on postnatal Day 3 ( -30% of adult values) and increases steadily toward adult values, which are achieved by about the third week of life. It is during this period of ontogenetic variation that environmental events can influence the development of the receptor population. In contrast to the glucocorticoid receptor, mineralocorticoid receptor density does not vary sig- nificantly with age: Hippocampal mineralocorticoid receptor density in early postnatal life is largely indistinguishable from that of adult rats (Rosenfeld et al., 1988; Sarrieau et al., 1988), and as mentioned above, handling has no effect on hippocampal mineralocorticoid receptor binding (Meaney et al. , 1989b; Sarrieau et al., 1988). Thus, it is tempting to consider the relationship between the developmental pattern in hippocampal glucocorticoid receptor density and both (1) the sensitivity of this receptor system to environmental stimuli and (2) timing of the critical period for these stimuli on glucocorticoid receptor development.
However, the developmental pattern for glucocorticoid receptor binding in regions not affected by handling, such as the hypothalamus, amygdala, and septum (Olpe and McEwen, 1976; Meaney et al., 1985a), is identical to that of the hippocampus (references cited above) and the frontal cortex (Meaney and Aitken, 1985b). Thus, it is unlikely that the handling effect on glucocorticoid receptor density in the hippocampus and the frontal cortex can be explained simply by the status of the glucocorticoid receptor system during the first weeks of life.
Handling during the first week of life can involve a mild and transient drop in the pup's body temperature [although this feature of the manipulation is not critical for the effect on HPA activation during stress (see Levine, 1970)]. This is likely associated with the relatively modest thermoregulatory abilities of young rats over the first 2 weeks of life. Such changes in body temperature and/or some other sensory component of the handling manipulation activate the hypothamalic-pituitary-thyroid axis, leading to increased levels of circulating thyroxine (T4) and increased intracellular levels of the biologically more potent T 4 metabolite, triiodothyronine (T3). Handling results in a modest but reliable ( -30%) increase in plasma thyroid hormones (Meaney and Aitken, unpubl- ished). Thus, we (Meaney et al., 1987) examined whether the effects of handling might be mediated by increased exposure to these thyroid hormones. Neonatal rat pups were exposed to elevated levels of either T4, T3, or the vehicle over the first week of life. Both T4 and T3 treatment resulted in significantly increased glucocorticoid receptor binding capacity in the hippocampus in animals examined as adults. Like the handling manipulation, neither T 4 nor T3 treatment affected hypothalamic or pituitary glucocorticoid receptor density. Moreover, administra- tion of the thyroid hormone synthesis inhibitor, propylthrouracil (PTU), to H

HPA Development 337
pups for the first 2 weeks of life completely blocked the effects of handling on hippocampal glucocorticoid receptor binding capacity. These data are consistent with the idea that the thyroid hormones might mediate, at least in part; the effects of neonatal handling on the development of the hippocampal glucocorticoid receptor system.
Systemic injections of neonatal rat pups represent a fairly crude manipula- tion, and while these data might implicate the thyroid hormones, there is no indication that the hippocampus is the actually critical site of action for thyroid hormone effect. In order to examine whether hormones might act directly on hippocampal cells, we have turned to an in vitro system, using primary cultures of dissociated hippocampal cells (Banker and Cowan, 1977). The hippocampal cells are taken from embryonic rat pups (E20), and beginning on the fifth day after plating the cultures were exposed to 0, 1, 10, or 100 nM T3. The cells were cultured in 10% fetal calf serum, stripped of thyroid hormones. Thusfar, the results of several experiments have failed to detect any effect of thyroid hormones on glucocorticoid receptor density in cultured hippocampal cells (Meaney, unpublished). These in vitro data suggest that (a) the effects of the thyroid hormones on the glucocorticoid receptor binding capacity occurs at some site distal to the hippocampal cells or (b) thyroid hormones interact at the level of the hippocampus with some other hormonal signal that is obligatory for the expression of the thyroid hormone effect.
The thyroid hormones have pervasive effects throughout the developing CNS and one such effect involves the regulation of central serotonergic neurons (e.g., Savard et al., 1983). Thyroid hormones increase serotonin (5-HT) turnover in the hippocampus of the neonatal rat (Mitchell et al., 1990a). Handling also increases hippocampal 5-HT turnover (Mitchell et al., 1990a; Smythe et al., 1991), and thus both manipulations increase serotonergic stimulation of hippocampal neurons. There is also direct evidence for an effect of 5-HT on glucocorticoid receptor density in the neonatal rat. 5,7-Dihydroxytyrptamine (5,7-DHT) lesions of the raphe 5-HT neurons dramatically reduce the ascending serotonergic input into the hippocampus. Rat pups administered 5,7,-DHT on Day 2 of life showed reduced hippocampal glucocorticoid receptor density as adults (Mitchell et al., 1990a). Similar lesions of ascending catecholaminergic systems, using 6- hydroxydopamine, had no effect on the development of glucocorticoid receptor density in the hippocampus (Meaney, unpublished).
The next question, then, is whether 5-HT might be mediating the effect of handling at the level of the hippocampal cells. Indeed, 5-HT has a profound effect on glucocorticoid receptor density in cultured hippocampal cells (Mitchell et al., 1990b, 1992). In hippocampal cells cultured in the presence of 10 nM 5-HT there was a twofold increase in glucocorticoid receptor binding. The effect of 5-HT was dose-related, with an ECs0 of 4-5 nM. Interestingly, about 4 days of treatment was required for the maximal effect of 5-HT to occur. Shorter periods of exposure were ineffective, suggesting that the effect of 5-HT may involve the increased synthesis of receptors. In support of this idea, we (O'Donnell and Meaney, unpublished) found that the effect of 5-HT on glucocorticoid receptor density in cultured hippocampal cells was blocked by either actionmycin D or cycloheximide.

338 Meaney et al.
The effect of 5-HT on glucocorticoid receptor binding appears to occur uniquely in the neuronal cell population. We found that there was no effect of 5-HT on glucocorticoid receptor binding in hippocampal glial-enriched cell cultures. This finding is not surprising, since our initial studies were performed with cultures comprised largely ( -75%) of neuron-like cells (Mitchell et al., 1990b). Moreover, the composition of the cultures is unaffected by 5-HT treatment.
There are two very intriguing features of this effect that bear directly on the question of the hippocampal cell cultures as a model for neural differentiation (Meaney et al., 1990). First, the effects of 5-HT on glucocorticoid receptor binding in hippocampal cell cultures are restricted to the first 3 weeks in culture. Thus, cultures treated with 10 nM 5-HT for 7 days at any time during the first 3 weeks in culture show a significant increase in glucocorticoid receptor density; however, the effect is lost after this point. Cultures treated with 10 nM 5-HT for 7 days during the third to the fourth week following plating show no increase in glucocorticoid receptor binding. Second, and perhaps most exciting, the increase in glucocorticoid receptor binding capacity following exposure to 10 nM 5-HT persists as long as 40 days following 5-HT removal from the medium (Meaney et al., 1990). Thus, the effect of 5-HT on glucocorticoid receptor density observed in hippocampal culture cells mimics the long term effects of neonatal handling.
The effects of 10 nM 5-HT on glucocorticoid receptor density in cultured hippocampal cells was completely blocked by the 5-HTe receptor antagonists, ketanserin and mianserin (Mitchell et al., 1990b). Moreover, the 5-HT2 agonists 1-(2,5-dimethoxy-4-iodophenyl)-2-aminopropane (DOI), m-trifluoromethyl- phenylpiperazine (TFMPP), and quipazine were also effective in increasing gluco- corticoid receptor binding in hippocampal culture. Selective agonists or antagonists of the 5-HT1~ or 5-HT3 receptors had no effect on glucocorticoid receptor binding. Using [125I]7-amino-8-iodo-ketanserin as radioligand, we have confirmed the presence of high-affinity 5-HT2 binding sites in our cultured hippocampal cells. Titeler et al. (1987) have provided evidence that the ketanserin-labeled 5-HT2 site may exist in two states: an agonist state (5-HT2H) with a high, nanomolar affinity for 5-HT and an antagonist state (5-HTzL) with a low, molecular affinity for 5-HT. In both states the receptor shows a high affinity for antagonists, such as ketanserin. The 5-HT2r~ site binds with a high affinity to DOI, TFMPP, and quipazine, and Titeler et al. (1987) have reported a Kd of --5 nM for 5-HT. This Kd closely approximates the ECso (4-5 nM) for the effect of 5-HT on glucocortic- oid receptors in cultured hippocampal cells. Taken together, these findings suggest that this effect of 5-HT appears to be mediated via high-affinity, 5-HT: receptor.
We have begun to examine the nature of the secondary messenger systems involved in this serotonergic effect on glucocorticoid receptor binding. Mitchell et al. (1992) found that low nanomolar concentrations of 5-HT (ECso = 7.2 nM) produce a fourfold increase in cAMP levels in cultured hippocampal cells, with no effect on cGMP levels. This increase in cAMP is blocked by ketanserin and mimicked by both quipazine and DOI. Moreover, treatment with the stable cAMP analogue, 8-bromo-cAMP, or with 10 ~M forskolin produce a significant

HPA Development 339
increase in glucocorticoid receptor density in cultured hippocampal neurons. The effect of 8-bromo cAMP is concentration-related, and the maximal effect of 8-bromo-cAMP ( -181%) is comparable to that for 5-HT (-195%). Interestingly, as with 5-HT, the effects of 8-bromo cAMP on glucocorticoid receptor density were not apparent until at least 4 days of treatment.
Taken together, these findings suggest that changes in cAMP concentrations may mediate the effects of 5-HT on glucocorticoid receptor synthesis in hippocampal cells. We (Mitchell et al., 1992) also found that the cyclic nucleotide-dependent protein kinase inhibitor, H8 (Hidaka et al., 1984), comp- letely blocked the effects of 10 nM 5-HT on glucocorticoid receptor binding 'in hippocampal cell cultures. In contrast, the protein kinase C inhibitor, H7, had no such effect. These data suggest that activation of protein kinase A might, at some point, be involved in the serotonergic regulation of hippocampal glucocorticoid receptor development.
We must also consider the fact that we are working with intact cells over long incubation periods, and there is ample possibility for an interaction between second-messenger systems. In v ivo 5-HT increases both diacylglycerol (DAG) levels and inositol phosphate metabolism (notably IP1) in hippocampal mem- branes from neonates (Parent, Sharma, and Meaney, in preparation). The effect is not quite as potent as in the frontal cortex (e.g., Claustre et al., 1985), perhaps reflecting the fact that 5-HT2 receptor density is greater in the cortex. Interest- ingly, our preliminary findings suggest that repeated handling of pups increases the amount of stimulation associated with 5-HT. We also have data suggesting that treatment with a phorbol ester also increases glucocorticoid receptor binding in the hippocampal cell cultures (O'Donnell and Meaney, unpublished). These findings suggest that 5-HT effects on glucocorticoid receptor density might, at some level, involve the mobilization of intracellular Ca 2+ stores. Interestingly, Ca-calmodulin activation has been found to increase intracellular cAMP levels. The postreceptor events involved in the regulation of glucocorticoid receptor development are likely to be complex, but these studies should ultimately lead to an understanding of how environmental events can regulate glucocorticoid receptor gene expression and neuronal differentiation in the hippocampus.
Recent studies (Smythe et al., 1991) of 5-HT turnover have provided some insight into why the hippocampus and frontal cortex are selectively affected by handling. We found that when rat pups were handled for the first 7 days of life and sacrified immediately following handling on Day 7, 5-HT turnover was significantly increased in the hippocampus and frontal cortex but not in the hypothalamus or amygdala (regions where handling has no effect on glucocortic- oid receptor density). These data suggest that handling selectively activates certain ascending 5-HT pathways and that it is this feature of the handling effect, together with the existence of a developing glucocorticoid receptor system, that underlies the sensitivity of this receptor system in specific brain regions to regulation by environmental events during the first week of life. In contrast, handling has no effect on 5-HT 2 receptor density in either the hippocampus on the frontal cortex during the first week of life (Smythe, 1991). Thus, handling increases 5-HT turnover but not 5-HT2 receptor sites in the hippocampus and

3411 Meaney et al.
frontal cortex. Nevertheless, the in v ivo data also suggest that 5-HT acts at the 5-HT2 receptor to alter hippocampal glucocorticoid receptor density. We found that effects of postnatal handling on glucocorticoid receptor binding are blocked by concurrent administration of the 5-HT2 receptor antagonist, ketanserin (Mitchell et al., 1990a).
WORKING MODEL
A model of the way in which neonatal handling might increase glucocorticoid receptor density in the hippocampus and frontal cortes is provided in Fig. 5. Handling, although a brief interlude in the routine of mother-pup interactions, provides for an increase in sensory stimulation which, in turn, elicits a set of physiologial responses in the pup. These responses and their relevance for the handling effect remain as a black box at the present. In addition, the mother is also subject to the manipulation. We have found that handling the mother alone (leaving the pups undisturbed) has no effect on hippocampal glucocorticoid receptor density (Aitken and Meaney, unpublished). However, handling the pups does alter the behavior of the mother toward the offspring, and the effect persists well beyond the immediate period following the return of the mother and pups to the nest. H pups show increased ultrasonic vocalizations (Bell et al., 1971) and such calls elicit maternal care (e.g., retrieval). Immediately following the return of the pups, mothers of H litters show increased care of their offspring. Over the
Han0n01 I Response°f the Mother j
S iern~uSul~!Y o n I ~, p h~S:p io~gsieC a ~
[ 5-HT2 receptor I
AP2 Regl~ation?? Inhibition J
of • [AP2 site' Glucocorticoid Receptor Gene CRF Neuron Fig. 5. A working model summarizing our current understanding of the processes by which handling alters glucocorticoid receptor levels in rat hippocampus (see text for description) and, subsequently, the regulation of CRF activity in the PVN.

HPA Development 341
long term, handling also increases maternal licking (Lee and Williams, 1974) and alters nest-building (Lee and Williams, 1975). Several aspects of the mother-pup interactions are of known importance in neural development, especially maternal licking (135-137). We (Woodside, Jans, and Meaney, unpublished) have ex- amined the effect of handling on the amount of time the mother spends with her offspring. Typically, a nesting bout begins when the mother approaches the litter, gathers the pups under her, and suckles and terminates when the mother licks the pups and leaves the nest (the termination of a nest bout is associated with a buildup in the mother's body temperature). Overall, mothers of H pups spend the same amount of time with their litters as mothers of NH pups. However, mothers of H litters have shorter but more frequent nest bouts and spend significantly more time manipulating their pups, especially licking of pups (Lee and Williams, 1974; Meaney, unpublished) than mothers of NH pups. The latter effect is likely due to the fact that mothers of H litters are getting on and off their pups more frequently, and licking is associated with the termination of each bout. In addition, handling of pups also elicits endocrine responses from the mother that do not occur as a function of simply being separated from her pups (Smotherman et al., 1977). The question that arises is whether some aspect of this altered pattern of maternal behavior/physiology may mediate, at least in part, the effects of handling on the development of the HPA axis (see Hennessy et al., 1982).
The handling procedure is associated with increased plasma thyroid hormone levels. Whether this is due to alterations in mother-pup interactions or to the actual manipulation of the pups is not clear. Thyroid hormones serve to increase 5-HT turnover in the hippocampus (see above). Both in vitro and in v i vo experiments suggest that the 5-HT on glucocorticoid receptor binding is mediated through a 5-HT2 receptor. Importantly, we recently found that the effects of T3 on hippocampal glucocorticoid receptor binding are blocked by concurrent treatment with ketanserin (Sharma, Aitken, and Meaney, in preparation). This finding strengthens the proposed link between thyroid hormones and 5-HT in the regulation of glucocorticoid receptor density in the hippocampus.
The postreceptor events involved in the 5-HT appear to involve increases in cAMP levels and the activation of cycle nucleotide-dependent protein kinases. Whether this cascade is initiated or faciltated by changes in inositol phosphate or diacylglycerol metabolism is not clear; however, there are numerous examples in the literature of such "cross-talk" between second-messenger systems (e.g., Yoshimasa et al., 1987). cAMP regulation of gene transcription has been shown to be mediated by at least two transcription factors: the cyclic nucleotide response element binding protein (CREB) and activator protein-2 [AP2 (Habener, 1990; Imagawa et al., 1987; Montminy et al., 1990; Roesler e ta l . , 1988)]. The promoter region of the glucocorticoid receptor gene has been mapped and contains numerous binding sites for the AP2 transcription factor. Interestingly, AP2 is induced by both cAMP and forskolin. Therefore, at this time it is interesting to speculate that the cascade of intracellular events may involve AP2 induction.
Then we arrive at the most interesting feature of these effects: tbe finding that the handling effect, as well as the in vitro effect of 5-HT, persists well beyond the period of the treatment. There are at least two possible explanations for this

342 Meaney et al.
finding. First, the increase in 5-HT turnover associated with the handling procedure might be accomplished by an increase in 5-HT innervarion of the hippocampus which persists throughout the life of the animal. The increased 5-HT innervation could then serve to maintain the handling effect. Seckl et al.
(1991) have shown that 5,7-DHT lesions in the hippocampus decrease glucocor- ticoid receptor mRNA level. However, such lesions also decrease mineralocortic- oid mRNA levels, which are unaffected by handling. Nevertheless, we are currently examining the 5-HT innervation into the hippocampus in H and NH animals at various stages of life. Second, handling could result in a "structural" change in the glucocorticoid receptor gene (and/or the promoter region) which sustains a difference in basal transcription rates throughout the life of the animal.
CONCLUSIONS
Handling during the early postnatal period leads to increased glucocorticoid receptor binding in the hippocampus and is associated with enhanced negative- feedback control over HPA function. It is likely that this plasticity reflects a basic process, whereby the early environment is able to "fine-tune" the sensitivity and efficiency of certain neuroendocrine systems that mediate the animal's response to stimuli that threaten homeostasis. Such plasticity is likely to be of considerable importance to a species such as the rat, which prospers in a tremendous range of ecological niches. In this sense, it is important to note that we do not consider H or NH animals to be superior to one another (or to be more "fit" in the biological sense). They differ as a function of the variation in early stimulation afforded the animals. What is of great interest to us is the possibility that the handling manipulation provides a paradigm for the study of the molecular processes through which the development of specific neuroendocrine systems is directed by the environment. This, of course, was the source of the considerable enthusiasm for handling studies in the early periods of developmental psychobiology.
ACKNOWLEDGMENTS
The research described in this paper was supported by grants from the Medical Research Council of Canada (MRCC) and from the National Institute on Aging (AG09488) to M.J.M.M.J.M. is the recipient of an MRCC Scientist career award. N.S. and J.S. were supported by postdoctoral fellowships from the Natural Sciences and Engineering Research Council of Canada (NSERC). S.B. is a graduate fellow of the Canadian Heart Foundation and the Medical Research Council of Canada.

HPA Development 343
REFERENCES
Ader, R., and Grota, L. J. (1969). Effects of early experience on adrenocortical reactivity. Physiol. Behav. 4:303-305.
Akana, S. F., Cascio, C. S., Du, J.-Z., Levin, N., and Dallman, M. F. (1986). Reset of feedback in the adrenocortical system: An apparent shift in sensitivity of adrenocorticotropin to inhibition by corticosterone between morning and evening. Endocrinology 119:2325-2332.
Akana, S. F., Jacobson, L., Cascio, C. S., Shinsako, J., and Dallman, M. F. (1988). Constant corticosterone replacement normalizes basal adrenocorticotropin (ACTH) but permits sustained ACTH hypersecretion stress in adrenalectomized rats. Endorcinology 122:1337-1342.
Antoni, F. A. (1986). Hypothalamic control of ACTH secretion: Advances since the discovery of 41-residue corticotropin-releasing factor. Endocr. Rev. 7:351-370.
Banker, G. A., and Cowan, W. M. (1977). Rat hippocampal neurons in dispersed cell culture. Brain Res. 126:397-425.
Beaumont, K., and Fanestil, D. D. (1983). Characterization of rat brain aldosterone receptors reveals high affinity for corticosterone. Endocrinology 113:2043-2051.
Bell, R. W., Nitschke, W., Gorry, T. H., and Zachman, T. A. (1971). Infantile stimulation and ultrasonic signaling: A possible mediator of early handing phenomena. Dev. Psychobiol. 4:181-191.
Beyer, H. S., Matta, S. G., and Sharp, B. M. (1988). Regulation of the messenger ribonucleic acid for corticotropin-releasing factor in the paraventricular nucleus and other brain sites of the rat. Endocrinology 123:2117-2123.
Bradbury, M., and Dallman, M. F. (1991). Effects of type 1 and type 2 glucocorticoid receptor antagonists on ACTH levels in the PM. Soc. Neurosci. Abstr. 19:716.
Brindley, D. N., and Rolland, Y. (1989). Possible connections between stress, diabetes, obesity, hypertension and altered lipoprotein metabolism that may result in atherosclerosis. Clin. Sci. 77:453-461.
Claustre, Y., Roquier, L., and Scatton, B. (1988). J. Pharmacol. Exp. Ther. 244:1051-1056. Dallman, M. F., Akana, S., Cascio, C. S., Darlington, D. N., Jacobson, L., and Levin, N. (1987).
Regulation of ACTH secretion: Variations on a theme of B. Rec. Prog. Horm. Res. 43:113-173. Dallman, M. F., Levin, N., Cascio, C. S., Akana, S. F., Jacobson, L., and Kuhn, R. W. (1989).
Pharmacological evidence that the diurnal regulation of adrenocorticotropin secretion by corticosteroids is mediated by type I B-preferring receptors. Endorcrinology 124:2844-2850.
Diorio, D., Viau, V., and Meaney, M. J. (1993). The role of the frontal cortex in the regulation of hypothalamic-pituitary-adrenal activity. J. Neuroscience (in press).
Emadian, S. M., Luttge, W. G., and Densmore, C. L. (1986). Chemical differentiation of Type I aed Type II receptors for adrenal steroids in brain cytosol. J. Steroid Biochem. Mol. Biol. 24:953-961.
Evans, R. M., and Arriza, J. L. (1989). A molecular framework for the actions of glucocorticoid hormones in the nervous system. Neuron 2:1105-1112.
Feldman, S., and Conforti, N. (1976). Feedback effects of dexamethasone on adrenocortical responses in rats with fornix lesions. Horm. Res. 7:56-60.
Feldman, S., and Conforti, N. (1980). Participation of the dorsal hippocampus in the glucocorticoid negative-feedback effect on adrenocortical activity. Neuroendocrinology 30:52-55.
Fischette, C. T., Komisurak, B. R., Ediner, H. M., Feder, H. H., and Siegal, A. (1980). Differential fornix ablations and the circadian rhythmicity of adrenal corticosterone secretion. Brain Res. 195:373-380.
Funder, J. W., and Sheppard, K. (1987). Adrenocortical steroids and the brain. Annu. Rev. Physiol. 49:397-412.
Gibbs, D. M. (1986). Vasopressin and oxytocin. Hypothalamic modulators of the stress response. Psychoneuroendocrinology 11:131-140.
Goldman, L., Winget, C., Hollinshead, G., and Levine, S. (1978). Postweaning development of negative feedback in the pituitary-adrenal system of the rat. Neuroendocrinology 12:199-211.
Grota, L. J. (1975). Effects of early experience on the metabolism and production of corticosterone in rats. Dev. Psychobiol. 9:211-215.
Habener, J. (1990). Cyclic AMP response element binding proteins: A cornucopia of transcription factors. Mol. Endocrinol. 4:1087-1094.
Harbuz, M. S., and Lightman, S. L. (1992). Stress and the hypothalamic-pituitary-adrenal axis, acute, chronic and imminological activation. J. Endocrinol. 134:327-339.
Hennessy, M. B., VogL J., and Levine, S. (1982). Strain of foster mother determines long-term effects of early handling: Evidence for maternal mediation. Pyshiol. Psychol. 10:153-157.

344 Meaney et al.
Herman, J. P., Schafer, M. K.-H., Young, E. A., Thompson, R., Douglass, J., Akil, H., and Watson, S. J. (1989). Evidence for hippocampal regulation of neuroendocrine neurons of the hypothalamo-pituitary-adrenocortical axis. J. Neurosci. 9:3072-3082.
Hess, J. L., Denenberg, V. H., Zarrow, M. X., and Pfeifer, W. D. (1960). Modification of the corticosterone response curve as a function of handling in infancy. Physiol. Behav. 4:109-112.
Hidaka, H., Inagaki, M., Kawamamoto, S., and Sasaki, Y. (1984). Isoquinolinesulfonamides, novel and potent inhibitors of cyclic nucleotide dependent protein kinase A and protein kinase C. Biochemistry 23:5036-5041.
Imagawa, M., Chiu, R., and Karin, M. (1987). Transcription factor AP-2 mediates induction by two different signal-transduction pathways: Protein kinase C and cAMP. Cell 51:251-260.
Jacobson, L., and Sapolsky, R. M. (1991). The role of the hippocampus in feedback regulation of the hypothalamic-pituitary-adrenal axis. Endocr. Rev. 12:118-134.
Jacobson, L., Akana, S., Cascio, C. S., Shinsako, J., and Dallman, M. F. (1988). Circadian variations in plasma B permit normal termination of adrenocorticotropin responses to stress. Endocrinology 122:1343-1349.
Jones, M. T., Gillham, B., Greenstein, B. D., Beckford, U., and Holmes, M. C. (1982). Feedback actions of adrenal steroid hormones. In Current Topics in Neuroendocrinology, Vol. 2 (D. Ganten and D. Pfaff, Eds.), Springer, New York, pp. 45-68.
Keller-Wood, M., and Dallman, M. F. (1984). Corticosteroid inhibition of ACTH secretion. Endocr. Reo. 5:1-24.
Kiss, J. Z., Mezey, E., and Skirboll, L. (1984). Corticotropin-releasing factor-immunoreactive neurons become vasopressin positive after adrenolactomy. Proc. Natl. Acad. Sci. USA 81:1854- 1858.
Kovacs, K. J., and Makara, G. B. (1988). B and dexamethosone act at different brain sites to inhibit adrenalectomy-induced adrenocorticotropin hypersection. Brain Res. 474:205-210.
LaRocque, S., O'Donnell, D., Gianoulakis, C., Seckl, J. R., and Meaney, M. J. (1992). Postnatal handling in the rat alters hippocampal glucocorticoid receptor gene expression. Soc. Neurosci. Abstr. 18:479.
Lee, M. H. S., and Williams, D. I. (1974). Changes in licking behavior of rat mother following handling of young. Anita. Behav. 22:679-681.
Lee, M. H. S., and Williams, D. I. (1975). Long term changes in nest condition and pup grouping following handling of rat litters. Dev. Psychobiol. 8:91-95.
Levin, N., Shinsako, J., and Dallman, M. F. (1987). B acts on the brain to inhibit adrenalectomy- induced adrenocorticotropin secretion. Endocrinology 122:694-701.
Levine, S. (1957). Infantile experience and resistence to physiological stress. Science 126:405-406. Levine, S. (1962). Plasma-free corticosteroid response to electric shock in rats stimulated in infancy.
Science 135:795-796. Levine, S. (1970). The pituitary-adrenal system and the developing brain. Prog. Brain Res.
32:79-102. Levine, S., Haltmeyer, G. C., Karas, G. G., and Denenberg, V. H. (1967). Physiological and
behavioral effects of infantile stimulation. Physiol. Behav. 2:55-63. Lightman, S. L., and Young, W. S., III (1987). Vasopressin, oxytocin, dynorphin, enkephalin, and
corticotropin-releasing factor mRNA stimulation in the rat. J. Physiol. 394:23-39. Linton, E. A., Tilders, F. J. H., Hodgkinson, S., Berkenbosch, F., Vermes, I., and Lowry, P. J.
(1985). Stress-induced secretion of adrenocorticotropin in rats is inhibited by antisera to corticotropin-releasing factor and vasopressin. Endocrinology 116:966-970.
McEwen, B. S., DeKloet, E. R., and Rostene, W. H. (1986). Adrenal steroid receptors and actions in the nervous system. Physiol. Rev. 66:1121-1150.
Meaney, M. J., and Aitken, D. H. (1985a). [3H]Dexamethasone binding in rat frontal cortex. Brain Res. 328:176-180.
Meaney, M. J., and Aitken, D. H. (1985b). The effects of early postnatal handling on the development of hippocampal glucocorticoid receptors: Temporal parameters. Dev. Brain. Res. 22:301-304.
Meaney, M. J., Aitken, D. H., Bodnoff, S. R., Iny, L. J., and Sapolsky, R. M. (1985a). The effects of postnatal handling on the development of the glucocorticoid receptor systems and stress recovery in the rat. Prog. Neuropsychopharm. Biol. Psychiat. 7:731-734.
Meaney, M. J., Aitken, D. H., Bodnoff, S. R., Iny, L. J., Tatarewicz, J. E., and Sapolsky, R. M. (1985b). Early, postnatal handling alters glucocorticoid receptor concentrations in selected brain regions. Behav. Neurosci. 99:760-765.
Meaney, M. J., Aitken, D. H., and Sapolsky, R. M. (1987). Thyroid hormones influence the development of hippocampal glucocorticoid receptors in the rat: A mechanism for the effects of

HPA Development 345
postnatal handling on the development of the adrenocortical stress response. Neuroendocrinology 45:278-283.
Meaney, M. J., Viau, V., Bhatnagar, S., and Aitken, D. H. (1988a). Occupancy and translocation of hippocampal glucocorticoid receptors during and following stress. Brain Res. 445:198-203.
Meaney, M. J., Aitken, D. H., Bhatnagar, S., Van Berkel, C., and Sapolsky, R. M. (1988b). Postnatal handling attenuates neuroendocrine, anatomical, and cognitive impairments related to the aged hippocampus. Science 238:766-768.
Meaney, M. J., Aitken, D. H., Sharma, S., Viau, V., and Sarrieau, A. (1989). Postnatal handling increases hippocampal type II, glucocorticoid receptors and enhances adrenocortical negative- feedback efficacy in the rat. Neuroendocrinology 50:597-604.
Meaney, M. J., Aitken, D. H., Sharma, S., and Viau, V. (1992). Basal ACTH, corticosterone, and corticosterone-binding globulin levels over the diurnal cycle, and hippocampal type I and type II corticosteroid receptors in young and old, handled and nonhandled rats. Neuroendocrinology 55:204-213.
Meaney, M. J., Bodnoff, S. R., O'Donnell, D., Rowe, W., Sarrieau, A., Rose, G. M., Poirier, J., and Seckl, J. R. (1993a). Glucocorticoids as regulators of neuron survival and repair in the aged brain. In A. C. Cuello (Ed.), Resorative Neurology, Vol. 6, Elsevier, Amsterdam (in press).
Meaney, M. J., O'Donnell, D., Viau, V., Bhatnagar, S., Sarrieau, A., Smythe, J. W., Shanks, N., and Walker, C.-D. (1993b). Corticosteroid receptors in rat brain and pituitary during develop- ment and hypothalamic-pituitary-adrenal (HPA) function. In Receptors and the Developing Nervous System (P. McLaughin and I. Zagon (Eds.), Chapman and Hall, London (in press).
Merchenthaler, I., Vigh, S., Petrusz, P., and Schally, A. V. (1983). Tha paraventriculo-infundibular corticotropin-releasing factor (CRF) pathway as revealed by immunochemistry in long-term hypophysectomized or adrenalectomized rats. Regul. Peptides 5:295-305.
Mitchell, J. B., Iny, L. J., and Meaney, M. J. (1990a). The role of serotonin in the development and environmental regulation of hippocampal type II corticosteroid receptors. Developmental Brain Res. 55:231-235.
Mitchell, J. B., Rowe, W., Boksa, P., and Meaney, M. J. (1990b). Serotonin regulates type If corticosteroid receptor binding in hippocampal cell cultures. J. Neurosci. 10:1745-1752.
Mitchell, J. B., Betito, K., Rowe, W., Boksa, P., and Meaney, M. J. (1993). Serotonergic regulation of type II corticosteroid receptor binding in cultured hippocampal cells: The role of serotonin- induced increases in cAMP levels. Neuroscience (in press).
Montminy, M. R., Gonzalez, G. A., and Yamamoto, K. K. (1990). Regulation of cAMP-inducible genes by CREB. Trends Neurosci., 184-188.
Munck, A., Guyre, P. M., and Holbrook, N. J. (1984). Physiological functions of glucocorticoids in stress and their relations to pharmacological actions. Endocr. Rev. 5:25-44.
Nakane, T., Aughya, T., Kanie, N., and Hollander, C. S. (1985). Evidence for the role of endogenous corticotropin-releasing factor in cold, ether, immobilization, and a traumatic stress. Proc. Natl. Acad. Sci. USA, 82-'1247-1251.
Olpe, H. R., and McEwen, B. S. (1976). Glucocorticoid binding to receptor-like proteins in rat brain and pituitary: Ontogenetic and experimentally-induced changes. Brain Res. 105:121-128.
Partridge, W. M., Sakiyama, R., and Judd, H. L. (1983). Protein-bound corticosterone in human serum is selectively transported into rat brain and liver in vivo. J. Clin. Endocrinol. Metabol. 57:160-166.
Plotsky, P. M. (1987). Regulation of hypophysiotropic factors mediating ACTH secretion. Ann. N.Y. Aead. Sci. 512:205-217.
Plotsky, P. M., and Meaney, M. J. (1993). Effects of early environment on hypothalamic corticotropin-releasing factor mRNA, content and stress-induced release. Mol. Brain Res. (in press).
Plotsky, P. M., and Vale, W. W. (1984). Hemorrhage-induced secretion of corticotropin-releasing factor-like immunoreactivity into the rat hypophysial portal circulation and its inhibition by glucocortieoids. Endorcinology 114:164-169.
Plotsky, P. M , and Sawchenko, P. E. (1987). Hypophysial plasma portal levels, median eminence content, and immunohistochernical staining of corticotropin-releasing factor, arginine vasopres- sin, and oxytocin and pharmacological adrenalectomy. Endocrinology 120:1361-1369.
Piotsky, P. M., Otto, S., and Sapolsky, R. M. (1987). Inhibition of immunoreactive corticotropin- releasing factor into the hypophysial-portal circulation by delayed glueocorticoid feedback. Endoreinology 119:1126-1130.
Ratka, A., Sutanto, W., BIoemers, M., and De Kloet, E. R. (1989). On the role of brain mineralocorticoid (type I) and glucocorticoid (type II) receptors in neuroendocrine regulation. Neuroendocrinology 50:117-123.

346 Meaney et al.
Reul, J. H. M., and DeKloet, E. R. (1985). Two receptor systems for corticosterone in rat brain: Microdistribution and differential occupation. Endocrinology 117:2505-2511.
Reui, J. M. H. M., and De Kloet, E. R. (1986). Anatomical resolution of two types of corticosterone receptor site in the rat brain with in vitro autoradiography and computerized image analysis. J. Steroid Biochem. Mol. Biol. 24:269-272.
Reul, J. M. H. M., van den Bosch, F. R., and DeKloet, E. R. (1987). Relative occupation of type-I and type-II corticosteroid receptors in rat brain following stress and dexamethasone treatment Functional implications. J. Endocrinol. 115:459-467.
Rivier, C., and Vale, W. W. (1983). Effects of angiotensin lI on ACTH release in vivo: Role of corticotropin-releasing factor (CRF). Regul. Peptides 7:253-258.
River, C., and Plotsky, P. M. (1986). Mediation by corticotropin-releasing factor of adenohypophysial hormone secretion. Annu. Rev. Physiol. 48:475-489.
Rivier, C., Brownstein, M., Spiess, J., Rivier, J., and Vale, W. W. (1982). In vivo corticotropin- releasing factor-induced secretion of adrenocorticotropin, /~-endorphin, and corticosterone. Endocrinology 110:272-278.
Roesler, W. J., Vandenbark, G. R., and Hanson, R. W. (1988). Cyclic AMP and the induction of eukaryotic gene transcription. J. Biol. Chem. 263:9063-9066.
Rosenfeld, P., Sutanto, W., Levine, S., and De Kloet, E. R. (1988a). Ontogeny of type I and type II corticosteroid receptors in the rat hippocampus. Dev. Brain Res. 42:113-138.
Rosenfeld, P., Van Eekelen, J. A. M., Levine, S., And DeKloet, E. R. (1988b). Ontogeny of the type 2 glucocorticoid receptor in discrete rat brain regions: An immunocytochemical study. Dev. Brain Res. 42:119-127.
Sapolsky, R. M. (1990). Glucocorticoids, hippocampal damage and the glutaminergic synapse. Prog. Brain Res. 86:13-23.
Sapolsky, R. M., and Meaney, M. J. (1986). The maturation of the adrenocortical stress response in the rat. Brain Res. Rev. 11:65-76.
Sapoisky, R. M., Krey, L. C., and McEwen, B. S. (1984a). Glucocorticoid-sensitive hippocampal neurons are involved in terminating the adrenocortical stress response. Proc. Natl. Acad. Sci. USA 81:6174-6177.
Sapolsky, R. M., Krey, L. C., and McEwen, B. S. (1984b). Stress down-regulated corticosterone receptors in a site-specific manner. Endocrinology 114:287-292.
Sapolsky, R. M., Krey, L. C., and McEwen, B. S. (1986). The neuroendocrinology of stress and aging: The glucocorticoid cascade hypothesis. Endocr. Rev. 7:284-301.
Sapolsky, R. M., Armanini, M. P., Packan, D. R., Sutton, S. W., and Plotsky, P. M. (1990). Glucocorticoid feedback inhibition of adrenocorticotropic hormone secretagogue release. Relati- onship to corticosteroid receptor occupancy in various limbic sites. Neuroendocrinology 51:328- 336.
Sarrieau, A., Sharma, S., and Meaney, M. J. (1988). Postnatal development and environmental regulation of hippocampal glucocorticoid and mineralocorticoid receptors in the rat. Dev. Brain Res. 43:158-162.
Savard, P., Merand, Y., Di Paolo, T., and Dupont, A. (1984). Thyroid hormone regulation of serotonin metabolism in developing rat brain. Brain Res. 292:99-108.
Seekl, J. R., Dickson, K. L., and Fink, G. (1990). Central 5,7-dihydroxytryptamine lesions decrease hippocampal glucocorticoid and mineralocorticoid receptor messenger ribonucleic acid expres- sion. J. Neuroenocrinol. 2:911-916.
Selye, H. (1950). The Physiology and Pathology o f Exposure to Stress. Acta, Montreal. Sheppard, K. E., and Funder, J. W. (1987). Equivalent affinity of aldosterone and corticosterone for
type I receptors in kidney and hippocampus: Direct binding studies. J. Steroid Biochem. Mol. Biol. 28:737-742.
Smotherman, W. P., Wiener, S. G., Mendoza, S. P., and Levine, S. (1977). Maternal pituitary- adrenal responsiveness as a function of differential treatment of rat pups. Dev. Psychobiol. 10:113-122.
Smythe, J. W., Rowe, W., and Meaney, M. J. (1991). The effects of neonatal handling on serotonin turnover and receptor binding. Soc. Neurosci. Abstr. 17:990.
Swanson, L. W., and Simmons, D. M. (1989). Differential steroid hormone and neural influences on peptide mRNA levels in CRH cells of the paraventricular nucleus: A hybridization histochemical study in the rat. J. Comp. Neurol., 285:413-435.
Swanson, L. W., Sawchenko, P. E., Lind, R. W., and Rho, J. H. (1983). The CRH motoneuron: Differential peptide regulation in neurons with possible synaptic, paracrine, and endocrine outputs. Ann. N.Y. Acad. Sci. 512:12-23.

HPA Development 347
Titeler, M., Lyon, R. A., Davis, K. H., and Glennon, R. A. (1987). Selectivity of serotoninergic drugs for multiple brain serotonin receptors. Biochem. Pharmacol. 36:3265-3271.
Tornello, S., Orti, E., DeNicola, A. F., Rainbow, T. C., and McEwen, B. S. (1982). Regulation of glucocorticoid receptors in brain by corticosterone treatment of adrenalectomized rats. Neuroendocrinology 35:411-417.
Van Loon, G. R., and DeSousa, E. B. (1987). Regulation of stress-induced secretion of POMC-derived peptides. Ann. N.Y. Acad. Sci. 512:300-307.
Viau, V., Sharma, S., Plotsky, p. M., and Meaney, M. J. (1993). Increased plasma ACTH responses to stress in nonhandled compared with handled rats require basal corticosterone levels and are associated with increased levels of ACTH secretagogues in the median eminence. J. Neurosci. (in press).
Wilson, M., Greer, M., and Roberts, L. (1980). Hippocampal inhibition of pituitary-adrenocortical function in female rats. Brain Res. 197:344-351.
Wolfson, B., Manning, R. W., Davis, L. G., Arentzen, R., and Baldrino, F. (1985). Co-localization of corticotropin-releasing factor and vasopressin mRNA in neurones after adrenalectomy. Nature 315:59-61.
Yoshimasa, T., Sibley, D. R., Bouvier, M., Lefkowitz, R. J., and Caron, M. G. (1987). Cross-talk between cellular signalling pathways suggested by phorbol-ester-induced adenylate cyclase phosphorylation. Nature 327:67-70.
Young, W. S., III, Mezey, E., and Siegel, R. E. (1986). Quantitative in situ hybridization histochemistry reveals increased corticotropin-releasing factor mRNA after adrenalectomy in rats. Neurosci. Lett. 70:198-203.
Zarrow, M. X., Campbell, P. S., and Denenberg, V. H. (1972). Handling in infancy: Increased levels of the hypothalamic corticotropin releasing factor (CRF) following exposure to a novel situation. Proc. Soc. Exp. Biol. Med. 356:141-143.




















