
Secretin: Hypothalamic Distribution and Hypothesized Neuroregulatory Role in Autism
23
Cellular and Molecular Neurobiology, Vol. 24, No. 2, April 2004 ( C 2004) Secretin: Hypothalamic Distribution and Hypothesized Neuroregulatory Role in Autism M. G. Welch, 1,2,4 J. D. Keune, 1,2 T. B. Welch-Horan, 1,2 N. Anwar, 1,2 M. Anwar, 1,2 R. J. Ludwig, 3 and D. A. Ruggiero 1,2,3 Received June 19, 2003; accepted July 18, 2003 SUMMARY 1. This study aims (1) to determine whether secretin is synthesized centrally, specifically by the HPA axis and (2) to discuss, on the basis of the findings in this and previous studies, secretin’s possible neuroregulatory role in autism. 2. An immunocytochemical technique with single-cell resolution was performed in 12 age/weight-matched male rats pretreated with stereotaxic microinjection of colchicine (0.6 μg/kg) or vehicle into the lateral ventricle. Following 2-day survival, rats were anes- thetized and perfused for immunocytochemistry. Brain segments were blocked and alternate frozen 30-μm sections incubated in rabbit antibodies against secretin, vasoactive intestinal peptide, glucagon, or pituitary-adenylate-cyclase-activating peptide. Adjacent sec- tions were processed for Nissl stain. Preadsorption studies were performed with members of the secretin peptide family to demonstrate primary antibody specificity. 3. Specificity of secretin immunoreactivity (ir) was verified by clear-cut preadsorption control data and relatively high concentrations and distinct topographic localization of se- cretin ir to paraventricular/supraoptic and intercalated hypothalamic nuclei. Secretin levels were upregulated by colchicine, an exemplar of homeostatic stressors, as compared with low constitutive expression in untreated rats. 4. This study provides the first direct immunocytochemical demonstration of secretiner- gic immunoreactivity in the forebrain and offers evidence that the hypothalamus, like the gut, is capable of synthesizing secretin. Secretin’s dual expression by gut and brain secretin cells, as well as its overlapping central distribution with other stress-adaptation neurohor- mones, especially oxytocin, indicates that it is stress-sensitive. A neuroregulatory relation- ship between the peripheral and central stress response systems is suggested, as is a dual role for secretin in conditioning both of those stress-adaptation systems. Colchicine-induced upregulation of secretin indicates that secretin may be synthesized on demand in response to stress, a possible mechanism of action that may underlie secretin’s role in autism. KEY WORDS: secretin; autism; HPA stress axis; secretinergic neurons; conditioned response; oxytocin; visceral; neuroregulatory. 1 Laboratories of Childhood Regulatory Disorders and Behavioral Neuroanatomy, Columbia University College of Physicians and Surgeons, Division of Neuroscience, NYSPI, 1051 Riverside Drive, New York, New York. 2 Department of Psychiatry, Columbia University College of Physicians and Surgeons, Division of Neu- roscience, NYSPI, 1051 Riverside Drive, New York, New York. 3 Department of Anatomy and Cell Biology, Columbia University College of Physicians and Surgeons, Division of Neuroscience, NYSPI, 1051 Riverside Drive, New York, New York. 4 To whom correspondence should be addressed at Department of Psychiatry, Columbia University Col- lege of Physicians and Surgeons, Division of Neuroscience, NYSPI, 1051 Riverside Drive, New York, New York 10032; e-mail: [email protected]. 219 0272-4340/04/0400-0219/0 C 2004 Plenum Publishing Corporation
Transcript of Secretin: Hypothalamic Distribution and Hypothesized Neuroregulatory Role in Autism

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
Cellular and Molecular Neurobiology, Vol. 24, No. 2, April 2004 ( C© 2004)
Secretin: Hypothalamic Distribution and HypothesizedNeuroregulatory Role in Autism
M. G. Welch,1,2,4 J. D. Keune,1,2 T. B. Welch-Horan,1,2 N. Anwar,1,2
M. Anwar,1,2 R. J. Ludwig,3 and D. A. Ruggiero1,2,3
Received June 19, 2003; accepted July 18, 2003
SUMMARY
1. This study aims (1) to determine whether secretin is synthesized centrally, specificallyby the HPA axis and (2) to discuss, on the basis of the findings in this and previous studies,secretin’s possible neuroregulatory role in autism.
2. An immunocytochemical technique with single-cell resolution was performed in12 age/weight-matched male rats pretreated with stereotaxic microinjection of colchicine(0.6 µg/kg) or vehicle into the lateral ventricle. Following 2-day survival, rats were anes-thetized and perfused for immunocytochemistry. Brain segments were blocked andalternate frozen 30-µm sections incubated in rabbit antibodies against secretin, vasoactiveintestinal peptide, glucagon, or pituitary-adenylate-cyclase-activating peptide. Adjacent sec-tions were processed for Nissl stain. Preadsorption studies were performed with membersof the secretin peptide family to demonstrate primary antibody specificity.
3. Specificity of secretin immunoreactivity (ir) was verified by clear-cut preadsorptioncontrol data and relatively high concentrations and distinct topographic localization of se-cretin ir to paraventricular/supraoptic and intercalated hypothalamic nuclei. Secretin levelswere upregulated by colchicine, an exemplar of homeostatic stressors, as compared withlow constitutive expression in untreated rats.
4. This study provides the first direct immunocytochemical demonstration of secretiner-gic immunoreactivity in the forebrain and offers evidence that the hypothalamus, like thegut, is capable of synthesizing secretin. Secretin’s dual expression by gut and brain secretincells, as well as its overlapping central distribution with other stress-adaptation neurohor-mones, especially oxytocin, indicates that it is stress-sensitive. A neuroregulatory relation-ship between the peripheral and central stress response systems is suggested, as is a dualrole for secretin in conditioning both of those stress-adaptation systems. Colchicine-inducedupregulation of secretin indicates that secretin may be synthesized on demand in responseto stress, a possible mechanism of action that may underlie secretin’s role in autism.
KEY WORDS: secretin; autism; HPA stress axis; secretinergic neurons; conditionedresponse; oxytocin; visceral; neuroregulatory.
1 Laboratories of Childhood Regulatory Disorders and Behavioral Neuroanatomy, Columbia UniversityCollege of Physicians and Surgeons, Division of Neuroscience, NYSPI, 1051 Riverside Drive, New York,New York.
2 Department of Psychiatry, Columbia University College of Physicians and Surgeons, Division of Neu-roscience, NYSPI, 1051 Riverside Drive, New York, New York.
3 Department of Anatomy and Cell Biology, Columbia University College of Physicians and Surgeons,Division of Neuroscience, NYSPI, 1051 Riverside Drive, New York, New York.
4 To whom correspondence should be addressed at Department of Psychiatry, Columbia University Col-lege of Physicians and Surgeons, Division of Neuroscience, NYSPI, 1051 Riverside Drive, New York,New York 10032; e-mail: [email protected].
219
0272-4340/04/0400-0219/0 C© 2004 Plenum Publishing Corporation

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
220 Welch, Keune, Welch-Horan, Anwar, Anwar, Ludwig, and Ruggiero
INTRODUCTION
To date, there is no comprehensive treatment for the broad range of autistic symp-tomatology: seizures (Park, 2003); attentional/arousal dysregulation and ADHD(Booth et al., 2003); obsessive–compulsive disorder (Hollander et al., 2003a); stereo-typies (Militerni et al., 2002); social isolation (Iqbal, 2002); attachment disorders(Kobayashi et al., 2001; Tinbergen and Tinbergen, 1983); face recognition deficits(Ogai et al., 2003, Schultz et al., 2003); gaze aversions (Richer and Coss 1976); gas-trointestinal disorders (Gershon, personal communication, 2003; Horvath et al., 1998;Horvath and Perman, 2002; Torrente et al., 2002); and altered heart rate variability(Corona et al., 1998; Graveling and Brooke, 1978). Current drugs directed at treatingthese symptoms have long-term side effects and efficacy not far above placebo rates(Posey and McDougle, 2000), resulting in motivation to seek new treatments. Thedevelopment of novel therapeutic measures targeting root causes of autism can ben-efit from combining understanding obtained at the bedside with knowledge gainedat the lab bench.
Clinical work, in collaboration with Nobel Laureate Niko Tinbergen (Tinbergenand Tinbergen, 1983, Welch, 1983a,b, 1989), has shown that autistic spectrum disor-ders are ameliorated by reinstating components of maternal nurturing, including theestablishment of synchronous attunement between mother and child (Welch 1983a,b,1987, 1988, 1989, Welch and Chaput, 1988). Mother–infant interaction appears to bea powerful stimulus to neuropeptide release (Matthiesen et al., 2001). Brain–gut neu-ropeptides contribute to developmental neuroregulation of growth, differentiationand regeneration, and the control of hormone release (Houben and Denef, 1994),as well as the resolution of visceral inflammation and brain activation in brain/gutdysregulation models (Welch et al., 2003b). These studies led to the hypothesis thatmaternal nurturing, as well as interventions that effectively replicate it, involvesameliorative mechanisms that stimulate neuropeptide release.
Our clinical and experimental work tests the hypothesis that some childhooddevelopmental abnormalities are spectrum disorders of brain/gut dysregulation thatcan be ameliorated by naturalistic and/or peptide therapy. Recent research has re-vealed pathology in the gastrointestinal tract of autistic children extending fromthe esophagus to the colon. This finding has led to investigations of impaired gut–immune system development, altered production of brain/gut peptides, increased in-testinal mucosal permeability, and inflammation (Nelson et al., 2001; Torrente et al.,2002; Warren et al., 1997; White, 2003). Other evidence has focused attention on thepossibility of homeostatic imbalance, such as altered central and peripheral energymetabolism in autistic children (Chugani et al., 1999), and dysregulation of pep-tide hormones that protect homeostasis (Gershon, personal communication, 2003;Hollander et al., 2003b; Nelson et al., 2001). It is our belief that effective clinical orpharmacokinetic intervention in autistic symptomatology will require a mechanismthat acts simultaneously upon the mind–brain–body stress axis to reestablish home-ostasis (Welch et al., 2003b). Metabolic imbalances in autism have been defined viafMRI (Haznedar et al., 2000). Naturalistic and/or peptide therapies (Hollander et al.,2003b; Horvath et al., 1998; Welch 1983a, b; Welch et al., 2003b) will be effective tothe extent that they address such imbalances.

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
Secretin: Hypothalamic Distribution and Hypothesized Neuroregulatory Role in Autism 221
Psychotherapeutic and pharmacologic measures (Diggle et al., 2003;Langworthy-Lam et al., 2002), including peptide neurohormone administration, havebeen attempted in autistic children, with limited outcomes (Coniglio et al., 2001;Dunn-Geier et al., 2000; Horvath et al., 1998; Kern et al., 2002; Koren, 2001; Lamsonand Plaza, 2001; Owley et al., 2001; Sandler et al., 1999; Roberts et al., 2001; Lightdaleet al., 2001). Research supports the importance of peptides in treating behavioral anddevelopmental disorders in autistics: at the bedside, through systemic peptide admin-istration (Hollander et al., 2003b; Horvath et al., 1998), in clinical studies (Matthiesenet al., 2001), and in experimental animals, through reinstating components of mater-nal nurturing, such as feeding, handling, and licking (Bredy et al., 2003; Francis et al.,2002). Experimental models show that feeding and handling ameliorate brain pathol-ogy resulting from the social-isolation stress of maternal deprivation (Anisman et al.,1998; Meaney et al., 1988, 1991). One peptide in particular, secretin, is associatedwith feeding and handling, a form of controlled restraint (Lauterbach et al., 1980,Mineo et al., 1990).
Secretin is a bioactive peptide synthesized by upper intestinal secretin S cells(Bloom et al., 1978; Chang et al., 1999; Miller et al., 1978; Paquette et al., 1982; Straussand Yalow, 1978). It is also synthesized in mice by the pancreas and colon (Lopezet al., 1995), and by flora that inhabit the gut (Gauthier et al., 2003). Whether secretinis synthesized by the forebrain is the subject of this study. Secretin belongs to thesecretin/vasoactive intestinal peptide (VIP)/glucagon receptor family with actionsat high and low-affinity secretin receptors (Ichihara et al., 1983). It is a 27 aminoacid peptide and an enterogastrone (Jin et al., 1994, Li et al., 1998). Secretin recep-tors couple to G proteins that stimulate adenylate cyclase and, in turn, lead to theproduction of cyclic adenosine monophosphate (cAMP) and the stimulation of as-sociated second messenger systems (Harmar, 2001). Secretin receptors concentratein brain regions (Itoh et al., 1991, Ohta et al., 1992) that are responsive to intracere-broventricular (i.c.v.) administration of the secretin peptide (Welch et al., 2002a,b,2003a). These regions are also sites of pathology in autism (Bauman and Kemper1985, Haznedar et al., 2000, Ogai et al., 2003, Schultz et al., 2003).
Secretin’s peripheral role as a gastric hormone has long been established (Baylissand Starling 1902); less is known about its central actions. Secretin regulates the cen-tral and peripheral stress axes via neurohumoral mechanisms (Ruggiero et al., 2003,Welch et al., 2002a,b, 2003a,b) that involve interactions with other signaling systemsacting at the level of the hypothalamus, such as secretin/angiotensin (Walker et al.,1999) and secretin/dopamine (Fuxe et al., 1979). Secretin functions to modulate HPAstress axis output, and, in contrast to VIP, increases norepinephrine and dopamineturnover in the hypothalamus and median eminence (Fuxe et al., 1979). Assays find-ing positive secretin bioactivity, radioimmunoreactivity, or secretin precursor mRNAexpression indicated high secretin levels in the hypothalamus and hypophysis, withthe preponderance of evidence suggesting that the hypothalamus is a site of origin ofendogenous secretin (Chang et al., 1985; Charleton et al., 1981; Fuxe et al., 1979; Itohet al., 1991; Mutt et al., 1979; Nussdorfer et al., 2000; O’Donohue et al., 1981; Ohta et al.,1992; Samson et al., 1984). Another study assessing the presence of secretin in the ratbrain and gut did not find central expression of the bioactive peptide (Kopin et al.,1990). These studies, however, lacked the single-cell resolution needed to precisely

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
222 Welch, Keune, Welch-Horan, Anwar, Anwar, Ludwig, and Ruggiero
delineate the organization of a presumptive secretinergic system. Recently, studiesfound secretin immunoreactivity in the brain stem and cerebellum (Koves et al.,2002; Ng et al., 2002; Yung et al., 2001), but not in the forebrain. According to Ng,“secretin is only present at detectable levels in the brain stem and cerebellum,” al-though unpublished data suggest “the presence of secretin-producing cells in thecerebral cortex” (Ng et al., 2002).
Prior studies led our laboratory to test the hypothesis that secretin regulatesstress response patterns via endogenous synthesis along the hypothalamic stressaxis. Central secretin administration (i.c.v.) activates the area postrema, nucleus ofthe solitary tract (NTS), and its terminal fields, including the parabrachial complex,amygdala, and hypothalamus. In addition, secretin activates the visceral thalamusand its insula/orbital and medial prefrontal cortical projection fields, which regu-late visceral reflex networks overlapping areas of pathology in autism (Welch et al.,2002a,b, 2003a). Corroborating some of these results was a report focused on the ef-fects of secretin-induced c-fos gene expression in the amygdala of rats (Goulet et al.,2003). In a subsequent study, long-term systemic administration of bioactive peptides,including trials with secretin, was found to resolve inflammatory bowel lesions andstress-related effects on specific CNS regions in mice and rats that correspond to sitesof pathology in autism (Welch et al., 2003b). Systemic exogenous secretin was foundto reestablish communicative and affiliative interactions in autistic children with gas-trointestinal abnormalities (Horvath et al., 1998), an observation supported in a sub-sequent case report (Lamson and Plaza, 2001). This interest in secretin has led to a cur-rent investigative effort to replicate Horvath’s novel peptide therapy (Wheeler 2003).
To date, there has been no study with single-cell resolution identifying secretiner-gic neurons in the forebrain. In this study, we sought to determine whether secretinis synthesized specifically in the forebrain, and whether its specificity and distribu-tion patterns might predict possible interactions of secretin with other hormonesinvolved in stress adaptation. Additionally, we investigated whether secretin is syn-thesized on demand in distinct areas of the HPA stress axis of rats. The centraldistribution of secretin immunoreactivity in adult male Sprague–Dawley rats wasmapped to single-cell resolution using immunocytochemistry. To demonstrate se-cretin’s specificity, the experiments included preadsorption controls, as well as stud-ies cross-comparing the distribution of secretin and other members of its peptidefamily, including VIP, glucagon, and pituitary-adenylate-cyclase-activating peptide(PACAP).
METHODS
This study was approved by the Columbia University Institutional Animal Careand Use Committee. Experiments were performed on 12 male Sprague–Dawley rats(250–300 g) housed in the New York State Psychiatric Institute animal facility andmaintained in a thermally controlled, light-cycled environment with lab chow andwater ad libitum. The experiments were performed according to NIH guidelines.
Colchicine pretreatment was necessary to identify secretin immunoreactivityproduct in neurons expressing constitutively low basal secretin immunolabeling as

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
Secretin: Hypothalamic Distribution and Hypothesized Neuroregulatory Role in Autism 223
shown in pilot studies of untreated animals (n = 6). Colchicine, a metabolic stressor,acts by disrupting the microtubule network and by blocking axonal transport, leadingto a buildup of cytoplasmic peptide content in quantities adequate for immunocy-tochemical detection (Aguado et al., 1999). Colchicine was injected into the lateralventricle; 2 days later, brain tissue sections were immunocytochemically processedto detect secretin.
Rats were deeply anesthetized by intraperitoneal (i.p.) administration of ke-tamine/xylazine, and 10µL of colchicine solution (15µg/µL, 0.9% saline) was slowlymicroinjected unilaterally into the right lateral ventricle through a glass pipette in-serted via a burr hole in the skull. Age-matched controls were administered equalvolumes of physiological saline. Wounds were closed and animals were returned totheir cages. After 48 h, animals were deeply reanesthetized with ketamine/xylazinei.p. and perfused transcardially with heparinized, physiological saline followed by afixative containing 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4). Theforebrain was blocked and removed from each animal, postfixed for 1 h in the samefixative and cryoprotected overnight at 4◦C in 0.1 M phosphate buffered saline (pH7.4) containing 10% sucrose. Frozen tissues were sectioned in the transverse planeat 30 µm on a sledge microtome and collected in phosphate buffer. Free-floatingserial sections were incubated for 18–24 h in commercially available, highly specificpolyclonal rabbit antibodies raised against secretin (Chemicon International, Inc.,Temecula, CA), VIP, PACAP, or glucagon (Phoenix Pharmaceuticals, Inc., Belmont,CA). The antisera were diluted (1:10,000) in Tris-buffered saline (TBS) containing0.1% bovine serum albumen to which 0.25% Triton X-100 was added to facilitatetissue penetration. Alternate tissue sections were rinsed in TBS (3× 15 min), in-cubated (1 h) in 1% BSA in TBS, and processed immunohistochemically, using animmunoperoxidase ABC technique. Tissues were sequentially incubated in biotiny-lated goat anti-rabbit immunoglobulin (IgG) secondary antibody (1:200, 60 min)and avidin–biotin peroxidase complex (1:100, 60 min) (Vector, Burlingame, CA).Immunocytochemically processed sections were washed in Tris (3× 15 min), andtreated with 0.05% diaminobenzidine hydrochloride in dilute phosphate buffer con-taining 0.003% hydrogen peroxide to develop the reaction. Sections were mountedon 1% gelatin-coated slides, air-dried, and coverslipped without counterstaining. Aset of adjacent sections were Nissl-stained to delineate nuclear borders. Sections wereexamined by light microscopy and digitized images were obtained with a DiagnosticInstruments SPOT-RT Slider camera mounted on a Nikon Microphot microscope.Forebrain nuclei were identified by reference to Nissl-stained sections and stereotaxicatlases of the rat brain (Paxinos and Watson 1998, Swanson 1998).
Controls
The specificity of the secretin staining pattern was determined by internal con-trols comparing the distribution patterns of secretin, VIP, PACAP, and glucagon.Controls for the specificity of the antibodies included (1) omission of the primaryantibodies and (2) substitution of normal rabbit serum for the primary antiserumand preincubation of the secretin antibody by preadsorption with secretin peptide(Sigma) or VIP or PACAP peptide (Phoenix Pharmaceuticals). Working solution

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
224 Welch, Keune, Welch-Horan, Anwar, Anwar, Ludwig, and Ruggiero
containing each of the peptides in 0.1 M TBS/1% bovine serum albumen, pH 7.55, wasincubated on a slow shaker table at 4◦C overnight with the primary secretin antibody(dilution 1:10,000). Incubation of the conjugate, 1:10 ratio of secretin Ab/peptide(secretin, VIP, or PACAP), was carried out and the reaction was allowed to pro-ceed for 24 h, followed by 10-min centrifugation. The tissue sections were incubatedin the resulting supernatant overnight at room temperature, followed by standardimmunocytochemical methods.
RESULTS
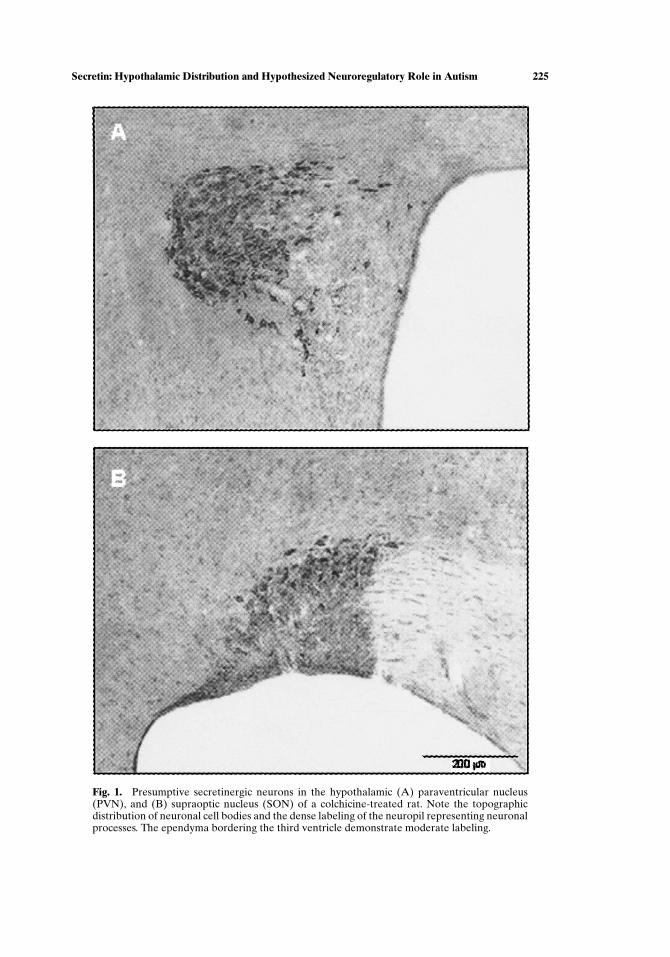
Secretinergic neurons were heavily labeled in colchicine-treated rats, as com-pared with the untreated group, which showed light to undetectable labeling. Secretinimmunoreactivity was cytoplasmic and restricted to neurons of the anterior and mid-dle regions of the hypothalamus and adjoining periventricular gray. The hindbrain,cerebellum, and spinal cord were not examined in this study. Presumptive secretiner-gic neurons were concentrated in precise loci within the paraventricular/supraopticand intercalated regions of the hypothalamus (Fig. 1). Secretinergic cells were heav-ily concentrated and intensely stained in both the peripheral and central core of theparaventricular nucleus magnocellularis, and labeled in its medial extensions intothe parvicellular and periventricular divisions. Secretinergic cells in the supraop-tic nucleus (SON) were concentrated dorsally and extended medially, arching overthe optic tract. Small numbers of cells were scattered among a heavily labeled neu-ropil in the ventral region of the SON. Arrays of cells were diagonally organizedin the nucleus intercalatus. High-power optics revealed secretinergic neurons withsubependymal processes in the periventricular gray deep to the ependyma (Fig. 2).
Controls: Preadsorption
The specificity of the secretin immunoreaction product was verified. Secretinimmunoreactivity was not detected on tissue sections incubated in secretin antibodythat had been preadsorbed with the secretin peptide as shown in the PVN (Fig. 3).Secretinergic neurons and ependyma were immunolabeled on alternate tissue sec-tions that were incubated in secretin antibody preadsorbed with another member ofthe secretin peptide family: VIP or PACAP.
Controls: Cross Comparisons
Distribution patterns of secretin immunoreactive neurons were distinct fromthose on tissues incubated in antibodies raised against the other members of thepeptide family (Fig. 4). These dramatic differences in topographic distribution anddensity served as an internal control in colchicine-treated animals. Their locations dif-fered from the distributions of other members of the secretin/VIP/glucagon/PACAPfamily, as delineated on serially sectioned alternate tissues processed with the re-spective antibodies raised against each peptide. Secretinergic neurons in PVN andSON far outnumbered cells staining for the other peptides, including VIP.
P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
Secretin: Hypothalamic Distribution and Hypothesized Neuroregulatory Role in Autism 225
Fig. 1. Presumptive secretinergic neurons in the hypothalamic (A) paraventricular nucleus(PVN), and (B) supraoptic nucleus (SON) of a colchicine-treated rat. Note the topographicdistribution of neuronal cell bodies and the dense labeling of the neuropil representing neuronalprocesses. The ependyma bordering the third ventricle demonstrate moderate labeling.

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
226 Welch, Keune, Welch-Horan, Anwar, Anwar, Ludwig, and Ruggiero
Fig. 2. Evidence suggestive of secretin exchange across the CSF/ependymal interface of the thirdventricle. A secretinergic neuron in the rat periventricular gray in proximity to the ependyma ischaracterized by diffuse cytoplasmic immunoreactivity. Preadsorption controls verified the labelingof neuronal cells. Theoretically, secretin may be secreted into, or extracted from, the CSF. Thepeptide is upregulated on demand and secreted into the CSF and the blood stream, and releasedby terminals of hypothalamic/pituitary/bulbar/spinal projections. Suspected interactions may occurwith classical neurotransmitter systems, such as serotonin and dopamine, and other stress-regulatorypeptides, such as CRH, AVP, and OT.
VIP labeling was expressed in superficial and deep layers of the cerebralcortex and striatum with a distinct distribution in the anterior cingulate and con-tiguous medial prefrontal cortex and lateral prefrontal cortex. VIP staining waslight and diffuse in the SON and weak in the PVN with the exception of intenselystained cells and neuropil in the hypothalamic suprachiasmatic nucleus (Fig. 5).The caudate and putamen exhibited small-spindle-shaped cells that resembledinterneurons.
Glucagon appeared in the retrochiasmatic nucleus and extended dorsally intothe subparaventricular zone of the hypothalamus. Glucagon immunoreactivity alsoappeared in the internal and external lamina of the median eminence, and in theneuropil of PVN, SON, caudate, and putamen. Further contrasting secretin-stainingpatterns was heavy labeling of the basolateral amygdala.

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
Secretin: Hypothalamic Distribution and Hypothesized Neuroregulatory Role in Autism 227
Fig. 3. Preadsorption study in paraventricular hypothalamus (PVN). (A) Secretinergic neurons inPVN on cross-sectioned tissue incubated in secretin antibody. Specificity was verified by absence ofimmunoreactivity on tissue incubated in the (B) secretin peptide/antibody complex, and the robustlabeling of secretinergic hypothalamic neurons on tissues incubated in antibody preadsorbed with(C) VIP or (D) PACAP.
PACAP staining was dramatically different from the secretin patterns, exhibit-ing high concentrations in processes of the basolateral and lateral amygdala andlighter labeling in the central nucleus. PACAP was present in PVN cells butskewed medially in the parvicellular region. Scattered labeling in the SON wasconcentrated ventrally, where the bulk of the labeling appeared to be in theneuropil.
Secretinergic cells were not detected in regions containing moderate to highconcentrations of glucagon, VIP, or PACAP. These other peptides had distinctivedistribution patterns and were localized to regions devoid of secretin or, where therewas overlap, were characterized by low levels of secretin immunoreactivity. Locienriched in secretin immunoreactivity were devoid of, or characterized by low levelsof, glucagon or PACAP immunoreaction products.

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
228 Welch, Keune, Welch-Horan, Anwar, Anwar, Ludwig, and Ruggiero
Fig. 4. Distribution patterns of the secretin peptide family in the paraventricular nucleus (PVN) ofa colchicine-treated rat. Secretinergic neurons in the hypothalamus on tissues incubated in a rabbitpolyclonal antibody raised against (A) secretin, (B) VIP, (C) glucagon, and (D) PACAP. They werecharacterized by different distributional and density patterns. Specificity of the antibody was verifiedby the cross-comparisons of the secretin family of peptides and by internal controls.
DISCUSSION
This study provides the first single-cell immunocytochemical demonstration ofsecretinergic immunoreactivity in the hypothalamus of a rat, extending earlier dataobtained using less sensitive techniques (Chang et al., 1985; Charlton et al., 1981; Muttet al., 1979; O’Donohue et al., 1981; Samson et al., 1984). On the basis of observationsmade in this study, theoretical stress-regulatory mechanisms of secretin are discussed.A novel concept for the neurobiological basis of secretin’s role in autism is createdby synthesizing previous medical and experimental literature with our findings.
Secretin Is Synthesized by Neurons in the Hypothalamus
Secretin-like neuronal immunoreaction product is restricted to the hypothala-mus and adjoining periventricular gray. Secretin immunolabeling in this study wasneuronal and restricted to precise loci within the paraventricular/supraoptic nucleiand adjoining periventricular gray (Fig. 1). Our control studies support specificityof secretin immunoreactivity. Dramatic differences in topographic distribution and

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
Secretin: Hypothalamic Distribution and Hypothesized Neuroregulatory Role in Autism 229
Fig. 5. Distribution patterns of secretin and VIP in suprachiasmatic hypothalamic nucleus (SCN) ina colchicine-treated rat. SCN demonstrates weak neuropil labeling on tissues incubated in the secretinantibody: (A) secretin under low power, (C) secretin under high power. By contrast, the SCN wascharacterized by dense VIP immunoreactive somata and processes: (B) VIP low power, (D) VIP highpower.
density of secretinergic neurons from the distribution patterns of other membersof the secretin/VIP/glucagon/PACAP family extend evidence of the existence of asecretinergic brain/gut stress-regulatory system. The number of cells staining for se-cretin far outnumbered those staining for VIP, glucagon, and PACAP. Yet anotherrelated peptide, hypocretin, shares substantial amino acid identities with secretin.However, hypocretin, in contrast to secretin, localizes to neuronal cell bodies in thedorsal posterior hypothalamus and amygdala (de Lecea et al., 1998, Ciriello et al.,2003).
Previous studies using immunocytochemical techniques failed to find secretinin the hypothalamus (Koves et al., 2002, Ng et al., 2002). Our data, however, whichwere supported by extensive control experiments, support earlier studies that foundhypothalamic secretin expression (Fuxe et al., 1979). Secretin precursor protein isabundant in the hypothalamus and hippocampus (Itoh et al., 1991), sites of pathol-ogy in autism (Bauman and Kemper 1985). These data suggest that secretin mayserve as a bioactive stress-regulatory peptide in the rat brain, specifically alongthe HPA stress axis. The fact that the hypothalamus plays a key role in regulat-ing brain/body functions leads us to consider secretin’s relationship to varioushomeostatic functions.

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
230 Welch, Keune, Welch-Horan, Anwar, Anwar, Ludwig, and Ruggiero
Fig. 6. Swanson brain maps: Schematic level 25. Secretinergic neurons shown in previous fig-ures were localized to the paraventricular (PVN) and supraoptic (SON) hypothalamic subnucleidepicted at this forebrain level.
Secretin Regulates Stress Centrally as Well as Peripherally
Secretin has been predicted to be a stress-regulatory peptide (Fuxe et al., 1979,Itoh et al., 1991). Secretin’s role as a peripheral stress-regulatory hormone and centralneuromodulator of stress-adaptation responses has been suggested by earlier struc-tural and functional studies (Chang et al., 1985; Charlton et al., 1983; Mutt et al., 1979;O’Donohue et al., 1981; Samson et al., 1984). Since secretin is expressed along theHPA stress axis, it is our contention that the neurohormone receptor expression andits interactions with other transmitters/modulators should be studied in relationship

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
Secretin: Hypothalamic Distribution and Hypothesized Neuroregulatory Role in Autism 231
to homeostasis. In particular, the fact that secretin was upregulated by colchicineprovides evidence of secretin’s central expression. In our study, secretin staining inthe brain was scant in intact rats, reflecting the fact that under normal physiologicalconditions the level of central secretin in the adult rat may be low. However, in ourstudy, the experimental rats were exposed to colchicine, which acts as a powerfuloxidative stressor. In cases of toxicity, it has been shown to cause respiratory andrenal failure in the treatment of gout, impairments of cognition, learning and mem-ory (Veerendra-Kumar and Gupta, 2002), and even death in humans (Jones et al.,2002). Secretin levels are elevated in other conditions of stress, such as severe dy-namic exercise (Bell et al., 1984; Oektedalen et al., 1982) and restraint stress/hypoxia(Lauterbach et al., 1980). This observation leads us to speculate that secretin mayhave been upregulated by colchicine because the latter compound is a physiologicalstressor.
Stress-Related Dysregulation of Brain–Gut Peptides in Autism
No study to date has measured bioactive peptide production levels in the autis-tic brain. In animal models, colchicine upregulation of secretin, as well as oxytocin(Bojanowska et al., 1999), may replicate the effects of stress on brain/gut peptides inautistics. Clinical evidence suggests dysregulation of peripheral VIP in neonates laterdiagnosed as autistic (Nelson et al., 2001). These findings lead us to question whetherother stress-regulatory peptides are dysregulated in autism. If so, such dysregulationcould explain the reported effectiveness of hormone replacement therapies, such assecretin (Horvath et al., 1998) and oxytocin administration (Hollander et al., 2003b),in ameliorating autistic symptoms. Autism, along with other developmental disor-ders, has long been linked to adverse intrauterine environmental conditions, suchas hypoxic–ischemic insult (Davis et al., 1992); such insults have been causally re-lated to brain/gut pathology (Barlow and Santulli, 1975). Since hypoxia stimulatessecretin release (Lauterbach et al., 1980), it is possible that the failure of autisticinfants to communicate and affiliate is in part a consequence of visceral disorders(Horvath et al., 1999), including abnormalities of secretinergic systems (Gershon,personal communication, 2003). In such cases, adverse visceral environmental con-ditioning of brain/gut response patterns may be related to the pathophysiology ofautism, including perceptual, social, and intellectual impairments, and stress-relatedsymptomatology.
It is well established that cognitive and emotional networks are dysregulatedin autism (Rumsey and Ernst, 2000), and that these networks control visceral re-flex function at the ventral and dorsal vagal complex (Loewy, 1991; Ruggiero et al.,1985). Moreover, the HPA stress axis and the sympathetic projections to the visceraare hyperdriven by high unremitting stress (McEwen, 2001; Steptoe et al., 1999).Secretinergic cells may be similar to epinephrine neurons, which exert powerful in-fluence over behavioral and visceral reflex functions (Sved et al., 1994). Both areexquisitely sensitive to chronic homeostatic perturbations, such as hypoxia, and areupregulated by colchicine (Ruggiero et al., 1985, 2003).
It is conceivable that stress-related hyperproduction of secretin mightremodel the secretin receptor complex. The endocytic process of secretin receptor

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
232 Welch, Keune, Welch-Horan, Anwar, Anwar, Ludwig, and Ruggiero
internalization, identified by Walker (Walker et al., 1999), could be compromisedover time by such remodeling, which in turn could compromise secretinergic func-tion. Some recent research on secretin-mediated gene delivery supports this concept(McKay et al., 2002), although much additional work needs to be done to test such ahypothesis conclusively.
Stress-Related Dysregulation of the GI Tract in Autism
Under conditions of unremitting stress, or in response to chronic homeostaticperturbations, the secretin S cells, like epinephrine cells, may be activated on de-mand (Sved et al., 1994) and thus driven to depletion or apoptosis through overuse.Indeed, Gershon found markedly fewer S cells in the upper gastrointestinal tracts ofautistics, as well as fewer cells colocalizing secretin and serotonin (Gershon, personalcommunication, 2003). In addition, secretin levels are elevated in diseases such ascystic fibrosis, hyaline membrane disease (Boccia et al., 2001), and Crohn’s disease(Teufel et al., 1986), all of which are associated with gastrointestinal abnormalities.Friedman found that hypoxic gut epithelial cells were impaired in their ability togenerate cAMP, and that pharmacologic elevation of cAMP in hypoxic cells bothnormalized polymorphonuclear-induced permeability changes and restored barrierfunction (Friedman et al., 1998). Such permeability changes have been found in autis-tic children (D’Eufemia et al., 1996), and Horvath reported that secretin, which hasbeen shown to elevate cAMP (Fremeau et al., 1986), decreased intestinal permeabil-ity in 13 of 20 autistic children (Horvath and Perman, 2002).
Clinical Support for the Hypothesis That Secretin Is Synthesizedon Demand in Response to Stressors
Secretin-releasing peptide and secretin are secreted as part of unified, vagallymediated behavioral and reflex response patterns (Chey and Chang, 2001). Thesepeptides are triggered by stress-related increases in gastrin and gastric acid output(Li et al., 1998). Interestingly, this reflex response pattern is viscerally conditioned bycomponents of maternal nurturing, such as breast-feeding and vocalization, whichcondition a child to alternate between swallowing and breathing, or between talkingand listening (Porges, 1995). Stimuli, such as sucking, feeding, and tactile visceroso-matic stimulation, activate periventricular/tegmental pathways which are predictablyinvolved in the development and differentiation of visceral memory. Behavioral, en-docrine and autonomic networks that are assembled postnatally (Rinaman et al.,2000) were found to respond to secretin in the present study (Welch et al., 2002a,b).Our findings indicate that these networks could have been dysregulated by visceraldisorders/stress, and call for further investigation.
Synthesis and secretion of secretin on demand may be operative in ameliorationof perinatal stress (Lucas et al., 1983; Rogers et al., 1975) via two mechanisms of action:as a vasodilator (Gandhi et al., 2002) and as a gastric protective hormone (Baylissand Starling, 1902). Brain stem tegmental pathways vasodilate cerebral blood vessels(Golanov et al., 2000). Human neonates exhibit excessive gastric acid and secretinoutput, as assayed in 2-day-old neonates, whereas secretin expression reaches the

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
Secretin: Hypothalamic Distribution and Hypothesized Neuroregulatory Role in Autism 233
mature pattern by the second postnatal week in healthy preterm infants (Lucas et al.,1980). Sick infants with hyaline membrane disease exhibit sustained upregulationof secretin secondary to both starvation (Lucas et al., 1980) and respiratory stressanalogous to that induced by colchicine (Jones et al., 2002).
Secretin is secreted in response to milk ingestion (Mineo et al., 1990). Thecolostrum content of breast milk during the immediate post-partum period is aneven more powerful stimulus to secretin than is milk itself (Guilloteau et al., 1992).There is a relationship between autism and maternal withdrawal from nursing: stud-ies on infantile autism revealed a lower incidence of breastfeeding (Tanoue, andOda, 1989). Brain/gut peptides involving a behaviorally modulated vagal reflex arereleased in response to breastfeeding (Eriksson et al., 1994); consequently, such pep-tides are implicated in the acute and long-term effects of breastfeeding. These find-ings suggest that early weaning may contribute to the etiology of autism. Further,they suggest that restoration of components of maternal nurturing (Tinbergen andTinbergen, 1983; Welch, 1983a,b, 1987, 1988, 1989; Welch and Chaput, 1988) and/orpeptide therapy (Welch et al., 2003b) might replicate the protective effects of holdingand/or breastfeeding, as found in Barlow’s studies (Barlow and Santulli, 1975). Thecombined therapies of reinstatement of components of maternal nurturing and pep-tide hormones should be evaluated by neuroimaging and assays of brain/gut peptidelevels in both mother and autistic child.
Interactions of Secretin With Other Peptides and Neurotransmitters
Contemporary research investigates the function of visceral reflex circuits re-sponsible for homeostasis (Agassandian et al., 2002, Talman and Kelkar, 1993,Wasserman et al., 2002). Signaling molecules interact in the regulation of homeosta-sis and the stress response, notably via the paraventricular hypothalamus (Leonget al., 2002). Relevant interactions between classical neurotransmitters and peptidesinclude dopamine and secretin, norepinephrine and secretin (Fuxe et al., 1979), an-giotensin and secretin (Walker et al., 1999), epinephrine, norepinephrine and an-giotensin (Jezova et al., 2003; Li and Guyenet, 1996), norepinephrine, serotonin andoxytocin (Vacher et al., 2002) and oxytocin and atrial natriuretic peptide (Chrigueret al., 2001).
An example relevant to gastrointestinal disorders is the relationship betweenangiotensin and secretin receptors. Angiotensin II AT(1) receptors and secretin re-ceptors are colocalized in endocytic vesicles (Walker et al., 1999). Angiotensin IIAT(1) receptors are linked to social isolation stress (Armando et al., 2001). Antago-nizing AT(1) receptors restores modulation of HPA stress axis function (Armandoet al., 2001) and prevents gastric mucosal injury (Bregonzio et al., 2003). Social isola-tion stress is a causal factor in hemorrhagic stress ulcers and hippocampal formationpathology among insubordinate vervets (Uno et al., 1989). Autistic children andinsubordinate vervets both experience social isolation and share noncompliance be-haviors (Breiner and Beck, 1984). The cingulate/hippocampal stress-adaptation net-work is also a major site of pathology for both autistic children and vervets (Baumanand Kemper, 1985, Uno et al., 1989). This network is implicated in the adverse con-ditioning of both gastrointestinal functions (Gabry et al., 2002; Uno et al., 1989) and

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
234 Welch, Keune, Welch-Horan, Anwar, Anwar, Ludwig, and Ruggiero
behavioral functions (Freeman et al., 1997; Jones and Gonzalez-Lima, 2001). Secretin,which activates the cingulate/hippocampal stress-adaptation network (Welch et al.,2003a), has been effective in ameliorating both gut and behavioral abnormalities ofautistic children (Horvath et al., 1998; Horvath and Perman, 2002; Lamson, 2001).Still to be investigated is whether secretin levels are altered by the social isolation ofautistics, and whether secretin might be therapeutic in Uno’s insubordinate primatestress-ulcer model (Uno et al., 1989).
Taken together, these findings suggest that secretin, a vasodilator (Gandhi et al.,2002), could be modulating the effect of angiotensin II AT(1), a vasoconstrictor(Helou et al., 2003). This modulation may take place, as Leong suggests, by “cross-talk” (interaction) of peptides at the level of the hypothalamus (Leong et al., 2002).If secretin is modulating AT(1) receptors, this interaction may explain why secretinameliorates both GI pathology (Horvath and Perman, 2002, Welch et al., 2003b) andsymptoms of autism (Horvath et al., 1998). These findings also support our theorythat autistic symptomatology is related to dysregulation of a single, unified mind–brain–body stress axis, one that is normally conditioned by peptide–peptide andpeptide–neurotransmitter interactions. If secretin is dysregulated, then we hypoth-esize that its central and peripheral networks and transmitter interactions, such aswith norepinephrine, dopamine (Fuxe et al., 1979), angiotensin (Walker et al., 1999),and serotonin (Gershon, 1998; Gershon, personal communication, 2003), could bedysregulated as well.
CONCLUSION
This study provides the first direct immunocytochemical demonstration of se-cretinergic immunoreactivity in the forebrain, and provides strong evidence thatthe hypothalamus, like the gut, is capable of synthesizing secretin. Since secretin isexpressed along the HPA stress axis, the neurohormone receptor expression andits interactions with other transmitters/modulators deserve study in relationship tohomeostasis.
The medical and experimental literatures, taken together with our data, suggestthat secretin, along with other neuropeptides, plays a role in conditioning stress-adaptation patterns. These findings lend support for the existence of a neuroregula-tory functional interdependence (synchronization) of the central/peripheral stress-response systems. Our study demonstrated localizations of secretin that overlap withthose of other stress-regulatory neurohormones, especially oxytocin (Vacher et al.,2002; Welch, unpublished data ). Given secretin’s expression by gut and brain cells,secretin may have an integrated peripheral and central stress-related function inmaintaining homeostasis. Colchicine, an oxidative stressor, induces upregulation ofsecretin. Therefore, we conclude that secretin may be synthesized on demand inresponse to homeostatic challenges. If secretin upregulates in response to visceralstressors, this finding would shed new light on an important potential mechanism ofaction in autism.
Although the etiology of autism is poorly understood, as is the link betweenautism and gastrointestinal abnormalities (Gershon, personal communication, 2003;Horvath and Perman, 2002; Torrente et al., 2002), our findings suggest that secretin

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
Secretin: Hypothalamic Distribution and Hypothesized Neuroregulatory Role in Autism 235
may in some cases ameliorate dysregulated gastrointestinal function in autism byacting upon or influencing the brain and visceral processing networks simultaneouslyas a single unit. Still in question is whether the autistic gut is dysregulated prior to, orsubsequent to, the onset of the cognitive disorder, or whether one or both systems aredevelopmentally abnormal. Also in question is whether secretin is altered in autisticsexperiencing social isolation.
Future Directions
Our laboratory’s preliminary studies of the brain/gut axis have identified re-gional brain activation patterns secondary to gastrointestinal inflammation in bothacquired and genetic animal models of inflammatory bowel disease (IBD), and haveassessed the extent of their resolution by peptide administration (Welch et al., 2003b).While these studies offer further support for secretin’s dual role as both a peripheralstress-regulatory hormone and a central neuromodulator of stress-induced responses,future studies must determine whether secretin peptide production is simultaneouslyaugmented in the brain and in the periphery. Whether other secretin/glucagon/VIPpeptide family members and other peptides act together as part of a mind–brain–body stress communication and regulatory network also remains to be investigated.
The relevance to autism and to the link between autism and visceral disordersmay emerge from future clinical and experimental studies (Welch et al., 2003b).For instance, in a large retrospective study, neonates later diagnosed with autismwere shown to have abnormally high levels of two gut–brain peptides, VIP, and/orcalcitonin-gene-related peptide, as well as neurotrophic factor (NT4), and/or brain-derived neurotrophic factor (Nelson et al., 2001). Peptide dysregulation is also as-sociated with IBD (Teufel et al., 1986). Since secretin was not evaluated in Nelson’sstudy of neonates, these findings invite a similar investigation of secretin. Also tobe studied are the pharmacokinetic administration of peptide combinations to bothautistic children and patients with IBD and the effects of secretin on other trans-mitters and peptides. Inasmuch as autistic children have responded to neuropeptidetreatment, as well as to restoration of components of maternal nurturing, as statedabove, these combined therapies should be evaluated via neuroimaging and assaysof brain/gut peptide levels before and after treatment.
ACKNOWLEDGMENTS
This work was supported by Columbia University Grants # 664613, 643622(mgw), and NIH RO1 NS #36363 (dar).
REFERENCES
Agassandian, K., Fazan, V. P., Adanina, V., and Talman W. T. (2002). Direct projections from the car-diovascular nucleus tractus solitarii to pontine preganglionic parasympathetic neurons: A link tocerebrovascular regulation. J. Comp. Neurol. 452(3):242–254.
Aguado, F., Pozas, E., and Blasi, J. (1999). Colchicine administration in the rat central nervous systeminduces SNAP-25 expression. Neuroscience 93(1):275–283.

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
236 Welch, Keune, Welch-Horan, Anwar, Anwar, Ludwig, and Ruggiero
Anisman, H., Zaharia, M. D., Meaney, M. J., and Merali, Z. (1998). Do early-life events permanently alterbehavioral and hormonal responses to stressors? Int. J. Dev. Neurosci. 16(3/4):149–164.
Armando, I., Carranza, A., Nishimura, Y., Hoe, K. L., Barontini, M., Terron, J. A., Falcon-Neri, A., Ito, T.,Juorio, A. V., and Saavedra, J. M. (2001). Peripheral administration of an angiotensin II AT(1) receptorantagonist decreases the hypothalamic–pituitary–adrenal response to isolation stress. Endocrinology142(9):3880–3889.
Barlow, B., and Santulli, T. V. (1975). Importance of multiple episodes of hypoxia or cold stress on thedevelopment of enterocolitis in an animal model. Surgery 77(5):687–690.
Bauman, M., and Kemper, T. L. (1985). Histoanatomic observations of the brain in early infantile autism.Neurology 35(6):866–874.
Bayliss, W. M., and Starling, E. H. (1902). The mechanism of pancreatic secretion. J. Physiol. (Lond.)28:325–353.
Bell, P. M., Henry, R. W., Buchanan, K. D., and Alberti, K. G. (1984). The effect of starvation on the gastro-entero-pancreatic hormonal and metabolic responses to exercise. (GEP hormones in starvation andexercise). Diabete Metab. 10(3):194–198.
Bloom, S. R., Mitchell, S. J., Greenberg, G. R., Christofides, N., Domschke, W., Domschke, S., Mitznegg,P., and Demling, L. (1978). Release of VIP, secretin and motilin after duodenal acidification in man.Acta Hepatogastroenterol. (Stuttg.) 25(5):365–368.
Boccia, D., Stolfi, I., Lana, S., and Moro, M. L. (2001). Nosocomial necrotising enterocolitis outbreaks:epidemiology and control measures. Eur. J. Pediatr. 160(6):385–391.
Bojanowska, E., Juszczak, M., Guzek, J. W., and Dabrowski, R. (1999). The pineal and oxytocin synthesis.J. Physiol. Pharmacol. 50(1):121–128.
Booth, R., Charlton, R., Hughes, C., and Happe, F. (2003). Disentangling weak coherence and executivedysfunction: Planning drawing in autism and attention-deficit/hyperactivity disorder. Philos. Trans.R. Soc. Lond. B Biol. Sci. 358(1430):387–392.
Bredy, T. W., Humpartzoomian, R. A., Cain, D. P., and Meaney, M. J. (2003). Partial reversal of the effectof maternal care on cognitive function through environmental enrichment. Neuroscience 118(2):571–576.
Bregonzio, C., Armando, I., Ando, H., Jezova, M., Baiardi, G., and Saavedra, J. M. (2003). Anti-inflammatory effects of angiotensin II AT1 receptor antagonism prevent stress-induced gastric injury.Am. J. Physiol. Gastrointest. Liver Physiol. 285(2):G414–G423.
Breiner, J., and Beck, S. (1984). Parents as change agents in the management of their developmentallydelayed children’s noncompliant behaviors: A critical review. Appl. Res. Ment. Retard. 5(2):259–278.
Chang, T. M., Berger-Ornstein, L., and Chey, W. Y. (1985). Presence of biologically and immunologicallyactive secretin-like substance in the mammalian brain. Peptides 6(2):193–198.
Chang, T. M., Chang, C. H., Wagner, D. R., and Chey, W. Y. (1999). Porcine pancreatic phospholipase A2stimulates secretin release from secretin-producing cells. J. Biol. Chem. 274(16):10758–10764.
Charlton, C. G., Miller, R. L., Crawley, J. N., Handelmann, G. E., and O’Donohue, T. L. (1983). Secretinmodulation of behavioral and physiological functions in the rat. Peptides 4(5):739–742.
Charlton, C. G., O’Donohue, T. L., Miller, R. L., Jacobowitz, D. M. (1981). Secretin immunoreactivity inrat and pig brain. Peptides 2(Suppl. 1):45–49.
Chey, W. Y., and Chang, T. (2001). Neural hormonal regulation of exocrine pancreatic secretion. Pancre-atology 1(4):320–335.
Chriguer, R. S., Rocha, M. J., Antunes-Rodrigues, J., and Franci, C. R. (2001). Hypothalamic atrial natri-uretic peptide and secretion of oxytocin. Brain Res. 889(1/2):239–242.
Chugani, D. C., Sundram, B. S., Behen, M., Lee, M. L., and Moore, G. J. (1999). Evidence of altered energymetabolism in autistic children. Prog. Neuropsychopharmacol. Biol. Psychiatry 23(4):635–641.
Ciriello, J., Rosas-Arellano, M. P., Solano-Flores, L. P., de Oliveira, C. V. (2003). Identification of neuronscontaining orexin-B (hypocretin-2) immunoreactivity in limbic structures. Brain Res. 967(1/2):123–131.
Coniglio, S. J., Lewis, J. D., Lang, C., Burns, T. G., Subhani-Siddique, R., Weintraub, A., Schub, H., andHolden, E. W. (2001). A randomized, double-blind, placebo-controlled trial of single-dose intra-venous secretin as treatment for children with autism. J. Pediatr. 138(5):649–655.
Corona, R., Dissanayake, C., Arbelle, S., Wellington, P., and Sigman, M. (1998). Is affect aversive toyoung children with autism? Behavioral and cardiac responses to experimenter distress. Child Dev.69(6):1494–1502.
Davis, E., Fennoy, I., Laraque, D., Kanem, N., Brown, G., and Mitchell, J. (1992). Autism and developmentalabnormalities in children with perinatal cocaine exposure. J. Natl. Med. Assoc. 84(4):315–319.
de Lecea, L., Kilduff, T. S., Peyron, C., Gao, X., Foye, P. E., Danielson, P. E., Fukuhara, C., Battenberg, E. L.,Gautvik, V. T., Bartlett, F. S., II, Frankel, W. N., van den Pol, A. N., Bloom, F. E., Gautvik, K. M., and

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
Secretin: Hypothalamic Distribution and Hypothesized Neuroregulatory Role in Autism 237
Sutcliffe, J. G. (1998). The hypocretins: Hypothalamus-specific peptides with neuroexcitatory activity.Proc. Natl. Acad. Sci. U.S.A. 95(1):322–327.
D’Eufemia, P., Celli, M., Finocchiaro, R., Pacifico, L., Viozzi, L., Zaccagnini, M., Cardi, E., and Giardini,O. (1996). Abnormal intestinal permeability in children with autism. Acta Paediatr. 85(9):1076–1079.
Diggle, T., McConachie, H. R., and Randle, V. R. (2003). Parent-mediated early intervention for youngchildren with autism spectrum disorder. Cochrane Database Syst. Rev. (1):CD003496.
Dunn-Geier, J., Ho, H. H., Auersperg, E., Doyle, D., Eaves, L., Matsuba, C., Orrbine, E., Pham, B., andWhiting, S. (2000). Effect of secretin on children with autism: A randomized controlled trial. Dev.Med. Child Neurol. 42(12):796–802.
Eriksson, M., Bjorkstrand, E., Smedh, U., Alster, P., Matthiesen, A. S., and Uvnas-Moberg, K. (1994).Role of vagal nerve activity during suckling. Effects on plasma levels of oxytocin, prolactin, VIP,somatostatin, insulin, glucagon, glucose and of milk secretion in lactating rats. Acta Physiol. Scand.151(4):453–459.
Francis, D. D., Young, L. J., Meaney, M. J., and Insel, T. R. (2002). Naturally occurring differences inmaternal care are associated with the expression of oxytocin and vasopressin (V1a) receptors: Genderdifferences. J. Neuroendocrinol. 14(5):349–353.
Freeman, J. H., Jr., Weible, A., Rossi, J., and Gabriel, M. (1997). Lesions of the entorhinal cortex disruptbehavioral and neuronal responses to context change during extinction of discriminative avoidancebehavior. Exp. Brain Res. 115(3):445–457.
Fremeau, R. T., Jr., Korman, L. Y., and Moody, T. W. (1986). Secretin stimulates cyclic AMP formation inthe rat brain. J. Neurochem. 46(6):1947–1955.
Friedman, G. B., Taylor, C. T., Parkos, C. A., and Colgan, S. P. (1998). Epithelial permeability inducedby neutrophil transmigration is potentiated by hypoxia: Role of intracellular cAMP. J. Cell Physiol.176(1):76–84.
Fuxe, K., Andersson, K., Hokfelt, T., Mutt, V., Ferland, L., Agnati, L. F., Ganten, D., Said, S., Eneroth,P., and Gustafsson, J. A. (1979). Localization and possible function of peptidergic neurons and theirinteractions with central catecholamine neurons, and the central actions of gut hormones. Fed. Proc.38(9):2333–2340.
Gabry, K. E., Chrousos, G. P., Rice, K. C., Mostafa, R. M., Sternberg, E., Negrao, A. B., Webster, E. L.,McCann, S. M., and Gold, P. W. (2002). Marked suppression of gastric ulcerogenesis and intestinalresponses to stress by a novel class of drugs. Mol. Psychiatry 7(5):474–483.
Gandhi, S., Tsueshita, T., Onyuksel, H., Chandiwala, R., and Rubinstein, I. (2002). Interactions of humansecretin with sterically stabilized phospholipid micelles amplify peptide-induced vasodilation in vivo.Peptides 23(8):1433–1439.
Gauthier, A., Puente, J. L., and Finlay, B. B. (2003). Secretin of the enteropathogenic Escherichia colitype III secretion system requires components of the type III apparatus for assembly and localization.Infect Immun. 71(6):3310–3319.
Gershon, M. D. (1998). The Second Brain: A Groundbreaking New Understanding of Nervous Disordersof the Stomach and Intestine, Harper Collins, New York.
Golanov, E. V., Ruggiero, D. A., and Reis, D. J. (2000). A brainstem area mediating cerebrovascular andEEG responses to hypoxic excitation of rostral ventrolateral medulla in rat. J. Physiol. 529(Pt. 2):413–429.
Goulet, M., Shiromani, P. J., Ware, C. M., Strong, R. A., Boismenu, R., Rusche, J. R. (2003). A secretini.v. infusion activates gene expression in the central amygdala of rats. Neuroscience 118(4):881–888.
Graveling, R. A., and Brooke, J. D. (1978). Hormonal and cardiac response of autistic children to changesin environmental stimulation. J. Autism Child Schizophr. 8(4):441–455.
Guilloteau, P., Chayvialle, J. A., Toullec, R., Grongnet, J. F., and Bernard, C. (1992). Early-life patternsof plasma gut regulatory peptide levels in calves: Effects of the first meals. Biol. Neonate. 61(2):103–109.
Harmar, A. J. (2001). Family-B G-protein-coupled receptors. Genome Biol. 2(12): Reviews 3013.Haznedar, M. M., Buchsbaum, M. S., Wei, T. C., Hof, P. R., Cartwright, C., Bienstock, C. A., and Hollander,
E. (2000). Limbic circuitry in patients with autism spectrum disorders studied with positron emissiontomography and magnetic resonance imaging. Am. J. Psychiatry 157(12):1994–2001.
Helou, C. M., Imbert-Teboul, M., Doucet, A., Rajerison, R., Chollet, C., Alhenc-Gelas, F., and Marchetti,J. (2003). Angiotensin receptor subtypes in thin and muscular juxtamedullary efferent arterioles ofrat kidney. Am. J. Physiol. Renal. Physiol. 285(3):F507–F514.
Hollander, E., King, A., Delaney, K., Smith, C. J., and Silverman, J. M. (2003a). Obsessive–compulsivebehaviors in parents of multiplex autism families. Psychiatry Res. 117(1):11–16.
Hollander, E., Novotny, S., Hanratty, M., Yaffe, R., DeCaria, C. M., Aronowitz, B. R., and Mosovich,S. (2003b). Oxytocin infusion reduces repetitive behaviors in adults with autistic and Asperger’sdisorders. Neuropsychopharmacology 28(1):193–198.

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
238 Welch, Keune, Welch-Horan, Anwar, Anwar, Ludwig, and Ruggiero
Horvath, K., Papadimitriou, J. C., Rabsztyn, A., Drachenberg, C., and Tildon, J. T. (1999). Gastrointestinalabnormalities in children with autistic disorder. J. Pediatr. 135(5):559–563.
Horvath, K., and Perman, J. A. (2002). Autism and gastrointestinal symptoms. Curr. Gastroenterol Rep.4(3):251–258.
Horvath, K., Stefanatos, G., Sokolski, K. N., Wachtel, R., Nabors, L., and Tildon, J. T. (1998). Improvedsocial and language skills after secretin administration in patients with autistic spectrum disorders.J. Assoc. Acad. Minor. Phys. 9(1):9–15.
Houben, H., and Denef, C. (1994). Bioactive peptides in anterior pituitary cells. Peptides 15(3):547–582.Ichihara, K., Eng, J., and Yalow, R. S. (1983). Ontogeny of immunoreactive CCK, VIP and secretin in rat
brain and gut. Biochem. Biophys. Res. Commun. 112(3):891–898.Iqbal Z. (2002). Ethical issues involved in the implementation of a differential reinforcement of inap-
propriate behaviour programme for the treatment of social isolation and ritualistic behaviour in anindividual with intellectual disabilities. J. Intellect. Disabil. Res. 46(Pt. 1):82–93.
Itoh, N., Furuya, T., Ozaki, K., Ohta, M., and Kawasaki, T. (1991). The secretin precursor gene. Structureof the coding region and expression in the brain. J. Biol. Chem. 266(19):12595–12598.
Jezova, M., Armando, I., Bregonzio, C., Yu, Z. X., Qian, S., Ferrans, V. J., Imboden, H., and Saavedra, J. M.(2003). Angiotensin II AT(1) and AT(2) receptors contribute to maintain basal adrenomedullarynorepinephrine synthesis and tyrosine hydroxylase transcription. Endocrinology 144(5):2092–tt22101.
Jin, H. O., Lee, K. Y., Chang, T. M., Chey, W. Y., and Dubois, A. (1994). Secretin: A physiological regulatorof gastric emptying and acid output in dogs. Am. J. Physiol. 267(4, Pt. 1):702–708.
Jones, D., and Gonzalez-Lima, F. (2001). Mapping Pavlovian conditioning effects on the brain: Blocking,contiguity, and excitatory effects. J. Neurophysiol. 86(2):809–823.
Jones, G. R., Singer, P. P., and Bannach, B. (2002). Application of LC-MS analysis to a colchicine fatality.J. Anal. Toxicol. 26(6):365–369.
Kern, J. K., Van Miller, S., Evans, P. A., and Trivedi, M. H. (2002). Efficacy of porcine secretin in childrenwith autism and pervasive developmental disorder. J. Autism Dev. Disord. 32(3):153–160.
Kobayashi, R., Takenoshita, Y., Kobayashi, H., Kamijo, A., Funaba, K., and Takarabe, M. (2001). Earlyintervention for infants with autistic spectrum disorders in Japan. Pediatr. Int. 43(2):202–208.
Kopin, A. S., Wheeler, M. B., and Leiter, A. B. (1990). Secretin: Structure of the precursor and tissuedistribution of the mRNA. Proc. Natl. Acad. Sci. U.S.A. 87(6):2299–2303.
Koren, G. (2001). Repeated doses of porcine secretin in the treatment of autism: A randomized, placebo-controlled trial. Pediatrics 107(5):E71.
Koves, K., Kausz, M., Reser, D., and Horvath, K. (2002). What may be the anatomical basis that secretincan improve the mental functions in autism? Regul. Pept. 109(1–3):167–172.
Lamson, D. W., and Plaza, S. M. (2001). Transdermal secretin for autism—A case report. Altern. Med.Rev. 6(3):311–313.
Langworthy-Lam, K. S., Aman, M. G., and Van Bourgondien, M. E. (2002). Prevalence and patterns ofuse of psychoactive medicines in individuals with autism in the Autism Society of North Carolina. J.Child Adolesc. Psychopharmacol. 12(4):311–321.
Lauterbach, H. H., Lehmann, E., Escobar-Jimenez, F., Mattes, P., and Raptis, S. (1980). Behavior ofsecretin in the hypoxia-immobilisation stress on the rat. Eur. Surg. Res. 12(2):103–107.
Leong, D. S., Terron, J. A., Falcon-Neri, A., Armando, I., Ito, T., Johren, O., Tonelli, L. H., Hoe, K. L.,and Saavedra, J. M. (2002). Restraint stress modulates brain, pituitary and adrenal expression ofangiotensin II AT(1A), AT(1B) and AT(2) receptors. Neuroendocrinology 75(4):227–240.
Li, P., Chang, T. M., and Chey, W. Y. (1998). Secretin inhibits gastric acid secretion via a vagal afferentpathway in rats. Am. J. Physiol. 275(1, Pt. 1):G22–G28.
Li, Y. W., and Guyenet, P. G. (1996). Angiotensin II decreases a resting K+ conductance in rat bulbospinalneurons of the C1 area. Circ. Res. 78(2):274–282.
Lightdale, J. R., Hayer, C., Duer, A., Lind-White, C., Jenkins, S., Siegel, B., Elliott, G. R., andHeyman, M. B. (2001). Effects of intravenous secretin on language and behavior of childrenwith autism and gastrointestinal symptoms: A single-blinded, open-label pilot study. Pediatrics108(5):90.
Loewy, A. D. (1991). Forebrain nuclei involved in autonomic control. Prog. Brain Res. 87:253–268.Lopez, M. J., Upchurch, B. H., Rindi, G., and Leiter, A. B. (1995). Studies in transgenic mice reveal
potential relationships between secretin-producing cells and other endocrine cell types. J. Biol. Chem.270(2):885–891.
Lucas, A., Adrian, T. E., Bloom, S. R., Aynsley-Green, A. (1980). Plasma secretin in neonates. ActaPaediatr. Scand. 69(2): 205–210.
Lucas, A., Bloom, S. R., and Aynsley-Green, A. (1983). Metabolic and endocrine consequences of depriv-ing preterm infants of enteral nutrition. Acta Paediatr. Scand. 72(2):245–249.

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
Secretin: Hypothalamic Distribution and Hypothesized Neuroregulatory Role in Autism 239
Matthiesen, A. S., Ransjo-Arvidson, A. B., Nissen, E., and Uvnas-Moberg, K. (2001). Postpartum maternaloxytocin release by newborns: Effects of infant hand massage and sucking. Birth 28(1):13–19.
McEwen, B. S. (2001). Plasticity of the hippocampus: Adaptation to chronic stress and allostatic load.Ann. N.Y. Acad. Sci. 933:265–277.
McKay, T., Reynolds, P., Jezzard, S., Curiel, D., and Coutelle, C. (2002). Secretin-mediated gene delivery,a specific targeting mechanism with potential for treatment of biliary and pancreatic disease in cysticfibrosis. Mol. Ther. 5(4):447–454.
Meaney, M. J., Aitken, D. H., Bhatnagar, S., and Sapolsky, R. M. (1991). Postnatal handling attenuatescertain neuroendocrine, anatomical, and cognitive dysfunctions associated with aging in female rats.Neurobiol. Aging. 12(1):31–38.
Meaney, M. J., Aitken, D. H., van Berkel, C., Bhatnagar, S., and Sapolsky, R. M. (1988). Effect of neonatalhandling on age-related impairments associated with the hippocampus. Science 239(4841, Pt. 1):766–768.
Militerni, R., Bravaccio, C., Falco, C., Fico, C., and Palermo, M. T. (2002). Repetitive behaviors in autisticdisorder. Eur. Child Adolesc. Psychiatry 11(5):210–218.
Miller, T. A., Llanos, O. L., Swierczek, J. S., Rayford, P. L., and Thompson, J. C. (1978). Concentrations ofgastrin and secretin in the alimentary tract of the cat. Surgery 83(1):90–93.
Mineo, H., Oyamada, T., and Kato, S. (1990). Effect of feeding on plasma secretin concentrations in sheep.Res. Vet. Sci. 49(2):157–179.
Mutt, V., Carlquist, M., and Tatemoto, K. (1979). Secretin-like bioactivity in extracts of porcine brain. LifeSci. 25(20):1703–1707.
Nelson, K. B., Grether, J. K., Croen, L. A., Dambrosia, J. M., Dickens, B. F., Jelliffe, L. L., Hansen, R. L.,and Phillips, T. M. (2001). Neuropeptides and neurotrophins in neonatal blood of children with autismor mental retardation. Ann. Neurol. 49(5):597–606.
Ng, S. S., Yung, W. H., and Chow, B. K. (2002). Secretin as a neuropeptide. Mol. Neurobiol. 26(1):97–107.Nussdorfer, G. G., Bahcelioglu, M., Neri, G., and Malendowicz, L. K. (2000). Secretin, glucagon, gas-
tric inhibitory polypeptide, parathyroid hormone, and related peptides in the regulation of thehypothalamus–pituitary–adrenal axis. Peptides 21(2):309–324.
O’Donohue, T. L., Charlton, C. G., Miller, R. L., Boden, G., and Jacobowitz, D. M. (1981). Identification,characterization, and distribution of secretin immunoreactivity in rat and pig brain. Proc. Natl. Acad.Sci. U.S.A. 78(8):5221–5224.
Oektedalen, O., Opstad, P. K., Schaffalitzky de Muckadell, O. B. (1982). Secretin—A new stress hormone?Regul. Pept. 4(4):213–219.
Ogai, M., Matsumoto, H., Suzuki, K., Ozawa, F., Fukuda, R., Uchiyama, I., Suckling, J., Isoda, H., Mori,N., and Takei, N. (2003). fMRI study of recognition of facial expressions in high-functioning autisticpatients. Neuroreport 14(4):559–563.
Ohta, M., Funakoshi, S., Kawasaki, T., and Itoh, N. (1992). Tissue-specific expression of the rat secretinprecursor gene. Biochem. Biophys. Res. Commun. 183(2):390–395.
Owley, T., McMahon, W., Cook, E. H., Laulhere, T., South, M., Mays, L. Z., Shernoff, E. S., Lainhart, J.,Modahl, C. B., Corsello, C., Ozonoff, S., Risi, S., Lord, C., Leventhal, B. L., and Filipek, P. A. (2001).Multisite, double-blind, placebo-controlled trial of porcine secretin in autism. J. Am. Acad. ChildAdolesc. Psychiatry 40(11):1293–1299.
Paquette, T. L., Shulman, D. F., Alpers, D. H., and Jaffe, B. M. (1982). Postnatal development of intestinalsecretin in rats and guinea pigs. Am. J. Physiol. 243(6):G511–G5117.
Park, Y. D. (2003). The effects of vagus nerve stimulation therapy on patients with intractable seizuresand either Landau–Kleffner syndrome or autism. Epilepsy Behav. 4(3):286–290.
Paxinos, G., and Watson, C. (1998). The Rat Brain in Stereotaxic Coordinates, Academic Press, New York.Porges, S. W. (1995). Orienting in a defensive world: Mammalian modifications of our evolutionary her-
itage. A Polyvagal Theory. Psychophysiology 32(4):301–318.Posey, D. J., and McDougle, C. J. (2000). The pharmacotherapy of target symptoms associated with autistic
disorder and other pervasive developmental disorders. Harv. Rev. Psychiatry 8(2):45–63.Richer, J. M., and Coss, R. G. (1976). Gaze aversion in autistic and normal children. Acta Psychiatr. Scand.
53(3):193–210.Rinaman, L., Levitt, P., and Card, J. P. (2000). Progressive postnatal assembly of limbic–autonomic cir-
cuits revealed by central transneuronal transport of pseudorabies virus. J. Neurosci. 20(7):2731–2741.
Roberts, W., Weaver, L., Brian, J., Bryson, S., Emelianova, S., Griffiths, A. M., MacKinnon, B., Yim, C.,Wolpin, J., and Robinson, T. W. (2001). Homeopathic secretin in autism: A clinical pilot study. Br.Homeopath. J. 90(2):86–91.
Rogers, I. M., Davidson, D. C., Lawrence, J., and Buchanan, K. D. (1975). Neonatal secretion of secretin.Arch. Dis. Child 50(2):120–122.

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
240 Welch, Keune, Welch-Horan, Anwar, Anwar, Ludwig, and Ruggiero
Ruggiero, D. A., Ross, C. A., Anwar, M., Park, D. H., Joh, T. H., and Reis, D. J. (1985). Distribution ofneurons containing phenylethanolamine N-methyltransferase in medulla and hypothalamus of rat.J. Comp. Neurol. 239(2):127–154.
Ruggiero, D. A., Welch-Horan, T. B., Keune, J. D., Anwar, N., Anwar, M., Ludwig, R. J., and Welch, M. G.(2003). Secretin: Distribution, Specificity Support Neuroregulatory Role in Autism. In 33rd AnnualMeeting, Neurosci Abstracts.
Rumsey, J. M., and Ernst, M. (2000). Functional neuroimaging of autistic disorders. Ment. Retard. Dev.Disabil. Res. Rev. 6(3):171–179.
Samson, W. K., Lumpkin, M. D., and McCann, S. M. (1984). Presence and possible site of action of secretinin the rat pituitary and hypothalamus. Life Sci. 34(2):155–163.
Sandler, A. D., Sutton, K. A., DeWeese, J., Girardi, M. A., Sheppard, V., and Bodfish, J. W. (1999). Lackof benefit of a single dose of synthetic human secretin in the treatment of autism and pervasivedevelopmental disorder. N. Engl. J. Med. 341(24):1801–1806.
Schultz, R. T., Grelotti, D. J., Klin, A., Kleinman, J., Van der Gaag, C., Marois, R., and Skudlarski, P.(2003). The role of the fusiform face area in social cognition: Implications for the pathobiology ofautism. Philos. Trans. R. Soc. Lond. B Biol. Sci. 358(1430):415–427.
Steptoe, A., Cropley, M., and Joekes, K. (1999). Job strain, blood pressure and response to uncontrollablestress. J. Hypertens. 17(2):193–200.
Strauss, E., and Yalow, R. S. (1978). Immunoreactive secretin in gastrointestinal mucosa of several mam-malian species. Gastroenterology 75(3):401–404.
Sved, A. F., Mancini, D. L., Graham, J. C., Schreihofer, A. M., and Hoffman, G. E. (1994). PNMT-containingneurons of the C1 cell group express c-fos in response to changes in baroreceptor input. Am J Physiol.266(2, Pt. 2):R361-R367.
Swanson, L. W. (1998). Brain Maps: Structure of the Rat Brain, 2nd Revision, Elsevier Science B. V.,Amsterdam, the Netherlands.
Talman, W. T., and Kelkar P. (1993). Neural control of the heart. Central and peripheral. Neurol. Clin.11(2):239–256.
Tanoue, Y., and Oda, S. (1989). Weaning time of children with infantile autism. J. Autism Dev. Disord.19(3):425–434.
Teufel, M., Luik, G., and Niessen, K. H. (1986). [Gastrin, secretin, VIP and motilin in children withmucoviscidosis and Crohn disease]. Monatsschr. Kinderheilkd. 134(3):132–137.
Tinbergen, N., and Tinbergen, E. A. (1983). Autistic Children—New Hope for a Cure, George, Allen andUnwin, London.
Torrente, F., Ashwood, P., Day, R., Machado, N., Furlano, R. I., Anthony, A., Davies, S. E., Wakefield,A. J., Thomson, M. A., Walker-Smith, J. A., and Murch, S. H. (2002). Small intestinal enteropathywith epithelial IgG and complement deposition in children with regressive autism. Mol. Psychiatry7(4):375–382.
Uno, H., Tarara, R., Else, J. G., Suleman, M. A., and Sapolsky, R. M. (1989). Hippocam-pal damage associated with prolonged and fatal stress in primates. J. Neurosci. 9(5):1705–1711.
Vacher, C. M., Fretier, P., Creminon, C., Calas, A., and Hardin-Pouzet, H. (2002). Activation by sero-tonin and noradrenaline of vasopressin and oxytocin expression in the mouse paraventricular andsupraoptic nuclei. J Neurosci. 22(5):1513–1522.
Veerendra-Kumar, M. H., and Gupta, Y. K. (2002). Intracerebroventricular administration of colchicineproduces cognitive impairment associated with oxidative stress in rats. Pharmacol. Biochem. Behav.73(3):565–571.
Walker, J. K., Premont, R. T., Barak, L. S., Caron, M. G., and Shetzline, M. A. (1999). Properties ofsecretin receptor internalization differ from those of the beta(2)-adrenergic receptor. J. Biol. Chem.274(44):31515–31523.
Warren, R. P., Odell, J. D., Warren, W. L., Burger, R. A., Maciulis, A., Daniels, W. W., and Torres, A. R.(1997). Brief Report: Immunoglobulin A deficiency in a subset of autistic subjects. J. Autism Dev.Disord. 27(2):187–192.
Wasserman, A. M., Ferreira, M., Jr., Sahibzada, N., Hernandez, Y. M., and Gillis, R. A. (2002). GABA-mediated neurotransmission in the ventrolateral NTS plays a role in respiratory regulation in the rat.Am. J. Physiol. Regul. Integr. Comp .Physiol. 283(6):R1423–R1441.
Welch, M. G. (1983a). Retrieval from autism through mother–child holding. In Tinbergen, N., andTinbergen, E. A., Autistic Children—New Hope for a Cure, George, Allen and Unwin, London.
Welch, M. G. (1983b). Retrieval from autism through mother–child holding therapy. In Call, J. D.,Galenson, E., and Tyson, R. L. (eds.), Frontiers of Infant Psychiatry, Basic Books, New York, 450.
Welch, M. G. (1987). Toward prevention of developmental disorders. Pa Med. 90(3):47–52.Welch, M. G. (1988). Holding Time, Simon and Schuster, New York.

P1: JWF
Cellular and Molecular Neurobiology [cemn] pp1089-cemn-478086 March 2, 2004 12:21 Style file version Oct 23, 2000
Secretin: Hypothalamic Distribution and Hypothesized Neuroregulatory Role in Autism 241
Welch, M. G. (1989). Holding Time: How When Why. In Proceedings of the First International Congressof Holding Therapy, Regensberg, Germany.
Welch, M. G., and Chaput P. (1988). Mother–child holding therapy and autism. Pa Med. 91(10):33–38.Welch, M. G., Keune, J. D., Welch-Horan, T. B., Anwar, M., Anwar, N., and Ruggiero, D. A. (2003a).
Secretin activates visceral brain regions in rat including areas abnormal in autism. Cell Mol Neurobiol.23(4/5):817–837.
Welch, M. G., Welch-Horan, T. B., Keune, J. D., Anwar, N., Anwar, M., Ludwig, R. J., and Ruggiero, D. A.(2003b). Neurohormonal resolution of genetic and acquired IBD and secondary brain activation inareas abnormal in autism. In 33rd Annual Meeting, Neuroscience Abstracts.
Welch, M. G., Keune, J. D., Welch-Horan, T. B., and Ruggiero, D. A. (2002a). Secretin in autism. Soci.Neurosci. Press Book, 32nd Annu. Meet. II:572–574.
Welch, M. G., Keune, J. D., Welch-Horan, T. B., and Ruggiero, D. A. (2002b). Secretin activates visceralbrain regions including areas abnormal in autism. In 32nd Annual Meeting, Neuroscience Abstracts.
Wheeler, G. (2003). RG-1068 RepliGen. Curr. Opin. Investig. Drugs 4(1):66–71.White, J. F. (2003). Intestinal pathophysiology in autism. Exp. Biol. Med. (Maywood) 228(6):639–649.Yung, W. H., Leung, P. S., Ng, S. S., Zhang, J., Chan, S. C., and Chow, B. K. (2001). Secretin facilitates
GABA transmission in the cerebellum. J. Neurosci. 21(18):7063–7068.




















