
Molecular Mechanisms of Signal Integration in Hypothalamic ...
11
AMER. ZOOL., 33:244-254 (1993) Molecular Mechanisms of Signal Integration in Hypothalamic Neurons 1 R. THOMAS ZOELLER 2 AND NISREEN KABEER Department of Anatomy and Neurobiology, University of Missouri, School of Medicine, Columbia, Missouri 65212 AND H. ELLIOTT ALBERS Laboratory of Neuroendocrinology and Behavior, Departments of Biology and Psychology, Georgia State University, Atlanta, Georgia 30303 SYNOPSIS. The purpose of this paper is to describe our studies focused on the mechanisms by which hypothalamic neurons process multiple signals and produce an integrated response. We illustrate our research strategy by reviewing our work on two separate neural systems: the hypo- thalamic paraventricular nucleus (PVN) and the suprachiasmatic nucleus (SCN). We have focused on different peptidergic subpopulations within these nuclei to address two issues. In the PVN, we concentrate on the population of neurons containing thyrotropin-releasing hormone (TRH). These neurons are inhibited by thyroid hormones, but activated by cold exposure. Using a molecular approach, we have demonstrated that these conflicting signals simultaneously act on the same population of TRH neurons. This system will continue to be a productive model to study the mechanisms by which neurons process multiple signals. In the SCN, we concentrate on the population of neurons containing vasoactive intestinal peptide (VIP), peptide histidine isoleucine (PHI) and gastrin releasing peptide (GRP). We have demonstrated that injection of all three peptides into the SCN of hamsters mimics the phase-delaying effects of light on circadian wheel running behavior. In addition, the genes encoding these peptides exhibit different 24-hour profiles of changes in neurons of the SCN. These data support the hypothesis that one mechanism by which these neurons produce an integrated response is by changing the concen- tration ratio of co-released peptides. INTRODUCTION focused on the mechanisms by which hypo- The brain coordinates behavioral, endo- thalamic neurons process information they crine and autonomic functions by process- receive from both internal and external ing a variety of internal and external stimuli, sources, and how they may produce an inte- Stimuli from both internal and external grated response. Considering that external sources can converge on a single neuron stimuli reach the brain through sensory sys- which must process these signals and pro- terns that synapse on specific neurons, and duce an integrated response. Our work is that internal stimuli can communicate with the brain in the form of hormones, we have identified a model system that will allow us 1 From the Symposium Neural Aspects of Repw- to examine how a single neuron can co-pro- ductive Endocrinology: Old Questions, New Approaches cess trans-synaptic and endocrine signals to presented at the Annual Meeting of the American Sod- regulate neuroendocrine function. In addi- ety of Zoologists, 27-30 December 1990 at San Anto- ^ tQ determine how hypothalamic neu- nio, 1 cx3.s. * • * = Present address: Department of Biology, Univer- rons ma y produce an integrated response to sity of Massachusetts at Amherst, Morrill Science Cen- multiple Signals, we have identified a group ter, Amherst, MA 01003. of neurons that contains three different pep- 244 Downloaded from https://academic.oup.com/icb/article/33/2/244/154040 by guest on 15 September 2022
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Molecular Mechanisms of Signal Integration in Hypothalamic ...

AMER. ZOOL., 33:244-254 (1993)
Molecular Mechanisms of Signal Integration inHypothalamic Neurons1
R. THOMAS ZOELLER2 AND NISREEN KABEER
Department of Anatomy and Neurobiology, University of Missouri,School of Medicine, Columbia, Missouri 65212
AND
H. ELLIOTT ALBERS
Laboratory of Neuroendocrinology and Behavior, Departments of Biology and Psychology,Georgia State University, Atlanta, Georgia 30303
SYNOPSIS. The purpose of this paper is to describe our studies focusedon the mechanisms by which hypothalamic neurons process multiplesignals and produce an integrated response. We illustrate our researchstrategy by reviewing our work on two separate neural systems: the hypo-thalamic paraventricular nucleus (PVN) and the suprachiasmatic nucleus(SCN). We have focused on different peptidergic subpopulations withinthese nuclei to address two issues. In the PVN, we concentrate on thepopulation of neurons containing thyrotropin-releasing hormone (TRH).These neurons are inhibited by thyroid hormones, but activated by coldexposure. Using a molecular approach, we have demonstrated that theseconflicting signals simultaneously act on the same population of TRHneurons. This system will continue to be a productive model to study themechanisms by which neurons process multiple signals. In the SCN, weconcentrate on the population of neurons containing vasoactive intestinalpeptide (VIP), peptide histidine isoleucine (PHI) and gastrin releasingpeptide (GRP). We have demonstrated that injection of all three peptidesinto the SCN of hamsters mimics the phase-delaying effects of light oncircadian wheel running behavior. In addition, the genes encoding thesepeptides exhibit different 24-hour profiles of changes in neurons of theSCN. These data support the hypothesis that one mechanism by whichthese neurons produce an integrated response is by changing the concen-tration ratio of co-released peptides.
INTRODUCTION focused on the mechanisms by which hypo-The brain coordinates behavioral, endo- thalamic neurons process information they
crine and autonomic functions by process- receive from both internal and externaling a variety of internal and external stimuli, sources, and how they may produce an inte-Stimuli from both internal and external grated response. Considering that externalsources can converge on a single neuron stimuli reach the brain through sensory sys-which must process these signals and pro- terns that synapse on specific neurons, andduce an integrated response. Our work is that internal stimuli can communicate with
the brain in the form of hormones, we haveidentified a model system that will allow us
1 From the Symposium Neural Aspects of Repw- to examine how a single neuron can co-pro-ductive Endocrinology: Old Questions, New Approaches cess trans-synaptic and endocrine signals topresented at the Annual Meeting of the American Sod- regulate neuroendocrine function. In addi-ety of Zoologists, 27-30 December 1990 at San Anto- ^ t Q d e t e r m i n e h o w hypothalamic neu-nio, 1 cx3.s. * • *
= Present address: Department of Biology, Univer- r o n s m a y produce an integrated response tosity of Massachusetts at Amherst, Morrill Science Cen- multiple Signals, we have identified a groupter, Amherst, MA 01003. of neurons that contains three different pep-
244
Dow
nloaded from https://academ
ic.oup.com/icb/article/33/2/244/154040 by guest on 15 Septem
ber 2022

HYPOTHALAMIC INTEGRATION 245
tides; we are investigating the functionalconsequences of these co-localized peptidesand whether the biosynthesis of these pep-tides by external and internal stimuli mightrepresent a mechanism by which these neu-rons produce an integrated response.
The purpose of this paper is to illustrateour research strategy by describing these twoseparate projects. These projects are focusedon well-defined subpopulations of neuronsin two hypothalamic nuclei of rats knownfor their role in integrating multiple signalsand in coordinating behavioral and physi-ological processes: the paraventricularnucleus (PVN) and the suprachiasmaticnucleus (SCN). We particularly emphasizethe necessity of validating certain charac-teristics of a neural system to be the focusof studies investigating mechanisms of sig-nal processing, and the experimental utilityof looking first at mechanisms regulatingcellular levels of neuroendocrine peptidemRNAs.
IDENTIFICATION OF A MODEL SYSTEM TOEXAMINE MOLECULAR MECHANISMS BY
WHICH HYPOTHALAMIC NEURONSPROCESS MULTIPLE SIGNALS
We have been guided by four criteria toidentify a "model" neuroendocrine systemthat would be accessible to the kinds ofexperiments we felt were required to under-stand the molecular and cellular mecha-nisms by which neurons can integrate mul-tiple signals. First, the neuronal systemshould be spatially restricted; that is, theneurons under study should be anatomi-cally clustered so that they can be "har-vested" from adult tissue for biochemicaland molecular analyses, as well as for cul-turing. Second, there should be a back-ground of information describing the phys-iological systems regulating this group ofneurons, and these systems should be easilymanipulated under laboratory conditions.Third, all neurons within this "cluster" mustbe regulated by the same set of physiologicalstimuli, so that mechanisms of integrationcan be reasonably interpreted as occurringwithin the same neuron. Finally, the neu-rons should be viable in vitro to allow accessto experiments focused on signal transduc-tion mechanisms. In this section, we describe
how these criteria have guided our approachto defining a neuroendocrine system thatwill be accessible to experimental manipu-lation.
Information on TRH neurons of the PVNTRH was the first releasing factor chem-
ically defined (Folkers et al, 1969; Burguset al, 1970; Nair et al, 1970); thus, thereis a wealth of information about its role asa regulator of thyrotropin synthesis andsecretion (Morley, 1981; Koenig et al, 1984;Taylor et al, 1986, 1990; Shupnik et al,1986), as well as the location of TRH-con-taining neurons within the brain. AlthoughTRH is found widespread throughout thebrain (Jackson and Lechan, 1985; Liao etal, 1985; Lechan et al, 1986), TRH in themedian eminence is produced exclusivelyby a cluster of parvocellular neurons in thePVN (Brownstein et al, 1975, 1982; Pal-kovits et al, 1982; Ishikawa et al, 1988).Thus, TRH neurons of the PVN met thefirst criterion: They are restricted in theirdistribution, and they appear to represent ahomogeneous population regulating thepituitary-thyroid axis. In addition, an abun-dance of work has demonstrated that TRHneurons of the PVN integrate multiple sig-nals; we have focused on two—thyroid hor-mone (T3) and cold exposure.
External and internal stimuliinteract to regulate TRHneurons of the PVN
Several recent studies have demonstratedthat T3 exerts a negative feedback effect onTRH neurons of the PVN. T3 reduces TRHconcentration in pituitary-portal blood(Rondeel et al, 1988), TRH content of themedian eminence and paraventricularnucleus (Mori and Yamada, 1987; Yamadaet al, 1989), and ultrastructural character-istics of neuronal activity in TRH-contain-ing neurons of the PVN (Hisano et al, 1986).In contrast, acute exposure to cold producesa rapid (within 15 min) increase in TRHrelease from median eminence (Rondeel etal, 1988; Arancibia et al, 1989) as well asin plasma TSH and T3 (Jobin et al, 1975,1976; Hefco et al, 1975; Mannisto, 1983).This effect of cold is blocked by PVN lesions
Dow
nloaded from https://academ
ic.oup.com/icb/article/33/2/244/154040 by guest on 15 Septem
ber 2022

246 R. T. ZOELLER ET AL.
(Ishikawa et al, 1984), passive immuniza-tion with TRH antisera (Szabo and Froh-man, 1977; Szabo et al, 1978; Mori et al,1978; Prasad et al, 1980), and local (to PVNor ME) or systemic administration of a-ad-renergic antagonists or inhibitors of cate-cholamine synthesis (Onaya and Hashi-zume, 1977; Krulich et al, 1977; Schettiniet al, 1979; Terry, 1986; Arancibia et al,1989). Considering that TRH neurons ofthe PVN receive adrenergic synapses(Liposits et al, 1987), these findings providestrong evidence that cold exposure producesa neural reflex that stimulates TRH neuronsof the PVN through adrenergic afferentsdespite elevated levels of T3. The detail ofthis background information and the rela-tive ease with which ambient temperatureand circulating levels of T3 can be regulatedestablished our third criterion for a "model"system.
Neural and endocrine factors interactwithin the same TRH neurons
In principle, cold exposure must eitheractivate a subset of TRH neurons in thePVN that are not suppressed by T3 or atten-uate the inhibitory effect of T3 within eachTRH neuron. Our approach to discrimi-nating between these possibilities was basedon the observation that many genes encod-ing neurotransmitter enzymes and peptidesare regulated in parallel with neuronal activ-ity. There is a rapidly growing literaturedemonstrating that cellular levels of neu-ropeptide mRNAs are regulated by trans-membrane mechanisms initiated by thesame stimulus {e.g., synaptic activity) thatdepolarizes the membrane and leads totransmitter release (see reviews by YoungandZoeller, 1987; Comb etal, 1987; Good-man, 1990; Habener, 1990; Uhl and Nishi-mori, 1990; Van Nguyen et al, 1990). Thiscoupling between transmitter secretion andcellular levels of mRNAs provides a molec-ular correlate of neuronal activity as well asa pivotal justification for our approach. Wereasoned that if changes in TRH mRNAlevels mirror changes in TRH neuronalactivity, then T3 will reduce TRH mRNAlevels in neurons of the PVN while coldexposure will elevate TRH mRNA levels.Further, if these two signals are regulating
the activity of the same population of neu-rons, then TRH mRNA levels should beelevated by cold exposure in the same neu-rons in which T3 reduces TRH mRNA lev-els.
As predicted, TRH mRNA levels are reg-ulated by T3: cellular levels of TRH mRNAin the PVN are elevated in hypothyroid rats,and daily injections of T3 reduce TRHmRNA levels in the PVN of both hypothy-roid and euthyroid rats (Koller etal, 1987;Segerson et al, 1987) (Fig. 1). This effectwas observed only in neurons of the PVN,not in TRH-containing cells of the ventro-lateral hypothalamus or in the reticularnucleus of the thalamus. In contrast, coldexposure increased TRH mRNA levels inthe PVN (Zoeller et al, 1990), but not inTRH-containing neurons of the reticularnucleus of the thalamus. Hence, we haveused well-developed and well-characterizedmethods of manipulating the activity ofTRH neurons of the PVN to confirm thatTRH mRNA levels are regulated in a man-ner predicted by the hypothesis that changesin TRH expression are coupled to changesin neuronal activity (Fig. 1).
To approach the analysis of possible sub-sets of TRH neurons differentially regulatedby cold exposure and T3, we postulated thatif cold-activated TRH neurons in the PVNare separate from T3-inhibited TRH neu-rons, TRH mRNA levels should becomeelevated only in a subset of TRH neuronsin an animal exposed to cold, and reducedonly in a subset of TRH neurons in an ani-mal treated with T3. We tested this hypoth-esis by performing single-cell analysis ofTRH mRNA levels in animals with differ-ing thyroid status and exposed to differentambient temperatures (Zoeller et al, 1990).We found no evidence of functional subsetsof TRH neurons in the PVN (Fig. 2).
These studies demonstrate that cellularlevels of TRH mRNA are elevated by coldexposure in the same PVN neurons thatTRH mRNA levels are reduced by T3. Fur-ther, because previous studies have so clearlydemonstrated that T3 inhibits, and coldexposure stimulates, TRH release from neu-rons whose perikarya reside in the PVN, ourstudies indicate that changes in cellular lev-els of TRH mRNA in neurons of the PVN
Dow
nloaded from https://academ
ic.oup.com/icb/article/33/2/244/154040 by guest on 15 Septem
ber 2022

HVPOTHALAMIC INTEGRATION
B 200
247
R N -
PVN —<
Cont PTU T3 Cont PTU T3Treatments
FIG. 1. Quantitative in situ hybridization reveals that TRH mRNA levels in the rat PVN are reduced by T3and elevated by environmental temperature. Techniques for manipulating circulating levels of T3 and ambienttemperature, and for in situ hybridization and quantitative analysis of resulting film autoradiograms are describedin detail by Zoeller et al. (1990). Film autoradiograms are shown in (A) for an animal exposed to 25°C, and in(B) for an animal maintained at 5°C. The hybridization signal appears as an area of film darkening over thereticular nucleus of the thalamus (RN), paraventricular nucleus (PVN), and ventrolateral hypothalamus (V).Note that the density of the signal over the PVN in (B) is much greater than that in (A). The graph (C) displaysthe results of an experiment designed to establish the interaction of T3 and cold exposure on TRH mRNA levelsin the rat PVN. Cont- (untreated), PTU- [treated with 6-n-propyl-2-thiouracil (0.05% wt/wt in chow) for 12days], T3- (PTU-treated animals given a daily injection of 20 Mg/kg T3 for the final 4 days of the experiment)treated animals (n = 10) were maintained at 25°C (left panel) or transferred to an adjacent environmentalchamber maintained at 5°C (right panel) for 6 hr beginning at 0900 hr. Note that PTU (which decreases circulatinglevels of T3) increases (a) and T3 decreases (b) TRH mRNA levels in the PVN of animals at both temperatures.The observation that cold exposure significantly elevated TRH mRNA levels (c) only in euthyroid animals(untreated) suggests that T3 is a primary regulatory signal while environmental temperature can attenuate theefficacy of T3 negative feedback. Defining the cellular processes by which environmental temperature producesthis effect on TRH neurons of the PVN may reveal a molecular mechanism by which the "set point" aroundwhich negative feedback functions can be modulated by environmental factors.
mirror changes in the activity of these neu-rons. Therefore, as we study the molecularmechanisms by which trans-synaptic andendocrine signals interact to regulate cel-lular levels of TRH mRNA, we will gaininsight into how TRH neurons process theseconvergent signals to produce an integratedresponse. To further investigate these mech-anisms, we must characterize the secondmessenger systems activated by cold-stim-ulated afferents synapsing on TRH neurons,and determine whether TRH mRNA levelsare being regulated transcriptionally or post-transcriptionally. The observation that TRHneurons of the PVN remain viable andaggregated in culture (Fig. 3) will be essen-tial to these experiments.
Application to comparative endocrinologyThere are several approaches to measur-
ing mRNA levels, including northerns, dot-and slot-blot, and solution hybridization(Sambrook et al., 1989). Each of theseapproaches presents a specific set of strengthsand weaknesses which must be considered
when designing a particular experiment. Forexample, we decided to map the distribu-tion of TRH expressing cells in Xenopuslaevis brain to determine whether the TRHneuronal system in Xenopus is organized ina manner similar to that of mammals(Zoeller and Conway, 1989). Hence, wedesigned a 48-base oligomer complemen-tary to the TRH mRNA originally describedby Richter et al. (1984), and used this probeto visualize the widespread TRH-express-ing neuronal system by in situ hybridiza-tion. However, we found by northern anal-ysis that the probe hybridizes to several sizeclasses of mRNAs (Fig. 4). Kuchler et al.(1990) have confirmed this observation andhave demonstrated that these different sizeclasses of mRNA encode a precursor con-taining seven copies each of the TRH tri-peptide. Thus, to clarify the physiologicalsignificance of these multiple size classes ofTRH mRNA in Xenopus, we must deter-mine 1) whether these transcripts are dif-ferentially expressed; that is, whether theyare produced by a subset of TRH neurons,
Dow
nloaded from https://academ
ic.oup.com/icb/article/33/2/244/154040 by guest on 15 Septem
ber 2022

248 R. T . ZOELLER ET AL.
Increasing mRNA Level
50
40
g & 30
f ! ! ^ i •..*••,
r-l
D
n „Increasing mRNA Level
Increasing mRNA Level Increasing mRNA Level
FIG. 2. Single cell analysis of TRH mRNA levels demonstrate that T3 and cold exposure are acting on thesame population of TRH neurons in the rat PVN. A) Bright field micrograph of the in situ hybridization signalin Kodak NTB3 emulsion for TRH mRNA in the PVN from an animal exposed to cold. The autoradiographicsignal appears as dense clusters of grains over individual parvocellular elements along the border of the thirdventricle (III). Note that grain density over individual neurons varies within a single PVN. This variation canarise from differences in TRH mRNA levels among neurons, and from differences in the amount the neuroncontained within this 12 jim-thick section. B) Histogram relating the proportion of labeled neurons (frequency;ordinate) to grain density (abscissa; a measure of TRH mRNA level). Single cell analysis was performed on 100cells/animal for the 10 animals exposed to cold. C) Bright field micrograph of the PVN from an animal maintainedat 25°C after in situ hybridization for TRH mRNA and exposure to Kodak NTB3 nuclear track emulsion. Theautoradiographic signal appears as clusters of grains over individual parvocellular elements along the border ofthe third ventricle (III). Note that grain density over individual neurons in C is less than that over individualneurons in A. This is reflected in the frequency histogram plotted for control animals in D. The purpose of thisexercise was to test whether cold exposure increased TRH mRNA levels in a subset of TRH neurons in the ratPVN that is separate from that in which T3 reduces TRH mRNA levels. Hence, we compared the frequencyhistograms following single cell analysis of TRH mRNA levels in animals maintained at 25°C (open bars) withthose of animals exposed to cold (hatched bars) (E), and in animals treated with PTU (open bars) with those ofanimals treated with T3 (hatched bars) (F). If cold exposure increased TRH mRNA levels in a subset of PVNneurons, cold exposure would produce a bimodal distribution in the frequency histogram. Similarly, if T, actedon a subset of PVN neurons, we expected to see a bimodal distribution for both groups in F. We found noevidence of a bimodal distribution in E or F, and thus conclude that T3 and cold exposure are acting on thesame population of TRH neurons within the PVN. (Data redrawn from Zoeller et ai. 1990.)
Dow
nloaded from https://academ
ic.oup.com/icb/article/33/2/244/154040 by guest on 15 Septem
ber 2022

HYPOTHALAMIC INTEGRATION 249
while the TRH molecule is identical amongvertebrates, and the organization of themRNA (Richter et al., 1984; Lechan et al.,1986) and the neuronal system (Zoeller andConway, 1989) is remarkably similarbetween Xenopus and mammals, there areclear differences in the mechanisms regu-lating TRH expression among vertebratesthat require the use of different technicalapproaches.
FIG. 3. In situ hybridization for TRH mRNA inorganotypic cultures of rat PVN demonstrate that TRHneurons of the PVN remain aggregated after 21 daysof culture. The signal on film (left panel) appears as anarea of darkening in the center of the culture whichappears as a circular region of background staining,while in emulsion (right panel) the signal appears asgrains clustered over individual cells aggregated in thecenter of the cultured tissue. Coronal sections of 5 day-old rat pups were cultured according to the proceduresdescribed in Wray et al. (1989). This culture was pro-vided by Dr. Susan Wray, Laboratory of Neurochem-istry, NINDS, Bethesda.
2) whether potential regulators {e.g., T3 orenvironmental temperature) exert an effectspecifically on a particular transcript, and3) whether these regulatory effects areobserved in specific brain regions. Hence,
IDENTIFICATION OF A MODEL SYSTEM TOEXAMINE THE ROLE OF CO-LOCALIZEDNEUROTRANSMITTERS AS A CELLULAR
MECHANISM OF HYPOTHALAMICINTEGRATION
Neurons processing multiple signals mustpossess flexibility in their signalling mech-anism so that they can produce an inte-grated response. In principle, the signallingpotential of a neuron containing severaltransmitters may be affected by the concen-tration ratio of co-released factors (Fischer-Colbrie et al., 1988). Hence, differential reg-ulation of co-localized transmitter synthesismay be an important mechanism by whichneurons can integrate multiple signals.However, despite an abundance of anatom-ical data documenting cellular co-localiza-tion of multiple neurotransmitters in thecentral and peripheral nervous systems(Hokfelt et al, 1986), little is known aboutthe functional consequences of neurotrans-mitter co-release.
To address this issue experimentally, wehave focused on a population of neurons inthe suprachiasmatic nucleus (SCN) thatcontain two peptides derived from the samegene, VIP and peptide histidine isoleucine(PHI) (Nishizawa et al., 1985) as well as athird peptide derived from a different gene,gastrin-releasing peptide (GRP) (Okamuraet al., 1987; Albers et al., 1991). We con-sidered these neurons a useful "model,"using the criteria we applied to the TRHsystem of the PVN. VIP-containing neuronsof the SCN are clustered along the ventro-lateral portion of the SCN (Card et ai, 1981),and there exists a large literature of back-ground information about the function ofthe SCN. These data support the view thatthe SCN acts as a circadian clock that bothgenerates rhythms in behavior and physi-
Dow
nloaded from https://academ
ic.oup.com/icb/article/33/2/244/154040 by guest on 15 Septem
ber 2022
250 R. T . ZOELLER ET AL.
VIP PHI GRP V/P V/G P/GTreatment
V/P/G
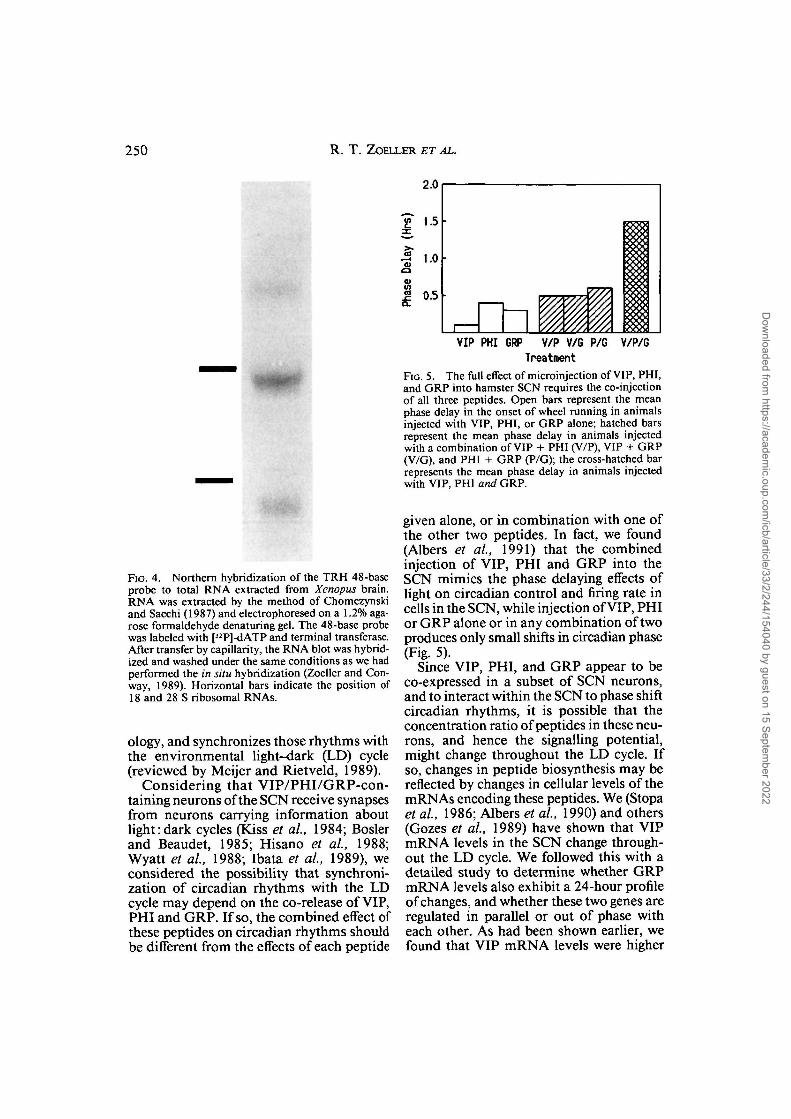
FIG. 5. The full effect of microinjection of VIP, PHI,and GRP into hamster SCN requires the co-injectionof all three peptides. Open bars represent the meanphase delay in the onset of wheel running in animalsinjected with VIP, PHI, or GRP alone; hatched barsrepresent the mean phase delay in animals injectedwith a combination of VIP + PHI (V/P), VIP + GRP(V/G), and PHI + GRP (P/G); the cross-hatched barrepresents the mean phase delay in animals injectedwith VIP, PHI and GRP.
FIG. 4. Northern hybridization of the TRH 48-baseprobe to total RNA extracted from Xenopus brain.RNA was extracted by the method of Chomczynskiand Sacchi (1987) and electrophoresed on a 1.2% aga-rose formaldehyde denaturing gel. The 48-base probewas labeled with [32P]-dATP and terminal transferase.After transfer by capillarity, the RNA blot was hybrid-ized and washed under the same conditions as we hadperformed the in situ hybridization (Zoeller and Con-way, 1989). Horizontal bars indicate the position of18 and 28 S ribosomal RNAs.
ology, and synchronizes those rhythms withthe environmental light-dark (LD) cycle(reviewed by Meijer and Rietveld, 1989).
Considering that VIP/PHI/GRP-con-taining neurons of the SCN receive synapsesfrom neurons carrying information aboutlight:dark cycles (Kiss et al, 1984; Boslerand Beaudet, 1985; Hisano et al, 1988;Wyatt et al, 1988; Ibata et al, 1989), weconsidered the possibility that synchroni-zation of circadian rhythms with the LDcycle may depend on the co-release of VIP,PHI and GRP. If so, the combined effect ofthese peptides on circadian rhythms shouldbe different from the effects of each peptide
given alone, or in combination with one ofthe other two peptides. In fact, we found(Albers et al, 1991) that the combinedinjection of VIP, PHI and GRP into theSCN mimics the phase delaying effects oflight on circadian control and firing rate incells in the SCN, while injection of VIP, PHIor GRP alone or in any combination of twoproduces only small shifts in circadian phase(Fig. 5).
Since VIP, PHI, and GRP appear to beco-expressed in a subset of SCN neurons,and to interact within the SCN to phase shiftcircadian rhythms, it is possible that theconcentration ratio of peptides in these neu-rons, and hence the signalling potential,might change throughout the LD cycle. Ifso, changes in peptide biosynthesis may bereflected by changes in cellular levels of themRNAs encoding these peptides. We (Stopaet al, 1986; Albers et al, 1990) and others(Gozes et al, 1989) have shown that VIPmRNA levels in the SCN change through-out the LD cycle. We followed this with adetailed study to determine whether GRPmRNA levels also exhibit a 24-hour profileof changes, and whether these two genes areregulated in parallel or out of phase witheach other. As had been shown earlier, wefound that VIP mRNA levels were higher
Dow
nloaded from https://academ
ic.oup.com/icb/article/33/2/244/154040 by guest on 15 Septem
ber 2022

HYPOTHALAMIC INTEGRATION 251
130
120
100
90
80
70
60
T
V0 4 8 12
Circadian16
Tine
V20 24
(Hrs)
FIG. 6. Twenty-four hours profile of changes in cel-lular levels of GRP (solid circles) and VIP/PHI (opencircles) mRNA in the SCN of rats. Points representmean (±SEM) mRNA levels standardized with respectto the value observed at circadian time 4 (CT-4).
during the dark phase than during the lightphase (Fig. 6); however, GRP mRNA levelswere higher during the light phase than dur-ing the dark phase.
These data support the hypothesis thatthe synchronization of circadian rhythmswith the LD cycle may depend on the co-release of VIP, PHI and GRP, and that theconcentration ratio of these co-localizedpeptides, and hence the signalling potential,might change throughout the LD cycle.Clearly, a complete test of this complexhypothesis must include experimentsdesigned to test questions about 1) the rela-tionship between VIP and GRP mRNA lev-els and the biosynthesis of their respectivepeptides, as well as 2) the physiologicaleffects of different concentration ratios ofthese peptides. However, this system isamenable to experimentation at both levels.The VIP/PHI/GRP neurons are spatiallyrestricted so that they can be harvested forbiochemical analyses and culturing. Thereis a wealth of information about the func-tion of the SCN; its role in the control ofcircadian activity rhythms, tracked by mea-suring wheel running behavior, provides aconvenient assay for SCN activity and therole these neurons may play in the controlof that activity. This assay for SCN activitywill allow us to determine whether all VIP/GRP neurons in the SCN are regulated bythe same set of stimuli. And finally, culture
techniques are available to study cellularmechanisms in the SCN.
CONCLUSIONS
Our view of the mechanisms by whichthe brain processes the myriad internal andexternal stimuli to coordinate behavioral,endocrine and autonomic processes is beinggreatly enhanced by progress in molecularbiology and the development of the kindsof hybridization techniques (e.g., in situhybridization) with the quantitative sensi-tivity and anatomical resolution requiredfor application to the central nervous sys-tem. As we apply these techniques to learnmore about the cellular mechanisms bywhich neurons integrate a variety of infor-mation, we will gain a clearer understandingof the underlying biology we observe in thelife histories of animals, including humans.However, to relate the molecular and cel-lular events occurring in the nervous systemto behavioral, endocrine and autonomicphenomena, we must combine these molec-ular techniques with the "traditional" tech-niques which have allowed us to so clearlydefine the major issues of today.
ACKNOWLEDGMENTS
We thank Dr. Sandra Petersen, ProfessorFrank L. Moore and members of his labo-ratory for helpful comments made on theinitial drafts of this manuscript.
REFERENCES
Albers, H. E., E. G. Stopa, R. T. Zoeller, J. S. Kauer,J. C. King, J. S. Fink, H. Mobtaker, and H. Wolfe.1990. Day-night variation in prepro vasoactiveintestinal peptide/peptide histidine isoleucinemRNA within the rat suprachiasmatic nucleus.Mol. Brain Res. 7:85-89.
Albers, H. E., S. Y. Liou, E. G. Stopa, and R. T. Zoeller.1991. Interaction of co-localized neuropeptides:Functional significance in the circadian timing sys-tem. J. Neurosci. 11:846-851.
Arancibia, S., L. Tapia-Arancibia, H. Astier, and I.Assenmacher. 1989. Physiological evidence foralpha 1-adrenergic facilitatory control of the cold-induced TRH release in the rat, obtained by push-pull cannulation of the median eminence. Neu-rosci. Lett. 100:169-174.
Bosler, O. and A. Beaudet. 1985. VIP neurons asprime synaptic targets for serotonin afferents in ratsuprachiasmatic nucleus: A combined radioau-tographic and immunocytochemical study. J.Neurocytol. 14:749-763.
Dow
nloaded from https://academ
ic.oup.com/icb/article/33/2/244/154040 by guest on 15 Septem
ber 2022

252 R. T. ZOELLER ET AL.
Brownstein, M. J., R. D. Utiger, M. Palkovits, and J.S. Kizer. 1975. Effect of hypothalamic deaffer-entation on thyrotropin-releasing hormone levelsin rat brain. Proc. Natl. Acad. Sci. U.S.A. 72:4177-4179.
Burgus, R., T. F. Dunn, D. Desiderio, D. N. Ward, W.Vale, and R. Guillemin. 1970. Characterizationof ovine hypothalamic hypophysiotropic TSH-releasing factor. Nature 226:321-323.
Card, J. P., N. Brecha, H. J. Karten, and R. Y. Moore.1981. Immunocytochemical localization of vaso-active intestinal polypeptide-containing cells andprocesses in the suprachiasmatic nucleus of therat: Light and electron microscopic analysis. J.Neurosci. 1:1289-1303.
Chomczynski, P. and N. Sacchi. 1987. Single-stepmethod of RNA isolation by acid guanidiniumthiocyanate-phenol-chloroform extraction. Anal.Biochem. 162:156-159.
Comb, M., S. E. Hyman, and H. M. Goodman. 1987.Mechanisms of trans-synaptic regulation of geneexpression. Trends Neurosci. 10:473-478.
Fischer-Colbrie, R., A. Iacangelo, and L. E. Eiden.1988. Neural and humoral factors separately reg-ulate neuropeptide Y, enkephalin, and chroma-granin A and B mRNA levels in rat adrenalmedulla. Proc. Natl. Acad. Sci. U.S.A. 85:3240-3244.
Folkers, K., F. Enzmann, J. Boler, C. Y. Bowers, andA. V. Schally. 1969. Discovery of modificationof the synthetic tripeptide-sequence of the thyro-tropin releasing hormone having activity. Bio-chem. Biophys. Res. Commun. 37:123-126.
Goodman, R. H. 1990. Regulation of neuropeptidegene expression. Annu. Rev. Neurosci. 13:111-127.
Gozes, I., Y. Shani, B. Liu, and J. P. H. Burbach. 1989.Diurnal variation in vasoactive intestinal peptidemessenger RNA in the suprachiasmatic nucleus ofthe rat. Neurosci. Res. Commun. 5:83-86.
Habener, J. F. 1990. Cyclic AMP response elementbinding proteins: A cornucopia of transcriptionfactors. Mol. Endocrinol. 4:1087-1094.
Hefco, E., L. Krulich, P. Illner, and P. R. Larsen. 1975.Effect of acute exposure to cold on the activity ofthe hypothalamic-pituitary-thyroid system. Endo-crinology 97:1185-1195.
Hisano, S., H. Ishizuka, T. Nishiyama, Y. Tsuruo, S.Katoh, and S. Daikoku. 1986. Immunoelectronmicroscopic observations of hypothalamic TRH-containing neurons in rats. Exp. Brain Res. 63:495-504.
Hisano, S., M. Chikamori-Aoyama, S. Katoh, Y.Kagotani, S. Daikoku, and K. Chihara. 1988.Suprachiasmatic nucleus neurons immunoreac-ti ve for vasoactive intestinal peptide have synapticcontacts with axons immunoreactive for neuro-peptide Y: An immunoelectronmicroscopic studyin the rat. Neurosci. Lett. 88:145-150.
Hokfelt, T., K. Fuxe, and B. Pernow. 1986. Coex-istence of neuronal messengers: A new principlein chemical transmission. Progress Brain Res. 68:3-404.
Ibata, Y., Y. Takahashi, H. Okamura, K. Fumio, H.Terubayashi, T. Kubo, and N. Yanaihara. 1989.Vasoactive intestinal peptide (VlP)-like immu-noreactive neurons located in the rat suprachias-matic nucleus receive a direct retinal projection.Neurosci. Lett. 97:1-5.
Ishikawa, K., Y. Taniguchi, K. Inoue, K. Kurosumi,and M. Suzuki. 1988. Immunocytochemicaldelineation of the thyrotrophic area: Origin of thy-rotropin-releasing hormone in the median emi-nence. Neuroendocrinology 47:384-388.
Jackson, I. M. D., P. Wu, and R. M. Lechan. 1985.Immunohistochemical localization in the rat brainof the precursor for thyrotropin-releasing hor-mone. Science 229:1097-1099.
Jobin, M., L. Ferland, J. Cote, and F. Labrie. 1975.Effect of exposure to cold on hypothalamic TRHactivity and plasma levels of TSH and prolactinin the rat. Neuroendocrinology 18:204-212.
Jobin, M., L. Ferland, and F. Labrie. 1976. Effect ofpharmacological blockade of ACTH and TSHsecretion on the acute stimulation of prolactinrelease by exposure to cold and ether stress. Endo-crinology 99:146-151.
Kiss, J., C. Leranth, and B. Halasz. 1984. Serotonin-ergic endings of VIP-neurons in the suprachias-matic nucleus and on ACTH-neurons in the arcu-ate nucleus of the rat hypothalamus. A combinationof high resolution autoradiography and electronmicroscopic immunocytochemistry. Neurosci.Lett. 44:119-124.
Koenig, R. J., D. Senator, and P. R. Larsen. 1984.Phorbol esters as probes of the regulation of thy-rotropin secretion. Biochem. Biophys. Res. Com-mun. 125:353-359.
Koller, K. J., R. S. Wolff, M. K. Warden, and R. T.Zoeller. 1987. Thyroid hormones regulate levelsof thyrotropin-releasing hormone mRNA in theparaventricular nucleus. Proc. Natl. Acad. Sci.U.S.A. 84:7329-7333.
Krulich, L., A. Giachetti, A. Marchlewska-Koj, E.Hefco, and H. E. Jameson. 1977. On the role ofthe central noradrenergic and dopaminergic sys-tems in the regulation of TSH secretion in the rat.Endocrinology 100:496-505.
Kuchler, K., K. Richter, J. Trnovsky, R. Egger, and G.Kreil. 1990. Two precursors of thyrotropin-releasing hormone from skin ofXenopus laevis. J.Biol. Chem. 265:11731-11733.
Lechan, R. M., P. Wu, and I. M. D. Jackson. 1986.Immunolocalization of the thyrotropin-releasinghormone prohormone in the rat central nervoussystem. Endocrinology 119:1210-1216.
Liao, N., M. Bulant, P. Nicolas, H. Vaudry, and G.Pelletier. 1985. Immunocytochemical distribu-tion of neurons containing a peptide derived fromthyrotropin-releasing hormone precursor in the ratbrain. Neurosci. Lett. 85:24-28.
Liposits, Z., W. K. Paull, P. Wu, I. M. D. Jackson, andR. M. Lechan. 1987. Hypophysiotrophic thy-rotropin releasing hormone (TRH) synthesizingneurons. Ultrastructure, adrenergic innervation andputative transmitter action. Histochemistry 88:1-10.
Dow
nloaded from https://academ
ic.oup.com/icb/article/33/2/244/154040 by guest on 15 Septem
ber 2022

HYPOTHALAMIC INTEGRATION 253
Mannisto, P. T. 1983. Central regulation of thyro-tropin secretion in rats: Methodological aspects,problems and some progress. Med. Biol. 61:92-100.
Meijer, J. H. and W. J. Rietveld. 1989. Neurophys-iology of the suprachiasmatic circadian pacemakerin rodents. Physiol. Rev. 69:671-707.
Mori, M. and M. Yamada. 1987. Thyroid hormonesregulate the amount of thyrotropin-releasing hor-mone in the hypothalamic median eminence ofthe rat. J. Endocrinol. 114:443-448.
Mori, M., I. Kobayashi, and K. Wakabayashi. 1978.Suppression of serum thyrotropin (TSH) concen-trations following thyroidectomy and cold-expo-sure by passive immunization with antiserum tothyrotropin-releasing hormone (TRH) in rats.Metabolism 27:1485-1490.
Morley, J. E. 1981. Neuroendocrine control of thy-rotropin secretion. Endocr. Rev. 2:396-436.
Nair, R. M., J. F. Barrett, C. Y. Bowers, and A. V.Schally. 1970. Structure of porcine thyrotropinreleasing hormone. Biochemistry 9:1103—1106.
Nishizawa, M., Y. Hayakawa, N. Yanaihara, and H.Okamoto. 1985. Nucleotide sequence diver-gence and functional constraint in VIP precursormRNA evolution between human and rat. FEBSLett. 183:55-59.
Okamura, H, Y. Takahashi, H. Terubayashi, S.Hamada, N. Yanaihara, and Y. Ibata. 1987.Coexistence of vasoactive intestinal peptide (VIP)-,peptide histidine isoleucine amide (PHI)-, and gas-trin releasing peptide (GRP)-like immunoreactiv-ity in neurons of the rat suprachiasmatic nucleus.Bio. Res. 7:295-299.
Onaya, T. and K. Hashizume. 1977. Effects of drugsthat modify brain biogenic amine concentrationson thyroid activation induced by exposure to cold.Neuroendocrinology 20:47-58.
Palkovits, M., R. L. Eskay, and M. J. Brownstein.1982. The course of thyrotropin-releasing hor-mone fibers to the median eminence in rats. Endo-crinology 110:1526-1528.
Prasad, C, J. J. Jacobs, and J. F. Wilber. 1980.Immunological blockade of endogenous thyrotro-pin-releasing hormone produces hypothermia inrats. Brain Res. 193:580-583.
Richter, K.., E. Kawashima, R. Egger, and G. Kreil.1984. Biosynthesis of thyrotropin-releasing hor-mone in the skin of Xenopus laevis: Partial sequenceof the precursor deduced from cloned cDNA.EMBOJ. 3:617-621.
Rondeel, J. M. M., W. J. deGreef, P. van der Schoot,B. Karels, W. Klootwijk, and T. J. Visser. 1988.Effect of thyroid status and paraventricular arealesions on the release of thyrotropin-releasing hor-mone and catecholamines into hypophysial portalblood. Endocrinology 123:523-527.
Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989.Molecular cloning. A laboratory manual. ColdSpring Harbor Laboratory Press, Cold Spring Har-bor.
Schettini, G., A. Quattrone, G. Di Renzo, G. Lom-bardi, and P. Preziosi. 1979. Effect of6-hydroxydopamine treatment on TSH secretion
in basal and cold-stimulated conditions in the rat.Eur. J. Pharmacol. 56:153-157.
Segerson, T. P., J. Kauer, H. C. Wolfe, H. Mobtaker,P. Wu, I. M. D. Jackson, and R. M. Lechan. 1987.Thyroid hormone regulates TRH biosynthesis inthe paraventricular nucleus of the rat hypothala-mus. Science 238:78-80.
Shupnik, M. A., S. L. Greenspan, and E. C. Ridgway.1986. Transcriptional regulation of thyrotropinsubunit genes by thyrotropin-releasing hormoneand dopamine in pituitary cell culture. J. Biol.Chem. 261:12675-12679.
Stopa, E. G., N. Minamitani, J. A. Jonassen, J. C. King,H. Wolfe, H. Mobtaker, and H. E. Albers. 1988.Localization of vasoactive intestinal peptide andpeptide histidine isoleucine immunoreacti vity andmRNA within the rat suprachiasmatic nucleus.Mol. Brain Res. 4:319-325.
Szabo, M. and L. A. Frohman. 1977. Suppression ofcold-stimulated thyrotropin secretion by antise-rum to thyrotropin-releasing hormone. Endocri-nology 101:1023-1033.
Szabo, M., N. Kovathana, K. Gordon, and L. A. Froh-man. 1978. Effect of passive immunization withan antiserum to thyrotropin (TSH)-releasing hor-mone on plasma TSH levels in thyroidectomizedrats. Endocrinology 102:799-805.
Taylor, T., N. Gesundheit, and B. D. Weintraub.1986. Effects of in vivo bolus versus continuousTRH administration on TSH secretion, biosyn-thesis, and glycosylation in normal and hypothy-roid rats. Mol. Cell Endocrinol. 46:253-261.
Taylor, T., F. E. Wondisford, T. Blaine, and B. D.Weintraub. 1990. The paraventricular nucleusof the hypothalamus has a major role in thyroidhormone feedback regulation of thyrotropin syn-thesis and secretion. Endocrinology 126:317-324.
Terry, L. C. 1986. Regulation of thyrotropin secre-tion by the central epinephrine system. Neuroen-docrinology 42:102-108.
Uhl, G. R. and T. Nishimori. 1990. Neuropeptidegene expression and neural activity: Assessing aworking hypothesis in nucleus caudalis and dorsalhorn neurons expressing preproenkephalin andpreprodynorphin. Cell. Mol. Neurobiol. 10:73-98.
Van Nguyen, T., L. Kobierski, M. Comb, and S. E.Hyman. 1990. The effect of depolarization onexpression of the human proenkephalin gene issynergistic with cAMP and dependent upon cAMP-inducible enhancer. J. Neurosci. 10:2825-2833.
Wray, S., R. T. Zoeller, and H. Gainer. 1989. Dif-ferential effects of estrogen on luteinizing hor-mone-releasing hormone gene expression in sliceexplant cultures prepared from specific rat fore-brain regions. Mol. Endocrinol. 3:1197-1206.
Wyatt, L. M., R. B. Norgren, and M. N. Lehman.1988. Retinal and neuropeptide Y innervation ofthe hamster suprachiasmatic nucleus: Light andelectron microscopic observations. Soc. Neurosci.Abstr. 14:50.
Yamada, M., D. Rogers, and J. F. Wilber. 1989.Exogenous triiodothyronine lowers thyrotropin-releasing concentrations in the specific hypotha-lamic nucleus (paraventricular) involved in thy-
Dow
nloaded from https://academ
ic.oup.com/icb/article/33/2/244/154040 by guest on 15 Septem
ber 2022

254 R. T . ZOELLER ET AL.
rotropin regulation and also in posterior nucleus.Neuroendocrinology 50:560-563.
Young, W. S., HI and R. T. Zoeller. 1987. Neuroen-docrine gene expression in the hypothalamus: Insitu hybridization histochemical studies. Cell. Mol.Neurobiol. 7:353-366.
Zoeller, R. T. and K. M. Conway. 1989. Neuronsexpressing thyrotropin-releasing hormone-likemessenger ribonucleic acid are widely distributed
in Xenopus laevis brain. Gen. Comp. Endocrinol.76:139-146.
Zoeller, R. T., N. Kabeer, and H. E. Albers. 1990.Cold exposure elevates cellular levels of messengerribonucleic acid encoding thyrotropin-releasinghormone in paraventricular nucleus despite ele-vated levels of thyroid hormones. Endocrinology127:2955-2962.
Dow
nloaded from https://academ
ic.oup.com/icb/article/33/2/244/154040 by guest on 15 Septem
ber 2022




















