
Environmental enrichment mitigates the sex-specific effects of gestational inflammation on social...
13
Environmental enrichment mitigates the sex-specific effects of gestational inflammation on social engagement and the hypothalamic pituitary adrenal axis-feedback system E.J. Connors a , A.N. Shaik b , M.M. Migliore b , A.C. Kentner a,⇑ a School of Arts & Sciences, Health Psychology Program, MCPHS University (formerly Massachusetts College of Pharmacy & Health Sciences), Boston, MA 02115, United States b School of Pharmacy, MCPHS University, Boston, MA 02115, United States article info Article history: Received 22 April 2014 Received in revised form 27 June 2014 Accepted 27 June 2014 Available online 7 July 2014 Keywords: Environmental enrichment Gestational inflammation Social interaction Spatial discrimination Hippocampus NR2B receptor subunit Glutamate Glucocorticoid receptor Corticosterone abstract Modest environmental enrichment (EE) is well recognized to protect and rescue the brain from the con- sequences of a variety of insults. Although animal models of maternal immune activation (MIA) are asso- ciated with several neurodevelopmental impairments in both the behavioral and cognitive functioning of offspring, the impact of EE in protecting or reversing these effects has not been fully evaluated. In the present study, female Sprague–Dawley rats were randomized into EE (pair-housed in a large multi-level cage with toys, tubes and ramps) or animal care control (ACC; pair-housed in standard cages) conditions. Each pair was bred, following assignment to their housing condition, and administered 100 lg/kg of lipo- polysaccharide (LPS) on gestational day 11. After birth, and until the end of the study, offspring were maintained in their respective housing conditions. EE protected against both the social and hypothalamic pituitary adrenal axis consequences of MIA in juvenile male rats, but surprisingly not against the spatial discrimination deficits or accompanying decrease in glutamate levels within the hippocampus (as mea- sured via LCMS–MS). Based on these preliminary results, the mechanisms that underlie the sex-specific consequences that follow MIA appear to be dependent on environmental context. Together, this work highlights the importance of environmental complexity in the prevention of neurodevelopmental deficits following MIA. Ó 2014 Elsevier Inc. All rights reserved. 1. Introduction Programming cues such as physical or psychogenic stressors, experienced during gestation, have a far reaching impact on gen- eral health (Lipkind et al., 2010; Nugent et al., 2011; see Pembrey, 2002; Soberanes-Chávez et al., 2013) and behavioral/ cognitive functioning (Brown, 2012; Brown and Derkits, 2010; Mueller and Bale, 2008). Preclinical studies are particularly sugges- tive of the involvement of maternal immune activation (MIA) in later social and cognitive impairments of offspring (see Patterson, 2009; Meyer et al., 2007). For example, MIA has been associated with impaired object recognition (Coyle et al., 2009; Graciarena et al., 2010) and spatial memory (Golan et al., 2005; Howland et al., 2012), decreased prepulse (Fortier et al., 2007; Wolff and Bilkey, 2010) and latent (Bitanihirwe et al., 2010; Meyer et al., 2006) inhibition, increased amphetamine induced locomotion (Fortier et al., 2004; Poinkewitz et al., 2011), and altered social behaviors (Bitanihirwe et al., 2010; Hava et al., 2006; Malkova et al., 2012). Additionally, in animal models of MIA, the associated brain changes are reported to parallel the pathogenesis of schizo- phrenia and autism including diminished NMDA receptor func- tioning, imbalances in dopamine regulation, reductions in the glycoprotein reelin, and general immune dysregulation (Coyle et al., 2003; Meyer et al., 2008; see Patterson, 2009; Shi et al., 2003). Moreover, the behavioral impairments observed in offspring following MIA may be reversed by schizophrenia drug treatments (Shi et al., 2003). However, it is rare that studies investigate inter- ventions that may protect against these neurodevelopmental effects of MIA. Animals models of MIA utilize pathogens such as gram negative bacteria (lipopolysaccharide; LPS) and viruses (polyinositic:polyci- tidylic acid; polyI:C) to activate Toll-like receptors located on immune cells distributed throughout the brain and periphery (reviewed in Meyer et al., 2007). Lipopolysaccharide (LPS) mimics Gram negative infection by activating Toll-like receptor 4, which in turn initiates intracellular signaling events that lead to the synthesis of a variety of pro-inflammatory molecules such as http://dx.doi.org/10.1016/j.bbi.2014.06.020 0889-1591/Ó 2014 Elsevier Inc. All rights reserved. ⇑ Corresponding author. Tel.: +1 617 274 3360 (O); fax: +1 617 732 2959. E-mail address: [email protected] (A.C. Kentner). Brain, Behavior, and Immunity 42 (2014) 178–190 Contents lists available at ScienceDirect Brain, Behavior, and Immunity journal homepage: www.elsevier.com/locate/ybrbi
Transcript of Environmental enrichment mitigates the sex-specific effects of gestational inflammation on social...

Brain, Behavior, and Immunity 42 (2014) 178–190
Contents lists available at ScienceDirect
Brain, Behavior, and Immunity
journal homepage: www.elsevier .com/locate /ybrbi
Environmental enrichment mitigates the sex-specific effects ofgestational inflammation on social engagement and the hypothalamicpituitary adrenal axis-feedback system
http://dx.doi.org/10.1016/j.bbi.2014.06.0200889-1591/� 2014 Elsevier Inc. All rights reserved.
⇑ Corresponding author. Tel.: +1 617 274 3360 (O); fax: +1 617 732 2959.E-mail address: [email protected] (A.C. Kentner).
E.J. Connors a, A.N. Shaik b, M.M. Migliore b, A.C. Kentner a,⇑a School of Arts & Sciences, Health Psychology Program, MCPHS University (formerly Massachusetts College of Pharmacy & Health Sciences), Boston, MA 02115, United Statesb School of Pharmacy, MCPHS University, Boston, MA 02115, United States
a r t i c l e i n f o
Article history:Received 22 April 2014Received in revised form 27 June 2014Accepted 27 June 2014Available online 7 July 2014
Keywords:Environmental enrichmentGestational inflammationSocial interactionSpatial discriminationHippocampusNR2B receptor subunitGlutamateGlucocorticoid receptorCorticosterone
a b s t r a c t
Modest environmental enrichment (EE) is well recognized to protect and rescue the brain from the con-sequences of a variety of insults. Although animal models of maternal immune activation (MIA) are asso-ciated with several neurodevelopmental impairments in both the behavioral and cognitive functioning ofoffspring, the impact of EE in protecting or reversing these effects has not been fully evaluated. In thepresent study, female Sprague–Dawley rats were randomized into EE (pair-housed in a large multi-levelcage with toys, tubes and ramps) or animal care control (ACC; pair-housed in standard cages) conditions.Each pair was bred, following assignment to their housing condition, and administered 100 lg/kg of lipo-polysaccharide (LPS) on gestational day 11. After birth, and until the end of the study, offspring weremaintained in their respective housing conditions. EE protected against both the social and hypothalamicpituitary adrenal axis consequences of MIA in juvenile male rats, but surprisingly not against the spatialdiscrimination deficits or accompanying decrease in glutamate levels within the hippocampus (as mea-sured via LCMS–MS). Based on these preliminary results, the mechanisms that underlie the sex-specificconsequences that follow MIA appear to be dependent on environmental context. Together, this workhighlights the importance of environmental complexity in the prevention of neurodevelopmental deficitsfollowing MIA.
� 2014 Elsevier Inc. All rights reserved.
1. Introduction
Programming cues such as physical or psychogenic stressors,experienced during gestation, have a far reaching impact on gen-eral health (Lipkind et al., 2010; Nugent et al., 2011; seePembrey, 2002; Soberanes-Chávez et al., 2013) and behavioral/cognitive functioning (Brown, 2012; Brown and Derkits, 2010;Mueller and Bale, 2008). Preclinical studies are particularly sugges-tive of the involvement of maternal immune activation (MIA) inlater social and cognitive impairments of offspring (see Patterson,2009; Meyer et al., 2007). For example, MIA has been associatedwith impaired object recognition (Coyle et al., 2009; Graciarenaet al., 2010) and spatial memory (Golan et al., 2005; Howlandet al., 2012), decreased prepulse (Fortier et al., 2007; Wolff andBilkey, 2010) and latent (Bitanihirwe et al., 2010; Meyer et al.,2006) inhibition, increased amphetamine induced locomotion(Fortier et al., 2004; Poinkewitz et al., 2011), and altered social
behaviors (Bitanihirwe et al., 2010; Hava et al., 2006; Malkovaet al., 2012). Additionally, in animal models of MIA, the associatedbrain changes are reported to parallel the pathogenesis of schizo-phrenia and autism including diminished NMDA receptor func-tioning, imbalances in dopamine regulation, reductions in theglycoprotein reelin, and general immune dysregulation (Coyleet al., 2003; Meyer et al., 2008; see Patterson, 2009; Shi et al.,2003). Moreover, the behavioral impairments observed in offspringfollowing MIA may be reversed by schizophrenia drug treatments(Shi et al., 2003). However, it is rare that studies investigate inter-ventions that may protect against these neurodevelopmentaleffects of MIA.
Animals models of MIA utilize pathogens such as gram negativebacteria (lipopolysaccharide; LPS) and viruses (polyinositic:polyci-tidylic acid; polyI:C) to activate Toll-like receptors located onimmune cells distributed throughout the brain and periphery(reviewed in Meyer et al., 2007). Lipopolysaccharide (LPS) mimicsGram negative infection by activating Toll-like receptor 4, whichin turn initiates intracellular signaling events that lead to thesynthesis of a variety of pro-inflammatory molecules such as

E.J. Connors et al. / Brain, Behavior, and Immunity 42 (2014) 178–190 179
interleukin (IL)-1b, IL-6 and tumor necrosis factor (TNF)-a (Blatteiset al., 2004; Dinarello, 1999; Steiner et al., 2006). These moleculesactivate the central nervous system and de novo synthesis ofcytokines within the brain that are responsible for the initiationof fever and its accompanying constellation of behavioral andcognitive disruptions (i.e., sleep changes, lethargy, performancedisruptions on memory tasks, reduced food and water intake,etc.), known as sickness behaviors (Maier et al., 1998). In animalmodels of MIA, these behavioral and physiological markers canbe used to verify the initiation of the sickness response (Frenchet al., 2013), without stressing the mother by collecting serologicalsamples to confirm inflammation. When evaluating gestationalinterventions that may mitigate the effects of MIA, such confirma-tion is necessary to determine if their success may be due to adirect prevention of sickness in the mother.
In contrast to the detrimental effects of MIA, environmentalenrichment (EE) has been shown to improve performance on avariety of learning and memory measures such as object recogni-tion (Leger et al., 2012) and spatial recall (Huang et al., 2006) tasks,and has even been reported to enhance social play behavior(Morley-Fletcher et al., 2003; Schneider et al., 2006). In the labora-tory, EE typically takes the form of a larger home cage filled withnovel toys of varying sizes, shapes and textures, which increasean animal’s opportunity for exploration and activity. This housingenvironment has also been attributed to the prevention and rever-sal of the detrimental effects of a variety of challenges to the adultnervous system. For example, EE has been reported to delay motordeficits in an animal model of Huntington’s Disease (Spires et al.,2004), rescue sensorimotor and spatial memory impairments afterstroke (Dahlqvist et al., 2004; Ohlsson and Johansson, 1995), atten-uate inflammation and the associated physiological and cognitivedisruptions that accompany infection (Jurgens and Johnson,2012; Kentner et al., 2008), and prevent functional deficits follow-ing traumatic brain injury (Johnson et al., 2013). There is also areport that EE counteracts the decreased social exploration andlatency that accompanies prenatal valproic acid exposure(Schneider et al., 2006), which attenuates proinflammatory cyto-kines and reactive oxygen species (Cardinale et al., 2010). How-ever, to-date, studies evaluating the effects of EE following early-life insults have focused on maternal separation (Francis et al.,2002; Vivinetto et al., 2013) and prenatal stressors such asrestraint (Li et al., 2012; Morley-Fletcher et al., 2003) or early expo-sure to drugs of abuse/misuse (i.e., nicotine; Mychasiuk et al.,2014), but not MIA.
Therefore, in the present study we employed an early gesta-tional LPS challenge and (1) confirmed MIA by passively quantify-ing sickness behaviors in the dam, and (2) assessed the protectiveeffects of combined pre- and post-weaning EE against MIA-inducedsocial and cognitive disruptions and (3) their associated markers ofhypothalamic pituitary adrenal (HPA) axis and glutamatergic func-tioning in her juvenile male and female offspring.
2. Materials and methods
All experimental procedures were carried out in accordancewith recommendations in the Guide for the Care and Use of Labo-ratory Animals of the National Institutes of Health and the studyprotocol approved by the Institutional Animal Care and Use Com-mittee at MCPHS.
2.1. Animals and housing
Male and female Sprague–Dawley rats were obtained fromCharles River (Wilmington, MA) and housed at 20 �C on a 12 hlight/dark cycle (0700–1900 light) with food and water available
ad libitum. Upon arrival, female rats were pair-housed in eitherEE (large multi-level cage with toys changed 2 times weekly, tubes,chew bone, Nestlets� and ramps; Critter Nation, Muncie IN) orAnimal Care Control (ACC; standard cage with tube, chew bone,and Nestlets�) housing. Male rats were pair-housed in standardconditions. Rats were differentiated by a non-scented/non-toxiccolored marking on their tail. A flowchart of the study procedurescan be found in Fig. 1.
2.2. Breeding
Following one-week of acclimatization, estrous cycles wereevaluated daily (Marcondes et al., 2002) for an additional weekin order to determine individual cycle length for later pregnancyverification. During proestrous, a male rat was introduced tofemales in a 2:1 female:male ratio. The next morning vaginal sam-ples were collected and breeding (gestational day 1) established bythe presence of both spermatozoa and vaginal cells typical ofestrus. Male breeders were then returned to their home cage. Preg-nancy was confirmed by observations of the diestrus phase fromvaginal samples, continued weight gain of more than 3–5 g/day,and visible teats during the later phase of gestation.
2.3. Gestational treatment and sickness evaluation
On the morning of gestational day (G)11 (between 9:00 and10:00 h), ACC dams were separated into individual clean cages sothat we could accurately measure food and water intake, and othersickness behaviors, following lipopolysaccharide (LPS) administra-tion. EE dams were separated from their partner by a physical divi-der for the same reason. The physical divider (build directly intothe EE cage) allowed for olfactory, auditory, and some tactile andvisual contact, important components of EE conditions. Between11:30 and 12:30 h, each dam was given an i.p. injection of eitherLPS (Escherichia coli, serotype 026:B6; L-3755, Sigma, St. Louis,MO; 100 lg/kg) in pyrogen-free saline, or an equivalent volumeof pyrogen-free saline. The LPS injection produces a short-livedinflammatory response (Steiner et al., 2006). For each housing pair,one dam received LPS and the other received saline.
In order to confirm the sickness-inducing effects of gestationalLPS, sickness behaviors were evaluated. Dams were passivelyassessed on measures of ptosis (droopy eyelids), piloerection (ruf-fled, unkempt coat appearance), and lethargy by an observer blindto the drug treatment. Each behavior was scored on a three pointscale (none = 0, mild = 1, or severe = 2), as previously described(Kentner et al., 2006, 2007) at 60, 90, 105, 120 and 210 min postinjection. Based on the presence and severity of these behaviors,rats were assigned a total composite sickness score at each timepoint (adapted from Hayley et al., 2002; Kentner et al., 2007) as fol-lows: 0 = ‘no sickness’ indicated by all behaviors being scored asnone, or one behavior scored as mild; 1 = two or more mild scores,no severe scores (i.e., mild lethargy and ptosis and/or piloerection);2 = severe score on one sickness behavior (mild or none scored onall other behaviors); 3 = severe score on two sickness behaviors;4 = ‘very sick appearance’ as indicated by 3 severe scores (severeptosis, lethargy and piloerection). Anorectic effects were evaluatedby weighing food and water bottles immediately before LPS treat-ment, and again 20 h later, in order to determine food and waterintake. Body weights were also evaluated 20 h after injection atwhich point dams were reunited with their cage-mate in theirrespective housing assignments.
2.4. Parturition, litter phenotype and maternal behavior
On G19, ACC dams were again separated into individual cageswhere they stayed alone with their litters until weaning. EE female

Fig. 1. Flow chart of study design.
180 E.J. Connors et al. / Brain, Behavior, and Immunity 42 (2014) 178–190
rats were separated by physical dividers (as described in Sec-tion 2.3), maintaining their housing condition. Rat dams were sep-arated at this time point to prevent the mixing of littersimmediately following birth. Day of birth was designated as post-natal (P)1.
On P2 all litters were adjusted to 10 pups, with approximatelyequal ratios of males to females, and each pup was identified asbelonging to a specific dam using a non-scented non-toxic marker.To do this, dams were removed from their home cage and pupswere marked and immediately replaced onto their nest. Damsreceived the same colored marking as their litter on their tail andwere then returned to the home cage. This process took approxi-mately 3 min per cage. All pups from each housing condition weremarked daily until their ears could be clipped on P14, clearly iden-tifying them. Pair-housed EE dams gave birth within two days ofeach other, at which point the partition separating litters wasremoved when the later litter reached P2. The ACC dams pairedduring breeding (saline and LPS) that successfully gave birth alsodid so within two days of each other.
Passive maternal observations took place in the home cage fromP3 to P6 at 06:00, 12:00, and 20:30 h. During the dark phase dimred lights were used to facilitate viewing. Maternal behaviors weremanually recorded for 10 one-minute intervals per observationsession, spaced at least 5 min apart. Examples of behaviors scoredincluded the frequency of licking/grooming, pup retrieval, mater-nal time on nest, in addition to the number of pups (and gesta-tional treatment distribution in the case of paired EE dams) on
the nest. Maternal evaluations only took place when the identityof each dam was accurately determined (in the case of the pair-housed EE animals) by the colored making on their tail.
Weaning took place on P22. This involved the removal of thedam and placing her offspring into a clean cage, maintaining theirrespective neonatal housing conditions. EE offspring were housedin same-sex groups of four-six while ACC animals were housedin pairs. Offspring remained in these housing conditions until P40.
2.5. Offspring physiology and behavior
All male and female offspring were weighed weekly betweenP22 and the end of the study. One male and one female rat per lit-ter was used for each behavioral test and tissue collection timepoint described below. Beginning on P36, animals were individu-ally habituated to a black Plexiglas arena (40 cm � 40 cm � 28 cm),on three separate days, for 10 min daily. Habituation to the arenaoccurred across four consecutive days. All equipment was cleanedusing Quatriside�TB (Waterbury, CT) between animals and testsessions. All behavioral tests were digitally recorded and laterscored using a manual behavioral scoring software program(ODLog™ 2.0, http://www.macropodsoftware.com/) by observersblind to the group designations. On P40, animals were evaluatedon the social-interaction and the object-in-place recognition testsbetween 9:00 and 1:00 h. These tests were completed in a counter-balanced manner with at least one hour separating each assess-ment. Both of these behavioral experiments took place in the

E.J. Connors et al. / Brain, Behavior, and Immunity 42 (2014) 178–190 181
arena that animals had been habituated to between P36 and P39,no more than 48 h prior to behavioral assessment.
2.5.1. Affiliative behaviorIn the social interaction test, each animal was paired with an
unknown test partner (stimulus rat) that did not differ from themby more than 15 g in weight. All male and female stimulus rats(n = 8) were raised under ACC-saline conditions and were alter-nated between trials to ensure at least 60 min of rest between eachpairing. Each session was 10 min long and the total frequency andduration of the following categories were recorded:
(i) Initiation of social engagement: Allogrooming (experimentalrat grooms/sniffs any part of stimulus rats’ body), mount-ing/crawling (experimental rat stands on hind legs and putsone or two forepaws on back of conspecific or climbs overconspecific), approaching/following (experimental rat fol-lows stimulus rat), nipping (experimental rat bites conspe-cific’s nape or tail).
(ii) Recipient of social engagement: Allogrooming (any part of theexperimental rat’s body is being groomed/sniffed by stimu-lus rat), being mounted (stimulus rat stands on hind legsand puts one or two forepaws on back of experimental rator climbs over them), being approached/followed (stimulusrat is actively seeking experimental rat), being nipped(experimental rat is being bitten on nape or tail by stimulusrat).
The total percentage of time spent in social engagement wasalso calculated [((total time initiating + total time as recipient)/600 s)�100].
2.5.2. Cognitive discriminationIn the object-in-place recognition test, a rats’ memory for spa-
tial locations is inferred to be intact if the animal spends more timeexploring a set of objects that have switched location compared toobjects that have not moved (Dere et al., 2007; Howland et al.,2012). Previous work has shown that rats treated with a viralmimetic during gestation demonstrate disrupted behavior in thistask (Howland et al., 2012). During the sample phase of this recog-nition test, rats were given the opportunity to explore four differ-ent objects. Objects were placed in each of the four corners of thearena and rats were allowed to explore for 5 min. Following a 1-hdelay, identical copies of the original objects presented in the sam-ple phase were used, except two of the objects switched location(i.e., the front object on either the right or the left-side becamethe back object, while the back object was repositioned in thefront. The right/left side of alternation was randomized to controlfor side preferences). Animals were again given 5 min to explorethe objects. From this test phase, a novel discrimination ratio(DR) was calculated [(total time exploring moved objects � totaltime exploring permanent objects)/(total time exploring bothobjects)]. Total time exploring the objects in both the sample andtest phases was also recorded. Objects were made of plastic, glassand metal and placement locations were counterbalanced acrossgroups.
2.5.3. Hedonic statusIn the early evening of P40, an overnight 5% sucrose preference
test was used to assess hedonic status. Prior to the test, animalswere placed into individual standard cages with a tube and Nest-lets� (EE animals were given additional toys). Rats were giventwo bottles, each containing either a 5% sucrose solution or tapwater. Intake and preference were recorded for each animal overa 16 h period (17:00–9:00 h). Intake was defined as the totalamount of fluid consumed, whereas preference was obtained by
calculating the ratio of sucrose intake (grams) to total fluid intake(grams) and converted into a percent score. At no time were ani-mals deprived of food or water. A sucrose preference score below60% has been defined previously as an indicator of anhedonia(Duncko et al., 2003). Rats were maintained in their new housingconditions until the completion of the second 5% overnight sucrosetest on P42. The placement of the sucrose and water bottles wascounterbalanced between tests to prevent side preferences.
2.6. Brain collection and analysis
On the morning of P42, male and female offspring were deeplyanesthetized with a mixture of Ketamine/Xylazine (40–80 mg/kg,i.p./5–10 mg/kg, i.p.). Subsequently, rats were perfused intracardi-ally with a phosphate buffer solution. Brains were removed andimmediately frozen on dry ice and stored at �75 �C until process-ing. At that point, brains were placed in a blocker and cut into2 mm coronal sections. Using the Paxinos and Watson (1998) brainatlas as a guide the hippocampus, prefrontal cortex, and amygdalawere collected for further analysis via liquid chromatography massspectrometry (LCMS–MS) and Western blot techniques.
2.6.1. LCMS–MSA CTC Pal auto sampler (Leap Technologies, Carrboro, NC,USA)
was used to inject 15 lL aliquots of the processed samples on aKinetex XB� C18 column (3 � 100 mm, 2.6 lm, Phenomenex,Torrance, CA, USA). A binary gradient mobile phase consisting of(A) 0.1% formic acid in Milli Q water (Millipore, Milford, MA,USA) and (B) 100% acetonitrile (HPLC Grade, JT Baker), was usedto deliver a gradient flow into the mass spectrometer’selectrospray ionization chamber. Quantification was achieved byMS–MS detection in both positive and negative ion modes, usinga AB Sciex API-5500 mass spectrometer (Framingham, MA, USA),equipped with a Turboionspray™ interface at 500 �C. The ion sprayvoltage was set at 4500 V. The other parameters such as curtaingas, collision gas, ion source gas 1 GS1 and GS2 were set at 32,medium, 60 and 70, respectively. Detection of all the ions was per-formed in the multiple reaction monitoring (MRM) mode. Thereagents for glutamate and corticosterone were obtained fromSigma–Aldrich. The quadrupoles Q1 and Q3 were set on unit reso-lution and the analytical data was processed by Multiquant soft-ware (version 2.0). Primary stock solutions (20 mM) of theanalytes were prepared in 100% dimethyl sulfoxide (DMSO) byweighing each of them separately. Subsequent secondary stocksolutions (10 mM) were prepared in 100% methanol and all thestock solutions were stored at �80 �C. Secondary stock solutionswere used for the preparation of standards (CC) and quality control(QC) samples. The calibration range was 2.44–2500 nM with 12concentrations, each of the calibration curves were prepared usinga control brain and acceptance criteria for calibration was a regres-sion of greater than 0.99 (r > 0.99). The samples were prepared byweighing each brain structure and adding 3 parts (w/v) of homog-enizing/extraction buffer 0.1% formic acid in 50% acetonitrile. Thesamples were homogenized using Fastprep 24 (MP Biomedicals)and 100 lL of the aliquot was then transferred to a 96 well plate(Waters 186002481). Acetonitrile (200 lL) containing 10 ng/mL8-cyclopentyl-1,3-dipropyl xanthine (CPDPX) as internal standardwas added, vortexed for 2 min and centrifuged at 4000�g for10 min. 100 lL of the supernatant was transferred to 96 deep wellanalysis plate with 2 mL volume capacity (Analytical; 59623-23) towhich 100 lL of diluent was added containing a mixture of aceto-nitrile, MilliQ water, and formic acid in the following ratio80:20:0.1. Calibration curves were generated by plotting peak arearatios of glutamate (m/z = 148.1?84.2) and corticosterone(m/z = 347.1?121.0) against the nominal concentration of calibrationstandards (glutamate: 20–625 nM, corticosterone = 2.4–2000 nM).

182 E.J. Connors et al. / Brain, Behavior, and Immunity 42 (2014) 178–190
Subsequently, blinded samples were injected followed by the gen-eration of a calibration curve for each analyte. Appropriate analyt-ical quality control samples as well as blank samples weresequentially run.
2.6.2. Western blot proceduresHippocampal tissue was homogenized, proteins were extracted
and homogenates (40 lg protein/well) were separated by 7.5%SDS–PAGE and transferred onto a nitrocellulose membrane. Mem-branes were then blocked overnight in 10% fat-free milk with Tris-buffered saline containing Tween 20 (TBST) at 4 �C. The next day,membranes were incubated in a 1:1000 dilution of a mouse mono-clonal IgG glucocorticoid receptor (GR) antibody (MA1–510; Piercefrom Thermo Fisher Scientific Inc., Rockford, IL) for 1.5 h. The mem-branes were washed three times for 5 min in TBS-T and incubatedat room temperature for 1 h with goat anti-mouse IgG secondaryconjugated with horseradish peroxidase (1:2000; sc-2031; SantaCruz Biotechnology, Santa Cruz, CA). Following another set of 3washes for 5 min, the membranes were exposed to a mouse anti-beta Actin antibody IgG conjugated with horseradish peroxidase(1:20,000; ab49900; Abcam, Cambridge, MA) for 1 h. After threewashes for 5 min with TBS-T, a chemiluminescence substratewas applied to the membrane to reveal bound antibodies(37,574; 1-step Ultra TMB-Blotting Solution; Pierce from ThermoFisher Scientific Inc, Rockford, IL). Following band detection, themembrane was stripped with Restore Western Blot Stripping Buf-fer (21,059; Pierce from Thermo Fisher Scientific Inc., Rockford, IL)and blocked overnight in 10% fat-free milk with Tris-buffered sal-ine containing Tween 20 (TBST) at 4 �C. The next morning, themembrane was reblotted with a mouse monoclonal IgG NMDAReceptor 2B (NR2B) antibody (MA1–2014; Pierce from ThermoFisher Scientific Inc, Rockford, IL) at a 1:1000 dilution for 1.5 h atroom temperature, washed three times for 5 min in TBS-T, andincubated at room temperature for 1 h with goat anti-mouse IgGsecondary conjugated with horseradish peroxidase (1:2000; sc-2031; Santa Cruz Biotechnology, Santa Cruz, CA). Please note thatprefrontal cortex tissue was treated similarly but only evaluatedfor NR2B. To obtain a semi-quantitative analysis of protein expres-sion we quantified proteins by densitometry to obtain GR/actinand NR2B/actin ratios.
2.7. Data analysis
All analyses were conducted using SPSS version 21.0 software.To confirm MIA, non-parametric Mann–Whitney U tests were usedto evaluate LPS and housing effects on composite sickness behaviorscores. Difference scores were calculated between maternal bodyweights on G11 and G12, following LPS treatment. For this variable,in addition to maternal behavior comparisons, a mixed analysis ofvariance (ANOVA) design was utilized. With respect to the ANOVAthere were two independent factors: ‘gestational treatment’ withtwo levels (saline and LPS) and ‘housing condition’ with two levels(ACC and EE).
Male and female juvenile offspring body weights and sucrosepreference were assessed using a three-factor repeated measuresANOVA with ‘time’ (body weight: P22, P29, P36, P40; sucrose:P40, P41) as the repeated factor and ‘gestational treatment’ (saline,LPS), ‘sex’ (male, female), and ‘housing condition’ (ACC, EE) as thebetween subject factors. All other treatment group comparisonswere made using three-way ANOVAS with ‘gestational treatment’,‘sex’, and ‘housing condition’ as the independent factors. Oneexception to this was for analysis of the object-in-place recognitiontest DR. In this case, DRs for each group were evaluated using onesample t-tests employing ‘0’ as a comparison value to indicateequal, or chance, performance in exploring the two objects (seeHowland et al., 2012). Alpha levels were set at a probability of
0.05 and the Greenhouse–Geisser correction procedure wasapplied for violations to the assumption of sphericity for repeatedmeasures designs (Howell, 2002), where appropriate. Significantmain effects were further assessed via pairwise t-tests andLevene’s test was applied in the occurrence of unequal varianceson the posthoc assessments. For all food and water intake and pref-erence data, body weight was used as a covariate. Cohen’s kappawas employed in order to measure inter-rater agreement betweentwo observers for each categorical variable and Chronbach’s alphawas calculated for each continuous measure. Individual Cohen’skappa data is provided while inter-rater reliability was found tobe >90% for each continuous measure described.
3. Results
3.1. Gestational LPS treatment
In order to verify that gestational LPS produced an inflamma-tory-associated sickness response, dams were evaluated on sick-ness behaviors, food and water intake, and body weight changes.Mann–Whitney U tests did not reveal an effect of housing condi-tion (ACC vs EE) across the observation period (p > 0.05) for totalsickness behavior. However there was a significant effect of gesta-tional treatment at 105 (z = �3.945; p = 0.0001), 120 (z = �2.537;p = 0.011) and 210 (z = �2.916; p = 0.004) min post treatment inthat LPS induced higher sickness scores, across housing conditionand time, compared to saline treated animals (Fig. 2A). The kappacoefficient for the total sickness behavior score was j = 0.699,T = 16.926; p = 0.0001, indicating a substantial level of inter-raterreliability.
No significant effects were observed in body weight change fol-lowing the G11 LPS challenge (p > 0.05). Using body weight on G11as a covariate, the overall two-way ANOVA for food(F(1,30) = 12.634; p = 0.001, g2 = 0.280; Fig. 2B) and water(F(1,30) = 4.450; p = 0.043, g2 = 0.129; Fig. 2C) intake revealed asignificant effect of gestational treatment, 20 h post injection. Pos-thoc tests showed a significant difference within the ACC and EEgroups in that LPS treated animals had a larger reduction in foodintake compared to saline treated dams (ACC; t(17) = �2.174;p = 0.044 and EE; t(14) = �2.713; p = 0.017), confirming the anorec-tic sickness effects of LPS treatment (Fig. 2B). Following the injec-tion procedure, three ACC dams treated with LPS, and one treatedwith saline lost their litters. Therefore, there was a total of 26 lit-ters (ACC-Saline = 7, ACC-LPS = 5, EE-Saline = 7, EE-LPS = 7) for allprocedures, run across two replicates of the study.
3.2. Litter health and behavior
There were no significant differences between the gestationaldrug treatment or housing condition on the length of gestation,the number of dams that successfully gave birth, the number ofpups born per litter, the sex-ratio composition of the litter, pupsurvival, or maternal body weight from P2 to P22 (p > 0.05). Therewere no differences between maternal housing or gestationaltreatment condition in terms of the frequency of licking/grooming,pup retrieval, or the number of pups maintained in the nests(p > 0.05). In the case of the EE dams, pairs would generally alter-nate between shared and individual nests across the postpartumperiod. During maternal behavior observation times when EE damsmaintained their own nests, there were no treatment differences(i.e., saline, LPS) in the distribution of pups within the nest(p > 0.05). Moreover, all EE mothers maintained visible teatsthroughout the study, evidence of continued nursing. Overall, EEdams cared for each other’s pups, regardless of their gestationaltreatment condition. Notably, ACC dams spent more time on the
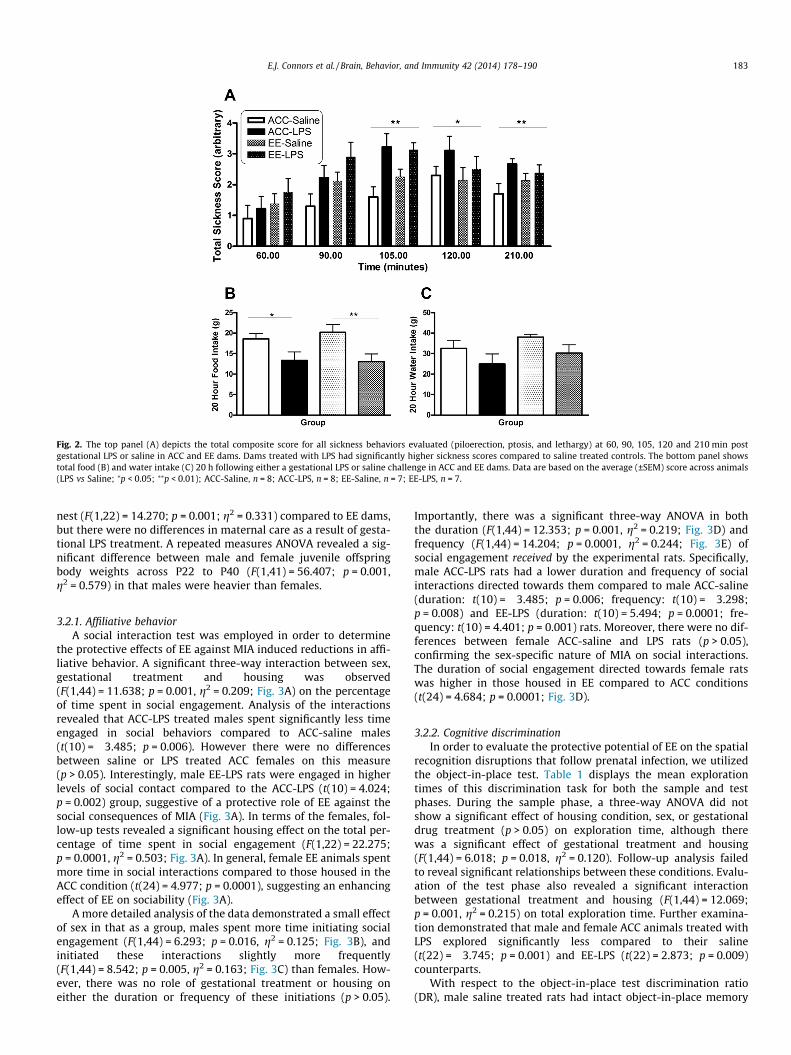
Fig. 2. The top panel (A) depicts the total composite score for all sickness behaviors evaluated (piloerection, ptosis, and lethargy) at 60, 90, 105, 120 and 210 min postgestational LPS or saline in ACC and EE dams. Dams treated with LPS had significantly higher sickness scores compared to saline treated controls. The bottom panel showstotal food (B) and water intake (C) 20 h following either a gestational LPS or saline challenge in ACC and EE dams. Data are based on the average (±SEM) score across animals(LPS vs Saline; ⁄p < 0.05; ⁄⁄p < 0.01); ACC-Saline, n = 8; ACC-LPS, n = 8; EE-Saline, n = 7; EE-LPS, n = 7.
E.J. Connors et al. / Brain, Behavior, and Immunity 42 (2014) 178–190 183
nest (F(1,22) = 14.270; p = 0.001; g2 = 0.331) compared to EE dams,but there were no differences in maternal care as a result of gesta-tional LPS treatment. A repeated measures ANOVA revealed a sig-nificant difference between male and female juvenile offspringbody weights across P22 to P40 (F(1,41) = 56.407; p = 0.001,g2 = 0.579) in that males were heavier than females.
3.2.1. Affiliative behaviorA social interaction test was employed in order to determine
the protective effects of EE against MIA induced reductions in affi-liative behavior. A significant three-way interaction between sex,gestational treatment and housing was observed(F(1,44) = 11.638; p = 0.001, g2 = 0.209; Fig. 3A) on the percentageof time spent in social engagement. Analysis of the interactionsrevealed that ACC-LPS treated males spent significantly less timeengaged in social behaviors compared to ACC-saline males(t(10) = �3.485; p = 0.006). However there were no differencesbetween saline or LPS treated ACC females on this measure(p > 0.05). Interestingly, male EE-LPS rats were engaged in higherlevels of social contact compared to the ACC-LPS (t(10) = 4.024;p = 0.002) group, suggestive of a protective role of EE against thesocial consequences of MIA (Fig. 3A). In terms of the females, fol-low-up tests revealed a significant housing effect on the total per-centage of time spent in social engagement (F(1,22) = 22.275;p = 0.0001, g2 = 0.503; Fig. 3A). In general, female EE animals spentmore time in social interactions compared to those housed in theACC condition (t(24) = 4.977; p = 0.0001), suggesting an enhancingeffect of EE on sociability (Fig. 3A).
A more detailed analysis of the data demonstrated a small effectof sex in that as a group, males spent more time initiating socialengagement (F(1,44) = 6.293; p = 0.016, g2 = 0.125; Fig. 3B), andinitiated these interactions slightly more frequently(F(1,44) = 8.542; p = 0.005, g2 = 0.163; Fig. 3C) than females. How-ever, there was no role of gestational treatment or housing oneither the duration or frequency of these initiations (p > 0.05).
Importantly, there was a significant three-way ANOVA in boththe duration (F(1,44) = 12.353; p = 0.001, g2 = 0.219; Fig. 3D) andfrequency (F(1,44) = 14.204; p = 0.0001, g2 = 0.244; Fig. 3E) ofsocial engagement received by the experimental rats. Specifically,male ACC-LPS rats had a lower duration and frequency of socialinteractions directed towards them compared to male ACC-saline(duration: t(10) = �3.485; p = 0.006; frequency: t(10) = �3.298;p = 0.008) and EE-LPS (duration: t(10) = 5.494; p = 0.0001; fre-quency: t(10) = 4.401; p = 0.001) rats. Moreover, there were no dif-ferences between female ACC-saline and LPS rats (p > 0.05),confirming the sex-specific nature of MIA on social interactions.The duration of social engagement directed towards female ratswas higher in those housed in EE compared to ACC conditions(t(24) = 4.684; p = 0.0001; Fig. 3D).
3.2.2. Cognitive discriminationIn order to evaluate the protective potential of EE on the spatial
recognition disruptions that follow prenatal infection, we utilizedthe object-in-place test. Table 1 displays the mean explorationtimes of this discrimination task for both the sample and testphases. During the sample phase, a three-way ANOVA did notshow a significant effect of housing condition, sex, or gestationaldrug treatment (p > 0.05) on exploration time, although therewas a significant effect of gestational treatment and housing(F(1,44) = 6.018; p = 0.018, g2 = 0.120). Follow-up analysis failedto reveal significant relationships between these conditions. Evalu-ation of the test phase also revealed a significant interactionbetween gestational treatment and housing (F(1,44) = 12.069;p = 0.001, g2 = 0.215) on total exploration time. Further examina-tion demonstrated that male and female ACC animals treated withLPS explored significantly less compared to their saline(t(22) = �3.745; p = 0.001) and EE-LPS (t(22) = 2.873; p = 0.009)counterparts.
With respect to the object-in-place test discrimination ratio(DR), male saline treated rats had intact object-in-place memory
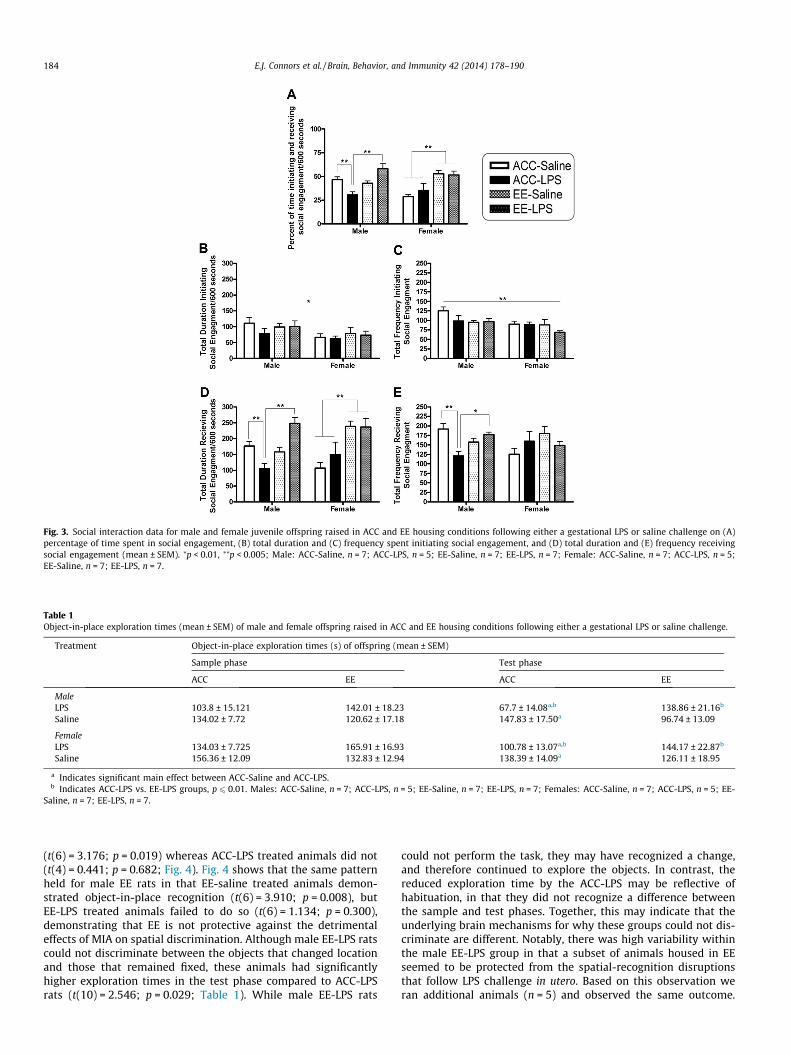
Fig. 3. Social interaction data for male and female juvenile offspring raised in ACC and EE housing conditions following either a gestational LPS or saline challenge on (A)percentage of time spent in social engagement, (B) total duration and (C) frequency spent initiating social engagement, and (D) total duration and (E) frequency receivingsocial engagement (mean ± SEM). ⁄p < 0.01, ⁄⁄p < 0.005; Male: ACC-Saline, n = 7; ACC-LPS, n = 5; EE-Saline, n = 7; EE-LPS, n = 7; Female: ACC-Saline, n = 7; ACC-LPS, n = 5;EE-Saline, n = 7; EE-LPS, n = 7.
Table 1Object-in-place exploration times (mean ± SEM) of male and female offspring raised in ACC and EE housing conditions following either a gestational LPS or saline challenge.
Treatment Object-in-place exploration times (s) of offspring (mean ± SEM)
Sample phase Test phase
ACC EE ACC EE
MaleLPS 103.8 ± 15.121 142.01 ± 18.23 67.7 ± 14.08a,b 138.86 ± 21.16b
Saline 134.02 ± 7.72 120.62 ± 17.18 147.83 ± 17.50a 96.74 ± 13.09
FemaleLPS 134.03 ± 7.725 165.91 ± 16.93 100.78 ± 13.07a,b 144.17 ± 22.87b
Saline 156.36 ± 12.09 132.83 ± 12.94 138.39 ± 14.09a 126.11 ± 18.95
a Indicates significant main effect between ACC-Saline and ACC-LPS.b Indicates ACC-LPS vs. EE-LPS groups, p 6 0.01. Males: ACC-Saline, n = 7; ACC-LPS, n = 5; EE-Saline, n = 7; EE-LPS, n = 7; Females: ACC-Saline, n = 7; ACC-LPS, n = 5; EE-
Saline, n = 7; EE-LPS, n = 7.
184 E.J. Connors et al. / Brain, Behavior, and Immunity 42 (2014) 178–190
(t(6) = 3.176; p = 0.019) whereas ACC-LPS treated animals did not(t(4) = 0.441; p = 0.682; Fig. 4). Fig. 4 shows that the same patternheld for male EE rats in that EE-saline treated animals demon-strated object-in-place recognition (t(6) = 3.910; p = 0.008), butEE-LPS treated animals failed to do so (t(6) = 1.134; p = 0.300),demonstrating that EE is not protective against the detrimentaleffects of MIA on spatial discrimination. Although male EE-LPS ratscould not discriminate between the objects that changed locationand those that remained fixed, these animals had significantlyhigher exploration times in the test phase compared to ACC-LPSrats (t(10) = 2.546; p = 0.029; Table 1). While male EE-LPS rats
could not perform the task, they may have recognized a change,and therefore continued to explore the objects. In contrast, thereduced exploration time by the ACC-LPS may be reflective ofhabituation, in that they did not recognize a difference betweenthe sample and test phases. Together, this may indicate that theunderlying brain mechanisms for why these groups could not dis-criminate are different. Notably, there was high variability withinthe male EE-LPS group in that a subset of animals housed in EEseemed to be protected from the spatial-recognition disruptionsthat follow LPS challenge in utero. Based on this observation weran additional animals (n = 5) and observed the same outcome.
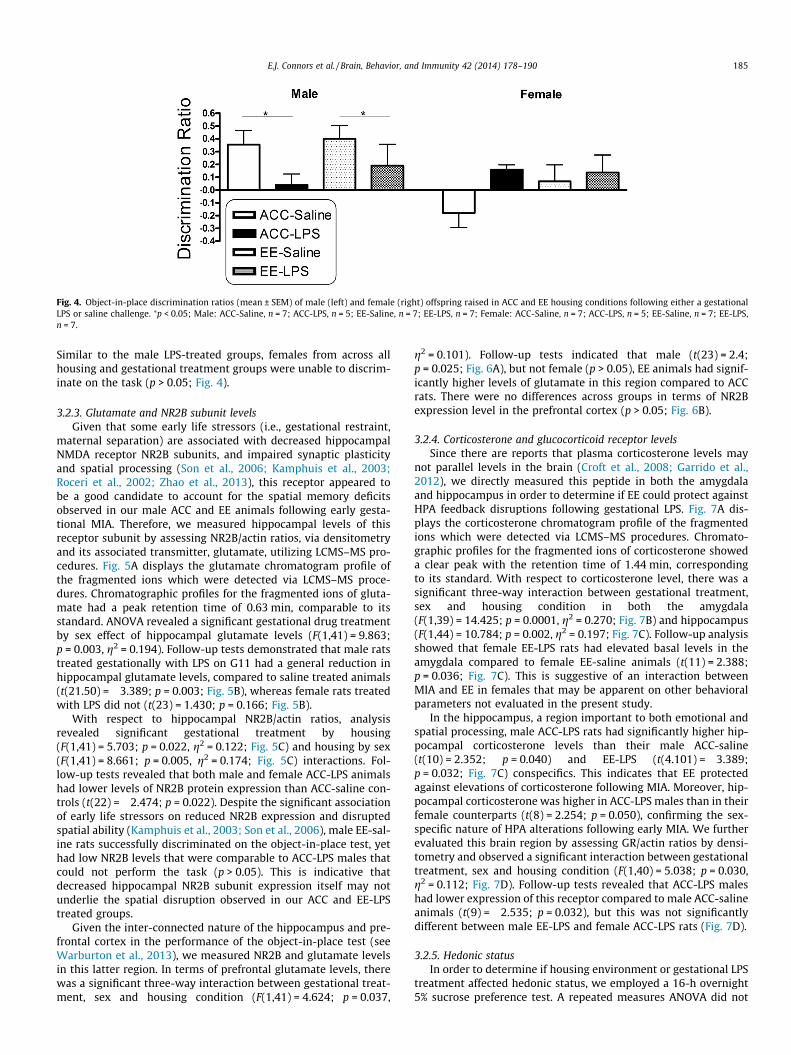
Fig. 4. Object-in-place discrimination ratios (mean ± SEM) of male (left) and female (right) offspring raised in ACC and EE housing conditions following either a gestationalLPS or saline challenge. ⁄p < 0.05; Male: ACC-Saline, n = 7; ACC-LPS, n = 5; EE-Saline, n = 7; EE-LPS, n = 7; Female: ACC-Saline, n = 7; ACC-LPS, n = 5; EE-Saline, n = 7; EE-LPS,n = 7.
E.J. Connors et al. / Brain, Behavior, and Immunity 42 (2014) 178–190 185
Similar to the male LPS-treated groups, females from across allhousing and gestational treatment groups were unable to discrim-inate on the task (p > 0.05; Fig. 4).
3.2.3. Glutamate and NR2B subunit levelsGiven that some early life stressors (i.e., gestational restraint,
maternal separation) are associated with decreased hippocampalNMDA receptor NR2B subunits, and impaired synaptic plasticityand spatial processing (Son et al., 2006; Kamphuis et al., 2003;Roceri et al., 2002; Zhao et al., 2013), this receptor appeared tobe a good candidate to account for the spatial memory deficitsobserved in our male ACC and EE animals following early gesta-tional MIA. Therefore, we measured hippocampal levels of thisreceptor subunit by assessing NR2B/actin ratios, via densitometryand its associated transmitter, glutamate, utilizing LCMS–MS pro-cedures. Fig. 5A displays the glutamate chromatogram profile ofthe fragmented ions which were detected via LCMS–MS proce-dures. Chromatographic profiles for the fragmented ions of gluta-mate had a peak retention time of 0.63 min, comparable to itsstandard. ANOVA revealed a significant gestational drug treatmentby sex effect of hippocampal glutamate levels (F(1,41) = 9.863;p = 0.003, g2 = 0.194). Follow-up tests demonstrated that male ratstreated gestationally with LPS on G11 had a general reduction inhippocampal glutamate levels, compared to saline treated animals(t(21.50) = �3.389; p = 0.003; Fig. 5B), whereas female rats treatedwith LPS did not (t(23) = 1.430; p = 0.166; Fig. 5B).
With respect to hippocampal NR2B/actin ratios, analysisrevealed significant gestational treatment by housing(F(1,41) = 5.703; p = 0.022, g2 = 0.122; Fig. 5C) and housing by sex(F(1,41) = 8.661; p = 0.005, g2 = 0.174; Fig. 5C) interactions. Fol-low-up tests revealed that both male and female ACC-LPS animalshad lower levels of NR2B protein expression than ACC-saline con-trols (t(22) = �2.474; p = 0.022). Despite the significant associationof early life stressors on reduced NR2B expression and disruptedspatial ability (Kamphuis et al., 2003; Son et al., 2006), male EE-sal-ine rats successfully discriminated on the object-in-place test, yethad low NR2B levels that were comparable to ACC-LPS males thatcould not perform the task (p > 0.05). This is indicative thatdecreased hippocampal NR2B subunit expression itself may notunderlie the spatial disruption observed in our ACC and EE-LPStreated groups.
Given the inter-connected nature of the hippocampus and pre-frontal cortex in the performance of the object-in-place test (seeWarburton et al., 2013), we measured NR2B and glutamate levelsin this latter region. In terms of prefrontal glutamate levels, therewas a significant three-way interaction between gestational treat-ment, sex and housing condition (F(1,41) = 4.624; p = 0.037,
g2 = 0.101). Follow-up tests indicated that male (t(23) = 2.4;p = 0.025; Fig. 6A), but not female (p > 0.05), EE animals had signif-icantly higher levels of glutamate in this region compared to ACCrats. There were no differences across groups in terms of NR2Bexpression level in the prefrontal cortex (p > 0.05; Fig. 6B).
3.2.4. Corticosterone and glucocorticoid receptor levelsSince there are reports that plasma corticosterone levels may
not parallel levels in the brain (Croft et al., 2008; Garrido et al.,2012), we directly measured this peptide in both the amygdalaand hippocampus in order to determine if EE could protect againstHPA feedback disruptions following gestational LPS. Fig. 7A dis-plays the corticosterone chromatogram profile of the fragmentedions which were detected via LCMS–MS procedures. Chromato-graphic profiles for the fragmented ions of corticosterone showeda clear peak with the retention time of 1.44 min, correspondingto its standard. With respect to corticosterone level, there was asignificant three-way interaction between gestational treatment,sex and housing condition in both the amygdala(F(1,39) = 14.425; p = 0.0001, g2 = 0.270; Fig. 7B) and hippocampus(F(1,44) = 10.784; p = 0.002, g2 = 0.197; Fig. 7C). Follow-up analysisshowed that female EE-LPS rats had elevated basal levels in theamygdala compared to female EE-saline animals (t(11) = 2.388;p = 0.036; Fig. 7C). This is suggestive of an interaction betweenMIA and EE in females that may be apparent on other behavioralparameters not evaluated in the present study.
In the hippocampus, a region important to both emotional andspatial processing, male ACC-LPS rats had significantly higher hip-pocampal corticosterone levels than their male ACC-saline(t(10) = 2.352; p = 0.040) and EE-LPS (t(4.101) = �3.389;p = 0.032; Fig. 7C) conspecifics. This indicates that EE protectedagainst elevations of corticosterone following MIA. Moreover, hip-pocampal corticosterone was higher in ACC-LPS males than in theirfemale counterparts (t(8) = 2.254; p = 0.050), confirming the sex-specific nature of HPA alterations following early MIA. We furtherevaluated this brain region by assessing GR/actin ratios by densi-tometry and observed a significant interaction between gestationaltreatment, sex and housing condition (F(1,40) = 5.038; p = 0.030,g2 = 0.112; Fig. 7D). Follow-up tests revealed that ACC-LPS maleshad lower expression of this receptor compared to male ACC-salineanimals (t(9) = �2.535; p = 0.032), but this was not significantlydifferent between male EE-LPS and female ACC-LPS rats (Fig. 7D).
3.2.5. Hedonic statusIn order to determine if housing environment or gestational LPS
treatment affected hedonic status, we employed a 16-h overnight5% sucrose preference test. A repeated measures ANOVA did not

Fig. 5. Markers of hippocampal glutamatergic functioning in juvenile offspring raised in ACC and EE housing following either an early gestational LPS or saline challenge. (A)Standard LCMS–MS ion chromatogram of glutamate, male (left) and female (right), (B) glutamate levels expressed as ng/mg, and (C) densitometric data for NR2B/actin ratiosare displayed. Data expressed as mean ± SEM; ⁄p < 0.05, ⁄⁄p < 0.01; bp < 0.05 between ACC-Saline and ACC-LPS; Male: ACC-Saline, n = 7; ACC-LPS, n = 5; EE-Saline, n = 7; EE-LPS, n = 7; Female: ACC-Saline, n = 7; ACC-LPS, n = 5; EE-Saline, n = 6–7; EE-LPS, n = 7.
186 E.J. Connors et al. / Brain, Behavior, and Immunity 42 (2014) 178–190
reveal hedonic differences in either male or female animals of anygroup (p > 0.05). All groups had an immediate high preference forsucrose, significantly above the criterion of 60% or below for anhe-donia (see Duncko et al., 2003).
4. Discussion
In the present work, we demonstrate that EE protects young off-spring against the sex-specific hypothalamic–pituitary–adrenalaxis (HPA) and social interaction disruptions that follow earlymaternal immune activation (MIA). Specifically, EE preventedagainst elevations in hippocampal corticosterone level, decreasedGR expression, and reductions in social contact in juvenile malerats treated with LPS in utero. Thus, these preliminary findingsdemonstrate that EE mitigates against the detrimental effects ofMIA. Since gestational LPS induced significant sickness, regardlessof housing condition, it appears that the protection offered by EEwas not due to a direct prevention of inflammation in the dam.Whether the success of EE as a behavioral intervention againstthe social consequences of MIA was due to a reduction in stresslevel, or some other factor of complex housing, is unknown.
Impairments in spatial learning are common in animal modelsof prenatal infection (Golan et al., 2005; Howland et al., 2012;Lanté et al., 2007), and are thought to be mediated in part throughthe inhibition of long-term potentiation (Escobar et al., 2011) and areduction in the contribution of NMDA receptors to basal glutama-tergic synaptic transmission (Lanté et al., 2007). As expected, our
ACC-LPS males demonstrated spatial impairments in an object-in-place spatial task and a reduction of glutamate levels andNR2B expression in the hippocampus. Despite the success of envi-ronmental enhancement protocols in promoting cognitive recov-ery (Dahlqvist et al., 2004; Jurgens and Johnson, 2012; Ohlssonand Johansson, 1995) EE was unable to protect against the spatialdeficits imposed by MIA. This suggests that some of the effects ofMIA are not dependent on the typical environment of the labora-tory rat, but that they may extend to species living in more com-plex and varied conditions. However, we do report individualdifferences in that a subset of EE animals were protected and themechanisms underlying this resilience is an opportunity for futurework.
Given the coordinated role of the hippocampus and prefrontalcortex in the object-in-place task, we also evaluated NR2B in thislatter region and found no difference in protein expression associ-ated with housing, sex, or MIA status. Interestingly, in an animalmodel of methamphetamine addiction, object-in-place recognitionwas disrupted without any changes in NR2B expression in eitherthe prefrontal cortex or hippocampus (Reichel et al., 2014).Although there was a reported decrease of NR2B in the perirhinalcortex, modafinil restoration of discrimination performance wasnot accompanied by a reversal of NR2B in this region (Reichelet al., 2014). Therefore, NR2B protein expression alone may notbe an adequate biomarker for the novel discrimination deficits inthese models. Overall, the diverse mechanisms underlying spatialimpairments following MIA appear to differ as a function of expe-

Fig. 6. Markers of prefrontal cortex glutamatergic functioning in juvenile offspring raised in ACC and EE housing following either an early gestational LPS or saline challenge.Male (right) and female (left) (A) glutamate levels expressed as ng/mg, and (B) densitometric data for NR2B/actin ratios are displayed. Data expressed as mean ± SEM; ⁄⁄p < .01between ACC and EE animals; Male: ACC-Saline, n = 7; ACC-LPS, n = 5; EE-Saline, n = 6; EE-LPS, n = 7; Female: ACC-Saline, n = 7; ACC-LPS, n = 5; EE-Saline, n = 7; EE-LPS, n = 6.
E.J. Connors et al. / Brain, Behavior, and Immunity 42 (2014) 178–190 187
rience. For example, elevated hippocampal corticosterone com-bined with, or independent of, changes in glutamate and NR2B lev-els may be related to spatial disruptions in ACC-LPS males;although EE prevented most of these neurophysiological conse-quences of MIA, complex rearing did not protect against a reduc-tion in hippocampal glutamate which may in part account forthe sustained learning impairment in this group. Disruptions inthe trafficking and assembly of NR2B (Kamphuis et al., 2003;Zhao et al., 2013), including synaptic NR2A/NR2B levels as medi-ated by kinesin superfamily motor protein 17 (Zhao et al., 2013),or changes in Ca2+/calmodulin-dependent protein kinase II (aCAM-KII) phosphorylation (Giese et al., 1998; Kamphuis et al., 2003) fol-lowing MIA should also be considered. With respect to lowerNR2B/actin ratios in the hippocampus of EE-saline rats, comparedto ACC-saline controls, it seems that our EE housing environmentmodifies the glutamatergic system differently between these twogroups. For example, initial findings with this model suggest thatEE-saline males had significantly higher glutamate levels in theprefrontal cortex, which may partially underlie their spatial dis-crimination competency, despite lower hippocampal NR2B expres-sion. Notably, it was expected that the EE experience would lead toelevated NR2B levels (Bredy et al., 2004), but most studies placeanimals in EE post weaning; perhaps novelty at this developmentalstage results in higher NR2B expression compared to our combinedpre- and post-weaning EE schedule. Moreover, different EE proto-cols affect region-specific protein changes (Lambert et al., 2005)and it is important to note that EE exposure during adolescencevs. adulthood has been reported to induce opposite organizationaleffects in some brain areas (see Mychasiuk et al., 2014).
Further evidence of cognitive differences between treatmentconditions can be found in the behavioral strategies that animalsused within the object-in-place task. Our preliminary data suggestthat EE-LPS rats had disruptions in retrieval while ACC-LPS rats haddifficulty with encoding and/or attention. For example, EE maleshabituate quickly to novelty (Zimmermann et al., 2001), whichwe observed in our EE-saline treated group. However EE-LPS rats
explored significantly longer in the test, but not sample, phase ofthe object-in-place test. This suggests that they recognized some-thing had changed, between phases, but could not identify whatit was. In contrast, lowered exploration times in the test phaseindicate that ACC-LPS males either did not correctly encode infor-mation during the sample phase, or they simply did not notice adifference. The latter may specifically relate to an issue of selectiveattention which accompanies early-life models of hypoglutamater-gic functioning, which are not reversible by memory promotingagents such as tacrine (Terranova et al., 2005).
Importantly, our results show that EE can rescue the brain fromthe social deficits that follow MIA. In line with other inflammatorymodels (Kirsten et al., 2010; Taylor et al., 2012), our male ACC-LPSrats had a reduction in the percentage of time they spent in socialcontact. However, these animals actively engaged in affiliativebehaviors at a level similar to other groups, although these interac-tions did not seem to be reciprocated, an effect prevented by EE.Since the quantity and quality of social interaction is dependenton how two animals engage and respond to each other (Pelliset al., 1997; Taylor et al., 2012), our choice of using mixed-treat-ment (i.e., ACC-LPS/ACC-saline, EE-saline/ACC-saline) as opposedto matched (i.e., ACC-LPS/ACC-LPS, EE-saline/EE-saline) pairs mayhave masked additional effects of LPS.
Moreover, there are important sensory, motor and motivationalcomponents that underlie social interaction (including play) (seePellis et al., 1997), and it is possible that gestational LPS treatmentdirectly affects one or more of these variables, ultimately affectingsocial interaction. Specifically, auditory cues may have influencedstimulus rats, resulting in stalled initiation or continuation of socialcontact with ACC-LPS males. For example, early MIA with eitherLPS or polyI:C reduced separation-induced ultrasonic vocalizationsin neonatal offspring (Baharnoori et al., 2012; Kirsten et al., 2012;Malkova et al., 2012), suggesting that the auditory cues from theseanimals may not appropriately signal prompts that stimulate inter-action. EE housing may have ameliorated this effect in our LPS-treated males; in one model of anxiety, rats emitted significantly

Fig. 7. Glucocorticoid functioning in juvenile offspring raised in ACC and EE housing following either an early gestational LPS or saline challenge. (A) standard LCMS–MS ionchromatogram of corticosterone, and male (left) and female (right), (B) amygdala and (C) hippocampal corticosterone levels expressed as ng/mg. (D) Male (left) female (right)densitometric data for hippocampal GR/actin ratios are also displayed. Data expressed as mean ± SEM; ⁄p < .05; Male: ACC-Saline, n = 6–7; ACC-LPS, n = 5; EE-Saline, n = 6–7;EE-LPS, n = 6–7; Female: ACC-Saline, n = 6–7; ACC-LPS, n = 5; EE-Saline, n = 6–7; EE-LPS, n = 6–7.
188 E.J. Connors et al. / Brain, Behavior, and Immunity 42 (2014) 178–190
lower levels of ‘positive’ 50-kHz vocalizations which recovered fol-lowing chronic exposure to EE (Perez-Sepulveda et al., 2013). Nota-bly, animals maintained in EE have been reported as less anxious,compared to standard housing conditions (Baldini et al., 2013;Ravenelle et al., 2013), which may explain the maintained socialcompetency of these animals following MIA. Overall, it is possiblethat anxiety underlies the reduced social interaction directedtowards the ACC-LPS males, whether mediated via vocalizations,or other motivational cues.
In terms of female animals housed in EE, there have been incon-sistencies on both the behavioral and neurochemical/hormonaloutcomes reported (see Girbovan and Plamondon, 2013) and thebenefits of this condition to females is underexplored. Like others,our females were unable to complete the object-in-place test(Howland et al., 2012) and we had hypothesized that the EE expe-rience would improve spatial discrimination on this task. We weresurprised to find this was not the case; combined with our findings,recent work suggests that improvement may be mediated through
hormones and parental experience (Cost et al., 2014), as opposedto spatially enhanced EE conditions. Importantly, our female ratswere more responsive to enrichment on measures of social engage-ment, suggesting that this housing condition increases the socialvalue of females housed in EE. Future work will need to evaluatethe role of specific elements of enrichment (i.e., running wheels,social enrichment) and the potential benefits of this housing condi-tion to females.
A potential limitation of our model is the daily coloring manip-ulation used to identify pup treatment groups. This effectivelyintroduced a neonatal handling procedure, to all animals, whichmay have raised basal corticosterone and GR levels (Wiener andLevine, 1978; Meaney et al., 1992, 1989), in addition to NR2B(Liu et al., 2000). Although corticosterone was only elevated inLPS treated animals and GR was diminished in ACC-LPS males, itis possible that we did not observe higher GR in EE animals asexpected (Baldini et al., 2013) because early handling may havemasked this effect. Alternatively, the small sample size of the data

E.J. Connors et al. / Brain, Behavior, and Immunity 42 (2014) 178–190 189
set could be a factor and may underlie some of our findings. Impor-tantly, EE was required to prevent changes in markers of HPA axisfunctioning, suggestive that handling alone is not sufficient to pro-tect against MIA. Maternal care, specifically frequency of lickingand grooming, is also known to affect GR expression and laterbehavior of offspring (Zhang et al., 2013). However, licking andgrooming patterns were comparable across all housing and gesta-tional treatment conditions. Notably, in the present study, the pro-tective effects of EE against MIA were only evaluated in younganimals; it will be interesting to determine whether these benefitspersist into adulthood.
5. Conclusion
Altogether, our initial findings demonstrate that the complexnature of EE conferred protection against MIA-associated disrup-tions in HPA axis functioning, and reductions in social contact,but not diminished hippocampal glutamate or spatial impair-ments. The success of our early life manipulation in rescuing thebrain from some consequences of MIA, but not others, suggeststhat the specific elements of EE that promote resilience againstdevelopmental adversity must be further delineated. This is rele-vant when considering the utility of environmental enhancementprotocols as an intervention against the behavioral and physiolog-ical consequences of MIA.
Author contributions
E.J.C., A.N.S., M.M.M. & A.C.K. ran the experiments and analyzeddata. A.C.K. designed the study and wrote the manuscript.
Disclosures and potential conflict of interests
Mr. A.N. Shaik worked as an intern in the Department of DrugMetabolism & Pharmacokinetics at Biogen Idec.
Acknowledgments
This work was funded by an MCPHS Summer UndergraduateResearch Fellowship awarded to (E.J.C.) and institutional supportprovided by MCPHS University. The authors would like to thankDr. Timothy Maher from the MCPHS School of Pharmacy for kindlyallowing us the use of his behavioral equipment. We would alsolike to extend our thanks to the Department of Drug Metabolism& Pharmacokinetics at Biogen Idec for donating time on theirLCMS–MS unit. In addition we thank Ms. Shelby Pillsbury andMs. Molly MacRae for their assistance in scoring blinded behavioralvideos.
References
Baharnoori, M., Bhardwaj, S.K., Srivastava, L.K., 2012. Neonatal behavioral changesin rats with gestational exposure to lipopolysaccharide: a prenatal infectionmodel for developmental neuropsychiatric disorders. Schizophr. Bull. 38, 444–456.
Baldini, S., Restani, L., Baroncelli, L., Coltelli, M., Franco, R., Cenni, M.C., Maffei, L.,Berardi, N., 2013. Enriched early life experiences reduce adult anxiety-likebehavior in rats: a role for insulin-like growth factor 1. J. Neurosci. 33 (28),11715–11723.
Bitanihirwe, B.K., Peleg-Raibstein, D., Mouttet, F., Feldon, J., Meyer, U., 2010. Lateprenatal immune activation in mice leads to behavioral and neurochemicalabnormalities relevant to the negative symptoms of schizophrenia.Neuropsychopharmacology 35, 2462–2478.
Blatteis, C.M., Li, S., Li, Z., Perlik, V., Feleder, C., 2004. Signaling the brain in systemicinflammation: the role of complement. Front. Biosci. 9, 915–931.
Bredy, T.W., Zhang, T.Y., Grant, R.J., Diorio, J., Meaney, M.J., 2004. Peripubertalenvironmental enrichment reverses the effects of maternal care onhippocampal development and glutamate receptor subunit expression. Eur. J.Neurosci. 20, 1355–1362.
Brown, A.S., 2012. Epidemiologic studies of exposure to prenatal infection and riskof schizophrenia and autism. Dev. Neurobiol. 72, 1272–1276.
Brown, A.S., Derkits, E.J., 2010. Prenatal infection and schizophrenia: a review ofepidemiologic and translational studies. Am. J. Psychiatry 167, 261–280.
Cardinale, J.P., Sriramula, S., Pariaut, R., Guggilam, A., Mariappan, N., Elks, C.M.,Francis, J., 2010. HDAC Inhibition attenuates inflammatory, hypertrophic andhypertensive responses in spontaneously hypertensive rats. Hypertension 56,437–444.
Cost, K.T., Lobell, T.D., Williams-Yee, Z.N., Henderson, S., Dohanich, G., 2014. Theeffects of pregnancy, lactation, and primiparity on object-in-place memory offemale rats. Horm. Behav. 65, 32–39.
Coyle, J.T., Tsai, G., Goff, D., 2003. Converging evidence of NMDA receptorhypofunction in the pathophysiology of schizophrenia. Ann. N.Y. Acad. Sci.1003, 318–327.
Coyle, P., Tran, N., Fung, J.N., Summers, B.L., Rofe, A.M., 2009. Maternal dietary zincsupplementation prevents aberrant behaviour in an object recognition task inmice offspring exposed to LPS in early pregnancy. Behav. Brain Res. 197, 210–218.
Croft, A.P., O’Callaghan, M.J., Shaw, S.G., Connolly, G., Jacquot, C., Little, H.J., 2008.Effects of minor laboratory procedures, adrenalectomy, social defeat or acutealcohol on regional brain concentrations of corticosterone. Brain Res. 1238, 12–22.
Dahlqvist, P., Rönnbäck, A., Bergström, S.A., Söderström, I., Olsson, T., 2004.Environmental enrichment reverses learning impairment in the Morris watermaze after focal cerebral ischemia in rats. Eur. J. Neurosci. 19, 2288–2298.
Dere, E., Huston, J.P., De Souza Silva, M.A., 2007. The pharmacology, neuroanatomyand neurogenetics of one-trial object recognition in rodents. Neurosci.Biobehav. Rev. 31, 673–704.
Dinarello, C.A., 1999. Cytokines as endogenous pyrogens. J. Infect. Dis. 179, S294–S304.
Duncko, R., Schwendt, M., Jezova, D., 2003. Altered glutamate receptor andcorticoliberin gene expression in brain regions related to hedonic behavior inrats. Pharmacol. Biochem. Behav. 76, 9–16.
Escobar, M., Crouzin, N., Cavalier, M., Quentin, J., Roussel, J., Lanté, F., Batista-Novais,A.R., Cohen-Solal, C., De Jesus Ferreira, M.-C., Guiramand, J., Barbanel, G., Vignes,M., 2011. Early, time dependent disturbances of hippocampal synaptictransmission and plasticity after in utero immune challenge. Biol. Psychiatry70, 992–999.
Fortier, M.E., Joober, R., Luheshi, G.N., Boksa, P., 2004. Maternal exposure to bacterialendotoxin during pregnancy enhances amphetamine-induced locomotion andstartle responses in adult rat offspring. J. Psychiatr. Res. 38, 335–345.
Fortier, M.E., Luheshi, G.N., Boksa, P., 2007. Effects of prenatal infection on prepulseinhibition in the rat depend on the nature of the infectious agent and the stageof pregnancy. Behav. Brain Res. 181, 270–277.
Francis, D.D., Diorio, J., Plotsky, P.M., Meaney, M.J., 2002. Environmental enrichmentreverses the effects of maternal separation on stress reactivity. J. Neurosci. 22,7840–7843.
French, S.S., Chester, E.M., Demas, G.E., 2013. Maternal immune activation affectslitter success, size and neuroendocrine responses related to behavior in adultoffspring. Physiol. Behav. 119, 175–184.
Garrido, P., de Blas, M., Del Arco, A., Segovia, G., Mora, F., 2012. Aging increases basalbut not stress-induced levels of corticosterone in the brain of the awake rat.Neurobiol. Aging 33, 375–382.
Giese, K.P., Fedorov, N.B., Filipkowski, R.K., Silva, A.J., 1998. Autophosphorylationand Thr286 of the a calcium-calmodulin kinase II in LTP and learning. Science279, 870–873.
Girbovan, C., Plamondon, H., 2013. Environmental enrichment in female rodents:considerations in the effects on behavior and biochemical markers. Behav. BrainRes. 253, 178–190.
Golan, H.M., Lev, V., Hallak, M., Sorokin, Y., Huleihel, M., 2005. Specificneurodevelopmental damage in mice offspring following maternalinflammation during pregnancy. Neuropharmacology 48, 903–917.
Graciarena, M., Depino, A.M., Pitossi, F.J., 2010. Prenatal inflammation impairs adultneurogenesis and memory related behavior through persistent hippocampalTGFb1 downregulation. Brain Behav. Immun. 24, 1301–1309.
Hava, G., Vered, L., Yael, M., Mordechai, H., Mahoud, H., 2006. Alterations inbehavior in adult offspring mice following maternal inflammation duringpregnancy. Dev. Psychobiol. 48, 162–168.
Hayley, S., Kelly, O., Anisman, H., 2002. Murine tumor necrosis factor-alphasensitizes plasma corticosterone activity and the manifestation of shock:modulation by histamine. J. Neuroimmunol. 131, 60–69.
Howell, D.C., 2002. Statistical Methods for Psychology, 5th ed. Duxbury Press,Belmont, Canada.
Howland, J.G., Cazakoff, B.N., Zhang, Y., 2012. Altered object-in-place recognitionmemory, prepulse inhibition, and locomotor activity in the offspring of ratsexposed to a viral mimetic during pregnancy. Neuroscience 201, 184–198.
Huang, F.L., Huang, K.P., Wu, J., Boucheron, C., 2006. Environmental enrichmentenhances neurogranin expression and hippocampal learning and memory butfails to rescue the impairments of neurogranin null mutant mice. J. Neurosci. 26,6230–6237.
Johnson, E.M., Traver, K.L., Hoffman, S.W., Harrison, C.R., Herman, J.P., 2013.Environmental enrichment protects against functional deficits caused bytraumatic brain injury. Front. Behav. Neurosci. 7, 44.
Jurgens, H.A., Johnson, R.W., 2012. Environmental enrichment attenuateshippocampal neuroinflammation and improves cognitive function duringinfluenza infection. Brain Behav. Immun. 26, 1006–1016.

190 E.J. Connors et al. / Brain, Behavior, and Immunity 42 (2014) 178–190
Kamphuis, P.J., Gardoni, F., Kamal, A., Croiset, G., Bakker, J.M., Cattabeni, F., Gispen,W.H., Van Bel, F., Di Luca, M., Wiegant, V.M., 2003. Long-lasting effects ofneonatal dexamethasone treatment on spatial learning and hippocampalsynaptic plasticity: involvement of the NMDA receptor complex. FASEB J. 17,911–913.
Kentner, A.C., Miguelez, M., James, J.S., Bielajew, C., 2006. Behavioral andphysiological effects of a single injection of rat interferon-alpha on maleSprague–Dawley rats: a long-term evaluation. Brain Res. 1095, 96–106.
Kentner, A.C., James, J.S., Miguelez, M., Bielajew, C., 2007. Investigating the hedoniceffects of interferon-a on female rats using brain-stimulation reward. Behav.Brain Res. 177, 90–99.
Kentner, A.C., Takeuchi, A., James, J.S., Miki, T., Seino, S., Hayley, S., Bielajew, C., 2008.The effects of rewarding ventral tegmental area stimulation and environmentalenrichment on lipopolysaccharide-induced sickness behavior and cytokineexpression in female rats. Brain Res. 1217, 50–61.
Kirsten, T.B., Taricano, M., Maiorka, P.C., Palermo-Neto, J., Bernardi, M.M., 2010.Prenatal lipopolysaccharide reduces social behavior in male offspring.NeuroImmunoModulation 17, 240–251.
Kirsten, T.B., Chaves-Kirsten, G.P., Chaible, L.M., Silva, A.C., Martins, D.O., Britto, L.R.,Dagli, M.L., Torrão, A.S., Palermo-Neto, J., Bernardi, M.M., 2012. Hypoactivity ofthe central dopaminergic system and autistic-like behavior induced by a singleearly prenatal exposure to lipopolysaccharide. J. Neurosci. Res. 90, 1903–1912.
Lambert, T.J., Fernandez, S.M., Frick, K.M., 2005. Different types of environmentalenrichment have discrepant effects on spatial memory and synaptophysinlevels in female mice. Neurobiol. Learn. Mem. 83, 206–216.
Lanté, F., Meunier, J., Guiramand, J., Maurice, T., Cavalier, M., de Jesus Ferreira, M.-C.,Aimar, R., Cohen-Solal, C., Vignes, M., Barbanel, G., 2007. Neurodevelopmentaldamage after prenatal infection: role of oxidative stress in the fetal brain. FreeRadical Biol. Med. 42, 1231–1245.
Leger, M., Quiedeville, A., Paizanis, E., Natkunarajah, S., Freret, T., Boulouard, M.,Schumann-Bard, P., 2012. Environmental enrichment enhances episodic-likememory in association with a modified neuronal activation profile in adultmice. Plos One, e48043.
Li, M., Wang, M., Ding, S., Li, C., Luo, X., 2012. Environmental enrichment duringgestation improves behavioral consequences and synaptic plasticity inhippocampus of prenatal-stressed offspring rats. Acta Histochem. Cythochem.45, 157–166.
Lipkind, H.S., Curry, A.E., Huynh, M., Thorpe, L.E., Matte, T., 2010. Birth outcomesamong offspring of women exposed to the September 11, 2001, terroristattacks. Obstet. Gynecol. 116, 917–925.
Liu, D., Diorio, J., Day, J.C., Francis, D.D., Meaney, M.J., 2000. Maternal care,hippocampal synaptogenesis and cognitive development in rats. Nat. Neurosci.3, 799–806.
Maier, S.F., Goehler, L.E., Fleshner, M., Watkins, L.R., 1998. The role of the vagusnerve in cytokine-to-brain communication. Ann. N.Y. Acad. Sci. 840, 289–300.
Malkova, N.V., Yu, C.Z., Hsiao, E.Y., Moore, M.J., Patterson, P.H., 2012. Maternalimmune activation yields offspring displaying mouse versions of the three coresymptoms of autism. Brain Behav. Immun. 26, 607–616.
Marcondes, F.K., Bianchi, F.J., Tanno, A.P., 2002. Determination of the estrous cyclephases of rats: some helpful considerations. Braz. J. Biol. 62, 609–614.
Meaney, M.J., Aitken, D.H., Viau, V., Sharma, S., Sarrieau, A., 1989. Neonatal handlingalters adreno cortical negative feedback sensitivity and hippocampal type IIglucocorticoid receptor binding in the rat. Neuroendocrinology 50, 597–604.
Meaney, M.J., Aitken, D.H., Sharma, S., Viau, V., 1992. Basal ACTH, corticosterone andcorticosterone-binding globulin levels over the diurnal cycle, and hippocampalcorti-costeroid receptors in young and aged, handled and non-handled rats.Neuroendocrinology 55, 204–213.
Meyer, U., Feldon, J., Schedlowski, M., Yee, B.K., 2006. Immunological stress at thematernal–foetal interface. A link between neurodevelopment and adultpsychopathology. Brain Behav. Immun. 20, 378–388.
Meyer, U., Yee, B.K., Feldon, J., 2007. The developmental impact of prenatalinfections at different times of pregnancy: the earlier the worse? Neuroscientist13, 241–256.
Meyer, U., Engler, A., Weber, L., Schedlowski, M., Feldon, J., 2008. Preliminaryevidence for a modulation of fetal dopaminergic development by maternalimmune activation during pregnancy. Neuroscience 154, 701–709.
Morley-Fletcher, S., Rea, M., Maccari, S., Laviola, G., 2003. Environmentalenrichment during adolescence reverses the effects of prenatal stress on playbehaviour and HPA axis reactivity in rats. Eur. J. Neurosci. 18, 3367–3374.
Mueller, B.R., Bale, T.L., 2008. Sex-specific programming of offspring emotionalityafter stress early in pregnancy. J. Neurosci. 28, 9055–9065.
Mychasiuk, R., Muhammad, A., Kolb, B., 2014. Environmental enrichment altersstructural plasticity of the adolescent brain but does not remediate the effectsof prenatal nicotine exposure. Synapse 68, 293–305.
Nugent, J.L., Khashan, A.S., Baker, P.N., 2011. Reduced infant birth weight in theNorth West of England consequent upon ‘maternal exposure’ to 7/7 terroristattacks on central London. J. Obstet. Gynaecol. 31, 118–121.
Ohlsson, A.L., Johansson, B.B., 1995. Environment influences functional outcome ofcerebral infarction in rats. Stroke 26, 644–649.
Patterson, P.H., 2009. Immune involvement in schizophrenia and autism: etiology,pathology, and animal models. Behav. Brain Res. 204, 313–321.
Paxinos, G., Watson, C., 1998. The Rat Brain in Stereotaxic coordinates, 2nd ed.Academic Press, Sydney.
Pellis, S.M., Field, E.F., Smith, L.K., Pellis, V.C., 1997. Multiple differences in the playfighting of male and female rats. Implications for the causes and functions ofplay. Neurosci. Biobehav. Rev. 21, 105–120.
Pembrey, M.E., 2002. Time to take epigenetic inheritance seriously. Eur. J. Hum.Genet. 10, 669–671.
Perez-Sepulveda, J.A., Flagel, S.B., Garcia-Fuster, M.J., Slusky, R.J., Aldridge, J.W.,Watson, S., Akil, H., 2013. Differential impact of a complex environment onpositive affect in an animal model of individual differences in emotionality.Neuroscience 248, 436–447.
Poinkewitz, Y., Arad, M., Weiner, I., 2011. Abnormal trajectories ofneurodevelopment and behavior following in utero insult to the rat. Biol.Psychiatry 70, 842–851.
Ravenelle, R., Byrnes, E.M., Byrnes, J.J., McInnis, C., Park, J.H., Donaldson, S.T., 2013.Environmental enrichment effects on the neurobehavioral profile of selectiveoutbred trait anxiety rats. Behav. Brain Res. 252, 49–57.
Reichel, C.M., Gilstrap, M.G., Ramsey, L.A., See, R.E., 2014. Modafinil restoresmethamphetamine induced object-in-place memory deficits in ratsindependent of glutamate N-methyl-D-aspartate receptor expression. DrugsAlcohol Depend. 134, 115–122.
Roceri, M., Hendriks, W., Racagni, G., Ellenbroek, B.A., Riva, M.A., 2002. Earlymaternal deprivation reduces the expression of BDNF and NMDA receptorsubunits in rat hippocampus. Mol. Psychiatry 7, 609–616.
Schneider, T., Turczak, J., Przewłocki, R., 2006. Environmental enrichment reversesbehavioral alterations in rats prenatally exposed to valproic acid: issues for atherapeutic approach in autism. Neuropsychopharmacology 31 (1), 36–46.
Shi, L., Fatemi, S.H., Sidwell, R.W., Patterson, P.H., 2003. Maternal influenza infectioncauses marked behavioral and pharmacological changes in the offspring. J.Neurosci. 23, 297–302.
Soberanes-Chávez, P., López-Rubalcava, C., de Gortari, P., Cruz, S.L., 2013. Exposureto toluene and stress during pregnancy impairs pups’ growth and dams’lactation. Neurotoxicol. Teratol. 40, 9–16.
Son, H.G., Geum, Dongho, Chung, S., Kim, E.J., Jo, J.-H., Kim, C.-M., Lee, K.H., Kim, H.,Choi, S., Kim, H.T., Lee, C.-J., Kim, K., 2006. Maternal stress produced learningdeficits associated with impairment of NMDA receptor-mediated synapticplasticity. J. Neurosci. 26, 2209–3318.
Spires, T.L., Grote, H.E., Varshney, N.K., Cordery, P.M., van Dellen, A., Blakemore, C.,Hannan, A.J., 2004. Environmental enrichment rescues protein deficits in amouse model of Huntington’s disease, indicating a possible disease mechanism.J. Neurosci. 24, 2270–2276.
Steiner, A.A., Ivanov, A.I., Serrats, J., Hosokawa, H., Phayre, A.N., Robbins, J.R.,Roberts, J.L., Kobayashi, S., Matsumura, K., Sawchenko, P.E., Romanovsky, A.A.,2006. Cellular and molecular bases of the initiation of fever. PLoS Biol. 4, e284.
Taylor, P.V., Veenema, A.H., Paul, M.J., Bredewold, R., Isaacs, S., de Vries, G.J., 2012.Sexually dimorphic effects of a prenatal immune challenge on social play andvasopressin expression in juvenile rats. Biol. Sex Differ. 3, 15–24.
Terranova, J.-P., Chabot, C., Barnouin, M.-C., Perrault, G., Depoortere, R., Griebel, G.,Scatton, B., 2005. SSR181507, a dopamine D2 receptor antagonist and 5-HT1Areceptor agonist, alleviates disturbances of novelty discrimination in a socialcontext in rats, a putative model of selective attention deficit.Psychopharmacology 181, 134–144.
Vivinetto, A.L., Suárez, M.M., Rivarola, M.A., 2013. Neurobiological effects ofneonatal separation and post-weaning environmental enrichment. Behav.Brain Res. 240, 110–118.
Warburton, E.C., Barker, G.R.I., Brown, M.W., 2013. Investigations into theinvolvement of NMDA mechanisms in recognition memory.Neuropharmacology 74, 41–47.
Wiener, S.G., Levine, S., 1978. Perinatal malnutrition and early handling: interactiveeffects on the development of the pituitary-adrenal system. Dev. Psychobiol. 11,335–352.
Wolff, A.R., Bilkey, D.K., 2010. The maternal immune activation (MIA) model ofschizophrenia produced pre-pulse inhibition deficits in both juvenile and adultrats but these effects are not associated with maternal weight loss. Behav. BrainRes. 213 (323), 327.
Zhang, T.Y., Labonté, B., Lan, X.W., Turecki, G., Meaney, M.J., 2013. Epigeneticmechanisms for the early environmental regulation of hippocampalglucocorticoid receptors gene expression in rodents and humans.Neuropsychopharmacol. Rev. 38, 111–123.
Zhao, D., Liu, D., Chen, X., Wang, K., Zhang, A., Kang, J., Zhou, Q., Duan, T., 2013.Prenatal stress disturbs hippocampal KIF17 and NR2B in spatial cognition inmale offspring. J. Neurosci. Res. 91, 535–544.
Zimmermann, A., Stauffacher, M., Langhans, W., Wurbel, H., 2001. Enrichment-dependent differences in novelty exploration in rats can be explained byhabituation. Behav. Brain Res. 121, 11–20.




















