
Molecular basis for checkpoints in the CD8 T cell response: Tolerance versus activation
16
Molecular Basis for Checkpoints in the CD8 T Cell Response: Tolerance versus Activation Matthew F. Mescher * , Pujya Agarwal, Kerry A. Casey, Christopher D. Hammerbeck, Zhengguo Xiao, and Julie M. Curtsinger Center for Immunology and Department of Laboratory Medicine and Pathology, 420 Delaware St. S.E., University of Minnesota, Minneapolis, MN 55455, USA Abstract CD8 T cells specific for self-antigens are present in the peripheral lymphoid system and can contribute to autoimmunity or transplant rejection. Whether recognition of Ag leads to full activation, or to induction of tolerance, depends upon availability of cytokine at critical stages of the response. Signals provided by IL-12 and/or IFN-α/β are required for activation of naïve CD8 T cells, and IL-2 is needed to sustain and further expand the effector cells if Ag persists. These critical signaling requirements provide new insights into the factors that regulate the CD8 T cell contributions to development of autoimmunity or rejection of transplants. Keywords CD8 T lymphocyte; activation; differentiation; anergy; peripheral tolerance 1. Introduction Central tolerance resulting from negative selection in the thymus eliminates the majority of self-reactive CD8 T cells, but is incomplete. Despite the presence of the AIRE system that provides for presentation of many self-Ags in the thymus [1], some self-reactive T cells do escape negative selection and enter the periphery. Thus, mature naïve CD8 T cells that can respond to self-Ag are present in the lymphoid organs where they can potentially contribute to autoimmunity and transplant rejection. On the positive side, however, these cells also have the potential to respond to self Ags expressed on tumor cells, and are the target of vaccine approaches to cancer immunotherapy. Thus, understanding the mechanisms that can result in the induction of peripheral tolerance in CD8 T cells is important for developing strategies to avoid responses that contribute to autoimmunity or transplant rejection, as well as for devising ways of circumventing the induction of tolerance to Ags on tumor cells. Mechanisms responsible for establishing peripheral tolerance can be broadly classified into three categories; ignorance, deletion and anergy. Clonal ignorance is the simplest form of peripheral tolerance; the naïve T cells circulating in the lymphoid system fail to gain access to the peripheral site where Ag is present, and sufficient Ag is not being carried to the lymph nodes draining the site to result in T cell activation in the nodes [2]. Tolerance due to ignorance is tenuous, however, because some peripheral self-Ags do reach the lymph nodes via migrating *Correspondence: Matthew F. Mescher, Phone: +1 612 626 2368, Fax: +1 612 625 2199, Email: [email protected]. Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. NIH Public Access Author Manuscript Semin Immunol. Author manuscript; available in PMC 2007 December 1. Published in final edited form as: Semin Immunol. 2007 June ; 19(3): 153–161. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Molecular basis for checkpoints in the CD8 T cell response: Tolerance versus activation

Molecular Basis for Checkpoints in the CD8 T Cell Response:Tolerance versus Activation
Matthew F. Mescher*, Pujya Agarwal, Kerry A. Casey, Christopher D. Hammerbeck,Zhengguo Xiao, and Julie M. CurtsingerCenter for Immunology and Department of Laboratory Medicine and Pathology, 420 Delaware St.S.E., University of Minnesota, Minneapolis, MN 55455, USA
AbstractCD8 T cells specific for self-antigens are present in the peripheral lymphoid system and cancontribute to autoimmunity or transplant rejection. Whether recognition of Ag leads to full activation,or to induction of tolerance, depends upon availability of cytokine at critical stages of the response.Signals provided by IL-12 and/or IFN-α/β are required for activation of naïve CD8 T cells, and IL-2is needed to sustain and further expand the effector cells if Ag persists. These critical signalingrequirements provide new insights into the factors that regulate the CD8 T cell contributions todevelopment of autoimmunity or rejection of transplants.
KeywordsCD8 T lymphocyte; activation; differentiation; anergy; peripheral tolerance
1. IntroductionCentral tolerance resulting from negative selection in the thymus eliminates the majority ofself-reactive CD8 T cells, but is incomplete. Despite the presence of the AIRE system thatprovides for presentation of many self-Ags in the thymus [1], some self-reactive T cells doescape negative selection and enter the periphery. Thus, mature naïve CD8 T cells that canrespond to self-Ag are present in the lymphoid organs where they can potentially contribute toautoimmunity and transplant rejection. On the positive side, however, these cells also have thepotential to respond to self Ags expressed on tumor cells, and are the target of vaccineapproaches to cancer immunotherapy. Thus, understanding the mechanisms that can result inthe induction of peripheral tolerance in CD8 T cells is important for developing strategies toavoid responses that contribute to autoimmunity or transplant rejection, as well as for devisingways of circumventing the induction of tolerance to Ags on tumor cells.
Mechanisms responsible for establishing peripheral tolerance can be broadly classified intothree categories; ignorance, deletion and anergy. Clonal ignorance is the simplest form ofperipheral tolerance; the naïve T cells circulating in the lymphoid system fail to gain access tothe peripheral site where Ag is present, and sufficient Ag is not being carried to the lymphnodes draining the site to result in T cell activation in the nodes [2]. Tolerance due to ignoranceis tenuous, however, because some peripheral self-Ags do reach the lymph nodes via migrating
*Correspondence: Matthew F. Mescher, Phone: +1 612 626 2368, Fax: +1 612 625 2199, Email: [email protected]'s Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customerswe are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resultingproof before it is published in its final citable form. Please note that during the production process errors may be discovered which couldaffect the content, and all legal disclaimers that apply to the journal pertain.
NIH Public AccessAuthor ManuscriptSemin Immunol. Author manuscript; available in PMC 2007 December 1.
Published in final edited form as:Semin Immunol. 2007 June ; 19(3): 153–161.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

dendritic cells (DC), and pathological insults can lead to increased levels of these Ags becomingavailable for T cell recognition. Peripheral tolerance can still occur, however, due to clonaldeletion or anergy that can result when the T cell recognizes Ag but the ensuing TCR-initiatedcell division and differentiation are incomplete, and lead to either death of the Ag-activatedcells (deletion), or to the induction of a long-lived non-responsive state (anergy) [3].
For CD8 T cells, there is now considerable evidence that the mechanisms responsible fortolerance induction by deletion or establishment of anergy involve the recognition of Ag in theabsence of additional signals that are required to support development of full responsiveness,and the molecular basis for these additional signaling requirements is beginning to be clarified.It appears that these additional requirements represent ‘checkpoints’ in the CD8 T cell responsethat help to insure that a productive response occurs only under appropriate circumstances, e.g.upon infection with a pathogen or when help from CD4 T cells is available. When Ag isrecognized and a checkpoint is reached, but the additional signal is not available, the result isestablishment of peripheral tolerance in the cells specific for the Ag.
2. Activation and differentiation of naïve CD8 T cellsIn most cases, naïve CD8 T cells first encounter Ag being cross-presented by DC in lymphnodes [4,5], and the outcome depends on the maturation state of the DC [6]. Presentation byimmature, quiescent DC results in tolerance induction by a mechanism involving deletion,induction of non-responsiveness (anergy), or both. In contrast, the T cells make a productiveresponse if the DC have been activated to mature as a result of either engagement of their Toll-like receptors (TLR) by pathogen-derived polymers (adjuvants), or by a CD40-dependentinteraction with CD4 T helper cells [7-9]. Thus, the DC must be ‘licensed’ or ‘conditioned’ topresent Ag in a manner that stimulates a productive response and avoids tolerance induction.This is often attributed to increased expression of Ag and/or costimulatory ligands B7-1 (CD80)and B7-2 (CD86) on the activated DC, but there is no direct evidence that these quantitativechanges are responsible for the different outcomes. Furthermore, there are reports of DC failingto activate productive CD8 T cell response even when they are expressing high levels ofcostimulatory ligands [10-12]. Considerable evidence has accumulated to indicate that survivaland differentiation of CD8 T cells responding to Ag and costimulation require activation of asignaling pathway(s) distinct from those activated by the TCR and CD28 receptors, and thatcytokines produced by mature DC can provide the ‘third signal’ needed to activate thispathway.
2.1. A ‘three signal’ model for CD8 T cell activationActivation of T cells requires both Ag engagement by the TCR and signals provided by acostimulatory receptor, usually CD28 binding to its B7 ligands [13-16]. These classical ‘twosignals’ are necessary for activation of naïve CD8 T cells, but alone are not sufficient to supportstrong clonal expansion, development of effector functions, or establishment of a long-lived,responsive memory population. Instead, a third signal is needed, and can be provided by eitherIL-12 or Type I IFN (IFN-α/β). This was initially suggested by in vitro studies showing thathighly purified naïve CD8 T cells could not expand and develop cytolytic activity in responseto artificial APC presenting just Ag and B7, but responded strongly if either IL-12 [17] or IFN-α/β [18] was present. In contrast, CD44hi memory cells bearing the same TCR responded tothe artificial APC presenting Ag and B7 by undergoing strong clonal expansion and developingeffector functions [17]; they did not require a third signal. Earlier work had shown that IL-12can augment generation of cytotoxic T lymphocyte (CTL) responses [19,20] and that IFN-α/β could promote survival [21] and IFN-γ [22] production by CD8 T cells, but the critical roleof these cytokines in determining whether Ag recognition leads to tolerance versus fullactivation and development of a memory population has only recently been realized.
Mescher et al. Page 2
Semin Immunol. Author manuscript; available in PMC 2007 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

In vivo studies also provide strong support for a model in which naïve CD8 T cells require athird signal to develop effector functions and transition to a memory population. Immunizationwith a CD8 epitope peptide results in tolerance induction unless an adjuvant is given alongwith the peptide [23-25]. Co-administration of either IL-12 [26,27] or IFN-α [28-30] alongwith peptide replaces the requirement for adjuvant, and supports generation of strong primaryclonal expansion, development of effector functions, and establishment of a long-lived,responsive memory population. It appears likely that the major role of adjuvants in supportingCD8 T cell responses to Ag, in addition to increasing expression levels of costimulatoryligands, is that of inducing IL-12 and/or Type I IFN production by DC and other cells throughengagement of TLR.
Experiments examining the responses of CD8 T cells lacking the IL-12 receptor (IL-12R) orthe Type I IFN receptor (IFN-IR) further support a three-signal model for CD8 T cell activation.Antigen-specific CD8 T cells that lack the IFN-IR exhibit a greater than 99% reduction in theirability to respond to lymphocytic choriomeningitis virus (LCMV) infection [28,29]. Incontrast, responses of the IFN-IR-deficient cells to vaccinia virus (VV), vesicular stomatitisvirus or Listeria monocytogenes (LM) are much less impaired [28,31]. We have found thatAg-specific CD8 T cells lacking the IL-12R exhibit a stronger reduction in response to VVand LM than do cells lacking the IFN-IR, and that generation of memory populations is onlyeliminated when the cells are deficient in both receptors (Xiao et.al., manuscript inpreparation). Thus, it appears that either IL-12 or Type I IFN can support in vivo responsesto pathogens, and that the levels of the cytokines produced in response to a particular pathogenwill dictate which makes the greater contribution. In contrast, CD8 T cell recognition of a self-Ag, which may normally occur under conditions where these cytokines are not being produced,will result in an abortive proliferative response leading to tolerance. Alternatively, productiveCD8 T cell responses to self-Ag that contribute to autoimmunity or transplant rejection mayresult in cases where IL-12, IFN-α/β, or potential alternate third signals, are present during theinitial period of interaction with the Ag. One way that this may occur is by CD4 T helper cellsstimulating production of these cytokines by DC.
2.2 CD4 T helper cells condition DC to provide signal 3One of the ways in which CD4 T cells can provide help for a CD8 T cell response is byinteracting with DC that are cross-presenting class I Ag to ‘condition’ these DC to effectivelyactivate naïve CD8 T cells, and this requires binding of CD40 ligand (CD40L; CD154)expressed on the CD4 T cell to CD40 on the DC [7-9]. Engagement of CD40 on DC stimulatesthe cells to produce IL-12 [32], suggesting that ‘conditioning’ of the DC by CD4 T cell mightinvolve stimulating the production of this cytokine so that the DC can provide the critical thirdsignal for CD8 T cell activation. Hernandez et.al. [12] provided evidence consistent with thispossibility by demonstrating that IL-12 could bypass a requirement for CD4 T cells in a CD8-dependent diabetes model. Generation of the CD8 response and development of diseasedepended on CD40L-dependent CD4 T cell help, but both occurred in the absence of CD4 helpwhen IL-12 was administered to the mice.
The involvement of IL-12 production by DC in response to CD4 T cell help for a CD8 T cellresponse was directly demonstrated in a study by Filatenkov et.al. [33] examining an ectopicheart transplant model. In this model, donor mice expressed a membrane form of ovalbumin(mOVA) as a transgene under the control of an actin promoter, so that OVA was expressed inall tissues, including the heart. Recipient mice first received, by adoptive transfer, TCRtransgenic OT-I CD8 and OT-II CD4 T cells, specific for H-2Kb/OVA257-264 and I-Ab/OVA323-339 respectively, and subsequently received heterotopic mOVA heart transplants.Thus, mOVA served as a model minor Ag, and the use of adoptively transferred CD4 and CD8T cells allowed direct visualization of the responses of these cells to the graft. Rapid graft
Mescher et al. Page 3
Semin Immunol. Author manuscript; available in PMC 2007 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

rejection required the presence of both OT-I and OT-II cells, and required that the OT-II cellsexpress CD40L. When both OT-I and OT-II cells were present, OT-I cells clonally expanded,migrated to the graft, expressed high granzyme B and IFN-γ levels, and mediated rapid graftrejection.
In contrast, OT-I cells underwent some clonal expansion and migrated to the graft in theabsence of OT-II cells, but expressed low levels of granzyme B and IFN-γ, and did not causerapid rejection, suggesting that a signal 3 cytokine was not available to support their response.Administration of IL-12 replaced the requirement for CD4 help, causing the OT-I cells toexpress high granzyme B and IFN-γ and mediate rapid graft rejection. Furthermore, even whenOT-II cells were present, OT-I cells that lacked the IL-12R failed to upregulate the expressionof effector proteins or mediate graft rejection. It was also shown that DC from mice havingOT-II cells produced IL-12, but only if the OT-II cells expressed CD40L. Thus, in this modelof minor Ag-mediated transplant rejection, the evidence very strongly suggests that CD4 Tcells provide help for CD8 T cells, at least in part, by upregulating IL-12 production by DC sothat they can provide the third signal to the CD8 T cells to support a productive response. TheCD4 T cells may also upregulate expression of costimulatory ligands on the DC, but this doesnot appear to be essential, since administration of IL-12 was sufficient to replace the need forhelp.
Under most circumstances, peripheral CD8 T cells with the potential to contribute toautoimmunity or graft rejection will undergo tolerance induction if they encounter the self-Agthey recognize, as a result of failing to receive a third signal needed to support theirdifferentiation. However, if the encounter happens at a time when there is sufficient IL-12 and/or IFN-α/β present to provide the third signal, then strong clonal expansion and developmentof effector functions may result to contribute to disease. This can occur if CD4 helper cells are‘conditioning’ DC to produce the necessary signal 3 cytokine, but may also occur if aconcomitant infection induces production of these cytokines triggered by TLR ligandsproduced by the pathogen.
2.3 Signaling pathways for IL-12 and IFN-α-dependent differentiationBrief interaction of CD8 T cells with Ag and costimulation provides sufficient signals toprogram the cells to proliferate; allowing cells to interact in vitro with artificial APC for evena few hours, followed by removal of the stimulus, results in multiple rounds of division overthe next three days [34]. Under these conditions, however, survival is limited so that clonalexpansion is minimal by day 3 despite extensive cell division, the cells produce little IFN-γupon re-stimulation, and they fail to develop cytolytic effector function [35]. When either IL-12or IFN-α are present, the rate of cell division does not change significantly but survival, andthus clonal expansion, are increased, and effector functions develop by day 3 .
In contrast to the brief interaction required for Ag and costimulation to program the cells todivide, optimum survival and development of effector functions requires more prolongedsignaling. In the case of IL-12, we found that optimal clonal expansion by 72 hr required thatAg and B7 were present for 24 hr, and that IL-12 was present from about 15 to 30 hr [35].More prolonged exposure to IL-12, up to 60 hr, was required for optimum development ofeffector functions. The timing requirements for stimulation by IFN-α have not been examined,but are likely to mirror those seen for IL-12. These results suggest that varying levels oftolerance may result for CD8 T cells responding to Ag, depending not only on whether a signal3 cytokine is available, but also on the duration of its availability. Thus, two signals aresufficient to rapidly program cell division, but the response is abortive in the absence of a thirdsignal. Only when the signals provided by IL-12 or IFN-α/β are present for a prolonged periodare the cells fully programmed to survive and become effectors.
Mescher et al. Page 4
Semin Immunol. Author manuscript; available in PMC 2007 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Recognition of Ag by the TCR activates the Ras-MAPK pathway, and costimulation providedby CD28 activates the phosphatidyl inositol-3-kinase pathway and synergizes to augment TCRinduced transcriptional regulation mediated by AP-1, NF-AT and NF-κB. In contrast tosignaling by the TCR and CD28, the Jak/Stat (Janus kinase/signal transducer and activator oftranscription) pathways play a major role in signaling by cytokine receptors. Upon cytokinebinding, Jaks are activated and phosphorylate the receptor to form docking sites for STATs,latent transcription factors present in the cytosol. Upon docking to the phosphorylated receptorthe Stats are phosphorylated by Jaks, resulting in dimerization and nuclear translocation, wherethe STATs then regulate gene transcription by binding to specific DNA motifs and recruitingother transcription factors and coactivators.
The IL-12 receptor, a hetrodimeric receptor composed of IL-12Rβ1 and β2 chains, recruitsJAK2 and TYK2, leading to phosphorylation of STAT-4 to form homodimers, and thisconstitutes the major IL-12 signaling pathway. Mice lacking the IL-12R have defects in cellmediated immunity and Th1 CD4 helper cell development, and Stat-4-deficient mice have avery similar phenotype [36,37]. Thus, not surprisingly, provision of the ‘third signal’ to naïveCD8 T cells by IL-12 requires Stat-4. When wild type BalbC cells are stimulated in vitro withanti-TCR mAb and B7-1 on artificial APC, addition of IL-12 supports development of cytolyicfunction and the ability to produce IFN-γ. In contrast, these effector functions fail to developwhen naïve CD8 T cells from Stat-4-deficient mice are stimulated in the same way [18].
IFN-α/β strongly activates Stat-1 and Stat-2 upon binding to the IFN-IR, but can variablyactivate other Stats, including Stat-3, 4 and 6, depending on the cell type being examined.Earlier studies suggested that IFN-α could activate Stat-4 in human T cells, but that Stat-4activation was impaired in murine CD4 T cells due to a mini-satellite insertion in murine Stat-2that resulted in impaired recruitment of Stat-4 to the receptor [38]. However, a subsequentstudy demonstrated that during an LCMV infection, IFN-α signaling led to Stat-4 being morestrongly activated in CD8 T cells than in CD4 T cells, and promoted Stat-4-dependent IFN-γproduction by the CD8 T cells [22]. Using naïve CD8 T cells from wild type and Stat-4 deficientmice, we found that IFN-α-dependent provision of signal 3 was largely dependent on Stat-4[18], as assessed by strongly decreased development of cytolytic activity and ability to produceIFN-γ, although loss of function was not as complete as in the case of stimulation with IL-12.
While development of effector functions demonstrated similar Stat-4-dependence in responseto IL-12 and IFN-α, more recent work suggests that the signaling pathways involved inpromoting CD8 T cell differentiation may differ substantially for these two cytokines (Agarwaland Mescher, unpublished results). Artificial APC presenting anti-TCR mAb and B7-1 wereused to stimulate wild type and Stat-4-deficient naïve CD8 T cells in the absence or presenceof either IL-12 or IFN-α for 72 hr, and oligonucleotide microarray analysis was then done. Incomparison to cells stimulated in the absence of a cytokine, IL-12 stimulated the up- or down-regulation of about 200 genes, and regulation of all but 2 was eliminated in the Stat-4-deficientcells. In contrast, IFN-α stimulated the up- or down-regulation of about 300 genes, and abouta third of these showed similar regulation in the STAT-4-deficient cells. Genes regulated incommon by both IL-12 and IFN-α are likely candidates for contributing importantly todifferentiation, since functional outcomes are very similar with the two cytokines, and therewere approximately 35 such genes in wild type cells. Of this commonly regulated subset,regulation of all of the genes was Stat-4-dependent in response to IL-12, but regulation of onlyabout one-third of them was Stat-4-dependent in response to IFN-α. Thus, these preliminaryresults examining the ‘third signal’ pathways at the level of gene expression regulation suggestthat while Stat-4 activation contributes in both cases, activation of additional Stats is likely tomake an important contribution to the response to IFN-α. In addition, the results suggest thatwhile either cytokine can support development of effector functions and a memory population,the phenotypes of the resulting cells may differ.
Mescher et al. Page 5
Semin Immunol. Author manuscript; available in PMC 2007 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

2.4. Alternate ‘third signals’The phenotype and functions of T cells can be modified in response to a wide array of cytokines,but IL-12 and IFN-α are distinguished by the fact that they can be essential for productive Ag-dependent activation of naïve CD8 T cells, i.e. they can act as the ‘switch’ that determineswhether a response to Ag will result in tolerance versus full activation leading to formation ofa responsive memory population. This, together with the fact that their action is mediatedthrough activation of a signaling pathway(s) distinct from those activated by the TCR andCD28, provide the rationale for viewing them as a ‘third signal’. A number of other cytokineshave been found to lack signal 3 activity based on their inability to support development ofeffector functions in naïve CD8 T cells stimulated in vitro with artificial APCs presenting Agand B7-1, including IL-1, -2, -4, -6, -7, -15, -18, -TNF-α and IFN-γ [18]. Some of thesecytokines can modify responses of developing CD8 T cells. IL-4, for example, can skeweffector cells to an IL-4-producing Tc2 phenotype [39], and IL-2 can promote proliferationand clonal expansion (see below). IFN-γ has been referred to as a signal 3 cytokine [40] basedon its ability to promote optimal expansion of CD8 T cells responding to LCMV [41,42] orimmunization with peptide and LPS [43], and on the fact that it does so by acting directly onthe CD8 T cell, at least in the case of the LCMV response [42]. While IFN-γ can clearly modifythe extent of proliferation/clonal expansion, it does not support development of effectorfunctions by naïve CD8 T cells in response to signals 1 and 2 ([18] and unpublished results).We would suggest that the term ‘signal three’ be reserved for cytokines or ligands that actcomparably to IL-12 and IFN-α, i.e. cytokines that act directly on naïve CD8 T cells respondingto Ag and costimulation to support development of effector functions in vitro, and to supportproductive activation and development of a memory population in vivo. Thus, signal 3cytokines would be defined as those that initiate the CD8 T cell differentiation program, ratherthan simply modifying the program once it is initiated.
While IL-12 and/or IFN-α appear to be the major contributors to several responses, as discussedabove, it is possible that other cytokines or cell surface ligands may have a comparable function.We have found that IL-21 [44], a member of the common gamma chain family of cytokinesthat is produced by activated CD4 T cells, promotes clonal expansion and supportsdevelopment of cytolytic effector function by naïve CD8 T cells stimulated in vitro with Agand B7-1 (Casey and Mescher, manuscript submitted), suggesting that it may act as a signal 3cytokine. Work is in progress to determine whether IL-21 also acts directly on CD8 T cells invivo to support productive activation, avoidance of tolerance and development of memory. Ifso, it could provide a means by which CD4 T cells provide help to initiate a CD8 T cell response,in addition to the CD4-dependent conditioning of DC discussed in the following section.
2.5 Long-term fate of CD8 T cells activated in the absence of signal 3Tolerance of CD8 T cells can involve deletion of the cells, induction of anergy, or both,depending on the response being examined [3]. The tolerance that results when naïve CD8 Tcell respond to Ag in the absence of signal 3 also appears likely to involve both of theseoutcomes. IL-12 and IFN-α promote survival of the responding cells as they become effectorcells, at least in part by upregulating expression of Bcl-3 [45]. Thus, the death of the majorityof the responding cells that occurs following the peak of clonal expansion is more rapid andprofound in the absence of a third signal ([26] and unpublished results). Nevertheless, someof the cells that respond in the absence of a third signal can survive long term, at least in thecase of immunization with peptide, and these cells are anergic in that they do not respond tore-challenge with a potent stimulus of Ag and adjuvant [26,27]. What determines the balancebetween deletion and survival of anergic cells is not clear, but there is considerable evidenceto suggest that the ‘strength’ of interaction with Ag (TCR affinity, Ag dose and duration) maybe an important factor, as Redmond and Sherman [3] have recently discussed.
Mescher et al. Page 6
Semin Immunol. Author manuscript; available in PMC 2007 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

3. Activation-induced non-responsiveness (AINR)When costimulation and a signal 3 cytokine are available, naïve CD8 T cells begin to divideabout 24 hr after initially encountering Ag, and proceed through multiple rounds of divisionso that clonal expansion peaks at about 72 hr, and the cells have developed effector functionsby this time. A second form of tolerance can occur at this stage of the response, however, ifthe resulting effector cell population is insufficient to clear the Ag. In the face of persistent Ag,the activated CD8 T cells undergo a gradual decline in number and functional capacity, asdemonstrated in studies examining responses to HY Ag [46] and LCMV [23,47]. There isconsiderable evidence to suggest that this results from another checkpoint in the CD8 T cellresponse.
Effector CD8 T cells develop AINRWhen CD8 T cells are stimulated in vitro they develop effector functions by day 3, but areanergic [48]. Signaling through the TCR still occurs, as evidenced by killing of Ag-bearingtarget cells and upregulation of IFN-γ production, but the cells can no longer produce IL-2 tosupport further proliferation in response to Ag and costimulation. This resembles the classicalanergy induced in CD4 T cells when they are stimulated by TCR engagement in the absenceof costimulation [16,49,50]. It clearly differs, however, in that the CD8 T cell anergy developseven when costimulation and a signal 3 cytokine are present along with the Ag stimulus. Thus,we have termed this ‘activation-induced non-responsiveness’ (AINR) to distinguish it fromclassical CD4 T cell anergy. Although CD8 T cells that have developed AINR cannot produceIL-2, they can still respond to the cytokine by proliferating. Furthermore, when exogenous IL-2is provided in vitro to drive proliferation of effector CTL for one to two days, the AINR stateis reversed and the cells regain the ability to produce IL-2 in response to Ag [51]. The AINRstate that develops during a response by naïve CD8 T cells resembles the ‘split anergy’described by Otten and Germain for cloned CTL lines [52].
Development of AINR appears to be a normal part of the CD8 T cell differentiation program.This non-responsive state develops in CD8 T cells responding to LCMV virus [53] orallogeneic tumor [48], where Ag is cleared, or to syngeneic tumor [54] or persistant virus[55], where Ag persists. For responses to both syngeneic tumor [56] and persistent virus [57],administration of IL-2 stimulates the AINR cells to proliferate, and AINR is reversed so thatthe cells regain the ability to produce IL-2. Furthermore, we showed that CD4 T cells couldprovide the IL-2 needed to reverse AINR in CD8 T cells responding to syngeneic tumor [58].Thus, it appears that AINR constitutes a CD4 helper T cell-dependent checkpoint in the CD8T cell response; the effector CD8 T cells cannot continue to expand in number without IL-2that can be provided by helper T cells.
In cases where Ag is successfully cleared by the initial response, AINR is reversed over timeas some of the effector cells survive to become responsive memory cells. Results from Kaechet.al. [53] indicate that this occurs one to two weeks after the peak of the effector response. Atthe peak of the response to LCMV, the effector CD8 T cells were unable to make IL-2 orproliferate upon re-exposure to Ag, but the cells remaining two weeks later had regained thiscapacity. IL-7 plays a role in the transition of effector CD8 T cells to memory cells [59,60],and may be involved in reversal of AINR. We have found that in vitro culture of AINR CD8effector cells in IL-7 stimulates their proliferation, and the AINR state is reversed within oneto two days (Hammerbeck and Mescher, unpublished results). Thus, following Ag clearanceand re-expression of the IL-7Rα receptor on some of the effector CD8 T cells ([61,62] andHammerbeck and Mescher, manuscript submitted), the low levels of IL-7 present in vivo maystimulate slow proliferation of the effectors and reversal of AINR over one to two weeks. Incontrast to Ag clearance and memory formation, if the effector CTL resulting from the initialphase of clonal expansion fail to clear the Ag, tolerance to the persisting Ag results if CD4 T
Mescher et al. Page 7
Semin Immunol. Author manuscript; available in PMC 2007 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

cell help in the form of IL-2 is not available to reverse AINR. The numbers and functionalcapacity of the AINR CD8 T cells then decline over time. CD8 T cell responses limited byAINR could potentially be a mechanism that contributes to chronic graft rejection, with newthymic emigrants having specificity for the graft making an initial response and causing sometissue damage and inflammation, but being limited by the development of AINR and lack ofCD4 T cell help so that the graft is not acutely rejected. The same mechanism may also beinvolved in chronic autoimmune diseases where CD8 T cells can contribute to the disease.
The molecular basis for the AINR stateWhile classical CD4 T cell anergy differs from AINR in CD8 T cells with respect to the modeof induction, lack of costimulation versus full activation, the consequences are similar; the cellscan signal through the TCR to upregulate effector functions, but fail to produce IL-2 in responseto TCR and CD28 signaling. The first signaling defects characterized in anergic CD4 T cellswere blocks in activation of the ras-MAP-kinase pathway. Anergic CD4 T cell clones failedto activate p21 ras in response to TCR and CD28 engagement [63] with a resulting block inERK and JNK upregulation [64,65]. Activated ERK and JNK act synergistically totransactivate AP-1, a transcription factor required for IL-2 production [66]. A third member ofthe MAPK family, p38, has also been implicated as being necessary for IL-2 production [67].Defects in the ras-MAP-kinase pathway have also been demonstrated in CD8 T cells in theAINR state, although this has been much less extensively studied than in CD4 T cells.
Signaling in naïve and AINR CD8 T cells was studied in vitro using artificial APC having justpeptide-MHC Ag complex or anti-TCR mAb immobilized alone on microspheres to providesignal 1, or along with immobilized B7-1 to provide signals 1 and 2 [68]. ERK was activatedin response to just TCR engagement, and activation was increased little, if at all, when B7-1was present (Fig. 1). Signal 1 alone also significantly increased phosphorylation of p38, butthere was a substantial further increase when B7-1 was also present, as Zhang et.al. observedfor mouse splenic T cells [67]. In contrast, JNK was only activated in response to both TCRand CD28 signals, as is the case for CD4 T cells [65,69].
Defects in activation of all three of these MAPKs were seen in AINR CD8 T cells [68]. Initialactivation of ERK (1 min) was comparable in normal and AINR cells, but activity declinedmuch more rapidly in AINR cells. At 10 min, ERK activity remained high in normal cells, buthad declined to almost basal level in AINR cells. JNK activity was reduced by greater than50% in AINR cells at all time points, and p38 activation was strongly inhibited, with little orno phosphorylation detected at any time. Specific inhibitors of ERK and p38 (PD98059 andSB202190, respectively) caused a dosedependent decrease in IL-2 mRNA upregulation andproliferation of naïve CD8 T cells responding to TCR and CD28 signaling, suggesting thatreduced activation of these kinases in AINR cells accounts, at least in part, for their inabilityto respond.
Activation of p21ras, which is upstream in the MAPK signaling pathway, is defective in anergicCD4 T cells [63], and this is likely to also be the case for CD8 T cells in the AINR state.Treatment of AINR cells with phorbol myristic acetate (PMA), an activator of ras and PKC,and ionomycin, a calcium ionophore, activated ERK, JNK and p38 and stimulated IL-2production and proliferation by the cells, and the proliferation was inhibited by ERK and p38inhibitors [68]. These results suggest that cells in the AINR state are unable to activate p21 rasvia TCR and CD28 signaling, although this has not yet been directly demonstrated. Thus, itappears that the AINR state of CD8 T cells closely resembles that of anergic CD4 T cells withrespect to defects in activation of the MAPK signaling pathway.
Uhlin et.al. [70] have recently shown that TCR engagement by anergic human CD8 T cellsresults in rapid degradation of lck, a src-family protein kinase involved in proximal TCR
Mescher et al. Page 8
Semin Immunol. Author manuscript; available in PMC 2007 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

signaling. Inhibition of lck degradation, or transfection with an lck expression vector, restoredresponsiveness of the cells. Whether the cells examined in these experiments were in a statecomparable to the AINR state seen in activated primary murine CD8 T cells is unclear, sincedevelopment and maintenance of non-responsiveness in the human CD8 T cell lines was notaffected by the exogenous addition of IL-2. A role for degradation of components of the T cellsignaling pathway in CD4 T cell anergy has also emerged from results showing that a numberof E3 ubiquitin ligases have important roles in the induction and maintenance of anergy [71],as discussed elsewhere in this volume. Whether degradation of signaling pathway components,by ubiquitination-dependent pathways or other pathways, has a role in the development ormaintenance of the AINR state in primary CD8 T cells following an initial response to Ag andcostimulatory signals remains to be determined.
As mentioned above, the AINR state can be reversed by culturing cells for one to two days inthe presence of exogenous IL-2. Following reversal, the cells can again respond to Ag byupregulating mRNA expression and proliferating [51]. Furthermore, in contrast to naïve cells,the reversed cells do not require a costimulatory signal to respond; engagement of just the TCRis sufficient for IL-2 mRNA and protein upregulation, and IL-2-dependent proliferation tooccur. This appears to result from some ‘re-wiring’ of the signaling pathways. While naïvecells require both Ag and costimulation to activate JNK, and to optimally activate p38, bothare strongly upregulated in reversed cells in response to just TCR engagement, and B7-1 ligandmakes no further contribution (Fig. 1). As is the case for naïve cells, the effects ofpharmacological inhibitors indicate that both ERK and p38 contribute to IL-2 upregulation andproliferation in the reversed cells. The decreased dependence on costimulation following AINRreversal should allow the cells to more effectively continue to expand in response to Ag thatis not expressed on professional APC. Memory cells also exhibit decreased dependence oncostimulation, in comparison to naïve cells, raising the possibility that the ‘re-wiring’ thatoccurs upon reversal of AINR by IL-2 may also occur when AINR cells undergo transition tomemory cells when Ag is no longer present.
Studies of signaling defects in AINR cells have primarily examined cells responding in vitro,but there is some evidence for defective activation of the MAPK signaling pathway in cellsthat have become AINR following an in vivo response to virus. As mentioned above, Kaechet.al. [53] showed that effector CD8 T cells present at the peak of the response to LCMV (day8) are unable to make IL-2 or proliferate upon re-exposure to Ag, but regained responsivenessat later times. In this same study, it was shown that the effector cells at the peak of the responsewere defective in their ability to phosphorylate ERK in response to Ag, and thus activate theenzyme, and the ability to phosphorylate ERK increased in parallel with increased IL-2production and proliferation as the cells transitioned to memory.
Whether development of AINR in effector CD8 T cells is intrinsic to the differentiationprogram initiated by TCR, CD28 and IL-12/IFN-α/β, or is, instead, the result of negativesignaling through a surface receptor is unclear. CTLA-4 can deliver inhibitory signals uponbinding to its B7 ligands, but blocking this interaction does not prevent or reverse developmentof the AINR state in vitro [48]. However, programmed death (PD)-1, another inhibitorymember of the CD28 family, can negatively regulate T cell responses upon binding to itsligands PD-L1 and PD-L2. The PD-1 pathway mediates peripheral tolerance [72,73], and micelacking PD-1 expression develop spontaneous autoimmune diseases [74,75]. The propertiesof PD-1-mediated inhibition of T cell responses resemble in many respects those of the AINRstate. Barber et.al. [76] showed that blockade of the PD-1/PD-L1 pathway restoredresponsiveness of exhausted CD8 T cells present in mice chronically infected with LCMV.PD-1 mediated inhibition results in decreased IL-2 production and proliferation, but the cellsremain responsive to IL-2 and exogenous IL-2 can overcome the inhibition and augment IL-2production [77]. PD-1 expression is upregulated upon activation of CD8 T cells, and the
Mescher et al. Page 9
Semin Immunol. Author manuscript; available in PMC 2007 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

similarities of PD-1-mediated inhibition and the AINR state suggest that PD-1 may mediateAINR. In the in vitro studies examining development of AINR in purified CD8 T cells usingartificial APC, a PD-1 ligand was not present on the Ag-presenting surface [48,51,68].However, in addition to expressing PD-1, activated T cells express the PD-1 ligand PD-L1[78]. Thus, it is possible that PD-1 might mediate development of AINR under these conditionsalso, by acting in trans, i.e. by PD-1 on one T cell binding to PD-L1 on other cells in the culture.Whether this is the case remains to be determined.
4. ConclusionThe requirement for a third signal to support differentiation and memory development, and therequirement for IL-2 to sustain effector CTL expansion if Ag persists, represent two distinctcheckpoints in the CD8 T cell response (Fig. 4). CD4 T helper cells can provide the signalsnecessary for CD8 T cells to pass these checkpoints, by stimulating DC to produce signal 3cytokines, IL-12 and/or IFN-α/β, and by producing IL-2 to reverse AINR and allow continuedexpansion. In the absence of CD4 T cell help, the first checkpoint can still be passed if DCbecome activated through engagement of TLR or other surface receptors to produce signal 3cytokines. Whether alternatives pathways exist to pass the AINR checkpoint is not clear,although costimulatory receptors of the TNFR family (OX40, 4-1BB) may be able to stimulatethe AINR cells that are unresponsive to CD28-mediated costimulation.
These checkpoints may be the major means by which self-reactive CD8 T cells are preventedfrom productively responding to cause autoimmune diseases or transplant rejection. Encounterwith self-Ag in a non-inflammatory environment where DC are not activated will result ininitial proliferation, but tolerance will ensue in the absence of a signal 3 cytokine. However,the self-reactive cells may successfully pass this initial checkpoint if they encounter Ag in aninflammatory environment, such as could occur during an infection. In this regard, theemerging roles of IFN-α in directly signaling to T cells to skew CD4 cells to the Th1 pathwayand drive differentiation of Ag-activated CD8 T cells are particularly interesting. Theimportance of Type I IFN in the pathogenesis of autoimmunity has been appreciated for sometime, and recent evidence is implicating genetic alterations that lead to Type I IFNoverproduction as contributing to susceptibility to systemic lupus erythematosus and possiblyother autoimmune diseases (reviewed in [79,80]). Type I IFNs have a broad and diverse impacton both innate and adaptive immune responses, and affect many cell types, but direct signalingto skew and activate CD4 and CD8 T cells is likely to be among their important roles incontributing to autoimmune diseases.
Even when CD8 T cells receive a full set of signals for initial activation and differentiation,AINR limits the extent to which they can expand in the face of persisting Ag. A CD8 T cellresponse to self-Ag or transplanted tissue under conditions where a full set of initial signals isavailable, but help in the form of IL-2 produced by CD4 T cells is not available to reverseAINR, may contribute to chronic autoimmune disease states or transplant rejection. As theeffector CTL in the AINR state become exhausted, disease may wane, but may recur if newlyemerged self-reactive CD8 T cells again encounter Ag under stimulatory conditions.
Signaling by cytokines clearly plays a critical role in initiating and maintaining CD8 T cellresponses, and the Jaks and Stats involved in the signaling pathways employed by thesecytokines are potential targets for immunosuppressive agents to treat autoimmunity or preventtransplant rejection [81]. Stat4 would appear to provide a particularly good target, given itsimportant role in both skewing CD4 T cells to the Th1 pathway and signaling for differentiationof CD8 T cells.
Mescher et al. Page 10
Semin Immunol. Author manuscript; available in PMC 2007 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Acknowledgements
This work was supported by NIH grants RO1 AI34824 and PO1 AI35296.
References1. Anderson MS, Venanzi ES, Chen Z, Berzins SP, Benoist C, Mathis D. The Cellular Mechanism of
Aire Control of T Cell Tolerance. Immunity 2005;23(2):227–239. [PubMed: 16111640]2. Ohashi P, Oehen S, Buerki K, Pircher H, Ohashi C, Odermatt B, Malissen B, Zinkernagel R, Hengartner
H. Ablation of tolerance and induction of diabetes by virus infection in viral antigen transgenic mice.Cell 1991;65:305–17. [PubMed: 1901764]
3. Redmond WL, Sherman LA. Peripheral tolerance in CD8 T lymphocytes. Immunity 2005;22:275–284. [PubMed: 15780985]
4. Bevan MJ. Cross-priming for a secondary cytotoxic response to minor H antigens with H-2 congeniccells which do not cross-react in the cytotoxic assay. J Exp Med 1976;143:1283–1288. [PubMed:1083422]
5. Heath WR, Carbone FR. Cross-presentation, dendritic cells, tolerance and immunity. Annu RevImmunol 2001;19:47–64. [PubMed: 11244030]
6. Steinman RM, Hawiger D, Nussenzweig MC. Tolerogenic dendritic cells. Ann Rev Immunol2003;21:685–711. [PubMed: 12615891]
7. Bennett SRM, Carbone FR, Karamalis F, Flavell RA, Miller JFAP, Heath WR. Help for cytotoxic T-cell responses is mediated by CD40 signaling. Nature 1998;393:478–480. [PubMed: 9624004]
8. Ridge JP, DiRosa F, Matzinger P. A conditioned dendritic cell can be a temporal bridge between aCD4+ T-helper and a T-killer cell. Nature 1998;393:474–478. [PubMed: 9624003]
9. Schoenberger SP, Toes REM, van der Voort EIH, Offringa R, Melief CJM. T-cell help for cytotoxicT lymphocytes is mediated by CD40-CD40L interactions. Nature 1998;393:480–483. [PubMed:9624005]
10. Albert ML, Jegathesan M, Darnell RB. Dendritic cell maturation is required for the cross-tolerizationof CD8+ T cells. Nature Immunol 2001;2:1010–1017. [PubMed: 11590405]
11. Fujii, S-i; Liu, K.; Smith, C.; Bonito, AJ.; Steinman, RM. The Linkage of Innate to Adaptive Immunityvia Maturing Dendritic Cells In Vivo Requires CD40 Ligation in Addition to Antigen Presentationand CD80/86 Costimulation. J Exp Med 2004;199(12):1607–1618. [PubMed: 15197224]
12. Hernandez J, Aung S, Marquardt K, Sherman LA. Uncoupling of proliferative potential and gain ofeffector function by CD8(+) T cells responding to self-antigens. J Exp Med 2002;196(3):323–33.[PubMed: 12163561]
13. Allison JP. CD28-B7 interactions in T cell activation. Curr Opin Immunol 1994;6:414–419. [PubMed:7522466]
14. Janeway CA, Bottomly K. Signals and signs for lymphocyte responses. Cell 1994;76:275–285.[PubMed: 7904901]
15. Lenschow DJ, Walunas TL, Bluestone JA. CD28/B7 system of T cell costimulation. Ann RevImmunol 1996;14:233. [PubMed: 8717514]
16. Mueller D, Jenkins M, Schwartz R. Clonal expansion vs functional clonal inactivation. Ann RevImmunol 1989;7:445–480. [PubMed: 2653373]
17. Curtsinger JM, Schmidt CS, Mondino A, Lins DC, Kedl RM, Jenkins MK, Mescher MF.Inflammatory cytokines provide third signals for activation of naive CD4+ and CD8+ T cells. JImmunol 1999;162:3256–3262. [PubMed: 10092777]
18. Curtsinger JM, Valenzuela JO, Agarwal P, Lins DC, Mescher MF. Cutting edge: Type I interferonsprovide a third signal to CD8 T cells to stimulate clonal expansion and differentiation. J ImmunolCE. 2005in press
19. Gately MK, Wolitzky AG, Quinn PM, Chizzonite R. Regulation of human cytolytic lymphocyteresponses by IL-12. Cell Immunol 1992;143:127–142. [PubMed: 1352483]
20. Trinchieri G. Interleukin-12: a cytokine at the interface of inflammation and immunity. Adv Immunol1998;70:83. [PubMed: 9755338]
Mescher et al. Page 11
Semin Immunol. Author manuscript; available in PMC 2007 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

21. Marrack P, Kappler J, Mitchell T. Type I interferons keep activated T cells alive. J Exp Med 1999;189(3):521–30. [PubMed: 9927514]
22. Nguyen KB, Watford WT, Salomon R, Hofmann SR, Pien GC, Morinobu A, Gadina M, O’Shea JJ,Biron CA. Critical role for STAT4 activation by type 1 interferons in the interferon-gamma responseto viral infection. Science 2002;297(5589):2063–6. [PubMed: 12242445]
23. Aichele P, Brduscha-Reim K, Zinkernagel R, Hengartner H, Pircher H. T cell priming versus T celltolerance induced by synthetic peptides. J Exp Med 1995;182:261–66. [PubMed: 7540654]
24. Aichele P, Kyburz D, Ohashi PS, Odermatt B, Zinkernagel RM, Hengartner H, Pircher H. Peptide-induced T-cell tolerance to prevent autoimmune diabetes in a transgenic mouse model. Proc NatlAcad Sci USA 1994;91:444. [PubMed: 8290546]
25. Kyburz D, Aichele P, Speiser D, Hengartner H, Zinkernagel R, Pircher H. T cell immunity after aviral infection versus T cell tolerance induced by soluble viral peptides. Eur J Immunol1993;23:1956–62. [PubMed: 8344359]
26. Curtsinger JM, Lins DC, Mescher MF. Signal 3 determines tolerance versus full activation of naiveCD8 T cells: dissociating proliferation and development of effector function. J Exp Med2003;197:1141–1151. [PubMed: 12732656]
27. Schmidt CS, Mescher MF. Adjuvant effect of IL-12: conversion of peptide antigen administrationfrom tolerizing to immunizing for CD8+ T cells in vivo. J Immunol 1999;163:2561–2567. [PubMed:10452994]
28. Aichele P, Unsoeld H, Koshella M, Schweier O, Kalinke U, Vucikuja S. Cutting Edge: CD8 T cellsspecific for lymphocytic choriomeningitis virus require Type I IFN receptor for clonal expansion. JImmunol 2006;176:4525–4529. [PubMed: 16585541]
29. Kolumam GA, Thomas S, Thompson LJ, Sprent J, Murali-Krishna K. Type I interferons act directlyon CD8 T cells to allow clonal expansion and memory formation in response to viral infection. J ExpMed 2005;202:637–650. [PubMed: 16129706]
30. Le Bon A, Durand V, Kamphuis E, Thompson C, Bulfone-Paus S, Rossmann C, Kalinke U, ToughDF. Direct stimulation of T cells by Type I IFN enhances the CD8+ T cell response during cross-priming. J Immunol 2006;176:4682–4689. [PubMed: 16585561]
31. Thompson LJ, Kolumam GA, Thomas S, Murali-Krishna K. Innate inflammatory signals induced byvarious pathogens differentially dictate the IFN-I dependence of CD8 T cells for clonal expansionand memory formation. J Immunol 2006;177:1746–1754. [PubMed: 16849484]
32. Cella M, Scheidegger D, Palmer-Lehmann K, Lane P, Lanzavecchia A, Alber G. Ligation of CD40on dendritic cells triggers production of high levels of interleukin-12 and enhances T cell stimulatorycapacity: T-T help via APC activation. J Exp Med 1996;184(2):747–52. [PubMed: 8760829]
33. Filatenkov AA, Jacovetty EL, Fischer UB, Curtsinger JM, Mescher MF, Ingulli E. CD4 T cell-dependent conditioning of dendritic cells to produce IL-12 results in CD8-mediated graft rejection.J Immunol. 2005in press
34. van Stipdonk MJ, Lemmens EE, Schoenberger SP. Naive CTLs require a single brief period ofantigenic stimulation for clonal expansion and differentiation. Nat Immunol 2001;2(5):423–9.[PubMed: 11323696]
35. Curtsinger JM, Johnson CM, Mescher MF. CD8 T cell clonal expansion and development of effectorfunction require prolonged exposure to antigen, costimulation, and signal 3 cytokine. J Immunol2003;171:5165–5171. [PubMed: 14607916]
36. Kaplan MH, Sun YL, Hoey T, Grusby MJ. Impaired IL-12 responses and enhanced development ofTh2 cells in Stat4-deficient mice. Nature 1996;382:174–177. [PubMed: 8700209]
37. Thierfelder WE, van Deursen JM, Yamamoto K, Tripp RA, Sarawar SR, Carson RT, Sangster MY,Vignali DA, Doherty PC, Grosveld GC, et al. Requirement for Stat4 in interleukin-12-mediatedresposes of natural killer cells and T cells. Nature 1996;382:171–174. [PubMed: 8700208]
38. Murphy TL, Geissal ED, Farrar JD, Murphy KM. Role of the Stat4 N domain in recptor proximaltyrosine phosphorylation. Mol Cell Biol 2000;20:7121–7131. [PubMed: 10982828]
39. Croft M, Carter L, Swain SL, Dutton RW. Generation of polarized antigen-specific CD8 effectorpopulations: reciprocal action of interleukin (IL)-4 and IL-12 in promoting type 2 versus type 1cytokine profiles. J Exp Med 1994;180:1715–1728. [PubMed: 7525836]
Mescher et al. Page 12
Semin Immunol. Author manuscript; available in PMC 2007 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

40. Haring JS, Badovinac VP, Harty JT. Inflaming the CD8+ T cell response. Immunity 2006;25:19–29.[PubMed: 16860754]
41. Badovinac VP, Tvinnereim AR, Harty JT. Regulation of antigen-specific CD8+ T cell homeostasisby perforin and interferon-gamma. Science 2000;290:1354–1358. [PubMed: 11082062]
42. Whitmire JK, Tan JT, Whitton JL. IFN-gamma acts directly on CD8+ T cells to increase theirabundance during virus infection. J Exp Med 2005;201:1053–1059. [PubMed: 15809350]
43. Sercan O, Hammerling GJ, Arnold B, Schuler T. Innate immune cells contribute to the IFN-gamma-dependent regulation of antigen-specific CD8+ T cell homeostasis. J Immunol 2006;176:735–739.[PubMed: 16393956]
44. Parrish-Novak J, Dillon SR, Nelson A, Hammond A, Sprecher C, Gross JA, Johnston J, Madden K,Xu W, West J, et al. Interleukin 21 and its receptor are involved in NK cell expansion and regulationof lymphocyte function. Nature 2000;408(6808):57–63. [PubMed: 11081504]
45. Valenzuela J, Hammerbeck CD, Mescher MF. Cutting Edge: Bcl-3 up-regulation by signal 3 cytokine(IL-12) prolongs survival of antigen-activated CD8 T cells. J Immunol 2005;174:600–604. [PubMed:15634875]
46. Rocha B, Grandien A, Freitas A. Anergy and exhaustion are independent mechanisms of peripheralT cell tolerance. J Exp Med 1995;181(3):993–1003. [PubMed: 7869056]
47. Zajac AJ, Blattman JN, Murali-Krishna K, Sourdive DJD, Suresh M, Altman JD, Ahmed R. Viralimmune evasion due to persistence of activated T cells without effector function. J Exp Med1999;188:2205–2213. [PubMed: 9858507]
48. Deeths MJ, Kedl RM, Mescher MF. CD8+ T cells become nonresponsive (anergic) followingactivation in the presence of costimulation. J Immunol 1999;163(1):102–10. [PubMed: 10384105]
49. Jenkins M, Schwartz R. Antigen presentation by chemically modified splenocytes induces antigen-specific T cell unresponsiveness in vitro and in vivo. J Exp Med 1987;165:302–319. [PubMed:3029267]
50. Mueller D, Jenkins M. Molecular mechanisms underlying function T-cell unresponsiveness. CurrentOpinion in Immunol 1995;7:375–81.
51. Tham EL, Shrikant P, Mescher MF. Activation-induced nonresponsiveness: A Th-dependentregulatory checkpoint in the CTL response. J Immunol 2002;168:1190–1197. [PubMed: 11801654]
52. Otten G, Germain R. Split anergy in a CD8+ T cell: Receptor-dependent cytolysis in the absence ofinterleukin-2 production. Science 1991;251:1228–1231. [PubMed: 1900952]
53. Kaech SM, Hemby S, Kersh E, Ahmed R. Molecular and Functional Profiling of Memory CD8 TCell Differentiation. Cell 2002;111(6):837–851. [PubMed: 12526810]
54. Shrikant P, Mescher MF. Control of syngeneic tumor growth by activation of CD8+ T cells: efficacyis limited by migration away from the site and induction of nonresponsiveness. J Immunol1999;162:2858–2866. [PubMed: 10072534]
55. Wherry EJ, Barber DL, Kaech SM, Blattman JN, Ahmed R. Antigen-independent memory CD8 Tcells do not develop during chronic viral infection. PNAS 2004;101(45):16004–16009. [PubMed:15505208]
56. Shrikant P, Mescher MF. Opposing effects of interleukin-2 in tumor immunotherapy: promoting CD8T cell growth and inducing apoptosis. J Immunol 2002;169:1753–1759. [PubMed: 12165496]
57. Blattman JN, Grayson JM, Wherry EJ, Kaech SM, Smith KA, Ahmed R. Therapeutic use of IL-2 toenhance antiviral T-cell responses in vivo. Nat Med 2003;9(5):540–547. [PubMed: 12692546]
58. Shrikant P, Khoruts A, Mescher MF. CTLA-4 blockade reverses CD8+ T cell tolerance to tumor bya CD4+ T cell and IL-2-dependent mechanism. Immunity 1999;11:483–493. [PubMed: 10549630]
59. Buentke E, Mathiot M, Tolaini M, Di Santo J, Zamoyska R, Seddon B. Do CD8 effector T cells needIL-7R expression to become resting memory cells? Blood 2006;108:1949–1956. [PubMed:16705084]
60. Prlic M, Lefrancois L, Jameson SC. Multiple choices: regulation of memory CD8 T cell generationand homeostasis by interleukin (IL)-7 and IL-15. J Exp Med 2002;195:F49–F52. [PubMed:12070294]
61. Kaech SM, Tan JT, Wherry EJ, Konieczny BT, Surh CD, Ahmed R. Selective expression of theinterleukin 7 receptor identifies effector CD8 T cells that give rise to long-lived memory cells. NatImmunol 2003;4(12):1191–8. [PubMed: 14625547]
Mescher et al. Page 13
Semin Immunol. Author manuscript; available in PMC 2007 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

62. Lacombe MH, Hardy MP, Rooney J, Labrecque N. IL-7 receptor expression levels do not identifyCD8 memory T lymphocyte precursors following peptide immunization. J Immunol 2005;175:4400–4407. [PubMed: 16177081]
63. Fields PE, Gajewski TF, Fitch FW. Blocked Ras activation in anergic CD4+ T cells. Science1996;271:1276–1278. [PubMed: 8638108]
64. DeSilva DR, Feeser WS, Tancula EJ, Scherle PA. Anergic T cells are defective in both jun NH2-terminal kinase and mitogen-activated protein kinase signaling pathways. J Exp Med 1996;183(5):2017–23. [PubMed: 8642312]
65. Li W, Whaley CD, Mondino A, Mueller DL. Blocked signal transduction to the ERK and JNK proteinkinases in anergic CD4+ T cells. Science 1996;271:1272–1276. [PubMed: 8638107]
66. Kang SM, Beverly B, Tran AC, Brorson K, Schwartz RH, Lenardo MJ. Transactivation by AP-1 isa molecular target of T cell clonal anergy. Science 1992;21:1134–1138. [PubMed: 1509265]
67. Zhang J, Salojin KV, Gao JX, Cameron MJ, Bergerot I, Delovitch TL. p38 mitogen-activated proteinkinase mediates signal integration of TCR/CD28 costimulation in primary murine T cells. J Immunol1999;162:3819–3829. [PubMed: 10201899]
68. Tham EL, Mescher MF. Signaling alterations in activation-induced non-responsive (AINR) CD8 Tcells. J Immunol 2001;167:2040–2048. [PubMed: 11489986]
69. Su B, Jacinto E, Hibi M, Kallunki T, Karin M, Ben-Neriah Y. JNK is involved in signal integrationduring costimulation of T lymphocytes. Cell 1994;77:727–736. [PubMed: 8205621]
70. Uhlin M, Masucci MG, Levitsky V. Regulation of lck degradation and refractory state in CD8+cytotoxic T lymphocytes. Proc Natl Acad Sci U S A 2005;102:9264–9269. [PubMed: 15958529]
71. Mueller DL. E3 ubiquitin ligases as T cell anergy factors. Nat Immunol 2004;5:883–890. [PubMed:15334084]
72. Greenwald RJ, Freeman GJ, Sharpe AH. The B7 family revisited. Ann Rev Immunol 2005;23:515–548. [PubMed: 15771580]
73. Okazaki T, Honjo T. The PD-1-PD-L pathway in immunological tolerance. Trends Immunol2006;27:195–201. [PubMed: 16500147]
74. Nishimura H, Nose M, Hiai H, Minato N, Honjo T. Development of lupus-like autoimmune diseasesby disruption of the PD-1 gene encoding an ITIM motif-carrying immunoreceptor. Immunity1999;11:141–151. [PubMed: 10485649]
75. Nishimura H, Okazaki T, Tanaka Y, Nakatani K, Hara M, Matsumori A, Sasayama S, Mizoguchi A,Hiai H, Minato N, et al. Autoimmune dilated cardiomyopathy in PD-1 receptor-deficient mice.Science 2001;291:319–322. [PubMed: 11209085]
76. Barber DL, Wherry EJ, Masopust D, Zhu B, Allison JP, Sharpe AH, Freeman GJ, Ahmed R. Restoringfunction in exhausted CD8 T cells during chronic viral infection. Nature 2006;439:682–687.[PubMed: 16382236]
77. Carter LL, Fouser LA, Jussif J, Fitz L, Deng B, Wood CR, Collins M, Honjo T, Freeman GJ, CarrenoBM. PD-1:PD-L inhibitory pathway affects both CD4+ and CD8+ T cells and is overcome by IL-2.Eur J Immunol 2002;32:634–643. [PubMed: 11857337]
78. Lactchman YE, Liang SC, Wu Y, Chernova T, Sobel RA, Klemm M, Kuchroo VK, Freeman GJ,Sharpe AH. PD-L1-deficient mice show that PD-L1 on T cells, antigen-presenting cells, and hosttissues negatively regulates T cells. Proc Natl Acad Sci U S A 2004;101:10691–10696. [PubMed:15249675]
79. Banchereau J, Pascual V. Type I interferon in Systemic Lupus erythematosus and other autoimmunediseases. Immunity 2006;25:383–392. [PubMed: 16979570]
80. Theofilopoulos AN, Baccala R, Beutler B, Kono DH. Type I interferons (alpha/beta) in immunityand autoimmunity. Ann Rev Immunol 2005;23:307–336. [PubMed: 15771573]
81. O’Shea JJ, Park H, Pesu M, Borie D, Changelian P. New strategies for immunosuppression: interferingwith cytokines by targeting the Jak/Stat pathway. Curr Op Rheumatol 2005;17:305–311.
Mescher et al. Page 14
Semin Immunol. Author manuscript; available in PMC 2007 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
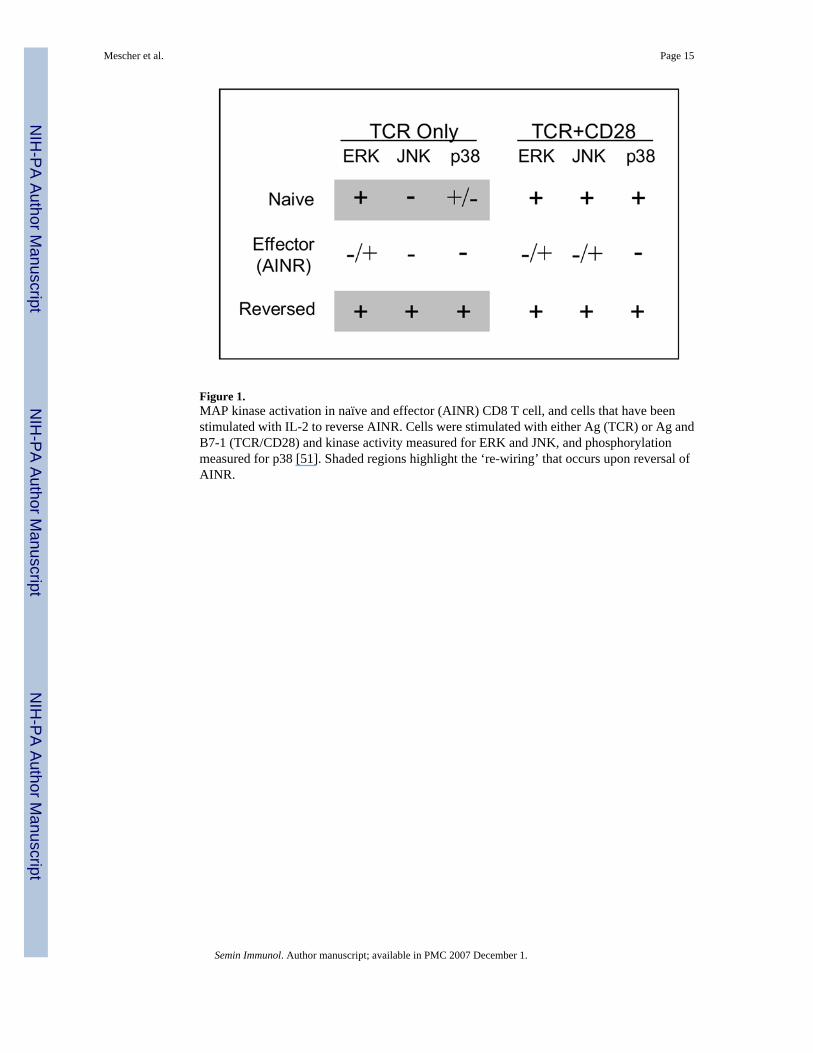
Figure 1.MAP kinase activation in naïve and effector (AINR) CD8 T cell, and cells that have beenstimulated with IL-2 to reverse AINR. Cells were stimulated with either Ag (TCR) or Ag andB7-1 (TCR/CD28) and kinase activity measured for ERK and JNK, and phosphorylationmeasured for p38 [51]. Shaded regions highlight the ‘re-wiring’ that occurs upon reversal ofAINR.
Mescher et al. Page 15
Semin Immunol. Author manuscript; available in PMC 2007 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
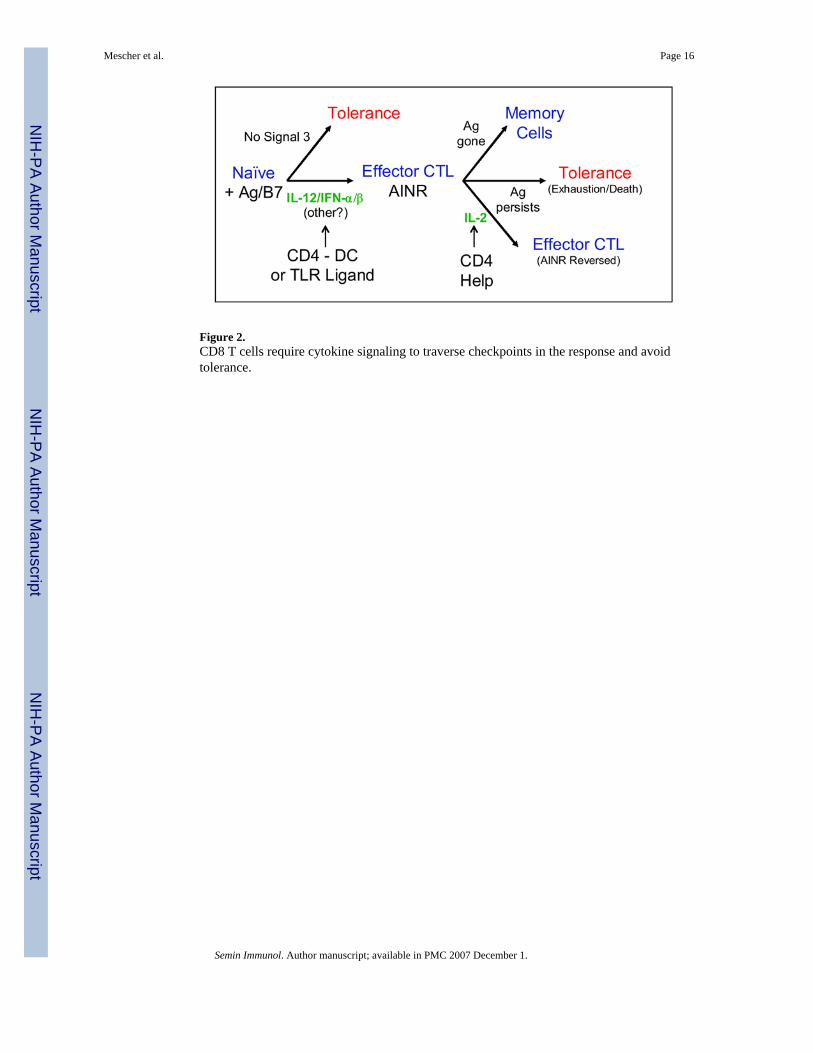
Figure 2.CD8 T cells require cytokine signaling to traverse checkpoints in the response and avoidtolerance.
Mescher et al. Page 16
Semin Immunol. Author manuscript; available in PMC 2007 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript




















