
That Produce TRAIL T Cells + Generating Helpless CD8 Apoptotic Cells Induce Tolerance by
10
of February 18, 2015. This information is current as Produce TRAIL T Cells That + Generating Helpless CD8 Apoptotic Cells Induce Tolerance by R. Green and Thomas A. Ferguson VanOosten, James K. Earle, Jr., John M. Herndon, Douglas Thomas S. Griffith, Hirotaka Kazama, Rebecca L. http://www.jimmunol.org/content/178/5/2679 doi: 10.4049/jimmunol.178.5.2679 2007; 178:2679-2687; ; J Immunol References http://www.jimmunol.org/content/178/5/2679.full#ref-list-1 , 26 of which you can access for free at: cites 59 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2007 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on February 18, 2015 http://www.jimmunol.org/ Downloaded from by guest on February 18, 2015 http://www.jimmunol.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of That Produce TRAIL T Cells + Generating Helpless CD8 Apoptotic Cells Induce Tolerance by

of February 18, 2015.This information is current as
Produce TRAIL T Cells That+Generating Helpless CD8
Apoptotic Cells Induce Tolerance by
R. Green and Thomas A. FergusonVanOosten, James K. Earle, Jr., John M. Herndon, Douglas Thomas S. Griffith, Hirotaka Kazama, Rebecca L.
http://www.jimmunol.org/content/178/5/2679doi: 10.4049/jimmunol.178.5.2679
2007; 178:2679-2687; ;J Immunol
Referenceshttp://www.jimmunol.org/content/178/5/2679.full#ref-list-1
, 26 of which you can access for free at: cites 59 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2007 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on February 18, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on February 18, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

Apoptotic Cells Induce Tolerance by Generating HelplessCD8� T Cells That Produce TRAIL1
Thomas S. Griffith,* Hirotaka Kazama,† Rebecca L. VanOosten,* James K. Earle, Jr.,*John M. Herndon,† Douglas R. Green,‡ and Thomas A. Ferguson2†
The decision to generate a productive immune response or immune tolerance following pathogenic insult often depends on thecontext in which T cells first encounter Ag. The presence of apoptotic cells favors the induction of tolerance, whereas immuneresponses generated with necrotic cells promote immunity. We have examined the tolerance induced by injection of apoptotic cells,a system in which cross-presentation of Ag associated with the dead cells induces CD8� regulatory (or suppressor) T cells. Weobserved that haptenated apoptotic cells induced CD8� suppressor T cells without priming CD4� T cells for immunity. TheseCD8� T cells transferred unresponsiveness to naive recipients. In contrast, haptenated necrotic cells stimulated immunity, butinduced CD8� suppressor T cells when CD4� T cells were absent. We further found that CD8� T cells induced by these treatmentsdisplayed a “helpless CTL” phenotype and suppress the immune response by producing TRAIL. Animals deficient in TRAIL wereresistant to tolerance induction by apoptotic cells. Thus, the outcome of an immune response taking place in the presence of celldeath can be determined by the presence of CD4�-mediated Th cell function. The Journal of Immunology, 2007, 178: 2679–2687.
D ead cells arise during normal tissue turnover or frompathogenic conditions, and the immune system mustdeal with these cells without the induction of autoim-
munity or suppression of productive immune responses. Althoughcells can be induced to die by various methods (e.g., death recep-tor, toxicity, etc.), there are generally two types of cell death–apoptotic and necrotic (1–3). Apoptosis is a highly regulated pro-cess induced by specific stimuli; apoptotic cells tend to induce atolerogenic or immunosuppressive response. Necrosis, conversely,results from the failure to control cellular homeostasis after injuryand can result in stimulation of immunity. A number of recentstudies have compared the effects of apoptotic and necrotic cellsand the consensus is that dendritic cells (DC)3 are central playersin determining the immunological outcome. Apoptotic cells canstimulate T cells without the benefit of DC maturation (and co-stimulation), leading to immune tolerance (4, 5). Necrotic cellsrelease proinflammatory molecules, such as high mobility groupbox protein 1 (6), uric acid (7), or heat shock protein 70 (8), which
can stimulate the DC to mature, leading to immunity. Mature DC,for example, stimulate CD4� cells to provide help for CD8� Tcells, encouraging their expansion into effector cells (5, 9, 10).
Although apoptotic cells can induce tolerance by deletionalmechanisms (11), recent studies suggest that tolerance followingpresentation of apoptotic cells is “infectious” and mediated by reg-ulatory T cells (Treg) (12). The phenomenon of infectious immu-nological tolerance was described over 30 years ago in Ag-mod-ified self and heterologous erythrocyte models (13–15). The ideathat T cells can regulate (or suppress) immunity unfortunatelynever progressed beyond the phenomenological stage, as the cellsand factors proposed to mediate tolerance escaped molecular char-acterization. Interestingly, many of the earliest descriptions of in-fectious tolerance described the function of CD8� regulatory Tcells (henceforth called T suppressor or Ts cells); however, withthe exception of a few studies with T cell vaccination (16, 17) andautoimmunity (18), little is known about the function of the CD8�
Ts cells. Mechanisms proposed to account for CD8� T cell sup-pression have been regulatory cytokines (IL-10) (19), as well asCD8 immune deviation (20). With the availability of new technol-ogy, there has been a renewed interest in T cell suppression, evi-denced by the many studies focused on the ability of CD4� Tregto suppress immunity in multiple systems (21).
Our investigation into the molecular mechanisms of infectioustolerance uses a classical tolerance system of presenting hapten-modified spleen cells to the immune system. Injection of thesecells i.v. leads to their apoptosis (via Fas/FasL), entry into thecross-priming pathway of the CD8�� DC, and presentation toCD8� T cells (12). The induced CD8� T cells are capable oftransferring tolerance to nontolerant individuals (i.e., infectioustolerance). In the current study, we explored the stimulation ofCD8� T cells with apoptotic cells and necrotic cells. Our resultsdemonstrate that haptenated apoptotic cells stimulate CD8� Tcells, which upon restimulation secrete TRAIL (22, 23) as an effectormolecule in the tolerance pathway. Apoptotic cells do not stimulateTh cell function. Interestingly, necrotic cells, which can stimulateCD4� T cell-mediated immunity, do not induce TRAIL productionby CD8� T cells unless the Ag is presented in the absence of CD4�
*Department of Urology and Interdisciplinary Graduate Program in Immunology,University of Iowa, Iowa City, IA 52242; †Department of Ophthalmology and VisualSciences, Washington University School of Medicine, St. Louis, MO 63110; and‡Department of Immunology, St. Jude Children’s Hospital, Memphis, TN 38105
Received for publication September 28, 2006. Accepted for publication December8, 2006.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by National Institutes of Health Grants CA109446 (toT.S.G.), EY06765 (to T.A.F.), EY015570 (to T.A.F.), and AI44848 (to D.R.G.), andthe Department of Ophthalmology and Visual Sciences Core Grant (EY08972). Sup-port was also received from the Foundation for Fighting Blindness (Owings Mills,MD) and from Research to Prevent Blindness (New York, NY).2 Address correspondence and reprint requests to Dr. Thomas A. Ferguson, Depart-ment of Ophthalmology and Visual Sciences, Washington University School of Med-icine, 660 South Euclid, Box 8096, St. Louis, MO 63110. E-mail address: [email protected] Abbreviations used in this paper: DC, dendritic cell; Ts, T suppressor cell; Treg,regulatory T cell; TNBS, 2,4,6 trinitrobenzene sulfonic acid; TNP, trinitrophenyl;TNP-spl, TNP-coupled spleen; AICD, activation-induced cell death.
Copyright © 2007 by The American Association of Immunologists, Inc. 0022-1767/07/$2.00
The Journal of Immunology
www.jimmunol.org
by guest on February 18, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
T cells. Thus, the response of the immune system after encounteringdead cells depends on the stimulation of CD4� T cell help.
Materials and MethodsAnimals
C57BL/6 mice were purchased from The Jackson Laboratory. C57BL/6TRAIL�/� mice were obtained from Amgen (24) and bred in our ownfacility at the University of Iowa (UI), according to UI Institutional AnimalCare and Use Committee (IACUC) guidelines. They are �10 generationsbackcrossed to C57BL/6. All animal procedures were performed accordingto National Institutes of Health guidelines and approved by the UI orWashington University IACUC.
Abs and reagents
Anti-CD4 (GK1.5) and anti-CD8 (clone 2.43) Ab for in vivo deletion werepurified from hybridoma supernatants. Mouse anti-TRAIL Ab (N2B2) andthe control mouse IgG2a were purchased from eBioscience. 2,4,6 trinitro-benzene sulfonic acid (TNBS) was purchased from Sigma-Aldrich. Re-combinant mouse TRAIL (rTRAIL) and zVAD-fmk were purchased fromBIOMOL. Anti-CD40 (FGK45) has been described (25) and mice receiveda single i.v. injection of 300 �g of FGK45.
Trinitrophenyl (TNP) coupling of spleen cells
Spleen cells were isolated and coupled with TNP as previously described(12). Briefly, 108 cells were incubated in 0.5 ml of HBSS and 0.5 ml of 10mM TNBS for 7–10 min at room temperature. After incubation, cells werewashed three times with HBSS before use. Cells were irradiated (3000 R)to induce apoptosis. Necrotic cells were prepared by alternating five freeze/thaw cycles using a liquid nitrogen/37°C water bath. TNP-coupled cellswere injected at 107 cells (apoptotic) or equivalence (necrotic) per mouse.All cells were delivered i.v. via the retro-orbital plexus.
Immune response
Mice were immunized with 0.1 ml of 10 mM TNBS s.c. Four days later,mice were challenged with 0.033 ml of 10 mM TNBS in PBS in the rightfootpad and 0.033 ml of PBS in the left footpad. Measurements were taken24 h postinjection by a masked observer. Values are expressed in microme-ters (�SE) and represent the difference between the right footpad (Agchallenge) and the left footpad (PBS challenge). Background values rep-resent the difference between the challenged and unchallenged footpad inunimmunized mice.
T cell isolation
T cell populations were obtained using Easy Sep Mouse CD4/CD8 T Cellenrichment kit(s) CD4/CD8 (Stem Cell Technologies) as per the manufac-turer’s instructions. Cell populations were used only when purity was�95% as determined by flow cytometry using anti-CD4 and anti-CD8 Abs.
Preparation of immune and tolerant supernatants
Mice were injected with TNP-apoptotic cells i.v. and immunized with 0.1ml of 10 mM TNBS s.c. 7 days later. Spleen cells were then isolated 1 daylater, cultured in vitro for 48 h in RPMI 1640 medium with 10% FBS.Supernatants from tolerant mice or immune mice were then mixed with4-day immune CD4� cells for 1 h. Following washing, the CD4� T cellswere transferred to naive mice that were challenged immediately. In somegroups, 1 �g of anti-TRAIL Ab (N2B2) or 1 �g of isotype control IgG2awas added to the tolerant supernatant.
Adoptive transfer of immunity
Mice were immunized with 0.1 ml of 10 mM TNBS. Four days later,spleens were harvested, erythrocyte-free suspensions were prepared (12),and the CD4� T cells were negatively selected and purified using the StemCell Technologies kit. Ten million CD4� cells per mouse were infused intonaive recipients. In some experiments, CD4� cells were incubated withimmune supernatant, tolerant supernatant, zVAD-fmk, or rTRAIL beforetransfer. Mice were immediately challenged and measurements were takenas described above.
Quantitative RT-PCR
Total RNA was harvested from cell populations with TRIzol reagent (In-vitrogen Life Technologies). Total RNA (2 �g) was reverse-transcribedusing Superscript II. The real-time quantitative RT-PCR primer/probe setsfor mouse TRAIL, FasL, TNF, FLIP, Bcl-xL, DR5, and rRNA were pur-
chased from Applied Biosystems. A total of 250 ng of cDNA was used asa template for TaqMan assays for all transcripts and the internal rRNAcontrol. The TaqMan PCR was conducted as described previously (26).
In vivo cytotoxicity
TNP-coupled spleen (TNP-spl) or normal spleen cells (spl) were suspendedat 1 � 107 cells/ml in warmed PBS/0.1% BSA. CFSE (1.25 �l/ml of 5 mMstock) was added to the TNP-spl for the CFSEhigh and a 10-fold dilutionwas used for the CFSElow population (reference population). Cells wereincubated 10 min at 37°C (water bath). The reaction was stopped by theaddition of ice-cold PBS containing 10% FBS. Cells were washed threetimes with PBS/FBS, counted, and resuspended to the appropriate vol-ume. Ten million target cells (CFSEhigh) and 10 million reference cells(CFSElow) were injected into naive or tolerant C57BL/6 or TRAIL�/�
recipients. Spleens were harvested 18 h later and analyzed by flowcytometry. A total of 3–5000 events in the reference population werecollected and the number of target cells recovered was enumerated.Unimmunized C57BL/6 mice were used as controls. The percent re-duction in the number of recovered CFSEhigh cells (TNP-spl) in theunimmunized vs the tolerant mice was considered the percent cytotox-icity. Individual mice were analyzed and the percent killing in eachgroup of mice was determined by the formula 100 � ([(% TNP-spltolerant/% spl tolerant)/(% TNP-spl unimmunized/% spl unimmu-nized)] � 100).
Statistical analysis
Significant differences between groups were evaluated using a two-tailedStudent’s t test ( p � 0.01).
ResultsNecrotic, but not apoptotic, haptenated cells prime for immunity
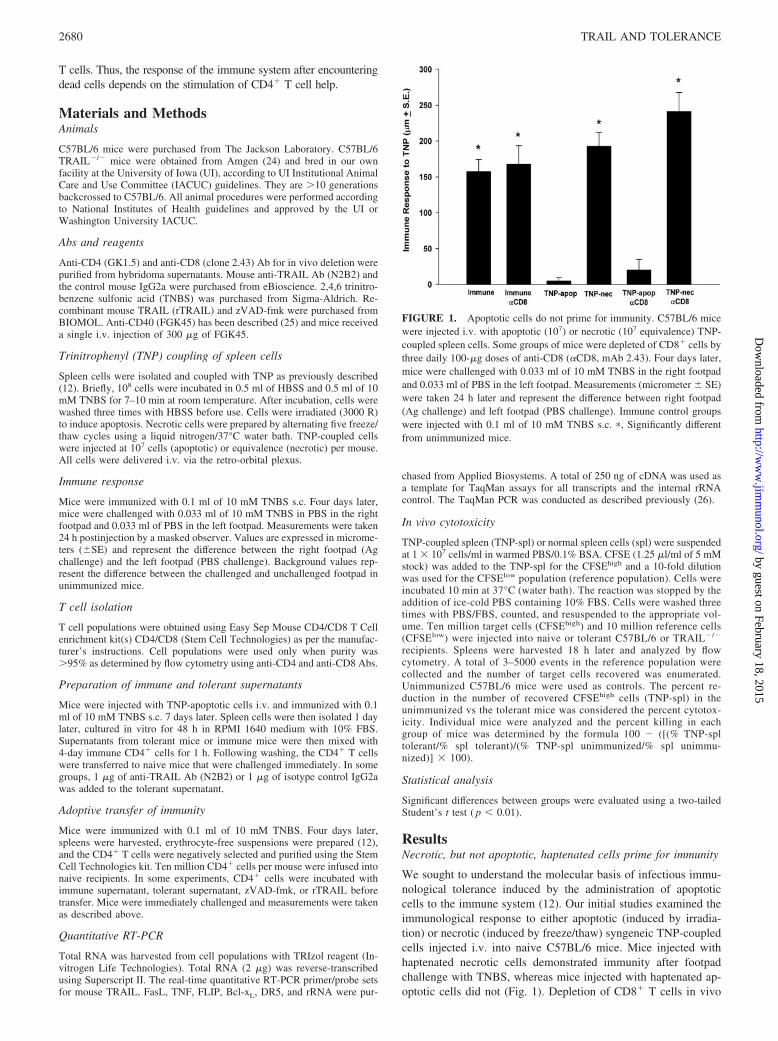
We sought to understand the molecular basis of infectious immu-nological tolerance induced by the administration of apoptoticcells to the immune system (12). Our initial studies examined theimmunological response to either apoptotic (induced by irradia-tion) or necrotic (induced by freeze/thaw) syngeneic TNP-coupledcells injected i.v. into naive C57BL/6 mice. Mice injected withhaptenated necrotic cells demonstrated immunity after footpadchallenge with TNBS, whereas mice injected with haptenated ap-optotic cells did not (Fig. 1). Depletion of CD8� T cells in vivo
FIGURE 1. Apoptotic cells do not prime for immunity. C57BL/6 micewere injected i.v. with apoptotic (107) or necrotic (107 equivalence) TNP-coupled spleen cells. Some groups of mice were depleted of CD8� cells bythree daily 100-�g doses of anti-CD8 (�CD8, mAb 2.43). Four days later,mice were challenged with 0.033 ml of 10 mM TNBS in the right footpadand 0.033 ml of PBS in the left footpad. Measurements (micrometer � SE)were taken 24 h later and represent the difference between right footpad(Ag challenge) and left footpad (PBS challenge). Immune control groupswere injected with 0.1 ml of 10 mM TNBS s.c. �, Significantly differentfrom unimmunized mice.
2680 TRAIL AND TOLERANCE
by guest on February 18, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

before immunization did not affect these results. These data showthat apoptotic cells do not prime for CD4-mediated immunitywhile necrotic cells are able to induce this type of immuneresponse.
CD8� T cells transfer tolerance induced by haptenatedapoptotic cells
We next determined whether injection of apoptotic cells led to thegeneration of the CD8� T cells that could regulate the immuneresponse. Haptenated apoptotic or necrotic cells were injected intountreated or CD4� T cell-depleted naive C57BL/6 mice. After 7days, CD8� T cells were isolated from the spleens of these miceand injected into naive C57BL/6 mice. These animals were im-
mediately immunized with TNBS, challenged after 4 days, andimmunity was measured 1 day later. The transfer of CD8� T cellsfrom mice that had received haptenated apoptotic cells made re-cipients unresponsive to Ag, while those receiving CD8� T cellsfrom mice that had received haptenated necrotic cells were readilyimmunized (Fig. 2A). The removal of CD4� T cells before in-jection of haptenated apoptotic cells did not affect the genera-tion of these CD8� Ts cells. However, when mice were de-pleted of CD4� T cells and then injected with haptenatednecrotic cells, this led to the generation of CD8� T cells thatinhibited responses upon transfer. Therefore, the generation ofregulatory CD8� Ts cells in this system appears to correlatewith the failure to activate CD4� T cells.
FIGURE 2. CD8� T cells transfer tolerance, produce TRAIL. A, C57BL/6 mice or C57BL/6 mice depleted of CD4� cells (anti-CD4 GK1.5 (100�g/day/mouse for 3 days) were injected i.v. with apoptotic (107) or necrotic (107 equivalence) TNP-coupled spleen cells. Seven days later, spleens wereremoved and CD8� T cells were isolated by negative selection. Purified CD8� T cells were then transferred to naive C57BL/6 mice, which were immunizedwith 0.1 ml of 10 mM TNBS s.c. Four days later, mice were challenged with 0.033 ml of 10 mM TNBS in the right footpad and 0.033 ml of PBS in theleft footpad. Measurements (micrometer � SE) were taken 24 h later and represent the difference between right footpad (Ag challenge) and left footpad(PBS challenge). �, Significantly different from the immune control. B, Quantitative RT-PCR. C57BL/6 mice were given apoptotic (107) TNP-coupledspleen cells i.v. 7 days before immunization with 0.1 ml of 10 mM TNBS s.c. On days 1, 2, 3, or 4 after immunization, spleens were removed and CD8�
T cells were purified by positive selection. RNA was isolated, reverse transcribed, and the amount of TRAIL, FasL, TNF, FLIP, Bcl-xL, and DR5 mRNAwas determined by quantitative PCR. The relative increase was determined by comparing RNA levels to CD8� T cells obtained from naive mice. C, TRAILproduction and CD4-mediated help. C57BL/6 or CD4-depleted C57BL/6 mice injected i.v. with TNP-apoptotic (107) or TNP-necrotic spleen cells (107
equivalence). Seven days later, mice were immunized with 0.1 ml of 10 mM TNBS s.c. One day later, CD8� T cells were isolated from the spleen bypositive selection and TRAIL levels determined by quantitative RT-PCR. The relative increase was determined by comparing RNA levels to CD8� T cellsobtained from naive mice. D, TRAIL production and restimulation. C57BL/6 mice were injected i.v. with 107 apoptotic TNP-coupled spleen cells. Sevendays later, mice were immunized with 0.1 ml of 10 mM TNBS s.c. Cohorts of mice were either tolerized, immunized, or both. One day later, CD8� T cellswere harvested from the spleen by positive selection and TRAIL levels were determined by quantitative RT-PCR. The relative increase was determinedby comparing RNA levels to CD8� T cells obtained from naive mice.
2681The Journal of Immunology
by guest on February 18, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

Apoptotic cells induce TRAIL production by CD8� T cells
Activation of CD8� T cells with Ag typically generates CTL thatkill target cells via perforin/granzyme or FasL pathways (27). Re-cent studies have indicated that CD8� CTL primed in the absenceof CD4� T cell help (“helpless CTL”) fail to undergo secondaryexpansion because they produce TRAIL and undergo activation-induced cell death (AICD) (28). CD8� cells primed in the pres-ence of CD4� T cell help, in contrast, do not produce TRAIL andexpand upon secondary exposure to Ag. Consequently, one pos-sible explanation for the results in Fig. 2A was that the CD8� Tcells primed in the absence of CD4� T cell help suppress anysubsequent immune response through the expression of TRAIL.To examine this possibility, mice were injected with haptenatedapoptotic cells and then immunized 7 days later. Cells from tol-erant or control immune mice were examined for expression ofseveral markers of “helped” vs “helpless” CTL (Fig. 2B). Al-though immune CD8� T cells showed elevated levels of c-FLIPand Bcl-xL, CD8� T cells from mice pretreated with haptenatedapoptotic cells (tolerant mice) showed elevated levels of TRAILmRNA. The gene expression pattern in the T cells from mice in-jected with haptenated apoptotic cells and then immunized resem-bled the pattern described for stimulated helpless CTL in thatTRAIL mRNA was elevated (28). Therefore, we examined theexpression of TRAIL in mice injected with haptenated apoptotic ornecrotic cells in the presence or absence of CD4� T cells. Micewere injected with apoptotic or necrotic cells and 7 days later theywere immunized with TNBS. Twenty-four hours after immuniza-tion, CD8� T cells were isolated and the level of TRAIL mRNAdetermined. Mice injected with apoptotic cells (CD4-depleted orintact) and CD4� T cell-depleted mice injected with haptenatednecrotic cells possessed CD8� T cells with elevated levels ofTRAIL mRNA (Fig. 2C). Other cell types from the spleen of tol-erant mice (e.g., CD4� T cells, B cells, etc.) did not show TRAILproduction (data not shown). Even though a variety of cells in theimmune system can be induced to express TRAIL under a numberof stimulatory conditions (29–31), these cells do not produceTRAIL under these tolerogenic conditions. We conclude that thegeneration of TRAIL-expressing CD8� T cells after i.v. immuni-zation with apoptotic cells is similar to that seen with necrotic cellsin the absence of CD4� T cell help, resulting in the activation ofcells resembling helpless CTL in both cases.
CTL stimulated in the absence of help have normal killing func-tion following primary stimulation; however, upon restimulationwith cognate Ag, the cells produce TRAIL and undergo AICD(28). Therefore, we examined our system for the requirement forrestimulation to induce TRAIL production. Mice were injectedwith apoptotic cells and 7 days later mice were immunized withTNBS or left untreated. One day later, the CD8� T cells wereremoved and TRAIL mRNA expression was determined. Fig. 2Dshows that CD8� T cells from tolerized mice (apoptotic cells/immunize) increased TRAIL mRNA, while mice that were immu-nized (no apoptotic cells) or injected with apoptotic cells (not im-munized) did not produce TRAIL. These data show that CD8� Tcells do not produce TRAIL from just a single encounter withapoptotic cells, but do so after a subsequent encounter with Ag(i.e., immunization).
Tolerant mice display TRAIL-mediated cellular cytotoxicity
Our data thus far suggest that apoptotic cells generate CD8� Tcells that regulate immunity and these cells resemble helpless CTLbased on production of TRAIL. CD8� T cells that regulate im-munity are generated when apoptotic cells pass through the cross-priming pathway of the DC (12). Similarly cross-priming in the
CD8� CTL compartment requires that Ags associated with apo-ptotic cells pass through the same class I pathway of the DC (4, 5,32). Because we observed the generation of TRAIL-producingCD8� T cells, it is possible that tolerant mice contain CTL thatmediate cytotoxicity via TRAIL. We examined this using an invivo killing system that can sensitively measure death (33). Thisassay monitored eradication of an adoptively transferred targetpopulation (TNP-spl) in tolerant and nontolerant mice. RecipientC57BL/6 or TRAIL�/� mice were injection with haptenated ap-optotic cells and immunized 1 wk later. Three days later, thesemice were infused with haptenated syngeneic spleen cells labeledwith a high concentration of CFSE (target cells) and unmodifiedsyngeneic spleen cells labeled with low CFSE (reference popula-tion) at a 1:1 ratio. Eighteen hours later, the remaining haptenatedspleen cells were enumerated and their numbers compared with thereference population. As controls, mice were untreated, immu-nized only, or given an injection of stimulatory anti-CD40 at thetime of injection of apoptotic cells as this treatment overcomestolerance in this system (12). These data are summarized in TableI and presented as percent (%) cytotoxicity. Immunization ofC57BL/6 or TRAIL�/� mice did not result in detectable cytotox-icity (group A), while injection of haptenated apoptotic cells alongwith anti-CD40 generated significant cytotoxicity in both strains(group D). However, mice rendered tolerant with apoptotic cellsfollowed by immunization demonstrated cytotoxicity in C57BL/6mice but not in TRAIL�/� mice (group C). Interestingly, miceinjected with haptenated apoptotic cells and not immunized (groupB) displayed some cytotoxicity in TRAIL�/� mice, suggestingsome influence of reencounter with Ag (see Fig. 2D). Thus, thesame procedure that generates tolerance also activates TRAIL-mediated cytotoxicity. These data show that apoptotic cells in-duce cellular cytotoxicity mediated by TRAIL and further sup-port the idea that apoptotic cells generate CD8� T cells with ahelpless CTL phenotype.
TRAIL is critical to tolerance induction
To confirm that TRAIL production was critical to the tolerance(i.e., immune suppression) mediated by “helpless” CD8� Tcells, we examined the generation of these cells in wild-typeC57BL/6 and TRAIL�/� mice. First, wild-type C57BL/6 miceor TRAIL�/� mice were injected with haptenated apoptoticcells. Animals were immunized 2 days later, the mice werechallenged after 4 days and the immune response was analyzedthe next day. Fig. 3A shows that TRAIL�/� mice were notrendered tolerant with apoptotic cells. Similarly, only CD8� Tcells obtained from tolerized C57BL/6 (TRAIL�/�) mice couldtransfer tolerance to naive mice (Fig. 3B). CD8� T cells from
Table I. In vivo killing of CFSE-labeled TNP-spl
Group TNP-spla Anti-CD40b Immunizec
% Cytotoxicityd
C57BL/6 TRAIL�/�
A � 0 0B � 26 � 4 12 � 2C � � 29 � 3 1D � � 0 0E � � � 100 � 10 100 � 14
a TNBS coupled spleen cells injected i.v.b Mice received a single injection of 300 �g of FGK45 (anti-CD40) at the time of
TNP-spl injection.c Mice were immunized with 0.1 ml of 10 mM TNBS s.c. 7 days following
TNP-spl.d Determined 3 days following infusion of CFSE-labeled cells. Values represent
the mean of five animals � SE.
2682 TRAIL AND TOLERANCE
by guest on February 18, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
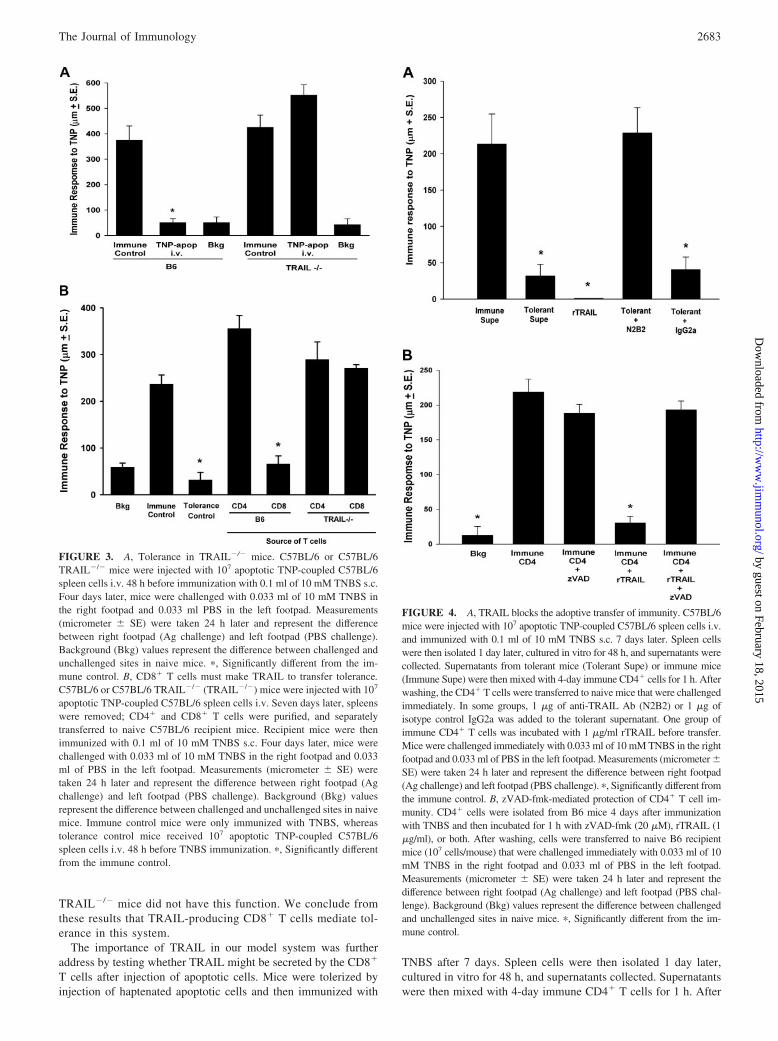
TRAIL�/� mice did not have this function. We conclude fromthese results that TRAIL-producing CD8� T cells mediate tol-erance in this system.
The importance of TRAIL in our model system was furtheraddress by testing whether TRAIL might be secreted by the CD8�
T cells after injection of apoptotic cells. Mice were tolerized byinjection of haptenated apoptotic cells and then immunized with
TNBS after 7 days. Spleen cells were then isolated 1 day later,cultured in vitro for 48 h, and supernatants collected. Supernatantswere then mixed with 4-day immune CD4� T cells for 1 h. After
FIGURE 3. A, Tolerance in TRAIL�/� mice. C57BL/6 or C57BL/6TRAIL�/� mice were injected with 107 apoptotic TNP-coupled C57BL/6spleen cells i.v. 48 h before immunization with 0.1 ml of 10 mM TNBS s.c.Four days later, mice were challenged with 0.033 ml of 10 mM TNBS inthe right footpad and 0.033 ml PBS in the left footpad. Measurements(micrometer � SE) were taken 24 h later and represent the differencebetween right footpad (Ag challenge) and left footpad (PBS challenge).Background (Bkg) values represent the difference between challenged andunchallenged sites in naive mice. �, Significantly different from the im-mune control. B, CD8� T cells must make TRAIL to transfer tolerance.C57BL/6 or C57BL/6 TRAIL�/� (TRAIL�/�) mice were injected with 107
apoptotic TNP-coupled C57BL/6 spleen cells i.v. Seven days later, spleenswere removed; CD4� and CD8� T cells were purified, and separatelytransferred to naive C57BL/6 recipient mice. Recipient mice were thenimmunized with 0.1 ml of 10 mM TNBS s.c. Four days later, mice werechallenged with 0.033 ml of 10 mM TNBS in the right footpad and 0.033ml of PBS in the left footpad. Measurements (micrometer � SE) weretaken 24 h later and represent the difference between right footpad (Agchallenge) and left footpad (PBS challenge). Background (Bkg) valuesrepresent the difference between challenged and unchallenged sites in naivemice. Immune control mice were only immunized with TNBS, whereastolerance control mice received 107 apoptotic TNP-coupled C57BL/6spleen cells i.v. 48 h before TNBS immunization. �, Significantly differentfrom the immune control.
FIGURE 4. A, TRAIL blocks the adoptive transfer of immunity. C57BL/6mice were injected with 107 apoptotic TNP-coupled C57BL/6 spleen cells i.v.and immunized with 0.1 ml of 10 mM TNBS s.c. 7 days later. Spleen cellswere then isolated 1 day later, cultured in vitro for 48 h, and supernatants werecollected. Supernatants from tolerant mice (Tolerant Supe) or immune mice(Immune Supe) were then mixed with 4-day immune CD4� cells for 1 h. Afterwashing, the CD4� T cells were transferred to naive mice that were challengedimmediately. In some groups, 1 �g of anti-TRAIL Ab (N2B2) or 1 �g ofisotype control IgG2a was added to the tolerant supernatant. One group ofimmune CD4� T cells was incubated with 1 �g/ml rTRAIL before transfer.Mice were challenged immediately with 0.033 ml of 10 mM TNBS in the rightfootpad and 0.033 ml of PBS in the left footpad. Measurements (micrometer �SE) were taken 24 h later and represent the difference between right footpad(Ag challenge) and left footpad (PBS challenge). �, Significantly different fromthe immune control. B, zVAD-fmk-mediated protection of CD4� T cell im-munity. CD4� cells were isolated from B6 mice 4 days after immunizationwith TNBS and then incubated for 1 h with zVAD-fmk (20 �M), rTRAIL (1�g/ml), or both. After washing, cells were transferred to naive B6 recipientmice (107 cells/mouse) that were challenged immediately with 0.033 ml of 10mM TNBS in the right footpad and 0.033 ml of PBS in the left footpad.Measurements (micrometer � SE) were taken 24 h later and represent thedifference between right footpad (Ag challenge) and left footpad (PBS chal-lenge). Background (Bkg) values represent the difference between challengedand unchallenged sites in naive mice. �, Significantly different from the im-mune control.
2683The Journal of Immunology
by guest on February 18, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
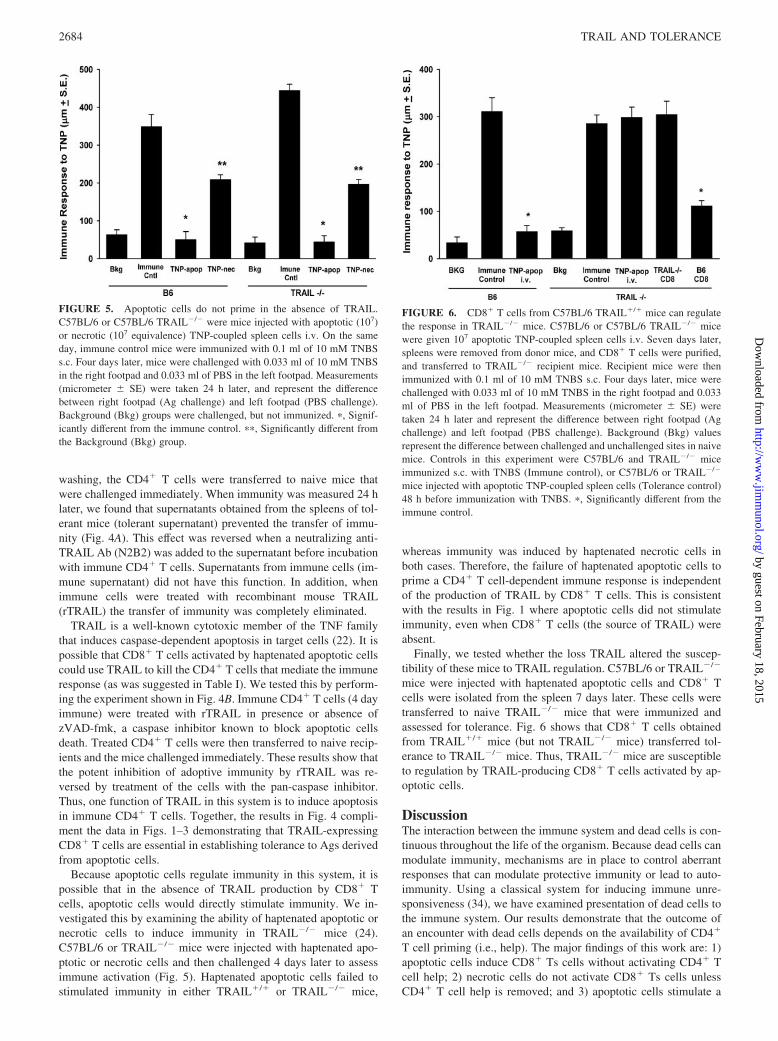
washing, the CD4� T cells were transferred to naive mice thatwere challenged immediately. When immunity was measured 24 hlater, we found that supernatants obtained from the spleens of tol-erant mice (tolerant supernatant) prevented the transfer of immu-nity (Fig. 4A). This effect was reversed when a neutralizing anti-TRAIL Ab (N2B2) was added to the supernatant before incubationwith immune CD4� T cells. Supernatants from immune cells (im-mune supernatant) did not have this function. In addition, whenimmune cells were treated with recombinant mouse TRAIL(rTRAIL) the transfer of immunity was completely eliminated.
TRAIL is a well-known cytotoxic member of the TNF familythat induces caspase-dependent apoptosis in target cells (22). It ispossible that CD8� T cells activated by haptenated apoptotic cellscould use TRAIL to kill the CD4� T cells that mediate the immuneresponse (as was suggested in Table I). We tested this by perform-ing the experiment shown in Fig. 4B. Immune CD4� T cells (4 dayimmune) were treated with rTRAIL in presence or absence ofzVAD-fmk, a caspase inhibitor known to block apoptotic cellsdeath. Treated CD4� T cells were then transferred to naive recip-ients and the mice challenged immediately. These results show thatthe potent inhibition of adoptive immunity by rTRAIL was re-versed by treatment of the cells with the pan-caspase inhibitor.Thus, one function of TRAIL in this system is to induce apoptosisin immune CD4� T cells. Together, the results in Fig. 4 compli-ment the data in Figs. 1–3 demonstrating that TRAIL-expressingCD8� T cells are essential in establishing tolerance to Ags derivedfrom apoptotic cells.
Because apoptotic cells regulate immunity in this system, it ispossible that in the absence of TRAIL production by CD8� Tcells, apoptotic cells would directly stimulate immunity. We in-vestigated this by examining the ability of haptenated apoptotic ornecrotic cells to induce immunity in TRAIL�/� mice (24).C57BL/6 or TRAIL�/� mice were injected with haptenated apo-ptotic or necrotic cells and then challenged 4 days later to assessimmune activation (Fig. 5). Haptenated apoptotic cells failed tostimulated immunity in either TRAIL�/� or TRAIL�/� mice,
whereas immunity was induced by haptenated necrotic cells inboth cases. Therefore, the failure of haptenated apoptotic cells toprime a CD4� T cell-dependent immune response is independentof the production of TRAIL by CD8� T cells. This is consistentwith the results in Fig. 1 where apoptotic cells did not stimulateimmunity, even when CD8� T cells (the source of TRAIL) wereabsent.
Finally, we tested whether the loss TRAIL altered the suscep-tibility of these mice to TRAIL regulation. C57BL/6 or TRAIL�/�
mice were injected with haptenated apoptotic cells and CD8� Tcells were isolated from the spleen 7 days later. These cells weretransferred to naive TRAIL�/� mice that were immunized andassessed for tolerance. Fig. 6 shows that CD8� T cells obtainedfrom TRAIL�/� mice (but not TRAIL�/� mice) transferred tol-erance to TRAIL�/� mice. Thus, TRAIL�/� mice are susceptibleto regulation by TRAIL-producing CD8� T cells activated by ap-optotic cells.
DiscussionThe interaction between the immune system and dead cells is con-tinuous throughout the life of the organism. Because dead cells canmodulate immunity, mechanisms are in place to control aberrantresponses that can modulate protective immunity or lead to auto-immunity. Using a classical system for inducing immune unre-sponsiveness (34), we have examined presentation of dead cells tothe immune system. Our results demonstrate that the outcome ofan encounter with dead cells depends on the availability of CD4�
T cell priming (i.e., help). The major findings of this work are: 1)apoptotic cells induce CD8� Ts cells without activating CD4� Tcell help; 2) necrotic cells do not activate CD8� Ts cells unlessCD4� T cell help is removed; and 3) apoptotic cells stimulate a
FIGURE 5. Apoptotic cells do not prime in the absence of TRAIL.C57BL/6 or C57BL/6 TRAIL�/� were mice injected with apoptotic (107)or necrotic (107 equivalence) TNP-coupled spleen cells i.v. On the sameday, immune control mice were immunized with 0.1 ml of 10 mM TNBSs.c. Four days later, mice were challenged with 0.033 ml of 10 mM TNBSin the right footpad and 0.033 ml of PBS in the left footpad. Measurements(micrometer � SE) were taken 24 h later, and represent the differencebetween right footpad (Ag challenge) and left footpad (PBS challenge).Background (Bkg) groups were challenged, but not immunized. �, Signif-icantly different from the immune control. ��, Significantly different fromthe Background (Bkg) group.
FIGURE 6. CD8� T cells from C57BL/6 TRAIL�/� mice can regulatethe response in TRAIL�/� mice. C57BL/6 or C57BL/6 TRAIL�/� micewere given 107 apoptotic TNP-coupled spleen cells i.v. Seven days later,spleens were removed from donor mice, and CD8� T cells were purified,and transferred to TRAIL�/� recipient mice. Recipient mice were thenimmunized with 0.1 ml of 10 mM TNBS s.c. Four days later, mice werechallenged with 0.033 ml of 10 mM TNBS in the right footpad and 0.033ml of PBS in the left footpad. Measurements (micrometer � SE) weretaken 24 h later and represent the difference between right footpad (Agchallenge) and left footpad (PBS challenge). Background (Bkg) valuesrepresent the difference between challenged and unchallenged sites in naivemice. Controls in this experiment were C57BL/6 and TRAIL�/� miceimmunized s.c. with TNBS (Immune control), or C57BL/6 or TRAIL�/�
mice injected with apoptotic TNP-coupled spleen cells (Tolerance control)48 h before immunization with TNBS. �, Significantly different from theimmune control.
2684 TRAIL AND TOLERANCE
by guest on February 18, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

helpless CD8� T cell phenotype that can function as CTL andregulate CD4� T cell-mediated immunity by the secretion ofTRAIL. Our results provide a molecular mechanism for infectioustolerance mediated by CD8� T cells and suggest how apoptosisin the immune system can promote immune tolerance.
The induction of immunological unresponsiveness by apoptoticcells has been attributed to a number of mechanisms. These in-clude the induction of immunosuppressive cytokines from phago-cytic cells (35), the production of inhibitors from the dead cellitself (36, 37), and effects on the maturation of the DC (4, 5). Wehave observed that apoptotic cells do not prime CD4� T cells, butdo stimulate CD8� cells. In recent years, a number of reports havehighlighted the pivotal role that CD4� T cells play in the inductionof CD8� T cell responses (4, 25, 28, 38). Although naive CD8� Tcells can be directly activated to proliferate in the absence ofCD4� T cells, the amount of IL-2 produced by CD8� T cells is�10-fold less than CD4� T cells, an amount that may be insuffi-cient to sustain a response (39). Thus, most CD8� T cell-mediatedresponses depend on concomitant priming of CD4� T cells to beeffective. In other studies, CD8� T cell priming in the absence ofCD4� T cell help leads to their deletion from the periphery, aneffect that can be overcome by supplying help during the initialpriming phase (9). Recently, it was shown that the priming ofCD8� T cells in the absence of CD4� T cell help alters the pro-gramming of these cells, which was only revealed upon secondarystimulation. Specifically, CD8� T cells activated without help (i.e.,helpless) produce TRAIL and undergo AICD upon secondary an-tigenic stimulation (28). We have observed a similar phenomenonfollowing encounter with apoptotic cells, i.e., the apoptotic cells donot prime CD4� cells, and the CD8� T cells acquire a helplessphenotype upon second stimulation. These cells mediate cellularcytotoxicity and suppress the response of CD4� T cells that me-diate delayed-type hypersensitivity. Thus, helpless CTL and CD8�
T cells that regulate immunity (i.e., Ts cells) may be the samecells.
Although the role of TRAIL in the concept of helpless CTL isan important observation when considering immunologic memory(28), the participation of TRAIL in activation-induced T cell deathwas first reported nearly 10 years ago by several laboratories (40,41) (42). Consequently, the immunoregulatory TRAIL-expressingCD8� T cells activated in response to hapten identified in oursystem may also act upon other CD8� T cells that would be in-volved in any subsequent Ag-specific immune response. CD8� Tcells are more sensitive to TRAIL-mediated regulation comparedwith CD4� T cells (31), which supports this possibility. However,since CD8� T cells are not involved in the transfer of immunity inthe present system, TRAIL effects on CD8� T cells may not in-fluence the immune response. Interestingly, an effect on CD8� Tcells might lead to the elimination of the induced Ts cells follow-ing TRAIL secretion.
One interesting finding from our studies was that haptenatednecrotic cells, which normally prime for immunity when given i.v.,can also generate a “helpless CTL” phenotype if CD4� T cells arefirst eliminated. It has been thought that what determines the out-come of the immune response to dead cells is influenced by surfacereceptors displayed by the dying cells. Apoptotic cells expressunique markers, such as CD36 and phosphatidylserine, and areefficiently phagocytized (43, 44). These markers help guide theapoptotic cells through the Ag-processing machinery leading totolerance by deletion and/or the production of immunosuppressivecytokines, such as IL-10 or TGF-� (36, 37). Conversely, necroticcells do not display such makers and are thus thought to not enterthe cross tolerance pathway. Necrotic cells can further stimulateimmunity by the release proinflammatory molecules, such as high
mobility group box protein 1 (6), uric acid (7), and/or heat shockprotein 70 (8), which prime CD4� T cell help and stimulate DCmaturation. Our data show that in the absence of CD4� T cell help,necrotic cells perform the same function as apoptotic cells, i.e.,induce immunologic tolerance and promote the development ofTRAIL-expressing CD8� Ts cells. Thus, necrotic cells must behandled similarly to apoptotic cells by APC, even though they canrelease proinflammatory molecules that can result in DC matura-tion. Perhaps more importantly, necrotic cells, even if they maturethe DC, can induce immunologic unresponsiveness should there beinadequate CD4� T cell help. CD4� T cell help is, therefore, a keydeterminant in the immunological outcome after the encounterwith dead cells.
TRAIL is becoming an increasingly important member of theTNF family of death-inducing ligands (22). Originally thought tobe a tumor-specific killer, recent studies have shown that TRAILis functional in a variety of other physiological systems (28, 45,46). Although the TRAIL�/� mice do not have an overt phenotype(24), studies with infectious agents and autoimmune models showthat TRAIL can play an important role in controlling the extent ofthe autoimmune reaction, as well as be an effector molecule incertain disease states (47, 48). In the experimental autoimmuneencephalomyelitis model, for example, the lack of TRAIL exac-erbates autoimmunity. Recent studies with TRAIL�/� mice in amodel of multiple sclerosis have suggested an important role forsuppressor CD8� T cells protecting against disease recurrence andexacerbation (49). In the NOD mouse, which is prone to autoim-mune diabetes, TRAIL blockade similarly exacerbates the onset oftype 1 diabetes (50). Also, treatment of mice undergoing experi-mental autoimmune thyroiditis with rTRAIL ameliorated the dis-ease (51). In our system, the failure of apoptotic cells to primeCD4� T cells results in CD8� cells that produce TRAIL, suggest-ing an important immunoregulatory role for TRAIL in the controlof potentially dangerous anti-self responses that could be inducedwith dead cells. TRAIL appears to function in our tolerance modelby inhibiting Ag-reactive CD4� cells through the induction of ap-optosis, which is consistent with previous reports of TRAIL inregulating T cell homeostasis (40, 42). Consistent with this is thefinding that functional TRAILR has been reported on mouse (52)and human (30) activated T cells, but not on resting T cells. To-gether, these results suggest that TRAIL may play a critical role incontrolling immune reactions.
TRAIL can also induce apoptosis in a number of other cells inthe immune system, including plasma cells, neutrophils, and DCs(53) (54) (55). Although we only examined the ability of theTRAIL-expressing CD8� Ts cells induced in our system to regu-late hapten-specific CD4� T cells, it is also possible that theTRAIL could be modulating the function of other cells, such asAPCs. TRAIL, or more specifically TRAILR, has been recentlyreported to act as a negative regulator of innate immune cell re-sponses (56). Using TRAILR-deficient mice, it was demonstratedthat these mice displayed enhanced innate immune responses afterbacterial or viral challenge. Further investigation revealed thatstimulation of TRAILR-deficient macrophages with various TLRagonists led to enhanced cytokine production compared with wildtype, suggesting that the TRAIL-TRAILR system has the capacityto regulate cytokine production and the resultant immune response.Similar findings were reported for Listeria-induced hepatitis (48)and for Listeria infections (47) in TRAIL-deficient mice. In bothsystems, the loss of TRAIL led to enhanced innate immunity.Thus, while the regulatory activity of TRAIL-expressing CD8� Tcells may be beneficial for protection in autoimmunity, their pres-ence may be detrimental during times of infection.
2685The Journal of Immunology
by guest on February 18, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

Some of the earliest descriptions of Treg examined the functionof CD8� Ts cells (13–15), which could regulate immune responsesthrough the secretion of a soluble factor that suppressed immunityby inhibiting T cell function (called T suppressor factor) (57, 58).Although our results are not intended to explain all observationswith T suppressor factor, it is interesting to speculate that at leastsome of the suppressor activity in the supernatants of CD8� Tscells was TRAIL. It is important to note that T cells that suppressimmune responses may not belong to a unique T cell subset withspecialized function, but may instead represent regulatory activi-ties of otherwise normal helper and cytotoxic CD8� T cells. Ourdata showing that the CD8 Treg is a helpless CTL support thisidea. This may explain why it was previously so difficult to iden-tify unique cell surface markers that identified the Ts cells as afunctional population (and one of the main issues that fueled the“suppressor cell” controversy years ago) (59). Perhaps our resultsalso demonstrate why it was so difficult to clone Ts cells, i.e., whengenerated they produced TRAIL and underwent AICD.
DisclosuresThe authors have no financial conflict of interest.
References1. Cohen, J. J. 1993. Apoptosis. Immunol. Today 14: 126–124.2. Vaux, D. L., and A. Strasser. 1996. The molecular biology of apoptosis. Proc.
Natl. Acad. Sci. USA 93: 2239–2244.3. Wyllie, A. H., J. F. Kerr, and A. R. Currie. 1980. Cell death: the significance of
apoptosis. Int. Rev. Cytol. 68: 251–306.4. Albert, M. L., M. Jegathesan, and R. B. Darnell. 2001. Dendritic cell maturation
is required for the cross-tolerization of CD8� T cells. Nat. Immunol. 2:1010–1017.
5. Steinman, R. M., S. Turley, I. Mellman, and K. Inaba. 2000. The induction oftolerance by dendritic cells that have captured apoptotic cells. J. Exp. Med. 191:411–416.
6. Scaffidi, P., T. Misteli, and M. E. Bianchi. 2002. Release of chromatin proteinHMGB1 by necrotic cells triggers inflammation. Nature 418: 191–195.
7. Shi, Y., J. E. Evans, and K. L. Rock. 2003. Molecular identification of a dangersignal that alerts the immune system to dying cells. Nature 425: 516–521.
8. Basu, S., R. J. Binder, R. Suto, K. M. Anderson, and P. K. Srivastava. 2000.Necrotic but not apoptotic cell death releases heat shock proteins, which delivera partial maturation signal to dendritic cells and activate the NF-�B pathway. Int.Immunol. 12: 1539–1546.
9. Kurts, C., F. R. Carbone, M. Barnden, E. Blanas, J. Allison, W. R. Heath, andJ. F. Miller. 1997. CD4� T cell help impairs CD8� T cell deletion induced bycross-presentation of self-antigens and favors autoimmunity. J. Exp. Med. 186:2057–2062.
10. Sauter, B., M. L. Albert, L. Francisco, M. Larsson, S. Somersan, andN. Bhardwaj. 2000. Consequences of cell death: exposure to necrotic tumor cells,but not primary tissue cells or apoptotic cells, induces the maturation of immu-nostimulatory dendritic cells. J. Exp. Med. 191: 423–434.
11. Steinman, R. M., D. Hawiger, and M. C. Nussenzweig. 2003. Tolerogenic den-dritic cells. Annu. Rev. Immunol. 21: 685–711.
12. Ferguson, T. A., J. Herndon, B. Elzey, T. S. Griffith, S. Schoenberger, andD. R. Green. 2002. Uptake of apoptotic antigen-coupled cells by lymphoid den-dritic cells and cross-priming of CD8� T cells produce active immune unrespon-siveness. J. Immunol. 168: 5589–5595.
13. Gershon, R. K., and K. Kondo. 1971. Infectious immunological tolerance. Im-munology 21: 903–914.
14. Sy, M. S., S. D. Miller, H. B. Kowach, and H. N. Claman. 1977. A splenicrequirement for the generation of suppressor T cells. J. Immunol. 119:2095–2099.
15. Greene, M. I., and B. Benacerraf. 1980. Studies on hapten specific T cell immu-nity and suppression. Immunol. Rev. 50: 163–186.
16. Ben-Nun, A., H. Wekerle, and I. R. Cohen. 1981. Vaccination against autoim-mune encephalomyelitis with T-lymphocyte line cells reactive against myelinbasic protein. Nature 292: 60–61.
17. Jiang, H., H. Kashleva, L. X. Xu, J. Forman, L. Flaherty, B. Pernis,N. S. Braunstein, and L. Chess. 1998. T cell vaccination induces T cell receptorV�-specific Qa-1- restricted regulatory CD8� T cells. Proc. Natl. Acad. Sci. USA95: 4533–4537.
18. Sarantopoulos, S., L. Lu, and H. Cantor. 2004. Qa-1 restriction of CD8� sup-pressor T cells. J. Clin. Invest. 114: 1218–1221.
19. Gilliet, M., and Y. J. Liu. 2002. Generation of human CD8 T regulatory cells byCD40 ligand-activated plasmacytoid dendritic cells. J. Exp. Med. 195: 695–704.
20. Seder, R. A., and G. G. Le Gros. 1995. The functional role of CD8� T helper type2 cells. J. Exp. Med. 181: 5–7.
21. Shevach, E. M. 2002. CD4� CD25� suppressor T cells: more questions thananswers. Nat. Rev. Immunol. 2: 389–400.
22. Wiley, S. R., K. Schooley, P. J. Smolak, W. S. Din, C. P. Huang, J. K. Nicholl,G. R. Sutherland, T. D. Smith, C. Rauch, C. A. Smith, et al. 1995. Identificationand characterization of a new member of the TNF family that induces apoptosis.Immunity 3: 673–682.
23. Pitti, R. M., S. A. Marsters, S. Ruppert, C. J. Donahue, A. Moore, andA. Ashkenazi. 1996. Induction of apoptosis by Apo-2 ligand, a new member ofthe tumor necrosis factor cytokine family. J. Biol. Chem. 271: 12687–12690.
24. Sedger, L. M., M. B. Glaccum, J. C. Schuh, S. T. Kanaly, E. Williamson,N. Kayagaki, T. Yun, P. Smolak, T. Le, R. Goodwin, and B. Gliniak. 2002.Characterization of the in vivo function of TNF-�-related apoptosis-inducingligand, TRAIL/Apo2L, using TRAIL/Apo2L gene-deficient mice. Eur. J. Immu-nol. 32: 2246–2254.
25. Schoenberger, S. P., R. E. Toes, E. I. van der Voort, R. Offringa, and C. J. Melief.1998. T-cell help for cytotoxic T lymphocytes is mediated by CD40-CD40Linteractions. Nature 393: 480–483.
26. VanOosten, R. L., J. M. Moore, B. Karacay, and T. S. Griffith. 2005. Histonedeacetylase inhibitors modulate renal cell carcinoma sensitivity to TRAIL/Apo-2L-induced apoptosis by enhancing TRAIL-R2 expression. Cancer Biol. Ther. 4:1104–1112.
27. Shresta, S., C. T. Pham, D. A. Thomas, T. A. Graubert, and T. J. Ley. 1998. Howdo cytotoxic lymphocytes kill their targets? Curr. Opin. Immunol. 10: 581–587.
28. Janssen, E. M., N. M. Droin, E. E. Lemmens, M. J. Pinkoski, S. J. Bensinger,B. D. Ehst, T. S. Griffith, D. R. Green, and S. P. Schoenberger. 2005. CD4�
T-cell help controls CD8� T-cell memory via TRAIL-mediated activation-in-duced cell death. Nature 434: 88–93.
29. Lunemann, J. D., S. Waiczies, S. Ehrlich, U. Wendling, B. Seeger, T. Kamradt,and F. Zipp. 2002. Death ligand TRAIL induces no apoptosis but inhibits acti-vation of human (auto)antigen-specific T cells. J. Immunol. 168: 4881–4888.
30. Herbeuval, J. P., J. C. Grivel, A. Boasso, A. W. Hardy, C. Chougnet, M. J. Dolan,H. Yagita, J. D. Lifson, and G. M. Shearer. 2005. CD4� T-cell death induced byinfectious and noninfectious HIV-1: role of type 1 interferon-dependent, TRAIL/DR5-mediated apoptosis. Blood 106: 3524–3531.
31. Bosque, A., J. Pardo, M. J. Martinez-Lorenzo, P. Lasierra, L. Larrad, I. Marzo,J. Naval, and A. Anel. 2005. Human CD8� T cell blasts are more sensitive thanCD4� T cell blasts to regulation by APO2L/TRAIL. Eur. J. Immunol. 35:1812–1821.
32. Belz, G. T., G. M. Behrens, C. M. Smith, J. F. Miller, C. Jones, K. Lejon,C. G. Fathman, S. N. Mueller, K. Shortman, F. R. Carbone, and W. R. Heath.2002. The CD8�� dendritic cell is responsible for inducing peripheral self-tol-erance to tissue-associated antigens. J. Exp. Med. 196: 1099–1104.
33. Herndon, J. M., P. M. Stuart, and T. A. Ferguson. 2005. Peripheral deletion ofantigen-specific T cells leads to long-term tolerance mediated by CD8� cytotoxiccells. J. Immunol. 174: 4098–4104.
34. Battisto, J. R., and B. R. Bloom. 1966. Dual immunological unresponsivenessinduced by cell membrane coupled hapten or antigen. Nature 212: 156–157.
35. Fadok, V. A., D. L. Bratton, A. Konowal, P. W. Freed, J. Y. Westcott, andP. M. Henson. 1998. Macrophages that have ingested apoptotic cells in vitroinhibit proinflammatory cytokine production through autocrine/paracrine mech-anisms involving TGF-�, PGE2, and PAF. J. Clin. Invest. 101: 890–898.
36. Gao, Y., J. M. Herndon, H. Zhang, T. S. Griffith, and T. A. Ferguson. 1998.Antiinflammatory effects of CD95 ligand (FasL)-induced apoptosis. J. Exp. Med.188: 887–896.
37. Chen, W., M. E. Frank, W. Jin, and S. M. Wahl. 2001. TGF-� released byapoptotic T cells contributes to an immunosuppressive milieu. Immunity 14:715–725.
38. Bennett, S. R., F. R. Carbone, F. Karamalis, J. F. Miller, and W. R. Heath. 1997.Induction of a CD8� cytotoxic T lymphocyte response by cross-priming requirescognate CD4� T cell help. J. Exp. Med. 186: 65–70.
39. Singer, A., T. I. Munitz, H. Golding, A. S. Rosenberg, and T. Mizuochi. 1987.Recognition requirements for the activation, differentiation and function of T-helper cells specific for class I MHC alloantigens. Immunol. Rev. 98: 143–170.
40. Katsikis, P. D., M. E. Garcia-Ojeda, J. F. Torres-Roca, I. M. Tijoe, C. A. Smith,and L. A. Herzenberg. 1997. Interleukin-1� converting enzyme-like proteaseinvolvement in Fas-induced and activation-induced peripheral blood T cell apo-ptosis in HIV infection: TNF-related apoptosis-inducing ligand can mediate ac-tivation-induced T cell death in HIV infection. J. Exp. Med. 186: 1365–1372.
41. Jeremias, I., I. Herr, T. Boehler, and K. M. Debatin. 1998. TRAIL/Apo-2-ligand-induced apoptosis in human T cells. Eur. J. Immunol. 28: 143–152.
42. Martinez-Lorenzo, M. J., M. A. Alava, S. Gamen, K. J. Kim, A. Chuntharapai,A. Pineiro, J. Naval, and A. Anel. 1998. Involvement of APO2 ligand/TRAIL inactivation-induced death of Jurkat and human peripheral blood T cells. Eur.J. Immunol. 28: 2714–2725.
43. Fadok, V. A., D. L. Bratton, S. C. Frasch, M. L. Warner, and P. M. Henson. 1998.The role of phosphatidylserine in recognition of apoptotic cells by phagocytes.Cell Death Differ. 5: 551–562.
44. Savill, J., I. Dransfield, C. Gregory, and C. Haslett. 2002. A blast from the past:clearance of apoptotic cells regulates immune responses. Nat. Rev. Immunol. 2:965–975.
45. Griffith, T. S., and D. H. Lynch. 1998. TRAIL: a molecule with multiple receptorsand control mechanisms. Curr. Opin. Immunol. 10: 559–563.
46. Lee, H. O., J. M. Herndon, R. Barreiro, T. S. Griffith, and T. A. Ferguson. 2002.TRAIL: a mechanism of tumor surveillance in an immune privileged site. J. Im-munol. 169: 4739–4744.
47. Zheng, S. J., J. Jiang, H. Shen, and Y. H. Chen. 2004. Reduced apoptosis andameliorated listeriosis in TRAIL-null mice. J. Immunol. 173: 5652–5658.
48. Zheng, S. J., P. Wang, G. Tsabary, and Y. H. Chen. 2004. Critical roles of TRAILin hepatic cell death and hepatic inflammation. J. Clin. Invest. 113: 58–64.
2686 TRAIL AND TOLERANCE
by guest on February 18, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

49. Hilliard, B., A. Wilmen, C. Seidel, T. S. Liu, R. Goke, and Y. Chen. 2001. Rolesof TNF-related apoptosis-inducing ligand in experimental autoimmune enceph-alomyelitis. J. Immunol. 166: 1314–1319.
50. Mi, Q. S., D. Ly, S. E. Lamhamedi-Cherradi, K. V. Salojin, L. Zhou, M. Grattan,C. Meagher, P. Zucker, Y. H. Chen, J. Nagle, et al. 2003. Blockade of tumornecrosis factor-related apoptosis-inducing ligand exacerbates type 1 diabetes inNOD mice. Diabetes 52: 1967–1975.
51. Wang, S. H., Z. Cao, J. M. Wolf, M. Van Antwerp, and J. R. Baker, Jr. 2005.Death ligand tumor necrosis factor-related apoptosis-inducing ligand inhibits ex-perimental autoimmune thyroiditis. Endocrinology 146: 4721–4726.
52. Zhang, X. R., L. Y. Zhang, S. Devadas, L. Li, A. D. Keegan, and Y. F. Shi. 2003.Reciprocal expression of TRAIL and CD95L in Th1 and Th2 cells: role of ap-optosis in T helper subset differentiation. Cell Death Differ. 10: 203–210.
53. Ursini-Siegel, J., W. Zhang, A. Altmeyer, E. N. Hatada, R. K. Do, H. Yagita, andS. Chen-Kiang. 2002. TRAIL/Apo-2 ligand induces primary plasma cell apopto-sis. J. Immunol. 169: 5505–5513.
54. Renshaw, S. A., J. S. Parmar, V. Singleton, S. J. Rowe, D. H. Dockrell,S. K. Dower, C. D. Bingle, E. R. Chilvers, and M. K. Whyte. 2003. Accelerationof human neutrophil apoptosis by TRAIL. J. Immunol. 170: 1027–1033.
55. Hayakawa, Y., V. Screpanti, H. Yagita, A. Grandien, H. G. Ljunggren,M. J. Smyth, and B. J. Chambers. 2004. NK cell TRAIL eliminates immaturedendritic cells in vivo and limits dendritic cell vaccination efficacy. J. Immunol.172: 123–129.
56. Diehl, G. E., H. H. Yue, K. Hsieh, A. A. Kuang, M. Ho, L. A. Morici, L. L. Lenz,D. Cado, L. W. Riley, and A. Winoto. 2004. TRAIL-R as a negative regulator ofinnate immune cell responses. Immunity 21: 877–889.
57. Yamauchi, K., D. Murphy, H. Cantor, and R. K. Gershon. 1981. Analysis of anantigen-specific H-2-restricted cell-free products(s) made by “I-J-” Ly-2 cells(Ly-2 TsF) that suppresses Ly-2 cell-depleted spleen cell activity. Eur. J. Immu-nol. 11: 913–918.
58. Flood, P., K. Yamauchi, and R. K. Gershon. 1982. Analysis of the interactionsbetween two molecules that are required for the expression of Ly-2 suppressorcell activity: three different types of focusing events may be needed to deliver thesuppressive signal. J. Exp. Med. 156: 361–371.
59. Green, D. R., and D. R. Webb. 1993. Saying the ‘S’ word in public. Immunol.Today 14: 523–525.
2687The Journal of Immunology
by guest on February 18, 2015http://w
ww
.jimm
unol.org/D
ownloaded from




















