
TRAIL mediates apoptosis in cancerous but not normal primary cultured cells of the human...
13
Apoptosis (2007) 12:73–85 DOI 10.1007/s10495-006-0492-z TRAIL mediates apoptosis in cancerous but not normal primary cultured cells of the human reproductive tract Anil Sadarangani · Sumie Kato · Natalia Espinoza · Soledad Lange · Carmen Llados · Marisol Espinosa · Manuel Villal´ on · Stanley Lipkowitz · Mauricio Cuello · Gareth I. Owen Published online: 21 November 2006 C Springer Science + Business Media, LLC 2006 Abstract Cancer of the reproductive tract encompasses ma- lignancies of the uterine corpus, cervix, ovary, Fallopian tube, among others and accounts for 15% of female can- cer mortalities. Tumour necrosis factor-related apoptosis- inducing ligand (TRAIL) mediates apoptosis by binding to death receptors and offers a promising cancer treatment. The goal of this study was to investigate and character- ize the effect of TRAIL in endometrial cancer cell lines and normal (non-cancerous) epithelial cells of endometrial ori- gin. We also examined the effect of TRAIL in other primary cultured cancers and normal cells of the human female re- productive tract and evaluated if TRAIL mediated apoptosis correlated with death receptors and decoy receptors 1 and 2. Herein, we demonstrate that TRAIL at concentrations which kill cancerous cells, does not mediate apoptosis or alter cell viability in normal human endometrium, ovary, cervix or Fallopian tube. The partial inhibition by a caspase 9 inhibitor This work was supported by a grant from the Wellcome Trust GR071469 (GIO) and the Chilean national science grants FONDECYT 1060495 (GIO) and 1020675 (MC). A. Sadarangani · N. Espinoza · S. Lange · C. Llados · M. Espinosa · M. Villal´ on · G. I. Owen () Department of Physiology, Faculty of Biological Sciences, Pontificia Universidad Cat´ olica de Chile, Alameda 340, Santiago, Chile e-mail: [email protected] S. Kato · M. Cuello Departament of Obstetrics and Gynecology, Faculty of Medicine, Pontificia Universidad Cat´ olica de Chile, Alameda 340, Santiago, Chile S. Lipkowitz Laboratory of Cellular and Molecular Biology, Center for Cancer Research, NCI, Bethesda, MD 20892, USA and the total inhibition by a caspase 8 inhibitor demonstrates the dependency on the extrinsic apoptotic pathway. The se- lective mortality does not correlate with the presence of death or decoy receptors. These results suggest that TRAIL may be an effective treatment for endometrial cancer and other female reproductive cancers, with minimal secondary effects on healthy tissue. Keywords Endometrium . Cervix . Fallopian . Ovarian cancer Abbreviations TRAIL Tumour necrosis factor-related apoptosis- inducing ligand TRAIL-R TRAIL receptor DR4/5 Death receptor 4/5 DcR decoy receptor T TRAIL C vehicle control kDa kilodalton FBS fetal bovine serum MTS 3-(4,5-Dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide Introduction Cancer of the female reproductive tract encompasses malignancies of the uterine corpus (endometrial cancer), uterine cervix, ovary, vulva, Fallopian tube, vagina and other related genital tissue. According to estimated values for 2005 by the American Cancer Society, cancer of the reproductive tract in the USA will account for 12% of all female cancer incidence (79,480 of 662,870 estimated cases) and approximately 15% of female cancer mortalities (28,910 of 275,000 estimated cases). Springer
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of TRAIL mediates apoptosis in cancerous but not normal primary cultured cells of the human...

Apoptosis (2007) 12:73–85DOI 10.1007/s10495-006-0492-z
TRAIL mediates apoptosis in cancerous but not normal primarycultured cells of the human reproductive tractAnil Sadarangani · Sumie Kato · Natalia Espinoza · Soledad Lange · Carmen Llados ·Marisol Espinosa · Manuel Villalon · Stanley Lipkowitz · Mauricio Cuello ·Gareth I. Owen
Published online: 21 November 2006C© Springer Science + Business Media, LLC 2006
Abstract Cancer of the reproductive tract encompasses ma-lignancies of the uterine corpus, cervix, ovary, Fallopiantube, among others and accounts for 15% of female can-cer mortalities. Tumour necrosis factor-related apoptosis-inducing ligand (TRAIL) mediates apoptosis by binding todeath receptors and offers a promising cancer treatment.
The goal of this study was to investigate and character-ize the effect of TRAIL in endometrial cancer cell lines andnormal (non-cancerous) epithelial cells of endometrial ori-gin. We also examined the effect of TRAIL in other primarycultured cancers and normal cells of the human female re-productive tract and evaluated if TRAIL mediated apoptosiscorrelated with death receptors and decoy receptors 1 and 2.
Herein, we demonstrate that TRAIL at concentrationswhich kill cancerous cells, does not mediate apoptosis or altercell viability in normal human endometrium, ovary, cervix orFallopian tube. The partial inhibition by a caspase 9 inhibitor
This work was supported by a grant from the Wellcome TrustGR071469 (GIO) and the Chilean national science grantsFONDECYT 1060495 (GIO) and 1020675 (MC).
A. Sadarangani · N. Espinoza · S. Lange · C. Llados ·M. Espinosa · M. Villalon · G. I. Owen (�)Department of Physiology, Faculty of Biological Sciences,Pontificia Universidad Catolica de Chile,Alameda 340, Santiago, Chilee-mail: [email protected]
S. Kato · M. CuelloDepartament of Obstetrics and Gynecology, Faculty of Medicine,Pontificia Universidad Catolica de Chile,Alameda 340, Santiago, Chile
S. LipkowitzLaboratory of Cellular and Molecular Biology, Center for CancerResearch, NCI,Bethesda, MD 20892, USA
and the total inhibition by a caspase 8 inhibitor demonstratesthe dependency on the extrinsic apoptotic pathway. The se-lective mortality does not correlate with the presence of deathor decoy receptors. These results suggest that TRAIL maybe an effective treatment for endometrial cancer and otherfemale reproductive cancers, with minimal secondary effectson healthy tissue.
Keywords Endometrium . Cervix . Fallopian . Ovariancancer
AbbreviationsTRAIL Tumour necrosis factor-related apoptosis-
inducing ligandTRAIL-R TRAIL receptorDR4/5 Death receptor 4/5DcR decoy receptorT TRAILC vehicle controlkDa kilodaltonFBS fetal bovine serumMTS 3-(4,5-Dimethylthiazol-2-yl)-2,5-
diphenyltetrazolium bromide
Introduction
Cancer of the female reproductive tract encompassesmalignancies of the uterine corpus (endometrial cancer),uterine cervix, ovary, vulva, Fallopian tube, vagina andother related genital tissue. According to estimated valuesfor 2005 by the American Cancer Society, cancer of thereproductive tract in the USA will account for 12% ofall female cancer incidence (79,480 of 662,870 estimatedcases) and approximately 15% of female cancer mortalities(28,910 of 275,000 estimated cases).
Springer

74 Apoptosis (2007) 12:73–85
Cancer of the uterine endometrium is the most commongynaecological cancer and predominantly affects post-menopausal women. The American Cancer Society reported40,320 cases which resulted in 7,090 deaths (3% all femalecancer deaths) in the USA in 2004. Fortuitously, threequarters of endometrial cancers are diagnosed early andsurgery at this stage achieves a 5 years overall survival rateof over 80%. However, there still exist a significant numberof advanced stage endometrial cancers where the currenttherapies offer poor survival rates. In line with first worldstatistical data, first hand experience from the authors hasobserved that in Chile, South America, approximately 500women die per year as a result of advanced disease status,despite receiving a combination of therapies; includingsurgery, radiotherapy, systemic chemotherapy and/or hor-mone therapy. Therefore, elucidating new strategies to treatadvanced uterine cancer remains paramount and will haveconsiderable impact on the treatment of gynecological cancerworldwide.
Diverse therapies against specific molecular targets areunder evaluation as treatment of epithelial tumors. Activationof the death receptors of the tumor necrosis factor receptorfamily have been shown to bring about apoptosis in a varietyof epithelial malignancies [1–4]. Death receptors activationoffers a promising method to induce apoptosis in cancer cellsand thereby an effective treatment of cancer. Tumor necrosisfactor-related apoptosis-inducing ligand (TRAIL) binds tothe death receptors, TRAIL-R1 and TRAIL-R2, and bringsabout caspase-mediated apoptosis [1]. Currently, TRAIL asa cancer treatment, is in Phase I studies and showing en-couraging results at doses that can be safely administered inhumans [5]. Furthermore, antibodies against the TRAIL-R1(also called DR4) and against TRAIL-R2 (also called DR5)are also showing promise in Phase I trails and Phase II Trialsrespectively [6, 7].
Apoptosis initiated by death ligands depends on the cell-extrinsic signaling pathway, which involves death receptorengagement, DISC formation, proteolytic activation of theapical caspases, caspase-8 and -10, and consequently, ac-tivation of effector caspases such as caspase-3, -6, and -7[2]. In certain types of cells, effector caspase activation re-quires amplification of DISC signals by engagement of thecell-intrinsic pathway, which is the main apoptosis signalingpathway that is activated by DNA-damaging agents such aschemo- and radiotherapeutic agents. A critical step in thecell-intrinsic pathway is the activation and translocation ofthe Bcl-2 family member Bax to the mitochondria, leadingto dissipation of the mitochondrial transmembrane poten-tial and cytochrome c release to the cytosol. This facilitatesassembly of the Apaf-1 apoptosome with recruitment andactivation of caspase-9, as an initiator caspase, and subse-quently the effector caspases [3].
TRAIL has been shown to induce apoptosis selectivelyin a variety of cancer cell lines but not in normal cells [1].Membrane localized TRAIL-R3 (also called decoy recep-tor 1, DcR1) and TRAIL-R4 (also called decoy receptor 2,DcR2) also bind TRAIL, but do not transmit an apoptotic sig-nal due to an absence of intracellular death domains [8, 9].Currently, the literature has failed to demonstrate a positivecorrelation between TRAIL-R1 & 2 in relation to TRAIL-R3 & 4 expression and response to TRAIL [10–12]. Animalstudies have also shown that TRAIL can cause regressionof cancer xenografts without significant toxicity to normaltissues [13–15]. However, to date few studies have exam-ined the response of TRAIL on normal and cancerous cellsoriginating from the human female reproductive tract.
The goal of this study was to investigate and characterizethe effect of TRAIL in several endometrial cancer cell linesand then examine if these effects are mirrored in normalepithelial cells of endometrial origin. Finally, we wish toinvestigate the effect of TRAIL in other primary culturedcancers and normal cells of the human female reproductivetract.
Materials and methods
Primary tissue
Primary tissue of human female reproductive tract was ob-tained with patient consent from the Gynaecology Depart-ment of the Hospital of the Pontificia Universidad Catolicade Chile and the Hospital Padre Hurtado, both in Santiago,Chile. Ethical approval for the study was obtained from boththe ethical committees of the Faculty of Medicine and theFaculty of Biological Sciences at the Pontificia UniversidadCatolica de Chile and the South Eastern Health Service ofSantiago, Chile.
Primary culture of human endometrial epithelial cells
Endometrial epithelial cell cultures were derived from tissuefrom pre-menopausal women undergoing routine hysterec-tomy or hysterectomy with bilateral salpingooophorectomyas described previously [16]. Briefly, immediately after hys-terectomy, approximately 1–2 cm3 of relevant tissue wasremoved from the endometrium at the endometrial cavity un-der sterile conditions and collected into warm saline serum.The stage of the menstrual cycle was determined from pa-tient menstrual history and endometrial histology. The tis-sue was washed twice in cold PBS and the endometriumseparated from the muscular layer. The tissue was dis-sected into 1mm3 cubes and incubated with collagenase-1(2 mg/ml, Worthington, Lorne Laboratories, Twyford, UK),
Springer

Apoptosis (2007) 12:73–85 75
hyaluronidase I (1 mg/ml, Sigma, Missouri, USA), DNAaseI (0.05 mg/ml, Boehringer Mannheim, Germany), antibiotic-antimycotic (Invitrogen, NY, USA) in calcium and magne-sium free HBSS solution (Invitrogen, NY, USA) for 1 ha 37◦C with agitation. After digestion the majority of theendometrial stromal and immune cells are visualised as in-dividual cells, while cells of endometrial epithelial originare present as aggregates. This digestion medium is filteredthough a 70 µm nylon mesh (PGC Scientific, Maryland,USA) and the remaining clumps (containing epithelial cells)washed in HBSS. Cells were recovered and subjected to asecond digestion for 15 min before being filtered througha 20 µm nylon mesh. Aggregated cells were recovered andincubated with trypsin (0.05%, Sigma-Aldrich Co., USA),EDTA (0.02%, Sigma-Aldrich Co., USA) and DNAase I(Boehringer Mannheim) for 5 min at 37◦C. The solution wascentrifuged at 3,000 rpm for 3 min, then washed in HBSS so-lution to remove residual trypsin. The cells were resuspendedin Dulbecco’s Modified Eagle Medium (DMEM)/F12 (In-vitrogen, NY, USA) with 10% fetal bovine serum (FBS;Invitrogen, NY, USA) containing antibiotic-antimycotic (In-vitrogen, NY, USA).
Primary culture of normal human ovarian epithelial cellsand ovarian cancer
Human ovarian tissue was obtained from patients undergo-ing oophorectomy for non malignant gynecological disor-ders. Tissue was washed with Hank’s balanced salt solutionwithout calcium, magnesium or phenol red (Invitrogen, NYUSA), containing penicillin and streptomycin. Slices con-taining surface epithelium and underlying stroma were cutin 2 mm thick and incubated in collagenase Type IV (Sigma-Aldrich Co., USA) for 30 min at 37◦C with agitation. Ovarianepithelial cells were scraped from the surface of the sliceswith a rubber policeman as previously described [17]. Thecells were suspended in Hanks and pelleted prior to sus-pension in DMEM/F12 medium supplemented with 10%FBS with penicillin and streptomycin. For the primary cul-ture of cancerous cells derived from ascites, approximately400 ml of ascitic fluid was obtained from two individualpatients presenting with cystadenocarcinoma of the ovary.The fluid was centrifuged at 2,000 g for 5 min, and 60%of the supernatant was replaced with the DMEM/F12 sup-plemented with 10% FBS and transferred to the cell culturedishes.
Primary culture of normal human cervical epithelial cells
Cervical tissue was obtained from patients undergoinghysterectomy for uterine leiomyoma. The patients, althoughrequiring uterine surgery, were otherwise healthy, aged
between 30 and 47 years with regular menstrual cycles andwere not recent users of hormonal contraceptives. Immedi-ately after removal of the uterus, the tissue incorporating theregion from the external cervical to the fundus was dissectedunder aseptic conditions. The surface mucosa was rinsedwith Hank’s solution (Sigma-Aldrich Co., USA) beforeendocervical biopsies were obtained. Each biopsy consistedof approximately 1 cm2 of epithelium and 2–3 mm of theunderlaying stroma. Epithelial secretory cells of the humancervix were isolated using a modification of a previouslydescribed protocol [18]. The endocervical biopsy wastreated with protease type 14 (0.05%), followed by trypsin(0.01%) (both from Sigma-Aldrich Co., USA), for 30 minat 4◦C. Each enzyme was dissolved in Hank’s solution.The tissue was washed in Hank’s solution and placed inWaymouth medium (Sigma-Aldrich Co., USA) containingpenicillin G (100 units/ml), streptomycin (100 µg/ml) andamphotericin B (1 µg/ml) (all from Sigma-Aldrich Co.,USA). At this time, the mucus was aspirated with a syringeand the biopsy epithelium was removed mechanically usinga scalpel. The epithelium was centrifuged at 300 g for 5 min,the pellet was resuspended in 1 ml Waymouth mediumand then dispersed mechanically with a 1 ml syringe. Thecells were added to the dishes and filled with Waymouthmedium (pH 7.4; Invitrogen, NY USA) supplemented with20% FBS, 1.2% L-glutamine, penicillin G (100 units/ml),streptomycin (100 µm/ml) and amphotericin B (1 µg/ml).Culture dishes were incubated in a controlled atmosphereof 5% CO2 at 37◦C.
Primary culture of normal human Fallopiantube epithelial cells
Human Fallopian tubes were obtained from patients under-going hysterectomy under benign conditions. Immediatelyafter removal the tubes were washed in calcium and mag-nesium free Hank’s solution (Invitrogen, NY, USA). Thefimbria and distal portion of the ampulla were removed fromthe Fallopian tube and placed in sterile DMEM/F12 mediumsupplemented with 25mM Hepes at pH 7.4. The tissue wascut into 4–8 mm pieces and transferred to a 5 mM EDTAin DMEM/F12 and incubated at 37◦C for 45 min. The cil-iated epithelium was then mechanically separated form therest of the tissue using fine forceps. The cell suspension waspassed through a tuberculin syringe into a centrifuge tubeand spun at 300 g for 5 min. The pellet was resuspendedin DMEM supplemented with 44 mM NaHCO3, 10% FBS,100U /ml penicillin and 100 ug/ml streptomycin (Invitrogen,NY, USA) at pH 7.4. The cell suspension was plated in tissueculture 10 cm2 Petri dishes (Falcon, BD Labware, NJ, USA)and incubated in a 5% CO2 atmosphere at 37◦C until theculture formed a monolayer of cells (approximately 4 days).
Springer

76 Apoptosis (2007) 12:73–85
Cell line cell culture
The endometrial cancer cell lines Ishikawa [19], HEC1a [20],the ovarian cancer line A2780 [21], and HeLa [22] cervicalcancer line were maintained in DMEM/F12 media supple-mented with 10% FBS. For protein and RNA experiments,cells were plated at 50% confluence in 10 cm2 Petri dishes(Falcon, BD Labware, NJ, USA) and in 6 cm2 Petri dishes forthe coagulation assay. The medium was changed to charcoal-treated medium containing 5% serum for 24 h before TRAILaddition.
TRAIL and chemotherapeutics
GST-TRAIL fusion protein was produced as described pre-viously [23] and stored in aliquots at −20◦C. An equal vol-ume of vehicle (DMEM/F12 medium) was added as a controlin each experiment. The chemotherapeutics drugs (doxoru-bicin, paclitaxel & cisplatin) were kindly given by the Can-cer Center affiliated to the Pontificia Universidad Catolicade Chile, and used at a final concentration of 5uM.
MTS assay
To assess TRAIL-mediated cytotoxicity, cells were plated at20.000 cells/well in 96-well microtiter plates and allowedto adhere to the plate overnight. GST-TRAIL fusion pro-teins at the indicated concentrations were added, and thecells were then incubated for an additional 18 h. Cell vi-ability was assessed by the MTS (3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) dye reduction assayas described previously [24]. The tetrapeptide caspase in-hibitor ZVAD-fmk, caspase inhibitors 2, 8 and 9 (EnzymeSystems Products., Livermore, CA) were resuspended inDMSO (Sigma-Aldrich Co., USA) and added to cells ata final concentration of 50 uM, 30 min before TRAIL.All experiments were performed in quadruplicate and re-peated at least three times. Statistical analysis was per-formed by Mann-Whitney analysis with significance set atp < 0.05. Bars represent ± SD (standard deviation) of themean.
Western blotting
Cells were harvested in cold PBS and the pellet resuspendedin lysis buffer (10 mM TRIS-HCl pH 7.4, 150 mM NaCl,0.5% Triton X-100) for 20 min at 4◦C. After this incubation,the lysate was centrifuged at 14,000 × g for 20 min at 4◦Cto separate membrane (pellet) and cytosolic (supernatant)fractions. Protein concentration was determined by Brad-ford assay. 100 µg of crude membrane extract was loadedin each lane, separated by 10% polyacrylamide gel elec-trophoresis in the presence of sodium dodecylsulfate, trans-
ferred to nitrocellulose membranes, and incubated overnightwith primary antibody (1:1000; PARP (Santa Cruz, USA),and β-actin (Santa Cruz, USA). Secondary antibody, goatanti-mouse/rabbit IgG secondary antibody coupled to hy-drogen peroxidase (1:5000, Bio-Rad Labs, CA, USA), wasapplied for one h at room temperature. The reaction wasdeveloped with chemiluminescence using ECL Western blotanalysis system (NEN, Western lightning, Perkin-Elmer).
Flow cytometry analysis
Cell cycle distribution and the detection of a sub-G1 apop-totic peak were analyzed by flow cytometry using propidiumiodide DNA staining [25]. Endometrial cancer cells wereharvested, centrifuged, washed and re-suspended in a coldsolution of 1 ml 1 × PBS and 4 ml 70% ethanol. The cellswere incubated overnight at 4◦C, washed in 1 × PBS andresuspended in a solution of 250 µl of propidium iodide(50 µg/ml) in 1 × PBS and 1 ul RNase (20 µg/ml). Cellswere incubated (protected from light) for 15 min at roomtemperature before analysis on a FACScan cytometer usingthe Cell Quest software (Becton Dickinson, Mountain View,CA, U.S.A).
DNA fragmentation assay
Cells (2 × 106) were washed in cold PBS and harvestedin cold lysis buffer containing 10 mM Tris-HCl, pH 8.0,150 mM NaCl, 2 mM MgCl2, 1 mM dithiothreitol (DTT),and 0.5% NP-40 on ice for 40 min. Lysates were centrifugedat 13,000 rpm and the pellets resuspended in cold buffercontaining 10 mM Tris-HCl, pH 8.0, 350 mM NaCl, 1 mMMgCl2, and 1 mM DTT before incubation on ice for 20 min.Lysates were then extracted once with phenol/chloroformand the DNA precipitated with 10 mM MgCl2 and 2.5 vol-umes of 100% ethanol overnight at –20◦C. DNA was col-lected by centrifugation at 14,000 g for 20 min, resuspendedin TE buffer (10 mM Tris-HCl, pH 8.0, 1 mM EDTA) plus0.1 mg/mL RNase A, and incubated at 37◦C for 1 h. Pro-teinase K (1 mg/mL) was added and the mixture was furtherincubated at 37◦C for 1 h. Fragmented DNA was separatedby electrophoresis in a 1.5% agarose gel and visualised withethidium bromide (0.5 µg/mL).
Measurement of caspase-3, 8 and 9 activity
Cultured cells were harvested and washed once in cold PBS.After a brief centrifugation (3,000 rpm, 5 min) cells wereincubated in lysis buffer containing 20 mM Hepes (pH 7.4),100 mM NaCl, 0.5% (v/v) NP-40 and10 mM DTT on ice for30 min. Following centrifugation at 13,000 rpm for 10 minat 4◦C, supernatants were collected, transferred to a 96 wellplate and corresponding substrate to Caspase 3, 8 and 9
Springer

Apoptosis (2007) 12:73–85 77
(Calbiochem-Novabiochem Corp., Ca., USA) added. Thesamples were incubated for 24 h at room temperature. Opti-cal density at 405 nm was measured using an ELISA platereader (EL310 Boots-Celtech). Casapase activity was ex-pressed as percentage in respect to control which was set at100%. Statistical analysis was performed by Mann-Whitneyanalysis with significance set at p < 0.05. Bars represent ±SD (standard deviation) of the mean.
RT-PCR
Total RNA was isolated using the Chomczynski method[26]. cDNA was generated using reverse transcriptase (Su-perscript II, Invitrogen, NY, USA). 40 PCR cycles were usedto detect the presence or absence of the transcripts. Pre-vious studies had demonstrated that this number of cycleswas saturating within this system (not shown). DR4 sense:5′-CCGCGGCCACACCCAGAAAGT-3′ DR4 antisense:5′-GTACATGGGAGGCAAGCAAACAAA-3′ (generates afragment of 415bp); DR5 sense: 5′GGGAGCCGCTCATGAGGAAGTTGG 3′ DR5 antisense: 3′GGCAAGTCTCTCTCCCAGCGTCTC5′ (generates a fragment of 181bp);DcR1 sense: 5′-GCCGGAA GTGTAGCAGGTG-3′ DcR1antisense: 5′-GAGATGTGTGGACCCCCA-3′ (generates afragment of 672bp); DcR2 sense: 5′-CCCCCGGCAGGACGAAGTT-3 DcR2 antisense: 5′-CTCCTCCGCTGCTGGGGTTTT-3′ (generates a fragment of 418bp) (BiosChile,Santiago, Chile) semi-quantitative PCR reactions were per-formed from cDNA generated from cell lines and primarycultured cells maintained under maintenance conditions (inthe absence of TRAIL), using Taq polymerase (Invitro-gen, NY, USA). As an internal control, primers amplify-ing a region of glyceraldehyde-3-phosphate dehydrogenase(GAPDH), were used to test the integrity of the startingcDNA as previously published [16].
Results
TRAIL reduces cellular viability of the human endometrialcancer cell line Ishikawa
To determine if TRAIL has potential as an agent against en-dometrial cancer, we tested cell viability of the well charac-terized endometrial cancer cell line Ishikawa in the presenceof TRAIL and currently used chemotherapeutic agents, dox-orubicin and paclitaxel. Cisplatin, a chemotherapeutic previ-ously reported to be ineffective in bringing about cell death inthis cell line was used as a negative control [27]. Cells weregrown in 5% charcoal treated FBS in DMEM/F12 mediumand were exposed to TRAIL, doxorubicin, paclitaxel andcisplatin for 18 h (overnight). As demonstrated in Fig. 1(A),TRAIL at a concentration of 200 ng/ml and at the time point
tested, is capable of reducing cellular viability as determinedusing the MTS assay to a greater extent that chemotherapeu-tic agents currently used in advanced endometrial disease. Tofirst establish the optimal dose of TRAIL, a concentration re-sponse (10 ng/ml to 1000 ng/ml) was performed in Ishikawacells. As demonstrated in Fig. 1(B), a decrease in cellularviability reached significant values at 200 ng/ml. Increasingconcentrations did not confer a further loss in viability. Hav-ing selected 200 ng/ml as a suitable concentration to bringabout apoptosis, we wished to determine the optimal expo-sure time of Ishikawa cells to the drug. Figure 1(C) showsthat a significant decrease in cellular viability occurs after18 h and this loss increases with time out to 72 h. TRAILtreatment for 18 h at 200 ng/ml was then chosen for all futureexperiments.
TRAIL mediates apoptosis in cell lines of humanendometrial cancer origin
To confirm that the reduction in cell viability measured bythe MTS assay (which measures mitochondrial redox po-tential) was due to apoptosis, protein extracted from bothIshikawa and an another well characterized endometrial can-cer cell line, HEC1a, was harvested after 18 h of 100, 200and 500 ng/ml TRAIL treatment. Apoptosis in the presenceof TRAIL in both endometrial cancer cell lines was con-firmed by the appearance of the 85 kDa cleaved PARP pro-tein (Fig. 2(A)). Furthermore, TRAIL at a concentration of100 ng/ml is sufficient to bring about apoptosis despite cellu-lar viability not reaching statistical significance as measuredby the MTS assay (Fig. 2(A)). Apoptosis was further con-firmed by the presence of DNA laddering (Fig. 2(B)). Due tothe discrepancy in the concentration of TRAIL which bringsabout cell death between the MTS assays and the PARPcleavage assay, we performed a concentration response inIshikawa cells and determined apoptosis by the presence ofa sub-G0/G1 peak by flow cytomety. This technique con-firmed that TRAIL brings about apoptosis in a concentrationdependent fashion, reducing the percentage of cells in eachphase of the cell cycle. Furthermore, flow cytometry con-firmed that 100 ng/ml of TRAIL can bring about apoptosisin this endometrial cancer cell line. Although not shown thepresence of apoptosis was confirmed by immunocytochem-istry utilizing an antibody against activate caspase 3 and bya characteristic change in morphology and the detachmentof cells in the presence of TRAIL.
TRAIL utilizes the intrinsic and extrinsic apoptoticpathways
To determine the pathway by which this time and concentra-tion of TRAIL was causing apoptosis we examined caspaseactivity. The involvement of caspases in TRAIL-mediated
Springer

78 Apoptosis (2007) 12:73–85
apoptosis is demonstrated by the maintenance of cell vi-ability in the presence of the tetrapeptide caspase inhibitorZVAD-fmk (Fig. 3(A)). To further dissect the apoptotic path-ways utilized by TRAIL, Ishikawa cells were exposed toinhibitors of caspase 2, 8 and 9 previous to the addition ofTRAIL. Figure 3(A) shows that caspase 2 is not utilizedby TRAIL, while caspase 8 inhibitor (an extrinsic pathwaymarker), abolishes TRAIL mediated apoptosis. However,caspase 9 inhibitor (intrinsic pathway marker) only partiallycounteracted the effect of TRAIL. To further consolidate theinvolvement of these caspases we assayed the activity ofcaspase 3, 8 and 9 in the presence and absence of TRAIL.
� Fig. 1 TRAIL reduces cellular viability of the human endometrialcancer cell line Ishikawa. (A) Cell viability was measured by the MTSassay in the presence of TRAIL (200 ng/ml), doxorubicin, paclitaxelor cisplatin (5 uM) and the data represented as percentage in respectto control. Standard deviation of the mean is shown from five repli-cates with the Mann-Whitney test demonstrating a = p < 0.05 withrespect to control (vehicle) and b = p < 0.05 in respect to TRAIL. (B)Concentration-response curve for Ishikawa cells with TRAIL (ng/mL)added for 18 h. Cell viability was measured by the MTS assay, and thedata represented as percentage in respect to control. Standard deviationof the mean is shown from a minimum of three experiments performed,each consisting of five replicates. Mann-Whitney test a = p < 0.05with respect to control (vehicle) (C) Time-course (in hours) measuringcell viability in response to TRAIL (200 ng/mL) in Ishikawa cells. Cellviability was measured by the MTS assay, and the data represented asa percentage in respect to control (vehicle). Standard deviation of themean is shown from a minimum of three experiments performed, eachconsisting of five replicates. Mann-Whitney test a = p < 0.05 withrespect to control, b = p < 0.05 with respect to previous time point
Figure 3(B) demonstrates an increase in the activity of eachcaspase in response to TRAIL.
TRAIL mediates apoptosis in cancerous but not normalcells of endometrial epithelial origin
As an effective cancer treatment requires drug specificity tocancer cells in comparison to normal tissue, we evaluated theeffect of TRAIL on primary cultured normal (non-cancerous)endometrial epithelial cells obtained from ten separate pa-tients undergoing routine hysterectomy. Written patient con-sent was obtained in all cases. Figure 4(A) demonstrates thatno PARP cleavage occurs in any of the primary cultures ob-tained from hysterectomies (three shown), while cleavageoccurs in the presence of TRAIL in the Ishikawa cancercell line. The lack of apoptosis in the presence of TRAIL(200 ng/ml) was also confirmed by absence of a sub G0/G1peak by flow cytometry (Fig. 4(B) and (C)) and a failure tosignificantly increase activated caspase 3 and 9 (Fig. 4(D)).No change in cell morphology was observed after TRAILtreatment (Fig. (E)).
TRAIL mediates apoptosis in cancerous but not normalcells of the human reproductive tract
As the concentration of TRAIL used in this study is causingapoptosis in both endometrial cancer cell lines, we evaluatedthe effect of TRAIL in other cancers of the reproductive tractand corresponding normal tissue. Figure 5 demonstrates that200 ng/ml of TRAIL for 18 h did not induce apoptosis in pri-mary cultured endometrial epithelium and stroma, ovarian,cervical or Fallopian tube cells of human origin. In contrastTRAIL brought about apoptosis in all cancer cell lines tested,along with primary cultured endometrial and ovarian tumorcells. A selection of these cultures were harvested for western
Springer

Apoptosis (2007) 12:73–85 79
Fig. 2 TRAIL mediates apoptosis in cell lines of human endometrialcancer origin. (A) Western blot analysis in Ishikawa and HEC-1A cellsdemonstrating the presence of cleaved PARP after the addition of vary-ing concentrations of TRAIL (ng/mL) for 18 h. (B) Apoptosis is furtherconfirmed by the presence of DNA fragmentation induced by TRAIL(200 ng/mL) for 18 h in both HEC-1A and Ishikawa endometrial cancer
cell lines in comparison to vehicle treated controls. (C) Flow cytome-try analysis demonstrating a reduction in each phase of the cell cycle(GO/G1 = M2, S = M3, G2/M = M4) and a corresponding increasein a sub GO/G1 apoptotic peak (M1) with increasing concentrations ofTRAIL. (D) Graphical representation of the changes in the percentageof cells in cell cycle phases and the appearance of a sub GO/G1 peak
Springer

80 Apoptosis (2007) 12:73–85
Fig. 3 TRAIL utilizes theintrinsic and extrinsic apoptoticpathways. (A) Cell viability wasmeasured by the MTS assay,and the data represented aspercentage in respect to control(vehicle). Standard deviation ofthe mean is shown from aminimum of three experimentsperformed, each consisting fivereplicates. The caspaseinhibitors 2, 8, 9 and z-vad (all50 uM) were added 30 minprevious to TRAIL (T) addition200 ng/mL for 18 h.Mann-Whitney test a =p < 0.05 with respect to control,b = p < 0.05 with respect toTRAIL. (B) Caspase activityassays (caspase 3, 8 & 9) inIshikawa cells after the additionof TRAIL 200 ng/mL for 18 h.Cells in the absence of TRAILare set at 100%. Mann-Whitneytest ∗ = p < 0.05 with respectto control
blot analysis, which confirmed the absence of cleaved PARPin all primary cultured cells originating from non-cancerousreproductive tissue (Fig. 5(B)).
Apoptosis in response to TRAIL does not correlatewith the presence of TRAIL receptors
In an attempt to understand the mechanism of preferentialTRAIL induced apoptosis in cancerous but not normal tis-
sue we examined the levels of TRAIL receptors (DR4, DR5,DcR1 and DcR2) mRNA. Figure 6 demonstrates that celllines and primary cultured cells examined possessed both oreither DR4 or DR5. All cells tested possessed both DcR2and the housekeeping RNA GAPDH. Although mRNA lev-els of DR4 and DcR1 showed the greatest variability, noobvious correlation was detected between the presence orabsence of these receptors and apoptosis in response toTRAIL.
Springer
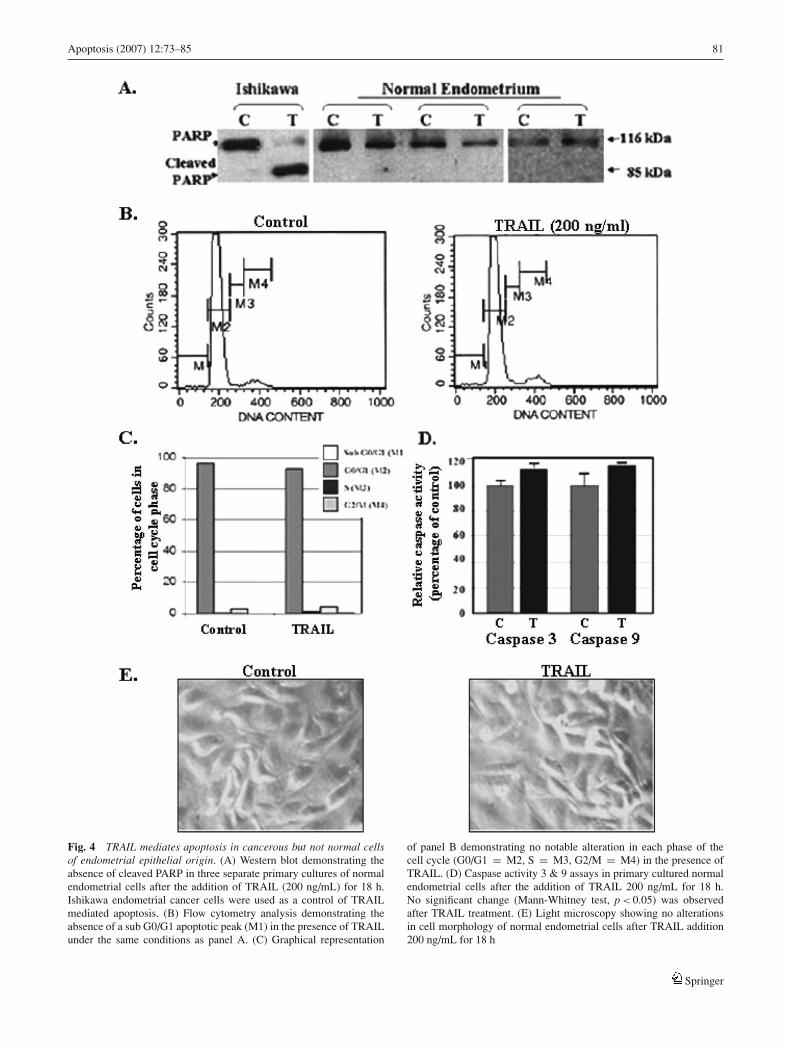
Apoptosis (2007) 12:73–85 81
Fig. 4 TRAIL mediates apoptosis in cancerous but not normal cellsof endometrial epithelial origin. (A) Western blot demonstrating theabsence of cleaved PARP in three separate primary cultures of normalendometrial cells after the addition of TRAIL (200 ng/mL) for 18 h.Ishikawa endometrial cancer cells were used as a control of TRAILmediated apoptosis. (B) Flow cytometry analysis demonstrating theabsence of a sub G0/G1 apoptotic peak (M1) in the presence of TRAILunder the same conditions as panel A. (C) Graphical representation
of panel B demonstrating no notable alteration in each phase of thecell cycle (G0/G1 = M2, S = M3, G2/M = M4) in the presence ofTRAIL. (D) Caspase activity 3 & 9 assays in primary cultured normalendometrial cells after the addition of TRAIL 200 ng/mL for 18 h.No significant change (Mann-Whitney test, p < 0.05) was observedafter TRAIL treatment. (E) Light microscopy showing no alterationsin cell morphology of normal endometrial cells after TRAIL addition200 ng/mL for 18 h
Springer

82 Apoptosis (2007) 12:73–85
Fig. 5 TRAIL mediates apoptosis in cancerous but not normal cellsof the human reproductive tract. (A) MTS cell viability assays afterTRAIL (200 ng/mL) addition for 18 h in cultured normal endometrialepithelium, normal endometrial, sarcoma (cancer originating in the en-dometrial stromal compartment), two cultured epithelial endometrialcancers, ovarian cells, two metastatic ovarian cancers, an ovarian can-cer cell line (A2780), normal cultured cervical cells, a cervical cancercell line (HeLa) and cultured normal epithelial cells of the Fallopiantube. Data represent percentage in respect to control. Standard deriva-
tion of the means is shown, each consisting of five replicates. All datashown are in the presence of TRAIL were cell viability is set at 100%in the presence of vehicle (first bar, C). Statistical significance (∗) inrespect to vehicle treated control for each cell type was set at p < 0.05using the Mann-Whitney test. (B) Western blot analysis demonstratingthe presence of cleaved PARP (apoptosis) in cancerous but not nor-mal cell cultures after the addition of TRAIL (200 ng/mL) for 18 h inparallel cultures of the cells used in panel A
Discussion
Herein, we demonstrate that endometrial cancer cell linesIshikawa and HEC1a but not normal endometrial epithe-lium in primary culture undergo apoptosis in the presenceof TRAIL. Moreover, cell viability was reduced by TRAILto a greater extent than doxorubicin and placitaxel, twochemotherapeutic agents currently used to treat advanced en-dometrial cancer. Furthermore, this selective apoptosis waspresent in cell lines and cells derived from cancerous andnormal ovarian, Fallopian and cervical tissue. We feel thisinformation has significant clinical relevance as endometrialcancer accounts for 3% all female cancer deaths and ovar-ian cancer remains the deadliest gynaecologic malignancy.Recently, Dolcet and colleagues [28] reported that Ishikawacells were resistant to TRAIL mediated apoptosis. Using thesame concentration of TRAIL our results confirm this, how-ever Ishikawa cells also show a 20–25% reduction in cell via-bility at concentration range between 200 ng/ml and 1 ug/mland an incubation time of less than 24 h (Fig. 1(A)). Compar-
ing the results obtained with the currently used chemother-apeutic agents, we feel that this level of cell death is highlysignificant to cancer treatment. Furthermore, at similar con-centrations, but at longer periods of incubation, we ob-served a significant increase in cell death induced by TRAIL(about 60% reduction of cell viability at 72 h of incubation,Fig. 1(C)) suggesting that in certain cell lines, resistancecould be overcome by prolonged TRAIL exposure.
The mechanism of TRAIL action is through both the ex-trinsic, as shown by caspase 8 activation and inhibition, andby the intrinsic pathway, as shown by caspase 9 activation andinhibition. The failure of caspase 9 inhibitor to completelyreverse TRAIL mediated apoptosis suggests that the intrin-sic pathway is involved, but is not acting alone in mediatingthe full apoptotic potential of TRAIL. Despite the mecha-nism still being elusive, the encouraging observation is thatTRAIL did not bring about apoptosis or a loss in cell viabilityin any normal primary cultured cells of endometrial, Fallop-ian tube, cervical or ovarian origin tested. In two endometrialadenocarcinomas in culture, TRAIL mediated a loss in cell
Springer

Apoptosis (2007) 12:73–85 83
Fig. 6 Apoptosis in response to TRAIL does not correlate with thepresence of TRAIL receptors. Conventional RT-PCR analysis of themRNA levels of TRAIL death receptors (DR4 and DR4), decoy re-ceptors (DcR1 and DcR2) and the mRNA of the housekeeping gene incultured normal endometrial epithelium, normal endometrial, sarcoma
(cancer originating in the endometrial stromal compartment), an ovar-ian cancer cell line (A2780), normal cultured cervical cells, a cervicalcancer cell line (HeLa) and cultured normal epithelial cells of the Fal-lopian tube. The presence or absence of TRAIL induced apoptosis ineach culture is marked by a “tick” or by an “x”, respectively
viability in only one culture. The non-responsive endome-trial tumor was of a lower stage (stage 1 as opposed to stageIII), and this may be due to lower stage cancers (well dif-ferentiated) maintaining many characteristics of the tissue oforigin. The ovarian tumors, each removed at advanced stages(isolated from ascites) both showed significant cell death inresponse to TRAIL. However, due to low sample numbers, noconclusion can be drawn at the present time and thus furtherexperiments are currently ongoing to correlate reproductivecancer stage/grade and response to TRAIL. To understandthe mechanisms by which TRAIL brings about apoptosis incancerous but not normal tissue of the reproductive tract,we analyzed the levels of death domain containing TRAILreceptors. In accordance with several other publications wecould not demonstrate a correlation between TRAIL recep-tors and TRAIL induced apoptosis [10–12]. Interestingly,the three cancerous endometrial samples (two cell lines andan endometrial sarcoma) possessed both the DR4 and DR5receptors, while corresponding cells from a non-cancerouspatient showed a lack of DR5 in the epithelium and the pres-ence of DR4 in both the stroma and the epithelium. This lat-ter result is against current thinking as several reports haveshown that a lack of DR4 expression, possibly by epige-
netic silencing, correlates with resistance to TRAIL-inducedapoptosis [29, 30]. However, other reports in cancer cellshave suggested that DR5 is the preferential route for TRAILinduced apoptosis [31, 32]. Although we demonstrate onlythe mRNA levels of DR5, which may not necessarily con-vert to protein expression, our results are at odds with thisreport and suggests that multiply factors, including DR4 andDR5 are responsible for the insensitivity of normal cells toTRAIL [33]. As with the death receptors, the levels of decoyreceptors also varied in cell lines, cultured primary tumorsand normal cells of the reproductive tract, tested. AlthoughDcR1 manifested the greatest variability between the sam-ples and was absent or reduced in three of the four cancercell lines tested and present in all TRAIL resistant cellstested, no correlation was found between TRAIL mediatedapoptosis and decoy receptor presence. While the absenceof DR4 and DcR1 may be a major factors and diagnostictools in cancer, other parallel mechanisms exist in whichtumors can overcome this sensitivity to TRAIL. To addressthis issue we specifically choose Ishikawa and HEC1a cellsas both endometrial adenocarcinoma cells are derived fromsimilar cancer stages, are phenotypically similar and expressreceptors for ovarian hormones. However, each cell line is
Springer

84 Apoptosis (2007) 12:73–85
known to be cancerous due to specific changes; Ishikawacells have native p53, EGFR, RAS and inactive PTEN whileHEC1a cells have mutated p53, RAS overexpress EGFRand have and wild-type PTEN [34–38]. Although these cellslines show slight differences in response to TRAIL, bothare similarly sensitive to TRAIL mediated cell death. Thisdemonstrates that at least in our endometrial cancer model,p53, PTEN, EGFR or RAS are not solely responsible for thedifferential sensitivity between cancerous and normal cells.Possible future targets in the search of this mechanism maybe the presence or absence of tyrosine kinase membranereceptors, (such as EGFR or Erb-B2) which may maintainanti-apoptotic factors (such as AKT) elevated [39, 40] orthe activation of the NF-kappaB pathway [41]. Some reportshave suggested that osteoprotegerin (OPG) is secreted at highlevels by non tumor cells [32]. The relationship between IAP,FLICE inhibitory protein, SMAC/Diablo, Bcl-2 and EphB4status are equally as contentious and thus further work is nec-essary to elucidate the selective nature of TRAIL-mediatedapoptosis [42–45].
Herein, we demonstrate for the first time that TRAILat concentrations which kill cancerous cells does not bringabout apoptosis or alter cell viability in normal cultures ofendometrium, ovary, cervix or Fallopian tube. This suggeststhat this agent may be an effective treatment for female repro-ductive cancers. Experiments currently ongoing in immuno-suppressed (nude) mice grafted with endometrial cancer celllines will confirm if our observations in vitro are reflectedin vivo. However, the nature of TRAIL to be used in thissetting is a main issue. Previously published safety studiesusing mice, monkeys, and chimpanzees did not show anydetectable toxic side effects [14, 15, 44]. Warning bells wereheard when human brain cells, keratinocytes and hepatocytessuffered cell death upon exposure to TRAIL, suggesting thata potential systemic toxicity would be observed in clinical tri-als [46, 47]. Assessment of these experiments suggested thatthis toxicity could be dependent on the form and source ofrecombinant TRAIL utilized. For example, toxicity in hepa-tocytes and keratinocytes was observed with the soluble ver-sion of the ligand, which contains a trimerizing leucine zip-per [44, 48]. Furthermore, in keratinocytes, toxicity was alsoobserved with the polyhistidine-tagged version of TRAIL.However, when the non-tagged soluble version was used notoxicity was observed in either cell types [49]. The intra-venous administration of non-tagged TRAIL was well tol-erated both in cynomolgus monkeys as chimpanzees, withno dangerous symptoms detected in laboratory parameters ororgan histology. In our experiments, presented herein, a GSTtagged TRAIL version was used. Although this form is notsoluble and is therefore unsuitable for use in a clinical setting,we have previously shown that this GST-TRAIL induces sim-ilar effects in terms of apoptosis as does the aforementionedrecombinant non-tagged soluble version of TRAIL [50].
Conclusion
We demonstrate that endometrial, ovarian and cervical can-cer cell lines, cultured primary tumors of endometrial (ep-ithelial and stromal) and ovarian origin, undergo apoptosis inthe presence of TRAIL. In contrast, normal (non-cancerous)primary cultures of corresponding tissue obtained from hys-terectomy with bilateral salpingooophorectomy presented noapoptosis or loss in viability. Therefore, we conclude that theselective mediation of apoptosis in cancerous but not healthytissue of the reproductive tract, suggests the use of recombi-nant non-tagged soluble TRAIL as an effective treatment offemale reproductive malignancies.
Acknowledgments We gratefully acknowledge the kind donation ofthe caspase inhibitors and the active caspase 3 antibody from Dr RicardoMoreno at the Pontificia Universidad Catolica de Chile.
References
1. Marsters SA, Pitti RA, Sheridan JP, Ashkenazi A (1999) Controlof apoptosis signalling by Apo2 ligand. Recent Prog Horm Res54:225–234
2. Gibson SB (2004) Epidermal growth factor and TRAIL interac-tions in epithelial-derived cells. Vitam Horm 67:207–227
3. Wajant H, Gerspach J, Pfizenmaier K (2005) Tumor therapeuticsby design: targeting and activation of death receptors. CytokineGrowth Factor Rev 16:55–76
4. de Thonel A, Eriksson JE (2005) Regulation of death receptors-Relevance in cancer therapies. Toxicol Appl Pharmaco 207:123–132
5. Ling J, Herbst RS, Mendelson DS et al (2006) Apo2L/TRAIL phar-macokinetics in a phase 1a trial in advanced cancer and lymphomaJournal of Clinical Oncology (Meeting Abstracts); ASCO AnnualMeeting Proceedings Part I 24: 3047
6. Kanzler S, Trarbach T, Heinemann V et al (2005) Results of aPhase 2 trial of HGS-ETR1 (agonistic human monoclonal anti-body to TRAIL receptor 1) in subjects with relapsed or refractorycolorectal cancer (CRC). ECCO 13 – the European Cancer Con-ference, Abstract 630
7. Bonomi P, Greco FA (2005). Results of a Phase 2 trial of HGS-ETR1 (agonistic human monoclonal antibody to TRAIL receptor1) in subjects with relapsed/recurrent non-small cell lung cancer.11th World Conference on Lung Cancer: Abstract 1851
8. Sheridan JP, Marsters SA, Pitti RM et al (1997) Control of TRAIL-induced apoptosis by a family of signaling and decoy receptors.Science 277:818–21
9. Baetu TM, Hiscott J (2002) On the TRAIL to apoptosis. CytokineGrowth Factor Rev 13:199–207
10. Cuello M, Ettenberg SA, Clark AS et al (2001) Down-regulationof the erbB-2 receptor by trastuzumab (herceptin) enhances tumornecrosis factor-related apoptosis-inducing ligand-mediated apop-tosis in breast and ovarian cancer cell lines that overexpress erbB-2.Cancer Res 61:4892–4900
11. Vignati S, Codegoni A, Polato F, Broggini M (2002) Trail activity inhuman ovarian cancer cells: potentiation of the action of cytotoxicdrugs. Eur J Cancer 38(1):177–183
12. Leverkus M, Neumann M, Mengling T et al (2000) Regulation oftumor necrosis factor-related apoptosis-inducing ligand sensitiv-ity in primary and transformed human keratinocytes. Cancer Res60:553–559
Springer

Apoptosis (2007) 12:73–85 85
13. Roth W, Isenmann S, Naumann U et al (1999) LocoregionalApo2L/TRAIL eradicates intracranial human malignant gliomaxenografts in athymic mice in the absence of neurotoxicity.Biochem Biophys Res Commun 265:479–483
14. Ashkenazi A, Pai RC, Fong S et al (1999) Safety and antitumoractivity of recombinant soluble Apo2 ligand. J Clin Invest 104:155–162
15. Walczak H, Miller RE, Ariail K et al (1999) Tumoricidal activityof tumor necrosis factor-related apoptosis-inducing ligand in vivo.Nat Med 5:157–163
16. Kato S, Pinto M, Carvajal A et al (2005) Tissue factor is regu-lated by epidermal growth factor in normal and malignant humanendometrial epithelial cells. Thromb Haemost 94:444–453
17. Auersperg N, Siemens CH, Myrdal SE (1984) Human ovariansurface epithelium in primary culture. In Vitro 20:743–755
18. Verdugo P, Langley L, Aitken ML, Villalon M (1990) Developmentof an in vitro model of primate cervical goblet cells. Biorheology27:465–470
19. Nishida M, Kasahara K, Kaneko M, Iwasaki H, Hayashi K (1985)Establishment of a new human endometrial adenocarcinoma cellline, Ishikawa cells, containing estrogen and progesterone recep-tors. Nippon Sanka Fujinka Gakkai Zasshi 37:1103–1111
20. Kuramoto H (1972) Studies of the growth and cytogenetic prop-erties of human endometrial adenocarcinoma in culture and itsdevelopment into an established line. Acta Obstet Gynaecol Jpn19:47–58
21. Louie KG, Behrens BC, Kinsella TJ et al (1985) Radiation sur-vival parameters of antineoplastic drug-sensitive and resistant hu-man ovarian cancer cell lines and their modification by buthioninesulfoximine. Cancer Res 45:2110–2115
22. Eagle H (1955) Propagation in a fluid medium of a human epider-moid carcinoma, strain KB. Proc Soc Exp Biol Med 89:362–364
23. Cuello M, Ettenberg SA, Nau MM, Lipkowitz S (2001) Synergisticinduction of apoptosis by the combination of trail and chemother-apy in chemoresistant ovarian cancer cells. Gynecol Oncol 81:380–390
24. KeaneMM, Ettenberg SA, Lowrey GA, Russell EK, Lipkowitz S(1996) Fas expression and function in normal and malignant breastcell lines. Cancer Res 56:4791–4798
25. Li L, Heldin NE, Grawe J, Ulmsten U, Fu X (2004) Inductionof apoptosis or necrosis in human endometrial carcinoma cells by2-methoxyestradiol. Anticancer Res 24:3983–3990
26. Chomczynski P, Sacchi N (1987) Single-step method of RNA iso-lation by acid guanidinium thiocyanate-phenol-chloroform extrac-tion. Anal Biochem 162:156–159
27. Thigpen T, Vance RB, Khansur T (1995) The platinum com-pounds and paclitaxel in the management of carcinomas of theendometrium and uterine cerviz. Semin Oncol 22:67–75
28. Dolcet X, Llobet D, Pallares J et al (2005) FLIP is frequentlyexpressed in endometrial carcinoma and has a role in resistance toTRAIL-induced apoptosis. Lab Invest 85:885–894
29. Horak P, Pils D, Haller G et al (2005) Contribution of epigeneticsilencing of tumor necrosis factor-related apoptosis inducing ligandreceptor 1 (DR4) to TRAIL resistance and ovarian cancer. MolCancer Res 3:335–343
30. Li CM, Peng ZL, Yang KX (2005) The expression of TRAIL,TRAIL-R (DR4, DcR1) and Fas-L in patients with ovarian cancerSichuan Da Xue Xue Bao Yi Xue Ban 36:341–343
31. Kelley RF, Totpal K, Lindstrom SH et al (2005) Receptor-selectivemutants of apoptosis-inducing ligand 2/tumor necrosis factor-related apoptosis-inducing ligand reveal a greater contribution ofdeath receptor (DR) 5 than DR4 to apoptosis signaling. J BiolChem 280:2205–2212
32. Vincent H, Claire PP, Marion T et al (2006) Sensitivity of prostatecells to TRAIL-induced apoptosis increases with tumor pro-gression: DR5 and caspase 8 are key players. Prostate 66:987–995
33. Reesink-Peters N, Hougardy BM, Van Den Heuvel FA et al (2005)Death receptors and ligands in cervical carcinogenesis: an im-munohistochemical study. Gynecol Oncol 96:705–713
34. Lelle RJ, Talavera F, Gretz et al (1993) Epidermal growth factorreceptor expression in three different human endometrial cancercell lines. Cancer 72:519–525
35. Yaginuma Y, Westphal H (1991) Analysis of the p53 gene in humanuterine carcinoma cell lines. Cancer Res 51: 6506–6509
36. Matsushima-Nishiu M, Unoki M, Ono K et al (2001) Growth andgene expression profile analysis of endometrial cancer cell linesexpressing exogenous PTEN. Cancer Res 61:3741–3749
37. Lilja JF, Wu D, Reynolds RK et al (2001) Growth suppressionactivity of the PTEN tumor suppressor gene in human endometrialcancer cells. Anticancer Res 21:1969–1974
38. Boyd J, Risinger JI (1991) Analysis of oncogene alterations inhuman endometrial carcinoma: prevalence of ras mutations. MolCarcinog 4:189–195
39. Nesterov A, Lu X, Johnson M et al (2001) Elevated AKT activityprotects the prostate cancer cell line LNCaP from TRAIL-inducedapoptosis. J Biol Chem 276:10767–10774
40. Tanaka Y, Gavrielides MV, Mitsuuchi Y, Fujii T, Kazanietz MG(2003) Protein kinase C promotes apoptosis in LNCaP prostatecancer cells through activation of p38 MAPK and inhibition of theAkt survival pathway. J Biol Chem 278:33753–33762
41. Keane MM, Rubinstein Y, Cuello M et al (2000) Inhibition ofNF-kappaB activity enhances TRAIL mediated apoptosis in breastcancer cell lines. Breast Cancer Res Treat 64:211–219
42. LeBlanc H, Lawrence D, Varfolomeev E et al (2002) Tumor-cellresistance to death receptor–induced apoptosis through mutationalinactivation of the proapoptotic Bcl-2 homolog Bax. Nat Med8:274–281
43. Sharp DA, Lawrence DA, Ashkenazi A (2005) Selective knock-down of the long variant of cellular FLICE inhibitory protein aug-ments death receptor-mediated caspase-8 activation and apoptosis.J Biol Chem 280:19401–19409
44. Leverkus M, Neumann M, Mengling T et al (2000) Regulation oftumor necrosis factor-related apoptosis-inducing ligand sensitiv-ity in primary and transformed human keratinocytes. Cancer Res60:553–559
45. Kumar SR, Singh J, Xia G et al (2006) Receptor tyrosine kinaseEphB4 is a survival factor in breast cancer. Am J Pathol 169:279–293.
46. Jo M, Kim TH, Seol DW (2000) Apoptosis induced in normalhepatocytes by tumor necrosis factor-realted apoptosis-inducingligand. Nat Med 6:564–567
47. Nitsch R, Bechmann I, Deisz RA (2000) Human brain-cell death in-duced by tumor-necrosis-factor-related apoptosis-inducing-ligand(TRAIL). Lancet 356:827–828
48. Lawrence D, Shahrokh Z, Marsters S (2001) Differential hepa-tocyte toxicity of recombinant Apo2L/TRAIL versions. Nat Med7:383–385
49. Qin J, Chaturvedi V, Bonish B, Nickoloff BJ (2001) Avoidingpremature apoptosis of normal epithelial cells. Nat Med 7:385–386
50. Cuello M, Coats AO, Darko I et al (2004) N-(4-hydroxyphenyl)retinamide (4HPR) enhances TRAIL-mediated apoptosis throughenhancement of a mitochondrial-dependent amplification loopin ovarian cancer cell lines. Cell Death Differ 11(5):527–541
Springer




















