
Liver NK cells expressing TRAIL are toxic against self hepatocytes in mice
11
Liver NK Cells Expressing TRAIL Are Toxic Against Self Hepatocytes in Mice Makoto Ochi, Hideki Ohdan, Hiroshi Mitsuta, Takashi Onoe, Daisuke Tokita, Hidetaka Hara, Kohei Ishiyama, Wendy Zhou, Yuka Tanaka, and Toshimasa Asahara Although it is known that activation of natural killer (NK) cells causes liver injury, the mecha- nisms underlying NK cell-induced killing of self-hepatocytes are not clear. We demonstrated that liver NK cells have cytotoxicity against normal syngeneic hepatocytes in mice. Polyinosinic- polycytidylic acid (poly I:C) treatment enhanced hepatocyte toxicity of liver NK cells but not that of spleen NK cells. Unlike NK cells in other tissues, approximately 30%– 40% of liver NK cells constitutively express tumor necrosis factor-related apoptosis-inducing ligand (TRAIL). An in vitro NK cell cytotoxic assay revealed that hepatocyte toxicity of liver NK cells from both naı ¨ve and poly I:C-treated mice was inhibited partially by an anti-TRAIL monoclonal antibody (mAb) alone and completely by the combination with anti-Fas ligand (FasL) mAb and a perforin inhibitor, concanamycin A, indicating contribution of TRAIL to NK cell-mediated hepatocyte toxicity. The majority of TRAIL NK cells lacked expression of Ly-49 inhibitory receptors recognizing self-major histocompatibility complex class I, indicating a propensity to targeting self-hepatocytes. Poly I:C treatment significantly upregulated the expression of Ly-49 receptors on TRAIL – NK cells. This might be a compensatory mechanism to protect self-class I-expressing cells from activated NK cell-mediated killing. However, such compensatory alteration was not seen at all in the TRAIL NK cell fraction. Thus, liver TRAIL NK cells have less capacity for self-recognition, and this might be involved in NK cell-dependent self-hepatocyte toxicity. In conclusion, our findings are consistent with a model in which TRAIL-expressing NK cells play a critical role in self-hepatocyte killing through poor recognition of MHC. (HEPATOLOGY 2004;39: 1321–1331.) N atural killer (NK) cells are thought to provide a first line of defense against invading infectious microbes by exerting an effector function with- out the necessity for priming. 1,2 In the liver, both conven- tional NK cells and NKT cells are quite abundant, in contrast to the relatively small percentage of these cells in the peripheral lymphatics. 2–6 Although the underlying reason for this anatomically biased distribution has not been fully elucidated, NK and NKT cells have been shown to mediate cytolytic activity and to secrete cyto- kines, enabling them to perform distinct functions in re- sponse to infected cells and neoplastic cells. 4,5,7 Under various pathophysiological conditions, such as viral hep- atitis, bacterial infection, and autoimmune liver disease, liver NK cells are activated by various cytokines, such as interleukin (IL)-2, IL-12, interferon (IFN)-/, and IFN-, secreted from a wide variety of cell types, includ- ing dendritic cells, macrophages, and NKT cells, and this activation enhances their cytotoxic capacity, 1,2,8 Activated liver NK cells target not only infected or transformed hepatocytes but also intact hepatocytes in a nonspecific fashion. 8 –10 Cytotoxicity of liver NK cells against normal hepatocytes seems excessive self-defense from the point of view of innate immunity. In general, the presence of self-major histocompatibility complex (MHC) class I molecules protects target cells from lysis mediated by NK cells via binding to inhibitory recep- Abbreviations: NK, natural killer; poly I:C, polyinosinic-polycytidylic acid; TRAIL, tumor necrosis factor-related apoptosis-inducing ligand; mAb, monoclonal antibody; FasL, Fas ligand; IL, interleukin; IFN, interferon; MHC, major histo- compatibility complex; SCID, severe combined immunodeficient disease; WT, wild-type; i.p., intraperitoneal; ALT, alanine aminotransferase; FITC, fluorescein isothiocyanate; PE, phycoerythrin; TCR, T cell receptor; FCM, flow cytometry. From the Department of Surgery, Division of Frontier Medical Science, Pro- grams for Biomedical Research, Graduate School of Biomedical Sciences, Hiroshima University, Hiroshima, Japan. Received June 5, 2003; accepted December 22, 2003. Supported by Grant-in-Aid for Creation of Innovations through Business-Aca- demic-Public Sector Cooperation. Address reprint requests to: Hideki Ohdan, MD, PhD, Department of Surgery, Division of Frontier Medical Science, Programs for Biomedical Research, Graduate School of Biomedical Sciences, Hiroshima University, 1-2-3 Kasumi, Minami-ku, Hiroshima 734-8551, Japan. E-mail: [email protected]; fax: 81 82 257 5224. Copyright © 2004 by the American Association for the Study of Liver Diseases. Published online in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/hep.20204 1321
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Liver NK cells expressing TRAIL are toxic against self hepatocytes in mice

Liver NK Cells Expressing TRAILAre Toxic Against Self Hepatocytes
in MiceMakoto Ochi, Hideki Ohdan, Hiroshi Mitsuta, Takashi Onoe, Daisuke Tokita, Hidetaka Hara, Kohei Ishiyama,
Wendy Zhou, Yuka Tanaka, and Toshimasa Asahara
Although it is known that activation of natural killer (NK) cells causes liver injury, the mecha-nisms underlying NK cell-induced killing of self-hepatocytes are not clear. We demonstrated thatliver NK cells have cytotoxicity against normal syngeneic hepatocytes in mice. Polyinosinic-polycytidylic acid (poly I:C) treatment enhanced hepatocyte toxicity of liver NK cells but not thatof spleen NK cells. Unlike NK cells in other tissues, approximately 30%–40% of liver NK cellsconstitutively express tumor necrosis factor-related apoptosis-inducing ligand (TRAIL). An invitro NK cell cytotoxic assay revealed that hepatocyte toxicity of liver NK cells from both naı̈veand poly I:C-treated mice was inhibited partially by an anti-TRAIL monoclonal antibody (mAb)alone and completely by the combination with anti-Fas ligand (FasL) mAb and a perforininhibitor, concanamycin A, indicating contribution of TRAIL to NK cell-mediated hepatocytetoxicity. The majority of TRAIL� NK cells lacked expression of Ly-49 inhibitory receptorsrecognizing self-major histocompatibility complex class I, indicating a propensity to targetingself-hepatocytes. Poly I:C treatment significantly upregulated the expression of Ly-49 receptorson TRAIL– NK cells. This might be a compensatory mechanism to protect self-class I-expressingcells from activated NK cell-mediated killing. However, such compensatory alteration was notseen at all in the TRAIL� NK cell fraction. Thus, liver TRAIL� NK cells have less capacity forself-recognition, and this might be involved in NK cell-dependent self-hepatocyte toxicity. Inconclusion, our findings are consistent with a model in which TRAIL-expressing NK cells play acritical role in self-hepatocyte killing through poor recognition of MHC. (HEPATOLOGY 2004;39:1321–1331.)
Natural killer (NK) cells are thought to provide afirst line of defense against invading infectiousmicrobes by exerting an effector function with-
out the necessity for priming.1,2 In the liver, both conven-tional NK cells and NKT cells are quite abundant, in
contrast to the relatively small percentage of these cells inthe peripheral lymphatics.2–6 Although the underlyingreason for this anatomically biased distribution has notbeen fully elucidated, NK and NKT cells have beenshown to mediate cytolytic activity and to secrete cyto-kines, enabling them to perform distinct functions in re-sponse to infected cells and neoplastic cells.4,5,7 Undervarious pathophysiological conditions, such as viral hep-atitis, bacterial infection, and autoimmune liver disease,liver NK cells are activated by various cytokines, such asinterleukin (IL)-2, IL-12, interferon (IFN)-�/�, andIFN-�, secreted from a wide variety of cell types, includ-ing dendritic cells, macrophages, and NKT cells, and thisactivation enhances their cytotoxic capacity,1,2,8
Activated liver NK cells target not only infected ortransformed hepatocytes but also intact hepatocytes in anonspecific fashion.8–10 Cytotoxicity of liver NK cellsagainst normal hepatocytes seems excessive self-defensefrom the point of view of innate immunity. In general, thepresence of self-major histocompatibility complex(MHC) class I molecules protects target cells from lysismediated by NK cells via binding to inhibitory recep-
Abbreviations: NK, natural killer; poly I:C, polyinosinic-polycytidylic acid;TRAIL, tumor necrosis factor-related apoptosis-inducing ligand; mAb, monoclonalantibody; FasL, Fas ligand; IL, interleukin; IFN, interferon; MHC, major histo-compatibility complex; SCID, severe combined immunodeficient disease; WT,wild-type; i.p., intraperitoneal; ALT, alanine aminotransferase; FITC, fluoresceinisothiocyanate; PE, phycoerythrin; TCR, T cell receptor; FCM, flow cytometry.
From the Department of Surgery, Division of Frontier Medical Science, Pro-grams for Biomedical Research, Graduate School of Biomedical Sciences, HiroshimaUniversity, Hiroshima, Japan.
Received June 5, 2003; accepted December 22, 2003.Supported by Grant-in-Aid for Creation of Innovations through Business-Aca-
demic-Public Sector Cooperation.Address reprint requests to: Hideki Ohdan, MD, PhD, Department of Surgery,
Division of Frontier Medical Science, Programs for Biomedical Research, GraduateSchool of Biomedical Sciences, Hiroshima University, 1-2-3 Kasumi, Minami-ku,Hiroshima 734-8551, Japan. E-mail: [email protected]; fax: 81 82 2575224.
Copyright © 2004 by the American Association for the Study of Liver Diseases.Published online in Wiley InterScience (www.interscience.wiley.com).DOI 10.1002/hep.20204
1321

tors.1,11,12 Although it is not clear at present how liver NKcells kill self-hepatocytes, it has been speculated that hepa-tocytes do not express sufficient MHC class I to inhibitNK cell-induced killing13 or that liver NK cells include acell population lacking sufficient inhibitory receptors.Another possibility is that liver NK cells include a cellpopulation expressing certain effector molecules that can-not be downregulated even by self-class I recognition viaNK inhibitory receptors. Enhanced cytotoxicity againstinfected or transformed targets is a feature of activatedNK cells, which is believed to be due to the elevatedexpression of tumor necrosis factor (TNF) family mem-bers, including Fas ligand (FasL), and the increased pro-duction of perforin, granzymes, and cytokines by thesecells.14–18 Among the TNF family members, TNF-relatedapoptosis-inducing ligand (TRAIL) has recently beenshown to be critical for NK cell-mediated anti-tumorfunction.19–21 Because TRAIL is constitutively expressedby NK cells in the liver, unlike in other tissues,21 thismolecule might be involved in NK cell-induced hepato-cyte killing.
In the present study, we proved that liver NK cells havecytotoxicity against freshly isolated syngeneic hepatocytesexpressing self-MHC class I molecules and that TRAILon the liver NK cells contribute to their hepatocyte tox-icity. We also have demonstrated that the majority of liverNK cells expressing TRAIL lack Ly-49 inhibitory recep-tors that recognize self-MHC molecules. These findingsare consistent with a model in which TRAIL-expressingliver NK cells, which are characterized as heretics in viewof the self-tolerance system, play a critical role in self-hepatocyte killing through poor recognition of self-MHC.
Materials and Methods
Mice. C57BL/6J (B6) (H-2b) mice were purchasedfrom Clea Japan, Inc. (Osaka, Japan). B6 severe com-bined immunodeficient disease (SCID) (H-2b) mice werepurchased from Jackson Laboratory (Bar Harbor, ME)and were bred at Hiroshima University. All animals weremaintained in a specific pathogen-free microenviron-ment. Mice were used between 8 and 12 weeks of age.
In Vivo Analysis of NK Cell-Mediated CytotoxicityAgainst Autologous Hepatocytes. To activate NK cells,B6 normal wild-type (WT) and SCID mice were treatedwith intraperitoneal (i.p.) injection of polyinosinic-poly-cytidylic acid (poly I:C) (Sigma, St. Louis, MO) (150�g/body). To deplete NK cells, mice were treated withi.p. injection of rabbit anti-asialo GM1 serum (anti-AsGM1) (Wako Chemicals, Osaka, Japan) (50 �L/body)two days before poly I:C injection. In our preliminary
experiment, the proportions of NK cells (NK1.1�
TCR�–) and NKT cells (NK1.1� TCR��) in liver leu-kocytes of the B6 mice treated with anti-AsGM1 were2.3 � 1.1% and 20.1 � 1.1%, respectively (n � 4) (3 dafter treatment), whereas those in liver leukocytes of un-treated control B6 mice were 11.5 � 2.1% and 20.9 �2.8%, respectively (n � 4). Peripheral blood was collectedfrom each mouse 1 day after poly I:C injection. The se-rum activity of alanine aminotransferase (ALT) was as-sayed by standard enzymatic methods.
Isolation of Liver and Spleen Leukocytes. Liver andspleen leukocytes were isolated from B6 WT and SCIDmice that were untreated or treated with i.p. injection ofpoly I:C (150 �g/body) 1 day before the assay. Liverleukocytes were prepared as described previously.22 Inbrief, after pre-perfusion via the portal vein using 1 mL ofphosphate-buffered saline (PBS) supplemented with 10%heparin, the liver was perfused with 50 mL of PBS sup-plemented with 0.1% ethylenediaminetetraacetic acid(EDTA), and the perfusate was collected and subjected toerythrocyte lysis using ammonium chloride/potassiumsolution.
Isolation and Culture of Hepatocytes. Hepatocyteswere prepared as described previously.23,24 Isolated hepa-tocytes were cultured in the hepatocyte growth medium:Dulbecco’s modified Eagle’s medium supplemented with10% fetal bovine serum and antibiotics of 100 IU/mLpenicillin G-100 �g/mL streptomycin (10% DMEM)containing 20 mmol/L N-2-hydroxyethylpiperazine-N�-2-ethane sulfonic acid, 30 �g/mL L-proline, 0.5 �g/mLinsulin, 10–7 mol/L dexamethasone, 44 mmol/LNaHCO3, 10 mmol/L nicotinamide, 10 ng/mL epider-mal growth factor (EGF), and 0.2 mmol/L L-ascorbic acid2-phosphate. In some experiments, the medium was sup-plemented with 300 U/mL IFN-� (Pepro Tech, Inc.,Rocky Hill, NJ). Viability of hepatocytes was determinedas more than 80% by the Trypan blue exclusion test.
Flow Cytometric Analysis. All analyses were per-formed on a FACSCalibur� cytometer (BD Biosciences,Mountain View, CA). For phenotyping NK cell surfacemarkers, liver and spleen leukocytes were stained with thefollowing mAbs: fluorescein isothiocyanate (FITC)-con-jugated anti-NK1.1 (PK136), phycoerythrin (PE)-conju-gated anti-T cell receptor (TCR)-� (H57-597), andbiotin-conjugated anti-Ly-49A (A1), anti-Ly-49C/I(5E6), anti-Ly-49G2 (4D11), anti-B220 (RA3-6B2),anti-CD69 (H1.2F3), anti-FasL (MLF4), or unconju-gated anti-TRAIL mAb (N2B2) (all from BD Pharmin-gen, San Diego, CA). Staining with unconjugated anti-TRAIL mAb was followed by staining with biotin-conjugated anti-rat immunoglobulin (Ig)G2a mAb(RG7/1.30). In some experiments, liver leukocytes were
1322 OCHI ET AL. HEPATOLOGY, May 2004

stained with FITC-conjugated anti-NK1.1, PE-conju-gated anti-TRAIL (N2B2) (eBioscience, San Diego, CA),and biotin-conjugated anti-CD69, anti-Ly-49A, anti-Ly-49C/I, anti-Ly-49G2 mAb, or a mixture of anti-Ly-49A,anti-Ly-49C/I, and anti-Ly-49G2 mAbs. The biotinyl-ated mAb was visualized with allophycocyanin-streptavi-din. For phenotyping hepatocyte MHC molecules,hepatocytes were stained with biotin-conjugated anti-H-2Db (KH95) (BD Pharmingen) and PE-streptavidin.Nonspecific Fc�R binding of labeled Abs was blocked byanti-CD16/32(2.4G2) (BD Pharmingen). Dead cellswere excluded from the analysis by light-scatter and/orpropidium iodide.
NK Cell Cytotoxicity Assay Against Self-Hepato-cytes. Freshly isolated or 7-day cultured hepatocytesfrom B6 WT mice were used as target cells. As effectorcells, liver and spleen leukocytes were obtained from B6WT or SCID mice that were untreated or injected withpoly I:C (150 �g, i.p.) 1 day before assay. In our prelim-inary studies, we confirmed that B6 SCID hepatocytesand congenic B6 WT hepatocytes express MHC class I atthe same levels and are equally susceptible to NK cell-mediated cytotoxicity (data not shown). After the hepa-tocytes (104 cells/well) had been cultured with thoseleukocytes at various ratios in 96-well round-bottom mi-crotiter plates containing 10% DMEM (total volume of200 �L) for 4 hours at 37°C in a 5% CO2 incubator, ALTactivity in each supernatant was assayed by standard en-zymatic methods. Percent cytotoxicity was calculated as(experimental ALT – spontaneous ALT) � 100 / (totalALT – spontaneous ALT). Total ALT activity was deter-mined by lysing the cells with 2.5% Nonidet� P-40 (Na-calai Tesque, Inc., Kyoto, Japan). In some experiments,the assay was performed in the presence of 10 �g/mL ofanti-TRAIL (N2B2) mAb, 10 �g/mL of anti-FasL(MFL3) mAb (all from BD Pharmingen), and/or 50nmol/L of concanamycin A (CMA) (Wako Chemicals,Osaka, Japan), which inhibited perforin-mediated cyto-toxicity.
Isolation of NK Cells. Liver and spleen leukocyteswere obtained from B6 WT mice treated with poly I:C 1day before the assay. NK cells were separated by high-gradient magnetic sorting using autoMACS� (MiltenyiBiotec, Auburn, CA). In brief, leukocytes were magneti-cally labeled by using a cocktail of biotin-conjugated anti-CD4 (RM4-5), CD5 (53-7.3), CD8a (53-6.7), CD19(1D3), Ly-6G (RB6-8C5), and Ter-119 mAbs (TER-119) (all from BD Pharmingen) and streptavidin mi-crobeads (Miltenyi Biotec). Isolation of unlabeled NKcells was achieved by depletion of the labeled cells.TRAIL– NK cells were further magnetically sorted usingunconjugated anti-TRAIL mAb, biotin-conjugated anti-
rat IgG2a mAb, and streptavidin microbeads in the neg-ative fraction. NK cells and TRAIL– NK cells were used aseffector cells in a cytotoxicity assay.
Hepatocyte Transplantation. Hepatocytes isolatedfrom B6 WT mice were labeled with 5-(and -6)-carboxy-fluorescein diacetate succinimidyl ester (CFSE) (Molecu-lar Probes, Eugene, OR) (5 �M), as describedpreviously.25 CFSE-labeled hepatocytes (1 � 106) in 0.5mL of medium 199 (Sigma, St. Louis, MO) were injectedthrough the superior mesenteric vein to B6 WT micetreated with poly I:C on day –1 with or without pretreat-ment of anti-AsGM1 on day –2. Those mice were sacri-ficed 7 days after hepatocyte transplantation. Dissectedlivers were examined by fluorescent microscopy to iden-tify CFSE-labeled hepatocytes.
Statistical Analysis. The results were analyzed byStudent’s t test of means. A P-value less than 0.05 wasconsidered to be statistically significant.
Results
Liver NK Cells Have Cytotoxicity Against NormalSelf-Hepatocytes. To determine whether activation ofNK cells leads to injury of normal self-hepatocytes,poly I:C, which activates NK cells primarily by induc-ing the production of type I (�, �) IFN and IL-12 froma wide variety of cell types,26 was injected into B6 WTmice. Administration of poly I:C resulted in mild butstatistically significant elevation of serum ALT values(Fig. 1). Such elevation of ALT values was abrogated byprior injection of anti-AsGM1 sera in the mice, indi-cating that cells expressing AsGM1 (either NK or NKTcells) are required for induction of hepatocyte injury.Similar results were obtained in B6 SCID mice lacking
Fig. 1. Administration of poly I:C resulted in mild but statisticallysignificant elevation of serum ALT values in mice. Poly I:C (150 �g, i.p.)was injected into B6 wild-type (WT) and SCID mice. To deplete NK cells,anti-mouse asialo GM1 serum (anti-AsGM1; 50 �L, i.p.) was injected 2days before poly I:C injection. The serum activity of ALT was determined1 day after injection of poly I:C. The results shown are the average �SEM of values from 6 or 7 mice in each group. *P � .05; #P � .01.Abbreviations; poly I:C, polyinosinic-polycytidylic acid; ALT, alanine ami-notransferase; i.p., intraperitoneally; WT, wild-type; SCID, severe com-bined immunodeficient disease; NK, natural killer.
HEPATOLOGY, Vol. 39, No. 5, 2004 OCHI ET AL. 1323
NKT cells, as well as T and B cells, but possessing NKcells, demonstrating that NK cells were the principaleffectors causing self-hepatocyte injury induced bypoly I:C.
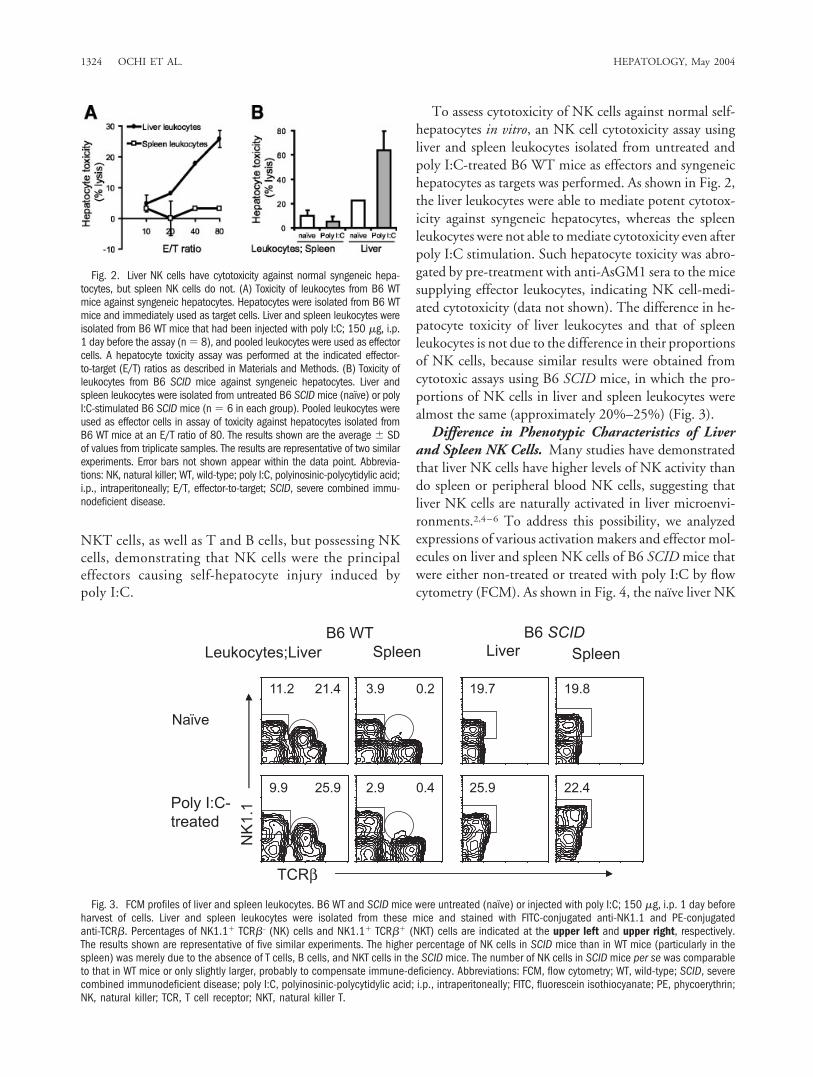
To assess cytotoxicity of NK cells against normal self-hepatocytes in vitro, an NK cell cytotoxicity assay usingliver and spleen leukocytes isolated from untreated andpoly I:C-treated B6 WT mice as effectors and syngeneichepatocytes as targets was performed. As shown in Fig. 2,the liver leukocytes were able to mediate potent cytotox-icity against syngeneic hepatocytes, whereas the spleenleukocytes were not able to mediate cytotoxicity even afterpoly I:C stimulation. Such hepatocyte toxicity was abro-gated by pre-treatment with anti-AsGM1 sera to the micesupplying effector leukocytes, indicating NK cell-medi-ated cytotoxicity (data not shown). The difference in he-patocyte toxicity of liver leukocytes and that of spleenleukocytes is not due to the difference in their proportionsof NK cells, because similar results were obtained fromcytotoxic assays using B6 SCID mice, in which the pro-portions of NK cells in liver and spleen leukocytes werealmost the same (approximately 20%–25%) (Fig. 3).
Difference in Phenotypic Characteristics of Liverand Spleen NK Cells. Many studies have demonstratedthat liver NK cells have higher levels of NK activity thando spleen or peripheral blood NK cells, suggesting thatliver NK cells are naturally activated in liver microenvi-ronments.2,4–6 To address this possibility, we analyzedexpressions of various activation makers and effector mol-ecules on liver and spleen NK cells of B6 SCID mice thatwere either non-treated or treated with poly I:C by flowcytometry (FCM). As shown in Fig. 4, the naı̈ve liver NK
Fig. 2. Liver NK cells have cytotoxicity against normal syngeneic hepa-tocytes, but spleen NK cells do not. (A) Toxicity of leukocytes from B6 WTmice against syngeneic hepatocytes. Hepatocytes were isolated from B6 WTmice and immediately used as target cells. Liver and spleen leukocytes wereisolated from B6 WT mice that had been injected with poly I:C; 150 �g, i.p.1 day before the assay (n � 8), and pooled leukocytes were used as effectorcells. A hepatocyte toxicity assay was performed at the indicated effector-to-target (E/T) ratios as described in Materials and Methods. (B) Toxicity ofleukocytes from B6 SCID mice against syngeneic hepatocytes. Liver andspleen leukocytes were isolated from untreated B6 SCID mice (naı̈ve) or polyI:C-stimulated B6 SCID mice (n � 6 in each group). Pooled leukocytes wereused as effector cells in assay of toxicity against hepatocytes isolated fromB6 WT mice at an E/T ratio of 80. The results shown are the average � SDof values from triplicate samples. The results are representative of two similarexperiments. Error bars not shown appear within the data point. Abbrevia-tions: NK, natural killer; WT, wild-type; poly I:C, polyinosinic-polycytidylic acid;i.p., intraperitoneally; E/T, effector-to-target; SCID, severe combined immu-nodeficient disease.
Fig. 3. FCM profiles of liver and spleen leukocytes. B6 WT and SCID mice were untreated (naı̈ve) or injected with poly I:C; 150 �g, i.p. 1 day beforeharvest of cells. Liver and spleen leukocytes were isolated from these mice and stained with FITC-conjugated anti-NK1.1 and PE-conjugatedanti-TCR�. Percentages of NK1.1� TCR�– (NK) cells and NK1.1� TCR�� (NKT) cells are indicated at the upper left and upper right, respectively.The results shown are representative of five similar experiments. The higher percentage of NK cells in SCID mice than in WT mice (particularly in thespleen) was merely due to the absence of T cells, B cells, and NKT cells in the SCID mice. The number of NK cells in SCID mice per se was comparableto that in WT mice or only slightly larger, probably to compensate immune-deficiency. Abbreviations: FCM, flow cytometry; WT, wild-type; SCID, severecombined immunodeficient disease; poly I:C, polyinosinic-polycytidylic acid; i.p., intraperitoneally; FITC, fluorescein isothiocyanate; PE, phycoerythrin;NK, natural killer; TCR, T cell receptor; NKT, natural killer T.
1324 OCHI ET AL. HEPATOLOGY, May 2004
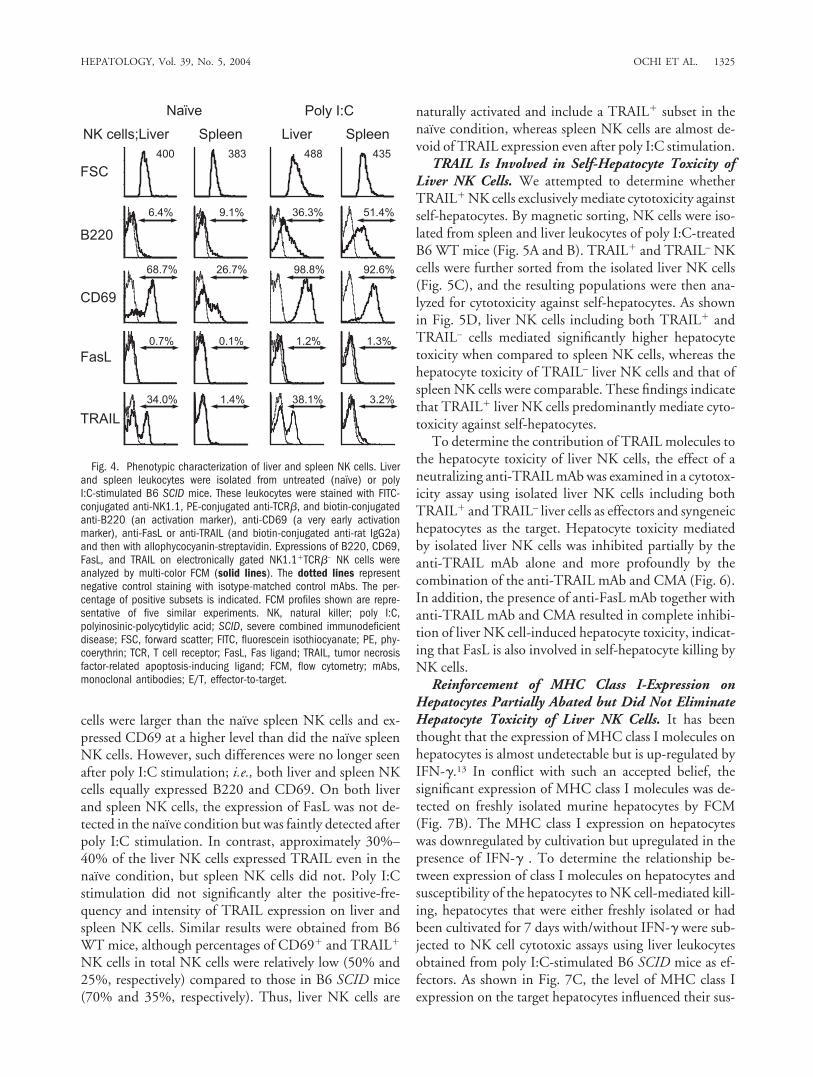
cells were larger than the naı̈ve spleen NK cells and ex-pressed CD69 at a higher level than did the naı̈ve spleenNK cells. However, such differences were no longer seenafter poly I:C stimulation; i.e., both liver and spleen NKcells equally expressed B220 and CD69. On both liverand spleen NK cells, the expression of FasL was not de-tected in the naı̈ve condition but was faintly detected afterpoly I:C stimulation. In contrast, approximately 30%–40% of the liver NK cells expressed TRAIL even in thenaı̈ve condition, but spleen NK cells did not. Poly I:Cstimulation did not significantly alter the positive-fre-quency and intensity of TRAIL expression on liver andspleen NK cells. Similar results were obtained from B6WT mice, although percentages of CD69� and TRAIL�
NK cells in total NK cells were relatively low (50% and25%, respectively) compared to those in B6 SCID mice(70% and 35%, respectively). Thus, liver NK cells are
naturally activated and include a TRAIL� subset in thenaı̈ve condition, whereas spleen NK cells are almost de-void of TRAIL expression even after poly I:C stimulation.
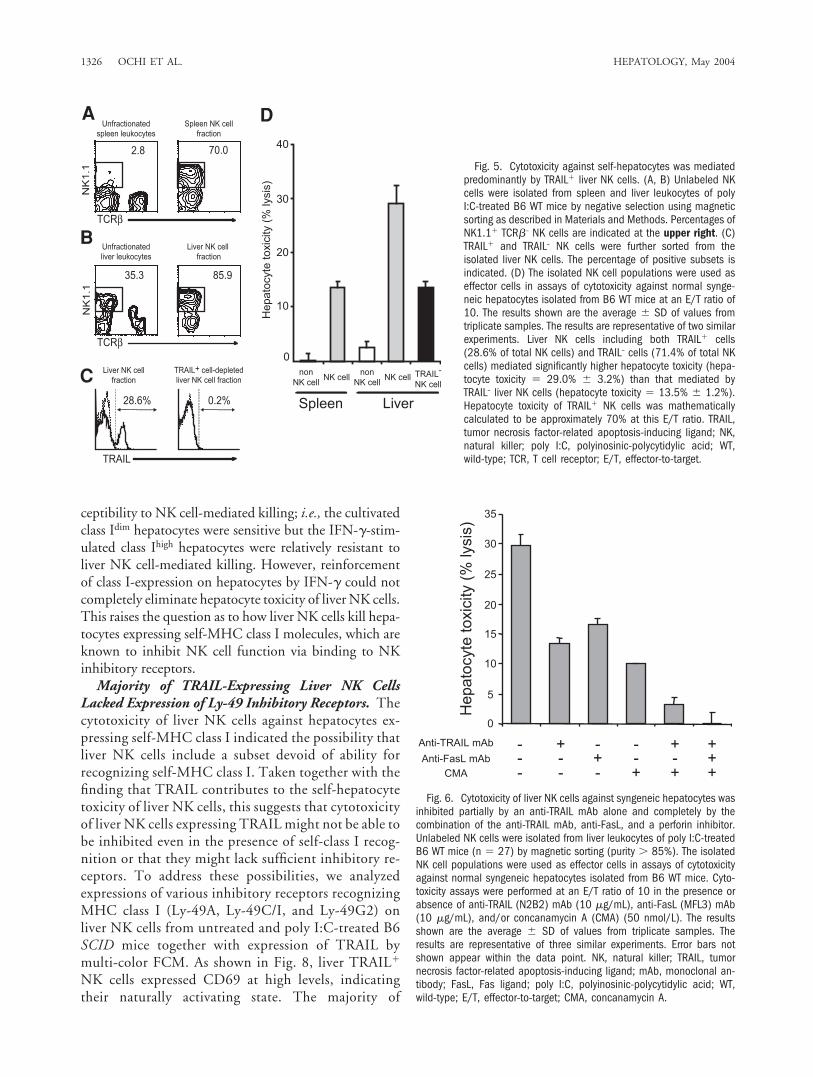
TRAIL Is Involved in Self-Hepatocyte Toxicity ofLiver NK Cells. We attempted to determine whetherTRAIL� NK cells exclusively mediate cytotoxicity againstself-hepatocytes. By magnetic sorting, NK cells were iso-lated from spleen and liver leukocytes of poly I:C-treatedB6 WT mice (Fig. 5A and B). TRAIL� and TRAIL– NKcells were further sorted from the isolated liver NK cells(Fig. 5C), and the resulting populations were then ana-lyzed for cytotoxicity against self-hepatocytes. As shownin Fig. 5D, liver NK cells including both TRAIL� andTRAIL– cells mediated significantly higher hepatocytetoxicity when compared to spleen NK cells, whereas thehepatocyte toxicity of TRAIL– liver NK cells and that ofspleen NK cells were comparable. These findings indicatethat TRAIL� liver NK cells predominantly mediate cyto-toxicity against self-hepatocytes.
To determine the contribution of TRAIL molecules tothe hepatocyte toxicity of liver NK cells, the effect of aneutralizing anti-TRAIL mAb was examined in a cytotox-icity assay using isolated liver NK cells including bothTRAIL� and TRAIL– liver cells as effectors and syngeneichepatocytes as the target. Hepatocyte toxicity mediatedby isolated liver NK cells was inhibited partially by theanti-TRAIL mAb alone and more profoundly by thecombination of the anti-TRAIL mAb and CMA (Fig. 6).In addition, the presence of anti-FasL mAb together withanti-TRAIL mAb and CMA resulted in complete inhibi-tion of liver NK cell-induced hepatocyte toxicity, indicat-ing that FasL is also involved in self-hepatocyte killing byNK cells.
Reinforcement of MHC Class I-Expression onHepatocytes Partially Abated but Did Not EliminateHepatocyte Toxicity of Liver NK Cells. It has beenthought that the expression of MHC class I molecules onhepatocytes is almost undetectable but is up-regulated byIFN-�.13 In conflict with such an accepted belief, thesignificant expression of MHC class I molecules was de-tected on freshly isolated murine hepatocytes by FCM(Fig. 7B). The MHC class I expression on hepatocyteswas downregulated by cultivation but upregulated in thepresence of IFN-� . To determine the relationship be-tween expression of class I molecules on hepatocytes andsusceptibility of the hepatocytes to NK cell-mediated kill-ing, hepatocytes that were either freshly isolated or hadbeen cultivated for 7 days with/without IFN-� were sub-jected to NK cell cytotoxic assays using liver leukocytesobtained from poly I:C-stimulated B6 SCID mice as ef-fectors. As shown in Fig. 7C, the level of MHC class Iexpression on the target hepatocytes influenced their sus-
Fig. 4. Phenotypic characterization of liver and spleen NK cells. Liverand spleen leukocytes were isolated from untreated (naı̈ve) or polyI:C-stimulated B6 SCID mice. These leukocytes were stained with FITC-conjugated anti-NK1.1, PE-conjugated anti-TCR�, and biotin-conjugatedanti-B220 (an activation marker), anti-CD69 (a very early activationmarker), anti-FasL or anti-TRAIL (and biotin-conjugated anti-rat IgG2a)and then with allophycocyanin-streptavidin. Expressions of B220, CD69,FasL, and TRAIL on electronically gated NK1.1�TCR�– NK cells wereanalyzed by multi-color FCM (solid lines). The dotted lines representnegative control staining with isotype-matched control mAbs. The per-centage of positive subsets is indicated. FCM profiles shown are repre-sentative of five similar experiments. NK, natural killer; poly I:C,polyinosinic-polycytidylic acid; SCID, severe combined immunodeficientdisease; FSC, forward scatter; FITC, fluorescein isothiocyanate; PE, phy-coerythrin; TCR, T cell receptor; FasL, Fas ligand; TRAIL, tumor necrosisfactor-related apoptosis-inducing ligand; FCM, flow cytometry; mAbs,monoclonal antibodies; E/T, effector-to-target.
HEPATOLOGY, Vol. 39, No. 5, 2004 OCHI ET AL. 1325
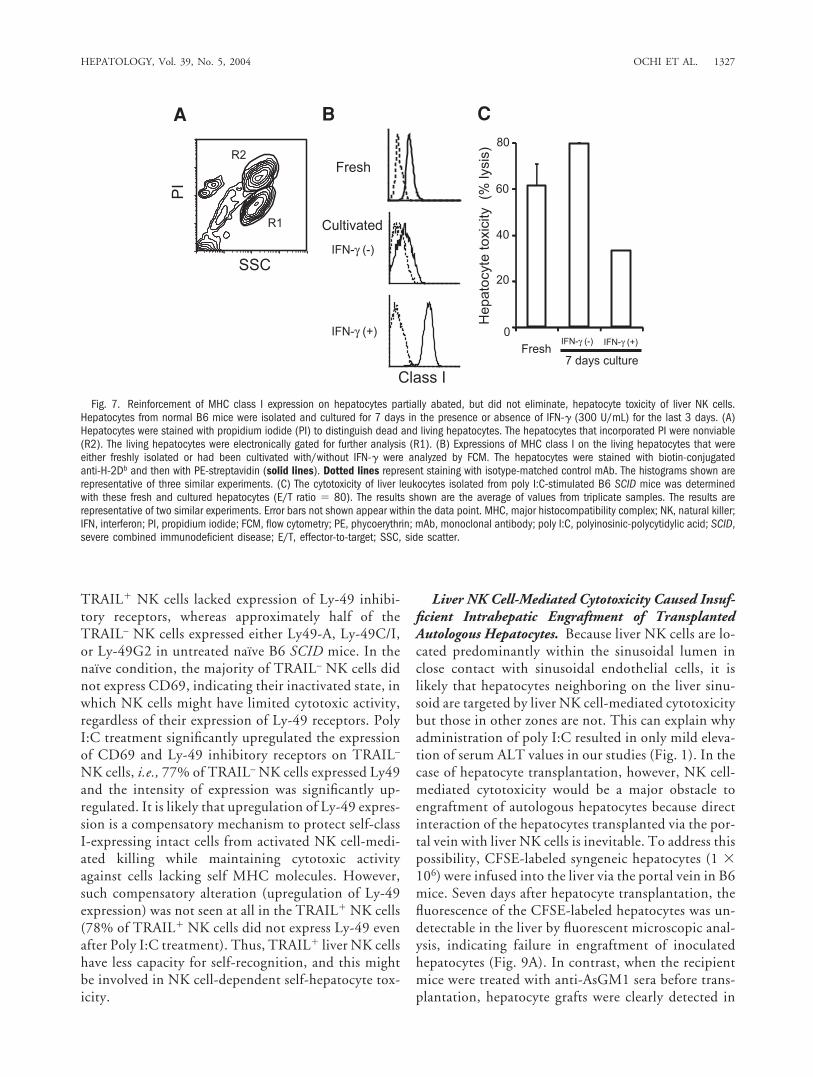
ceptibility to NK cell-mediated killing; i.e., the cultivatedclass Idim hepatocytes were sensitive but the IFN-�-stim-ulated class Ihigh hepatocytes were relatively resistant toliver NK cell-mediated killing. However, reinforcementof class I-expression on hepatocytes by IFN-� could notcompletely eliminate hepatocyte toxicity of liver NK cells.This raises the question as to how liver NK cells kill hepa-tocytes expressing self-MHC class I molecules, which areknown to inhibit NK cell function via binding to NKinhibitory receptors.
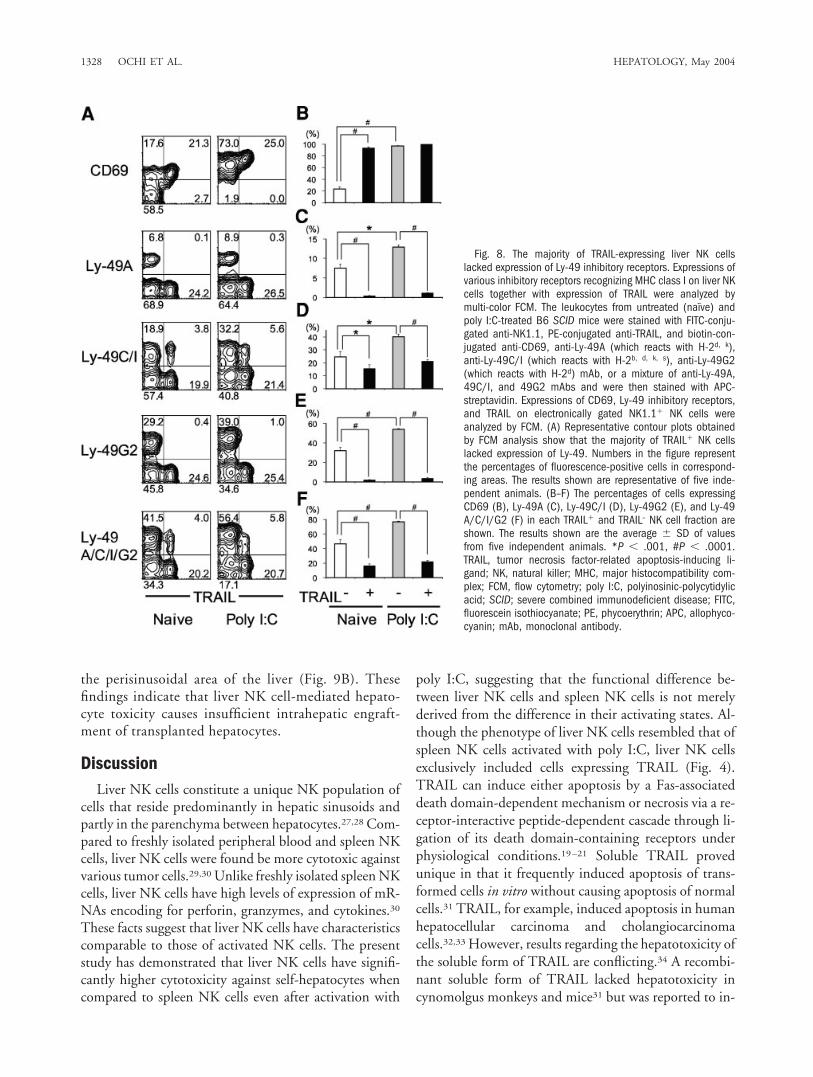
Majority of TRAIL-Expressing Liver NK CellsLacked Expression of Ly-49 Inhibitory Receptors. Thecytotoxicity of liver NK cells against hepatocytes ex-pressing self-MHC class I indicated the possibility thatliver NK cells include a subset devoid of ability forrecognizing self-MHC class I. Taken together with thefinding that TRAIL contributes to the self-hepatocytetoxicity of liver NK cells, this suggests that cytotoxicityof liver NK cells expressing TRAIL might not be able tobe inhibited even in the presence of self-class I recog-nition or that they might lack sufficient inhibitory re-ceptors. To address these possibilities, we analyzedexpressions of various inhibitory receptors recognizingMHC class I (Ly-49A, Ly-49C/I, and Ly-49G2) onliver NK cells from untreated and poly I:C-treated B6SCID mice together with expression of TRAIL bymulti-color FCM. As shown in Fig. 8, liver TRAIL�
NK cells expressed CD69 at high levels, indicatingtheir naturally activating state. The majority of
Fig. 5. Cytotoxicity against self-hepatocytes was mediatedpredominantly by TRAIL� liver NK cells. (A, B) Unlabeled NKcells were isolated from spleen and liver leukocytes of polyI:C-treated B6 WT mice by negative selection using magneticsorting as described in Materials and Methods. Percentages ofNK1.1� TCR�– NK cells are indicated at the upper right. (C)TRAIL� and TRAIL– NK cells were further sorted from theisolated liver NK cells. The percentage of positive subsets isindicated. (D) The isolated NK cell populations were used aseffector cells in assays of cytotoxicity against normal synge-neic hepatocytes isolated from B6 WT mice at an E/T ratio of10. The results shown are the average � SD of values fromtriplicate samples. The results are representative of two similarexperiments. Liver NK cells including both TRAIL� cells(28.6% of total NK cells) and TRAIL– cells (71.4% of total NKcells) mediated significantly higher hepatocyte toxicity (hepa-tocyte toxicity � 29.0% � 3.2%) than that mediated byTRAIL– liver NK cells (hepatocyte toxicity � 13.5% � 1.2%).Hepatocyte toxicity of TRAIL� NK cells was mathematicallycalculated to be approximately 70% at this E/T ratio. TRAIL,tumor necrosis factor-related apoptosis-inducing ligand; NK,natural killer; poly I:C, polyinosinic-polycytidylic acid; WT,wild-type; TCR, T cell receptor; E/T, effector-to-target.
Fig. 6. Cytotoxicity of liver NK cells against syngeneic hepatocytes wasinhibited partially by an anti-TRAIL mAb alone and completely by thecombination of the anti-TRAIL mAb, anti-FasL, and a perforin inhibitor.Unlabeled NK cells were isolated from liver leukocytes of poly I:C-treatedB6 WT mice (n � 27) by magnetic sorting (purity � 85%). The isolatedNK cell populations were used as effector cells in assays of cytotoxicityagainst normal syngeneic hepatocytes isolated from B6 WT mice. Cyto-toxicity assays were performed at an E/T ratio of 10 in the presence orabsence of anti-TRAIL (N2B2) mAb (10 �g/mL), anti-FasL (MFL3) mAb(10 �g/mL), and/or concanamycin A (CMA) (50 nmol/L). The resultsshown are the average � SD of values from triplicate samples. Theresults are representative of three similar experiments. Error bars notshown appear within the data point. NK, natural killer; TRAIL, tumornecrosis factor-related apoptosis-inducing ligand; mAb, monoclonal an-tibody; FasL, Fas ligand; poly I:C, polyinosinic-polycytidylic acid; WT,wild-type; E/T, effector-to-target; CMA, concanamycin A.
1326 OCHI ET AL. HEPATOLOGY, May 2004
TRAIL� NK cells lacked expression of Ly-49 inhibi-tory receptors, whereas approximately half of theTRAIL– NK cells expressed either Ly49-A, Ly-49C/I,or Ly-49G2 in untreated naı̈ve B6 SCID mice. In thenaı̈ve condition, the majority of TRAIL– NK cells didnot express CD69, indicating their inactivated state, inwhich NK cells might have limited cytotoxic activity,regardless of their expression of Ly-49 receptors. PolyI:C treatment significantly upregulated the expressionof CD69 and Ly-49 inhibitory receptors on TRAIL–
NK cells, i.e., 77% of TRAIL– NK cells expressed Ly49and the intensity of expression was significantly up-regulated. It is likely that upregulation of Ly-49 expres-sion is a compensatory mechanism to protect self-classI-expressing intact cells from activated NK cell-medi-ated killing while maintaining cytotoxic activityagainst cells lacking self MHC molecules. However,such compensatory alteration (upregulation of Ly-49expression) was not seen at all in the TRAIL� NK cells(78% of TRAIL� NK cells did not express Ly-49 evenafter Poly I:C treatment). Thus, TRAIL� liver NK cellshave less capacity for self-recognition, and this mightbe involved in NK cell-dependent self-hepatocyte tox-icity.
Liver NK Cell-Mediated Cytotoxicity Caused Insuf-ficient Intrahepatic Engraftment of TransplantedAutologous Hepatocytes. Because liver NK cells are lo-cated predominantly within the sinusoidal lumen inclose contact with sinusoidal endothelial cells, it islikely that hepatocytes neighboring on the liver sinu-soid are targeted by liver NK cell-mediated cytotoxicitybut those in other zones are not. This can explain whyadministration of poly I:C resulted in only mild eleva-tion of serum ALT values in our studies (Fig. 1). In thecase of hepatocyte transplantation, however, NK cell-mediated cytotoxicity would be a major obstacle toengraftment of autologous hepatocytes because directinteraction of the hepatocytes transplanted via the por-tal vein with liver NK cells is inevitable. To address thispossibility, CFSE-labeled syngeneic hepatocytes (1 �106) were infused into the liver via the portal vein in B6mice. Seven days after hepatocyte transplantation, thefluorescence of the CFSE-labeled hepatocytes was un-detectable in the liver by fluorescent microscopic anal-ysis, indicating failure in engraftment of inoculatedhepatocytes (Fig. 9A). In contrast, when the recipientmice were treated with anti-AsGM1 sera before trans-plantation, hepatocyte grafts were clearly detected in
Fig. 7. Reinforcement of MHC class I expression on hepatocytes partially abated, but did not eliminate, hepatocyte toxicity of liver NK cells.Hepatocytes from normal B6 mice were isolated and cultured for 7 days in the presence or absence of IFN-� (300 U/mL) for the last 3 days. (A)Hepatocytes were stained with propidium iodide (PI) to distinguish dead and living hepatocytes. The hepatocytes that incorporated PI were nonviable(R2). The living hepatocytes were electronically gated for further analysis (R1). (B) Expressions of MHC class I on the living hepatocytes that wereeither freshly isolated or had been cultivated with/without IFN-� were analyzed by FCM. The hepatocytes were stained with biotin-conjugatedanti-H-2Db and then with PE-streptavidin (solid lines). Dotted lines represent staining with isotype-matched control mAb. The histograms shown arerepresentative of three similar experiments. (C) The cytotoxicity of liver leukocytes isolated from poly I:C-stimulated B6 SCID mice was determinedwith these fresh and cultured hepatocytes (E/T ratio � 80). The results shown are the average of values from triplicate samples. The results arerepresentative of two similar experiments. Error bars not shown appear within the data point. MHC, major histocompatibility complex; NK, natural killer;IFN, interferon; PI, propidium iodide; FCM, flow cytometry; PE, phycoerythrin; mAb, monoclonal antibody; poly I:C, polyinosinic-polycytidylic acid; SCID,severe combined immunodeficient disease; E/T, effector-to-target; SSC, side scatter.
HEPATOLOGY, Vol. 39, No. 5, 2004 OCHI ET AL. 1327
the perisinusoidal area of the liver (Fig. 9B). Thesefindings indicate that liver NK cell-mediated hepato-cyte toxicity causes insufficient intrahepatic engraft-ment of transplanted hepatocytes.
DiscussionLiver NK cells constitute a unique NK population of
cells that reside predominantly in hepatic sinusoids andpartly in the parenchyma between hepatocytes.27,28 Com-pared to freshly isolated peripheral blood and spleen NKcells, liver NK cells were found be more cytotoxic againstvarious tumor cells.29,30 Unlike freshly isolated spleen NKcells, liver NK cells have high levels of expression of mR-NAs encoding for perforin, granzymes, and cytokines.30
These facts suggest that liver NK cells have characteristicscomparable to those of activated NK cells. The presentstudy has demonstrated that liver NK cells have signifi-cantly higher cytotoxicity against self-hepatocytes whencompared to spleen NK cells even after activation with
poly I:C, suggesting that the functional difference be-tween liver NK cells and spleen NK cells is not merelyderived from the difference in their activating states. Al-though the phenotype of liver NK cells resembled that ofspleen NK cells activated with poly I:C, liver NK cellsexclusively included cells expressing TRAIL (Fig. 4).TRAIL can induce either apoptosis by a Fas-associateddeath domain-dependent mechanism or necrosis via a re-ceptor-interactive peptide-dependent cascade through li-gation of its death domain-containing receptors underphysiological conditions.19–21 Soluble TRAIL provedunique in that it frequently induced apoptosis of trans-formed cells in vitro without causing apoptosis of normalcells.31 TRAIL, for example, induced apoptosis in humanhepatocellular carcinoma and cholangiocarcinomacells.32,33 However, results regarding the hepatotoxicity ofthe soluble form of TRAIL are conflicting.34 A recombi-nant soluble form of TRAIL lacked hepatotoxicity incynomolgus monkeys and mice31 but was reported to in-
Fig. 8. The majority of TRAIL-expressing liver NK cellslacked expression of Ly-49 inhibitory receptors. Expressions ofvarious inhibitory receptors recognizing MHC class I on liver NKcells together with expression of TRAIL were analyzed bymulti-color FCM. The leukocytes from untreated (naı̈ve) andpoly I:C-treated B6 SCID mice were stained with FITC-conju-gated anti-NK1.1, PE-conjugated anti-TRAIL, and biotin-con-jugated anti-CD69, anti-Ly-49A (which reacts with H-2d, k),anti-Ly-49C/I (which reacts with H-2b, d, k, s), anti-Ly-49G2(which reacts with H-2d) mAb, or a mixture of anti-Ly-49A,49C/I, and 49G2 mAbs and were then stained with APC-streptavidin. Expressions of CD69, Ly-49 inhibitory receptors,and TRAIL on electronically gated NK1.1� NK cells wereanalyzed by FCM. (A) Representative contour plots obtainedby FCM analysis show that the majority of TRAIL� NK cellslacked expression of Ly-49. Numbers in the figure representthe percentages of fluorescence-positive cells in correspond-ing areas. The results shown are representative of five inde-pendent animals. (B–F) The percentages of cells expressingCD69 (B), Ly-49A (C), Ly-49C/I (D), Ly-49G2 (E), and Ly-49A/C/I/G2 (F) in each TRAIL� and TRAIL– NK cell fraction areshown. The results shown are the average � SD of valuesfrom five independent animals. *P � .001, #P � .0001.TRAIL, tumor necrosis factor-related apoptosis-inducing li-gand; NK, natural killer; MHC, major histocompatibility com-plex; FCM, flow cytometry; poly I:C, polyinosinic-polycytidylicacid; SCID; severe combined immunodeficient disease; FITC,fluorescein isothiocyanate; PE, phycoerythrin; APC, allophyco-cyanin; mAb, monoclonal antibody.
1328 OCHI ET AL. HEPATOLOGY, May 2004

duce apoptosis in cultured human hepatocytes.35 Theseobservations raise the possibility that human hepatocytesare uniquely sensitive to TRAIL compared to nonhumanprimate and rodent hepatocytes. There are also conflict-ing results regarding the hepatotoxicity of the membrane-bound form of TRAIL delivered by gene-transfectionusing an adenoviral vector.36 Despite a number of studiesdealing with exogenous TRAIL of either the soluble formor membrane-bound form expressed by genomic manip-ulation, hepatotoxicity of TRAIL endogenously expressedon NK cells has not yet been investigated. In the presentstudy, hepatocyte toxicity of liver NK cells was inhibitedpartially by anti-TRAIL mAb alone and further pro-foundly by the combination of anti-TRAIL mAb andCMA, indicating that TRAIL contributes to NK cell-mediated hepatocyte toxicity.
In addition to NK cell-mediated hepatocyte toxicity,NKT cells also cause hepatocyte injury in certain immu-nopathological conditions. A recent study has indicatedthat NKT cells activated by �-galactosylceramide (�-Gal-Cer) induce hepatocyte damage through the Fas-FasLpathway.10 Different from hepatocyte damage induced by�-GalCer, NK cells were predominant effectors of hepa-tocyte damage induced by poly I:C kit given that polyI:C-treatment triggered hepatocyte injury in SCID micelacking NKT cells. In the present study, poly I:C treat-ment did not increase the expression level of Fasl on NKTcells (data not shown) but slightly increased the expres-
sion of Fasl level of FasL on both spleen and liver NK cells(Fig. 4). It is likely that the Fas-FasL pathway plays a rolein the hepatocyte toxicity mediated by NK cells, includ-ing TRAIL– NK cells. In support of this possibility, thepresence of anti-FasL mAb in addition to anti-TRAILmAb and CMA resulted in complete inhibition of liverNK cell-induced hepatocyte toxicity (Fig. 6). Thus, al-though TRAIL� NK cells are predominant cells that havethe ability to kill self-hepatocytes, it is likely that TRAILmolecules per se partially contribute to their killing. Eitherperforin and FasL are also involved in TRAIL� NK cell-mediated hepatocyte toxicity or TRAIL– NK cells par-tially contribute to hepatocyte toxicity via perforin andFasL. To distinguish these possibilities, additional studiesare needed.
It is well known that the presence of MHC class Imolecules protects normal self-cells from NK cell-medi-ated cytotoxicity via binding to NK inhibitory recep-tors.11,12 A “missing-self” hypothesis has been proposed toexplain the protective effect of target cell MHC class I onNK cell-mediated lysis; when tissues lack expression ofMHC class I or express abnormal forms of it, such asduring tumorigenesis or infection, this inhibitory influ-ence is released, permitting target lysis mediated by NKcells. Inconsistent with the missing-self hypothesis, wedemonstrated in the present study that normal hepato-cytes suffered from liver NK cell-induced killing evenwhen the hepatocytes expressed normal MHC class I. Weassumed that liver NK cells include a cell population lack-ing sufficient inhibitory receptors that can recognize self-MHC class I molecules, and we analyzed the expressionsof various inhibitory receptors recognizing class I on liverNK cells. Ly-49A binds the class I molecules H-2Dd, Dk,and Dp, and Ly-49G2 binds H-2Dd and H-2Ld.37–41
These bindings mediate a negative signal to the NK cells,resulting in inhibition of lysis. Recent studies have sug-gested that these members of the Ly-49 family may alsointeract with H-2b molecules.42–45 TRAIL� liver NK cellsalmost completely lacked expression of Ly-49A and Ly-49G2 inhibitory receptors, whereas approximately 10%and 30% of TRAIL– liver NK cells expressed Ly-49A andLy-49G2, respectively. In addition, TRAIL� liver NKcells less frequently express Ly-49 C/I, which binds H-2b
molecules, compared to TRAIL– NK cells. These findingsindicate TRAIL� liver NK cells have less capacity forself-recognition, and this might be involved in NK cell-dependent self-hepatocyte toxicity. Notably, poly I:Ctreatment significantly upregulated expression of Ly-49inhibitory receptors on TRAIL– NK cells but did not alterLy-49 expression on TRAIL� NK cells. It is likely thatnormal hepatocytes may be protected from killing medi-ated by activated TRAIL– NK cells via binding to upregu-
Fig. 9. Fluorescent photomicrographs of transplanted autologoushepatocytes. CFSE-labeled syngeneic hepatocytes (1 � 106) were in-fused into the liver via portal vein in B6 mice. The recipient mice werestimulated with poly I:C; 150 �g, i.p. 1 day before hepatocyte trans-plantation. (A) The fluorescence of the CFSE-labeled hepatocytes wasundetectable in the liver 7 days after hepatocyte transplantation. (B)Transplanted hepatocytes were clearly detected in the liver 7 days aftertransplantation when the recipient mice were treated with anti-AsGM1sera 2 days before hepatocyte transplantation. The results shown arerepresentative of three experimental animals in each group. CFSE, 5-(and-6)-carboxyfluorescein diacetate succinimidyl ester; poly I:C, polyi-nosinic-polycytidylic acid; i.p. intraperitoneally.
HEPATOLOGY, Vol. 39, No. 5, 2004 OCHI ET AL. 1329

lated Ly-49 inhibitory receptors but are susceptible tokilling mediated by activated TRAIL� NK cells becauseof the insufficiency of MHC class I recognition.
Such mechanisms for NK cell-mediated toxicityagainst autologous hepatocytes need to be taken into ac-count in a case of autologous hepatocyte transplantation,because liver NK cells caused a major problem obstruct-ing its success, i.e., insufficient intrahepatic engraftmentof transplanted hepatocytes. Hepatocyte transplantationhas been proposed as an alternative to whole-organ trans-plantation to support many forms of hepatic insuffi-ciency. Previous studies have demonstrated therapeuticefficacy of hepatocyte transplantation through a variety ofliver disease models.46,47 Particularly for the treatment ofinherited metabolic disease, a technique for transplanta-tion of genetically modified autologous hepatocytes,which have the advantage of no T cell-mediated rejection,is being developed. Although the host liver represents anideal site for transplanted hepatocytes in terms of theunique hepatic organization and interactions with non-parenchymal liver cells,48 insufficient intrahepatic en-graftment of transplanted hepatocytes, even those that areautologous, remains a major obstacle. It has been demon-strated that transplanted hepatocytes become stacked atthe portal vein radices, resulting in deposition of some ofthe cells at the hepatic sinusoid.49 Although the majorityof hepatocytes are cleared from these areas, a portion ofthe cells start to translocate into the space of Disse bydisrupting the sinusoidal endothelium and finally joinadjacent host hepatocytes. Because liver NK cells are lo-cated predominantly within the sinusoidal lumen, directinteraction of the transplanted hepatocytes with liver NKcells is inevitable. Therefore, to establish intrahepaticplacement of hepatocyte grafts, inhibition of liver NKcell-mediated hepatocyte toxicity might be necessary.
Acknowledgment: The authors thank Drs. T. Itamotoand H. Tashiro for their advice and encouragement.
References1. Yokoyama WM. Natural killer cells. In: Paul EW, ed. Fundamental Im-
munology. 4th ed. Philadelphia, Pa: Lippincott-Raven; 1999:575–603.2. Seki S, Habu Y, Kawamura T, Takeda K, Dobashi H, Ohkawa T, Hiraide
H. The liver as a crucial organ in the first line of host defense: the roles ofKupffer cells, natural killer (NK) cells and NK1.1 Ag� T cells in T helper1 immune responses. Immunol Rev 2000;174:35–46.
3. Goossens PL, Jouin H, Marchal G, Milon G. Isolation and flow cytometricanalysis of the free lymphomyeloid cells present in murine liver. J ImmunolMethods 1990;132:137–144.
4. Takahashi M, Ogasawara K, Takeda K, Hashimoto W, Sakihara H,Kumagai K, et al. LPS induces NK1.1� alpha beta T cells with potentcytotoxicity in the liver of mice via production of IL-12 from Kupffer cells.J Immunol 1996;156:2436–2442.
5. Crispe IN, Mehal WZ. Strange brew: T cells in the liver. Immunol Today1996;17:522–525.
6. Minagawa M, Oya H, Yamamoto S, Shimizu T, Bannai M, Kawamura H,et al. Intensive expansion of natural killer T cells in the early phase ofhepatocyte regeneration after partial hepatectomy in mice and its associa-tion with sympathetic nerve activation. HEPATOLOGY 2000;31:907–915.
7. Takeda K, Seki S, Ogasawara K, Anzai R, Hashimoto W, Sugiura K, et al.Liver NK1.1� CD4� alpha beta T cells activated by IL-12 as a majoreffector in inhibition of experimental tumor metastasis. J Immunol 1996;156:3366–3373.
8. Liu ZX, Govindarajan S, Okamoto S, Dennert G. NK cells cause liverinjury and facilitate the induction of T cell-mediated immunity to a viralliver infection. J Immunol 2000;164:6480–6486.
9. Nishikage T, Seki S, Toyabe S, Abo T, Kagata Y, Iwai T, et al. Inhibitionof concanavalin A-induced hepatic injury of mice by bacterial lipopolysac-charide via the induction of IL-6 and the subsequent reduction of IL-4: thecytokine milieu of concanavalin A hepatitis. J Hepatol 1999;31:18–26.
10. Nakagawa R, Nagafune I, Tazunoki Y, Ehara H, Tomura H, Iijima R, et al.Mechanisms of the antimetastatic effect in the liver and of the hepatocyteinjury induced by alpha-galactosylceramide in mice. J Immunol 2001;166:6578–6584.
11. Lanier LL. NK cell receptors. Annu Rev Immunol 1998;16:359–393.12. Yokoyama WM. Natural killer cell receptors. Curr Opin Immunol 1998;
10:298–305.13. Skoskiewicz MJ, Colvin RB, Schneeberger EE, Russell PS. Widespread
and selective induction of major histocompatibility complex-determinedantigens in vivo by gamma interferon. J Exp Med 1985;162:1645–1664.
14. Kagi D, Seiler P, Pavlovic J, Ledermann B, Burki K, Zinkernagel RM, et al.The roles of perforin- and Fas-dependent cytotoxicity in protection againstcytopathic and noncytopathic viruses. Eur J Immunol 1995;25:3256–3262.
15. Schneider P, Holler N, Bodmer JL, Hahne M, Frei K, Fontana A, et al.Conversion of membrane-bound Fas(CD95) ligand to its soluble form isassociated with downregulation of its proapoptotic activity and loss of livertoxicity. J Exp Med 1998;187:1205–1213.
16. Kashii Y, Giorda R, Herberman RB, Whiteside TL, Vujanovic NL. Con-stitutive expression and role of the TNF family ligands in apoptotic killingof tumor cells by human NK cells. J Immunol 1999;163:5358–5366.
17. Smyth MJ, Thia KY, Street SE, MacGregor D, Godfrey DI, Trapani JA.Perforin-mediated cytotoxicity is critical for surveillance of spontaneouslymphoma. J Exp Med 2000;192:755–760.
18. Street SE, Cretney E, Smyth MJ. Perforin and interferon-gamma activitiesindependently control tumor initiation, growth, and metastasis. Blood2001;97:192–197.
19. Zamai L, Ahmad M, Bennett IM, Azzoni L, Alnemri ES, Perussia B.Natural killer (NK) cell-mediated cytotoxicity: differential use of TRAILand Fas ligand by immature and mature primary human NK cells. J ExpMed 1998;188:2375–2380.
20. Kayagaki N, Yamaguchi N, Nakayama M, Takeda K, Akiba H, Tsutsui H,et al. Expression and function of TNF-related apoptosis-inducing ligandon murine activated NK cells. J Immunol 1999;163:1906–1913.
21. Smyth MJ, Cretney E, Takeda K, Wiltrout RH, Sedger LM, Kayagaki N,et al. Tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)contributes to interferon gamma-dependent natural killer cell protectionfrom tumor metastasis. J Exp Med 2001;193:661–670.
22. Bouwens L, Remels L, Baekeland M, Van Bossuyt H, Wisse E. Largegranular lymphocytes or “pit cells” from rat liver: isolation, ultrastructuralcharacterization and natural killer activity. Eur J Immunol 1987;17:37–42.
23. Seglen PO. Preparation of isolated rat liver cells. Methods Cell Biol 1976;13:29–83.
24. Tateno C, Takai-Kajihara K, Yamasaki C, Sato H, Yoshizato K. Hetero-geneity of growth potential of adult rat hepatocytes in vitro. HEPATOLOGY
2000;31:65–74.25. Ostrowska A, Karrer FM, Bilir BM. Histological identification of purified
and cryopreserved allogeneic hepatocytes following transplantation in amurine model without host immunosuppression. Transpl Int 1999;12:188–194.
1330 OCHI ET AL. HEPATOLOGY, May 2004

26. Djeu JY, Heinbaugh JA, Holden HT, Herberman RB. Augmentation ofmouse natural killer cell activity by interferon and interferon inducers.J Immunol 1979;122:175–181.
27. Wiltrout RH, Mathieson BJ, Talmadge JE, Reynolds CW, Zhang SR,Herberman RB, et al. Augmentation of organ-associated natural killeractivity by biological response modifiers. Isolation and characterization oflarge granular lymphocytes from the liver. J Exp Med 1984;160:1431–1449.
28. Vujanovic NL, Polimeno L, Azzarone A, Francavilla A, Chambers WH,Starzl TE, et al. Changes of liver-resident NK cells during liver regenera-tion in rats. J Immunol 1995;154:6324–6338.
29. Vanderkerken K, Bouwens L, Wisse E. Characterization of a phenotypi-cally and functionally distinct subset of large granular lymphocytes (pitcells) in rat liver sinusoids. HEPATOLOGY 1990;12:70–75.
30. Luo D, Vanderkerken K, Chen MC, Vermijlen D, Asosingh K, Willems E,et al. Rat hepatic natural killer cells (pit cells) express mRNA and proteinsimilar to in vitro interleukin-2 activated spleen natural killer cells. CellImmunol 2001;210:41–48.
31. Walczak H, Miller RE, Ariail K, Gliniak B, Griffith TS, Kubin M, et al.Tumoricidal activity of tumor necrosis factor-related apoptosis- inducingligand in vivo. Nat Med 1999;5:157–163.
32. Tanaka S, Sugimachi K, Shirabe K, Shimada M, Wands JR. Expressionand antitumor effects of TRAIL in human cholangiocarcinoma. HEPATOL-OGY 2000;32:523–527.
33. Yamanaka T, Shiraki K, Sugimoto K, Ito T, Fujikawa K, Ito M, et al.Chemotherapeutic agents augment TRAIL-induced apoptosis in humanhepatocellular carcinoma cell lines. HEPATOLOGY 2000;32:482–490.
34. Gores GJ, Kaufmann SH. Is TRAIL hepatotoxic? HEPATOLOGY 2001;34:3–6.
35. Jo M, Kim TH, Seol DW, Esplen JE, Dorko K, Billiar TR, et al. Apoptosisinduced in normal human hepatocytes by tumor necrosis factor- relatedapoptosis-inducing ligand. Nat Med 2000;6:564–567.
36. Ichikawa K, Liu W, Zhao L, Wang Z, Liu D, Ohtsuka T, et al. Tumori-cidal activity of a novel anti-human DR5 monoclonal antibody withouthepatocyte cytotoxicity. Nat Med 2001;7:954–960.
37. Daniels BF, Karlhofer FM, Seaman WE, Yokoyama WM. A natural killercell receptor specific for a major histocompatibility complex class I mole-cule. J Exp Med 1994;180:687–692.
38. Stoneman ER, Bennett M, An J, Chesnut KA, Wakeland EK, Scheerer JB,et al. Cloning and characterization of 5E6(Ly-49C), a receptor moleculeexpressed on a subset of murine natural killer cells. J Exp Med 1995;182:305–313.
39. Brennan J, Lemieux S, Freeman JD, Mager DL, Takei F. Heterogeneityamong Ly-49C natural killer (NK) cells: characterization of highly relatedreceptors with differing functions and expression patterns. J Exp Med1996;184:2085–2090.
40. Hanke T, Takizawa H, McMahon CW, Busch DH, Pamer EG, Miller JD,et al. Direct assessment of MHC class I binding by seven Ly49 inhibitoryNK cell receptors. Immunity 1999;11:67–77.
41. Mason LH, Ortaldo JR, Young HA, Kumar V, Bennett M, Anderson SK.Cloning and functional characteristics of murine large granular lympho-cyte-1: a member of the Ly-49 gene family (Ly-49G2). J Exp Med 1995;182:293–303.
42. Michaelsson J, Achour A, Salcedo M, Kase-Sjostrom A, Sundback J, HarrisRA, et al. Visualization of inhibitory Ly49 receptor specificity with solublemajor histocompatibility complex class I tetramers. Eur J Immunol 2000;30:300–307.
43. Held W, Raulet DH. Ly49A transgenic mice provide evidence for a majorhistocompatibility complex-dependent education process in natural killercell development. J Exp Med 1997;185:2079–2088.
44. Brawand P, Lemonnier FA, MacDonald HR, Cerottini JC, Held W.Transgenic expression of Ly49A on T cells impairs a specific antitumorresponse. J Immunol 2000;165:1871–1876.
45. Fahlen L, Lendahl U, Sentman CL. MHC class I-Ly49 interactions shapethe Ly49 repertoire on murine NK cells. J Immunol 2001;166:6585–6592.
46. Ohashi K, Park F, Kay MA. Hepatocyte transplantation: clinical and ex-perimental application. J Mol Med 2001;79:617–630.
47. Yoshida Y, Tokusashi Y, Lee GH, Ogawa K. Intrahepatic transplantationof normal hepatocytes prevents Wilson’s disease in Long-Evans cinnamonrats. Gastroenterology 1996;111:1654–1660.
48. Gupta S, Chowdhary JR. Hepatocyte transplantation: back to the future.HEPATOLOGY 1992;15:156–162.
49. Gupta S, Rajvanshi P, Sokhi R, Slehria S, Yam A, Kerr A, et al. Entry andintegration of transplanted hepatocytes in rat liver plates occur by disrup-tion of hepatic sinusoidal endothelium. HEPATOLOGY 1999;29:509–519.
HEPATOLOGY, Vol. 39, No. 5, 2004 OCHI ET AL. 1331




















