
Modulation of proteomic profile in H295R adrenocortical cell line induced by mitotane
10
COMMENTARY Modulation of proteomic profile in H295R adrenocortical cell line induced by mitotane A Stigliano 1,2 , L Cerquetti 1,2 , M Borro 3 , G Gentile 3 , B Bucci 2 , S Misiti 1,2 , P Piergrossi 2 , E Brunetti 2 , M Simmaco 3 and V Toscano 1 1 Endocrinology, II Faculty of Medicine, S Andrea Hospital, ‘Sapienza’ University of Rome, 00189 Rome, Italy 2 Research Center, S Pietro Hospital, Rome, Italy 3 Diagnostica Molecolare Avanzata, II Faculty of Medicine, S Andrea Hospital, ‘Sapienza’ University of Rome, 00189 Rome, Italy (Correspondence should be addressed to A Stigliano Endocrinology, II Faculty of Medicine, Research Center, ‘Sapienza’ University of Rome, 00189 Rome, Italy; Email: [email protected]) A Stigliano and L Cerquetti equally contributed to this work Abstract Mitotane, 1,1-dichloro-2-(o-chlorophenyl)-2-(p-chloro-phenyl) ethane (o,p 0 -DDD), is a compound that represents the effective agent in the treatment of the adrenocortical carcinoma (ACC), able to block cortisol synthesis. In this type of cancer, the biological mechanism induced by this treatment remains still unknown. In this study, we have already shown a greater impairment in the first steps of the steroidogenesis and recognized a little effect on cell cycle. We also evaluated the variation of proteomic profile of the H295R ACC cell line, either in total cell extract or in mitochondria-enriched fraction after treatment with mitotane. In total cell extracts, triose phosphate isomerase, a-enolase, D-3-phosphoglycerate dehydrogenase, peroxiredoxin II and VI, heat shock protein 27, prohibitin, histidine triad nucleotide-binding protein, and profilin-1 showed a different expression. In the mitochondrial fraction, the following proteins appeared to be down regulated: aldolase A, peroxiredoxin I, heterogenous nuclear ribonucleoprotein A2/B1, tubulin-b isoform II, heat shock cognate 71 kDa protein, and nucleotide diphosphate kinase, whereas adrenodoxin reductase, cathepsin D, and heat shock 70 kDa protein 1A were positively up-regulated. This study represents the first proteomic study on the mitotane effects on ACC. It permits to identify some protein classes affected by the drug involved in energetic metabolism, stress response, cytoskeleton structure, and tumorigenesis. Endocrine-Related Cancer (2008) 15 1–10 Introduction Mitotane is an adrenocorticolytic drug used for primary treatment and the recurrence of disease in patients affected by adrenocortical carcinoma (ACC; Hahner & Fassnacht 2005). Mitotane acts selectively on the adrenal cortex leading to cell destruction and the impairment of steroidogenesis (Fang 1979, Martz & Straw 1980). At higher concentrations, mitotane produces a dose-related cellular toxic effect with damage on the fasciculata/reticularis areas causing rupture of mitochondrial membranes, but with a minimal effect in the glomerulosa area (Schteingart et al. 1993). It is usually well tolerated in the plasmatic narrow range between 14 and 20 mg/l. Unfortunately, in some cases, its use is limited by a strong toxicity and a relative higher percentage of treated patients show side effects, particularly gastrointestinal and neuro- logical ones (Cai et al. 1997). Recently, there has been a strong interest in applying proteomics to foster a better understanding of disease processes, mechanisms of action, and new pharma- cological drug targets (Hanash 2003). Analyzing the protein expression by comparing the two-dimensional gel electrophoresis patterns of proteomes under different conditions enabled to identify the proteins whose levels significantly vary after treatment with specific compounds. In this study, we have described the effects of mitotane on growth, steroidogenesis, and proteomic profile on H295R cells, a model of ACC able to produce all the adrenocortical steroids Endocrine-Related Cancer (2008) 15 1–10 Endocrine-Related Cancer (2008) 15 1–10 1351–0088/08/015–001 q 2008 Society for Endocrinology Printed in Great Britain DOI: 10.1677/ERC-07-0003 Online version via http://www.endocrinology-journals.org
Transcript of Modulation of proteomic profile in H295R adrenocortical cell line induced by mitotane

COMMENTARYEndocrine-Related Cancer (2008) 15 1–10
Modulation of proteomic profile in H295Radrenocortical cell line induced by mitotane
A Stigliano1,2, L Cerquetti1,2, M Borro3, G Gentile3, B Bucci 2, S Misiti1,2,P Piergrossi2, E Brunetti2, M Simmaco3 and V Toscano1
1Endocrinology, II Faculty of Medicine, S Andrea Hospital, ‘Sapienza’ University of Rome, 00189 Rome, Italy2Research Center, S Pietro Hospital, Rome, Italy3Diagnostica Molecolare Avanzata, II Faculty of Medicine, S Andrea Hospital, ‘Sapienza’ University of Rome, 00189 Rome, Italy
(Correspondence should be addressed to A Stigliano Endocrinology, II Faculty of Medicine, Research Center, ‘Sapienza’ University
of Rome, 00189 Rome, Italy; Email: [email protected])
A Stigliano and L Cerquetti equally contributed to this work
Abstract
Mitotane, 1,1-dichloro-2-(o-chlorophenyl)-2-(p-chloro-phenyl) ethane (o,p0-DDD), is a compound thatrepresents the effective agent in the treatment of the adrenocortical carcinoma (ACC), able to blockcortisol synthesis. In this type of cancer, the biological mechanism induced by this treatment remainsstill unknown. In this study, we have already shown a greater impairment in the first steps of thesteroidogenesis and recognizeda little effecton cell cycle.We also evaluated the variation of proteomicprofile of the H295R ACC cell line, either in total cell extract or in mitochondria-enriched fraction aftertreatment with mitotane. In total cell extracts, triose phosphate isomerase, a-enolase,D-3-phosphoglycerate dehydrogenase, peroxiredoxin II and VI, heat shock protein 27, prohibitin,histidine triad nucleotide-binding protein, and profilin-1 showed a different expression. In themitochondrial fraction, the following proteins appeared to bedown regulated: aldolase A, peroxiredoxinI, heterogenous nuclear ribonucleoprotein A2/B1, tubulin-b isoform II, heat shock cognate 71 kDaprotein, and nucleotide diphosphate kinase, whereas adrenodoxin reductase, cathepsin D, and heatshock 70 kDa protein 1A were positively up-regulated. This study represents the first proteomic studyon the mitotane effectson ACC. It permits to identifysome protein classes affected by the drug involvedin energetic metabolism, stress response, cytoskeleton structure, and tumorigenesis.
Endocrine-Related Cancer (2008) 15 1–10
Introduction
Mitotane is an adrenocorticolytic drug used for
primary treatment and the recurrence of disease in
patients affected by adrenocortical carcinoma (ACC;
Hahner & Fassnacht 2005). Mitotane acts selectively
on the adrenal cortex leading to cell destruction and the
impairment of steroidogenesis (Fang 1979, Martz &
Straw 1980). At higher concentrations, mitotane
produces a dose-related cellular toxic effect with
damage on the fasciculata/reticularis areas causing
rupture of mitochondrial membranes, but with a
minimal effect in the glomerulosa area (Schteingart
et al. 1993). It is usually well tolerated in the plasmatic
narrow range between 14 and 20 mg/l. Unfortunately,
in some cases, its use is limited by a strong toxicity and
Endocrine-Related Cancer (2008) 15 1–10
1351–0088/08/015–001 q 2008 Society for Endocrinology Printed in Great
a relative higher percentage of treated patients show
side effects, particularly gastrointestinal and neuro-
logical ones (Cai et al. 1997).
Recently, there has been a strong interest in applying
proteomics to foster a better understanding of disease
processes, mechanisms of action, and new pharma-
cological drug targets (Hanash 2003). Analyzing the
protein expression by comparing the two-dimensional
gel electrophoresis patterns of proteomes under
different conditions enabled to identify the proteins
whose levels significantly vary after treatment with
specific compounds. In this study, we have described
the effects of mitotane on growth, steroidogenesis,
and proteomic profile on H295R cells, a model of
ACC able to produce all the adrenocortical steroids
Britain
DOI: 10.1677/ERC-07-0003
Online version via http://www.endocrinology-journals.org
A Stigliano, L Cerquetti et al.: Proteomic profile in H295R by mitotane induction
(Rainey et al. 1994). Mitotane-induced different
expression of proteins involved in energetic meta-
bolism, stress response, cytoskeleton structure, and
tumorigenesis. This work represents the first proteomic
study performed on an ACC cell line and the effects
induced by the main drug used for the treatment of this
neoplasia.
Materials and methods
Cell culture and treatments
H295R adrenocortical cells were supplied from the
ATCC (Rockville, MD, USA). Cells were cultured in
DMEM/HAM’S F-12 and medium supplemented with
penicillin/streptomycin 50 U/ml. 24 h post seeding, the
cells were treated with mitotane at 10K5 M final
concentration. This dose has been used to evaluate the
mitotane effect on cell growth and cell cycle at
different times (24, 48, 72, 96 and 120 h). Cell viability
was evaluated by using trypan blue dye exclusion test.
Cell cycle analysis
The cell cycle was studied by using propidium iodide
(PI) staining. Treated and untreated cells were
harvested, washed in cold PBS, fixed in 70% ethanol,
and stained with a solution containing 50 mg/ml PI
(Sigma Chemical) and 75 U/ml RNase (Sigma
Chemical) in PBS. Samples were then measured at a
different time after mitotane treatment by using a
FACScan cytofluorimeter (Becton Dickinson, Sunny-
vale, CA, USA).
Steroid determination
Hormone levels were determined in the cell super-
natant. Progesterone, testosterone and cortisol were
measured by ECLIA (Roche). Aldosterone was
measured by immunoenzymatic assay (DiaMetra,
Milan, Italy).
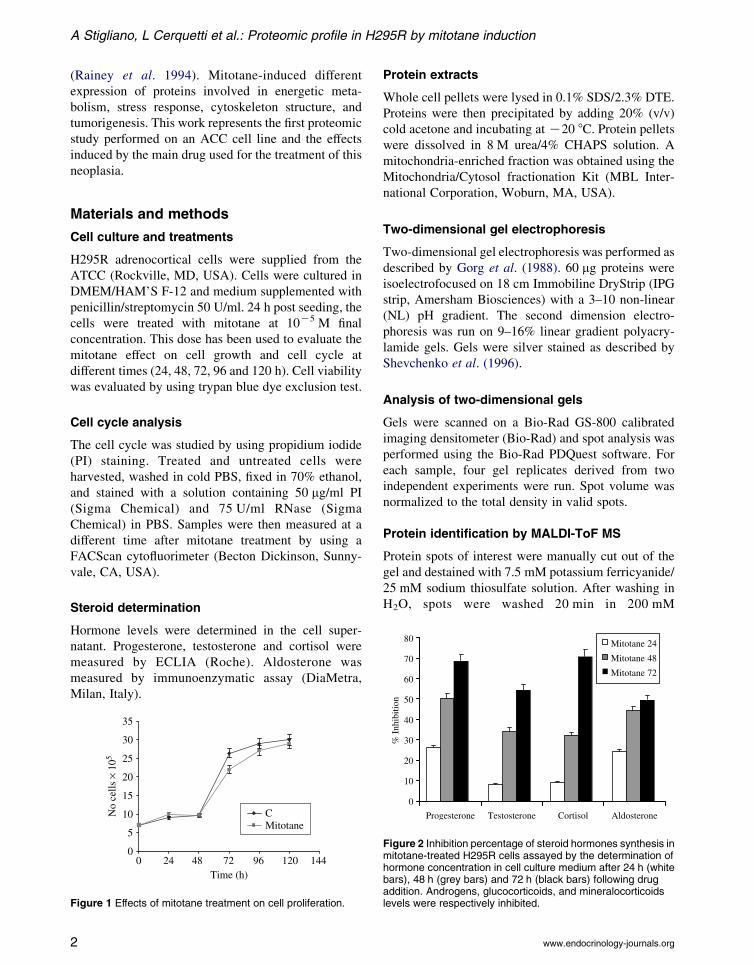
Figure 1 Effects of mitotane treatment on cell proliferation.
2
Protein extracts
Whole cell pellets were lysed in 0.1% SDS/2.3% DTE.
Proteins were then precipitated by adding 20% (v/v)
cold acetone and incubating at K20 8C. Protein pellets
were dissolved in 8 M urea/4% CHAPS solution. A
mitochondria-enriched fraction was obtained using the
Mitochondria/Cytosol fractionation Kit (MBL Inter-
national Corporation, Woburn, MA, USA).
Two-dimensional gel electrophoresis
Two-dimensional gel electrophoresis was performed as
described by Gorg et al. (1988). 60 mg proteins were
isoelectrofocused on 18 cm Immobiline DryStrip (IPG
strip, Amersham Biosciences) with a 3–10 non-linear
(NL) pH gradient. The second dimension electro-
phoresis was run on 9–16% linear gradient polyacry-
lamide gels. Gels were silver stained as described by
Shevchenko et al. (1996).
Analysis of two-dimensional gels
Gels were scanned on a Bio-Rad GS-800 calibrated
imaging densitometer (Bio-Rad) and spot analysis was
performed using the Bio-Rad PDQuest software. For
each sample, four gel replicates derived from two
independent experiments were run. Spot volume was
normalized to the total density in valid spots.
Protein identification by MALDI-ToF MS
Protein spots of interest were manually cut out of the
gel and destained with 7.5 mM potassium ferricyanide/
25 mM sodium thiosulfate solution. After washing in
H2O, spots were washed 20 min in 200 mM
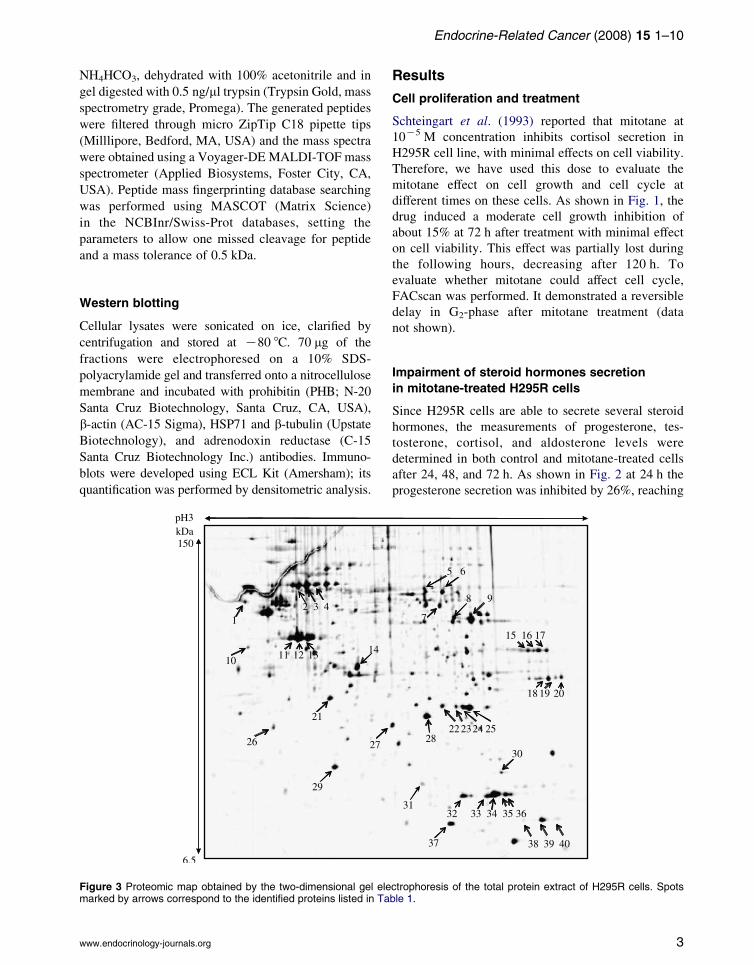
Figure 2 Inhibition percentage of steroid hormones synthesis inmitotane-treated H295R cells assayed by the determination ofhormone concentration in cell culture medium after 24 h (whitebars), 48 h (grey bars) and 72 h (black bars) following drugaddition. Androgens, glucocorticoids, and mineralocorticoidslevels were respectively inhibited.
www.endocrinology-journals.org
Endocrine-Related Cancer (2008) 15 1–10
NH4HCO3, dehydrated with 100% acetonitrile and in
gel digested with 0.5 ng/ml trypsin (Trypsin Gold, mass
spectrometry grade, Promega). The generated peptides
were filtered through micro ZipTip C18 pipette tips
(Milllipore, Bedford, MA, USA) and the mass spectra
were obtained using a Voyager-DE MALDI-TOF mass
spectrometer (Applied Biosystems, Foster City, CA,
USA). Peptide mass fingerprinting database searching
was performed using MASCOT (Matrix Science)
in the NCBInr/Swiss-Prot databases, setting the
parameters to allow one missed cleavage for peptide
and a mass tolerance of 0.5 kDa.
Western blotting
Cellular lysates were sonicated on ice, clarified by
centrifugation and stored at K80 8C. 70 mg of the
fractions were electrophoresed on a 10% SDS-
polyacrylamide gel and transferred onto a nitrocellulose
membrane and incubated with prohibitin (PHB; N-20
Santa Cruz Biotechnology, Santa Cruz, CA, USA),
b-actin (AC-15 Sigma), HSP71 and b-tubulin (Upstate
Biotechnology), and adrenodoxin reductase (C-15
Santa Cruz Biotechnology Inc.) antibodies. Immuno-
blots were developed using ECL Kit (Amersham); its
quantification was performed by densitometric analysis.
Figure 3 Proteomic map obtained by the two-dimensional gel elemarked by arrows correspond to the identified proteins listed in Ta
www.endocrinology-journals.org
Results
Cell proliferation and treatment
Schteingart et al. (1993) reported that mitotane at
10K5 M concentration inhibits cortisol secretion in
H295R cell line, with minimal effects on cell viability.
Therefore, we have used this dose to evaluate the
mitotane effect on cell growth and cell cycle at
different times on these cells. As shown in Fig. 1, the
drug induced a moderate cell growth inhibition of
about 15% at 72 h after treatment with minimal effect
on cell viability. This effect was partially lost during
the following hours, decreasing after 120 h. To
evaluate whether mitotane could affect cell cycle,
FACscan was performed. It demonstrated a reversible
delay in G2-phase after mitotane treatment (data
not shown).
Impairment of steroid hormones secretion
in mitotane-treated H295R cells
Since H295R cells are able to secrete several steroid
hormones, the measurements of progesterone, tes-
tosterone, cortisol, and aldosterone levels were
determined in both control and mitotane-treated cells
after 24, 48, and 72 h. As shown in Fig. 2 at 24 h the
progesterone secretion was inhibited by 26%, reaching
ctrophoresis of the total protein extract of H295R cells. Spotsble 1.
3
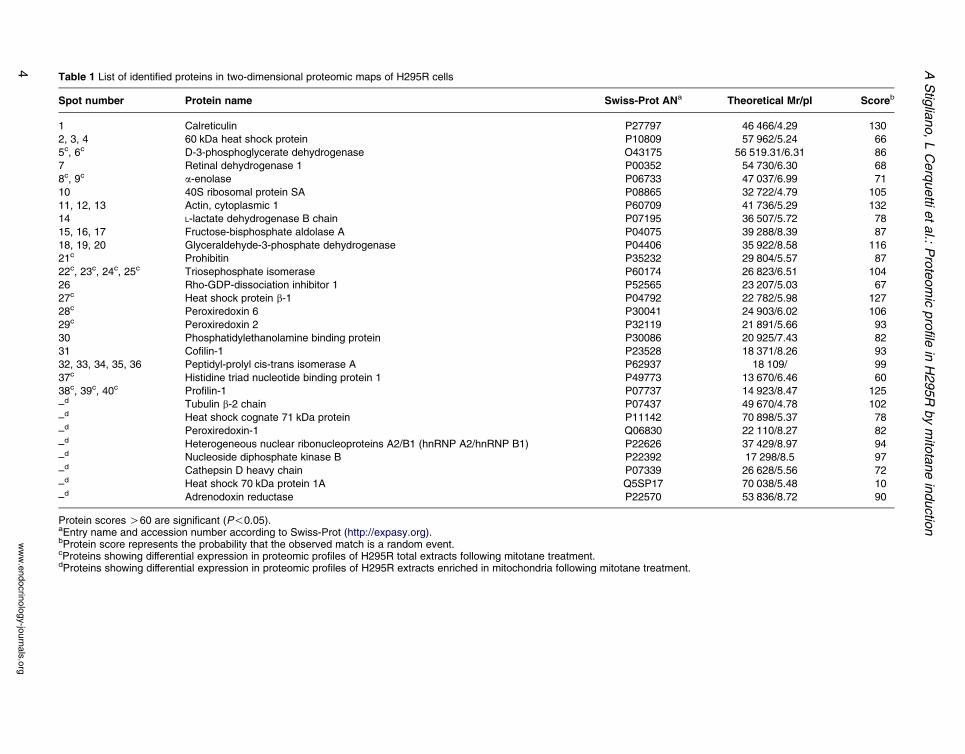
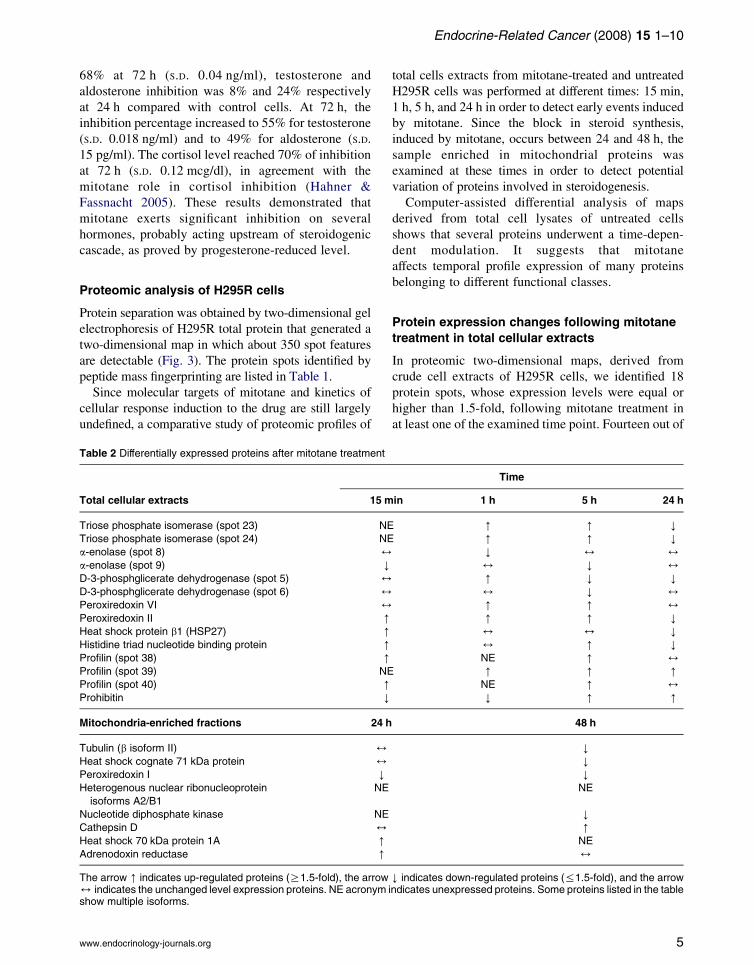
Table 1 List of identified proteins in two-dimensional proteomic maps of H295R cells
Spot number Protein name Swiss-Prot ANa Theoretical Mr/pI Scoreb
1 Calreticulin P27797 46 466/4.29 130
2, 3, 4 60 kDa heat shock protein P10809 57 962/5.24 66
5c, 6c D-3-phosphoglycerate dehydrogenase O43175 56 519.31/6.31 86
7 Retinal dehydrogenase 1 P00352 54 730/6.30 68
8c, 9c a-enolase P06733 47 037/6.99 71
10 40S ribosomal protein SA P08865 32 722/4.79 105
11, 12, 13 Actin, cytoplasmic 1 P60709 41 736/5.29 132
14 L-lactate dehydrogenase B chain P07195 36 507/5.72 78
15, 16, 17 Fructose-bisphosphate aldolase A P04075 39 288/8.39 87
18, 19, 20 Glyceraldehyde-3-phosphate dehydrogenase P04406 35 922/8.58 116
21c Prohibitin P35232 29 804/5.57 87
22c, 23c, 24c, 25c Triosephosphate isomerase P60174 26 823/6.51 104
26 Rho-GDP-dissociation inhibitor 1 P52565 23 207/5.03 67
27c Heat shock protein b-1 P04792 22 782/5.98 127
28c Peroxiredoxin 6 P30041 24 903/6.02 106
29c Peroxiredoxin 2 P32119 21 891/5.66 93
30 Phosphatidylethanolamine binding protein P30086 20 925/7.43 82
31 Cofilin-1 P23528 18 371/8.26 93
32, 33, 34, 35, 36 Peptidyl-prolyl cis-trans isomerase A P62937 18 109/ 99
37c Histidine triad nucleotide binding protein 1 P49773 13 670/6.46 60
38c, 39c, 40c Profilin-1 P07737 14 923/8.47 125
–d Tubulin b-2 chain P07437 49 670/4.78 102
–d Heat shock cognate 71 kDa protein P11142 70 898/5.37 78
–d Peroxiredoxin-1 Q06830 22 110/8.27 82
–d Heterogeneous nuclear ribonucleoproteins A2/B1 (hnRNP A2/hnRNP B1) P22626 37 429/8.97 94
–d Nucleoside diphosphate kinase B P22392 17 298/8.5 97
–d Cathepsin D heavy chain P07339 26 628/5.56 72
–d Heat shock 70 kDa protein 1A Q5SP17 70 038/5.48 10
–d Adrenodoxin reductase P22570 53 836/8.72 90
Protein scores O60 are significant (P!0.05).aEntry name and accession number according to Swiss-Prot (http://expasy.org).bProtein score represents the probability that the observed match is a random event.cProteins showing differential expression in proteomic profiles of H295R total extracts following mitotane treatment.dProteins showing differential expression in proteomic profiles of H295R extracts enriched in mitochondria following mitotane treatment.
AStig
liano,LCerquetti
etal.:
Proteomic
profile
inH295R
bymito
taneinductio
n
ww
w.e
ndocrin
olo
gy-jo
urn
als
.org
4
Endocrine-Related Cancer (2008) 15 1–10
68% at 72 h (S.D. 0.04 ng/ml), testosterone and
aldosterone inhibition was 8% and 24% respectively
at 24 h compared with control cells. At 72 h, the
inhibition percentage increased to 55% for testosterone
(S.D. 0.018 ng/ml) and to 49% for aldosterone (S.D.
15 pg/ml). The cortisol level reached 70% of inhibition
at 72 h (S.D. 0.12 mcg/dl), in agreement with the
mitotane role in cortisol inhibition (Hahner &
Fassnacht 2005). These results demonstrated that
mitotane exerts significant inhibition on several
hormones, probably acting upstream of steroidogenic
cascade, as proved by progesterone-reduced level.
Proteomic analysis of H295R cells
Protein separation was obtained by two-dimensional gel
electrophoresis of H295R total protein that generated a
two-dimensional map in which about 350 spot features
are detectable (Fig. 3). The protein spots identified by
peptide mass fingerprinting are listed in Table 1.
Since molecular targets of mitotane and kinetics of
cellular response induction to the drug are still largely
undefined, a comparative study of proteomic profiles of
Table 2 Differentially expressed proteins after mitotane treatment
Total cellular extracts 15 m
Triose phosphate isomerase (spot 23) NE
Triose phosphate isomerase (spot 24) NE
a-enolase (spot 8) 4a-enolase (spot 9) YD-3-phosphglicerate dehydrogenase (spot 5) 4D-3-phosphglicerate dehydrogenase (spot 6) 4Peroxiredoxin VI 4Peroxiredoxin II [Heat shock protein b1 (HSP27) [Histidine triad nucleotide binding protein [Profilin (spot 38) [Profilin (spot 39) NE
Profilin (spot 40) [Prohibitin Y
Mitochondria-enriched fractions 24 h
Tubulin (b isoform II) 4Heat shock cognate 71 kDa protein 4Peroxiredoxin I YHeterogenous nuclear ribonucleoprotein
isoforms A2/B1
NE
Nucleotide diphosphate kinase NE
Cathepsin D 4Heat shock 70 kDa protein 1A [Adrenodoxin reductase [
The arrow [ indicates up-regulated proteins (R1.5-fold), the arrow4 indicates the unchanged level expression proteins. NE acronymshow multiple isoforms.
www.endocrinology-journals.org
total cells extracts from mitotane-treated and untreated
H295R cells was performed at different times: 15 min,
1 h, 5 h, and 24 h in order to detect early events induced
by mitotane. Since the block in steroid synthesis,
induced by mitotane, occurs between 24 and 48 h, the
sample enriched in mitochondrial proteins was
examined at these times in order to detect potential
variation of proteins involved in steroidogenesis.
Computer-assisted differential analysis of maps
derived from total cell lysates of untreated cells
shows that several proteins underwent a time-depen-
dent modulation. It suggests that mitotane
affects temporal profile expression of many proteins
belonging to different functional classes.
Protein expression changes following mitotane
treatment in total cellular extracts
In proteomic two-dimensional maps, derived from
crude cell extracts of H295R cells, we identified 18
protein spots, whose expression levels were equal or
higher than 1.5-fold, following mitotane treatment in
at least one of the examined time point. Fourteen out of
Time
in 1 h 5 h 24 h
[ [ Y[ [ YY 4 44 Y 4[ Y Y4 Y 4[ [ 4[ [ Y4 4 Y4 [ YNE [ 4[ [ [
NE [ 4Y [ [
48 h
YYY
NE
Y[
NE
4
Y indicates down-regulated proteins (%1.5-fold), and the arrowindicates unexpressed proteins. Some proteins listed in the table
5
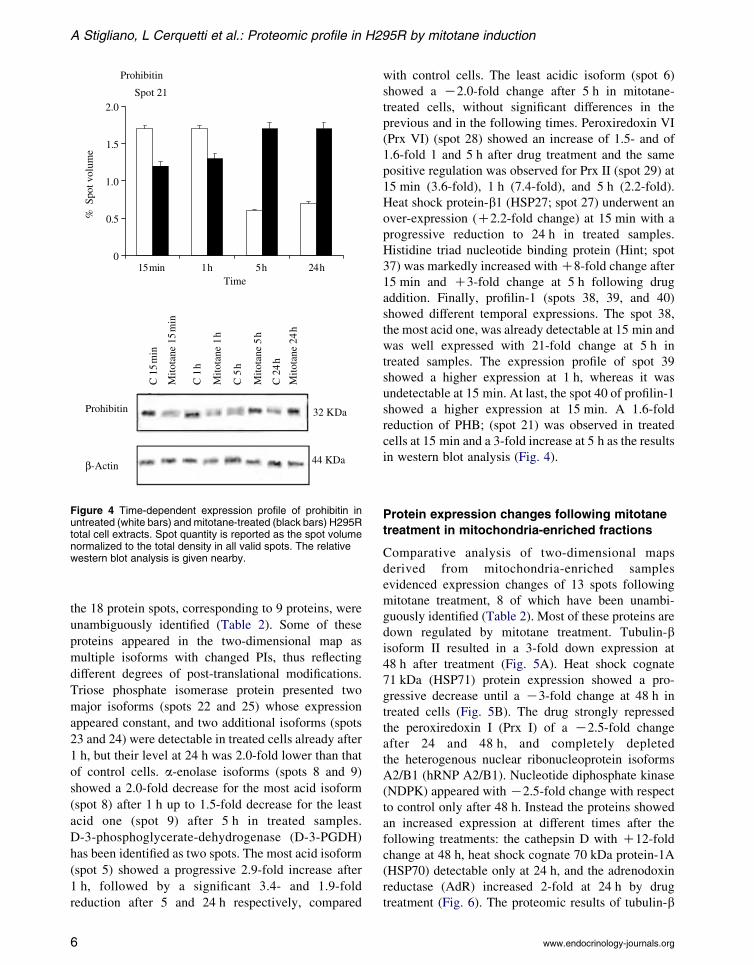
Figure 4 Time-dependent expression profile of prohibitin inuntreated (white bars) and mitotane-treated (black bars) H295Rtotal cell extracts. Spot quantity is reported as the spot volumenormalized to the total density in all valid spots. The relativewestern blot analysis is given nearby.
A Stigliano, L Cerquetti et al.: Proteomic profile in H295R by mitotane induction
the 18 protein spots, corresponding to 9 proteins, were
unambiguously identified (Table 2). Some of these
proteins appeared in the two-dimensional map as
multiple isoforms with changed PIs, thus reflecting
different degrees of post-translational modifications.
Triose phosphate isomerase protein presented two
major isoforms (spots 22 and 25) whose expression
appeared constant, and two additional isoforms (spots
23 and 24) were detectable in treated cells already after
1 h, but their level at 24 h was 2.0-fold lower than that
of control cells. a-enolase isoforms (spots 8 and 9)
showed a 2.0-fold decrease for the most acid isoform
(spot 8) after 1 h up to 1.5-fold decrease for the least
acid one (spot 9) after 5 h in treated samples.
D-3-phosphoglycerate-dehydrogenase (D-3-PGDH)
has been identified as two spots. The most acid isoform
(spot 5) showed a progressive 2.9-fold increase after
1 h, followed by a significant 3.4- and 1.9-fold
reduction after 5 and 24 h respectively, compared
6
with control cells. The least acidic isoform (spot 6)
showed a K2.0-fold change after 5 h in mitotane-
treated cells, without significant differences in the
previous and in the following times. Peroxiredoxin VI
(Prx VI) (spot 28) showed an increase of 1.5- and of
1.6-fold 1 and 5 h after drug treatment and the same
positive regulation was observed for Prx II (spot 29) at
15 min (3.6-fold), 1 h (7.4-fold), and 5 h (2.2-fold).
Heat shock protein-b1 (HSP27; spot 27) underwent an
over-expression (C2.2-fold change) at 15 min with a
progressive reduction to 24 h in treated samples.
Histidine triad nucleotide binding protein (Hint; spot
37) was markedly increased withC8-fold change after
15 min and C3-fold change at 5 h following drug
addition. Finally, profilin-1 (spots 38, 39, and 40)
showed different temporal expressions. The spot 38,
the most acid one, was already detectable at 15 min and
was well expressed with 21-fold change at 5 h in
treated samples. The expression profile of spot 39
showed a higher expression at 1 h, whereas it was
undetectable at 15 min. At last, the spot 40 of profilin-1
showed a higher expression at 15 min. A 1.6-fold
reduction of PHB; (spot 21) was observed in treated
cells at 15 min and a 3-fold increase at 5 h as the results
in western blot analysis (Fig. 4).
Protein expression changes following mitotane
treatment in mitochondria-enriched fractions
Comparative analysis of two-dimensional maps
derived from mitochondria-enriched samples
evidenced expression changes of 13 spots following
mitotane treatment, 8 of which have been unambi-
guously identified (Table 2). Most of these proteins are
down regulated by mitotane treatment. Tubulin-bisoform II resulted in a 3-fold down expression at
48 h after treatment (Fig. 5A). Heat shock cognate
71 kDa (HSP71) protein expression showed a pro-
gressive decrease until a K3-fold change at 48 h in
treated cells (Fig. 5B). The drug strongly repressed
the peroxiredoxin I (Prx I) of a K2.5-fold change
after 24 and 48 h, and completely depleted
the heterogenous nuclear ribonucleoprotein isoforms
A2/B1 (hRNP A2/B1). Nucleotide diphosphate kinase
(NDPK) appeared withK2.5-fold change with respect
to control only after 48 h. Instead the proteins showed
an increased expression at different times after the
following treatments: the cathepsin D with C12-fold
change at 48 h, heat shock cognate 70 kDa protein-1A
(HSP70) detectable only at 24 h, and the adrenodoxin
reductase (AdR) increased 2-fold at 24 h by drug
treatment (Fig. 6). The proteomic results of tubulin-b
www.endocrinology-journals.org
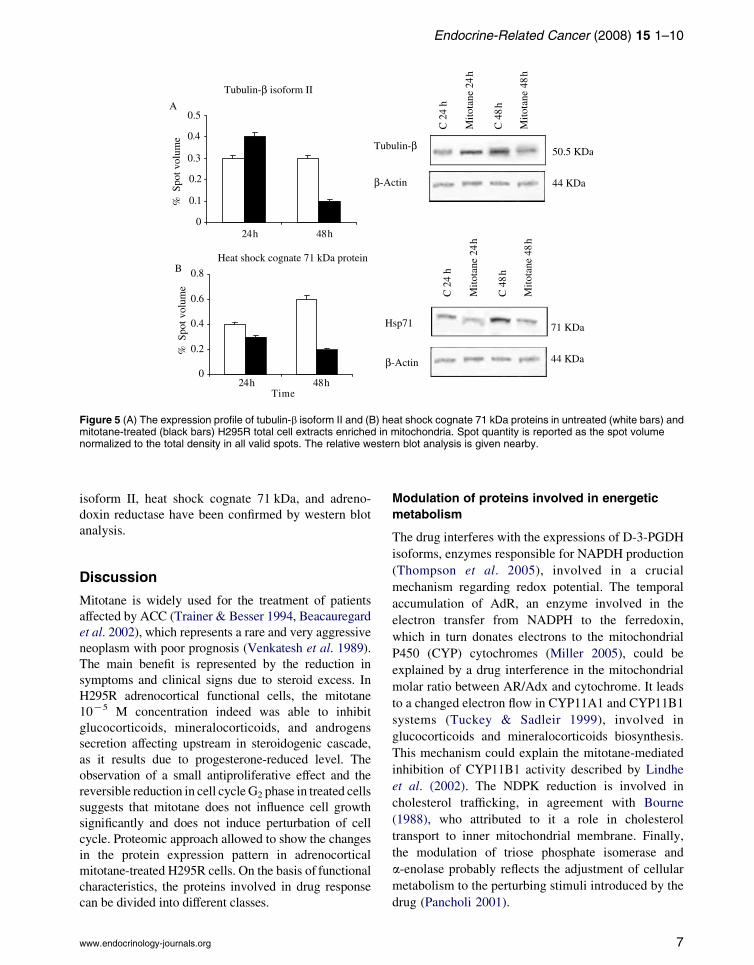
Figure 5 (A) The expression profile of tubulin-b isoform II and (B) heat shock cognate 71 kDa proteins in untreated (white bars) andmitotane-treated (black bars) H295R total cell extracts enriched in mitochondria. Spot quantity is reported as the spot volumenormalized to the total density in all valid spots. The relative western blot analysis is given nearby.
Endocrine-Related Cancer (2008) 15 1–10
isoform II, heat shock cognate 71 kDa, and adreno-
doxin reductase have been confirmed by western blot
analysis.
Discussion
Mitotane is widely used for the treatment of patients
affected by ACC (Trainer & Besser 1994, Beacauregard
et al. 2002), which represents a rare and very aggressive
neoplasm with poor prognosis (Venkatesh et al. 1989).
The main benefit is represented by the reduction in
symptoms and clinical signs due to steroid excess. In
H295R adrenocortical functional cells, the mitotane
10K5 M concentration indeed was able to inhibit
glucocorticoids, mineralocorticoids, and androgens
secretion affecting upstream in steroidogenic cascade,
as it results due to progesterone-reduced level. The
observation of a small antiproliferative effect and the
reversible reduction in cell cycleG2 phase in treated cells
suggests that mitotane does not influence cell growth
significantly and does not induce perturbation of cell
cycle. Proteomic approach allowed to show the changes
in the protein expression pattern in adrenocortical
mitotane-treated H295R cells. On the basis of functional
characteristics, the proteins involved in drug response
can be divided into different classes.
www.endocrinology-journals.org
Modulation of proteins involved in energetic
metabolism
The drug interferes with the expressions of D-3-PGDH
isoforms, enzymes responsible for NAPDH production
(Thompson et al. 2005), involved in a crucial
mechanism regarding redox potential. The temporal
accumulation of AdR, an enzyme involved in the
electron transfer from NADPH to the ferredoxin,
which in turn donates electrons to the mitochondrial
P450 (CYP) cytochromes (Miller 2005), could be
explained by a drug interference in the mitochondrial
molar ratio between AR/Adx and cytochrome. It leads
to a changed electron flow in CYP11A1 and CYP11B1
systems (Tuckey & Sadleir 1999), involved in
glucocorticoids and mineralocorticoids biosynthesis.
This mechanism could explain the mitotane-mediated
inhibition of CYP11B1 activity described by Lindhe
et al. (2002). The NDPK reduction is involved in
cholesterol trafficking, in agreement with Bourne
(1988), who attributed to it a role in cholesterol
transport to inner mitochondrial membrane. Finally,
the modulation of triose phosphate isomerase and
a-enolase probably reflects the adjustment of cellular
metabolism to the perturbing stimuli introduced by the
drug (Pancholi 2001).
7
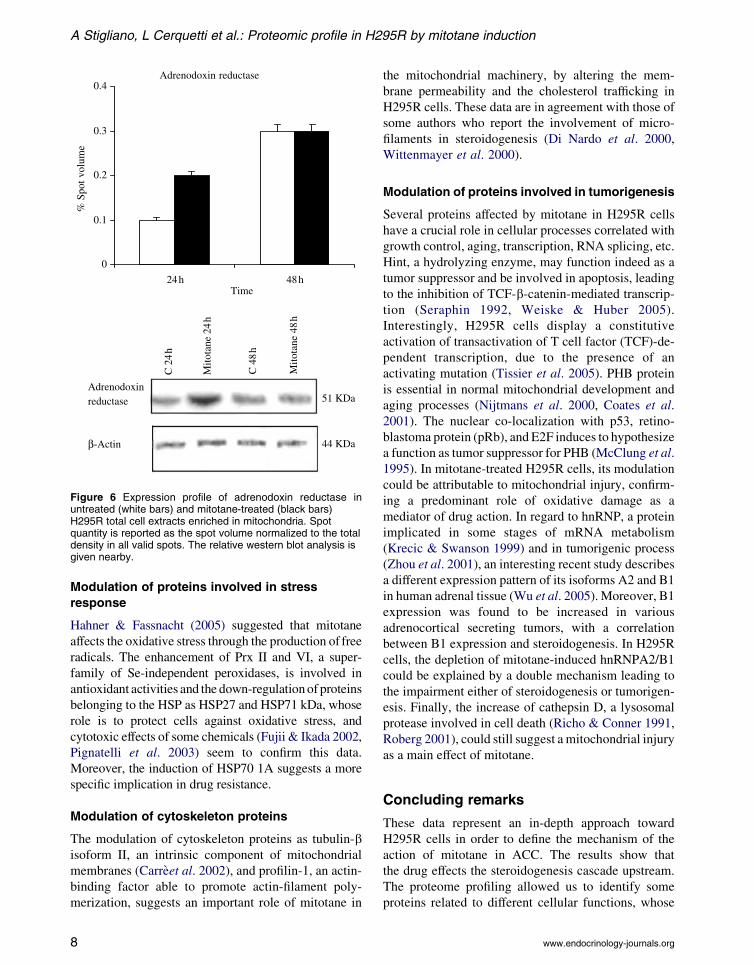
Figure 6 Expression profile of adrenodoxin reductase inuntreated (white bars) and mitotane-treated (black bars)H295R total cell extracts enriched in mitochondria. Spotquantity is reported as the spot volume normalized to the totaldensity in all valid spots. The relative western blot analysis isgiven nearby.
A Stigliano, L Cerquetti et al.: Proteomic profile in H295R by mitotane induction
Modulation of proteins involved in stress
response
Hahner & Fassnacht (2005) suggested that mitotane
affects the oxidative stress through the production of free
radicals. The enhancement of Prx II and VI, a super-
family of Se-independent peroxidases, is involved in
antioxidant activities and the down-regulationof proteins
belonging to the HSP as HSP27 and HSP71 kDa, whose
role is to protect cells against oxidative stress, and
cytotoxic effects of some chemicals (Fujii & Ikada 2002,
Pignatelli et al. 2003) seem to confirm this data.
Moreover, the induction of HSP70 1A suggests a more
specific implication in drug resistance.
Modulation of cytoskeleton proteins
The modulation of cytoskeleton proteins as tubulin-bisoform II, an intrinsic component of mitochondrial
membranes (Carreet al. 2002), and profilin-1, an actin-
binding factor able to promote actin-filament poly-
merization, suggests an important role of mitotane in
8
the mitochondrial machinery, by altering the mem-
brane permeability and the cholesterol trafficking in
H295R cells. These data are in agreement with those of
some authors who report the involvement of micro-
filaments in steroidogenesis (Di Nardo et al. 2000,
Wittenmayer et al. 2000).
Modulation of proteins involved in tumorigenesis
Several proteins affected by mitotane in H295R cells
have a crucial role in cellular processes correlated with
growth control, aging, transcription, RNA splicing, etc.
Hint, a hydrolyzing enzyme, may function indeed as a
tumor suppressor and be involved in apoptosis, leading
to the inhibition of TCF-b-catenin-mediated transcrip-
tion (Seraphin 1992, Weiske & Huber 2005).
Interestingly, H295R cells display a constitutive
activation of transactivation of T cell factor (TCF)-de-
pendent transcription, due to the presence of an
activating mutation (Tissier et al. 2005). PHB protein
is essential in normal mitochondrial development and
aging processes (Nijtmans et al. 2000, Coates et al.
2001). The nuclear co-localization with p53, retino-
blastoma protein (pRb), and E2F induces to hypothesize
a function as tumor suppressor for PHB (McClung et al.
1995). In mitotane-treated H295R cells, its modulation
could be attributable to mitochondrial injury, confirm-
ing a predominant role of oxidative damage as a
mediator of drug action. In regard to hnRNP, a protein
implicated in some stages of mRNA metabolism
(Krecic & Swanson 1999) and in tumorigenic process
(Zhou et al. 2001), an interesting recent study describes
a different expression pattern of its isoforms A2 and B1
in human adrenal tissue (Wu et al. 2005). Moreover, B1
expression was found to be increased in various
adrenocortical secreting tumors, with a correlation
between B1 expression and steroidogenesis. In H295R
cells, the depletion of mitotane-induced hnRNPA2/B1
could be explained by a double mechanism leading to
the impairment either of steroidogenesis or tumorigen-
esis. Finally, the increase of cathepsin D, a lysosomal
protease involved in cell death (Richo & Conner 1991,
Roberg 2001), could still suggest amitochondrial injury
as a main effect of mitotane.
Concluding remarks
These data represent an in-depth approach toward
H295R cells in order to define the mechanism of the
action of mitotane in ACC. The results show that
the drug effects the steroidogenesis cascade upstream.
The proteome profiling allowed us to identify some
proteins related to different cellular functions, whose
www.endocrinology-journals.org

Endocrine-Related Cancer (2008) 15 1–10
expressions were altered by mitotane treatment. Even
if further studies are needed in order to improve the
understanding of mitotane action in ACC therapy,
the identified proteins might represent good targets for
the development of strategies directed to contrast ACC.
Acknowledgements
This work was partially financed by research grants
(progetti di rilevante interesse nazionale) fromMinistero
dell’Universita e della Ricerca Scientifica e Tecnologica
(MURST) and ‘Sapienza’ Universita di Roma. The
authors declare that there is no conflict of interest that
would prejudice the impartiality of this scientific work.
References
Beacauregard C, Dickstein G & Lacroix A 2002 Classic and
recent etiologies of Cushing’s syndrome: diagnosis and
therapy. Treatments in Endocrinology 1 79–94.
Bourne HR 1988 Do GTPases direct membrane traffic in
secretion? Cell 53 669–671.
Cai W, Counsell RE, Schteingart DE, Sinsheimer JE,
Vaz AD & Wotring LL 1997 Adrenal proteins bound
by a reactive intermediate of mitotane. Cancer
Chemotherapy and Pharmacology 39 537–540.
Carre M, Andre N, Carles G, Borghi H, Brichese L,
Briand C & Braguer D 2002 Tubulin is an inherent
component of mitochondrial membranes that interacts
with the voltage-dependent anion channel. Journal of
Biological Chemistry 277 33664–33669.
Coates PJ, Nenutil R, McGregor A, Picksley SM, Crouch DH,
Hall PA&Wright EG 2001Mammalian prohibitin proteins
respond tomitochondrial stress and decrease during cellular
senescence. Experimental Cell Research 265 262–273.
Fang VS 1979 Cytotoxic activity of 1-(o-chlorophenyl)-1-
(p-chlorophenyl)-2,2-dichloroethane (mitotane) and its
analogs on feminizing adrenal neoplastic cells in culture.
Cancer Research 39 139–145.
Fujii J & Ikeda Y 2002 Advances in our understanding of
peroxiredoxin, a multifunctional, mammalian redox
protein. Redox Report 7 123–130.
Gorg A, Postel W&Gunther S 1988 The current state of two-
dimensional electrophoresis with immobilized pH
gradients. Electrophoresis 9 531–546.
Hahner S & Fassnacht M 2005 Mitotane for adrenocortical
carcinoma treatment. Current Opinion in Investigational
Drugs 6 386–394.
Hanash S 2003 Disease proteomics. Nature 13 226–232.
Krecic AM & Swanson MS 1999 hnRNP complexes:
composition, structure, and function. Current Opinion in
Cell Biology 11 363–371.
Lindhe O, Skogseid B & Brandt I 2002 Cytochrome P450-
catalyzed binding of 3- methylsulfonyl-DE and o,p 0-DDD
in human adrenal zona fasciculata/reticularis. Journal of
Clinical Endocrinology and Metabolism 87 1319–1326.
www.endocrinology-journals.org
Martz F & Straw JA 1980Metabolism and covalent binding of
1-(o-chlorophenyl)-1-(p- chlorophenyl)-2,2-dichloroethane
(o,p0-DDD). Correlation between adrenocorticolytic
activity and metabolic activation by adrenocortical mito-
chondria. Drug Metabolism and Disposition 8 127–130.
McClung JK, Jupe ER, Liu XT & Dell’Orco RT 1995
Prohibitin: potential role in senescence, development, and
tumor suppression. Experimental Gerontology 30 99–124.
Miller WL 2005 Minireview: regulation of steroidogenesis
by electron transfer. Endocrinology 146 2544–2550.
Di Nardo A, Gareus R, Kwiatkowski D & Witke W 2000
Alternative splicing of the mouse profilin II gene
generates functionally different profilin isoforms.
Journal of Cell Science 113 3795–3803.
NijtmansLG,NijtmansLG, de JongL,ArtalSanzM,CoatesPJ,
Berden JA, Back JW, Muijsers AO, van der Spek H &
Grivell LA 2000 Prohibitins act as a membrane-bound
chaperone for the stabilization of mitochondrial proteins.
EMBO Journal 19 2444–2451.
Pancholi V 2001 Multifunctional alpha-enolase: its role in
diseases.Cellular andMolecular Life Sciences 58 902–920.
Pignatelli D, Ferriera J, Soares P, Costa MJ &Magalhaes MC
2003 Immunohistochemical study of heat shock proteins
27, 60 and 70 in the normal human adrenal and in adrenal
tumors with suppressed ACTH production. Microscopy
Research and Technique 15 315–323.
RaineyWE,Bird IM&Mason JI 1994TheNCI-H295 cell line:
a pluripotent model for human adrenocortical studies.
Molecular and Cellular Endocrinology 100 45–50.
Richo G & Conner GE 1991 Proteolytic activation of human
procathepsin D. Advances in Experimental Medicine and
Biology 306 289–296.
Roberg K 2001 Relocalization of cathepsin D and cyto-
chrome c early in apoptosis revealed by immunoelectron
microscopy. Laboratory Investigation 81 149–158.
Schteingart DE, Sinsheimer JE, Counsell RE, Abrams GD, Mc
Clellan N, Djanegara T, Hines J, Ruangwises N, Benitez R
&Wotring LL 1993 Comparison of the adrenalytic activity
of mitotane and a methylated homolog on normal adrenal
cortex and adrenal cortical carcinoma. Cancer Chemo-
therapy and Pharmacology 31 459–466.
Seraphin B 1992 The HIT protein family: a new family of
proteins present in prokaryotes, yeast and mammals. DNA
Sequence 3 177–179.
Shevchenko A, Wilm M, Vorm O & Mann M 1996 Mass
spectrometric sequencing of proteins silver-stained poly-
acrylamide gels. Analytical Chemistry 68 850–858.
Thompson JR, Bell JK, Bratt J, Grant GA & Banaszak LJ
2005 Vmax regulation through domain and subunit
changes. The active form of phosphoglycerate dehydro-
genase. Biochemistry 44 5763–5773.
Tissier F, Cavard C, Groussin L, Perlemoine K, Fumey G,
Hagnere AM, Rene-Corail F, Jullian E, Gicquel C,
Bertagna X et al. 2005 Mutations of beta-catenin in
adrenocortical tumors: activation of the Wnt signaling
pathway is a frequent event in both benign and malignant
adrenocortical tumors. Cancer Research 65 7622–7627.
9

A Stigliano, L Cerquetti et al.: Proteomic profile in H295R by mitotane induction
Trainer PJ & Besser M 1994 Cushing’s syndrome.
Therapy directed at the adrenal glands. Endocrinology
and Metabolism Clinics of North America 23
571–584.
Tuckey RC & Sadleir J 1999 The concentration of
adrenodoxin reductase limits cytochrome p450scc
activity in the human placenta. European Journal of
Biochemistry 263 319–325.
Venkatesh S, Hickey RC, Sellin RV, Fernandez JF &
Samaan NA 1989 Adrenal cortical carcinoma. Cancer
64 765–769.
Weiske J & Huber O 2005 The histidine triad protein Hint1
interacts with Pontin and Reptin and inhibits TCF-b-catenin-mediated transcription. Journal of Cell Science
118 3117–3129.
10
Wittenmayer N, Rothkegel M, Jockusch BM & Schluter K
2000 Functional characterization of green fluorescent
protein-profilin fusion proteins. European Journal of
Biochemistry 267 5247–5256.
Wu W, Kamma H, Fujiwara M, Yano Y, Satoh H, Hara H,
Yashiro T, Ueno E & Aiyoshi Y 2005 Altered expression
patterns of heterogeneous nuclear ribonucleoproteins A2
and B1 in the adrenal cortex. Journal of Histochemistry
and Cytochemistry 53 487–495.
Zhou J, Allred DC, Avis I, Martinez A, Vos MD, Smith L,
Treston AM & Mulshine JL 2001 Differential expression
of the early lung cancer detection marker, heterogeneous
nuclear ribonucleoprotein-A2/B1 (hnRNP- A2/B1) in
normal breast and neoplastic breast cancer. Breast Cancer
Research and Treatment 66 217–224.
www.endocrinology-journals.org




















