
A model of spike initiation in neocortical pyramidal neurons

Available online at www.sciencedirect.com
www.elsevier.com/locate/brainres
b r a i n r e s e a r c h ] ( ] ] ] ] ) ] ] ] – ] ] ]
http://dx.doi.org/100006-8993/& 2014 Pu
nCorresponding autE-mail addresses1MP and CMF ar
Please cite this arfollowing prenabrainres.2014.06.
Research Report
Modified neocortical and cerebellar proteinexpression and morphology in adult rats followingprenatal inhibition of the kynurenine pathway
Mazura Pisara,1, Caroline M. Forresta,1, Omari S. Khalila, Kara McNaira,Maria C.J. Vincentena, Susana Qasema, L. Gail Darlingtonb, Trevor W. Stonea,n
aInstitute of Neuroscience and Psychology, West Medical Building, University of Glasgow, Glasgow G12 8QQ, UKbAshtead Hospital, The Warren, Ashtead, Surrey KT11 2SB, UK
a r t i c l e i n f o
Article history:
Accepted 13 June 2014
Inhibition of the kynurenine pathway of tryptophan metabolism during gestation can
lead to changes in synaptic transmission, neuronal morphology and plasticity in the rat
Keywords:
Kynurenines
Kynurenic acid
Neurodevelopment
Glutamate transport
GABA transport
Doublecortin
Sonic hedgehog
.1016/j.brainres.2014.06.01blished by Elsevier B.V.
hor. Fax: þ44 141 330 548: [email protected] equal first authors.
ticle as: Pisar, M., et al.,tal inhibition of t016
a b s t r a c t
hippocampus. This suggests a role for the kynurenine pathway in early brain development,
probably caused by kynurenine modulation of N-methyl-D-aspartate (NMDA) glutamate
receptors which are activated by the tryptophan metabolite quinolinic acid and blocked by
kynurenic acid. We have now examined samples of neocortex and cerebellum of adult
animals to assess the effects of a prenatally administered kynurenine-3-monoxygenase
inhibitor (Ro61-8048) on protein and mRNA expression, dendritic structure and immuno-
histochemistry. No changes were seen in mRNA expression using quantitative real-time
polymerase chain reaction. Changes were detected in the expression of several proteins
including the GluN2A subunit, unco-ordinated-5H3 (unc5H3), doublecortin, cyclo-oxyge-
nase, sonic hedgehog and Disrupted in schizophrenia-1 (DISC1), although no differences in
immunoreactive cell numbers were observed. In the midbrain, dependence receptor
expression was also changed. The numbers and lengths of individual dendritic regions
were not changed but there were significant increases in the overall complexity values
of apical and basal dendritic trees. The data support the hypothesis that constitutive
kynurenine metabolism plays a critical role in early, embryonic brain development,
although fewer effects are produced in the neocortex and cerebellum than in the
hippocampus and the nature of the changes seen are qualitatively different. The
significant changes in DISC1 and unc5H3 may be relevant to cerebellar dysfunction and
schizophrenia respectively, in which these proteins have been previously implicated.
& 2014 Published by Elsevier B.V.
6
1.c.uk, [email protected] (T.W. Stone).
Modified neocortical and cerebellar protein expression and morphology in adult ratshe kynurenine pathway. Brain Research (2014), http://dx.doi.org/10.1016/j.

b r a i n r e s e a r c h ] ( ] ] ] ] ) ] ] ] – ] ] ]2
1. Introduction
It is now recognised that glutamate receptors are cruciallyinvolved in the earliest phases of brain development, espe-cially those sensitive to the synthetic agonist N-methyl-D-aspartate (NMDA). Activation or blockade of these receptorscan affect the generation of neurites and axon branches aswell as the guidance of axons towards their appropriatecellular targets and the formation, stabilisation and functio-nalization of synaptic contacts (Rajan and Cline, 1998;Colonnese et al., 2005; Alvarez et al., 2007; Ultanir et al.,2007). These various facets of neuronal development deter-mine the details of neuronal structure, connectivity andsynaptic plasticity in the offspring (Iwasato et al., 2000;Ramoa et al., 2001).
Under the influence of external factors during pregnancy,many of these aspects of neuronal development can bemodified, the best studied influences being exposure toinfection or stress (Hornig et al., 1999; Meyer and Feldon,2010) with strong evidence that CNS function, includingbehaviour, can be modified permanently after brief exposurein utero. Such changes may contribute to the production ofneurological or psychiatric disorders at maturity (Horniget al., 1999; Pearce, 2003; Brown, 2006, 2011; Meyer andFeldon, 2010) although the molecular events mediating thelink between infection or stress and abnormal developmentare not clear.
The kynurenine pathway of tryptophan oxidation includesseveral compounds with marked neuroactive propertiesincluding the NMDA receptor agonist quinolinic acid (Stoneand Perkins, 1981; Stone and Darlington, 2002) andthe antagonist kynurenic acid (Perkins and Stone, 1982;Stone et al., 2013). Although kynurenic acid can also blocknon-NMDA receptors, this is a less potent action thanthe blockade of NMDA receptors. Kynurenic acid may alsoblock receptors for acetylcholine (Hilmas et al., 2001) butseveral groups have been unable to confirm the existence orpotency of this activity (Stone, 2007; Mok et al., 2009; Dobeliset al., 2012).
The probable involvement of the kynurenine pathwayin mediating external modulation of neurodevelopment isstrengthened by the fact that it is activated during infectionby interferons and pro-inflammatory cytokines which acti-vate the initial enzyme indoleamine-2,3-dioxygenase (IDO)and during exposure to stress by the corticosteroid inductionand activation of tryptophan-2,3-dioxygenase (TDO). Thisproposal has been supported by our demonstration thatprenatal interruption of the kynurenine pathway using theselective inhibitor of kynurenine-3-monoxygenase (KMO)Ro61-8048 can generate changes of protein expression andlocalisation, neurogenesis, neuronal structure and synaptictransmission in the hippocampus (Forrest et al., 2013a,2013b). Changes in NMDA subunit expression in the embryosare apparent as early as 5 h after treatment but the functionaland morphological effects persist into adulthood. We havenow sought to determine whether the changes described inthe hippocampus are representative of the whole brain orwhether there are regional variations throughout the CNS inthe effects of kynurenine pathway inhibition.
Please cite this article as: Pisar, M., et al., Modified neocortical anfollowing prenatal inhibition of the kynurenine pathwbrainres.2014.06.016
2. Results
2.1. Neocortex
2.1.1. Protein expressionIn order to compare the changes occurring in the neocortexand cerebellum with those observed previously in the hippo-campus, a similar range of proteins was examined in thisstudy, including proteins involved in neuronal migration,axon guidance, neurogenesis and dendrite formation. Of 19proteins examined, the majority showed no differencebetween control and drug-exposed animals. In neocortexthe presynaptic proteins synaptophysin, synaptotagmin andVesicle Associated Membrane Protein-1 (VAMP-1; synapto-brevin) were unchanged, as were the cytoskeletal regulatoryRhoGTPases RhoA (p¼0.95) and RhoB (p¼0.31) even thoughthere was a substantial increase in RhoB at P21 (Forrest et al.,2013a). This may reflect the increasing degree of CNS matura-tion and stability compared with the P21 stage.
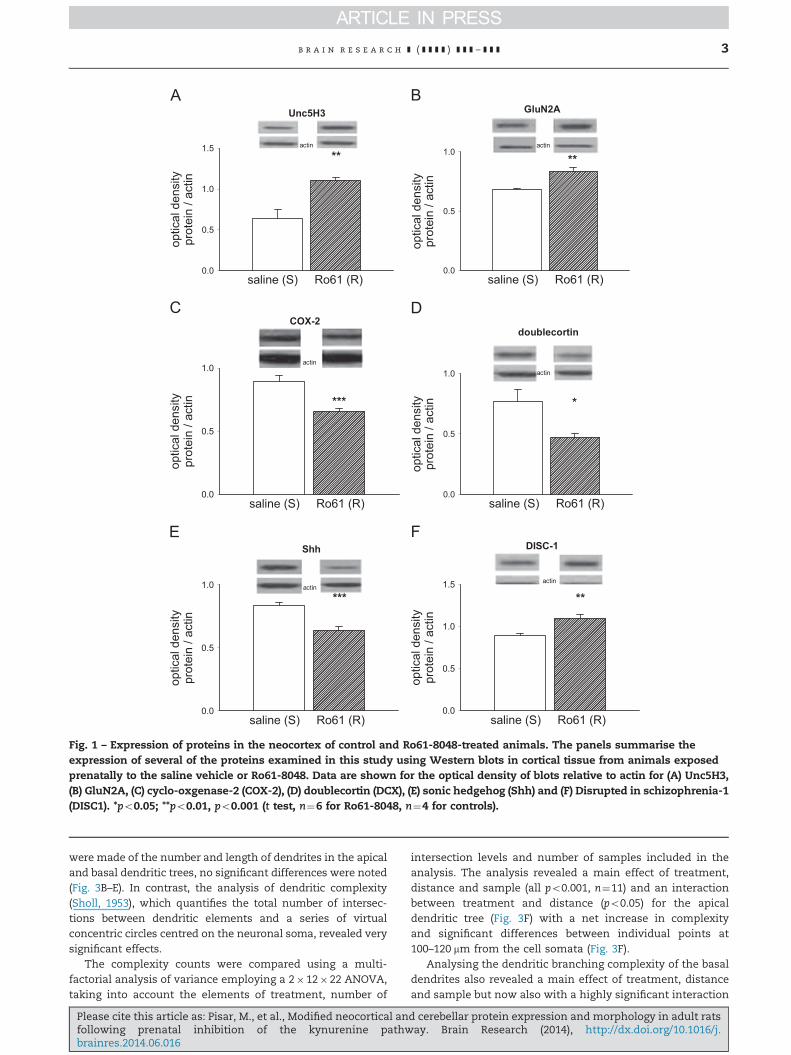
Of three dependence receptors for the axon guidance proteinnetrin: Unc5H1, Unc5H3 and Deleted in Colorectal Cancer (DCC),only Unc5H3 showed any significant change, increasing by over40% compared with control animals (po0.001; Fig. 1A).
In addition to Unc5H3, five other proteins exhibited signifi-cant changes in the neocortex of Ro61-8048-treated animals.A significant difference was noted in the expression of theGluN2A subunit for which expression was increased 20%(p¼o0.01; Fig. 1B), although there were no changes detected inthe GluN1 or GluN2B subunits, or in the NMDA receptor-associated post-synaptic density protein PSD-95. Two proteinsintimately reflecting inflammatory processes were examined,with a highly significant, 30% decrease observed in cyclo-oxygenase-2 (COX-2) expression (po0.001; Fig. 1C) but no changein Nuclear Factor kappa-B (NFkB) (data not shown).
Three of the proteins tested are linked with neurogenesis.Proliferating Cell Nuclear Antigen (PCNA) binds to a nuclearprotein in mature cells but its expression was not changedin the treated tissues. On the other hand doublecortin, aprotein produced in newly-formed neurons was substantiallydecreased by around 50% (po0.05, Fig. 1D). Expression of themorphogenetic protein sonic hedgehog (Shh) was alsodecreased very significantly by 30% (p¼o 0.001, Fig. 1E).
The postsynaptic surface contact protein EphA4 wasdecreased by a relatively small but significant amount (10%,po0.05, not shown) while the putatively schizophrenia-related protein Disrupted in schizophrenia-1 (DISC1) wasraised by over 20% (po0.01, Fig. 1F)
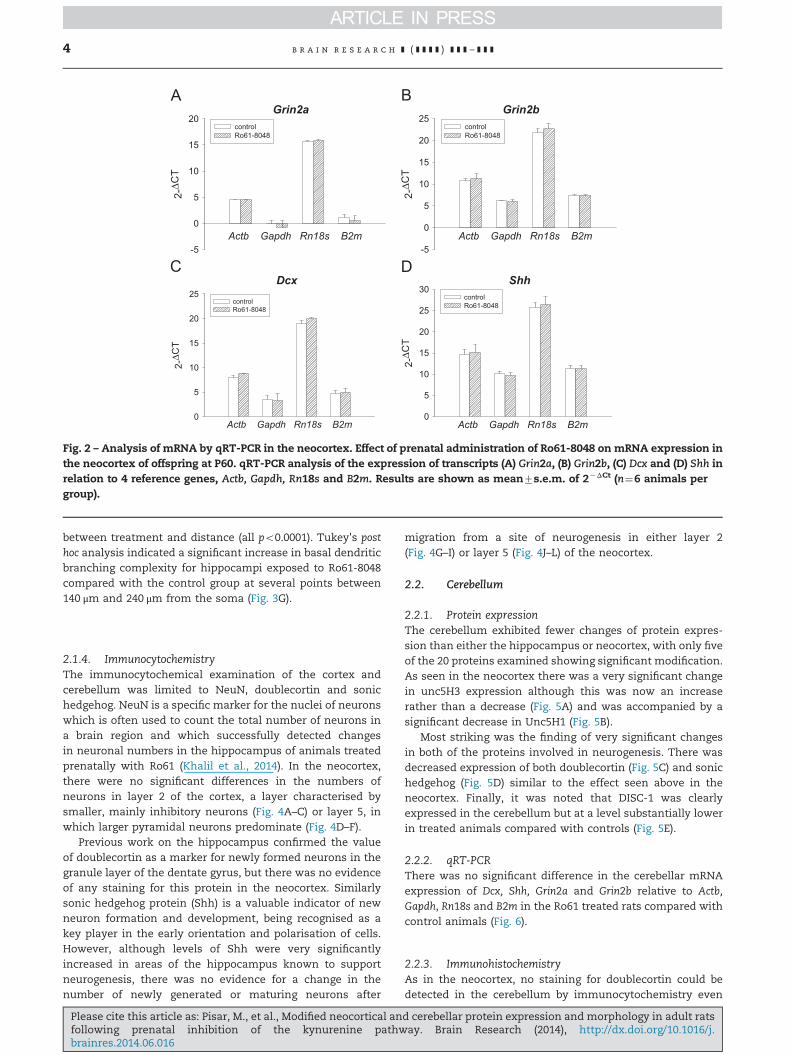
2.1.2. Quantitative real-time polymerase chain reaction (qRT-PCR)mRNA expression of Dcx, Shh, Grin2a and Grin2b transcriptsdid not change relative to Actb, Gapdh, Rn18s and B2m in theneocortex of Ro61 treated offspring compared with controlanimals (Fig. 2).
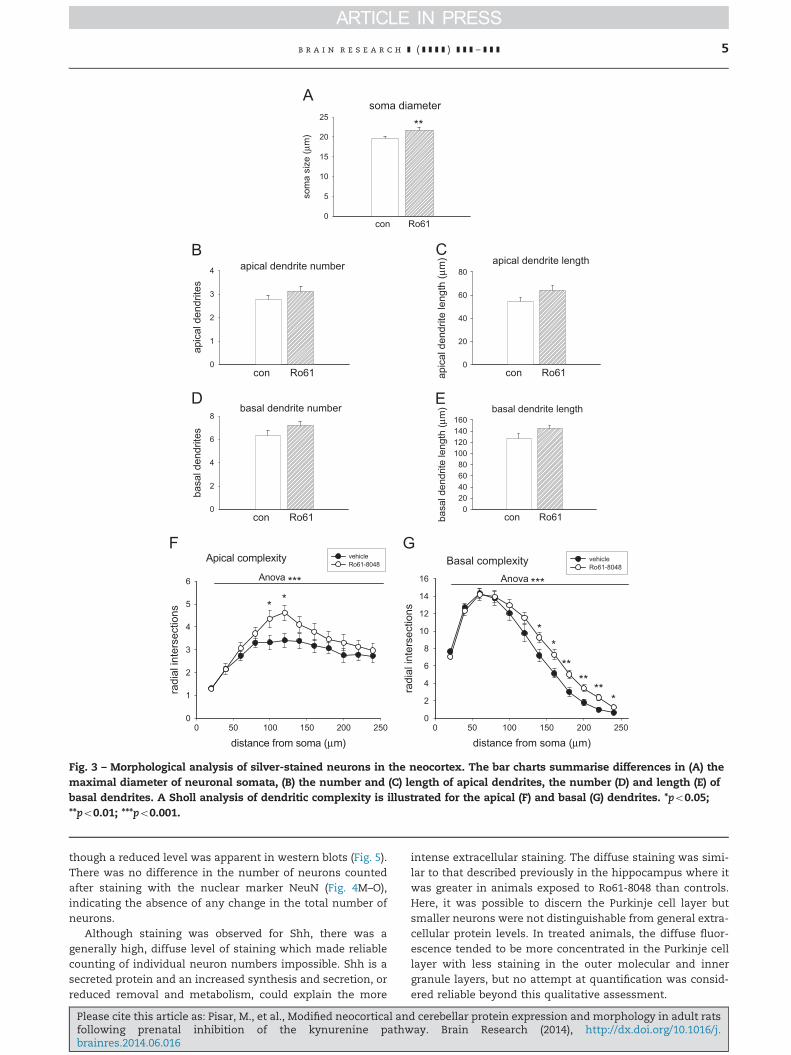
2.1.3. Dendrite and spine analysisThere was a small but highly significant difference of approxi-mately 10% in the maximum somatic diameters of theneurons examined (p¼0.006; Fig. 3A). When measurements
d cerebellar protein expression and morphology in adult ratsay. Brain Research (2014), http://dx.doi.org/10.1016/j.
optic
al d
ensi
typr
otei
n / a
ctin
0.0
0.5
1.0
1.5
saline (S) Ro61 (R)
**
optic
al d
ensi
typr
otei
n / a
ctin
0.0
0.5
1.0
1.5
saline (S) Ro61 (R)
**
Unc5H3
actin
actin
actin
actin
DISC-1
optic
al d
ensi
typr
otei
n / a
ctin
0.0
0.5
1.0
saline (S) Ro61 (R)
*
doublecortin
optic
al d
ensi
typr
otei
n / a
ctin
0.0
0.5
1.0
saline (S) Ro61 (R)
**
GluN2A
optic
al d
ensi
typr
otei
n / a
ctin
0.0
0.5
1.0
saline (S) Ro61 (R)
***actin
Shh
optic
al d
ensi
typr
otei
n / a
ctin
0.0
0.5
1.0
saline (S) Ro61 (R)
***
COX-2
actin
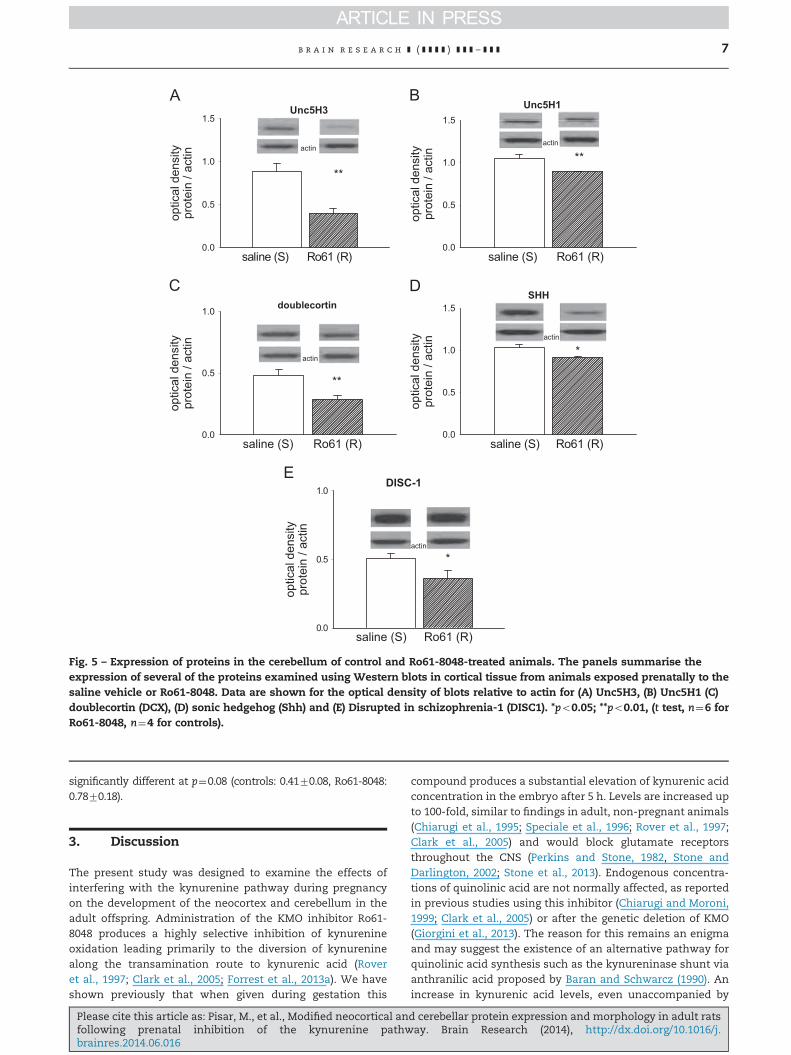
Fig. 1 – Expression of proteins in the neocortex of control and Ro61-8048-treated animals. The panels summarise theexpression of several of the proteins examined in this study using Western blots in cortical tissue from animals exposedprenatally to the saline vehicle or Ro61-8048. Data are shown for the optical density of blots relative to actin for (A) Unc5H3,(B) GluN2A, (C) cyclo-oxgenase-2 (COX-2), (D) doublecortin (DCX), (E) sonic hedgehog (Shh) and (F) Disrupted in schizophrenia-1(DISC1). *po0.05; **po0.01, po0.001 (t test, n¼6 for Ro61-8048, n¼4 for controls).
b r a i n r e s e a r c h ] ( ] ] ] ] ) ] ] ] – ] ] ] 3
were made of the number and length of dendrites in the apicaland basal dendritic trees, no significant differences were noted(Fig. 3B–E). In contrast, the analysis of dendritic complexity(Sholl, 1953), which quantifies the total number of intersec-tions between dendritic elements and a series of virtualconcentric circles centred on the neuronal soma, revealed verysignificant effects.
The complexity counts were compared using a multi-factorial analysis of variance employing a 2�12�22 ANOVA,taking into account the elements of treatment, number of
Please cite this article as: Pisar, M., et al., Modified neocortical anfollowing prenatal inhibition of the kynurenine pathwbrainres.2014.06.016
intersection levels and number of samples included in theanalysis. The analysis revealed a main effect of treatment,distance and sample (all po0.001, n¼11) and an interactionbetween treatment and distance (po0.05) for the apicaldendritic tree (Fig. 3F) with a net increase in complexityand significant differences between individual points at100–120 mm from the cell somata (Fig. 3F).
Analysing the dendritic branching complexity of the basaldendrites also revealed a main effect of treatment, distanceand sample but now also with a highly significant interaction
d cerebellar protein expression and morphology in adult ratsay. Brain Research (2014), http://dx.doi.org/10.1016/j.
Grin2a
Actb Gapdh Rn18s B2m
2-ΔC
T
-5
0
5
10
15
20controlRo61-8048
Grin2b
Actb Gapdh Rn18s B2m
2-ΔC
T
-5
0
5
10
15
20
25controlRo61-8048
Shh
Actb Gapdh Rn18s B2m
2-Δ C
T0
5
10
15
20
25
30controlRo61-8048
Dcx
Actb Gapdh Rn18s B2m
2-Δ C
T
0
5
10
15
20
25controlRo61-8048
Fig. 2 – Analysis of mRNA by qRT-PCR in the neocortex. Effect of prenatal administration of Ro61-8048 on mRNA expression inthe neocortex of offspring at P60. qRT-PCR analysis of the expression of transcripts (A) Grin2a, (B) Grin2b, (C) Dcx and (D) Shh inrelation to 4 reference genes, Actb, Gapdh, Rn18s and B2m. Results are shown as mean7s.e.m. of 2�ΔCt (n¼6 animals pergroup).
b r a i n r e s e a r c h ] ( ] ] ] ] ) ] ] ] – ] ] ]4
between treatment and distance (all po0.0001). Tukey's posthoc analysis indicated a significant increase in basal dendriticbranching complexity for hippocampi exposed to Ro61-8048compared with the control group at several points between140 μm and 240 μm from the soma (Fig. 3G).
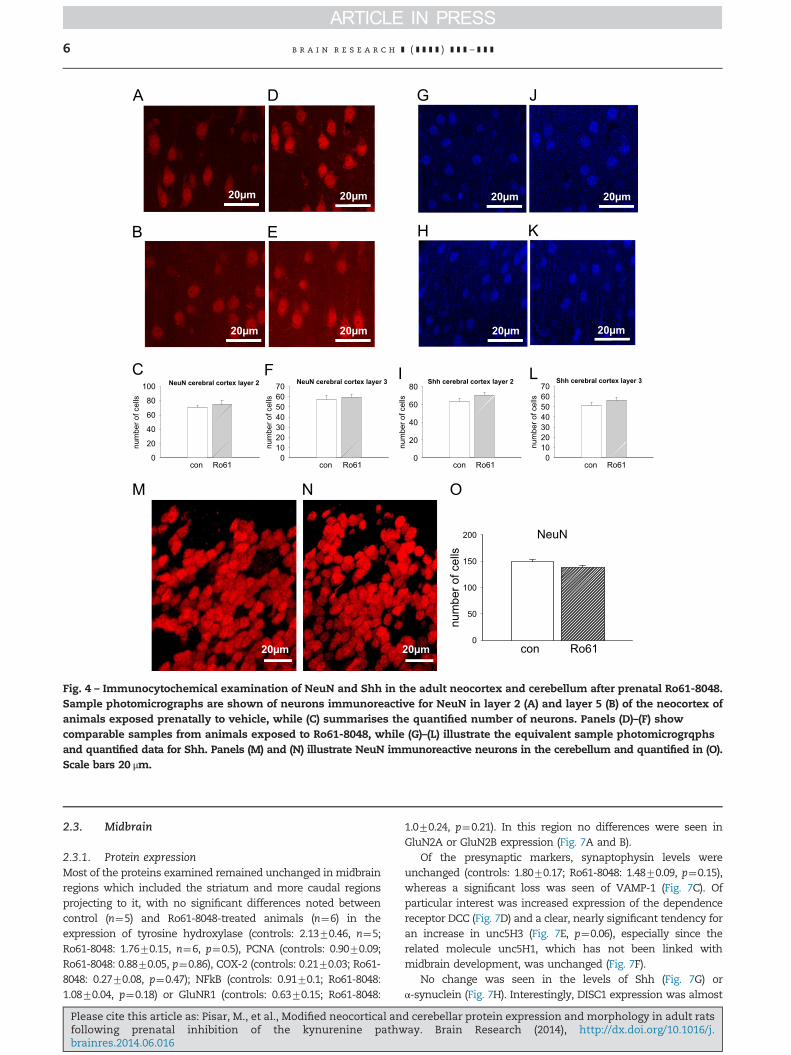
2.1.4. ImmunocytochemistryThe immunocytochemical examination of the cortex andcerebellum was limited to NeuN, doublecortin and sonichedgehog. NeuN is a specific marker for the nuclei of neuronswhich is often used to count the total number of neurons ina brain region and which successfully detected changesin neuronal numbers in the hippocampus of animals treatedprenatally with Ro61 (Khalil et al., 2014). In the neocortex,there were no significant differences in the numbers ofneurons in layer 2 of the cortex, a layer characterised bysmaller, mainly inhibitory neurons (Fig. 4A–C) or layer 5, inwhich larger pyramidal neurons predominate (Fig. 4D–F).
Previous work on the hippocampus confirmed the valueof doublecortin as a marker for newly formed neurons in thegranule layer of the dentate gyrus, but there was no evidenceof any staining for this protein in the neocortex. Similarlysonic hedgehog protein (Shh) is a valuable indicator of newneuron formation and development, being recognised as akey player in the early orientation and polarisation of cells.However, although levels of Shh were very significantlyincreased in areas of the hippocampus known to supportneurogenesis, there was no evidence for a change in thenumber of newly generated or maturing neurons after
Please cite this article as: Pisar, M., et al., Modified neocortical anfollowing prenatal inhibition of the kynurenine pathwbrainres.2014.06.016
migration from a site of neurogenesis in either layer 2(Fig. 4G–I) or layer 5 (Fig. 4J–L) of the neocortex.
2.2. Cerebellum
2.2.1. Protein expressionThe cerebellum exhibited fewer changes of protein expres-sion than either the hippocampus or neocortex, with only fiveof the 20 proteins examined showing significant modification.As seen in the neocortex there was a very significant changein unc5H3 expression although this was now an increaserather than a decrease (Fig. 5A) and was accompanied by asignificant decrease in Unc5H1 (Fig. 5B).
Most striking was the finding of very significant changesin both of the proteins involved in neurogenesis. There wasdecreased expression of both doublecortin (Fig. 5C) and sonichedgehog (Fig. 5D) similar to the effect seen above in theneocortex. Finally, it was noted that DISC-1 was clearlyexpressed in the cerebellum but at a level substantially lowerin treated animals compared with controls (Fig. 5E).
2.2.2. qRT-PCRThere was no significant difference in the cerebellar mRNAexpression of Dcx, Shh, Grin2a and Grin2b relative to Actb,Gapdh, Rn18s and B2m in the Ro61 treated rats compared withcontrol animals (Fig. 6).
2.2.3. ImmunohistochemistryAs in the neocortex, no staining for doublecortin could bedetected in the cerebellum by immunocytochemistry even
d cerebellar protein expression and morphology in adult ratsay. Brain Research (2014), http://dx.doi.org/10.1016/j.
con Ro61
som
a si
ze (μ
m)
0
5
10
15
20
25soma diameter
**
con Ro61
apic
al d
endr
ites
0
1
2
3
4 apical dendrite number
con Ro61 apic
al d
endr
ite le
ngth
(μm
)
0
20
40
60
80apical dendrite length
con Ro61
basa
l den
drite
s
0
2
4
6
8basal dendrite number
con Ro61 basa
l den
drite
leng
th (μ
m)
020406080
100120140160
basal dendrite length
Apical complexity
distance from soma (μm)0 50 100 150 200 250
radi
al in
ters
ectio
ns
0
1
2
3
4
5
6
vehicleRo61-8048
***Anova
* *
Basal complexity
distance from soma (μm)0 50 100 150 200 250
radi
al in
ters
ectio
ns
0
2
4
6
8
10
12
14
16
vehicleRo61-8048
***
**
**
***Anova
**
Fig. 3 – Morphological analysis of silver-stained neurons in the neocortex. The bar charts summarise differences in (A) themaximal diameter of neuronal somata, (B) the number and (C) length of apical dendrites, the number (D) and length (E) ofbasal dendrites. A Sholl analysis of dendritic complexity is illustrated for the apical (F) and basal (G) dendrites. *po0.05;**po0.01; ***po0.001.
b r a i n r e s e a r c h ] ( ] ] ] ] ) ] ] ] – ] ] ] 5
though a reduced level was apparent in western blots (Fig. 5).There was no difference in the number of neurons countedafter staining with the nuclear marker NeuN (Fig. 4M–O),indicating the absence of any change in the total number ofneurons.
Although staining was observed for Shh, there was agenerally high, diffuse level of staining which made reliablecounting of individual neuron numbers impossible. Shh is asecreted protein and an increased synthesis and secretion, orreduced removal and metabolism, could explain the more
Please cite this article as: Pisar, M., et al., Modified neocortical anfollowing prenatal inhibition of the kynurenine pathwbrainres.2014.06.016
intense extracellular staining. The diffuse staining was simi-lar to that described previously in the hippocampus where itwas greater in animals exposed to Ro61-8048 than controls.Here, it was possible to discern the Purkinje cell layer butsmaller neurons were not distinguishable from general extra-cellular protein levels. In treated animals, the diffuse fluor-escence tended to be more concentrated in the Purkinje celllayer with less staining in the outer molecular and innergranule layers, but no attempt at quantification was consid-ered reliable beyond this qualitative assessment.
d cerebellar protein expression and morphology in adult ratsay. Brain Research (2014), http://dx.doi.org/10.1016/j.
num
ber o
f cel
ls
0
50
100
150
200
con Ro61
NeuN
num
ber o
f cel
ls
010203040506070 NeuN cerebral cortex layer 3
num
ber o
f cel
ls
0
20
40
60
80
100 NeuN cerebral cortex layer 2
20µm20µm
20µm
num
ber o
f cel
ls
0
20
40
60
80Shh cerebral cortex layer 2
con Ro61 con Ro61 con Ro61 con Ro61
num
ber o
f cel
ls
010203040506070
Shh cerebral cortex layer 3
20µm
20µm
20µm
20µm
20µm20µm
20µm
Fig. 4 – Immunocytochemical examination of NeuN and Shh in the adult neocortex and cerebellum after prenatal Ro61-8048.Sample photomicrographs are shown of neurons immunoreactive for NeuN in layer 2 (A) and layer 5 (B) of the neocortex ofanimals exposed prenatally to vehicle, while (C) summarises the quantified number of neurons. Panels (D)–(F) showcomparable samples from animals exposed to Ro61-8048, while (G)–(L) illustrate the equivalent sample photomicrogrqphsand quantified data for Shh. Panels (M) and (N) illustrate NeuN immunoreactive neurons in the cerebellum and quantified in (O).Scale bars 20 μm.
b r a i n r e s e a r c h ] ( ] ] ] ] ) ] ] ] – ] ] ]6
2.3. Midbrain
2.3.1. Protein expressionMost of the proteins examined remained unchanged inmidbrainregions which included the striatum and more caudal regionsprojecting to it, with no significant differences noted betweencontrol (n¼5) and Ro61-8048-treated animals (n¼6) in theexpression of tyrosine hydroxylase (controls: 2.1370.46, n¼5;Ro61-8048: 1.7670.15, n¼6, p¼0.5), PCNA (controls: 0.9070.09;Ro61-8048: 0.8870.05, p¼0.86), COX-2 (controls: 0.2170.03; Ro61-8048: 0.2770.08, p¼0.47); NFkB (controls: 0.9170.1; Ro61-8048:1.0870.04, p¼0.18) or GluNR1 (controls: 0.6370.15; Ro61-8048:
Please cite this article as: Pisar, M., et al., Modified neocortical anfollowing prenatal inhibition of the kynurenine pathwbrainres.2014.06.016
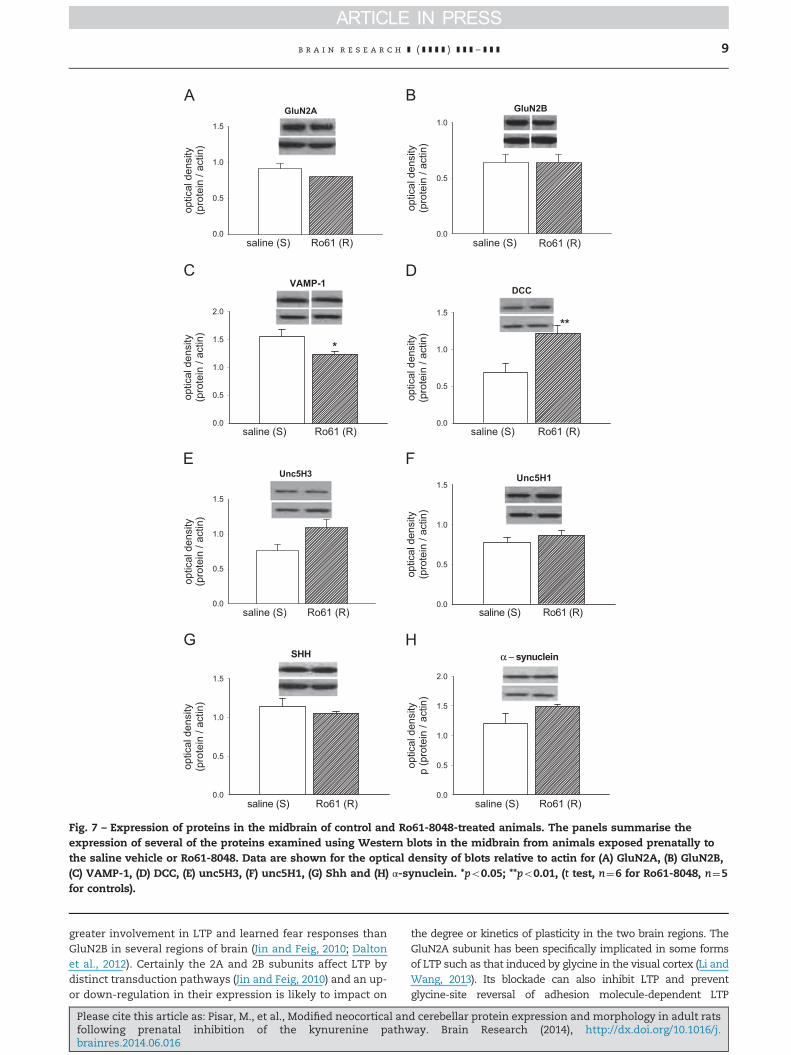
1.070.24, p¼0.21). In this region no differences were seen inGluN2A or GluN2B expression (Fig. 7A and B).
Of the presynaptic markers, synaptophysin levels wereunchanged (controls: 1.8070.17; Ro61-8048: 1.4870.09, p¼0.15),whereas a significant loss was seen of VAMP-1 (Fig. 7C). Ofparticular interest was increased expression of the dependencereceptor DCC (Fig. 7D) and a clear, nearly significant tendency foran increase in unc5H3 (Fig. 7E, p¼0.06), especially since therelated molecule unc5H1, which has not been linked withmidbrain development, was unchanged (Fig. 7F).
No change was seen in the levels of Shh (Fig. 7G) orα-synuclein (Fig. 7H). Interestingly, DISC1 expression was almost
d cerebellar protein expression and morphology in adult ratsay. Brain Research (2014), http://dx.doi.org/10.1016/j.
optic
al d
ensi
typr
otei
n / a
ctin
0.0
0.5
1.0
saline (S) Ro61 (R)
doublecortin
optic
al d
ensi
typr
otei
n / a
ctin
0.0
0.5
1.0
1.5
saline (S) Ro61 (R)
Unc5H1
**actin
actin
**
optic
al d
ensi
typr
otei
n / a
ctin
0.0
0.5
1.0
saline (S) Ro61 (R)
DISC-1
actin
*
optic
al d
ensi
typr
otei
n / a
ctin
0.0
0.5
1.0
1.5
saline (S) Ro61 (R)
Unc5H3
actin
**
optic
al d
ensi
typr
otei
n / a
ctin
0.0
0.5
1.0
1.5
saline (S) Ro61 (R)
SHH
*actin
Fig. 5 – Expression of proteins in the cerebellum of control and Ro61-8048-treated animals. The panels summarise theexpression of several of the proteins examined using Western blots in cortical tissue from animals exposed prenatally to thesaline vehicle or Ro61-8048. Data are shown for the optical density of blots relative to actin for (A) Unc5H3, (B) Unc5H1 (C)doublecortin (DCX), (D) sonic hedgehog (Shh) and (E) Disrupted in schizophrenia-1 (DISC1). *po0.05; **po0.01, (t test, n¼6 forRo61-8048, n¼4 for controls).
b r a i n r e s e a r c h ] ( ] ] ] ] ) ] ] ] – ] ] ] 7
significantly different at p¼0.08 (controls: 0.4170.08, Ro61-8048:0.7870.18).
3. Discussion
The present study was designed to examine the effects ofinterfering with the kynurenine pathway during pregnancyon the development of the neocortex and cerebellum in theadult offspring. Administration of the KMO inhibitor Ro61-8048 produces a highly selective inhibition of kynurenineoxidation leading primarily to the diversion of kynureninealong the transamination route to kynurenic acid (Roveret al., 1997; Clark et al., 2005; Forrest et al., 2013a). We haveshown previously that when given during gestation this
Please cite this article as: Pisar, M., et al., Modified neocortical anfollowing prenatal inhibition of the kynurenine pathwbrainres.2014.06.016
compound produces a substantial elevation of kynurenic acidconcentration in the embryo after 5 h. Levels are increased upto 100-fold, similar to findings in adult, non-pregnant animals(Chiarugi et al., 1995; Speciale et al., 1996; Rover et al., 1997;Clark et al., 2005) and would block glutamate receptorsthroughout the CNS (Perkins and Stone, 1982, Stone andDarlington, 2002; Stone et al., 2013). Endogenous concentra-tions of quinolinic acid are not normally affected, as reportedin previous studies using this inhibitor (Chiarugi and Moroni,1999; Clark et al., 2005) or after the genetic deletion of KMO(Giorgini et al., 2013). The reason for this remains an enigmaand may suggest the existence of an alternative pathway forquinolinic acid synthesis such as the kynureninase shunt viaanthranilic acid proposed by Baran and Schwarcz (1990). Anincrease in kynurenic acid levels, even unaccompanied by
d cerebellar protein expression and morphology in adult ratsay. Brain Research (2014), http://dx.doi.org/10.1016/j.

Grin2a
Actb Gapdh Rn18s B2m
2-C
T
0
5
10
15
20
25controlRo61-8048
Grin2b
Actb Gapdh Rn18s B2m
2-C
T
-5
0
5
10
15controlRo61-8048
Dcx
Actb Gapdh Rn18s B2m
2-C
T
0
5
10
15
20
25controlRo61-8048
Shh
Actb Gapdh Rn18s B2m
2-C
T
0
5
10
15
20
25controlRo61-8048
Fig. 6 – Analysis of mRNA by qRT-PCR in the cerebellum. Effect of prenatal administration of Ro61-8048 on mRNA expressionin the cerebellum of offspring at P60. qRT-PCR analysis of the expression of transcripts (A) Grin2a, (B) Grin2b, (C) Dcx and (D) Shhin relation to 4 reference genes, Actb, Gapdh, Rn18s and B2m. Results are shown as mean7s.e.m. of 2�ΔCt (n¼6 animals pergroup).
b r a i n r e s e a r c h ] ( ] ] ] ] ) ] ] ] – ] ] ]8
any change in quinolinic acid concentration, would still shiftthe overall neuronal effect of the kynurenic acid:quinolinicacid ratio away from neurotoxicity and towards that ofneuroprotection.
The only region of CNS examined so far in detail, however,has been the hippocampus, in which electrophysiologicalresponses, protein expression and localisation, dendriticarchitecture and spine properties were shown to be affectedinto adulthood of the offspring. Since NMDAR are importantplayers in the regulation of many aspects of neuronal andsynaptic development (Sin et al., 2002; P.Y. Wang et al., 2011;Kwon and Sabatini, 2011; Ultanir et al., 2007), the blockade ofthese receptors in the hippocampus is likely to explain manyif not all of these changes (Forrest et al., 2013a, 2013b), as theymay do following immune system activation of TLR4 recep-tors (Forrest et al., 2012; Khalil et al., 2013). There remainsthe important question of whether the changes seen inthe hippocampus are unique, whether they are representa-tive of the CNS as a whole, or whether they reflect changesthat vary between the different brain regions. The presentobjective was to compare changes in the neocortex andcerebellum with those already described in the hippocampus.
3.1. Protein expression
The changes in protein expression found here after prenatalRo61-8048 are different between the neocortex and cerebel-lum and from those seen in the hippocampus. One of theoutstanding similarities between cortex and hippocampus is
Please cite this article as: Pisar, M., et al., Modified neocortical anfollowing prenatal inhibition of the kynurenine pathwbrainres.2014.06.016
the altered expression of GluN2A, since this was one of theearliest changes noted in the embryo and at the stageof weaning the offspring at P21. The direction of change,however, is opposite in the adult neocortex – an increasedexpression compared with a decrease in hippocampus. Adetailed analysis of the results of this difference wouldrequire careful behavioural testing based on the physiologicaland pathological roles of the GluN2A subunits (Wyllie et al.,2013).
Normally, there is a well-recognised switch betweenprimarily GluN2B-containing NMDA receptors in the earlieststages of neuronal formation and the appearance of largelyGluN2A-containing receptors in later stages of development(see Gray et al. (2011) and Matta et al. (2011)). Reduced expres-sion of GluN2A subunits increases the efficacy of newly-formedsynapses. The differential expression of GluN2A found heremay therefore imply an accelerated development of corticalregions but retarded formation of hippocampal circuits. Thisview could have significant implications for neuronal structureand function in these regions since GluN2A subunits regulateaspects of neuronal dendrite formation (Henle et al., 2012)which are distinct from the effects of GluN2B subunits(Sepulveda et al., 2010).
These differences could in turn affect major behaviouralroles of the cortex and hippocampus, such as their involve-ment in learning and memory. Although the respective rolesof GluN2A and GluN2B subunit-containing receptors in long-term potentiation (LTP) continue to be unclear (Wyllie et al.,2013), there is an evidence that GluN2A subunits have a
d cerebellar protein expression and morphology in adult ratsay. Brain Research (2014), http://dx.doi.org/10.1016/j.
0.0
0.5
1.0
GluN2B
optic
al d
ensi
ty(p
rote
in /
actin
)
optic
al d
ensi
ty(p
rote
in /
actin
)
0.0
0.5
1.0
1.5
GluN2A
optic
al d
ensi
ty(p
rote
in /
actin
)
0.0
0.5
1.0
1.5
SHH
optic
al d
ensi
typ
(pro
tein
/ ac
tin)
0.0
0.5
1.0
1.5
2.0
α − synuclein
optic
al d
ensi
ty(p
rote
in /
actin
)
0.0
0.5
1.0
1.5Unc5H1
0.0
0.5
1.0
1.5
Unc5H3
optic
al d
ensi
ty(p
rote
in /
actin
)
optic
al d
ensi
ty(p
rote
in /
actin
)0.0
0.5
1.0
1.5
DCC
**
optic
al d
ensi
ty(p
rote
in /
actin
)
0.0
0.5
1.0
1.5
2.0
saline (S) Ro61 (R)saline (S) Ro61 (R)
saline (S) Ro61 (R) saline (S) Ro61 (R)
saline (S) Ro61 (R)saline (S) Ro61 (R)
saline (S) Ro61 (R)saline (S) Ro61 (R)
VAMP-1
*
Fig. 7 – Expression of proteins in the midbrain of control and Ro61-8048-treated animals. The panels summarise theexpression of several of the proteins examined using Western blots in the midbrain from animals exposed prenatally tothe saline vehicle or Ro61-8048. Data are shown for the optical density of blots relative to actin for (A) GluN2A, (B) GluN2B,(C) VAMP-1, (D) DCC, (E) unc5H3, (F) unc5H1, (G) Shh and (H) α-synuclein. *po0.05; **po0.01, (t test, n¼6 for Ro61-8048, n¼5for controls).
b r a i n r e s e a r c h ] ( ] ] ] ] ) ] ] ] – ] ] ] 9
greater involvement in LTP and learned fear responses thanGluN2B in several regions of brain (Jin and Feig, 2010; Daltonet al., 2012). Certainly the 2A and 2B subunits affect LTP bydistinct transduction pathways (Jin and Feig, 2010) and an up-or down-regulation in their expression is likely to impact on
Please cite this article as: Pisar, M., et al., Modified neocortical anfollowing prenatal inhibition of the kynurenine pathwbrainres.2014.06.016
the degree or kinetics of plasticity in the two brain regions. TheGluN2A subunit has been specifically implicated in some formsof LTP such as that induced by glycine in the visual cortex (Li andWang, 2013). Its blockade can also inhibit LTP and preventglycine-site reversal of adhesion molecule-dependent LTP
d cerebellar protein expression and morphology in adult ratsay. Brain Research (2014), http://dx.doi.org/10.1016/j.

b r a i n r e s e a r c h ] ( ] ] ] ] ) ] ] ] – ] ] ]10
(Kochlamazashvili et al., 2012). However, it seems not to beneeded for LTP in organotypic slices of hippocampus, where itmay actually suppress LTP via its cytoplasmic tail (Foster et al.,2010). GluN2A subunits play a positive role in earlier phases oflearning mediated in the rat forebrain (Jung and Suh, 2010). Onthe other hand, increased expression in cortex has been shownto adversely affect aspects of recognition and olfactory memory(Jacobs and Tsien, 2014) and to modulate fear memory inprefrontal cortex (Gilmartin et al., 2013). A loss of GluN2Asubunits generally produces antidepressant and anxiolytic beha-viours (Inta et al., 2013).
When NMDAR are blocked pharmacologically in neonatalrats, increased neuronal apoptosis results in both the pre-frontal cortex and hippocampus. These effects are associatedwith deficits in learning and memory (Owczarek et al., 2011)and are paralleled by increased expression of both GluN2Aand GluN2B subunits in adulthood. Both GluN2A and GluN2Bsubunits seem to be involved in excitotoxicity (Lujan et al.,2012; Zhou et al., 2013), although GluN2A subunit-containingreceptors tend to be neuroprotective (Xiao et al., 2010) andcan inhibit the phosphorylation of tau proteins in the hippo-campus (de Montigny et al., 2013). The greater expression ofGluN2A in cortex may therefore imply a greater resistance toneurodegenerative insults, while the hippocampus may bemore susceptible.
Of great interest is the substantial and highly significantincrease in expression of the dependence receptor Unc5H3 inneocortex and decrease in cerebellum. The special interest inthis observation stems from the fact that Un5H3 is a criticalfactor in cerebellar development, with a deficiency in Unc5H3being associated with gross abnormalities of cerebellar mor-phology and motor control (Kuramoto et al., 2004). Thisprobably arises from the role of Unc5 proteins in balancingattraction and repulsion of growing axons, dendrites andpostsynaptic targets (Srinivasan et al., 2012).
In the midbrain, although expression of most of theproteins examined were unaffected at P60 by the prenatalexposure to Ro61-8048, it was of great interest that expres-sion of unc5H3 almost increased to significance, although itsclose relative unc5H1 was unaffected. Since both DCC andunc5H3 have been linked to the development and maturationof dopaminergic projections and the efficacy of dopaminergictransmission in adult animals (Osborne et al., 2005; Manittet al., 2011), these changes may carry implications fordisorders involving aberrant dopaminergic neurotransmis-sion, consistent with the changes noted in the expressionof DISC1.
Levels of DISC1 were increased in the neocortex anddecreased in cerebellum. Both regions have been suggestedto have an involvement in schizophrenia, and there is asubstantial body of evidence linking increased levels of kynure-nic acid in the brain with symptoms of schizophrenia, both inhumans and animal models (Schwarcz et al., 2001; Erhardtet al., 2009; Sathyasaikumar et al., 2011; Linderholm et al., 2012;reviewed in Stone and Darlington, 2013). It is possible, there-fore, that the effects of chronically increased levels of kynure-nic acid (produced here by Ro61-8048) could alter theexpression of DISC1 as part of the mechanism of theirinvolvement in schizophrenia. It should be noted that thereare some reports of lowered kynurenic acid concentrations in
Please cite this article as: Pisar, M., et al., Modified neocortical anfollowing prenatal inhibition of the kynurenine pathwbrainres.2014.06.016
schizophrenic patients (Myint et al., 2011) and in a socialisolation model of schizophrenia in rats (Moller et al., 2012).The kynurenine analyses in those works, however, were madeusing plasma rather than CSF or brain tissue, and may not bereflecting changes in CNS function. The social isolation modelmay also be less appropriate than Ro61-8048 treatment as amodel of schizophrenia, in that raised levels of quinolinic acidwere observed in plasma although comparable findings havenot been seen in patients with schizophrenia (Schwarcz et al.,1988). Stress in prenatal and neonatal life can also inducebehavioural features of depression and anxiety (Sun et al.,2013a, 2013b) that could complicate any attempt to interpretthe results in terms of schizophrenia alone
Two proteins were quantified as indicators of inflamma-tion in the CNS, since that might have an influence on severalof the parameters examined here. However, there was nochange in the pro-inflammatory transcription factor NuclearFactor-kappa-B (NFkB) in neocortex or cerebellum and adecreased expression of the inducible cyclo-oxygenase-2(COX2) in neocortex, suggesting the absence of any localisedinflammation.
Most striking of all was the marked decline in expression ofthe two proteins involved in early post-mitotic developmentand maturation: doublecortin and sonic hedgehog. The formeris found primarily in newly formed, undifferentiated cells,while Shh is encountered mainly post-differentiation as cellsare becoming polarised and oriented into region-specificarrangements of cell types and layers. Expression of both ofthese proteins was readily detected in cortex and hippocam-pus, suggesting that a degree of cell generation is continuingeven at this young adult (P60) age point, or that neuronsgenerated elsewhere are still maturing after migrating fromtheir site of origin. Since the major neurochemical effect ofRo61-8048 is to raise levels of kynurenic acid, the decreasedexpression of these proteins would be entirely consistent withthe ability of selective NMDA antagonists to produce a loss ofneurons in the developing CNS (Ikonomidou et al., 1999;Dikranian et al., 2001; Harris et al., 2003; Vincent et al., 2004).
3.2. Spine density and neuronal morphology
The number, length and complexity of dendrites is widelyconsidered to reflect the level of neuronal interaction andnetwork integration corresponding to cognitive function.Even minor changes in the size and thickness of dendriteshave a significant influence on synaptic efficacy (Ferranteet al., 2013). This is especially so in the neocortex, where ahigh proportion of cognitive behaviour is organised andinitiated. The prenatal administration of Ro61-8048 has beenshown to alter dendritic parameters in the hippocampus, butmost of the simpler measures of dendritic morphology in theneocortex were not significantly affected by this treatment.Thus, neither the numbers nor the total lengths of dendriteswere altered in treated animals. This applied to both apicaland basal trees even though they are believed to acceptdifferent types of synaptic input with a different balance ofexcitation and inhibition, neurotransmitter profiles (Choet al., 2008; Leung and Peloquin, 2010), electrical propertiesand plasticity of synaptic connections (Sajikumar and Korte,2011; Gordon et al., 2006). However, despite the lack of
d cerebellar protein expression and morphology in adult ratsay. Brain Research (2014), http://dx.doi.org/10.1016/j.

b r a i n r e s e a r c h ] ( ] ] ] ] ) ] ] ] – ] ] ] 11
changes in individual aspects of the dendritic tree, ananalysis of overall dendritic complexity (Sholl, 1953) whichassesses the number of dendritic intersections with progres-sively distant targets, irrespective of location, revealed sig-nificant differences. In both the apical and basal dendritictrees of neocortical layer 3 pyramidal neurons there weresignificantly increased degrees of complexity. These wereseen closer to the soma on apical dendrites (100–120 μm)compared with the basal tree (over 180–240 μm distant) andmight reflect a difference balance in the ratio of excitatoryand inhibitory inputs to the cells examined since differenttypes of synapse are known to occur at different regions ofthe dendritic network.
3.3. Protein localisation
As a marker of neuronal nuclei, NeuN has become usedwidely to facilitate the counting of neurons rather than othercell types found in the CNS but in this work it revealed nochange in the total number of neurons occurring in theneocortex or cerebellum.
As a microtubule-associated protein, doublecortin isimportant in neuronal migration. It is particularly involvedin the migration of inhibitory neurons (Friocourt et al., 2007;Xiong et al., 2008; Cai et al., 2009), leading to the expectationthat it would reveal changes of total neuronal number in thecerebellum where the majority of neurons are inhibitory. Thefailure to observe any staining for doublecortin thereforesuggests a high degree of resistance to change in cerebellardevelopment. This would not be surprising given the funda-mentally important role of this organ for the survival ofneonatal animals immediately after parturition, where anysmall influences of diet or environment on neuronal functioncould have serious consequences. The absence of change,however, is entirely consistent with the absence of change inNeuN labelling, since doublecortin expression coincides withearly phases of cell maturation after formation (Couillard-Despres et al., 2005; Jin et al., 2010; Spampanato et al., 2012).It is also consistent with the absence of obvious changes inShh immunoreactivity given that this protein is also mostprominent in the early stages of cellular generation.
It is perhaps surprising that more changes were notobserved in cell number or maturation state since therewas a clear and highly significant increase in the expressionof the GluN2A subunit. It has been noted previously that theprenatal exposure of rats to Ro61-8048 produces very sig-nificant changes in the GluN2A or GluN2B subunits as early as5 h after administration, and changes are still evident in thepostnatal brains or the hippocampi of adult offspring. Sincemanipulation of NMDA receptor activity directly, using selec-tive agonists or antagonists, can influence neuron numbersand brain development (Ikonomidou et al., 1999; Harris et al.,2003; C.-C. Wang et al., 2011) it would be expected thatmaintained changes in subunit expression would be asso-ciated with altered neuron numbers and connectivity. Thatthis is not so in the neocortex and cerebellum suggests thatadditional factors may be required to generate changes ofneuronal production and maturation.
The glycoprotein sonic hedgehog (Shh) is involved in cellcycle control (Alvarez-Medina et al., 2009), cellular orientation
Please cite this article as: Pisar, M., et al., Modified neocortical anfollowing prenatal inhibition of the kynurenine pathwbrainres.2014.06.016
and tissue polarisation (Jessell, 2000; Palma et al., 2005;Traiffort et al., 2010), cell proliferation and migration(Traiffort et al., 2001; Charytoniuk et al., 2002; Palma et al.,2005; Balordi and Fishell, 2007). Despite its early importancein cellular development, Shh persists into adulthood inseveral regions of the CNS including the neocortex andcerebellum (Traiffort et al., 1998; Charytoniuk et al., 2002). Itis the only protein examined to date that has been signifi-cantly affected in all brain regions examined, suggesting thatit is particularly susceptible to the influence of kynureninepathway activity or the changes in glutamate receptor activa-tion that the pathway regulates. Unfortunately the amount ofextracellular secreted protein prevented the quantification ofimmunoreactive cell numbers, preventing visual confirma-tion of the altered protein expression as had been possible inthe hippocampus (Forrest et al., 2013a, 2013b)
Despite this limitation, the change in Shh protein maymake a significant contribution to the changes in dendriticcomplexity noted above, since it influences the balance ofsynaptic attraction and repulsion at dendritic synapses dur-ing development (Hor and Tang, 2010; Angot et al., 2008).Certainly, since Shh expression has been associated particu-larly with inhibitory neurons such as the cerebellar granulecells (Spassky et al., 2008), its decreased expression is con-sistent with the reduced expression of doublecortin.
4. Summary and conclusion
These results support the concept that the kynurenine path-way plays a key role in the early development of the CNS. Thepathway is certainly present in the embryonic brain (Bealet al., 1992; Walker et al., 1999; Schwarcz and Pellicciari, 2002;Guillemin et al., 2001, 2005), with its ability to modifyglutamate receptor activation via the balance between theagonist quinolinic acid and the antagonist kynurenic acid.A primary reason for investigating the role of kynurenines inearly brain development is that elements of the pathway,especially the major enzymes indoleamine-2,3-dioxygenase(IDO) and KMO are induced and activated by infection or pro-inflammatory cytokines. Indeed any immune challenge tothe mother or neonate which results, directly or indirectly, inthe activation of central glia or peripheral macrophages, orchanges in the levels of cytokines or kynurenines in the foetalor neonatal CNS would alter the balance of quinolinic acidand kynurenic acid concentrations and could seriously per-turb neural development and plasticity. This could in turnincrease the risk of CNS disorders, such as schizophrenia,Alzheimer's and Huntington's disease and would be consis-tent with evidence that genetic abnormalities of the kynur-enine pathway are linked to disorders such as schizophrenia(Sathyasaikumar et al., 2011; Holtze et al., 2012; Stone andDarlington, 2013). The blockade of NMDA receptors bykynurenic acid produces neurochemical and behaviouralchanges which have been likened to those seen in schizo-phrenia (Harris et al., 2003; du Bois and Huang, 2007) andrelated to altered levels of kynurenic acid in patients asdiscussed above.
Overall, the present data strongly support the concept thatconstitutive activity of the kynurenine pathway during
d cerebellar protein expression and morphology in adult ratsay. Brain Research (2014), http://dx.doi.org/10.1016/j.

b r a i n r e s e a r c h ] ( ] ] ] ] ) ] ] ] – ] ] ]12
embryogenesis is intimately involved in early brain develop-ment. This conclusion has important implications for theconverse possibility that external factors promoting or sup-pressing that basal activity may interfere with brain devel-opment and, as a result, brain structure and function in theadult. Infection and stress activate the pathway and maternalexposure to these factors during pregnancy has been linkedto the emergence of disorders such as schizophrenia inpostnatal life. The kynurenine pathway could be a majorcontributor to that association.
5. Experimental procedures
Female Wistar rats were mated (at least 3 per treatmentgroup) and then housed separately with access to food andwater ad libitum. All experiments were approved by theUniversity of Glasgow Research Ethical Committee and werefully licensed and conducted according to the Animals(Scientific Procedures) act 1986 of the UK as administeredand monitored by the UK Home Office.
Inhibition of the kynurenine pathway was achieved using3,4-dimethoxy-N-[4-(3-nitrophenyl)thiazol-2-yl]benzenesul-phonamide (Ro61-8048) (Rover et al., 1997) which inhibitskynurenine-3 monoxygenase (KMO) (Stone and Darlington,2002, 2013). The compound was administered 100 mg/kgintra-peritoneally (i.p.) in late gestation on embryonic daysE14, E16 and E18 in order to spread the developmental periodover which the kynurenine pathway would be inhibited.Gestation, parturition and weaning then proceeded normally.Offspring were subsequently euthanized at postnatal day60 (P60) for the removal of brains. Each measurementdescribed has been obtained from an analysis of young fromat least 3 pregnant females used for each treatment, yieldingat least 20 offspring in total for analysis.
5.1. Western immunoblotting
Brain sample homogenates were prepared in RIPA buffer(50mM Tris, 150 mM NaCl, 0.1% SDS, 0.5% Triton X-100, 1%IGEPAL, and a Roche complete protease inhibitor tablet) andcentrifuged at 18,000g for 5 min at 4 1C. Supernatants werecollected for protein concentration determination using theBio-Rad Coomassie Blue protein assay (Bio-Rad, Hemel Hemp-stead, UK). Samples were then normalised to 10 μg and pre-pared as; 65% protein sample, 25% sample buffer and 10%reducing agent (Life Technologies, Paisley, UK) and heated at70 1C for 10 min. The protein samples were loaded ontoNuPAGE Novex 4–12% Bis–Tris (1.0 mm) 15 lane gels (LifeTechnologies, Paisley, UK) and run at 150 V for 80 min toseparate proteins according to their molecular weight. SeeBluepre-stained standard (Life Technologies, Paisley, UK) wasincluded on each gel as a molecular weight marker. Theseparated proteins were then blotted onto Invitrolon poly(vinylidene difluoride) membranes (Life Technologies, Paisley,UK) at 30 V for 60 min. The membranes were blocked for 1 h in5% non-fat dried milk solution in Tris-buffered saline contain-ing 0.05% Tween (TBST) before overnight incubation at 4 1Cwith the appropriate primary antibody (diluted in 5% milk-TBST). Membranes were then washed 3 times for 15 min with
Please cite this article as: Pisar, M., et al., Modified neocortical anfollowing prenatal inhibition of the kynurenine pathwbrainres.2014.06.016
TBST and incubated with the appropriate horseradish perox-idase (HRP) conjugated secondary antibody (prepared in 5%milk-TBST) for 1 h at room temperature. Following secondaryantibody incubation, blots were washed 3 times for 15 minwith TBST then visualised using Enhanced Chemilumines-cence Plus detection kit (GE Healthcare, Chalfont St Giles, UK).
Western blot analysis was carried out using the followingprimary antibodies:
From Millipore, Watford, UK: GluN1 (mouse monoclonal, 05-432, 1:1000 dilution); synaptophysin (mouse monoclonal,MAB368, 1: 40,000 dilution);From R&D Systems, Abingdon, UK: GluN2A (rabbit polyclonal,PPS012, 1:5000 dilution); GluN2B (rabbit polyclonal, PPS013,1:5000); VAMP-1/synaptobrevin (goat polyclonal, AF4828,1:10,000 dilution), synaptotagmin (MAB 43641, 1:5000dilution);From Cell Signalling, New England Biolabs, Hitchin, Herts, UK:PSD-95 (rabbit monoclonal, #3450, 1:10,000 dilution);From Santa Cruz, Insight Biotechnology, Wembley, UK: dou-blecortin (goat polyclonal, sc-8066, 1:1000 dilution); α-synuclein (mouse monoclonal, sc-65500, 1:1000 dilution);actin (goat polyclonal, sc-1615, 1:10,000 dilution); DISC-1(goat polyclonal, sc-47990, 1:1000 dilution); Unc5H1 (goatpolyclonal, sc-67902, 1:1000 dilution), Unc5H3 (goat poly-clonal, sc-54442, 1:1000 dilution), SHH (goat polyclonal, sc-1194, 1:1000 dilution); NFκB (rabbit polyclonal, sc-372,1:5000 dilution); COX-2 (goat polyclonal, sc-1745, 1:1000dilution); RhoA (mouse monoclonal, sc-418, 1:1000 dilu-tion); RhoB (mouse monoclonal, sc-8048, 1:1000 dilution);EphA4 (rabbit polyclonal, sc-921, 1:5000); PCNA (mousemonocloncal, sc-56, 1:1000 dilution);From BD Biosciences, Oxford, UK: DCC (mouse monoclonal,#544223, 1:5000 dilution)
The following secondary HRP-conjugated antibodies wereused at 1:5000 dilution: goat anti-rabbit HRP (12-348) (Milli-pore, Watford, UK); donkey anti-goat HRP (sc-2020) and goatanti-mouse (sc-2005) (Santa Cruz, Insight Biotechnology,Wembley, UK)
All western blots were quantified using the Image J soft-ware (http://rsb.info.nih.gov/ij/) and comparisons were madestatistically between groups of pups born to mothers treatedwith Ro61-8048 and groups born to mothers injected withsaline vehicle. This protocol allowed the use of a t-test toexamine differences between the two groups. To control forvariations in the total amount of protein loaded onto gels allsamples were examined after staining with Ponceau S stain.In addition, actin levels were examined in each series of blotsand the ratio taken of the intensity of target protein to theintensity of actin. A probability value of 0.05 was adopted asthe criterion for significance.
5.2. Quantitative real-time polymerase chain reaction(qRT-PCR)
Total RNA was isolated from snap frozen neocortex andcerebellum from P60 rats (n¼2/litter, n¼3 litters/treatment
d cerebellar protein expression and morphology in adult ratsay. Brain Research (2014), http://dx.doi.org/10.1016/j.

b r a i n r e s e a r c h ] ( ] ] ] ] ) ] ] ] – ] ] ] 13
group, n¼6 control and n¼6 treated) using the Qiagen RNeasylipid midi kit with DNaseI treatment, according to themanufacturer's instructions. The RNA concentration of eachsample was determined using a NanoDrop (Thermo Scienti-fic) and the integrity of each mRNA sample was assessedusing the Agilent 2100 Bioanalyser (Glasgow Polyomics Facil-ity, University of Glasgow). Only samples with RNA integritynumber (RIN) greater than 8 were used. For cortex RIN valuesranged from 8.50 to 9.20, and for cerebellum the values were8.10–9.10.
First strand cDNAs were synthesised from total RNA usingrandom hexamers and Superscript III Reverse transcriptasein the VILO cDNA synthesis kit (Life Technologies) followingthe manufacturer's instructions.
qRT-PCR was carried out in an ABI cycler (SDS7000) using theTaqMan Gene expression assay system for the followingassays: doublecortin (Dcx; Rn00670392_m1), sonic hedge-hog (Shh; Rn00568129_m1), NMDA receptor subunit 2A(Grin2a; Rn00561341_m1) and NMDA receptor subunit 2B (Grin2b;Rn00680474_m1), (Applied Biosystems). In addition, four refer-ence genes were used: beta actin (Actb; Rn00667869_m1), glycer-aldehyde-3-phosphate dehydrogenase (Gapdh; Rn01775763_g1),beta-2 microglobulin (B2m; Rn00560865_m1) and 18S ribosomalRNA (Rn18s; Mm03928990_g1), (Applied Biosystems).
PCR Ct values and reaction efficiencies were obtained fromthe raw fluorescent data using LinReg PCR (Ramakers et al.,2003). TaqMan assay reaction efficiencies were Dcx: cortex1.873, cerebellum 1.850, Shh: cortex 1.800, cerebellum 1.828,Grin2a: cortex 1.867, cerebellum 1.797, Grin2b: cortex 1.805,cerebellum 1.882, Actb: cortex 1.833, cerebellum 1.822, Gapdh:cortex 1.726, cerebellum 1.750, B2m: cortex 1.872, cerebellum1.956 and Rn18s: cortex 1.660, cerebellum 1.714. Samples wererun in triplicate and Ct outliers were determined using theGrubb's test (http://graphpad.com/quickcalcs/Grubbs). Signif-icant Ct outliers (p40.05) were removed before statisticalanalysis. The mean Ct values for each sample were analysedusing REST 2009 v2.0.13 (Pfaffl et al., 2002). REST calculates pvalues on the basis of 2000 permutations of a pair-wise fixedreallocation randomisation test in which Ct values for thereference and target gene are reallocated to sample andcontrol groups and calculates the resulting expression ratioson the basis of the mean values.
REST analysis of relative gene expression confirmed thatfor the reference genes Actb, Gapdh, Rn18s and B2m these werenot significantly different between control and treated rats.qRT-PCR analysis of transcripts for Grin2a, Grin2b, Dcx andShh are therefore expressed relative to each of the fourreference genes.
5.3. Golgi–Cox silver staining
All tissues were taken from offspring at P60 which wereeuthanized by an i.p. injection of sodium pentobarbitone andthe brains removed. Golgi silver staining was performed usingthe FD Rapid GolgiStain Kit (FD Neurotechnologies Inc.), follow-ing the manufacturer's instructions. Briefly, brains wereimmersed in the impregnation solution for 2–3 weeks, pro-tected from light, at room temperature. The solution waschanged after the initial 24 h of immersion and was transferred
Please cite this article as: Pisar, M., et al., Modified neocortical anfollowing prenatal inhibition of the kynurenine pathwbrainres.2014.06.016
to solution C at 4 1C for at least 48 h, the solution again beingreplenished after the first 24 h. Brains were then cut into200 μm coronal slices using a Leica VT1200 series Vibratome(Leica, UK) and sections were mounted on gelatine-coatedslides, dried at room temperature shielded from light. Thedried sections were rinsed twice for 2 min before transferringto a mixture of solutions D and E for 10 min. Sections werethen rinsed again twice for 4 min and progressively dehydratedin ethanol 50%, 75% and 95% (4min each) followed by ethanol100% for four 4 min periods. Sections were cleared in Histoclearthree times for 4 min each and covered by a coverslip usingHistomount (ThermoFisher Scientific).
5.4. Dendritic morphology
An average of 6–8 neurons was examined in each of 3–4coronal sections from each animal, using a Nikon EclipseE400 microscope by an investigator blind to the treatment ofthe animal. Neurons were examined under a 40� objectivelens and copied using a camera lucida system onto a two-dimensional plane measuring 420�297 mm. The criteria forneuron selection were that neurons were clearly identifiablealong their entire length and breadth with a uniform andclearly visible level of silver staining and that each neuroncould be distinguished clearly enough from adjacent impreg-nated neurons to avoid contamination of the measurementprocess by nearby cells. Neurons were selected towards thecentre of sections so as to maximise the probability ofassessing cells that were completely intact.
Morphological data were analysed using an unpaired t-testto assess differences in the number of dendritic branches andtheir length between control and Ro61-8048 treated groups ofanimals. The analysis of dendritic complexity (Sholl, 1953)was subjected to a two-way analysis of variance (ANOVA)followed by Tukey's post hoc analysis to examine the con-tributions of drug treatment, sample number and distancefrom the cell soma. A probability value of 0.05 was taken asthe criterion for significance.
5.5. Immunohistochemistry
Groups of 7–9 animals were used from litters born to damstreated with vehicle or Ro61-8048. They were deeply anaesthe-sized using Euthatal (sodium pentobarbitone) and perfusedtranscardially via the left ventricle with 50ml artificial cere-brospinal fluid (in mM: NaCl 115; NaHCO3 25; KH2PO4 2.2; KCl 2;MgSO4 1.2; CaCl2 2.5; D-glucose 10; gassed with 5% CO2 in O2)followed by approximately 100ml of 4% paraformaldehyde (PFA)in 0.2 M phosphate buffered saline (PBS), pH 7.2. Immediatelyfollowing perfusion, the brains were removed and fixed in thebuffered paraformaldehyde solution for a further 4 h at 4 1C.After thorough rinsing in 0.1 M PBS, the brains were protected ina 30% sucrose solution at 4 1C until saturated. Sections (60 μM)were cut coronally using a vibratome (Leica VT1200, Leica, UK)and collected serially in 0.1 M PBS The sections were incubatedwith 50% ethanol for 30min before being washed 3 times with0.3 M (PBS) and stored in glycerol at �20 1C.
For analyses of neurogenesis, serial free-floating sectionswere first washed in 0.3 M PBS before incubation withprimary antibodies in PBS containing 0.1% Triton (PBST;
d cerebellar protein expression and morphology in adult ratsay. Brain Research (2014), http://dx.doi.org/10.1016/j.

b r a i n r e s e a r c h ] ( ] ] ] ] ) ] ] ] – ] ] ]14
pH 7.4) for 72 h at 4 1C with continuous agitation. The primaryantibodies used were mouse monoclonal anti-NeuN 1:500 (forneuronal nuclei, MAB377 Millipore, Watford, UK), goat poly-clonal anti-doublecortin 1:250 (for doublecortin, sc-8066 SantaCruz via Insight Biotechnology, Wembley, UK) and rabbitpolyclonal anti-Shh 1:100 (for sonic hedgehog, sc-9024 SantaCruz via Insight Biotechnology, Wembley, UK). Followingprimary antibody treatment, sections were incubated over-night at 4 1C in species specific Alexa Fluor-tagged secondaryantibodies (Alexa 488, Alexa 647, Molecular Probes, LifeTechnologies Ltd., Paisley, UK) and rhodamine-conjugatedsecondary antibody (Jackson Immunoresearch LaboratoriesEurope Ltd. Suffolk, UK). Rinses were performed between allsteps using 0.3 M PBS before or after the primary andsecondary antibody incubation. Sections were mounted andcover-slipped with the aqueous mounting anti-fade mediumH-1000:Vectashield (Vector Laboratories Ltd., Peterborough,UK) before storage at �201C.
Neurons were scanned with most parameters similar tothose outlined above but using a 40� oil immersion objectivelens. Each field consisted of a stack of 20 optical sections at anincrement of 1 μm z-separation. Immunopositive stainingwas manually counted in each subfield in an area measuring50 μm�50 μm from the 1024�1024 pixel images. Each chan-nel for Shh, NeuN and doublecortin was analysed separatelyto identify and count manually immunopositive stainingusing ImageJ software. The overall dendrite length ofdoublecortin-labelled granule cells in the dentate gyrus wasmeasured using the ImageJ plug-in, NeuronJ.
Statistical comparisons were made using InStat software,between pups born to mothers injected with vehicle andthose treated with Ro61-8048. This protocol allowed the useof an unpaired t-test to examine differences between the twogroups for each target protein of interest. A probability valueof 0.05 was adopted as the working criterion for significance.
Conflict of interest
The authors declare that they have no conflicts of interest inrelation to the publication of this work.
Acknowledgements
MP is supported by funding from the Malaysian Governmentgrant PGS-MP and OSK is in receipt of a BBSRC, UnitedKingdom Postgraduate Scholarship in Organismal Biologygrant BB/F016700. We are grateful to Prof. A. Todd, Dr. D.I.Hughes, Dr. C. Daly, Dr. S. Cobb and Dr. C. Winchester foradvice and the use of facilities, and to Epsom MedicalResearch (grant EMR-TWS09), the Peacock Trust (grant TWS-K1) and the Haddon Family Trust (grant WH1) for financialassistance.
r e f e r e n c e s
Alvarez, V.A., Ridenour, D.A., Sabatini, B.L., 2007. Distinctstructural and ionotropic roles of NMDA receptors in
Please cite this article as: Pisar, M., et al., Modified neocortical anfollowing prenatal inhibition of the kynurenine pathwbrainres.2014.06.016
controlling spine and synapse stability. J. Neurosci. 27,7365–7376.
Alvarez-Medina, R., Le Dreau, G., Ros, M., Marti, E., 2009.Hedgehog activation is required upstream of Wnt signalling tocontrol neural progenitor proliferation. Development 136,3301–3309.
Angot, E., Loulier, K., Nguyen-Ba-Charvet, K.T., Gadeau, A.-P.,Ruat, M., Traiffort, E., 2008. Chemoattractive activity of sonichedgehog in the adult subventricular zone modulates thenumber of neural precursors reaching the olfactory bulb. StemCells 26, 2311–2320.
Balordi, F., Fishell, G., 2007. Hedgehog signaling in thesubventricular zone is required for both the maintenance ofstem cells and the migration of newborn neurons. J. Neurosci.27, 5936–5947.
Baran, H., Schwarcz, R., 1990. Presence of 3-hydroxyanthranilicacid in rat tissues and evidence for its production fromanthranilic acid in the brain. J. Neurochem. 55, 738–744.
Beal, M.F., Swartz, K.J., Isacson, O., 1992. Developmental changesin brain kynurenic acid concentrations. Dev. Brain Res. 68,136–139.
Brown, A.S., 2006. Prenatal infection as a risk factor inschizophrenia. Schizophr. Bull. 32, 200–202.
Brown, A.S., 2011. The environment and susceptibility toschizophrenia. Prog. Neurobiol. 93, 23–58.
Cai, Y., Xiong, K., Chu, Y., Luo, D.-W., Luo, X.-G., Yuan, X.-Y.,Struble, R.G., Clough, R.W., Spencer, D.D., Williamson, A.,Kordower, J.H., Patrylo, P.R., Yan, X.X., 2009. Doublecortinexpression in adult cat and primate cerebral cortex relates toimmature neurons that develop into GABAergic subgroups.Exp. Neurol. 216, 342–356.
Charytoniuk, D., Porcel, B., Gomez, J.R., Faure, H., Ruat, M.,Traiffort, E., 2002. Sonic hedgehog signalling in the developingand adult brain. J. Physiol. (Paris) 96, 9–16.
Chiarugi, A., Carpenedo, R., Molina, M., Mattoli, L., Pellicciari, R.,Moroni, F., 1995. Comparison of the neurochemical andbehavioral-effects resulting from the inhibition of kynureninehydroxylase and/or kynureninase. J. Neurochem. 65,1176–1183.
Chiarugi, A., Moroni, F., 1999. Quinolinic acid formation inimmune-activated mice: studies with (m-nitrobenzoyl)-alanine (mNBA) and 3,4-dimethoxy-[-N-4-(-3-nitrophenyl)thiazol-2yl]-benzenesulfonamide (Ro 61-8048), two potent andselective inhibitors of kynurenine hydroxylase.Neuropharmacology 38, 1225–1233.
Cho, K.H., Jang, H.-J., Lee, E.-H., Yoon, S.H., Hahn, S.J., Jo, Y.-H.,Kim, M.-S., Rhie, D.-J., 2008. Differential cholinergicmodulation of Ca2þ transients evoked by back propagatingaction potentials in apical and basal dendrites of corticalpyramidal neurons. J. Neurophysiol. 99, 2833–2843.
Clark, C.J., Mackay, G.M., Smythe, G.A., Bustamante, S., Stone, T.W.,Phillips, S.R., 2005. Prolonged survival of a murine model ofcerebral malaria by kynurenine synthesis inhibition. Infect.Immun. 73, 5249–5251.
Colonnese, M.T., Zhao, J.P., Constantine-Paton, M., 2005. NMDAreceptor currents suppress synapse formation on sproutinaxons in vivo. J. Neurosci. 25, 1291–1303.
Couillard-Despres, S., Winner, B., Schaubeck, S., Aigner, R.,Vroemen, M., Weidner, N., Bogdahn, U., Winkler, J., Kuhn, H.G.,Aigner, L., 2005. Doublecortin expression levels in adult brainreflect neurogenesis. Eur. J. Neurosci. 21, 1–14.
Dalton, G.L., Wu, D.C., Huang, Y.T., Floresco, S.B., Phillips, A.G.,2012. NMDA GluN2A and GluN2B receptors play separate rolesin the induction of LTP and LTD in the amygdala and in theacquisition and extinction of conditioned fear.Neuropharmacology 62, 797–806.
De Montigny, A., Elhiri, I., Allyson, J., Cyr, M., Massicotte, G., 2013.NMDA reduces tau phosphorylation in rat hippocampal slices
d cerebellar protein expression and morphology in adult ratsay. Brain Research (2014), http://dx.doi.org/10.1016/j.

b r a i n r e s e a r c h ] ( ] ] ] ] ) ] ] ] – ] ] ] 15
by targeting NR2A receptors, GSK3 beta, and PKC activities.Neural Plast., 261593.
Dikranian, K., Ishimaru, M.J., Tenkova, T., Labruyere, J., Qin, Y.Q.,Ikonomidou, C., Olney, J.W., 2001. Apoptosis in the in vivomammalian forebrain. Neurobiol. Dis. 8, 359–379.
Dobelis., P., Staley, K.J., Cooper, D.C., 2012. Lack of modulation ofnicotinic acetylcholine alpha-7 receptor currents by kynurenicacid in adult hippocampal interneurons. PLoS One 7, e41108.
du Bois, T.M., Huang, X.F., 2007. Early brain developmentdisruption from NMDA receptor hypofunction: relevance toschizophrenia. Brain Res. Rev. 53, 260–270.
Erhardt, S., Olsson, S.K., Engberg, G., 2009. Pharmacologicalmanipulation of kynurenic acid potential in the treatment ofpsychiatric disorders. CNS Drugs 23, 91–101.
Ferrante, M., Migliore, M., Ascoli, G.A., 2013. Functional impact ofdendritic branch-point morphology. J. Neurosci. 33, 2156–2165.
Forrest, C.M., Khalil, O.S., Pisar, M., Smith, R.A., Darlington, L.G.,Stone, T.W., 2012. Prenatal activation of Toll-like receptors-3by administration of the viral mimetic poly(I:C) changessynaptic proteins, N-methyl-D-aspartate receptors andneurogenesis markers in offspring. Mol. Brain 5 (art.22).
Forrest, C.M., Khalil, O.S., Pisar, M., Darlington, L.G., Stone, T.W.,2013a. Prenatal inhibition of the tryptophan–kynureninepathway alters synaptic plasticity and protein expression inthe rat hippocampus. Brain Res. 1504, 1–15.
Forrest, C.M., Khalil, O.S., Pisar, P., McNair, K., Kornisiuk, E.,Snitcofsky, M., Gonzalez, N., Jerusalinsky, D., Darlington, L.G.,Stone, T.W., 2013b. Changes in synaptic transmission andprotein expression in the brains of adult offspring afterprenatal inhibition of the kynurenine pathway. Neuroscience254, 241–259.
Foster, K.A., McLaughlin, N., Edbauer, D., Phillips, M., Bolton, A.,Constantine-Paton, M., Sheng, M., 2010. Distinct roles of NR2Aand NR2B cytoplasmic tails in long-term potentiation. J.Neurosci. 30, 2676–2685.
Friocourt, G., Liu, J.S., Antypa, M., Rakic, S., Walsh, C.A.,Parnavelas, J.G., 2007. Both doublecortin and doublecortin-likekinase play a role in cortical interneuron migration. J.Neurosci. 27, 3875–3883.
Gilmartin, M.R., Kwapis, J.L., Helmstetter, F.J., 2013. NR2A-andNR2B-containing NMDA receptors in the prelimbic medialprefrontal cortex differentially mediate trace, delay, andcontextualfear conditioning. Learn. Mem. 20, 290–294.
Giorgini, F., Huang, S.-Y., Sathyasaikumar, K.V., Notarangelo, F.M.,Thomas, M.A.R., Tararina, M., Wu, Hui-Qiu, Schwarcz, R.,Muchowski, P.J., 2013. Targeted deletion of kynurenine 3-monooxygenase in mice: a new tool for studying kynureninepathway metabolism in periphery and brain. J. Biol. Chem.288, 36554–36566.
Gordon, U., Polsky, A., Schiller, J., 2006. Plasticity compartments inbasal dendrites of neocortical pyramidal neurons. J. Neurosci.26, 12717–12726.
Gray, J.A., Shi, Y., Usui, H., During, M.J., Sakimura, K., Nicoll, R.A.,2011. Distinct Modes of AMPA receptor suppression atdeveloping synapses by GluN2a and GluN2b: single-cell NMDAreceptor subunit deletion in vivo. Neuron 71, 1085–1101.
Guillemin, G.J., Kerr, S.J., Smythe, G.A., Smith, D.G., Kapoor, V.,Armati, P.J., Croitoru, J., Brew, B.J., 2001. Kynurenine pathwaymetabolism in human astrocytes: a paradox for neuronalprotection. J. Neurochem. 78, 842–853.
Guillemin, G.J., Smythe, G., Takikawa, O., Brew, B.J., 2005.Expression of indoleamine 2,3-dioxygenase and production ofquinolinic acid by human microglia, astrocytes, and neurons.Glia 49, 15–23.
Harris, L.W., Sharp, T., Gartlon, J., Jones, D.N.C., Harrison, P.J.,2003. Long-term behavioural, molecular and morphologicaleffects of neonatal NMDA receptor antagonism. Eur. J.Neurosci. 18, 1706–1710.
Please cite this article as: Pisar, M., et al., Modified neocortical anfollowing prenatal inhibition of the kynurenine pathwbrainres.2014.06.016
Henle, F., Dehmel, M., Leemhuis, J., Fischer, C., Meyer, D.K., 2012.Role of GluN2A and GluN2B subunits in the formation offilopodia and secondary dendrites in cultured hippocampalneurons. Naunyn–Schmied. Arch. Pharmacol. 385, 171–180.
Hilmas, C., Pereira, E.F.R., Alkondon, M., Rassoulpour, A.,Schwarcz, R., Albuquerque, E.X., 2001. The brain metabolitekynurenic acid inhibits alpha 7 nicotinic receptor activity andincreases non-alpha 7 nicotinic receptor expression:physiopathological implications. J. Neurosci. 21, 7463–7473.
Holtze, M., Saetre, P., Engberg, G., Schwieler, L., Werge, T.,Andreassen, O.A., Hall, H., Terenius, L., Agartz, I., Jonsson, E.G.,Schalling, M., Erhardt, S., 2012. Kynurenine 3-monooxygenasepolymorphisms: relevance for kynurenic acid synthesis inpatients with schizophrenia and healthy controls. J. PsychiatryNeurosci. 37, 53–57.
Hor, C.H.H., Tang, B.L., 2010. Sonic hedgehog as achemoattractant for adult NPCs. Cell Adhes. Migr. 4, 1–3.
Hornig, M., Weissenbock, H., Horscroft, N., Lipkin, W.I., 1999. Aninfection-based model of neurodevelopmental damage. Proc.Natl. Acad. Sci. USA 96, 12102–12107.
Ikonomidou, C., Bosch, F., Miksa, M., Bittigau, P., Vockler, J.,Dikranian, K., Tenkova, T.I., Stefovska, V., Turski, L.,Olney, J.W., 1999. Blockade of NMDA receptors and apoptoticneurodegeneration in the developing brain. Science 283,70–74.
Inta, D., Vogt, M.A., Pfeiffer, N., Koehr, G., Gass, P., 2013.Dichotomy in the anxiolytic versus antidepressant effect of C-terminal truncation of the GluN2A subunit of NMDAreceptors. Behav. Brain Res. 247, 227–231.
Iwasato, T., Datwani, A., Wolf, A.M., Nishiyama, H., Taguchi, Y.,Tonegawa, S., Knopfel, T., Erzurumlu, R.S., Itohara, S., 2000.Cortex-restricted disruption of NMDAR1 impairs neuronalpatterns in the barrel cortex. Nature 406, 726–731.
Jacobs, S.A., Tsien, J.Z., 2014. Overexpression of the NR2A subunitin the forebrain impairs long-term social recognition and non-social olfactory memory. Genes, Brain Behav. 13, 376–384.
Jessell, T.M., 2000. Neuronal specification in the spinal cord:Inductive signals and transcriptional codes. Nat. Rev. Genet. 1,20–29.
Jin, S.-X., Feig, L.A., 2010. Long-term potentiation in the CA1hippocampus induced by NR2A subunit-containing NMDAglutamate receptors is mediated by Ras-GRF2/Erk Map kinasesignaling. PLOS ONE 5, e11732.
Jin, K., Wang, X., Xie, L., Mao, X.O., Greenberg, D.A., 2010.Transgenic ablation of doublecortin-expressing cellssuppresses adult neurogenesis and worsens stroke outcomein mice. Proc. Natl. Acad. Sci. USA 107, 7993–7998.
Jung, Y.-H., Suh, Y.-H., 2010. Differential functions of NR2A andNR2B in short-term and long-term memory in rats.NeuroReport 21, 808–811.
Khalil, O.S., Forrest, C.M., Pisar, M., Smith, R.A., Darlington, L.G.,Stone, T.W., 2013. Prenatal activation of maternal TLR3receptors by viral-mimetic poly(I:C) modifies GluN2Bexpression in embryos and sonic hedgehog in offspring in theabsence of kynurenine pathway activation. Immunopharm.Immunotoxicol. 35, 581–593.
Khalil, O.S., Pisar, M., Forrest, C.M., Vincenten, M.C.J., Darlington,L.G., Stone, T.W., 2014. Prenatal inhibition of the kynureninepathway leads to structural changes in the hippocampus ofadult rat offspring. Eur. J. Neurosci. 39, 1558–1571.
Kochlamazashvili, G., Bukalo, O., Senkov, O., Salmen, B., Gerardy-Schahn, R., Engel, A.K., Schachner, M., Dityatev, A., 2012.restoration of synaptic plasticity and learning in young andaged NCAM-deficient mice by enhancing neurotransmissionmediated by GluN2A-containing NMDA receptors. J. Neurosci.32, 2263–2275.
Kuramoto, T., Kuwamura, M., Serikawa, T., 2004. Rat neurologicalmutations cerebellar vermis defect and hobble are caused by
d cerebellar protein expression and morphology in adult ratsay. Brain Research (2014), http://dx.doi.org/10.1016/j.

b r a i n r e s e a r c h ] ( ] ] ] ] ) ] ] ] – ] ] ]16
mutations in the netrin-1 receptor gene unc5H3. Mol. BrainRes. 122, 103–108.
Kwon, H.-B., Sabatini, B.L., 2011. Glutamate induces de novogrowth of functional spines in developing cortex. Nature 474,100–104.
Leung, L.S., Peloquin, P., 2010. Cholinergic modulation differsbetween basal and apical dendritic excitation of hippocampalCA1 pyramidal cells. Cereb. Cortex 20, 1865–1877.
Li, Y.-H., Wang, J., 2013. Membrane insertion of new AMPAreceptors and LTP induced by glycine is prevented by blockingNR2A-containing NMDA receptors in the rat visual cortexin vitro. Curr. Neurovasc. Res. 10, 70–75.
Linderholm, K.R., Skogh, E., Olsson, S.K., Dahl, M.-L., Holtze, M.,Engberg, G., Samuelsson, M., Erhardt, S., 2012. Increased levelsof kynurenine and kynurenic acid in the CSF of patients withschizophrenia. Schizophr. Bull. 38, 426–432.
Lujan, B., Liu, X., Wan, Q., 2012. Differential roles of GluN2A- andGluN2B-containing NMDA receptors in neuronal survival anddeath. Intern. J. Physiol. Pathophysiol. Pharmacol. 4, 211–218.
Manitt, C., Mimee, A., Eng, C., Pokinko, M., Stroh, T., Cooper, H.M.,Kolb, B., Flores, C., 2011. The netrin receptor DCC is required inthe pubertal organization of mesocortical dopamine circuitry.J. Neurosci. 31, 8381–8394.
Matta, J.A., Ashby, M.C., Sanz-Clemente, A., Roche, K.W., Isaac, J.T.R., 2011. mGluR5 and NMDA receptors drive the experience-and activity-dependent NMDA Receptor NR2B to NR2Asubunit switch. Neuron 70, 339–351.
Meyer, U., Feldon, J., 2010. Epidemiology-drivenneurodevelopmental animal models of schizophrenia. Prog.Neurobiol. 90, 285–326.
Moeller, M., Du Preez, J.L., Emsley, R., Harvey, B.H., 2012. Socialisolation rearing in rats alters plasma tryptophan metabolismand is reversed by sub-chronic clozapine treatment.Neuropharmacology 62, 2499–2506.
Mok, M.H.S., Fricker, A.C., Weil, A., Kew, J.N.C., 2009.Electrophysiological characterisation of the actions ofkynurenic acid at ligand-gated ion channels.Neuropharmacology 57, 242–249.
Myint, A.M., Schwarz, M.J., Verkerk, R., Mueller, H.H., Zach, J.,Scharpe, S., Steinbusch, H.W.M., Leonard, B.E., Kim, Y.K., 2011.Reversal of imbalance between kynurenic acid and 3-hydroxykynurenine by antipsychotics in medication-naiveand medication-free schizophrenic patients. Brain Behav.Immun. 25, 1576–1581.
Osborne, P.B., Halliday, G.M., Cooper, H.M., Keast, J.R., 2005.Localization of immunoreactivity for deleted in colorectalcancer (DCC), the receptor for the guidance factor netrin-1, inventral tier dopamine projection pathways in adult rodents.Neuroscience 131, 671–681.
Owczarek, S., Hou, J., Secher, T., Kristiansen, L.V., 2011.Phencyclidine treatment increases NR2A and NR2B N-methyl-D-aspartate receptor subunit expression in rats. NeuroReport22, 935–938.
Palma, V., Lim, D.A., Dahmane, N., Sanchez, P., Brlonne, T.C.,Herzberg, C.D., Gitton, Y., Carleton, A., Alvarez-Buylla, A.,Altaba, A.R.I., 2005. Sonic hedgehog controls stem cellbehavior in the postnatal and adult brain. Development 132,335–344.
Pearce, B.D., 2003. Modeling the role of infections in the etiologyof mental illness. Clin. Neurosci. Res. 3, 271–282.
Perkins, M.N., Stone, T.W., 1982. An iontophoretic investigation ofthe action of convulsant kynurenines and their interactionwith the endogenous excitant quinolinic acid. Brain Res. 247,184–187.
Pfaffl, M.W., Horan, G.W., Dempfle, L., 2002. Relative expressionsoftware tool (REST) for group-wise comparison and statisticalanalysis of relative expression results in real-time PCR.Nucleic Acids Res. 30, e36.
Please cite this article as: Pisar, M., et al., Modified neocortical anfollowing prenatal inhibition of the kynurenine pathwbrainres.2014.06.016
Rajan, I., Cline, H.T., 1998. Glutamate receptor activity is requiredfor normal development of tectal cells dendrites in vivo.J. Neurosci. 18, 7836–7846.
Ramakers, C., Ruijter, J.M., Deprez, R.H.L., Moorman, A.F.M., 2003.Assumption-free analysis of quantitative real-timepolymerase chain reaction (PCR) data. Neurosci. Lett. 339,62–66.
Ramoa, A.S., Mower, A.F., Liao, D., Safri, S.I.A., 2001. Suppressionof cortical NMDA receptor function prevents development oforientation selectivity in the primary visual cortex. J.Neurosci. 21, 4299–4309.
Rover, S., Cesura, A.M., Huguenin, P., Kettler, R., Szente, A., 1997.Synthesis and biochemical evaluation of N-(4-phenylthiazol-2-yl)benzenesulfonamides as high-affinity inhibitors ofkynurenine 3-hydroxylase. J. Med. Chem. 40, 4378–4385.
Sajikumar, S., Korte, M., 2011. Different compartments of apicalCA1 dendrites have different plasticity thresholds forexpressing synaptic tagging and capture. Learn. Mem. 18,327–331.
Sathyasaikumar, K.V., Stachowski, E.K., Wonodi, I., Roberts, R.C.,Rassoulpour, A., McMahon, R.P., Schwarcz, R., 2011. Impairedkynurenine pathway metabolism in the prefrontal cortex ofindividuals with schizophrenia. Schizophr. Bull. 37, 1147–1156.
Schwarcz, R., Pellicciari, R., 2002. Manipulation of brainkynurenines: glial targets, neuronal effects, and clinicalopportunities. J. Pharmacol. Exp. Ther. 303, 1–10.
Schwarcz, R., Ramminga, C.A., Kurlan, R., Shoulson, I., 1988.Cerebrospinal fluid levels of quinolinic acid in Huntington’sdisease and schizophrenia. Ann. Neurol. 24m, 580–582.
Schwarcz, R., Rassoulpour, A., Wu, H.Q., Medoff, D., Tamminga, C.A., Roberts, R.C., 2001. Increased cortical kynurenate contentin schizophrenia. Biol. Psychiatry 50, 521–530.
Sepulveda, F.J., Bustos, F.J., Inostroza, E., Zuniga, F.A., Neve, R.L.,Montecino, M., van Zundert, B., 2010. Differential roles ofNMDA receptor subtypes NR2A and NR2B in dendritic branchdevelopment and requirement of RasGRF1. J. Neurophysiol.103, 1758–1770.
Sholl, D.A., 1953. Dendritic organization in the neurons of thevisual and motor cortices of the cat. J. Anat. 87, 387–406.
Sin, W.C., Haas, K., Ruthazer, E.S., Cline, H.T., 2002. Dendritegrowth increased by visual activity requires NMDA receptorand Rho GTPases. Nature 419, 475–480.
Spampanato, J., Sullivan, R.K., Turpin, F.R., Bartlett, P.F., Sah, P.,2012. Properties of doublecortin expressing neurons in theadult mouse dentate gyrus. PLOS One 7, e41029.
Spassky, N., Han, Y.-G., Aguilar, A., Strehl, L., Besse, L., Laclef, C.,Ros, M.R., Garcia-Verdugo, J.M., Alvarez-Buylla, A., 2008.Primary cilia are required for cerebellar development andShh-dependent expansion of progenitor pool. Dev. Biol. 317,246–259.
Speciale, C., Wu, H.Q., Cini, M., Marconi, M.A., Varasi, M.,Schwarcz, R., 1996. (R,S)-3,4-dichlorobenzoylalanine (FCE28833A) causes a large and persistent increase in brainkynurenic acid levels in rats. Eur. J. Pharmacol. 315, 263–267.
Srinivasan, K., Leone, D.P., Bateson, R.K., Dobreva, G., Kohwi, Y.,Kohwi-Shigematsu, T., Grosschedl, R., McConnell, S.K., 2012.A network of genetic repression and derepression specifiesprojection fates in the developing neocortex. Proc. Natl. Acad.Sci. USA 109, 19071–19078.
Stone, T.W., 2007. Kynurenic acid blocks nicotinic synaptictransmission to hippocampal interneurons in young rats.Eur. J. Neurosci. 25, 2656–2665.
Stone, T.W., Darlington, L.G., 2002. Endogenous kynurenines astargets for drug discovery and development. Nat. Rev. DrugDiscov. 1, 609–620.
Stone, T.W., Darlington, L.G., 2013. The kynurenine pathway as atherapeutic target in cognitive and neurodegenerativedisorders. Br. J. Pharmacol. 169, 1211–1277.
d cerebellar protein expression and morphology in adult ratsay. Brain Research (2014), http://dx.doi.org/10.1016/j.

b r a i n r e s e a r c h ] ( ] ] ] ] ) ] ] ] – ] ] ] 17
Stone, T.W., Perkins, M.N., 1981. Quinolinic acid: a potentendogenous excitant at amino acid receptors in CNS. Eur. J.Pharmacol. 72, 411–412.
Stone, T.W., Stoy, N., Darlington, L.G., 2013. An expanding rangeof targets for kynurenine metabolites of tryptophan. TrendsPharmacol. Sci. 34, 136–143.
Sun, H., Guan, L., Zhu, Z., Li, H., 2013a. Reduced Levels of NR1and NR2A with depression-like behavior in different brainregions in prenatally stressed juvenile offspring. PLOS One 8,e81775.
Sun, H., Jia, N., Guan, L., Su, Q., Wang, D., Li, H., Zhu, Z., 2013b.Involvement of NR1, NR2A different expression in brainregions in anxiety-like behavior of prenatally stressedoffspring. Behav. Brain Res. 257, 1–7.
Traiffort, E., Charytoniuk, D.A., Faure, H., Ruat, M., 1998. Regionaldistribution of sonic hedgehog, patched, and smoothenedmRNA in the adult rat brain. J. Neurochem. 70, 1327–1330.
Traiffort, E., Moya, K.L., Faure, H., Hassig, R., Ruat, M., 2001. Highexpression and anterograde axonal transport ofaminoterminal sonic hedgehog in the adult hamster brain.Eur. J. Neurosci. 14, 839–850.
Traiffort, E., Angot, E., Ruat, M., 2010. Sonic hedgehog in themammalian brain. J. Neurochem. 113, 576–590.
Ultanir, S.K., Kim, J.E., Hall, B.J., Deerinck, T., Ellisman, M., Ghosh,A., 2007. Regulation of spine morphology and spine density byNMDA receptor signaling in vivo. Proc. Natl. Acad. Sci. USA104, 19553–19558.
Vincent, A., Kessler, J.P., Baude, A., Dipasquale, E., Tell, F., 2004.N-methyl-D-aspartate receptor activation exerts a dual control
Please cite this article as: Pisar, M., et al., Modified neocortical anfollowing prenatal inhibition of the kynurenine pathwbrainres.2014.06.016
on postnatal development of nucleus tractus solitarii neuronsin vivo. Neuroscience 126, 185–194.
Walker, D.W., Curtis, B., Lacey, B., Nitsos, I., 1999. Kynurenic acidin brain and cerebrospinal fluid of fetal, newborn, and adultsheep and effects of placental embolization. Pediatr. Res. 45,820–826.
Wang, C.-C., Held, R.G., Chang, S.-C., Yang, L., Delpire, E., Ghosh,A., Hall, B.J., 2011. A critical role for gluN2B-containing NMDAreceptors in cortical development and function. Neuron 72,789–805.
Wang, P.Y., Petralia, R.S., Wang, Y.X., Wenthold, R.J., Brenowitz, S.D., 2011. Functional NMDA receptors at axonal growth conesof young hippocampal neurons. J. Neurosci. 31, 9289–9297.
Wyllie, D.J.A., Livesey, M.R., Hardingham, G.E., 2013. Influence ofGluN2 subunit identity on NMDA receptor function.Neuropharmacology 74, 4–17.
Xiao, L., Feng, C., Chen, Y., 2010. Glucocorticoid rapidly enhancesNMDA-evoked neurotoxicity by attenuating the NR2A-containing NMDA receptor-mediated ERK1/2 activation.Mol. Endocrinol. 24, 497–510.
Xiong, K., Luo, D.-W., Patrylo, P.R., Luo, X.-G., Struble, R.G., Clough,R.W., Yan, X.-X., 2008. Doublecortin-expressing cells arepresent in layer II across the adult guinea pig cerebral cortex:partial colocalization with mature interneuron markers. Exp.Neurol. 211, 271–282.
Zhou, X., Ding, Q., Chen, Z., Yun, H., Wang, H., 2013. Involvementof the GluN2A and GluN2B subunits in synaptic andextrasynaptic N-methyl-D-aspartate receptor function andneuronal excitotoxicity. J. Biol. Chem. 288, 24151–24159.
d cerebellar protein expression and morphology in adult ratsay. Brain Research (2014), http://dx.doi.org/10.1016/j.
Copyright © 2022 FDOKUMEN




















