
Melanoma, Nevogenesis, and Stem Cell Biology
16
Melanoma, Nevogenesis, and Stem Cell Biology James M. Grichnik 1 It is now well established that a subpopulation of tumor stem cells (TSCs) are present within cancer tissues. This suggests that tumors evolve from stem cells; however, the exact cell of tumor origin, the potential role of dedifferentiation, and the role of plasticity in tumor development are largely unknown. A model cancer for the study of the oncologic process is melanoma. The developmental biology of melanocytes is relatively well understood, the cells pigment as they differentiate making them easy to identify, and benign and malignant tumors develop on the skin surface allowing direct observation of growth features, early detection, and removal. This ready access to early-stage tumors will facilitate study of the early oncologic processes and the role of tissue stem cells. Melanomas, like other cancers, include a subpopulation of TSCs. These TSCs have access to embryologic developmental programs, including the capacity to differentiate along multiple cell lineages. For example, melanomas can activate germ–cell pathways with major implications for TSC self-renewal through the activation of telomerase and genomic instability through the collision of meiotic and mitotic pathways (meiomitosis). The TSC model is still evolving, but the existence of TSCs has significant ramifications for tumor development, diagnosis, prognosis, and treatment of melanoma and other cancers. Journal of Investigative Dermatology (2008) 128, 2365–2380; doi:10.1038/jid.2008.166 Introduction Cancer development, progression, and therapeutics remain at the forefront of medical research. The recognition of a subpopulation of tumor stem cells (TSCs) in solid cancers has reinvigo- rated the field. These cells have the capacity to self-renew and give rise to more differentiated cell forms (Reya et al., 2001). The existence of these TSCs implicates a pluripotent stem cell as the cell of origin for cancer. How- & 2008 The Society for Investigative Dermatology www.jidonline.org 2365 PERSPECTIVE Editor’s Note Malignant melanoma has long been identified as one of the most important and inscrutable of all skin diseases. In his 1941 review DeCholnoky concluded ‘‘Radical surgery is the treatment of choice, and should consist of wide local excision... followed by regional lymph node dissection....’’ While the recommended extent of surgery may have decreased over the last 60+ years, it is remarkable that our knowledge of the basic biology of malignant melano- ma has not resulted in significant changes in treatment; indeed, metastatic melanoma remains one of the malig- nancies most resistant to treatment. In the next two issues of the JID, recent advances in our understanding of the biology of malignant melanoma are featured. In this issue, Grichnik discusses how melanoma stem cells may play a critical role in the pathogenesis of melanoma and its resistance to conventional therapy. Zaidi and co-workers explore the role of UV light in the pathogenesis of melanoma and how animal models can advance our understanding of both initiation and metastasis. Next month, Hocker and co-workers review recent advances in the genetics of melanoma and how they may lead to new specific therapeutic interventions. Finally, Fang and co-workers discuss how recent advances in immunology have resulted in new strategies for using the immune system to treat life-threatening metastatic disease. In 1930 Dr James Ewing wrote, ‘‘The problems of melanoma maintain their position as the most interesting and complex of any department of oncology..... Possibly there are other important data lying within easy reach of the alert observer.’’ While the ‘‘problems of melanoma’’ persist, these Perspectives demonstrate the significant progress and challenges that characterize our understanding of melanoma in the 21st century; I hope they will present new directions for study by the ‘‘alert observer’’. Russell P. Hall, III, Deputy Editor Received 31 October 2007; revised 9 January 2008; accepted 10 January 2008 1 Departments of Medicine and Cell Biology, Duke University Medical Center, Durham, North Carolina, USA Correspondence: Dr James M. Grichnik, Division of Dermatology, Department of Medicine, Duke University Medical Center, 4032 Hospital South, Box 3135, Durham, North Carolina 27710, USA. E-mail: [email protected] Abbreviations: bFGF, basic fibroblast growth factor; DCT, dopachrome tautomerase; EPI-NCSC, epidermal neural crest epidermal stem cells; ET-3, endothelin 3; ESC, embryonic stem cell; SCF, stem cell factor; SKP, skin-derived precursor; TA, transiently amplifying; TSC, tumor stem cell
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Melanoma, Nevogenesis, and Stem Cell Biology

Melanoma, Nevogenesis, and Stem Cell BiologyJames M. Grichnik1
It is now well established that a subpopulation of tumor stem cells (TSCs) are present within cancer tissues. Thissuggests that tumors evolve from stem cells; however, the exact cell of tumor origin, the potential roleof dedifferentiation, and the role of plasticity in tumor development are largely unknown. A model cancer forthe study of the oncologic process is melanoma. The developmental biology of melanocytes is relatively wellunderstood, the cells pigment as they differentiate making them easy to identify, and benign and malignanttumors develop on the skin surface allowing direct observation of growth features, early detection, andremoval. This ready access to early-stage tumors will facilitate study of the early oncologic processes and therole of tissue stem cells. Melanomas, like other cancers, include a subpopulation of TSCs. These TSCs haveaccess to embryologic developmental programs, including the capacity to differentiate along multiple celllineages. For example, melanomas can activate germ–cell pathways with major implications for TSC self-renewalthrough the activation of telomerase and genomic instability through the collision of meiotic and mitoticpathways (meiomitosis). The TSC model is still evolving, but the existence of TSCs has significant ramificationsfor tumor development, diagnosis, prognosis, and treatment of melanoma and other cancers.
Journal of Investigative Dermatology (2008) 128, 2365–2380; doi:10.1038/jid.2008.166
Introduction
Cancer development, progression, andtherapeutics remain at the forefront ofmedical research. The recognition of a
subpopulation of tumor stem cells(TSCs) in solid cancers has reinvigo-rated the field. These cells have thecapacity to self-renew and give rise to
more differentiated cell forms (Reyaet al., 2001). The existence of theseTSCs implicates a pluripotent stem cellas the cell of origin for cancer. How-
& 2008 The Society for Investigative Dermatology www.jidonline.org 2365
PERSPECTIVE
Editor’s Note
Malignant melanoma has long been identified as one of themost important and inscrutable of all skin diseases. In his1941 review DeCholnoky concluded ‘‘Radical surgery isthe treatment of choice, and should consist of wide localexcision... followed by regional lymph node dissection....’’While the recommended extent of surgery may havedecreased over the last 60+ years, it is remarkable thatour knowledge of the basic biology of malignant melano-ma has not resulted in significant changes in treatment;indeed, metastatic melanoma remains one of the malig-nancies most resistant to treatment. In the next two issuesof the JID, recent advances in our understanding of thebiology of malignant melanoma are featured. In this issue,Grichnik discusses how melanoma stem cells may play acritical role in the pathogenesis of melanoma and itsresistance to conventional therapy. Zaidi and co-workersexplore the role of UV light in the pathogenesis ofmelanoma and how animal models can advance our
understanding of both initiation and metastasis. Nextmonth, Hocker and co-workers review recent advancesin the genetics of melanoma and how they may lead tonew specific therapeutic interventions. Finally, Fang andco-workers discuss how recent advances in immunologyhave resulted in new strategies for using the immunesystem to treat life-threatening metastatic disease. In 1930Dr James Ewing wrote, ‘‘The problems of melanomamaintain their position as the most interesting and complexof any department of oncology..... Possibly there are otherimportant data lying within easy reach of the alertobserver.’’ While the ‘‘problems of melanoma’’ persist,these Perspectives demonstrate the significant progress andchallenges that characterize our understanding of melanomain the 21st century; I hope they will present new directionsfor study by the ‘‘alert observer’’.
Russell P. Hall, III, Deputy Editor
Received 31 October 2007; revised 9 January 2008; accepted 10 January 2008
1Departments of Medicine and Cell Biology, Duke University Medical Center, Durham, North Carolina, USA
Correspondence: Dr James M. Grichnik, Division of Dermatology, Department of Medicine, Duke University Medical Center, 4032 Hospital South, Box 3135,Durham, North Carolina 27710, USA. E-mail: [email protected]
Abbreviations: bFGF, basic fibroblast growth factor; DCT, dopachrome tautomerase; EPI-NCSC, epidermal neural crest epidermal stem cells; ET-3, endothelin 3;ESC, embryonic stem cell; SCF, stem cell factor; SKP, skin-derived precursor; TA, transiently amplifying; TSC, tumor stem cell

ever, ‘‘dedifferentiation’’ from a maturetissue cell has not been completelyexcluded. Although the cell of origincan still be questioned, there is nodoubt that the TSCs exist and play acritical role in tumor maintenance.TSCs, although only a small fractionof the tumor bulk, have been shown tobe the cells with the capacity to giverise to new tumor nodules. Tumori-genic (TSCs) versus non-tumorigenicpopulations were initially defined inhematologic malignancies (Lapidotet al., 1994; Bonnet and Dick, 1997)and have recently been identified inbreast (Al-Hajj et al., 2003), pancreatic(Li et al., 2007), colon (Dalerba et al.,2007; O’Brien et al., 2007; Ricci-Vitiani et al., 2007), squamous cell(Prince et al., 2007), brain (Singh et al.,2003; Kondo et al., 2004), and lungcancers (Ho et al., 2007). TSC popula-tions have also been shown to be ableto differentiate along multiple celllineages; for example, in glioma,TSCs have been shown to differentiatealong both the neural and glial path-ways (Kondo et al., 2004). The findingssuggest that TSCs have access toembryologic developmental programs,including the capacity to differentiatealong multiple cell lineages.
Melanoma, like other cancers, in-cludes a subpopulation of TSCs. Thetumor often visibly develops on thesurface of the skin, and with the adventof dermoscopy, and developingtechnologies such as confocal micro-scopy, the early development ofthese lesions will be able to be betterstudied. In addition to the malignantlesions, benign nevi also develop onthe skin surface and may provideadditional critical information aboutthe role of tissue stem cells and/ormature melanocytes in the early stagesof tumor evolution. The relativelywell-defined developmental biology ofmelanocytes, as well as the readyaccess to normal and diseased tissue,makes melanoma a model system forthe study of stem cell biology andcarcinogenesis. Tumorigenesis is nowbest viewed as an aberrant develop-mental process. This review examinesselected aspects of normal embryologicand melanocytic development, home-ostasis, nevogenesis, and tumorgenesis,
as they may apply to melanocytic TSCbiology.
Normal embryologic development
The fertilized egg is the ultimate‘‘stem cell’’. During cell division, theprotoplasm is asymmetrically distribu-ted due to the influence of cellular andextracellular signal gradients. As cellnumbers expand and the cells areexposed to local environmental signals,selective differentiation occurs and avast array of tissue structures is formed.During the cellular developmental pro-cess, nuclear DNA is modified (altera-tion of histones and DNA methylation)and segregated within the nuclearmatrix structure controlling gene ex-pression and cell fate.
The developing cell is dependent onaccurate DNA primary sequence, andtherefore DNA mutations are activelyidentified and repaired. If mutationscannot be repaired, the cell is depen-dent on the intact apoptotic pathwaysfor elimination, as otherwise a mosaicdisorder would develop. Once a defec-tive cell has been eliminated, its posi-tion in the developmental gradient willbe occupied by a neighboring cell,which will differentiate appropriatelyas long as it has sufficient plasticity.Early embryonic cells have remarkableplasticity as up to one-fourth of the8-cell embryo can be removed withoutappreciable long-term sequella (Hardyet al., 1990; Pierce et al., 1997). Withfurther development, the degree ofplasticity retained by cells is most likelyto be dependent on its ultimate func-tion. Some cells would be expected notto maintain the capacity to differentiatealong other cell lineages. Obviousexamples would include cells in whichthe nucleus has been reabsorbed suchas red blood cells or corneocytes, cellsthat have undergone meiotic eventssuch as egg or sperm cells, or cells inwhich DNA recombination has occur-red such as T and B immunoregulatorycells. However, to sustain and repairtissue, a considerable number of cellsmust maintain significant plasticity intoadulthood. Presumably, these cells areretained as specialized tissue stemcells, but it is possible that some well-differentiated cells retain intrinsic plas-ticity. The fact that some cell nuclei can
be used in cloning experiments suggeststhat nuclear changes in some cells aresufficiently reversible to generate a newembryo (Oback and Wells, 2007).
The human developmental processproceeds quite quickly, and within 9months a single cell gives rise to morethan a trillion individual cells with atotal mass of more than 3 kg. Thiscellular mass is elegantly organized,including a multitude of different celltypes appropriately interacting withtightly controlled proliferation/apopto-tic and mitotic/meiotic mechanisms,with the capacity and plasticity to growand repair tissues for about a century. ATSC inherently has access to all thesecapacities.
Embryonic development of melanocytes
Melanocytic cells appear to be primar-ily derived from the developing neuralcrest. The migration of the neural cresthas been elegantly studied through theutilization of a reporter system thatlabels cells with an activated dopa-chrome tautomerase (DCT) promoter,which is expressed in melanocytic andneural progenitors (Mackenzie et al.,1997; Jiao et al., 2006). In the murinesystem, melanocytic precursor cells canbe first recognized in the neural crest atapproximately embryonic day 8.5,these cells migrate between the derma-tome and the overlying ectodermin the dorsolateral pathway, migrationcontinues ventrally through the devel-oping dermis at day 10.5, the cellsbegin to insert into the epidermis andthe developing hair follicles at day14.5, these cells proliferate and differ-entiate with pigment synthesis observedaround postnatal day 4 (Wilkie et al.,2002).
The melanocytic developmental pro-cess is very dependent on stem cellfactor (SCF) and its receptor, KIT. Dis-tinct waves of SCF/KIT dependence andindependence play a role in melanocytesurvival, migration, and epidermal in-sertion (Nishikawa et al., 1991; Okuraet al., 1995; Yoshida et al., 1996). Thesewaves of KIT dependence and indepen-dence are also seen during the haircycle (Nishikawa et al., 1991). Inaddition, endothelin 3 (ET-3) and theendothelin B receptor play a critical rolein melanocyte development, as loss of
2366 Journal of Investigative Dermatology (2008), Volume 128
JM GrichnikMelanoma, Nevogenesis, and Stem Cell Biology

these pathways also results in the loss ofcutaneous melanocytes (Baynash et al.,1994; Hosoda et al., 1994). A number ofother cytokine pathways have also beendocumented to play a role, includinga-melanocyte stimulating hormone,basic fibroblast growth factor (bFGF),nerve growth factor, endothelin 1,granulocyte macrophage-colony stimu-lating factor, and hepatocyte growthfactor (for review see Halaban, 2000;Hirobe, 2005).
Melanocytic development can alsobe driven in vitro through the use ofembryonic stem cells (ESC). Usingmouse ESCs cultivated on a feederlayer of ST2 cells, dexamethasone wasnoted to induce the formation ofmelanocytes (Yamane et al., 1999).Inhibition of the KIT pathway (withACK2) blocked melanocyte produc-tion, whereas the addition of ET3enhanced the number of melanocyticcells. Another group noted that theaddition of retinoic acid to the ESCculture increased the production ofmelanocytes (Motohashi et al., 2007).KITþ cells developed were isolated bycell sorting and were shown to bemultipotent with the capacity to giverise to melanocytes, glia, and neurons.The development of melanocytes couldbe antagonized by an ET-3 inhibitor(BQ788). A third group has demon-strated the need for Wnt3a for thedevelopment of melanocytes in theirESC system (Fang et al., 2006). In thepresence of WNT3a and ET-3, melano-cytes could be produced. Melanocyteproduction was further augmented bythe addition of SCF.
Thus, there is a well-orchestrated setof events within the embryonic massthat exposes embryonic cells to theappropriate factors to differentiatethe proper number of melanocytes inthe correct location. Theoretically, thisspecific series of developmental expo-sures reorganizes the DNA nuclearmatrix to initiate the appropriate ex-pression of melanocytic genes. Themelanocytic differentiation processcan also be induced in vitro by expos-ing ESCs to the appropriate cytokines,potentially allowing a more straightfor-ward study of nuclear changes.Although this melanocytic develop-mental process is normally driven by
external environmental signals, it ismost likely that the internal abnormalactivation of intracellular signalingpathways, such as the activation of B-Raf, in a stem cell could also drivemelanocytic differentiation.
Adult melanocytes
Melanocytes integrate into the epider-mis and function to provide melanin asa UV protectant to neighboring kerati-nocytes. The melanocytes may alsoperform other functions, including en-docrine (Slominski et al., 2007; Takedaet al., 2007), metal chelation (Farmeret al., 2003; Meyskens et al., 2004;Hong and Simon, 2007), and immunefunctions, yet to be fully explored(Burkhart and Burkhart, 2005). In per-forming these duties, the melanocyte issusceptible to UV and oxidative stress,which may create genetic mutations.Ideally, damaged melanocytes wouldbe eliminated. Although normal mela-nocytic turnover is generally impercep-tible, melanocytes being dischargedthrough the epidermis (‘‘pagetoid’’melanocytes) can be seen after acutesun exposure, over nevi, and melano-mas (Pharis and Zitelli, 2001; Petronic-Rosic et al., 2004).
The number of melanocytes in theepidermis is tightly controlled and thisis primarily due to their interaction withlocal keratinocytes. A major homeo-static factor in the adult skin is SCF.Epidermal keratinocytes produce SCFin both a membrane bound and solubleform, but local fibroblasts may alsocontribute to SCF levels. SCF/KIT sti-mulation or inhibition has been shownto drive proliferation or loss, respec-tively, of melanocytes in human skin(Grichnik et al., 1995, 1998). Expres-sion of SCF in the mouse epidermisresults in the retention of melanocyteswhen they would have otherwise beenlost (Kunisada et al., 1998). Signalingthrough SCF/KIT and other pathwaysprovides the homeostatic signaling tomaintain the appropriate number ofepidermal melanocytes in adult skin.
Melanocytes can be cultured fromisolated epidermal sheets. Multiplegrowth factors or chemical stimulantsare necessary to promote and maintainmelanocyte proliferation in vitro (Ha-laban, 2000). The response of the cells
to SCF in vitro can be modulated, withSCF functioning as a survival factorunder low bFGF conditions, a prolif-erative factor with intermediate levelsof bFGF, but having no additionalproliferative effect over bFGF whenhigh levels of bFGF are present (Linet al., 2000). Thus, the homeostatic roleof SCF can be partially recapitulatedin vitro.
Melanocytes obtained from neonatalforeskins grow robustly while melano-cytes from older human skin samplesgrow poorly. Eventually, melanocyteswill senesce in culture (Bennett andMedrano, 2002). This suggests that theseepidermal melanocyte cultures either donot include a stem cell component orthe existing culture conditions do notpromote or maintain melanocytic stemcells. However, the melanocytes inculture may have some plasticity, as‘‘fibroblast’’ outgrowth is not uncom-mon in melanocyte cultures. Althoughthis is assumed to be due to contamina-tion, it is possible that these cells arederived from melanocytic precursors.
As the human ages, the capacity tomake and retain melanocytic cellsappears to diminish. The most obviousexample of this is the graying ofhair. Although this loss of melanocytesis generally considered permanent,it is interesting to note that there arecase reports describing spontaneoushair repigmentation (Etienne et al.,2002; Kavak et al., 2005).
Thus, epidermal melanocytesclearly play a critical role in UVprotection. As a consequence of thisfunction, these cells may be particu-larly susceptible to mutation and needto be regularly turned over to preventthe accumulation of damaged DNA.Presumably, these damaged cells aredischarged through the epidermis. Thenumber of epidermal melanocytes istightly controlled by local factors. Withaging, the total number of melanocytesgradually diminishes.
Stem cell populations in adult skin
Melanocytes, like cells in many othertissues, are turned over and replenishedfrom immature precursors. Diseaseprocesses such as vitiligo give us awindow into this process. In activevitiligo, differentiated melanocytes are
www.jidonline.org 2367
JM GrichnikMelanoma, Nevogenesis, and Stem Cell Biology

destroyed. During recovery, new mel-anocytes migrate into the epidermisfrom a protected population of cells(Figure 1). A similar phenomenon canbe driven experimentally in zebrafish.Treatment with 4-(4-morpholinobu-tylthio)phenol results in the death ofdifferentiated melanocytes, and thesecells are replaced by cells derived froma precursor population (Yang andJohnson, 2006). Thus, differentiatedmelanocytes are replaced by cells thatdifferentiate from an immature precur-sor population.
A major reservoir for melanocyticprecursors is the hair follicle. Precursorcells have been identified in this loca-tion through the expression of KIT(Grichnik et al., 1996). These cells alsoexpressed Bcl-2, which may play a rolein the survival of the stem cell compart-ment, because mice defective forBcl-2 loose melanocytic stem cells(Nishimura et al., 2005). Melanocyteisolation from the hair follicle hasrevealed the presence of heavily pig-mented mature and amelanotic imma-ture cell populations (Na et al., 2006).Consistent with stem cell biology, theamelanotic population was shown tohave a slower doubling rate but en-hanced a long-term proliferation capa-city compared with the pigmented cellpopulation. With age and graying ofthe hair, residual melanocytic stem cellpopulations have been noted to bedecreased (Nishimura et al., 2005).
The melanocytic stem cell compart-ment in the murine hair follicle hasbeen studied using a DCT reportersystem to identify the melanocyticlineage cells (Nishimura et al., 2002).These cells gave rise to transientlyamplifying (TA) population of melano-cytes that populated the hair bulb;however, some of the cells were ableto return to a quiescent state in the hairbulge area. KIT receptor expressionappears to be downregulated in aquiescent subpopulation but upregu-lated on migratory melanocytic cellsduring the hair cycle (Botchkarevaet al., 2001; Peters et al., 2002). Inthe quiescent cells, although a fewpathways were upregulated includingthe Notch pathway, there appeared tobe a global suppression of transcrip-tion, including decreased expression ofmelanocytic genes (Nishikawa andOsawa, 2007). At the end of each haircycle, differentiated melanocytes inthe hair bulb are presumed to undergoapoptosis (Tobin et al., 1998).
It would be easier to model the stemcell system if it were only unidirec-tional. However, the capacity for a TAcell to become a quiescent stem cellsuggests that some degree of dediffer-entiation is possible. A dramatic exam-ple of the potential for a melanocyteto dedifferentiate in vitro has beendemonstrated in the quail system(Real et al., 2006; Dupin et al., 2007).Quail skin melanocytes have been
shown to give rise to Sox-10, Pax3,and slug-expressing stem cells. Further-more, the stem cells were pluripotent,that is they had the capacity to give riseto glia, myofibroblasts, and melano-cytes. Because stem cells are present inthe skin, the extent to which melano-cyte dedifferentiation occurs in vivo isunclear but clearly the potential exists.
Stem cells are present in normalskin. A number of groups have focusedon epidermal stem cells and their rolein giving rise to keratinocytes (Bick-enbach and Grinnell, 2004; Blanpainand Fuchs, 2006; Cotsarelis, 2006). Theseven groups presented below haveexamined skin stem cell populationscapable of producing melanocytes and/or other neural crest derivatives.
One group isolated a cell populationof small size and low Hoechst 33342staining from green fluorescence pro-tein (GFP)-labeled murine epidermisand injected the GFPþ cells intoblastocysts. They were able to showthat the GFPþ cells gave rise toectodermal, mesenchymal, and neuralcrest-derived tissues (Liang and Bick-enbach, 2002).
A second group isolated cells fromhuman hair follicles in conditionedhuman embryonic stem cell media(Yu et al., 2006). These ‘‘hair folliclestem cells’’ were noted to expressnestin, slug, snail, twist, sox-9, andbmp4. Focal Nanog and Oct4, a toti-potent embryonic stem and germ cellsmarker, expressions were also noted.This group was able to differentiatethese cells into melanocytes, neuronal,and smooth muscle cells.
A third group using a nestin-GFPconstruct was able to isolate cells fromthe bulge area of the murine hairfollicle (Amoh et al., 2005). The GFPþcells were found to be CD34þ andkeratin 15�. They were able to differ-entiate these cells into keratinocytes,melanocytes, neurons, glia, andsmooth muscle cells. In vivo, theformation of blood vessels and neuraltissue was shown. The ability to gen-erate melanocytes and keratinocytesfrom the same bulge stem cell popula-tion is quite intriguing.
A fourth group identified sphere-forming cells from skin culture (theepidermis and the dermis) that were
Figure 1. Repigmentation during recovery from vitiligo. During the active phase of vitiligo, cutaneous
melanocytes are destroyed. During recovery, as seen in this clinical image of a patient’s leg (a), the
destroyed melanocytic cells can be replaced. These new melanocytes appear to be derived from a
protected population of cells within the hair follicle as shown in these dermoscopic images (b and c) from
two patients recovering from vitiligo.
2368 Journal of Investigative Dermatology (2008), Volume 128
JM GrichnikMelanoma, Nevogenesis, and Stem Cell Biology

noted to express P75 and Sox-10(Wong et al., 2006). These cells alsoproduced multiple lineages but notkeratinocytes. With the addition ofSCF and ET-3, they were able to inducethe formation of a few melanocytes. Itwas noted that the skin from the faceproduced spheres with more potential,whereas spheres formed from the trunkskin grew less copiously and weremore restricted in their cell productionyielding only glial and melanocyticlineages. A further experiment utilizeda murine line with GFP expression incells in which the DCT promoterwas active. These mice made GFPþspheres that lacked pigmented melano-cytes, suggesting that the DCT promo-ter was active in p75 and Sox-10-positive stem cells.
A fifth group has isolated multipotentstem cells that they term ‘‘skin-derivedprecursors’’ (SKPs) from the rodent skin(Toma et al., 2001). The SKPs werederived from the dermis, but could notbe derived from the epidermis, andwere found to express nestin andfibronectin but not vimentin or cytoker-atin. The SKPs were shown to differ-entiate into neurons, glia, smoothmuscle, and adipocytes. A similar po-pulation of cells from the adult humanscalp skin was also derived (Toma et al.,2001). Further work on these SKPs hasrevealed that they also express snail/slug, twist, and Pax3 (Fernandes et al.,2004). Based on localization ofexpressed markers, they suggest a majorniche for the SKPs is in the dermalpapilla rather than in the hair bulge.
A sixth group attempted todetermine the molecular signature of‘‘neural crest epidermal stem cells’’(EPI-NCSC) isolated from the hair bulgearea (Hu et al., 2006). Nestin proteinexpression was present in 2- to 7-dayculture with KIT expression present atday 5. At day 7, autonomic neurons,Schwann cells, melanocytes, bone/cartilage cells, and myofibroblasts couldbe identified. EPI-NCSCs were definedas the cells emigrating from hair bulgemurine whisker while in culture. Thegene pattern of the EPI-NCSC cells (day2 in culture) was shown to be differentthan that of the epidermal stem cells(Tumbar et al., 2004) and the stem cellsthat may be increased in the dermal
papilla (Toma et al., 2001; Fernandeset al., 2004). Their work suggests atleast three different stem cell popula-tions may be present in the skin.
A seventh group has identifiedCD133þ (also Thy-1þ and CD34 low)cells in human skin. These multipotentstem cells have the capacity to differ-entiate into neurons, astrocytes, andoligodendrocytes (Belicchi et al.,2004). The CD133þ cells constituteabout 6% fetal skin cells but are onlyabout 1% of adult cells. Cell prolifera-tion and plasticity was more attenuatedin adult cells than in the fetal counterparts. These cells expressed c-kit andCD34 (by reverse transcription-PCR)(Belicchi et al., 2004). CD133þ cellshave also been previously noted in thehuman epidermis (Yu et al., 2002) andin a subpopulation of cells in cultureshuman foreskin melanocytes (Franket al., 2003).
The CD133þ cells are particularlyinteresting, as they have been noted toidentify neural crest stem cells (Uchidaet al., 2000), endothelia precursor cells(Gehling et al., 2000), and mast cellprecursor cells (Dahl et al., 2002). Inbone marrow transplantation experi-ments, the precursors for mast cellshave been noted to insert into the hairfollicle (Kumamoto et al., 2003).Although difficult to comprehend givenour current models, it is possible thatboth melanocytes and mast cells arereplenished from a common circulatingCD133þ precursor. Furthermore, it ispossible that these are the cells respon-sible for the spontaneous repigmenta-tion of gray hair.
Thus, the skin contains cells withstem cell characteristics, and many ofthese defined populations have theproven capacity to give rise to melano-cytes and/or at neural crest lineage cellpopulations. Although it could beargued that, in some of these experi-ments, the cells dedifferentiated from amore mature precursor, clearly inothers, the stem cells expressed mar-kers defining them as present prior toculture manipulation. The interrelation-ships between these different stem cellpopulations, their differentiation statein tissue, and the extent to which thedifferences experimental techniquesmodify the cells have yet to be fully
defined. However, it is clear thatthere are several potential stem cellpopulations in human skin with thecapacity to give rise to melanocytes. Itis even possible that melanocytes couldbe replenished from a circulatingCD133þ pluripotent precursor.
Nevogenesis
Origin and direction of melanocyticnevus growth. Nevi are benign clonalproliferations of cells expressing themelanocytic phenotype (Robinsonet al., 1998; Hui et al., 2001). Incongenital nevi, it is presumed that aprecursor cell is mutated, possibly bythe activation of N-Ras (Bauer et al.,2007), and an excessive number ofdaughter cells result. The migratingcells populate subcutaneous, dermal,and epidermal structures, as they at-tempt to complete normal migration tothe skin surface. Some of the excessmelanocytes completing migration maybe discharged through the epidermis as‘‘pagetoid’’ cells in early congenitalnevi (Figure 2). Transepidermal elimi-nation of nevus cells may also play arole in nevus involution (Kantor andWheeland, 1987). The nevus cells inthe epidermis or in the superficialdermis are more likely to be largerand express melanocytic differentiationantigens compared with the deepercells consistent with developmentalmodels. A similar antigen expressionpattern is seen in acquired nevi.
Although there is consensus thatcongenital nevi grow along develop-mental pathways toward the epidermis,for acquired nevi there is an activedebate as to the direction of migration.Unna described the process of Abtrop-fung, the dropping off of melanocyticcells into the dermis from the epider-mis; whereas Cramer (1991) has pro-posed the process of Hochsteigerung,migration toward the epidermis. Heproposes that the precursor cells areassociated with nerve tips and thatthere are four discrete migration/differ-entiation stages that may be involvedin normal melanocytic turnover, nevus,and melanoma formation (Cramer,1984).
One of the ways to determine thedirection of growth is to evaluate nevias they develop. The majority of the
www.jidonline.org 2369
JM GrichnikMelanoma, Nevogenesis, and Stem Cell Biology

nevi removed from children clearlyinclude a dermal component (Worretand Burgdorf, 1998). Dermoscopically,nevi developing in children often havea globular pattern, which is also mostconsistent with nevi that include adermal component (Zalaudek et al.,2006). Thus, if these nevi do not evolvein the dermis, dermal migration from
the epidermis must be a very earlyprocess.
In our own studies on early growingmelanocytic neoplasms, utilizing onlythe residual tissue in the block, anobvious dermal component was detect-able in 58% of the lesions, Ki67þ cellswere rare but could be identified in thedermal component, and upwardly
mobile ‘‘pagetoid’’ cells could beidentified in 32% of the growinglesions (Molino et al., 2003). Largejunctional nests (475% of the epider-mal thickness) were present in 42% ofthe lesions, and some of these nestswere clearly being expelled throughthe epidermis (Figure 3). These findingswere consistent with dermal originand epidermal migration. Given thepresence of pagetoid cells in growingnevi, we need to be careful not tooverinterpret growing benign nevi asmelanoma. Ever improving surfacemicroscopic technologies, such as con-focal, will be able to give us a clearerpicture as to the features to expectduring normal nevus growth.
The direction of melanocyte migra-tion has been evaluated in skin grafts.It has been demonstrated that KIT-activated melanocytic cells injectedinto the dermis of a graft migrate intothe epidermis (Alexeev and Yoon,2006; Grichnik, 2006b). This ability ofmelanocytic cells to migrate could alsoexplain how nevus cells washed into alymph node could migrate into thelymph node’s capsule. Although devel-opmentally this direction of migrationmakes sense, it is important to point outthat experiments with transformed cul-tured melanocytes have demonstratedthat these cells can penetrate into thedermal component of the graft (Chud-novsky et al., 2005); therefore, migra-tion from the epidermis into the dermisis also possible.
The direction of melanocytic migra-tion during nevus development willeventually be definitively determinedthrough sequential imaging. Confocalmicroscopy and optical coherence to-mography are likely to play criticalroles in defining this process. Dermo-scopy may also provide informationbased on color features. Owing to lightscattering, blue coloration representsdermal melanin. Both decreasing bluefeatures suggesting upward growthand increasing blue features suggestingdownward growth have been noted(Pizzichetta et al., 2006; Zalaudeket al., 2007). However, because ame-lanotic cells in the dermis couldbecome more pigmented as they ma-ture, increasing blue pigmentation doesnot necessarily reflect migration into
a b
dc
Figure 3. Growing melanocytic neoplasms have features suggesting epidermal tumor loss and dermal
proliferation. Growing melanocytic neoplasms can be detected clinically (a) by comparing the patient’s
lesion on a follow-up visit (lower left panel) with photographs from a prior visit (upper left panel).
This particular lesion was approximately 5 mm in longest diameter (inset) and was pathologically
interpreted as an atypical (dysplastic) nevus with severe atypia. In growing melanocytic neoplasms,
melanocytic cells could be detected in the upper layers of the epidermis and in the stratum corneum
(b, arrows, size bar¼ 100mm). In other lesions, melanocytic nests of cells could be identified in the
stratum corneum (c, arrow, size bar¼ 100mm) and proliferating dermal cells could be identified with
immunohistochemical staining (red) for Ki67 (d, arrow, size bar¼ 200mm, 50 mm (inset)).
Figure 2. Pagetoid melanocytic cells in congenital nevus. Photomicrograph of hematoxylin and
eosin-stained section of a congenital melanocytic nevus excised from the left buttock of a 10-month-old
female child. Two pagetoid melanocytes are clearly visualized in the mid-spinous and granular layers
of the epidermis (black arrows).
2370 Journal of Investigative Dermatology (2008), Volume 128
JM GrichnikMelanoma, Nevogenesis, and Stem Cell Biology

the dermis, and thus further studies willbe required. Beyond just direction,these imaging techniques will allowfurther study into the melanocyticdevelopmental process. There aremany different patterns of nevi, andtheoretically, the growth pattern shouldbe a reflection of the underlying muta-tion and the local environmental influ-ences on cell growth and survival(Figure 4). B-Raf and N-Ras mutationshave been associated with acquirednevi but specific dermoscopic patternshave yet to be delineated.
B-Raf-activating mutations are com-monly seen in acquired nevi and N-Rasmutations are commonly seen in con-genital nevi (Ichii-Nakato et al., 2006;Bauer et al., 2007). N-Ras not onlyactivates a number of pathways, inclu-ding phosphoinositide 3 kinase andRal, but also activates B-Raf, leadingto a common activation of downstreammitogen activated protein kinase andERK pathways (Arbiser, 2003), redu-
cing the need for stimulatory cytokines.In culture, nevus cells are also lessdependent on growth factors whencompared with normal melanocytes(Mancianti et al., 1993; Alanko et al.,1999). This suggests that although theunderlying mutations are not sufficientto drive malignancy, they are sufficientto allow some independence from thenormal homeostatic mechanisms.
Thus, nevi develop due to mutationsin a precursor cell that activate prolif-erative pathways, or suppress apoptoticpathways, allowing for the accumula-tion of melanocytic cells in the skin.The specific pattern of this accumula-tion is anticipated to be a reflection ofthe underlying mutation in the precur-sor cell. The direction of nevus growthis still in debate; however, similar tocongenital nevi, it is reasonable toassume that acquired melanocytic nevi(with a dermal component) developfrom a mutant dermal precursor andmigrate and differentiate along normal
melanocytic developmental pathwaystoward the epidermis. Once in theepidermis, there may be an increasedproliferative response due to localgrowth factors and excess melano-cytic cells would be shed through thestratum corneum.
Melanoma
Role of nevi in melanoma develop-ment. The development of melanomahas been modeled as a stepwise processfrom a cutaneous melanocyte throughnevus and dysplastic nevus stages to insitu and eventually invasive melanoma.However, approximately three-fourths ofmelanomas develop in normal skin andless than one-half of the nevi associatedwith melanoma are dysplastic (Bevonaet al., 2003). Thus, it is not clear that adysplastic nevus is really any more likelyto develop a melanoma than any othertype of nevus, and further it is quite clearthat a nevus precursor is not required forthe majority of melanomas. This phe-
CongenitalLentigo
Reed
Combined blue
Blue
Melanocyticstem cell
Common/dysplastic
Spitz
Der
mis
Epi
derm
is
Figure 4. Melanocytic neoplasm growth patterns. Melanocytic neoplasm demonstrate numerous different repeatable patterns on patient’s skin surface.
Presumably, each of distinct patterns represents an underlying distinct mutation, developmental phase, or local signaling environment. Repeatable distinct
patterns can be noted for acquired nevi (common and dysplastic), Spitz nevi, ‘‘congenital’’ nevi, lentigines, Reed nevi, blue nevi, and combined blue nevi.
Furthermore, distinct patterns can be noted for different melanoma types, indicated here in the red boxes, superficial spreading, lentigo maligna, acral lentiginous,
and others. Theoretically, these lesions could be derived from a common precursor (brown oval) that follows normal developmental pathways to the epidermis.
Dependent on the mutation and local environmental factors, the neoplasm would expand and migrate in the dermal and the epidermal compartments.
www.jidonline.org 2371
JM GrichnikMelanoma, Nevogenesis, and Stem Cell Biology

nomenon is difficult to explain utilizingcurrent dogma. However, it is relativelystraightforward to explain based on thestem cell theory. Melanoma could de-velop directly from a quiescent precur-sor cell that had accumulated amalignant complement of mutations.The one-fourth of melanomas associatedwith nevi may be due to a secon-dary mutation developing in a growingstem cell population giving rise to abenign nevus.
Increased mole burden clearly in-creases melanoma risk. For patientswith large congenital nevi, the mela-noma risk appears to be correlated withthe volume of the nevus and malignanttransformation often appears to occurwithin the nevus at a young age(Krengel et al., 2006). For patients withnumerous acquired nevi, the bulk ofthe risk may be in the otherwise normalappearing skin. It is not clear whatdrives this risk but there are a numberof possibilities. First, the patient’s nevusprecursor (stem) cells may be increasedin number, have had increased expo-sure to mutagenic events, or increasedsensitivity to mutagenic events. Second,the patient may have less effectiveimmunosurveillance (or other mechan-isms) to inhibit the growth of melano-mas and/or nevi from the mutantprecursor cells. Finally, the precursorcells may have an inherent increase inself-renewal capacity, leading to anincreased window of time for muta-tions to occur. Interestingly, it has beennoted that the telomeres are longer inthe white blood cells of patients withnumerous nevi (Bataille et al., 2007),suggesting increased longevity andproliferative potential.
Another interesting issue is differ-ence in age for nevus versus melanomadevelopment. Most nevi develop dur-ing late childhood and early adulthood,whereas melanoma incidence con-tinues to increase in the later years oflife (Banky et al., 2005). It is not clearwhat drives this phenomenon but thereare a number of possibilities. First, thelonger the period the precursor (stem)cell remains quiescent, the more themutations it may accumulate, resultingin a greater chance for malignancy.Second, mutant stem cells with anti-apoptotic mechanisms may be retained
and expanded with age. Additionalmutations in these cells would be morelikely to create a malignancy. Third,melanomas that are slow growing maysimply be easier to detect at older ages.Fourth, as the patient ages, they maylose effective immunosurveillance (orother mechanism) for inhibiting thegrowth of melanoma. Finally, it ispossible that somehow the body re-cruits normal stem cells first and onlylater relies on the mutated stem cells.
Thus, there is a clear association ofnevi with melanoma risk. Some of thisrisk is due to the potential for second-ary mutations within nevi. However,the majority of the risk may be due tothe inherent properties of the stem cellpopulation in individuals with numer-ous moles.
Origin and direction of malignantmelanoma growth. Similar to acquirednevi, the traditional model suggests thatmelanomas initially develop in theepidermis and then invade the dermis.This is largely based on pathologicfeatures in which the lowest risk in situmelanoma tumors are noted to bepresent entirely in the epidermis,whereas high-risk tumors are present indeeper dermal tissues. Thus, it is reas-onable to assume based on progressionmodels that the tumor must first arise inthe epidermis. These models also sug-gest that the cell of origin for the tumoris the epidermal melanocyte. This isalso supported by research that revealsthat melanocytes can be transformedwith oncogenes and acquire malignant/invasive characteristics (Chudnovskyet al., 2005). Furthermore, the tyrosi-nase promoter, expressed in melano-cytes, has been shown to inducemelanoma when driving oncogenicproteins SV40E (Kelsall and Mintz,1998) and N-Ras (Wong and Chin,2000). Thus, it is possible to generatea melanoma from a cell expressingmelanocytic proteins. The extent towhich these melanocytes are fullydifferentiated and the degree of plasti-city maintained in the cell undergoingthe transformation event have yet to befully clarified.
The stem cell model is essentially theconverse of the traditional model.Although it does not rule out epidermal
evolution of melanoma, which couldvery well be the case especially forlentigo maligna, it favors migration tothe epidermis from a dermal precursor.In this model, low-risk, in situ or thininvasive, melanomas would be highlyattracted to the epidermis and migratetoward this location and thus minimizingspread from the tissue. As additionalmutations accumulated in the tumor(due to genomic instability), the melano-ma cells would become less growthfactor-dependent and gain an increasedcapacity to grow in deeper levels of thedermis and other tissues. It is importantto note that on review of in situtumors, 29% were found to have anintradermal component (Megahed et al.,2002), so it is not entirely inconceivablethat many of these tumors have a dermalderivation. As the dermal TSC accumu-lated more mutations, the capacity toproduce cells that could continue todifferentiate along normal melanocyticpathways would be reduced, giving theimpression of dedifferentiation whenactually it would be due to the unmask-ing of the immature malignant stem cellpopulation.
Although early melanomas are diffi-cult to grow in culture, a significantproportion of metastatic melanomascan grow readily in culture withoutthe need to add exogenous growthfactors (Halaban et al., 1986). A fewmelanomas do exist that have KIT-activating mutations and, therefore,may be sensitive to KIT-inhibitingagents (Curtin et al., 2006), but mosthave downregulated this pathway pre-sumably because its homeostatic rolecould be detrimental to the viability ofthe tumor. Inhibition of the ET-3/endothelin B receptor pathway hasbeen reported to inhibit the growth ofmelanoma lines (Lahav et al., 1999).However, for the most part, melanomasare largely growth factor-independent,presumably due to mutations, such asN-Ras activation or B-Raf/PTEN, thatpromote proliferation and survival(Tsao et al., 2004) or from autocrinesignaling loops due to activation ofpathways leading to the production oftheir own growth factors. This inde-pendence from homeostatic pathwaysallows the tumors to thrive in non-epidermal environments.
2372 Journal of Investigative Dermatology (2008), Volume 128
JM GrichnikMelanoma, Nevogenesis, and Stem Cell Biology

Thus, at this point, whether melano-mas develop in the epidermis andmigrate into the dermis or develop inthe dermis with migration to theepidermis is still a matter of debate.However, in either scenario, the deeperthe tumor in the dermal tissue, thelower the requirement for epidermalfactors and the greater the risk thattumor will be able to proliferate outsideof the skin surface and be potentiallydeadly.
Stem cell/developmental behavior.Regardless of origin, it is clear that asubpopulation of TSCs exist in melano-mas (Figure 5). Studies on metastaticmelanoma lines have revealed that thecultures are heterogeneous even whenderived from a single cell (Grichniket al., 2006). The different cell popula-tions within these cultures appear tohave diverse characteristics. A small-cell phenotype has been noted thatappears to enter the cell cycle at alower rate but has an increased capa-city to expand the culture. The smallcells had the capacity to give riseto larger, more proliferative cells(TA cells). The TA cells then eventuallygave rise to still larger, more melanizedcells. Furthermore, some of thelarger, more melanized cells appearedto have terminally differentiated andwere eventually lost from culture.These experiments suggested a generalforward developmental flow of theprocess. However, even after doublecell flow purification of the larger cellforms, some small cell forms reap-peared in culture. This could havebeen due to incomplete purification,but it could also be due to thepossibility that some of the TA cellswere able to re-occupy the stem cellniche, reverting to a small-cell pheno-type. Thus, this suggests that there mayalso be a degree of backward flowin the culture system to achieve theappropriate embryonic developmentalbalance. It is also important to note thatthe small cell forms were very weaklyadherent and were readily releasedfrom the plate with simple mediaremoval. Despite being relatively freefloating, these cells were very effectiveat establishing new cultures. Tumorspheroids reminiscent of those
described for stem cells also readilydeveloped under dense culture condi-tions. A small subpopulation of nestin(pluripotent stem cell marker)-positivecells could be identified in densemelanoma cultures, suggesting the pre-sence of a stem cell phenotype.
Other groups have also identifiedpotential TSC markers. Fang et al.(2005) noted that non-adherent spher-oids developed in approximately 20%of melanomas cultured in ESC media.Cells from these spheroids could bedifferentiated into melanocytic, adipo-cytic, osteocytic, and chondrocyticlineages. Compared with the adherentcells, the spheroid cells were moretumorgenic in animal models. Thenon-adherent spheroid cells were foundto include CD20 B-cell markers. Thesorted CD20 fraction were found to bemore spheroidogenic (more likely tomake sphere or make larger spheres),and had more potential for mesenchy-mal differentiation. The CD20 markerwas also noted to be present in approxi-mately 20% of human metastatic mela-nomas by immunohistochemisty.
A second group has identifiedABCB5 in melanomas (tissue sampleand in cell lines) as preferentiallymarking a subset of CD133þ -expres-sing cells in melanomas (Frank et al.,2005). The ABCB5þ cells were alsoCD166þ . Inhibition of the ABCB5
pump significantly reversed resistanceto doxorubicin. Cells double labelingfor ABCB5 and CD133 were approxi-mately 2% of the G3361 line. Thesemarkers also stained melanoma tissuesamples (positive in all tissue sam-ples—overall staining in each sampleo50% six tumors examined).
A third group has reported thatCD166, CD133, and nestin are immu-nohistochemically expressed at signi-ficantly higher levels in melanomacompared with banal nevi (Kleinet al., 2007). Further, nestin was foundto be significantly increased in meta-static melanoma compared with pri-mary melanoma.
A fourth group has specifically iso-lated CD133þ cells from human mel-anomas (Monzani et al., 2007). Thisgroup noted that the CD133þ cellswere present at levels ranging from 0.2to 0.8% of the total cells in sevenmelanoma specimens. The CD133þmelanoma cells all grew tumors inNOD-SCID mice whereas CD133�melanoma cells failed to grow.
Melanoma cultures are quite dyna-mic and antigen expression can beerratic. In our hands, the greatestdegree of heterogeneity exists in long-term dense cultures with partial butfrequent media changes (Grichniket al., 2006). We have noted waves ofpatterning between nestin and KIT
a b
c d
Figure 5. Metastatic melanomas include cells with stem cell-like features. Some melanoma cultures
display a heterogeneous population of tumor cells, including small weakly adherent cells (a, line DM3N)
and larger adherent phenotypes (b, line DM3N). Although some of the larger cell phenotypes (b) do not
appear to be significantly proliferative, low-confluence cultures of the small weakly adherent cells
(a) have the capacity to perpetuate heterogeneous colonies in culture (c, line DM3N) while maintaining
a subpopulation of small weakly adherent cells (arrow). Melanoma line DM1N (d) plated at low
confluence also give rise to heterogeneous colonies after transfer as well as maintaining a subpopulation
of small weakly adherent cells (arrows). Magnification is the same for all photos; size bar¼200 mm.
www.jidonline.org 2373
JM GrichnikMelanoma, Nevogenesis, and Stem Cell Biology
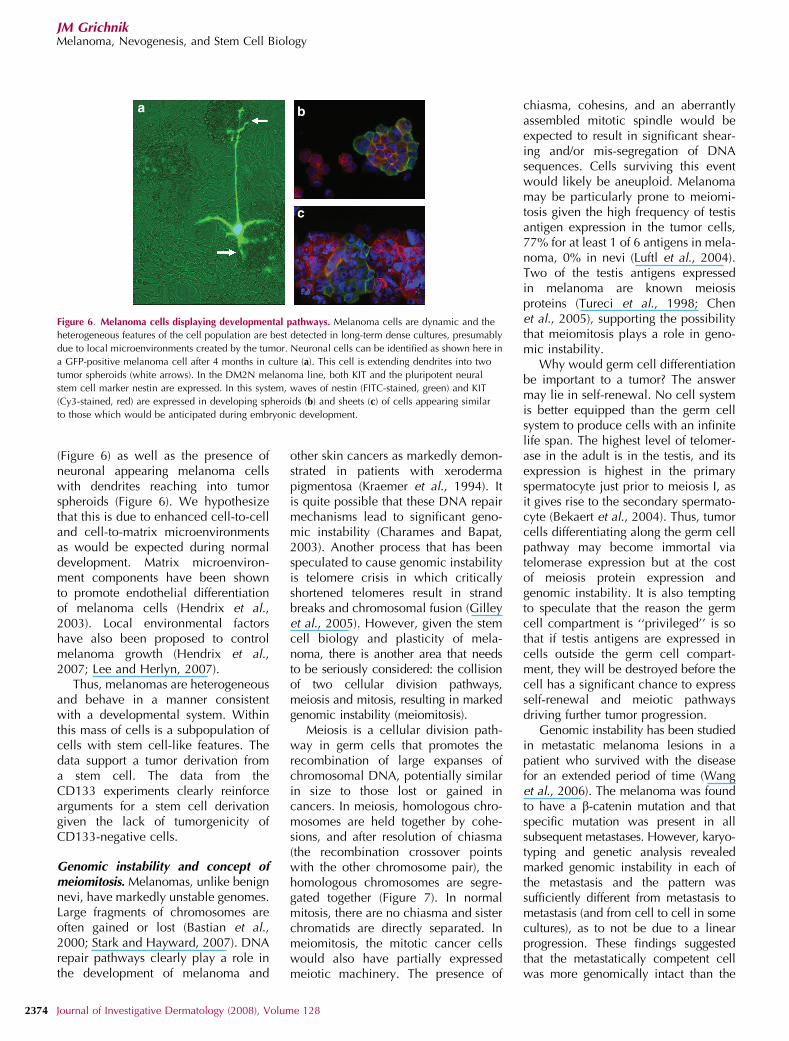
(Figure 6) as well as the presence ofneuronal appearing melanoma cellswith dendrites reaching into tumorspheroids (Figure 6). We hypothesizethat this is due to enhanced cell-to-celland cell-to-matrix microenvironmentsas would be expected during normaldevelopment. Matrix microenviron-ment components have been shownto promote endothelial differentiationof melanoma cells (Hendrix et al.,2003). Local environmental factorshave also been proposed to controlmelanoma growth (Hendrix et al.,2007; Lee and Herlyn, 2007).
Thus, melanomas are heterogeneousand behave in a manner consistentwith a developmental system. Withinthis mass of cells is a subpopulation ofcells with stem cell-like features. Thedata support a tumor derivation froma stem cell. The data from theCD133 experiments clearly reinforcearguments for a stem cell derivationgiven the lack of tumorgenicity ofCD133-negative cells.
Genomic instability and concept ofmeiomitosis. Melanomas, unlike benignnevi, have markedly unstable genomes.Large fragments of chromosomes areoften gained or lost (Bastian et al.,2000; Stark and Hayward, 2007). DNArepair pathways clearly play a role inthe development of melanoma and
other skin cancers as markedly demon-strated in patients with xerodermapigmentosa (Kraemer et al., 1994). Itis quite possible that these DNA repairmechanisms lead to significant geno-mic instability (Charames and Bapat,2003). Another process that has beenspeculated to cause genomic instabilityis telomere crisis in which criticallyshortened telomeres result in strandbreaks and chromosomal fusion (Gilleyet al., 2005). However, given the stemcell biology and plasticity of mela-noma, there is another area that needsto be seriously considered: the collisionof two cellular division pathways,meiosis and mitosis, resulting in markedgenomic instability (meiomitosis).
Meiosis is a cellular division path-way in germ cells that promotes therecombination of large expanses ofchromosomal DNA, potentially similarin size to those lost or gained incancers. In meiosis, homologous chro-mosomes are held together by cohe-sions, and after resolution of chiasma(the recombination crossover pointswith the other chromosome pair), thehomologous chromosomes are segre-gated together (Figure 7). In normalmitosis, there are no chiasma and sisterchromatids are directly separated. Inmeiomitosis, the mitotic cancer cellswould also have partially expressedmeiotic machinery. The presence of
chiasma, cohesins, and an aberrantlyassembled mitotic spindle would beexpected to result in significant shear-ing and/or mis-segregation of DNAsequences. Cells surviving this eventwould likely be aneuploid. Melanomamay be particularly prone to meiomi-tosis given the high frequency of testisantigen expression in the tumor cells,77% for at least 1 of 6 antigens in mela-noma, 0% in nevi (Luftl et al., 2004).Two of the testis antigens expressedin melanoma are known meiosisproteins (Tureci et al., 1998; Chenet al., 2005), supporting the possibilitythat meiomitosis plays a role in geno-mic instability.
Why would germ cell differentiationbe important to a tumor? The answermay lie in self-renewal. No cell systemis better equipped than the germ cellsystem to produce cells with an infinitelife span. The highest level of telomer-ase in the adult is in the testis, and itsexpression is highest in the primaryspermatocyte just prior to meiosis I, asit gives rise to the secondary spermato-cyte (Bekaert et al., 2004). Thus, tumorcells differentiating along the germ cellpathway may become immortal viatelomerase expression but at the costof meiosis protein expression andgenomic instability. It is also temptingto speculate that the reason the germcell compartment is ‘‘privileged’’ is sothat if testis antigens are expressed incells outside the germ cell compart-ment, they will be destroyed before thecell has a significant chance to expressself-renewal and meiotic pathwaysdriving further tumor progression.
Genomic instability has been studiedin metastatic melanoma lesions in apatient who survived with the diseasefor an extended period of time (Wanget al., 2006). The melanoma was foundto have a b-catenin mutation and thatspecific mutation was present in allsubsequent metastases. However, karyo-typing and genetic analysis revealedmarked genomic instability in each ofthe metastasis and the pattern wassufficiently different from metastasis tometastasis (and from cell to cell in somecultures), as to not be due to a linearprogression. These findings suggestedthat the metastatically competent cellwas more genomically intact than the
a b
c
Figure 6. Melanoma cells displaying developmental pathways. Melanoma cells are dynamic and the
heterogeneous features of the cell population are best detected in long-term dense cultures, presumably
due to local microenvironments created by the tumor. Neuronal cells can be identified as shown here in
a GFP-positive melanoma cell after 4 months in culture (a). This cell is extending dendrites into two
tumor spheroids (white arrows). In the DM2N melanoma line, both KIT and the pluripotent neural
stem cell marker nestin are expressed. In this system, waves of nestin (FITC-stained, green) and KIT
(Cy3-stained, red) are expressed in developing spheroids (b) and sheets (c) of cells appearing similar
to those which would be anticipated during embryonic development.
2374 Journal of Investigative Dermatology (2008), Volume 128
JM GrichnikMelanoma, Nevogenesis, and Stem Cell Biology

rest of the locally produced tumorbulk (Grichnik, 2006a). Thus, ongoingmarked genomic instability might hin-der the capacity of daughter cells tosuccessfully metastasize.
The stem cell characteristics of mel-anoma and the expression of testisantigens should lead us to consider thepotential role of germ cell differentiationin enhancing self-renewal while creat-ing genomic instability through meio-mitosis. Given the destruction wreakedupon the cell with genomic instability,only a subpopulation of cells mayremain metastatically competent.
Melanoma stem cell progression model.Melanoma progression and metastasisis traditionally modeled as a stepwiseprocess with the initial mutagenicevent occurring in a melanocyte inthe epidermis, with further mutationresulting in the proliferation passingthrough nevus and dysplastic nevusphases. The malignant cells eventuallyacquire the ability to digest their waythrough the basement membrane. Thenwith additional mutations, the cellsacquire a migratory form and even-tually physically penetrate the lympha-tic vessel walls and travel to the locallymph node where they remain trappeduntil they expand sufficiently to popu-late the next node in the chain.
There are a number of questions thetraditional model cannot readily explain.These include: (1) If dermal invasion is asecond malignant step in melanomadevelopment, how do we write off thefact that benign nevi readily ‘‘invade’’the dermis? (2) If precursor nevus anddysplastic nevus stages are critical, thenwhy do 75% of melanomas arise innormal skin? (3) How can we explainthe fact that benign nevus cells can alsobe found in lymph nodes? (4) If thelymph node traps the tumor cells, whydoes sentinel lymph biopsy and regionallymph node dissection fail to improvesurvival? (5) Why despite an exuberantimmunologic response to the primarymelanoma or to the vaccine therapydoes the immune system fail to eliminatemetastatic disease? And finally (6) Howcan an aggressive tumor remain dormantfor decades and then reoccur?
The stem cell model (Figure 8) morereadily addresses many of these issues.In contrast to the traditional theory ofmelanoma derivation from a differen-tiated melanocyte, the stem cell modelsuggests that the initial mutations accu-mulate in a quiescent stem cell. Even-tually, environmental signaling wouldactivate the stem cell, but due to themutations, proliferation would not beappropriately controlled. Benign tu-mors would be growth restricted
whereas malignant tumors would not.Secondary mutations could occur inthe expanding benign cells creating thedevelopment of a melanoma within anevus, but the nevus step would not berequired. The neoplastic cells producedwould attempt to follow normal mela-nocytic differentiation pathways in-cluding migration into the epidermis.In the epidermis, local growth factorswould likely drive further proliferation,and the excess tumor cells would bedischarged through the epidermis intothe stratum corneum. The stem cellcomponent in the dermis would con-tinue to expand, and some TSCs wouldinappropriately express germ cell path-ways not only allowing for increasedself-renewal but also driving genomicinstability due to meiomitosis. In themalignant tumors, this would result indifferent subclones of cells and woulddisplay the potentially diagnostic non-uniform surface features (Lucas et al.,2003). Although lymph node metastasisis often modeled as an active process, itis important to recall that carbon andtattoo ink can readily pass from the skinto the lymph node. Some of the dermalcells from both the benign and themalignant tumors could be washed intothe lymph system. Thus, although moreadherent melanoma cells would bind inthe lymph node and possibly migratetoward the capsule, the weakly adhe-rent TSCs may pass straight throughthe nodal chain and enter the circulatorysystem. The small quiescent TSCs wouldbe overlooked by the immune systemand could continue to circulate longterm, but eventually would reinsert intissues in accordance with normal stemcell regenerative pathways. These TSCswould again activate when the environ-mental signals where favorable and theprocess would continue.
Treatment
Regardless of the model applied, it isclear that melanomas are heteroge-neous and include a distinct subpopu-lation of TSCs. These TSCs may bedifficult to target with conventionaltreatments, as they may lack significantproliferation decreasing the effective-ness of anti-proliferative agents, lacksignificant antigen expression inhibit-ing vaccine efforts, and may not be
Mitosis Meiosis Meiomitosis
Figure 7. Meiomitosis. During normal mitosis sister chromatids are separated into different cells. During
meiosis, homologous chromosomes are held together by cohesions (purple lines) and are segregated
together after resolution of chiasma (recombination crossover points) with the other chromosome pair.
A second cycle separates the sister chromatids. The term ‘‘meiomitosis’’ is used for mitotic cells, in which
meiotic machinery is partially expressed. In these cells, chiasma occur and sister chromatids are
partially linked together with cohesions. During cell division, the attempt to separate the sister
chromatids results in shearing and/or mis-segregation of DNA sequences and creation of an
aneuploid state.
www.jidonline.org 2375
JM GrichnikMelanoma, Nevogenesis, and Stem Cell Biology

dependent on a targetable mutatedpathway while in a quiescent state. Inaddition, even if the TSCs were suscep-tible to a particular agent, they mayhave an enhanced ability to pump theagent out of the cell minimizing itseffect (as noted with ATP bindingcassette transporters—Frank et al.,2005). Furthermore, even if it waspossible to fully eradicate the TSCs, itis also possible that some of the TA cellscould revert to TSCs, similar to thatnoted in the murine melanocytic stemcell experiments (Nishimura et al.,2002). Thus, treatment of this hetero-geneous population of melanoma cellswill be quite a challenge, requiringeradication of the TSC subpopulationas well as any cells capable of givingrise to more TSCs. For the purpose of
discussion, we will consider the TAcells, the only population with thepotential capacity to revert to a TSCstate, the term TSC/TAs will be used todescribe the tumorigenic cell populationwithin the entire heterogeneous popula-tion of melanoma cells.
There are a number of treatmentapproaches that may be considered forthe TSC/TAs. One approach would be toactivate the TSCs and differentiate theentire TSC/TA population to a chemo-therapy sensitive or non-proliferative celltype. A second approach would be atargeted multiantigenic approach againstthe TSC/TA population either by activat-ing the immune system or by utilizingtargeted reagent antibodies. It is possiblethat targeting the CD133þ melanomacells in a specific manner will destroy all
TSC/TAs, but it is likely that additionalapproaches will be required. A thirdapproach would be to target a specificmutation or aberrantly activated path-way within the TSC/TA population.However, this therapy might only killthe TAs in which it was active, and thisapproach might need to be coupled witha reagent activating the quiescent TSCsto make them susceptible. A fourthapproach would be to force all theTSC/TA cells into a quiescent state,preventing further development ofdownstream tumor. These patientswould require long-term suppressivetherapies.
Thus, TSCs have inherited a numberof protective properties that will makethem difficult to destroy. It is clearthat any potentially successful therapywill need to find creative methodsto eradicate the TSC subpopulation aswell as eliminating any cells capableof making TSCs (TAs).
CONCLUSIONStem cells function to replenish termi-nally differentiated cell populations inadult tissues. It is these stem cells thatare the most likely source for cancers.The ultimate stem cell, the fertilizedegg, can give rise to over a trillion cellscomprising numerous different cellpopulations. A cancer stem cell poten-tially has access to this same cellularmachinery. The pigmentary systemserves as an ideal model for the studyof developmental biology and the roleof stem cells in tumorigenesis. Adiscrete series of developmental expo-sures, in vivo or in vitro, can differen-tiate stem cells toward the melanocyticlineage. Certain mutations in stemcells, such as B-Raf activation, maydrive melanocytic differentiation with-out the need for external signalingpathways. Both benign and malignantmelanocytic tumors develop directlyon the skin surface allowing for theevaluation of early events. Through thestudy of melanoma and nevus devel-opment, it is likely we will be able tobetter understand the early events andthe role of stem cell biology in tumori-genesis.
Numerous studies have demon-strated the existence of TSCs withinhuman melanomas. Given the pre-
Melanocyticstem cell 9
8
76
5
43
2
1
Blo
odFa
tD
erm
isE
pide
rmis
Figure 8. Stem cell-based model of melanocytic neoplasia. In this model, stem cell lineage cells (1)
implant into the dermis (2). When necessary to replenish melanocytes in the epidermal compartment,
these cells (2) migrate and implant in the basal layer of the epidermis (3). The cells in the epidermis
proliferate and differentiate giving rise to mature melanocytes (4). Eventually, the mature melanocytes are
shed through the stratum corneum (5). Mutations could occur at any level, but for the tumors with a
dermal component, mutations would occur in the dermal cells (2). For tumors confined to the epidermis,
mutations could occur in the epidermal cells (3) or in a dermal cell (2) that completely migrated into the
epidermis. The mutated cells would attempt to follow normal differentiation pathways, including the
shedding of excess cells through the stratum corneum. Some mutations could also result in aberrant
dermal migration, including along nerves and blood vessels (6). Cells in the dermis could accumulate
additional mutations (7). If the initial stem cell included only mutation for a benign nevus, the additional
mutation could then result in a focus of melanoma within the nevus. If the original stem cell included a
complement of mutations to create a melanoma, then these additional mutations would create new
tumor subpopulations. Loose non-adherent cells (from benign or malignant process) could ‘‘wash’’ into
the lymph system (8). Some of these cells would stick in the lymph node and cells retaining migratory
pathways may migrate into the capsule. The non-adherent stem-like cells may pass through the lymph
node and circulate systemically (9) as a mutated stem-like cell (1). This cell could then under the
appropriate conditions re-enter the dermis (2) (or some other tissue) and reinitiate the process.
2376 Journal of Investigative Dermatology (2008), Volume 128
JM GrichnikMelanoma, Nevogenesis, and Stem Cell Biology

sence of TSC in melanomas and thepresence of stem cell populations inhuman skin, it is reasonable to assumethat the skin stem cells serve as the cellsof origin for melanoma development.However, the specific stem cell typeinvolved in the transformation processand its location in the skin remainsundefined. Although many of the stemcell population studies have focused onfollicular stem cells, most melanomasdo not appear to have a follicularorigin. Given the normal developmen-tal direction of migration, it seemslikely that invasive melanomas arebased on a stem cell within the dermis.Nevi would also be based on thisdermal stem cell, and existence of bluenevi (heavily pigmented melanocyticneoplasm that originates in the dermiswithout any epidermal or follicularinvolvement) also supports a derivationfrom a melanocytic stem cell present inthe dermis.
A promising marker for dermal stemcells is CD133. CD133þ cells havebeen shown to give rise to endothelialcells (Gehling et al., 2000), mast cells(Dahl et al., 2002), and neurons andglial cells (Uchida et al., 2000). Adipo-cytes, osteocytes, and chondrocyteshave also been generated (Tondreauet al., 2005). Although melanocyteshave not yet been reported to differen-tiate from this population, the presenceof CD133þ melanoma TSC is highlysuggestive. Therefore, it is possiblethat different mutations in a CD133þstem cell could result in different tumortypes, including malignant melanoma,benign nevi, hemangiomas, lipomas,neurofibromas, and mastocytomas.CD133þ cells have been noted to beincreased in circulation after trauma(Liu et al., 2007). It is also feasible thatmutant (possibly B-Raf) CD133þ stemcells could be mobilized from apatient’s bone marrow due to theirimmunosuppressed state, and presentin the skin as eruptive nevi. The factthat only the CD133þ melanoma cellsare tumorigenic (Monzani et al., 2007)supports the role of a stem cell-drivenprocess.
Although melanoma derivation froma stem cell seems likely, the potentialrole for dedifferentiation from a mela-nocyte cannot be ruled out. Support for
a dedifferentiation process includesthat fact that melanized quail cellscan dedifferentiate into multipotentstem cells (Real et al., 2006), onco-genes added to cultured melanocytescan create tumors (Chudnovsky et al.,2005), and genes driven from thetyrosinase promoter can give rise tomelanomas in transgenic mice(Kelsall and Mintz, 1998; Wong andChin, 2000). Certainly, developmentalmodels include plasticity, allowingneighboring cells to assume the func-tion of cells lost during development.Thus, normal developmental biologydoes allow for both ‘‘stem cell’’ and‘‘dedifferentiation/plasticity’’ mechan-isms. At this point, it is reasonableto assume that both mechanisms maybe at play to varying degrees inmelanoma.
Melanocytic tumors are probablybest viewed as a pliable developmentalsystem with a TSC component. Thesetumors may be best defined by fourbiologic parameters. The first para-meter is the nature of the critical path-way(s) that have been mutated in thetumor cells. These pathway(s) are likelyto define the direction of cellulardifferentiation as well as the regulationover the proliferative and apoptoticprocess. The second parameter is therange of cell’s differentiation/plasticitycapacity. The extent of this capacitymay be, in part, defined by the mutatedpathway(s), but it may also be definedby the state of differentiation (organiza-tion of the nuclear matrix) in the cell oforigin. Theoretically, tumor origin in astem cell would allow for a far greaterrange of differentiation/plasticity capa-city. This capacity would allow cells tobetter survive under different environ-mental conditions and allow for greaterdiversity in expression programs, suchas allowing germ cell pathway differen-tiation, which may be advantageousfor the expression of telomerase buthazardous due to the potential formeiomitosis. The third parameter isthe intactness of the genome. Increas-ing genomic instability will result inthe loss of primary DNA sequencedata. Surviving cells may benefitinitially from acceleration of certainpathways but at the same time willloose other potentially useful capabil-
ities. The fourth and final parameteris telomere length. As telomeres short-en, the cells ability to continue toreplicate is diminished. Thus, whenviewed as a pliable, but mutated,developmental system, it may be ne-cessary to consider these four differentfacets in defining the neoplasm’s ulti-mate behavior.
In summary, melanoma tumors,similar to other solid cancers, clearlyinclude a subpopulation of TSCs. Thesecells appear to have the unique capa-city to perpetuate the tumor as well asthe capacity to differentiate alongdifferent cellular developmental lines.The melanocytic system is a modelsystem for normal and abnormal adulttissue stem cell biology. The develop-ment of these tumors on the skinsurface and the advance of skin surfaceimaging technologies will help us tostudy the complexities of the oncologicprocess. The lessons learned will beable to be applied across many solidtumor types and will have ramificationsfor tumor development, diagnosis,prognosis, and treatment.
CONFLICT OF INTERESTFounder and major shareholder—Digital DermInc.—MoleMapCD (total body photography).Consulting and Grants—Electro-Optical SciencesInc.—Melafind (melanoma detection device) andSpectral Image Inc.—Dermatologic Diagnostics.
ACKNOWLEDGMENTSDivision of Dermatology and the Benefactors tothe Duke Comprehensive Cancer center, espe-cially Fred and Sharon Matt (in honor of JamesMatt), Carolyn Holding, Carmen Ames, andRebecca Hundley for their generous financialsupport, Harold Rabinovitz for his image of themelanoma with blue features, and KP Grichnik,KK Hipp, and JA Burch for their editorial review.
REFERENCES
Alanko T, Rosenberg M, Saksela O (1999) FGF
expression allows nevus cells to survive in
three-dimensional collagen gel under condi-
tions that induce apoptosis in normal human
melanocytes. J Invest Dermatol 113:111–6
Alexeev V, Yoon K (2006) Distinctive role of the
cKit receptor tyrosine kinase signaling in
mammalian melanocytes. J Invest Dermatol
126:1102–10
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ, Clarke MF (2003) Prospective
identification of tumorigenic breast cancer
cells. Proc Natl Acad Sci USA 100:3983–8
Amoh Y, Li L, Katsuoka K, Penman S, Hoffman
RM (2005) Multipotent nestin-positive,
www.jidonline.org 2377
JM GrichnikMelanoma, Nevogenesis, and Stem Cell Biology

keratin-negative hair-follicle bulge stem cellscan form neurons. Proc Natl Acad Sci USA102:5530–4
Arbiser JL (2003) Activation of B-raf in non-malignant nevi predicts a novel tumorsuppressor gene in melanoma (MAP kinasephosphatase). J Invest Dermatol 121:xiv
Banky JP, Kelly JW, English DR, Yeatman JM,Dowling JP (2005) Incidence of new andchanged nevi and melanomas detected usingbaseline images and dermoscopy in patientsat high risk for melanoma. Arch Dermatol141:998–1006
Bastian BC, Kashani-Sabet M, Hamm H, Godfrey T,Moore DH II, Brocker EB et al. (2000)Gene amplifications characterize acral mela-noma and permit the detection of occulttumor cells in the surrounding skin. CancerRes 60:1968–73
Bataille V, Kato BS, Falchi M, Gardner J, KimuraM, Lens M et al. (2007) Nevus size andnumber are associated with telomere lengthand represent potential markers of a decrea-sed senescence in vivo. Cancer EpidemiolBiomarkers Prev 16:1499–502
Bauer J, Curtin JA, Pinkel D, Bastian BC (2007)Congenital melanocytic nevi frequently har-bor NRAS mutations but no BRAF mutations.J Invest Dermatol 127:179–82
Baynash AG, Hosoda K, Giaid A, Richardson JA,Emoto N, Hammer RE et al. (1994) Inter-action of endothelin-3 with endothelin-Breceptor is essential for development ofepidermal melanocytes and enteric neurons.Cell 79:1277–85
Bekaert S, Derradji H, Baatout S (2004) Telomerebiology in mammalian germ cells and duringdevelopment. Dev Biol 274:15–30
Belicchi M, Pisati F, Lopa R, Porretti L, FortunatoF, Sironi M et al. (2004) Human skin-derivedstem cells migrate throughout forebrain anddifferentiate into astrocytes after injectioninto adult mouse brain. J Neurosci Res 77:475–86
Bennett DC, Medrano EE (2002) Molecularregulation of melanocyte senescence.Pigment Cell Res 15:242–50
Bevona C, Goggins W, Quinn T, Fullerton J,Tsao H (2003) Cutaneous melanomas asso-ciated with nevi. Arch Dermatol 139:1620–4
Bickenbach JR, Grinnell KL (2004) Epidermalstem cells: interactions in developmentalenvironments. Differentiation 72:371–80
Blanpain C, Fuchs E (2006) Epidermal stem cells ofthe skin. Annu Rev Cell Dev Biol 22:339–73
Bonnet D, Dick JE (1997) Human acute myeloidleukemia is organized as a hierarchy thatoriginates from a primitive hematopoieticcell. Nat Med 3:730–7
Botchkareva NV, Khlgatian M, Longley BJ,Botchkarev VA, Gilchrest BA (2001) SCF/c-kit signaling is required for cyclic regene-ration of the hair pigmentation unit. FASEB J15:645–58
Burkhart CG, Burkhart CN (2005) The moletheory: primary function of melanocytesand melanin may be antimicrobial defense
and immunomodulation (not solar protec-tion). Int J Dermatol 44:340–2
Charames GS, Bapat B (2003) Genomic instabilityand cancer. Curr Mol Med 3:589–96
Chen YT, Venditti CA, Theiler G, Stevenson BJ,Iseli C, Gure AO et al. (2005) Identifi-cation of CT46/HORMAD1, an immuno-genic cancer/testis antigen encoding aputative meiosis-related protein. CancerImmun 5:9
Chudnovsky Y, Adams AE, Robbins PB, Lin Q,Khavari PA (2005) Use of human tissue toassess the oncogenic activity of melanoma-associated mutations. Nat Genet 37:745–9
Cotsarelis G (2006) Epithelial stem cells: afolliculocentric view. J Invest Dermatol 126:1459–68
Cramer SF (1984) The histogenesis of acquiredmelanocytic nevi. Based on a new conceptof melanocytic differentiation. Am J Derma-topathol 6:289–98
Cramer SF (1991) The origin of epidermalmelanocytes. Implications for the histogen-esis of nevi and melanomas. Arch Pathol LabMed 115:115–9
Curtin JA, Busam K, Pinkel D, Bastian BC (2006)Somatic activation of KIT in distinct subtypesof melanoma. J Clin Oncol 24:4340–6
Dahl C, Saito H, Nielsen HV, Schiotz PO (2002)The establishment of a combined serum-freeand serum-supplemented culture method ofobtaining functional cord blood-derivedhuman mast cells. J Immunol Methods 262:137–43
Dalerba P, Dylla SJ, Park IK, Liu R, Wang X, ChoRW et al. (2007) Phenotypic characterizationof human colorectal cancer stem cells. ProcNatl Acad Sci USA 104:10158–63
de Cholnoky T (1941) Malignant Melanoma: Aclinical study of one hundred seventeencases. Ann Surg 113:392–410
Dupin E, Calloni G, Real C, Goncalves-Trentin A,Le Douarin NM (2007) Neural crest progeni-tors and stem cells. C R Biol 330:521–9
Etienne G, Cony-Makhoul P, Mahon FX (2002)Imatinib mesylate and gray hair. N Engl JMed 347:446
Ewing J (1930) The Problems of Melanoma. B JMed 11:852–6
Fang D, Leishear K, Nguyen TK, Finko R, Cai K,Fukunaga M et al. (2006) Defining theconditions for the generation of melanocytesfrom human embryonic stem cells. StemCells 24:1668–77
Fang D, Nguyen TK, Leishear K, Finko R, KulpAN, Hotz S et al. (2005) A tumorigenicsubpopulation with stem cell properties inmelanomas. Cancer Res 65:9328–37
Farmer PJ, Gidanian S, Shahandeh B, Di Bilio AJ,Tohidian N, Meyskens FL Jr (2003) Melaninas a target for melanoma chemotherapy: pro-oxidant effect of oxygen and metals onmelanoma viability. Pigment Cell Res 16:273–9
Fernandes KJ, McKenzie IA, Mill P, Smith KM,Akhavan M, Barnabe-Heider F et al. (2004) Adermal niche for multipotent adult skin-
derived precursor cells. Nat Cell Biol6:1082–93
Frank NY, Margaryan A, Huang Y, Schatton T,Waaga-Gasser AM, Gasser M et al. (2005)ABCB5-mediated doxorubicin transport andchemoresistance in human malignant mela-noma. Cancer Res 65:4320–33
Frank NY, Pendse SS, Lapchak PH, Margaryan A,Shlain D, Doeing C et al. (2003) Regulationof progenitor cell fusion by ABCB5 P-glycoprotein, a novel human ATP-bindingcassette transporter. J Biol Chem 278:47156–65
Gehling UM, Ergun S, Schumacher U, WagenerC, Pantel K, Otte M et al. (2000) In vitrodifferentiation of endothelial cells fromAC133-positive progenitor cells. Blood95:3106–12
Gilley D, Tanaka H, Herbert BS (2005) Telomeredysfunction in aging and cancer. Int JBiochem Cell Biol 37:1000–13
Grichnik JM (2006a) Genomic instability andtumor stem cells. J Invest Dermatol 126:1214–6
Grichnik JM (2006b) Kit and melanocyte migra-tion. J Invest Dermatol 126:945–7
Grichnik JM, Ali WN, Burch JA, Byers JD, GarciaCA, Clark RE et al. (1996) KIT expressionreveals a population of precursor melano-cytes in human skin. J Invest Dermatol106:967–71
Grichnik JM, Burch JA, Burchette J, Shea CR(1998) The SCF/KIT pathway plays a criticalrole in the control of normal human mela-nocyte homeostasis. J Invest Dermatol111:233–8
Grichnik JM, Burch JA, Schulteis RD, Shan S, LiuJ, Darrow TL et al. (2006) Melanoma, atumor based on a mutant stem cell? J InvestDermatol 126:142–53
Grichnik JM, Crawford J, Jimenez F, Kurtzberg J,Buchanan M, Blackwell S et al. (1995)Human recombinant stem-cell factor in-duces melanocytic hyperplasia in suscepti-ble patients. J Am Acad Dermatol 33:577–83
Halaban R (2000) The regulation of normal melano-cyte proliferation. Pigment Cell Res 13:4–14
Halaban R, Ghosh S, Duray P, Kirkwood JM,Lerner AB (1986) Human melanocytes cul-tured from nevi and melanomas. J InvestDermatol 87:95–101
Hardy K, Martin KL, Leese HJ, Winston RM,Handyside AH (1990) Human preimplanta-tion development in vitro is not adverselyaffected by biopsy at the 8-cell stage. HumReprod 5:708–14
Hendrix MJ, Seftor EA, Hess AR, Seftor RE (2003)Vasculogenic mimicry and tumour-cell plas-ticity: lessons from melanoma. Nat RevCancer 3:411–21
Hendrix MJ, Seftor EA, Seftor RE, Kasemeier-Kulesa J, Kulesa PM, Postovit LM (2007)Reprogramming metastatic tumour cells withembryonic microenvironments. Nat RevCancer 7:246–55
Hirobe T (2005) Role of keratinocyte-derivedfactors involved in regulating the proli-feration and differentiation of mammalian
2378 Journal of Investigative Dermatology (2008), Volume 128
JM GrichnikMelanoma, Nevogenesis, and Stem Cell Biology

epidermal melanocytes. Pigment Cell Res18:2–12
Ho MM, Ng AV, Lam S, Hung JY (2007) Sidepopulation in human lung cancer cell linesand tumors is enriched with stem-like cancercells. Cancer Res 67:4827–33
Hong L, Simon JD (2007) Current understandingof the binding sites, capacity, affinity, andbiological significance of metals in melanin.J Phys Chem B 111:7938–47
Hosoda K, Hammer RE, Richardson JA, BaynashAG, Cheung JC, Giaid A et al. (1994)Targeted and natural (piebald-lethal) muta-tions of endothelin-B receptor gene producemegacolon associated with spotted coatcolor in mice. Cell 79:1267–76
Hu YF, Zhang ZJ, Sieber-Blum M (2006) Anepidermal neural crest stem cell (EPI-NCSC)molecular signature. Stem Cells 24:2692–702
Hui P, Perkins A, Glusac E (2001) Assessment ofclonality in melanocytic nevi. J Cutan Pathol28:140–4
Ichii-Nakato N, Takata M, Takayanagi S, Takashi-ma S, Lin J, Murata H et al. (2006) Highfrequency of BRAFV600E mutation in ac-quired nevi and small congenital nevi, butlow frequency of mutation in medium-sizedcongenital nevi. J Invest Dermatol 126:2111–8
Jiao Z, Zhang ZG, Hornyak TJ, Hozeska A, ZhangRL, Wang Y et al. (2006) Dopachrometautomerase (Dct) regulates neural progeni-tor cell proliferation. Dev Biol 296:396–408
Kantor GR, Wheeland RG (1987) Transepidermalelimination of nevus cells. A possible me-chanism of nevus involution. Arch Dermatol123:1371–4
Kavak A, Akcan Y, Korkmaz U (2005) Hairrepigmentation in a hepatitis C patienttreated with interferon and ribavirin. Dermato-logy 211:171–2
Kelsall SR, Mintz B (1998) Metastatic cutaneousmelanoma promoted by ultraviolet radiationin mice with transgene-initiated lowmelanoma susceptibility. Cancer Res 58:4061–5
Klein WM, Wu BP, Zhao S, Wu H, Klein-SzantoAJ, Tahan SR (2007) Increased expression ofstem cell markers in malignant melanoma.Mod Pathol 20:102–7
Kondo T, Setoguchi T, Taga T (2004) Persistenceof a small subpopulation of cancer stem-likecells in the C6 glioma cell line. Proc NatlAcad Sci USA 101:781–6
Kraemer KH, Lee MM, Andrews AD, Lambert WC(1994) The role of sunlight and DNA repairin melanoma and nonmelanoma skin can-cer. The xeroderma pigmentosum paradigm.Arch Dermatol 130:1018–21
Krengel S, Hauschild A, Schafer T (2006) Mela-noma risk in congenital melanocytic naevi:a systematic review. Br J Dermatol 155:1–8
Kumamoto T, Shalhevet D, Matsue H, MummertME, Ward BR, Jester JV et al. (2003) Hairfollicles serve as local reservoirs of skin mastcell precursors. Blood 102:1654–60
Kunisada T, Lu SZ, Yoshida H, Nishikawa S,Nishikawa S, Mizoguchi M et al. (1998)Murine cutaneous mastocytosis and epider-
mal melanocytosis induced by keratinocyteexpression of transgenic stem cell factor.J Exp Med 187:1565–73
Lahav R, Heffner G, Patterson PH (1999) Anendothelin receptor B antagonist inhibitsgrowth and induces cell death in humanmelanoma cells in vitro and in vivo. ProcNatl Acad Sci USA 96:11496–500
Lapidot T, Sirard C, Vormoor J, Murdoch B,Hoang T, Caceres-Cortes J et al. (1994) A cellinitiating human acute myeloid leukaemiaafter transplantation into SCID mice. Nature367:645–8
Lee JT, Herlyn M (2007) Microenvironmentalinfluences in melanoma progression. J CellBiochem 101:862–72
Li C, Heidt DG, Dalerba P, Burant CF, Zhang L,Adsay V et al. (2007) Identification of pancrea-tic cancer stem cells. Cancer Res 67:1030–7
Liang L, Bickenbach JR (2002) Somatic epidermalstem cells can produce multiple cell lineagesduring development. Stem Cells 20:21–31
Lin F, Burch JA, Singer S, Grichnik JM (2000) SCFinduced proliferation of human melanocytesis masked in vitro by bFGF. J Invest Dermatol114:859 (abstract)
Liu L, Liu H, Jao J, Liu H, Bergeron A, Dong JFet al. (2007) Changes in circulating humanendothelial progenitor cells after braininjury. J Neurotrauma 24:936–43
Lucas CR, Sanders LL, Murray JC, Myers SA, HallRP, Grichnik JM (2003) Early melanomadetection: nonuniform dermoscopic featuresand growth. J Am Acad Dermatol 48:663–71
Luftl M, Schuler G, Jungbluth AA (2004) Mela-noma or not? Cancer testis antigens mayhelp. Br J Dermatol 151:1213–8
Mackenzie MA, Jordan SA, Budd PS, Jackson IJ(1997) Activation of the receptor tyrosinekinase Kit is required for the proliferation ofmelanoblasts in the mouse embryo. Dev Biol192:99–107
Mancianti ML, Gyorfi T, Shih IM, Valyi-Nagy I,Levengood G, Menssen HD et al. (1993)Growth regulation of cultured human nevuscells. J Invest Dermatol 100:281S–7S
Megahed M, Schon M, Selimovic D, Schon MP(2002) Reliability of diagnosis of melanomain situ. Lancet 359:1921–2
Meyskens Jr FL, Farmer PJ, Anton-Culver H (2004)Etiologic pathogenesis of melanoma: a uni-fying hypothesis for the missing attributablerisk. Clin Cancer Res 10:2581–3
Molino A, Lucas C, Benitez-Graham A, Burch JA,Herman C, Selim A et al. (2003) Immaturedermal melanocytic cells are present ingrowing melanocytic neoplasms. J InvestDermatol 121:1174 (abstract)
Monzani E, Facchetti F, Galmozzi E, Corsini E,Benetti A, Cavazzin C et al. (2007) Melano-ma contains CD133 and ABCG2 positivecells with enhanced tumourigenic potential.Eur J Cancer 43:935–46
Motohashi T, Aoki H, Chiba K, Yoshimura N,Kunisada T (2007) Multipotent cell fate ofneural crest-like cells derived from embryo-nic stem cells. Stem Cells 25:402–10
Na GY, Paek SH, Park BC, Kim DW, Lee WJ, LeeSJ et al. (2006) Isolation and characterizationof outer root sheath melanocytes of humanhair follicles. Br J Dermatol 155:902–9
Nishikawa S, Kusakabe M, Yoshinaga K, OgawaM, Hayashi S, Kunisada T et al. (1991)In utero manipulation of coat color forma-tion by a monoclonal anti-c-kit antibody:two distinct waves of c-kit-dependencyduring melanocyte development. EMBOJ 10:2111–8
Nishikawa S, Osawa M (2007) Generating quies-cent stem cells. Pigment Cell Res 20:263–70
Nishimura EK, Granter SR, Fisher DE (2005)Mechanisms of hair graying: incompletemelanocyte stem cell maintenance in theniche. Science 307:720–4
Nishimura EK, Jordan SA, Oshima H, Yoshida H,Osawa M, Moriyama M et al. (2002)Dominant role of the niche in melanocytestem-cell fate determination. Nature 416:854–60
Oback B, Wells DN (2007) Donor cell differ-entiation, reprogramming, and cloning effi-ciency: elusive or illusive correlation? MolReprod Dev 74:646–54
O’Brien CA, Pollett A, Gallinger S, Dick JE (2007)A human colon cancer cell capable ofinitiating tumour growth in immunodeficientmice. Nature 445:106–10
Okura M, Maeda H, Nishikawa S, Mizoguchi M(1995) Effects of monoclonal anti-c-kit anti-body (ACK2) on melanocytes in newbornmice. J Invest Dermatol 105:322–8
Peters EM, Tobin DJ, Botchkareva N, Maurer M,Paus R (2002) Migration of melanoblasts intothe developing murine hair follicle is accom-panied by transient c-Kit expression.J Histochem Cytochem 50:751–66
Petronic-Rosic V, Shea CR, Krausz T (2004)Pagetoid melanocytosis: when is it signifi-cant? Pathology (Phila) 36:435–44
Pharis DB, Zitelli JA (2001) Sunburn, trauma, andthe timing of biopsies of melanocytic nevi.Dermatol Surg 27:835–6
Pierce KE, Michalopoulos J, Kiessling AA, SeibelMM, Zilberstein M (1997) Preimplantationdevelopment of mouse and human embryosbiopsied at cleavage stages using a modifieddisplacement technique. Hum Reprod12:351–6
Pizzichetta MA, Massone C, Grandi G, Pelizzo G,Soyer HP (2006) Morphologic changes ofacquired melanocytic nevi with eccentricfoci of hyperpigmentation (‘‘Bolognia sign’’)assessed by dermoscopy. Arch Dermatol142:479–83
Prince ME, Sivanandan R, Kaczorowski A, WolfGT, Kaplan MJ, Dalerba P et al. (2007)Identification of a subpopulation of cellswith cancer stem cell properties in head andneck squamous cell carcinoma. Proc NatlAcad Sci USA 104:973–8
Real C, Glavieux-Pardanaud C, Le Douarin NM,Dupin E (2006) Clonally cultured differen-tiated pigment cells can dedifferentiate andgenerate multipotent progenitors with self-renewing potential. Dev Biol 300:656–69
www.jidonline.org 2379
JM GrichnikMelanoma, Nevogenesis, and Stem Cell Biology

Reya T, Morrison SJ, Clarke MF, Weissman IL(2001) Stem cells, cancer, and cancer stemcells. Nature 414:105–11
Ricci-Vitiani L, Lombardi DG, Pilozzi E, BiffoniM, Todaro M, Peschle C et al. (2007)Identification and expansion of human colon-cancer-initiating cells. Nature 445:111–5
Robinson WA, Lemon M, Elefanty A, Harrison-Smith M, Markham N, Norris D (1998)Human acquired naevi are clonal. Melano-ma Res 8:499–503
Singh SK, Clarke ID, Terasaki M, Bonn VE,Hawkins C, Squire J et al. (2003) Identifica-tion of a cancer stem cell in human braintumors. Cancer Res 63:5821–8
Slominski A, Wortsman J, Tuckey RC, Paus R(2007) Differential expression of HPA axishomolog in the skin. Mol Cell Endocrinol265–266:143–9
Stark M, Hayward N (2007) Genome-wide loss ofheterozygosity and copy number analysis inmelanoma using high-density single-nucleo-tide polymorphism arrays. Cancer Res67:2632–42
Takeda K, Takahashi NH, Shibahara S (2007)Neuroendocrine functions of melanocytes:beyond the skin-deep melanin maker. Toho-ku J Exp Med 211:201–21
Tobin DJ, Hagen E, Botchkarev VA, Paus R (1998)Do hair bulb melanocytes undergo apoptosisduring hair follicle regression (catagen)?J Invest Dermatol 111:941–7
Toma JG, Akhavan M, Fernandes KJ, Barnabe-Heider F, Sadikot A, Kaplan DR et al. (2001)Isolation of multipotent adult stem cells fromthe dermis of mammalian skin. Nat Cell Biol3:778–84
Tondreau T, Meuleman N, Delforge A, DejeneffeM, Leroy R, Massy M et al. (2005) Mesen-chymal stem cells derived from CD133-positive cells in mobilized peripheral bloodand cord blood: proliferation, Oct4 expres-sion, and plasticity. Stem Cells 23:1105–12
Tsao H, Goel V, Wu H, Yang G, Haluska FG(2004) Genetic interaction between NRASand BRAF mutations and PTEN/MMAC1inactivation in melanoma. J Invest Dermatol122:337–41
Tumbar T, Guasch G, Greco V, Blanpain C,Lowry WE, Rendl M et al. (2004) Definingthe epithelial stem cell niche in skin. Science303:359–63
Tureci O, Sahin U, Zwick C, Koslowski M, SeitzG, Pfreundschuh M (1998) Identification ofa meiosis-specific protein as a member of theclass of cancer/testis antigens. Proc NatlAcad Sci USA 95:5211–6
Uchida N, Buck DW, He D, Reitsma MJ, MasekM, Phan TV et al. (2000) Direct isolation ofhuman central nervous system stem cells.Proc Natl Acad Sci USA 97:14720–5
Wang E, Voiculescu S, Le Poole IC, El-Gamil M,Li X, Robbins PF et al. (2006) Clonalpersistence and evolution during a decade ofrecurrent melanoma. J Invest Dermatol 126:1372–7
Wilkie AL, Jordan SA, Jackson IJ (2002) Neuralcrest progenitors of the melanocyte lineage:coat colour patterns revisited. Development129:3349–57
Wong AK, Chin L (2000) An inducible melanomamodel implicates a role for RAS in tumormaintenance and angiogenesis. CancerMetastasis Rev 19:121–9
Wong CE, Paratore C, Dours-Zimmermann MT,Rochat A, Pietri T, Suter U et al. (2006)Neural crest-derived cells with stem cellfeatures can be traced back to multiplelineages in the adult skin. J Cell Biol 175:1005–15
Worret WI, Burgdorf WH (1998) Which directiondo nevus cells move? Abtropfung reexa-mined. Am J Dermatopathol 20:135–9
Yamane T, Hayashi S, Mizoguchi M, Yamazaki H,Kunisada T (1999) Derivation of melano-cytes from embryonic stem cells in culture.Dev Dyn 216:450–8
Yang CT, Johnson SL (2006) Small molecule-induced ablation and subsequent regene-ration of larval zebrafish melanocytes.Development 133:3563–73
Yoshida H, Kunisada T, Kusakabe M, NishikawaS, Nishikawa SI (1996) Distinct stagesof melanocyte differentiation revealed byanalysis of nonuniform pigmentation pat-terns. Development 122:1207–14
Yu H, Fang D, Kumar SM, Li L, Nguyen TK, Acs Get al. (2006) Isolation of a novel populationof multipotent adult stem cells from humanhair follicles. Am J Pathol 168:1879–88
Yu Y, Flint A, Dvorin EL, Bischoff J (2002) AC133-2, a novel isoform of human AC133 stem cellantigen. J Biol Chem 277:20711–6
Zalaudek I, Ferrara G, Argenziano G (2007)Dermoscopy insights into nevogenesis:‘‘Abtropfung’’ vs ‘‘Hochsteigerung’’. ArchDermatol 143:284
Zalaudek I, Grinschgl S, Argenziano G, MarghoobAA, Blum A, Richtig E et al. (2006) Age-related prevalence of dermoscopy patterns inacquired melanocytic naevi. Br J Dermatol154:299–304
2380 Journal of Investigative Dermatology (2008), Volume 128
JM GrichnikMelanoma, Nevogenesis, and Stem Cell Biology




















