
Meiotic behavior and pollen viability of tetraploid Arachis glabrata and A. nitida species (Section...
11
ORIGINAL ARTICLE Meiotic behavior and pollen viability of tetraploid Arachis glabrata and A. nitida species (Section Rhizomatosae, Leguminosae): implications concerning their polyploid nature and seed set production Alejandra Marcela Ortiz • Jose ´ Guillermo Seijo • Aveliano Ferna ´ndez • Graciela Ine ´s Lavia Received: 11 September 2010 / Accepted: 20 December 2010 Ó Springer-Verlag 2011 Abstract The meiotic behavior and pollen viability of the tetraploids (2n = 4x = 40) Arachis glabrata and A. nitida were analyzed aiming to provide insights into the nature of these polyploids and into the mechanism that determines the low seed production of these species. Meiotic analysis revealed 21 different chromosome configurations at diaki- nesis-metaphase I in A. glabrata (from 20 II to 4 II ? 8 IV) and 7 in A. nitida (from 20 II to 2 I ? 12 II ? 2 III ? 2 IV). The multivalent associations (up to 8 IV) observed in some A. glabrata metaphases suggest that this species may have an autopolyploid origin. However, the fact that the mean number of bivalents varied among accessions indi- cates different degrees of diploidization among them. In contrast, the multivalents (up to 2 III ? 2 IV) observed in very low frequency in A. nitida indicate that this species may be either a largely diploidized autopolyploid or a segmental allopolyploid. A great diversity, although in low frequency, of meiotic abnormalities (abnormal chromo- some orientation and segregation, chromosome bridges, irregular spindles, micronuclei, aneuploid nuclei, restitu- tion nuclei, microcytes, monads, dyads, triads, and hexads) was detected in both species. The meiotic indexes were over 95%, and pollen viabilities ranged from 83.20 to 95.99%. These results suggest that chromosome behavior during meiosis would not severely affect pollen viability. Thus, the irregular chromosome behavior caused by the autopolyploid or segmental allopolyploid nature of these species may not be related to their low seed set production. Keywords Arachis glabrata Á A. nitida Á Chromosome associations Á Diploidization Á Low seed production Á Meiotic abnormalities Á Polyploidy Introduction The genus Arachis (Leguminosae) is native to South America and is distributed eastwards of the Andes moun- tains, from the Amazon River to the La Plata River. It has 80 formally recognized species characterized by having a geocarpic fruit and autogamy (Krapovickas and Gregory 1994; Valls and Simpson 2005). Based on their morpho- logical characters, as well as on the cross compatibility and fertility of the hybrids, the species have been arranged into nine taxonomic sections (Krapovickas and Gregory 1994; Valls and Simpson 2005). The section Rhizomatosae has been defined on mor- phological features exclusively by the presence of rhi- zomes in all the entities (Krapovickas and Gregory 1994). From a biological point of view, this section is very interesting because it has the highest frequency of poly- ploid species (75%), including three tetraploids (2n = 4x = 40), A. glabrata Benth., A. nitida Valls, Krapov. et C.E. Simpson, and A. pseudovillosa (Chodat et Hassl.) Krapov. et W.C. Gregory, and only one diploid taxon (2n = 2x = 20), A. burkartii Handro (Lavia et al. 2008 and references therein). In spite of the high frequency of polyploids in the section, their nature remains unknown. Cytogenetic information for these taxa is limited to the Part of a Doctoral Thesis that will be presented to the Facultad de Ciencias Exactas, Fı ´sicas y Naturales de Co ´rdoba. A. M. Ortiz Á J. G. Seijo Á A. Ferna ´ndez Á G. I. Lavia (&) Instituto de Bota ´nica del Nordeste (UNNE-CONICET), Casilla de Correo 209, 3400 Corrientes, Argentina e-mail: [email protected] J. G. Seijo Á A. Ferna ´ndez Á G. I. Lavia Facultad de Ciencias Exactas y Naturales y Agrimensura (UNNE), Av. Libertad 5460, 3400 Corrientes, Argentina 123 Plant Syst Evol DOI 10.1007/s00606-010-0397-8
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Meiotic behavior and pollen viability of tetraploid Arachis glabrata and A. nitida species (Section...

ORIGINAL ARTICLE
Meiotic behavior and pollen viability of tetraploid Arachisglabrata and A. nitida species (Section Rhizomatosae,Leguminosae): implications concerning their polyploidnature and seed set production
Alejandra Marcela Ortiz • Jose Guillermo Seijo •
Aveliano Fernandez • Graciela Ines Lavia
Received: 11 September 2010 / Accepted: 20 December 2010
� Springer-Verlag 2011
Abstract The meiotic behavior and pollen viability of the
tetraploids (2n = 4x = 40) Arachis glabrata and A. nitida
were analyzed aiming to provide insights into the nature of
these polyploids and into the mechanism that determines
the low seed production of these species. Meiotic analysis
revealed 21 different chromosome configurations at diaki-
nesis-metaphase I in A. glabrata (from 20 II to 4 II ? 8 IV)
and 7 in A. nitida (from 20 II to 2 I ? 12 II ? 2 III ? 2
IV). The multivalent associations (up to 8 IV) observed in
some A. glabrata metaphases suggest that this species may
have an autopolyploid origin. However, the fact that the
mean number of bivalents varied among accessions indi-
cates different degrees of diploidization among them. In
contrast, the multivalents (up to 2 III ? 2 IV) observed in
very low frequency in A. nitida indicate that this species
may be either a largely diploidized autopolyploid or a
segmental allopolyploid. A great diversity, although in low
frequency, of meiotic abnormalities (abnormal chromo-
some orientation and segregation, chromosome bridges,
irregular spindles, micronuclei, aneuploid nuclei, restitu-
tion nuclei, microcytes, monads, dyads, triads, and hexads)
was detected in both species. The meiotic indexes were
over 95%, and pollen viabilities ranged from 83.20 to
95.99%. These results suggest that chromosome behavior
during meiosis would not severely affect pollen viability.
Thus, the irregular chromosome behavior caused by the
autopolyploid or segmental allopolyploid nature of these
species may not be related to their low seed set production.
Keywords Arachis glabrata � A. nitida � Chromosome
associations � Diploidization � Low seed production �Meiotic abnormalities � Polyploidy
Introduction
The genus Arachis (Leguminosae) is native to South
America and is distributed eastwards of the Andes moun-
tains, from the Amazon River to the La Plata River. It has
80 formally recognized species characterized by having a
geocarpic fruit and autogamy (Krapovickas and Gregory
1994; Valls and Simpson 2005). Based on their morpho-
logical characters, as well as on the cross compatibility and
fertility of the hybrids, the species have been arranged into
nine taxonomic sections (Krapovickas and Gregory 1994;
Valls and Simpson 2005).
The section Rhizomatosae has been defined on mor-
phological features exclusively by the presence of rhi-
zomes in all the entities (Krapovickas and Gregory 1994).
From a biological point of view, this section is very
interesting because it has the highest frequency of poly-
ploid species (75%), including three tetraploids (2n =
4x = 40), A. glabrata Benth., A. nitida Valls, Krapov. et
C.E. Simpson, and A. pseudovillosa (Chodat et Hassl.)
Krapov. et W.C. Gregory, and only one diploid taxon
(2n = 2x = 20), A. burkartii Handro (Lavia et al. 2008 and
references therein). In spite of the high frequency of
polyploids in the section, their nature remains unknown.
Cytogenetic information for these taxa is limited to the
Part of a Doctoral Thesis that will be presented to the Facultad de
Ciencias Exactas, Fısicas y Naturales de Cordoba.
A. M. Ortiz � J. G. Seijo � A. Fernandez � G. I. Lavia (&)
Instituto de Botanica del Nordeste (UNNE-CONICET),
Casilla de Correo 209, 3400 Corrientes, Argentina
e-mail: [email protected]
J. G. Seijo � A. Fernandez � G. I. Lavia
Facultad de Ciencias Exactas y Naturales y Agrimensura
(UNNE), Av. Libertad 5460, 3400 Corrientes, Argentina
123
Plant Syst Evol
DOI 10.1007/s00606-010-0397-8

somatic chromosome numbers of the species and to the
morphology of the SAT chromosomes of A. glabrata and
A. nitida (Lavia et al. 2008 and references therein). Pre-
vious meiotic analysis included only A. glabrata, and, in
those reports, the authors just mentioned a few quadrivalent
associations in meiosis I (Raman 1981; Mallikarjuna and
Sastri 2002). Therefore, a detailed analysis of the chro-
mosome associations and behavior during the meiosis of
the tetraploid species is still lacking.
Cross-compatibility experiments between rhizomatous
species have revealed that none of the tetraploids produce
F1 hybrids with A. burkartii (Krapovickas and Gregory
1994). In addition, in spite of the many attempts to cross
polyploid species, only two hybrid plants were obtained
between A. pseudovillosa and A. glabrata (Krapovickas
and Gregory 1994). These incompatibilities evidenced the
existence of strong genetic barriers between the tetraploid
species and between them and A. burkartii, thus ques-
tioning the monophyly of the section Rhizomatosae.
Arachis nitida, which grows naturally in NE Paraguay
(Amambay and Concepcion Departments) and South of
Mato Grosso do Sul State in Brazil (Valls and Simpson
2005), was initially treated as A. glabrata, but was segre-
gated as a new species by Valls and Simpson (2005). From
a biological point of view, this taxon has great interest
because its morphology resembles that of A. burkartii,
although their current geographical areas are segregated by
a long distance (1,000 km). In this context, it is expected
that the genetic and cytogenetic analysis of A. nitida may
provide some hints about the origin of tetraploids among
the section.
Arachis glabrata has the most widespread distribution
of all the Arachis species, growing spontaneously from NE
Argentina to the Tocantins State in Brazil, and from San
Paulo and Mina Gerais States in Brazil up to the Paraguay
River (Krapovickas and Gregory 1994). It is the most
agronomically important species of section Rhizomatosae
since it produces high-quality forage and is also used as
cover crop and ornamental turf (Prine et al. 1981; Prine
et al. 1986; Rouse et al. 2004). Additionally, peanut
breeders have great interest in A. glabrata because it is
resistant to many diseases, insects, and nematodes that
constitute severe problems to A. hypogaea L. (Mallikarjuna
and Sastri 2002).
In spite of the agronomic potential of A. glabrata, all
commercial cultivars have been developed only from the
selection of a few accessions (French et al. 1994), and
genetic improvement by hybridization between the rhizo-
matous accessions with desirable traits has not yet been
reported. The available gene pool of A. glabrata is con-
siderably large, with more than 300 accessions, kept
mainly as live collections in germplasm banks (Valls et al.
1994). Biochemical and molecular markers, such as
isozymes (Maass and Ocampo 1995) and microsatellites
(Angelici et al. 2008), have revealed a high genetic poly-
morphism among these accessions, which reflect, to some
extent, the wide morphological variability exhibited by the
species in traits like color and size of leaves and flowers,
and biomass in natural populations (Krapovickas and
Gregory 1994; G. Lavia, pers. obs.). Therefore, these
materials constitute a great source of new alleles for the
genetic improvement of the existing commercial cultivars
and for the development of new ones.
The main feature that has constrained a greater diffusion
of A. glabrata as subtropical legume forage is its low to
null seed production (Simpson et al. 1994). Establishment
of this species in grass fields depends on its asexual
propagation by rhizomes, which is time-consuming and
expensive. Considering that, the understanding of the
mechanisms that restrict or inhibit seed production is one
of the main keystones for the development of improved
cultivars and for their massive adoption as forage. In dif-
ferent groups of plant species, reduction of the seed set has
been related to meiotic irregularities (Pagliarini 2000; Luan
et al. 2009), pollen-stigma incompatibilities (Nielsen et al.
2003), and postzygotic abortion (Diggle et al. 2002).
However, to date, the causes that determine the low seed
set in A. glabrata remain unknown.
In this context, in the present work, we carried out a
detailed meiotic analysis of six accessions of A. glabrata
and one of A. nitida in order to provide insights into the
nature of these two rhizomatous tetraploids. Additionally,
we evaluated the influence of different meiotic events on
pollen viability as a first approach to understand the
mechanisms that determine the reduction of seed produc-
tion in these species.
Materials and methods
Suitable floral buds of six Arachis glabrata and one
A. nitida accessions were obtained from plants growing
under greenhouse conditions at the Instituto de Botanica
del Nordeste, Corrientes, Argentina. Vouchers of original
collections are kept at the CTES herbarium, and living
collections are kept under greenhouse conditions. The
original provenances and voucher specimens of the
accessions studied are listed in Table 1.
Meiotic analysis was carried out in pollen mother cells
(PMCs) from young flower buds, either fresh or fixed in
ethanol:acetic acid (3:1). PMCs were stained and squashed
in 2% lactopropionic orcein (Dyer 1963). Permanent slides
were prepared using Euparal as mounting medium.
Chromosome associations at diakinesis-metaphase I
were evaluated at 20–49 PMCs per accession. These data
were normalized and compared by one-way analysis of
A. M. Ortiz et al.
123

variance (ANOVA) followed by Tukey’s test (InfoStat
2008) in order to detect whether the A. glabrata and A.
nitida accessions have significant intraspecific and inter-
specific variations in their mean chromosome associations.
The meiotic behavior was evaluated from metaphase I to
sporad stage in at least 1,848 PMCs per accession
(Table 4), and the frequency of each abnormality was
recorded. The meiotic index was calculated as the ratio
between the number of normal sporads/total sporads ana-
lyzed per accession 9100. Tetrads were considered normal
when they had four equal-sized cells.
Stainability with carmine:glycerine (1:1) was used as an
estimation of pollen viability (Pittenger and Frolik 1951).
Three floral buds of different inflorescences were analyzed,
and at least 500 pollen grains per bud were counted for
each accession. Pollen grains were considered viable when
the cytoplasm was uniformly darkly stained, and, accord-
ing to their size and shape, classified into four categories:
large, normal, small, and abnormally shaped grains.
Unviable pollen grains were grouped into three categories:
micro (small spheroids), shrunken (prolate with less than
50% of stained cytoplasm), and empty (prolate without
stained cytoplasm) grains. Pollen viability was expressed
as an average percentage of the stained pollen grains/total
pollen grains analyzed.
Results and discussion
Chromosome pairing analysis and evidence
of polyploid nature
The analysis of 149 PMCs at diakinesis-metaphase I in all
A. glabrata accessions revealed 21 different meiotic con-
figurations, including univalent (I), bivalent (II), trivalent
(III), quadrivalent (IV), pentavalent (V), and hexavalent (VI)
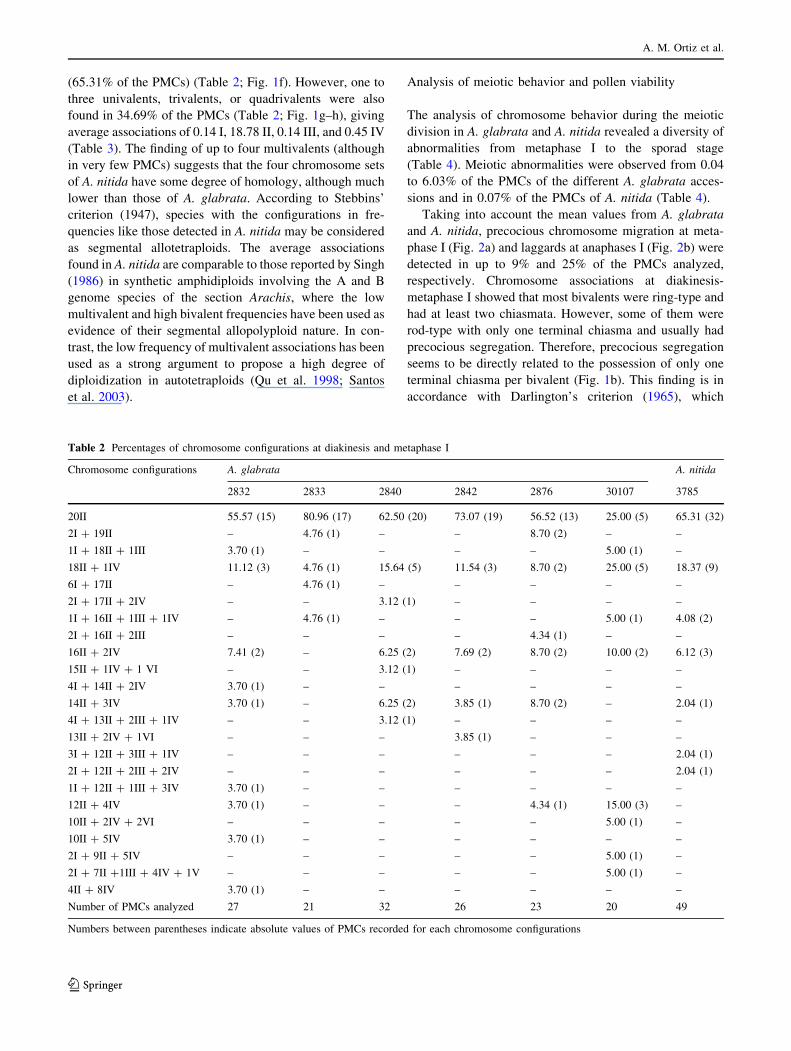
associations (Tables 2, 3, Fig. 1a–e). The configurations
most frequently observed in all accessions were 20 II and
18 II ? 1 IV. The former ranged from 25 to 80.96% of the
PMCs in different accessions, whereas the latter ranged
from 4.76 to 25% (Table 2). The remaining configurations
were generally detected in less than 15% of the PMCs. All
accessions showed a prevalence of bivalent associations,
with averages that ranged from 15.90 to 19.52 II (Table 3).
A low frequency of univalents, trivalents, pentavalents, and
hexavalents was also recorded in some accessions. The
most frequent multivalent association detected was quad-
rivalent (IV), which ranged from one to eight per PMC
(Table 3). The ANOVA showed significant differences for
quadrivalent associations (p = 0.0003, F = 4.48) between
the accessions. Arachis glabrata var. hagenbeckii 30107 and
A. glabrata var. glabrata 2832 had the maximum number of
quadrivalents (five and eight per PMC, respectively). The
former presented from four to six multivalents (quadriva-
lents plus occasional trivalents, pentavalents and hexava-
lents) in 30% of the PMCs analyzed (Table 2). The finding of
several multivalent associations (up to 8 IV) in most of
A. glabrata accessions indicates that its four chromosome
sets have a high degree of homology. Following Stebbins’
criterion (1947), the results here obtained suggest that this
species may probably be an autopolyploid, although a seg-
mental allopolyploid nature cannot be fully ruled out.
Besides the polyploid nature, the accessions of
A. glabrata showed significant differences (Table 3) in the
frequency of bivalents (p = 0.0002, F = 4.55). All the
accessions of A. glabrata var. glabrata (except for 2832)
showed similar values among themselves, but significantly
higher values (18.35–19.52 II) than those of Arachis
glabrata var. hagenbeckii 30107 (15.90 II). Arachis gla-
brata var. glabrata 2832 is between these two groups.
Therefore, these results show that the accessions analyzed
have different degrees of diplodization.
The analyses of chromosome pairing in A. nitida showed
that the most common chromosome configuration was 20II
Table 1 Provenance and collection number of the Arachis glabrata and A. nitida accessions
Species Accession and
collection numberaProvenanceb
A. glabrata var. glabrata SeLaSo2832 Argentina, Prov. Corrientes, Dpto. Ituzaingo, RP 34 to San Carlos, 1 km S from RN 12
SeLaSo 2833 Argentina, Prov. Corrientes, Dpto. Ituzaingo, RP 34 to San Carlos, 12 km S from RN 12
SeLaSo 2840 Argentina, Prov. Misiones, Dpto. Capital, RP 105 km 32.5
SeLaSo 2842 Argentina, Prov. Misiones, Dpto. Candelaria, 5 km SW from RN 12.
SeLaSo 2876 Argentina, Prov. Corrientes, Dpto. Concepcion, RP 117 km 28, 1 km E from Paso Naranjito
A. glabrata var. hagenbeckii KGPSc 30107 Paraguay, Dpto. Paraguarı, 2 km N from Caapucu
A. nitida SvPzHn 3785 Paraguay, Dpto. Amambay, 21 km W from Bella Vista
a G = W.C. Gregory, Hn = R. Heyn, K = A. Krapovickas, La = G. Lavia, P = J.R. Pietrarelli, Pz = E. Pizarro, Se = J.G. Seijo, Sc = A.
Schinini, So = V. Solıs Neffa, Sv = G.P. Silvab Prov = Province, Dpto = Department, RN = National route, RP = Provincial route
Meiotic behavior and pollen viability of tetraploid Arachis glabrata and A. nitida species
123
(65.31% of the PMCs) (Table 2; Fig. 1f). However, one to
three univalents, trivalents, or quadrivalents were also
found in 34.69% of the PMCs (Table 2; Fig. 1g–h), giving
average associations of 0.14 I, 18.78 II, 0.14 III, and 0.45 IV
(Table 3). The finding of up to four multivalents (although
in very few PMCs) suggests that the four chromosome sets
of A. nitida have some degree of homology, although much
lower than those of A. glabrata. According to Stebbins’
criterion (1947), species with the configurations in fre-
quencies like those detected in A. nitida may be considered
as segmental allotetraploids. The average associations
found in A. nitida are comparable to those reported by Singh
(1986) in synthetic amphidiploids involving the A and B
genome species of the section Arachis, where the low
multivalent and high bivalent frequencies have been used as
evidence of their segmental allopolyploid nature. In con-
trast, the low frequency of multivalent associations has been
used as a strong argument to propose a high degree of
diploidization in autotetraploids (Qu et al. 1998; Santos
et al. 2003).
Analysis of meiotic behavior and pollen viability
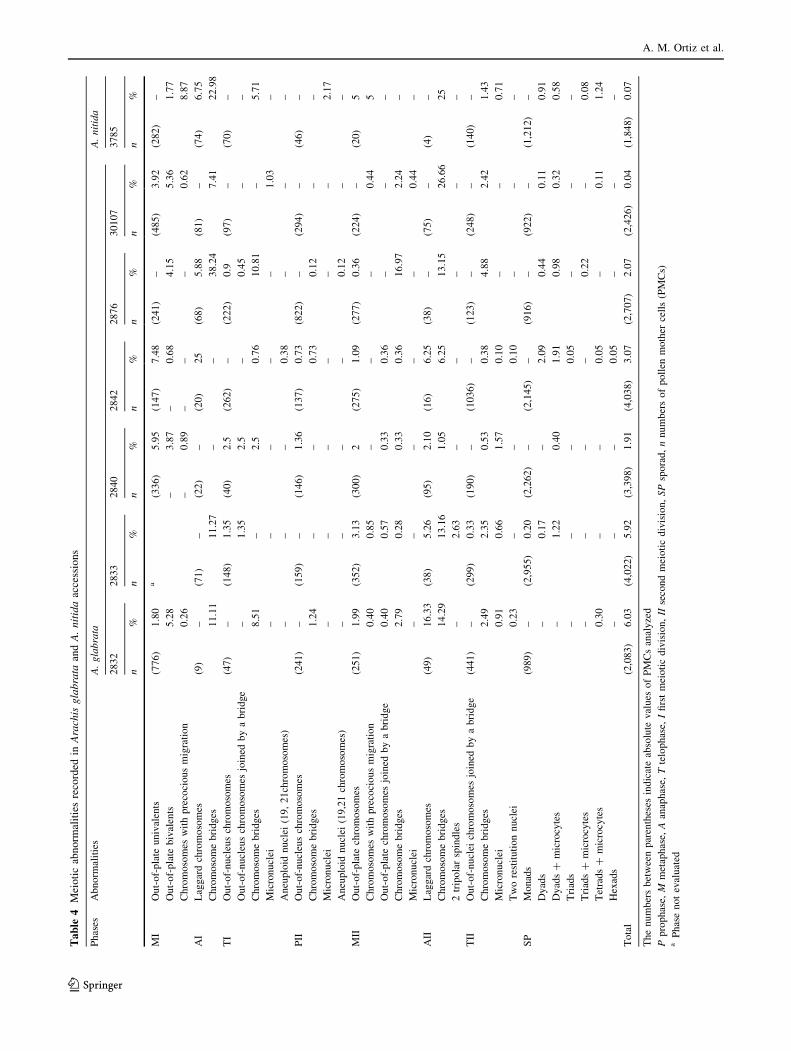
The analysis of chromosome behavior during the meiotic
division in A. glabrata and A. nitida revealed a diversity of
abnormalities from metaphase I to the sporad stage
(Table 4). Meiotic abnormalities were observed from 0.04
to 6.03% of the PMCs of the different A. glabrata acces-
sions and in 0.07% of the PMCs of A. nitida (Table 4).
Taking into account the mean values from A. glabrata
and A. nitida, precocious chromosome migration at meta-
phase I (Fig. 2a) and laggards at anaphases I (Fig. 2b) were
detected in up to 9% and 25% of the PMCs analyzed,
respectively. Chromosome associations at diakinesis-
metaphase I showed that most bivalents were ring-type and
had at least two chiasmata. However, some of them were
rod-type with only one terminal chiasma and usually had
precocious segregation. Therefore, precocious segregation
seems to be directly related to the possession of only one
terminal chiasma per bivalent (Fig. 1b). This finding is in
accordance with Darlington’s criterion (1965), which
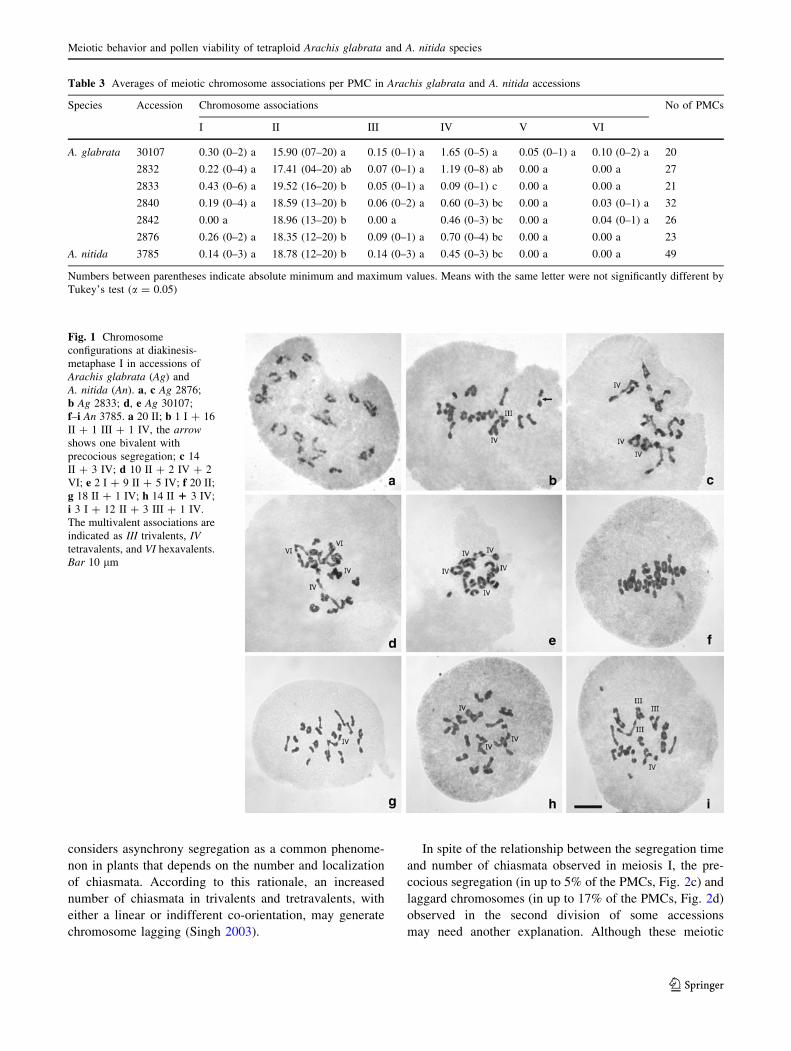
Table 2 Percentages of chromosome configurations at diakinesis and metaphase I
Chromosome configurations A. glabrata A. nitida
2832 2833 2840 2842 2876 30107 3785
20II 55.57 (15) 80.96 (17) 62.50 (20) 73.07 (19) 56.52 (13) 25.00 (5) 65.31 (32)
2I ? 19II – 4.76 (1) – – 8.70 (2) – –
1I ? 18II ? 1III 3.70 (1) – – – – 5.00 (1) –
18II ? 1IV 11.12 (3) 4.76 (1) 15.64 (5) 11.54 (3) 8.70 (2) 25.00 (5) 18.37 (9)
6I ? 17II – 4.76 (1) – – – – –
2I ? 17II ? 2IV – – 3.12 (1) – – – –
1I ? 16II ? 1III ? 1IV – 4.76 (1) – – – 5.00 (1) 4.08 (2)
2I ? 16II ? 2III – – – – 4.34 (1) – –
16II ? 2IV 7.41 (2) – 6.25 (2) 7.69 (2) 8.70 (2) 10.00 (2) 6.12 (3)
15II ? 1IV ? 1 VI – – 3.12 (1) – – – –
4I ? 14II ? 2IV 3.70 (1) – – – – – –
14II ? 3IV 3.70 (1) – 6.25 (2) 3.85 (1) 8.70 (2) – 2.04 (1)
4I ? 13II ? 2III ? 1IV – – 3.12 (1) – – – –
13II ? 2IV ? 1VI – – – 3.85 (1) – – –
3I ? 12II ? 3III ? 1IV – – – – – – 2.04 (1)
2I ? 12II ? 2III ? 2IV – – – – – – 2.04 (1)
1I ? 12II ? 1III ? 3IV 3.70 (1) – – – – – –
12II ? 4IV 3.70 (1) – – – 4.34 (1) 15.00 (3) –
10II ? 2IV ? 2VI – – – – – 5.00 (1) –
10II ? 5IV 3.70 (1) – – – – – –
2I ? 9II ? 5IV – – – – – 5.00 (1) –
2I ? 7II ?1III ? 4IV ? 1V – – – – – 5.00 (1) –
4II ? 8IV 3.70 (1) – – – – – –
Number of PMCs analyzed 27 21 32 26 23 20 49
Numbers between parentheses indicate absolute values of PMCs recorded for each chromosome configurations
A. M. Ortiz et al.
123
considers asynchrony segregation as a common phenome-
non in plants that depends on the number and localization
of chiasmata. According to this rationale, an increased
number of chiasmata in trivalents and tretravalents, with
either a linear or indifferent co-orientation, may generate
chromosome lagging (Singh 2003).
In spite of the relationship between the segregation time
and number of chiasmata observed in meiosis I, the pre-
cocious segregation (in up to 5% of the PMCs, Fig. 2c) and
laggard chromosomes (in up to 17% of the PMCs, Fig. 2d)
observed in the second division of some accessions
may need another explanation. Although these meiotic
Table 3 Averages of meiotic chromosome associations per PMC in Arachis glabrata and A. nitida accessions
Species Accession Chromosome associations No of PMCs
I II III IV V VI
A. glabrata 30107 0.30 (0–2) a 15.90 (07–20) a 0.15 (0–1) a 1.65 (0–5) a 0.05 (0–1) a 0.10 (0–2) a 20
2832 0.22 (0–4) a 17.41 (04–20) ab 0.07 (0–1) a 1.19 (0–8) ab 0.00 a 0.00 a 27
2833 0.43 (0–6) a 19.52 (16–20) b 0.05 (0–1) a 0.09 (0–1) c 0.00 a 0.00 a 21
2840 0.19 (0–4) a 18.59 (13–20) b 0.06 (0–2) a 0.60 (0–3) bc 0.00 a 0.03 (0–1) a 32
2842 0.00 a 18.96 (13–20) b 0.00 a 0.46 (0–3) bc 0.00 a 0.04 (0–1) a 26
2876 0.26 (0–2) a 18.35 (12–20) b 0.09 (0–1) a 0.70 (0–4) bc 0.00 a 0.00 a 23
A. nitida 3785 0.14 (0–3) a 18.78 (12–20) b 0.14 (0–3) a 0.45 (0–3) bc 0.00 a 0.00 a 49
Numbers between parentheses indicate absolute minimum and maximum values. Means with the same letter were not significantly different by
Tukey’s test (a = 0.05)
Fig. 1 Chromosome
configurations at diakinesis-
metaphase I in accessions of
Arachis glabrata (Ag) and
A. nitida (An). a, c Ag 2876;
b Ag 2833; d, e Ag 30107;
f–i An 3785. a 20 II; b 1 I ? 16
II ? 1 III ? 1 IV, the arrowshows one bivalent with
precocious segregation; c 14
II ? 3 IV; d 10 II ? 2 IV ? 2
VI; e 2 I ? 9 II ? 5 IV; f 20 II;
g 18 II ? 1 IV; h 14 II 1 3 IV;
i 3 I ? 12 II ? 3 III ? 1 IV.
The multivalent associations are
indicated as III trivalents, IVtetravalents, and VI hexavalents.
Bar 10 lm
Meiotic behavior and pollen viability of tetraploid Arachis glabrata and A. nitida species
123
Ta
ble
4M
eio
tic
abn
orm
alit
ies
reco
rded
inA
rach
isg
lab
rata
and
A.
nit
ida
acce
ssio
ns
Ph
ases
Ab
no
rmal
itie
sA
.g
lab
rata
A.
nit
ida
28
32
28
33
28
40
28
42
28
76
30
107
37
85
n%
n%
n%
n%
n%
n%
n%
MI
Ou
t-of-
pla
teu
niv
alen
ts(7
76
)1
.80
a(3
36
)5
.95
(14
7)
7.4
8(2
41
)–
(48
5)
3.9
2(2
82
)–
Ou
t-of-
pla
teb
ival
ents
5.2
8–
3.8
7–
0.6
84
.15
5.3
61
.77
Ch
rom
oso
mes
wit
hp
reco
cio
us
mig
rati
on
0.2
6–
0.8
9–
––
0.6
28
.87
AI
Lag
gar
dch
rom
oso
mes
(9)
–(7
1)
–(2
2)
–(2
0)
25
(68
)5
.88
(81
)–
(74
)6
.75
Ch
rom
oso
me
bri
dg
es1
1.1
11
1.2
7–
–3
8.2
47
.41
22
.98
TI
Out-
of-
nucl
eus
chro
moso
mes
(47)
–(1
48)
1.3
5(4
0)
2.5
(262)
–(2
22)
0.9
(97)
–(7
0)
–
Out-
of-
nucl
eus
chro
moso
mes
join
edby
abri
dge
–1.3
52.5
–0.4
5–
–
Ch
rom
oso
me
bri
dg
es8
.51
–2
.50
.76
10
.81
–5
.71
Mic
ron
ucl
ei–
––
––
1.0
3–
An
eup
loid
nu
clei
(19
,2
1ch
rom
oso
mes
)–
––
0.3
8–
––
PII
Out-
of-
nucl
eus
chro
moso
mes
(241)
–(1
59)
–(1
46)
1.3
6(1
37)
0.7
3(8
22)
–(2
94)
–(4
6)
–
Ch
rom
oso
me
bri
dg
es1
.24
––
0.7
30
.12
––
Mic
ron
ucl
ei–
––
––
–2
.17
An
eup
loid
nu
clei
(19
,21
chro
mo
som
es)
––
––
0.1
2–
–
MII
Ou
t-of-
pla
tech
rom
oso
mes
(25
1)
1.9
9(3
52
)3
.13
(30
0)
2(2
75
)1
.09
(27
7)
0.3
6(2
24
)–
(20
)5
Ch
rom
oso
mes
wit
hp
reco
cio
us
mig
rati
on
0.4
00
.85
––
–0
.44
5
Ou
t-of-
pla
tech
rom
oso
mes
join
edb
ya
bri
dg
e0
.40
0.5
70
.33
0.3
6–
––
Ch
rom
oso
me
bri
dg
es2
.79
0.2
80
.33
0.3
61
6.9
72
.24
–
Mic
ron
ucl
ei–
––
––
0.4
4–
AII
Lag
gar
dch
rom
oso
mes
(49
)1
6.3
3(3
8)
5.2
6(9
5)
2.1
0(1
6)
6.2
5(3
8)
–(7
5)
–(4
)–
Ch
rom
oso
me
bri
dg
es1
4.2
91
3.1
61
.05
6.2
51
3.1
52
6.6
62
5
2tr
ipo
lar
spin
dle
s–
2.6
3–
––
––
TII
Ou
t-of-
nu
clei
chro
mo
som
esjo
ined
by
ab
rid
ge
(44
1)
–(2
99
)0
.33
(19
0)
–(1
03
6)
–(1
23
)–
(24
8)
–(1
40
)–
Ch
rom
oso
me
bri
dg
es2
.49
2.3
50
.53
0.3
84
.88
2.4
21
.43
Mic
ron
ucl
ei0
.91
0.6
61
.57
0.1
0–
–0
.71
Tw
ore
stit
uti
on
nu
clei
0.2
3–
–0
.10
––
–
SP
Mo
nad
s(9
89
)–
(2,9
55)
0.2
0(2
,26
2)
–(2
,14
5)
–(9
16
)–
(92
2)
–(1
,21
2)
–
Dy
ads
–0
.17
–2
.09
0.4
40
.11
0.9
1
Dy
ads
?m
icro
cyte
s–
1.2
20
.40
1.9
10
.98
0.3
20
.58
Tri
ads
––
–0
.05
––
–
Tri
ads
?m
icro
cyte
s–
––
–0
.22
–0
.08
Tet
rads
?m
icro
cyte
s0
.30
––
0.0
5–
0.1
11
.24
Hex
ads
––
–0
.05
––
–
To
tal
(2,0
83)
6.0
3(4
,02
2)
5.9
2(3
,39
8)
1.9
1(4
,03
8)
3.0
7(2
,70
7)
2.0
7(2
,42
6)
0.0
4(1
,84
8)
0.0
7
Th
en
um
ber
sb
etw
een
par
enth
eses
ind
icat
eab
solu
tev
alu
eso
fP
MC
san
alyze
d
Pp
rop
has
e,M
met
aphas
e,A
anap
has
e,T
telo
phas
e,I
firs
tm
eio
tic
div
isio
n,
IIse
con
dm
eio
tic
div
isio
n,
SP
spo
rad
,n
nu
mb
ers
of
po
llen
moth
erce
lls
(PM
Cs)
aP
has
en
ot
eval
uat
ed
A. M. Ortiz et al.
123
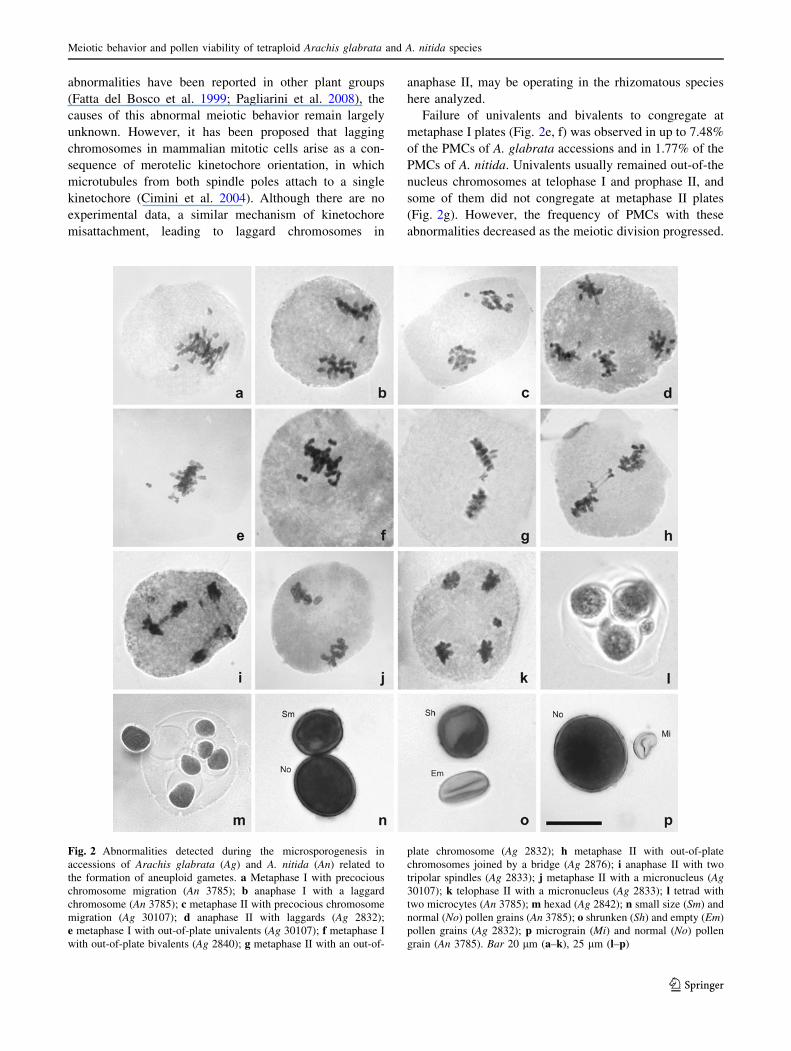
abnormalities have been reported in other plant groups
(Fatta del Bosco et al. 1999; Pagliarini et al. 2008), the
causes of this abnormal meiotic behavior remain largely
unknown. However, it has been proposed that lagging
chromosomes in mammalian mitotic cells arise as a con-
sequence of merotelic kinetochore orientation, in which
microtubules from both spindle poles attach to a single
kinetochore (Cimini et al. 2004). Although there are no
experimental data, a similar mechanism of kinetochore
misattachment, leading to laggard chromosomes in
anaphase II, may be operating in the rhizomatous species
here analyzed.
Failure of univalents and bivalents to congregate at
metaphase I plates (Fig. 2e, f) was observed in up to 7.48%
of the PMCs of A. glabrata accessions and in 1.77% of the
PMCs of A. nitida. Univalents usually remained out-of-the
nucleus chromosomes at telophase I and prophase II, and
some of them did not congregate at metaphase II plates
(Fig. 2g). However, the frequency of PMCs with these
abnormalities decreased as the meiotic division progressed.
Fig. 2 Abnormalities detected during the microsporogenesis in
accessions of Arachis glabrata (Ag) and A. nitida (An) related to
the formation of aneuploid gametes. a Metaphase I with precocious
chromosome migration (An 3785); b anaphase I with a laggard
chromosome (An 3785); c metaphase II with precocious chromosome
migration (Ag 30107); d anaphase II with laggards (Ag 2832);
e metaphase I with out-of-plate univalents (Ag 30107); f metaphase I
with out-of-plate bivalents (Ag 2840); g metaphase II with an out-of-
plate chromosome (Ag 2832); h metaphase II with out-of-plate
chromosomes joined by a bridge (Ag 2876); i anaphase II with two
tripolar spindles (Ag 2833); j metaphase II with a micronucleus (Ag30107); k telophase II with a micronucleus (Ag 2833); l tetrad with
two microcytes (An 3785); m hexad (Ag 2842); n small size (Sm) and
normal (No) pollen grains (An 3785); o shrunken (Sh) and empty (Em)
pollen grains (Ag 2832); p micrograin (Mi) and normal (No) pollen
grain (An 3785). Bar 20 lm (a–k), 25 lm (l–p)
Meiotic behavior and pollen viability of tetraploid Arachis glabrata and A. nitida species
123
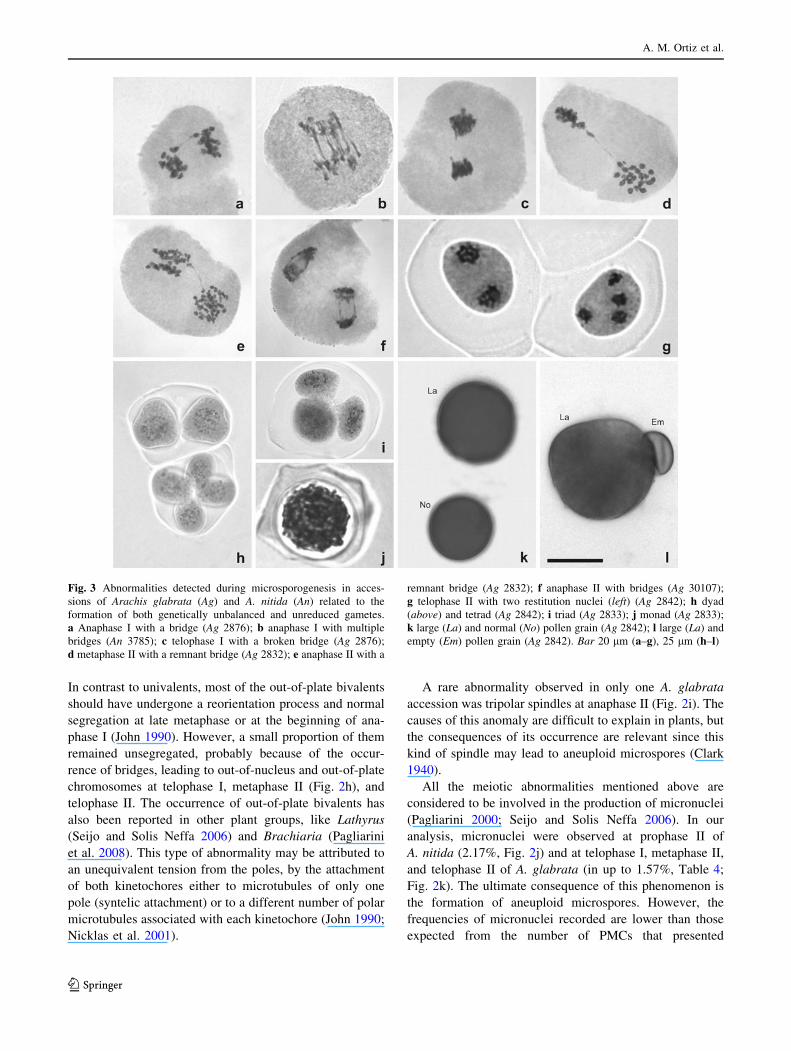
In contrast to univalents, most of the out-of-plate bivalents
should have undergone a reorientation process and normal
segregation at late metaphase or at the beginning of ana-
phase I (John 1990). However, a small proportion of them
remained unsegregated, probably because of the occur-
rence of bridges, leading to out-of-nucleus and out-of-plate
chromosomes at telophase I, metaphase II (Fig. 2h), and
telophase II. The occurrence of out-of-plate bivalents has
also been reported in other plant groups, like Lathyrus
(Seijo and Solis Neffa 2006) and Brachiaria (Pagliarini
et al. 2008). This type of abnormality may be attributed to
an unequivalent tension from the poles, by the attachment
of both kinetochores either to microtubules of only one
pole (syntelic attachment) or to a different number of polar
microtubules associated with each kinetochore (John 1990;
Nicklas et al. 2001).
A rare abnormality observed in only one A. glabrata
accession was tripolar spindles at anaphase II (Fig. 2i). The
causes of this anomaly are difficult to explain in plants, but
the consequences of its occurrence are relevant since this
kind of spindle may lead to aneuploid microspores (Clark
1940).
All the meiotic abnormalities mentioned above are
considered to be involved in the production of micronuclei
(Pagliarini 2000; Seijo and Solis Neffa 2006). In our
analysis, micronuclei were observed at prophase II of
A. nitida (2.17%, Fig. 2j) and at telophase I, metaphase II,
and telophase II of A. glabrata (in up to 1.57%, Table 4;
Fig. 2k). The ultimate consequence of this phenomenon is
the formation of aneuploid microspores. However, the
frequencies of micronuclei recorded are lower than those
expected from the number of PMCs that presented
Fig. 3 Abnormalities detected during microsporogenesis in acces-
sions of Arachis glabrata (Ag) and A. nitida (An) related to the
formation of both genetically unbalanced and unreduced gametes.
a Anaphase I with a bridge (Ag 2876); b anaphase I with multiple
bridges (An 3785); c telophase I with a broken bridge (Ag 2876);
d metaphase II with a remnant bridge (Ag 2832); e anaphase II with a
remnant bridge (Ag 2832); f anaphase II with bridges (Ag 30107);
g telophase II with two restitution nuclei (left) (Ag 2842); h dyad
(above) and tetrad (Ag 2842); i triad (Ag 2833); j monad (Ag 2833);
k large (La) and normal (No) pollen grain (Ag 2842); l large (La) and
empty (Em) pollen grain (Ag 2842). Bar 20 lm (a–g), 25 lm (h–l)
A. M. Ortiz et al.
123
segregational and spindle abnormalities. This fact suggests
that most of the chromosomes irregularly segregated
should have been included at least into one of the main
nuclei. However, those inclusions did not guarantee the
restitution of the euploid chromosome number of the main
nuclei. In this sense, counting at telophase II and prophases
II revealed PMCs having one nucleus with 19 and the other
with 21 chromosomes (Table 4).
Chromosome bridges without fragments were the most
frequent type of abnormality observed in both divisions of
most of the accessions analyzed (Table 4). Anaphase I
bridges (in up to 38.24% of the PMCs; Fig. 3a, b) either
broke at telophase I (Fig. 3c) or persisted as bridges in later
meiotic phases (Fig. 3d, e). Some of the bridges observed
in anaphase-telophase II were between brother poles
(Fig. 3f) and probably formed de novo in the second
division. The origin of bridges without fragments has been
related to failures in the chiasma resolution (Seijo 2002).
According to the modern chromosome model, the origin of
bridges between homologous (anaphase I) or sister chro-
matids (anaphase II) without fragments (all considered as
‘‘x’’ type exchanges in the subchromatid model of chro-
mosomes, Brandham 1970) may rely on failures in the
resolution of the Holliday intermediates or alternatively on
an altered dynamics of the cohesin proteins.
Regardless of the origin of the bridges, a large reduction
in the frequency of chromosome bridges at telophase II was
observed in all accessions (Table 4), suggesting that most
of them were broken along the meiotic division. Thereby,
the main consequence of bridges in these species should be
the production of genetically unbalanced microspores,
containing chromosome deficiencies or duplications.
Alternatively, although in low frequency, the persistence of
some bridges could have led to the generation of restitution
nuclei, such as those observed at telophases II in accessions
2832 and 2842 of A. glabrata (Fig. 3g). This mechanism of
nuclear restitution has been previously reported in other
genera such as Lathyrus (Seijo 2002) and Begonia (Dewitte
et al. 2010). In both cases, these restitution nuclei have
been involved in the origin of unreduced gametes.
All the accessions analyzed here had meiotic indexes
higher than 95%. However, several types of abnormal
sporads were detected (Table 4). These abnormalities
included sporads with additional microcytes, hexads,
monads, dyads, and triads. Microcytes (Fig. 2l) resulted
from the micronuclei observed in the second meiotic divi-
sion, whereas hexads (Fig. 2m) resulted from chromosome
segregation in multipolar spindles. Monads, dyads, and
triads (Fig. 3h–j) could have arisen from the restitution
nuclei induced by persistent bridges.
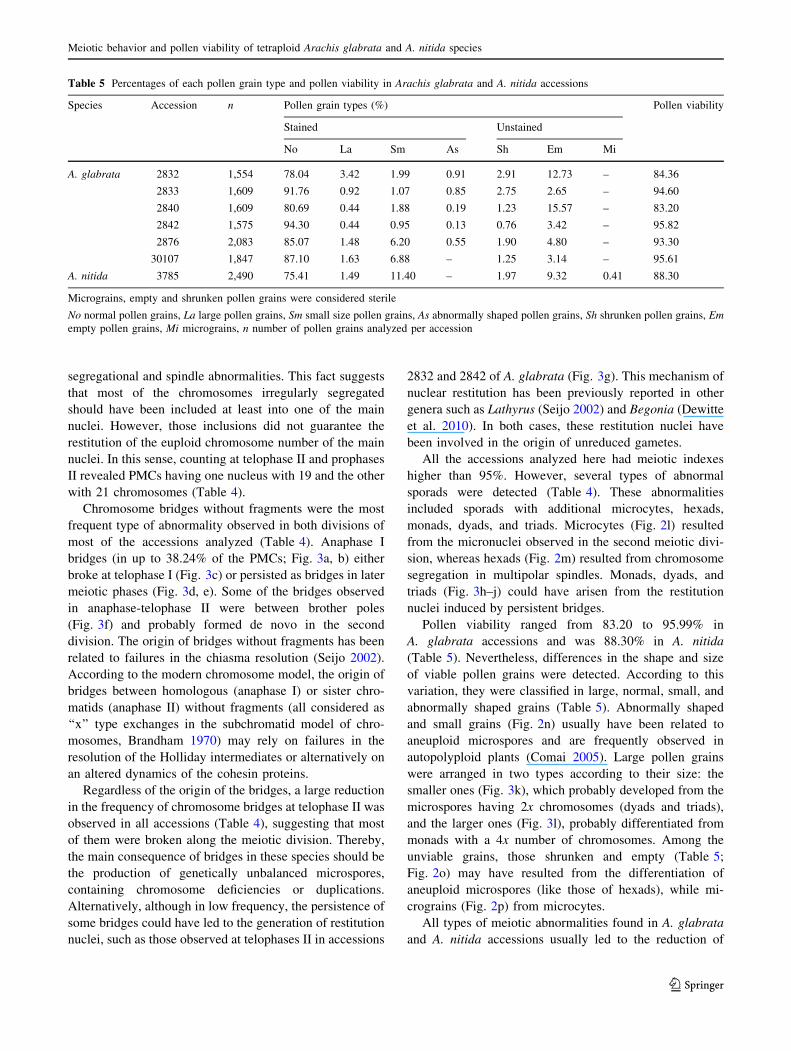
Pollen viability ranged from 83.20 to 95.99% in
A. glabrata accessions and was 88.30% in A. nitida
(Table 5). Nevertheless, differences in the shape and size
of viable pollen grains were detected. According to this
variation, they were classified in large, normal, small, and
abnormally shaped grains (Table 5). Abnormally shaped
and small grains (Fig. 2n) usually have been related to
aneuploid microspores and are frequently observed in
autopolyploid plants (Comai 2005). Large pollen grains
were arranged in two types according to their size: the
smaller ones (Fig. 3k), which probably developed from the
microspores having 2x chromosomes (dyads and triads),
and the larger ones (Fig. 3l), probably differentiated from
monads with a 4x number of chromosomes. Among the
unviable grains, those shrunken and empty (Table 5;
Fig. 2o) may have resulted from the differentiation of
aneuploid microspores (like those of hexads), while mi-
crograins (Fig. 2p) from microcytes.
All types of meiotic abnormalities found in A. glabrata
and A. nitida accessions usually led to the reduction of
Table 5 Percentages of each pollen grain type and pollen viability in Arachis glabrata and A. nitida accessions
Species Accession n Pollen grain types (%) Pollen viability
Stained Unstained
No La Sm As Sh Em Mi
A. glabrata 2832 1,554 78.04 3.42 1.99 0.91 2.91 12.73 – 84.36
2833 1,609 91.76 0.92 1.07 0.85 2.75 2.65 – 94.60
2840 1,609 80.69 0.44 1.88 0.19 1.23 15.57 – 83.20
2842 1,575 94.30 0.44 0.95 0.13 0.76 3.42 – 95.82
2876 2,083 85.07 1.48 6.20 0.55 1.90 4.80 – 93.30
30107 1,847 87.10 1.63 6.88 – 1.25 3.14 – 95.61
A. nitida 3785 2,490 75.41 1.49 11.40 – 1.97 9.32 0.41 88.30
Micrograins, empty and shrunken pollen grains were considered sterile
No normal pollen grains, La large pollen grains, Sm small size pollen grains, As abnormally shaped pollen grains, Sh shrunken pollen grains, Emempty pollen grains, Mi micrograins, n number of pollen grains analyzed per accession
Meiotic behavior and pollen viability of tetraploid Arachis glabrata and A. nitida species
123

pollen viability (Pagliarini 2000; Taschetto and Pagliarini
2004; Diao et al. 2009). In this sense, a major cause of
sterility in auto and segmental allopolyploid species is the
occurrence of irregular chromosomal segregation of univ-
alents and multivalents during meiosis (Stebbins 1947;
Ramsey and Schemske 2002). In spite of the variable fre-
quency of univalent and multivalent associations and of the
different meiotic abnormalities from metaphase I to sporad
stage detected in A. glabrata and in A. nitida accessions, it
seems that these do not contribute to a significant reduction
of pollen viability.
If we assume that pollen viability is a good estimation of
pollen fertility, the low seed production of these species
could not be explained by numerical or structural chro-
mosome aberrations. Therefore, the possible causes of
limited seed production in the species analyzed here must
be looked for in relation to other factors such as: (1)
abnormalities in pistil development (e.g., Teng et al. 2006),
(2) abnormalities in post-fertilization development (e.g.,
Diggle et al. 2002), or (3) self-incompatibility mechanisms
(e.g., Nielsen et al. 2003). The latter appears to be the least
probable phenomenon since, based on their floral structure,
the Arachis species are considered as highly autogamous
(Krapovickas and Gregory 1994).
In summary, the data provided in this report suggest that
A. nitida is either a largely diploidized autopolyploid or a
segmental allopolyploid and that A. glabrata is an auto-
polyploid with different degrees of diploidization. Our
results also suggest that chromosome behavior during
meiosis would not severely affect pollen viability. Thus,
the irregular chromosome behavior caused by the auto-
polyploid or segmental allopolyploid nature of these spe-
cies may not be related with their low seed set production.
Acknowledgments This work was supported by grants from Sec-
retarıa General de Ciencia y Tecnica (UNNE) PI 47/04, Consejo
Nacional de Investigaciones Cientıficas y Tecnicas (CONICET) PIP
6265, and Agencia Nacional de Promocion Cientıfica y Tecnologica
PICTO 2007-00099, Argentina.
References
Angelici C, Hoshino AA, Nobile PM, Palmieri DA, Valls JFM,
Gimenes MA, Lopes CR (2008) Genetic diversity in Rhizo-matosae section of the genus Arachis (Fabaceae) based on
microsatellite markers. Genet Mol Biol 31:79–88
Brandham PE (1970) Chromosome behavior in the Aloineae III
correlations between spontaneous chromatid and sub-chromatid
aberrations. Chromosoma 31:1–17
Cimini D, Cameron LA, Salmon ED (2004) Anaphase spindle
mechanics prevent mis-segregation of merotelically oriented
chromosomes. Curr Biol 14:2149–2155
Clark FJ (1940) Cytogenetics studies of divergent meiotic spindle
formation in Zea mays. Amer J Bot 27:547–559
Comai L (2005) The advantages and disadvantages of being
polyploid. Nat Rev Genet 89:836–846
Darlington CD (1965) Recent advances in cytology. J & A Churchill
Ltd., London
Dewitte A, Eackhaut T, Van Huylenbroeck J, Van Bockstaele E
(2010) Meiotic aberrations during 2n pollen formation in
Begonia. Heredity 104:215–223
Diao WP, Bao SY, Jiang B, Cui L, Qian CT, Chen JF (2009)
Cytogenetic studies on microsporogenesis and male gameto-
phyte development in autotriploid cucumber (Cucumis sativusL.): implication for fertility and production of trisomics. Plant
Syst Evol 279:87–92
Diggle PK, Meixner MA, Carrol AB, Aschwanden CF (2002) Barriers
to sexual reproduction in Polygonum viviparum: a comparative
developmental analysis of P. viviparum and P. bistortoides. Ann
Bot 89:145–156
Dyer AF (1963) The use of lacto-propionic orcein in rapid squash
methods for chromosome preparations. Stain Tech 38:85–90
Fatta Del Bosco S, Tusa N, Conicella C (1999) Microsporogenesis in
a Citrus interspecific tetraploid somatic hybrid and its fusion
parents. Heredity 83:373–377
French EC, Prine GM, Ocumpaugh WR, Rice RW (1994) Regional
experience with forage Arachis in the United States. In: Kerridge
PC, Hardy B (eds) Biology and agronomy of forage Arachis.
CIAT, Cali, pp 169–186
InfoStat (2008) InfoStat version 2008. Grupo InfoStat, FCA,
Universidad Nacional de Cordoba, Argentina
John B (1990) Meiosis. Cambridge University Press, Cambridge
Krapovickas A, Gregory WC (1994) Taxonomıa del genero Arachis(Leguminosae). Bonplandia 8:1–186
Lavia GI, Fernandez A, Seijo JG (2008) Cytogenetic and molecular
evidences on the evolutionary relationships among Arachisspecies. In: Sharma AK, Sharma A (eds) Plant genome:
biodiversity and evolution, vol 1E. Science Publishers, Calcutta,
pp 101–134
Luan L, Wang X, Long WB, Liu YH, Tu SB, Xiao XY, Kong FL
(2009) A comparative cytogenetic study of the rice (Oryza sativaL.) autotetraploid restorers and hybrids. Russ J Genet
45:1074–1081
Maass BL, Ocampo CH (1995) Isozyme polymorphism provides for
germplasm of Arachis glabrata Bentham. Genet Resour Crop
Evol 42:77–82
Mallikarjuna N, Sastri DC (2002) Morphological, cytological and
disease resistance studies of the intersectional hybrid between
Arachis hypogaea L. and A. glabrata Benth. Euphytica
126:161–167
Nicklas RB, Waters JC, Salmon ED, Ward SC (2001) Checkpoint
signals in grasshopper meiosis are sensitive to microtubule
attachment, but tension is still essential. J Cell Sci 114:4173–4183
Nielsen LR, Siegismund HR, Philipp M (2003) Partial self-incom-
patibility in the polyploid endemic species Scalesia affinis(Asteraceae) from the Galapagos: remnants of a self-incompat-
ibility system? Bot J Linn Soc 142:93–101
Pagliarini MS (2000) Meiotic behavior of economically important
plant species: the relationship between fertility and male
sterility. Genet Mol Biol 23:997–1002
Pagliarini MS, Risso-Pascotto C, Souza-Kaneshima AM (2008)
Analysis of meiotic behavior in selecting potential genitors
among diploid and artificially induced tetraploid accessions of
Brachiaria ruziziensis (Poaceae). Euphytica 164:181–187
Pittenger TH, Frolik EF (1951) Temporary mounts for pollen abortion
determinations. Stain Tech 26:181–184
Prine GM, Dunavin LS, Moore JE, Roush RD (1981) ‘‘Florigraze’’
rhizoma peanut: a perennial forage legume. Agr Exp Sta, Univ of
Florida, Gainsville, Circular S-275
Prine GM, Dunavin LS, Glennon RJ, Roush RD (1986) ‘‘Arbrook’’
rhizoma peanut: a perennial forage legume. Agr Exp Sta, Univ of
Florida, Gainsville, Circular S-332
A. M. Ortiz et al.
123

Qu L, Hancock JF, Whallon JH (1998) Evolution in an autopolyploid
group displaying predominantly bivalent pairing at meiosis:
genomic similarity of diploid Vaccinium darrowi and autotetra-
ploid V. corymbosum (Ericaceae). Amer J Bot 85:698–703
Raman VS (1981) Nature of chromosome pairing in allopolyploids of
Arachis and their stability. Cytologia 46:307–321
Ramsey J, Schemske DW (2002) Neopolyploidy in flowering plants.
Annu Rev Ecol Syst 33:589–639
Rouse RE, Roka F, Miavitz-Brown EM (2004) Guide for establishing
perennial peanut as a landscape groundcover. Proc Fla State Hort
Soc 117:289–290
Santos JL, Alfaro D, Sanchez-Moran E, Armstrong SJ, Franklin FCH,
Jones GH (2003) Partial diploidization of meiosis in autotetra-
ploid Arabidopsis thaliana. Genetics 165:1533–1540
Seijo JG (2002) Estudios citogeneticos en especies sudamericanas del
genero Lathyrus, seccion Notolathyrus (Leguminosae). Doctoral
thesis, Facultad de Ciencias Exactas, Fısicas y Naturales,
Universidad de Cordoba, Argentina
Seijo JG, Solis Neffa VG (2006) Cytogenetic studies in the rare South
american Lathyrus hasslerianus Burk. (Leguminosae). Cytologia
71:11–19
Simpson CE, Valls JFM, Miles JM (1994) Reproductive biology and
the potential for genetic recombination in Arachis. In: Kerridge
PC, Hardy B (eds) Biology and agronomy of forage Arachis.
CIAT, Cali, pp 43–52
Singh AK (1986) Utilization of wild relatives in the genetic
improvement of Arachis hypogaea L. Theor Appl Genet
72:433–439
Singh RJ (2003) Plant cytogenetics, 2nd edn. CRC Press, Boca Raton
Stebbins GL (1947) Types of polyploids, their classification and
significance. Adv Genet 1:403–429
Taschetto OM, Pagliarini MS (2004) Meiotic behavior in tetraploid
populations of Pfaffia tuberosa (Amaranthaceae). Acta Bot Croat
63:17–24
Teng N, Chen T, Jin B, Wu X, Huang Z, Li X, Wang Y, Mu X, Li J
(2006) Abnormalities in pistil development result in low seed set
in Leymus chinensis (Poaceae). Flora 201:658–667
Valls JFM, Simpson CE (2005) New species of Arachis (Legumino-
sae) from Brazil, Paraguay and Bolivia. Bonplandia 14:35–64
Valls JFM, Maas BL, Lopez CR (1994) Genetic resources of wild
Arachis and genetic diversity. In: Kerridge PC, Hardy B (eds)
Biology and Agronomy of Forage Arachis. CIAT, Cali, pp 28–40
Meiotic behavior and pollen viability of tetraploid Arachis glabrata and A. nitida species
123




















