
Mechanism of action of chromogranin A on catecholamine release: molecular modeling of the catestatin...
11
Regulatory Peptides 77 (1998) 43–53 Mechanism of action of chromogranin A on catecholamine release: molecular modeling of the catestatin region reveals a b-strand / loop / b- strand structure secured by hydrophobic interactions and predictive of activity a a ,b a ,b a ,b a ,b Igor Tsigelny , Sushil K. Mahata , Laurent Taupenot , Nicholas E. Preece , Manjula Mahata , a a ,b a ,b , * Imran Khan , Robert J. Parmer , Daniel T. O’Connor a Departments of Medicine, Pharmacology, and Chemistry, Center for Molecular Genetics, University of California, San Diego, CA, USA b VA San Diego Healthcare System, San Diego, CA, USA Received 25 March 1998; received in revised form 4 May 1998; accepted 4 May 1998 Abstract A novel fragment of chromogranin A, known as ‘catestatin’ (bovine chromogranin A ), inhibits catecholamine release from 344–364 chromaffin cells and noradrenergic neurons by acting as a non-competitive nicotinic cholinergic antagonist, and may therefore constitute an endogenous autocrine feedback regulator of sympathoadrenal activity. To characterize how this activity depends on the peptide’s structure, we searched for common 3-dimensional motifs for this primary structure or its homologs. Catestatin’s primary structure bore significant (29–35.5% identity, general alignment score 44–57) sequence homology to fragment sequences within three homologs of known 3-dimensional structures, based on solved X-ray crystals: 8FAB, 1PKM, and 2IG2. Each of these sequences exists in nature as a b-strand / loop / b-strand structure, stabilized by hydrophobic interactions between the b-strands. The catestatin structure was stable during molecular dynamics simulations. The catestatin loop contains three Arg residues, whose electropositive side chains form the terminus of the structure, and give rise to substantial uncompensated charge asymmetry in the molecule. A hydrophobic moment plot revealed that catestatin is the only segment of chromogranin A predicted to contain amphiphilic b-strand. Circular dichroism in the far ultraviolet showed substantial (63%) b-sheet structure, especially in a hydrophobic environment. Alanine-substitution mutants of catestatin 351 353 358 established a crucial role for the three central arginine residues in the loop (Arg , Arg , and Arg ), though not for two arginine 125 residues in the strand region toward the amino-terminus. [ I]Catestatin bound to Torpedo membranes at a site other than the nicotinic agonist binding site. When the catestatin structure was ‘docked’ with the extracellular domain of the Torpedo nicotinic cholinergic receptor, it interacted principally with the b and d subunits, in a relatively hydrophobic region of the cation pore extracellular orifice, and the complex of ligand and receptor largely occluded the cation pore, providing a structural basis for the non-competitive nicotinic cholinergic antagonist properties of the peptide. We conclude that a homology model of catestatin correctly predicts actual features of the peptide, both physical and biological. The model suggests particular spatial and charge features of the peptide which may serve as starting points in the development of non-peptide mimetics of this endogenous nicotinic cholinergic antagonist. 1998 Elsevier Science B.V. All rights reserved. Keywords: Adrenal medulla; Chromaffin; Chromogranin A; Catestatin; Nicotinic cholinergic receptor; Mutagenesis; Homology modeling; Molecular dynamics; Alanine scan; Catecholamine; Synthetic peptide; Circular dichroism 1. Introduction * Chromogranin A, a 48-kilodalton acidic, soluble protein Corresponding author. Tel.: 1 1-619-552-8585, ext. 7373; fax: 1 1- 619-552-7549; e-mail: [email protected] [36], is found in the core of catecholamine storage vesicles 0167-0115 / 98 / $ – see front matter 1998 Elsevier Science B.V. All rights reserved. PII: S0167-0115(98)00040-8
-
Upload
greenwichuniversity -
Category
Documents
-
view
0 -
download
0
Transcript of Mechanism of action of chromogranin A on catecholamine release: molecular modeling of the catestatin...

Regulatory Peptides 77 (1998) 43–53
Mechanism of action of chromogranin A on catecholamine release:molecular modeling of the catestatin region reveals a b-strand/ loop/b-strand structure secured by hydrophobic interactions and predictive of
activity
a a ,b a ,b a ,b a ,bIgor Tsigelny , Sushil K. Mahata , Laurent Taupenot , Nicholas E. Preece , Manjula Mahata ,a a ,b a ,b ,*Imran Khan , Robert J. Parmer , Daniel T. O’Connor
aDepartments of Medicine, Pharmacology, and Chemistry, Center for Molecular Genetics, University of California, San Diego, CA, USAbVA San Diego Healthcare System, San Diego, CA, USA
Received 25 March 1998; received in revised form 4 May 1998; accepted 4 May 1998
Abstract
A novel fragment of chromogranin A, known as ‘catestatin’ (bovine chromogranin A ), inhibits catecholamine release from344–364
chromaffin cells and noradrenergic neurons by acting as a non-competitive nicotinic cholinergic antagonist, and may therefore constitutean endogenous autocrine feedback regulator of sympathoadrenal activity. To characterize how this activity depends on the peptide’sstructure, we searched for common 3-dimensional motifs for this primary structure or its homologs. Catestatin’s primary structure boresignificant (29–35.5% identity, general alignment score 44–57) sequence homology to fragment sequences within three homologs ofknown 3-dimensional structures, based on solved X-ray crystals: 8FAB, 1PKM, and 2IG2. Each of these sequences exists in nature as ab-strand/ loop/b-strand structure, stabilized by hydrophobic interactions between the b-strands. The catestatin structure was stable duringmolecular dynamics simulations. The catestatin loop contains three Arg residues, whose electropositive side chains form the terminus ofthe structure, and give rise to substantial uncompensated charge asymmetry in the molecule. A hydrophobic moment plot revealed thatcatestatin is the only segment of chromogranin A predicted to contain amphiphilic b-strand. Circular dichroism in the far ultravioletshowed substantial (63%) b-sheet structure, especially in a hydrophobic environment. Alanine-substitution mutants of catestatin
351 353 358established a crucial role for the three central arginine residues in the loop (Arg , Arg , and Arg ), though not for two arginine125residues in the strand region toward the amino-terminus. [ I]Catestatin bound to Torpedo membranes at a site other than the nicotinic
agonist binding site. When the catestatin structure was ‘docked’ with the extracellular domain of the Torpedo nicotinic cholinergicreceptor, it interacted principally with the b and d subunits, in a relatively hydrophobic region of the cation pore extracellular orifice, andthe complex of ligand and receptor largely occluded the cation pore, providing a structural basis for the non-competitive nicotiniccholinergic antagonist properties of the peptide. We conclude that a homology model of catestatin correctly predicts actual features of thepeptide, both physical and biological. The model suggests particular spatial and charge features of the peptide which may serve as startingpoints in the development of non-peptide mimetics of this endogenous nicotinic cholinergic antagonist. 1998 Elsevier Science B.V. Allrights reserved.
Keywords: Adrenal medulla; Chromaffin; Chromogranin A; Catestatin; Nicotinic cholinergic receptor; Mutagenesis; Homology modeling;Molecular dynamics; Alanine scan; Catecholamine; Synthetic peptide; Circular dichroism
1. Introduction
* Chromogranin A, a 48-kilodalton acidic, soluble proteinCorresponding author. Tel.: 1 1-619-552-8585, ext. 7373; fax: 1 1-619-552-7549; e-mail: [email protected] [36], is found in the core of catecholamine storage vesicles
0167-0115/98/$ – see front matter 1998 Elsevier Science B.V. All rights reserved.PI I : S0167-0115( 98 )00040-8

44 I. Tsigelny et al. / Regulatory Peptides 77 (1998) 43 –53
of chromaffin cells and sympathetic nerves [3,29], from ensure the homogeneous distribution of water molecules.which it is co-released, by exocytosis, with epinephrine After that, MD of the entire system were studied. Snap-and norepinephrine [30]. Chromogranin A is a prohor- shots of trajectories were saved every 0.5 ps.mone, subject to proteolytic cleavage during and aftersecretion [2,10], yielding several biologically active pep- 2.1.2. Electrostatic energy calculationstides, including the mid-molecule fragment pancreastatin Models then underwent calculation of total electrostatic(porcine chromogranin A ), which inhibits glucose-240–288 energy, including Coulombic and solvent energy com-stimulated insulin release from pancreatic islet b-cells [32] ponents. The electrostatic energies were calculated byand parathyroid hormone release from chief cells [11], and solving the linearized Poisson–Boltzmann equation usingthe amino-terminal fragment vasostatin (human chromog- the program DelPhi 2.5 [12], as distributed by Molecularranin A ; [1]), which relaxes vascular smooth muscle1–76 Simulations Inc. The dielectric constant of the solvent was[1] and inhibits parathyroid hormone release [7]). set to 80, and that of the protein interior was set to 4; the
The recently described chromogranin A mid-molecule ionic strength was equivalent to 0.145 M sodium chloride.proteolytic fragment catestatin (bovine chromogranin The Poisson–Boltzmann equation was solved numericallyA ), formed within chromaffin granules, is a potent344–364 ˚on a 0.9 A grid. The ionizable residues Glu, Arg, Lys, andinhibitor of catecholamine release [20], acting as a non- Asp were assigned full charge, as expected at pH 7. Thecompetitive antagonist at the nicotinic cholinergic receptor, Amber partial charge distribution [35] was used to placethe physiologic trigger for catecholamine release from partial charges on all atoms in the system.chromaffin cells and sympathetic axons.
Little is known about the 3-dimensional structure of the2.1.3. Hydrophobic moment plot431 amino acid full-length chromogranin A molecule.
A hydrophobic moment plot (MacVector; Oxford Molec-Although chromogranin A is the best-characterizedular, Oxford, UK) searched chromogranin A’s entiremember of the chromogranin /secretogranin protein family,primary structure for regions suggesting amphiphilic b-it seems to have an overall extended structure [23,28], andsheet, by testing for alternating polar and apolar aminono proteins with known structure have a high level ofacid side chains at 1608 to 1808 intervals [8,9]. At eachhomology to chromogranin A as a whole.residue, results were averaged over an 11-amino-acidTo better understand the basis of catestatin’s catechol-window.amine release-inhibitory effect, we investigated whether its
primary structure was predictive of particular 3-dimension-al conformations. We found several primary structural 2.1.4. Dockinghomologs of catestatin whose 3-dimensional structure was Since catestatin is a non-competitive nicotinic choliner-a consistent b-sheet / loop/b-sheet motif. Such a model for gic antagonist, we evaluated its interaction with its likelycatestatin correctly predicted several of its structural and target on the nicotinic receptor: the extracellular domain inpharmacological features. the region of the cation pore. We have recently homology-
modeled the structure of the extracellular domain of theTorpedo nicotinic receptor [34]. Calculations were done
2. Materials and methods using the Grid-Grid docking program in Insight-II (Molec-˚ular Simulations Inc.). The grid unit size was 1 A on a
2.1. Molecular modeling and computational methods 128 3 128 3 128 matrix for the pentameric nicotiniccholinergic receptor as the stationary target, and catestatin
2.1.1. Homology modeling as the movable ligand. The program used van der WaalsTo perform sequence alignments, we used the programs and electrostatic energies as parameters to minimize
BLAST (for preliminary screening) and FASTA [24] for docking energy.the following pairwise alignments. The program Homol-ogy (Molecular Simulations Inc., San Diego, CA; 1997)
2.2. Experimental procedureswas used to build a catestatin model based on template(homolog) structures found using multi-template correla-tions. After homology modeling, the structure was energy- 2.2.1. Cell culture and catecholamine secretionminimized for 2000 iterations in a water shell (thickness, PC12 rat pheochromocytoma cells [13] were cultured (at
˚10 A) using the steepest-descent method. Then, 300 ps passage 8–20) as previously described [20]. PC12 cells’3molecular dynamics simulations in the same water shell chromaffin granules were pre-loaded with [ H]-L-norepi-
were performed on the peptide structure, using the follow- nephrine, and catecholamine secretion was monitored overing strategy at T 5 300 K. Initially the peptide was fixed a 30-min time course, in the presence or absence of
3and a new water shell was introduced. Molecular dynamics secretagogues (see below), by efflux of [ H]-L-norepineph-(MD) for 10 ps followed by a 1000 iteration energy rine. Tritium counts in release medium and cell lysate wereminimization were then performed on water alone to determined by liquid scintillation, and norepinephrine

I. Tsigelny et al. / Regulatory Peptides 77 (1998) 43 –53 45
secretion was expressed as % of cell total norepinephrine 2.3. Statisticsreleased.
Results are shown as mean values 6 1 standard error.2.2.2. Pharmacology and synthetic peptides Significance was inferred at P , 0.05. Peptide IC values50
The nicotinic cholinergic secretory pathway was trig- for nicotinic cholinergic inhibition were determined bygered by nicotine (Sigma, St. Louis, MO), 1–1000 mM; interpolation (Kaleidagraph) on dose-response curves atpeak secretory responses typically occur at 60–100 mM 0.1, 1, and 10 mM peptide.nicotine. Synthetic peptides (20–100 mmol scale) wereprepared by the solid phase method, using f-moc or t-bocprotection chemistry. Purification was by C-18 RP-HPLC. 3. Results and discussionAuthenticity of the resulting peptides was confirmed bymass spectrometry, using either MALDI (matrix-assisted 3.1. Sequence alignments and homology-modeled tertiarylaser-desorption ionization) or electrospray ionization. structureSome peptides were prepared by small (1 mmol) scalesimultaneous synthesis on pins (PepSet; Chiron, Mel- Fig. 1 shows the multiple sequence alignments built onbourne, Australia), then cleaved and solvent-washed prior the basis of pairwise alignments of the 8FAB (a pepsin-to use. digest Fab fragment of a human myeloma IgG1 immuno-
globulin [26]) 1PKM (cat muscle pyruvate kinase M12.2.3. Circular dichroism [22]) and 2IG2 (human monoclonal IgG1 immunoglobulin
Circular dichroism [37] tested peptide secondary struc- Kol [21]) primary structures with the catestatin region ofture by circularly polarized light rotation in the far chromogranin A. One can see that the overall sequenceultraviolet (195–270 nm), on a Cary instrument, at 0.54 identity of each of these homologs to catestatin is no lessmg/ml peptide concentration. Percent secondary structure than 29% and, in the case of 8FAB, 35.5%. Sequence(a-helix, b-sheet, or random coil) was estimated as de- identity between template proteins in these regions isscribed [4]. similarly congruent, with a high degree of 3-dimensional
structural similarity (Fig. 2): two anti-parallel b-strands125 ˚2.2.4. Equilibrium radioligand binding of [ I]catestatin about 10 A from each other, linked by a loop. This
to Torpedo nicotinic receptor-containing membranes structure is similar to a motif in neuraminidase [5] created[Cys ]-human catestatin (C - from four antiparallel b-strands joined by hairpin loops.0 0
SSMKLSFRARAYGFRGPGPQL) was iodinated on its Such antiparallel motifs frequently contain hydrophobicendogenous tyrosine using the chloramine T method [31] residues within the hairpin regions [5], and the hydro-to a specific activity of | 300 Ci /mmol. The phobic residues’ side chains may stabilize the overall
125[ I]catestatin was purified on a Sep-Pak C-18 silica structure of the motif by ‘hydrophobic collapse’. Thecolumn with a CH CN/H O gradient. The solvent system catestatin region of chromogranin A is thus predicted to3 2
consisted of 0.1% trifluoroacetic acid in water (solvent A) have such an anti-parallel structure.and 0.1% trifluoroacetic acid in 80% CH CN (solvent B). The portion of chromogranin A corresponding to the3
Nicotinic cholinergic receptors were partially purified biologically active catestatin (bovine chromograninfrom Torpedo californica electroplax membranes [18]; the A ) is thus framed by a loop joining two anti-parallel344–364
final preparation contained nicotinic receptors as | 20% of b-sheets, as shown in Fig. 3a, to display the orientation oftotal protein, with 1.32 nmol acetylcholine binding sites / the strands about the connecting loop in the modeled
342 370mg protein, at 8.4 mM concentration. region, bovine chromogranin A (Pro → Pro ;342–370
Receptor-enriched membranes (10 nM receptor final PDRSMRLSFRARGYGFRGPGLQLRRGWRP). Twoconcentration) were incubated at room temperature for 90 predictions might be considered here. Firstly, beforemin in 600 ml of assay buffer (10 mM sodium phosphate catestatin is cleaved from chromogranin A, its constraintpH 7.4, 5 mM EDTA, 0.1% Triton X-100, 0.03 mg/ml within the overall chromogranin A molecule may preventbovine albumin, 3 mM NaN , 100 mM phenylmethyl- its biological activity. Indeed, both we [20] and Simon et3
125sulfonyl fluoride) containing [ I]catestatin (120 nCi / al. [27] have found that the nicotinic cholinergic antagonisttube), in the presence or the absence of unlabelled cates- (i.e. catestatin) activity in chromogranin A is liberated bytatin or the nicotinic agonist carbachol. The incubation was proteolysis. Secondly, if the b-strands flanking the cates-terminated by rapid filtration under vacuum through Whit- tatin region (Fig. 2) are removed, the force constrainingman DE-81 cation exchange filter paper previously equili- the loop from unfolding consists of hydrophobic interac-
348brated with wash buffer (10 mM sodium phosphate buffer tions especially among apolar side chains of Leu ,350 355 360 370 362 364pH 7.4, 0.1% Triton X-100, 0.03 mg/ml bovine albumin). Phe , Tyr , Phe , Pro , Leu , and Leu ; the
Filters were then washed twice with 3 ml of ice-cold wash inter-carbon distances between these side chains are suffi-˚buffer, and counted in an LKB-Wallac Ria-gamma 1274 ciently proximate ( # 4.5 A) to form a hydrophobic ‘node’
counter. (Fig. 3a). Residue pairs whose side chains may so as-
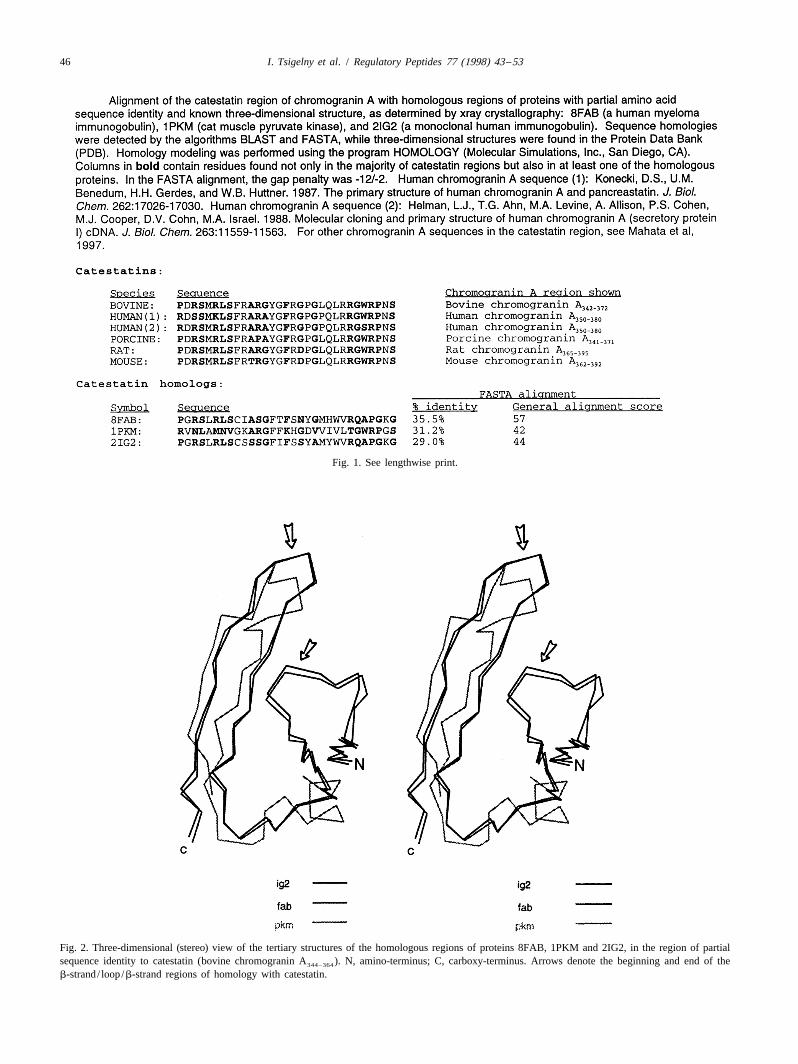
46 I. Tsigelny et al. / Regulatory Peptides 77 (1998) 43 –53
Fig. 1. See lengthwise print.
Fig. 2. Three-dimensional (stereo) view of the tertiary structures of the homologous regions of proteins 8FAB, 1PKM and 2IG2, in the region of partialsequence identity to catestatin (bovine chromogranin A ). N, amino-terminus; C, carboxy-terminus. Arrows denote the beginning and end of the344–364
b-strand/ loop/b-strand regions of homology with catestatin.
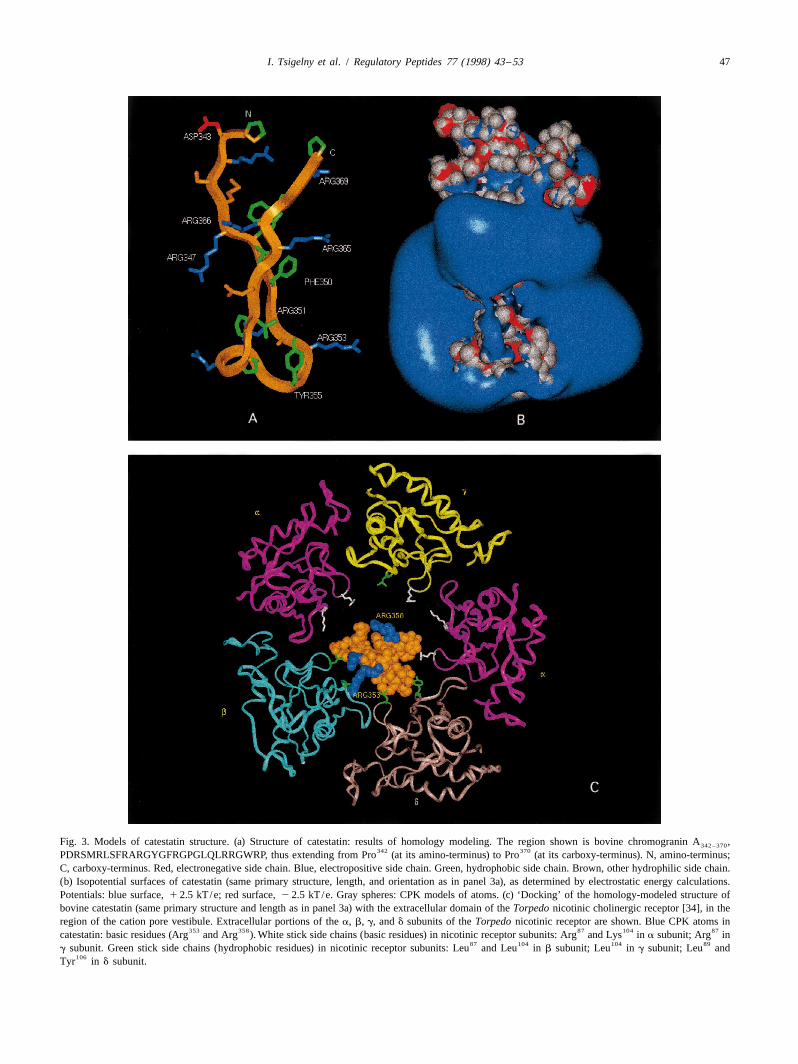
I. Tsigelny et al. / Regulatory Peptides 77 (1998) 43 –53 47
Fig. 3. Models of catestatin structure. (a) Structure of catestatin: results of homology modeling. The region shown is bovine chromogranin A ,342–370342 370PDRSMRLSFRARGYGFRGPGLQLRRGWRP, thus extending from Pro (at its amino-terminus) to Pro (at its carboxy-terminus). N, amino-terminus;
C, carboxy-terminus. Red, electronegative side chain. Blue, electropositive side chain. Green, hydrophobic side chain. Brown, other hydrophilic side chain.(b) Isopotential surfaces of catestatin (same primary structure, length, and orientation as in panel 3a), as determined by electrostatic energy calculations.Potentials: blue surface, 1 2.5 kT/e; red surface, 2 2.5 kT/e. Gray spheres: CPK models of atoms. (c) ‘Docking’ of the homology-modeled structure ofbovine catestatin (same primary structure and length as in panel 3a) with the extracellular domain of the Torpedo nicotinic cholinergic receptor [34], in theregion of the cation pore vestibule. Extracellular portions of the a, b, g, and d subunits of the Torpedo nicotinic receptor are shown. Blue CPK atoms in
353 358 87 104 87catestatin: basic residues (Arg and Arg ). White stick side chains (basic residues) in nicotinic receptor subunits: Arg and Lys in a subunit; Arg in87 104 104 89
g subunit. Green stick side chains (hydrophobic residues) in nicotinic receptor subunits: Leu and Leu in b subunit; Leu in g subunit; Leu and106Tyr in d subunit.
48 I. Tsigelny et al. / Regulatory Peptides 77 (1998) 43 –53
355 360sociate are (in order from the loop tip): Tyr with Pro ;355 362 360 362 357Tyr with Leu ; Pro with Leu ; Phe with362 350 362 350 364 364Leu ; Phe with Leu ; Phe with Leu ; Leu
368 348 364with Trp ; and Leu with Leu .
3.2. Stability of the structure
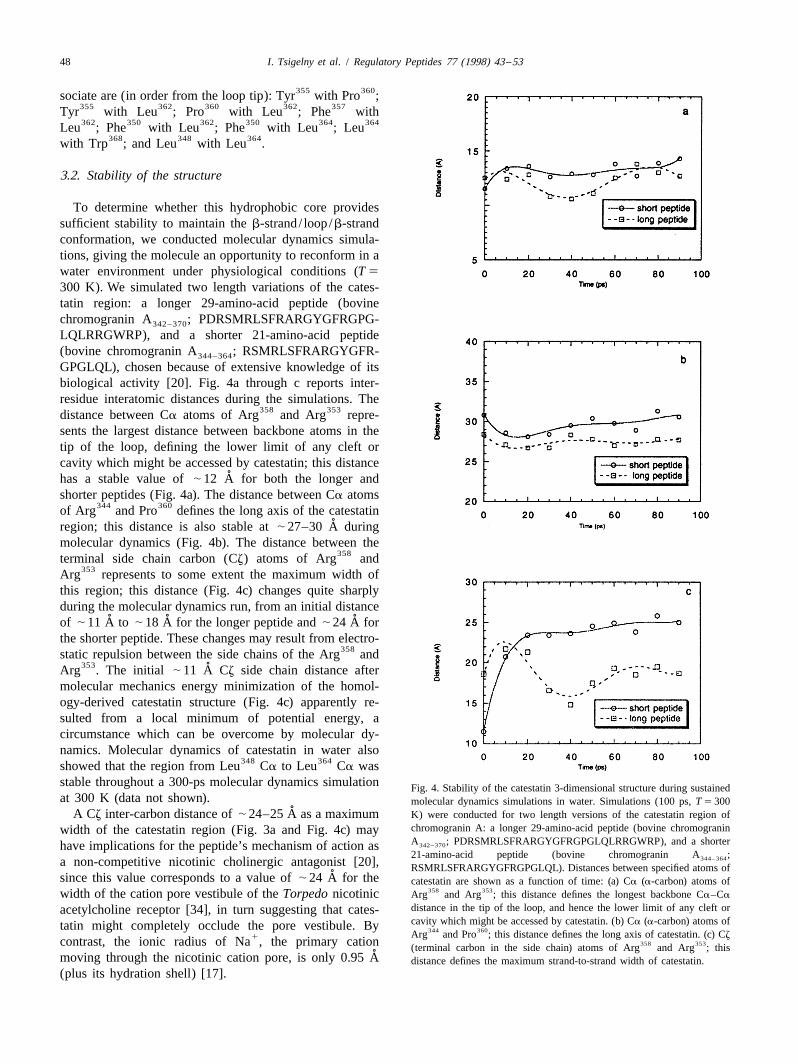
To determine whether this hydrophobic core providessufficient stability to maintain the b-strand/ loop/b-strandconformation, we conducted molecular dynamics simula-tions, giving the molecule an opportunity to reconform in awater environment under physiological conditions (T 5
300 K). We simulated two length variations of the cates-tatin region: a longer 29-amino-acid peptide (bovinechromogranin A ; PDRSMRLSFRARGYGFRGPG-342–370
LQLRRGWRP), and a shorter 21-amino-acid peptide(bovine chromogranin A ; RSMRLSFRARGYGFR-344–364
GPGLQL), chosen because of extensive knowledge of itsbiological activity [20]. Fig. 4a through c reports inter-residue interatomic distances during the simulations. The
358 353distance between Ca atoms of Arg and Arg repre-sents the largest distance between backbone atoms in thetip of the loop, defining the lower limit of any cleft orcavity which might be accessed by catestatin; this distance
˚has a stable value of | 12 A for both the longer andshorter peptides (Fig. 4a). The distance between Ca atoms
344 360of Arg and Pro defines the long axis of the catestatin˚region; this distance is also stable at | 27–30 A during
molecular dynamics (Fig. 4b). The distance between the358terminal side chain carbon (Cz) atoms of Arg and
353Arg represents to some extent the maximum width ofthis region; this distance (Fig. 4c) changes quite sharplyduring the molecular dynamics run, from an initial distance
˚ ˚ ˚of | 11 A to | 18 A for the longer peptide and | 24 A forthe shorter peptide. These changes may result from electro-
358static repulsion between the side chains of the Arg and353 ˚Arg . The initial | 11 A Cz side chain distance after
molecular mechanics energy minimization of the homol-ogy-derived catestatin structure (Fig. 4c) apparently re-sulted from a local minimum of potential energy, acircumstance which can be overcome by molecular dy-namics. Molecular dynamics of catestatin in water also
348 364showed that the region from Leu Ca to Leu Ca wasstable throughout a 300-ps molecular dynamics simulation Fig. 4. Stability of the catestatin 3-dimensional structure during sustainedat 300 K (data not shown). molecular dynamics simulations in water. Simulations (100 ps, T 5 300
˚A Cz inter-carbon distance of | 24–25 A as a maximum K) were conducted for two length versions of the catestatin region ofchromogranin A: a longer 29-amino-acid peptide (bovine chromograninwidth of the catestatin region (Fig. 3a and Fig. 4c) mayA ; PDRSMRLSFRARGYGFRGPGLQLRRGWRP), and a shorter342–370have implications for the peptide’s mechanism of action as21-amino-acid peptide (bovine chromogranin A ;344–364a non-competitive nicotinic cholinergic antagonist [20], RSMRLSFRARGYGFRGPGLQL). Distances between specified atoms of
˚since this value corresponds to a value of | 24 A for the catestatin are shown as a function of time: (a) Ca (a-carbon) atoms of358 353width of the cation pore vestibule of the Torpedo nicotinic Arg and Arg ; this distance defines the longest backbone Ca–Ca
distance in the tip of the loop, and hence the lower limit of any cleft oracetylcholine receptor [34], in turn suggesting that cates-cavity which might be accessed by catestatin. (b) Ca (a-carbon) atoms oftatin might completely occlude the pore vestibule. By 344 360Arg and Pro ; this distance defines the long axis of catestatin. (c) Cz1contrast, the ionic radius of Na , the primary cation 358 353(terminal carbon in the side chain) atoms of Arg and Arg ; this
˚moving through the nicotinic cation pore, is only 0.95 A distance defines the maximum strand-to-strand width of catestatin.(plus its hydration shell) [17].
I. Tsigelny et al. / Regulatory Peptides 77 (1998) 43 –53 49
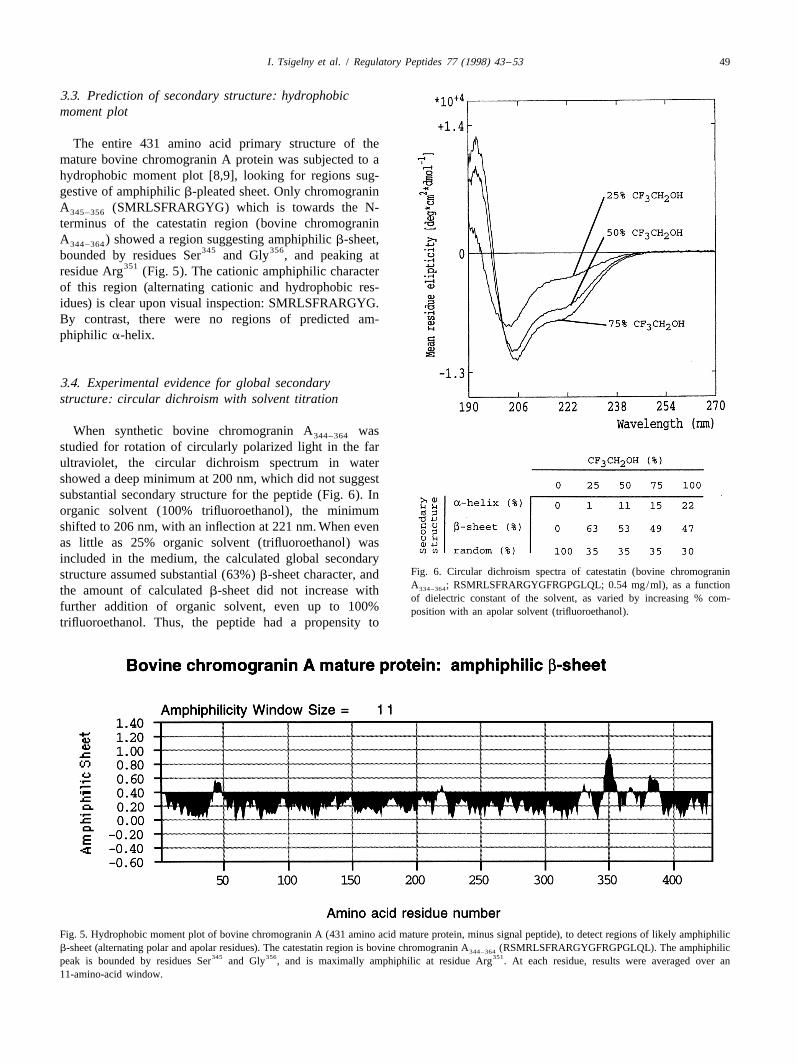
3.3. Prediction of secondary structure: hydrophobicmoment plot
The entire 431 amino acid primary structure of themature bovine chromogranin A protein was subjected to ahydrophobic moment plot [8,9], looking for regions sug-gestive of amphiphilic b-pleated sheet. Only chromograninA (SMRLSFRARGYG) which is towards the N-345–356
terminus of the catestatin region (bovine chromograninA ) showed a region suggesting amphiphilic b-sheet,344–364
345 356bounded by residues Ser and Gly , and peaking at351residue Arg (Fig. 5). The cationic amphiphilic character
of this region (alternating cationic and hydrophobic res-idues) is clear upon visual inspection: SMRLSFRARGYG.By contrast, there were no regions of predicted am-phiphilic a-helix.
3.4. Experimental evidence for global secondarystructure: circular dichroism with solvent titration
When synthetic bovine chromogranin A was344–364
studied for rotation of circularly polarized light in the farultraviolet, the circular dichroism spectrum in watershowed a deep minimum at 200 nm, which did not suggestsubstantial secondary structure for the peptide (Fig. 6). Inorganic solvent (100% trifluoroethanol), the minimumshifted to 206 nm, with an inflection at 221 nm. When evenas little as 25% organic solvent (trifluoroethanol) wasincluded in the medium, the calculated global secondary
Fig. 6. Circular dichroism spectra of catestatin (bovine chromograninstructure assumed substantial (63%) b-sheet character, andA ; RSMRLSFRARGYGFRGPGLQL; 0.54 mg/ml), as a function334–364the amount of calculated b-sheet did not increase withof dielectric constant of the solvent, as varied by increasing % com-
further addition of organic solvent, even up to 100% position with an apolar solvent (trifluoroethanol).trifluoroethanol. Thus, the peptide had a propensity to
Fig. 5. Hydrophobic moment plot of bovine chromogranin A (431 amino acid mature protein, minus signal peptide), to detect regions of likely amphiphilicb-sheet (alternating polar and apolar residues). The catestatin region is bovine chromogranin A (RSMRLSFRARGYGFRGPGLQL). The amphiphilic344–364
345 356 351peak is bounded by residues Ser and Gly , and is maximally amphiphilic at residue Arg . At each residue, results were averaged over an11-amino-acid window.

50 I. Tsigelny et al. / Regulatory Peptides 77 (1998) 43 –53
assume b-sheet conformation with only modest reductionin solvent dielectric constant.
By contrast, an a-helical conformation could beprovoked only by inclusion of very high concentrations oforganic solvent (22% a-helix at 100% trifluoroethanol).
3.5. Predictions of the catestatin model: functionalresults of point mutagenesis
Catestatin (bovine chromogranin A ) inhibits nico-344–364
tinic cholinergic-stimulated PC12 cell catecholamine re-lease, with IC | 200–500 nM peptide [20]. Selective50
alanine substitution (‘alanine scanning’) mutagenesis wasaccomplished by replacement of individual amino acidresidues by alanine (methylglycine), the smallest aminoacid which retains chirality (in this case, [L]-alanine).Alanine scanning revealed important functional roles forthe three central arginine residues in the tip of the
351 353 358catestatin loop (Fig. 3a): Arg , Arg , or Arg . Re-placement of any of these three central arginine residuesby alanine (to R351A, R353A, or R358A) resulted insubstantial increases in catestatin’s IC values (that is,50
decreases in its molar potency) (Fig. 7). By contrast,alanine replacement of the arginine residues in the amino-
344terminal strand region of catestatin (Fig. 3a), Arg (in387R344A) or Arg (in R347A), had little effect on cates-
360tatin’s IC ; nor did replacement of Pro by alanine (in50
P360A) (Fig. 7). Thus, particular amino acids contribute Fig. 7. Point mutants of catestatin: functional results of selective alaninespecifically to catestatin’s function, and residues in the substitutions (alanine scan) on nicotinic cholinergic-stimulated catechol-
amine secretion from PC12 pheochromocytoma cells. IC , the con-loop tip (Fig. 3a) are especially crucial. Lack of effect of 50360 centration of peptide required to inhibit 60 mM nicotine-stimulatedthe P360A mutation suggests that Pro is not absolutely
catecholamine secretion by 50% (derived from dose-response curvesrequired in formation of the turn in the catestatin loop (Fig.determined at 0.1, 1, and 10 mM peptide). Mutant alanine residues are in
7). bold.
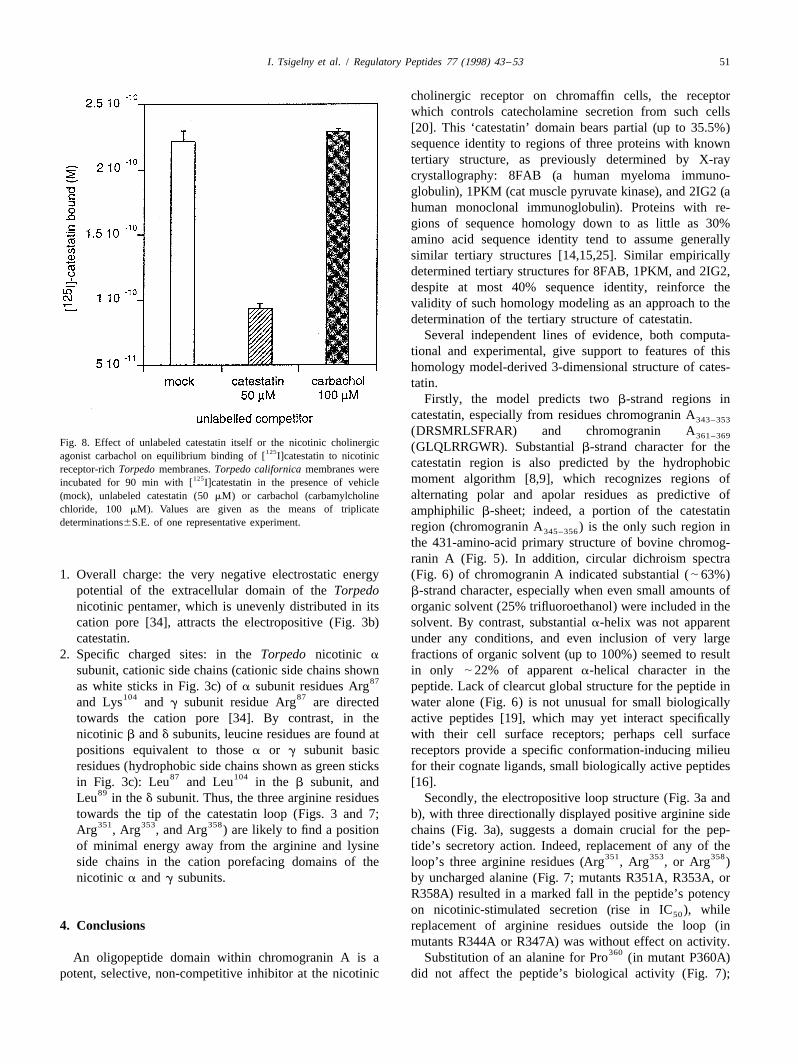
3.6. Binding of catestatin to Torpedo membranes
3.8. Docking of catestatin with the extracellular domain125[ I]Catestatin bound to Torpedo membranes, and an of the nicotinic cholinergic receptorexcess (50 mM) of unlabeled catestatin displaced the tracerfrom the membranes, defining specific binding (Fig. 8). By The directional positive potential of catestatin (Fig. 3b)contrast, the nicotinic agonist carbachol, even at 100 mM, also suggested that catestatin might interact with the125did not displace [ I]catestatin. Thus, catestatin seems to electronegative extracellular domain of the nicotinic re-bind to a site other than the agonist binding site. For such ceptor [34].non-competitive nicotinic cholinergic antagonists [20], that When catestatin was ‘docked’ to the extracellular face ofbinding site is typically in the cation channel vestibule or the Torpedo nicotinic receptor, the energy of interactionpore [6]. was minimized as catestatin approached the extracellular
orifice of the cation pore. In the final state, catestatin most3.7. Electrostatic potential closely approached the b and d subunits of the receptor,
with deep minima of both electrostatic energy and van derElectrostatic field calculations (Fig. 3b) show that a Waals energy (Fig. 3c). In this configuration of minimal
strong maximum of positive potential is located at the tip energy, it can be seen that catestatin almost completely˚of the catestatin loop between two b-strands, largely the occludes the | 24–25 A extracellular orifice of the nico-
351 353 358result of three arginines: Arg , Arg and Arg . This tinic cation pore (Fig. 3c).positive potential suggests a directional orientation of the The final docked position of catestatin on the nicotinicpeptide’s binding to the highly electronegative extracellu- receptor may be influenced by two major charge factors onlar domain of the acetylcholine nicotinic receptor [34]. the receptor.
I. Tsigelny et al. / Regulatory Peptides 77 (1998) 43 –53 51
cholinergic receptor on chromaffin cells, the receptorwhich controls catecholamine secretion from such cells[20]. This ‘catestatin’ domain bears partial (up to 35.5%)sequence identity to regions of three proteins with knowntertiary structure, as previously determined by X-raycrystallography: 8FAB (a human myeloma immuno-globulin), 1PKM (cat muscle pyruvate kinase), and 2IG2 (ahuman monoclonal immunoglobulin). Proteins with re-gions of sequence homology down to as little as 30%amino acid sequence identity tend to assume generallysimilar tertiary structures [14,15,25]. Similar empiricallydetermined tertiary structures for 8FAB, 1PKM, and 2IG2,despite at most 40% sequence identity, reinforce thevalidity of such homology modeling as an approach to thedetermination of the tertiary structure of catestatin.
Several independent lines of evidence, both computa-tional and experimental, give support to features of thishomology model-derived 3-dimensional structure of cates-tatin.
Firstly, the model predicts two b-strand regions incatestatin, especially from residues chromogranin A343–353
(DRSMRLSFRAR) and chromogranin A361–369Fig. 8. Effect of unlabeled catestatin itself or the nicotinic cholinergic (GLQLRRGWR). Substantial b-strand character for the125agonist carbachol on equilibrium binding of [ I]catestatin to nicotinic
catestatin region is also predicted by the hydrophobicreceptor-rich Torpedo membranes. Torpedo californica membranes were125 moment algorithm [8,9], which recognizes regions ofincubated for 90 min with [ I]catestatin in the presence of vehicle
alternating polar and apolar residues as predictive of(mock), unlabeled catestatin (50 mM) or carbachol (carbamylcholinechloride, 100 mM). Values are given as the means of triplicate amphiphilic b-sheet; indeed, a portion of the catestatindeterminations6S.E. of one representative experiment. region (chromogranin A ) is the only such region in345–356
the 431-amino-acid primary structure of bovine chromog-ranin A (Fig. 5). In addition, circular dichroism spectra
1. Overall charge: the very negative electrostatic energy (Fig. 6) of chromogranin A indicated substantial ( | 63%)potential of the extracellular domain of the Torpedo b-strand character, especially when even small amounts ofnicotinic pentamer, which is unevenly distributed in its organic solvent (25% trifluoroethanol) were included in thecation pore [34], attracts the electropositive (Fig. 3b) solvent. By contrast, substantial a-helix was not apparentcatestatin. under any conditions, and even inclusion of very large
2. Specific charged sites: in the Torpedo nicotinic a fractions of organic solvent (up to 100%) seemed to resultsubunit, cationic side chains (cationic side chains shown in only | 22% of apparent a-helical character in the
87as white sticks in Fig. 3c) of a subunit residues Arg peptide. Lack of clearcut global structure for the peptide in104 87and Lys and g subunit residue Arg are directed water alone (Fig. 6) is not unusual for small biologically
towards the cation pore [34]. By contrast, in the active peptides [19], which may yet interact specificallynicotinic b and d subunits, leucine residues are found at with their cell surface receptors; perhaps cell surfacepositions equivalent to those a or g subunit basic receptors provide a specific conformation-inducing milieuresidues (hydrophobic side chains shown as green sticks for their cognate ligands, small biologically active peptides
87 104in Fig. 3c): Leu and Leu in the b subunit, and [16].89Leu in the d subunit. Thus, the three arginine residues Secondly, the electropositive loop structure (Fig. 3a and
towards the tip of the catestatin loop (Figs. 3 and 7; b), with three directionally displayed positive arginine side351 353 358Arg , Arg , and Arg ) are likely to find a position chains (Fig. 3a), suggests a domain crucial for the pep-
of minimal energy away from the arginine and lysine tide’s secretory action. Indeed, replacement of any of the351 353 358side chains in the cation porefacing domains of the loop’s three arginine residues (Arg , Arg , or Arg )
nicotinic a and g subunits. by uncharged alanine (Fig. 7; mutants R351A, R353A, orR358A) resulted in a marked fall in the peptide’s potencyon nicotinic-stimulated secretion (rise in IC ), while50
4. Conclusions replacement of arginine residues outside the loop (inmutants R344A or R347A) was without effect on activity.
360An oligopeptide domain within chromogranin A is a Substitution of an alanine for Pro (in mutant P360A)potent, selective, non-competitive inhibitor at the nicotinic did not affect the peptide’s biological activity (Fig. 7);

52 I. Tsigelny et al. / Regulatory Peptides 77 (1998) 43 –53
perhaps the turn typically induced by a proline residue is membranes. Circular dichroism experiments were per-not essential to the ability of the peptide to form its central formed with the assistance of Joseph Taulane in thecationic loop structure, especially as the peptide ap- laboratory of Murray Goodman, Ph.D., University ofproaches its receptor, where ‘induced fit’ might prevail California, San Diego. The authors acknowledge support[16]. from the Department of Veterans Affairs, National Insti-
The electropositive nature of the catestatin loop structure tutes of Health and the American Heart Association.corresponds to an overall directionality of charge of thecatestatin molecule, as evidenced by the electrostaticpotential calculations (Fig. 3b). Several features suggested Referencesa model for the electrostatic interaction of catestatin withthe extracellular orifice of the nicotinic receptor: (1) [1] Aardal S, Helle KB. The vasoinhibitory activity of bovine chromo-
granin A fragment (vasostatin) and its independence of extracell-catestatin’s pharmacological action as a non-competitiveular calcium in isolated segments of human blood vessels. Regulnicotinic cholinergic antagonist to limit cation flux throughPept 1992;41:9–18.
the nicotinic ligand-gated cation pore [20], (2) the lack of [2] Barbosa JA, Gill BM, Takiyyuddin MA, O’Connor DT. Chromog-competition of a nicotinic agonist for catestatin binding to ranin A: posttranslational modifications in secretory granules. Endo-nicotinic receptors (Fig. 8), and (3) the marked elec- crinology 1991;128:174–90.
[3] Blaschko H, Comline RS, Schneider FH, Silver M, Smith AD.tronegativity of the extracellular domain of the nicotinicSecretion of a chromaffin granule protein, chromogranin, from thereceptor in the region of its cation pore orifice [34].adrenal gland after splanchnic stimulation. Nature 1967;215:58–9.
‘Docking’ of catestatin to the Torpedo nicotinic receptor [4] Bloemendal M, Johnson Jr WC. Structural information on proteinsextracellular domain, with both electrostatic and van der from circular dichroism spectroscopy: possibilities and limitations.Waals energy minima, revealed a complex of catestatin at Pharm Biotechnol 1995;7:65–100.
[5] Branden C, Tooze J. Introduction to protein structure. New York:the orifice of the cation pore, with catestatin’s electroposi-Garland, 1991:302 pp.tive loop closely abutting the b and d subunits of the
[6] Conley EC. The ion channel facts book. I. Extracellular ligand-gatednicotinic receptor, away from those positively charged channels. San Diego: Academic Press, 1996:426 pp.receptor subunit side chains in the vestibule (Fig. 3c); [7] Drees BM, Rouse J, Johnson J, Hamilton JW. Bovine parathyroidindeed, the complex virtually occluded the extracellular glands secrete a 26-kDa N-terminal fragment of chromogranin A
which inhibits parathyroid cell secretion. Endocrinologyorifice of the cation pore, thereby providing a structural1991;129:3381–7.basis for the non-competitive nicotinic antagonist prop-
[8] Eisenberg D, Weiss RM, Terwilliger TC. The hydrophobic momenterties of the peptide [20]. detects periodicity in protein hydrophobicity. Proc Natl Acad Sci
In conclusion, we developed a homology model for the USA 1984;81:140–4.endogenous nicotinic cholinergic antagonist catestatin. The [9] Eisenberg D, Schwarz E, Komaromy M, Wall R. Analysis of
membrane and surface protein sequences with the hydrophobicmodel predicts a central cationic loop tethered by twomoment plot. J Mol Biol 1984;179:125–42.flanking b-strands. Several predictions of the model have
[10] Eskeland NL, Zhou A, Dinh TQ, Wu H, Parmer RJ, Mains RE,been fulfilled by experimental data, including circular O’Connor DT. Chromogranin A processing and secretion: specificdichroism and functional activity of crucial deletion and role of endogenous and exogenous prohormone convertases in thepoint-mutants of the peptide sequence, as well as covalent regulated secretory pathway. J Clin Invest 1996;98:148–56.
[11] Fasciotto BH, Gorr SU, Bourdeau AM, Cohn DV. Autocrinecrosslinking to specific nicotinic subunits. This 3-dimen-regulation of parathyroid secretion: inhibition of secretion bysional structure may serve as a useful guide in designingchromogranin-A (secretory protein-I) and potentiation of secretion
further experiments to probe crucial links between this by chromogranin-A and pancreastatin antibodies. Endocrinologypeptide’s structure and function, and may serve as a 1990;127(3):1329–35.starting point in the development of nonpeptide mimetics [12] Gilson M, Honig B. Calculations of electrostatic potentials in an
enzyme active state. Proteins 1988;4:7–18.of this endogenous nicotinic cholinergic antagonist. In-[13] Greene LA, Tischler AS. Establishment of a noradrenergic clonaldeed, synthetic non-competitive nicotinic cholinergic an-
line of rat adrenal pheochromocytoma cells which respond to nervetagonists (or ganglionic blockers) such as trimethaphan or growth factor. Proc Natl Acad Sci USA 1976;73(7):2424–8.hexamethonium are useful in antihypertensive management [14] Greer J. Comparative modeling of homologous proteins. Methods[33], though their side effects currently preclude wide- Enzymol 1991;202:239–52.
[15] Greer J. Comparative modeling methods: application to the familyspread use; hence, ganglionic blockers with enhancedof the mammalian serine proteases. Proteins 1990;7(4):317–34.potency or selectivity would be attractive.
[16] Gschwend DA, Good AC, Kuntz ID. Molecular docking towardsdrug discovery. J Mol Recogn 1996;9(2):175–86.
[17] Hughes MN. The inorganic chemistry of biological processes, 2ndAcknowledgements edn. New York: Wiley, 1981:259.
[18] Johnson DA, Ayres S. Quinacrine noncompetitive inhibitor bindingsite localized on the Torpedo acetylcholine receptor in the openWe appreciate the assistance of the laboratory of Palmerstate. Biochemistry 1996;35(20):6330–6.
Taylor, Ph.D., University of California, San Diego, in [19] MacKerell Jr AD. Molecular modeling and dynamics of biologicallydesigning these experiments. David Johnson, Ph.D., Uni- active peptides: application to neuropeptide Y. Methods Enzymolversity of California, Riverside, kindly supplied Torpedo 1991;202:449–70.

I. Tsigelny et al. / Regulatory Peptides 77 (1998) 43 –53 53
[20] Mahata SK, O’Connor DT, Mahata M, Yoo SH, Taupenot L, Wu H, protein from chromaffin granules of bovine adrenal medulla. Bio-Gill BM, Parmer RJ. Novel autocrine feedback control of catechol- chem J 1967;103:483–92.amine release: a discrete chromogranin A fragment is a non- [29] Takiyyuddin MA, Cervenka JH, Hsiao RJ, Barbosa JA, Parmer RJ,competitive nicotinic cholinergic antagonist. J Clin Invest O’Connor DT. Chromogranin A: storage and release in hyperten-1997;100:1623–33. sion. Hypertension 1990;15:237–46.
[21] Marquart M, Deisenhofer J, Huber R, Palm W. Crystallographic [30] Takiyyuddin MA, Cervenka JH, Sullivan PA, Pandian MR, Barbosarefinement and atomic models of the intact immunoglobulin mole- JA, O’Connor DT. Is physiologic sympathoadrenal catecholamine
˚ ˚cule Kol and its antigen-binding fragment at 3.0 A and 1.0 A release exocytotic in humans? Circulation 1990;81:185–95.resolution. J Mol Biol 1980;141(4):369–91. [31] Takiyyuddin MA, DeNicola L, Gabbai FB, Dinh TQ, Kennedy B,
[22] Muirhead H, Clayden DA, Barford D, Lorimer CG, Fothergill- Ziegler MG, Sabban EL, Parmer RJ, O’Connor DT. CatecholamineGilmore LA, Schiltz E, Schmitt W. The structure of cat muscle secretory vesicles. Augmented chromogranins and amines in sec-pyruvate kinase. EMBO J 1986;5(3):475–81. ondary hypertension. Hypertension 1993;21:674–9.
[23] O’Connor DT, Frigon RP. Chromogranin A, the major catechol- [32] Tatemoto K, Efendic S, Mutt V, Makk G, Feistner GJ, Barchas JD.amine storage vesicle soluble protein: multiple size forms, subcellu- Pancreastatin, a novel pancreatic peptide that inhibits insulin secre-lar storage and regional distribution in chromaffin and nervous tissue tion. Nature 1986;324:476–8.elucidated by radioimmunoassay. J Biol Chem 1984;259:3237–47. [33] Taylor P. Agents acting at the neuromuscular junction and au-
[24] Pearson WR, Lipman DJ. Improved tools for biological sequence tonomic ganglia. In: Hardman JG, Limbird LE, editors. Goodmancomparison. Proc Natl Acad Sci 1988;85:2444–8. and Gilman’s the pharmacological basis of therapeutics, 9th edn.
[25] Rost B, Sander C. Bridging the protein sequence-structure gap by New York: McGraw-Hill, 1996;Chapter 9:177–97.structure predictions. Annu Rev Biophys Biomol Struct [34] Tsigelny I, Sugiyama N, Sine SM, Taylor P. A model of the1996;25:113–36. nicotinic receptor extracellular domain based on sequence identity
[26] Saul FA, Poljak RJ. Crystal structure of the Fab fragment from the and residue location. Biophys J 1997;73(1):52–66.human myeloma immunoglobulin Igg Hil at 1.8 angstroms res- [35] Wiener SI, Kollman PA, Case D, Singh UC, Glaio C, Alagana G,olution. Unpublished, 1992. Available as file 8FAB at , http: Profeta S, Weiner P. A new force field for molecular mechanical/ /www.pdb.bnl.gov . . simulation of nucleic acids and proteins. J Am Chem Soc
[27] Simon J-P, Bader M-F, Aunis D. Secretion from chromaffin cells is 1984;106:765–84.controlled by chromogranin A-derived peptides. Proc Natl Acad Sci [36] Winkler H, Fischer-Colbrie R. The chromogranins A and B: the firstUSA 1988;85:1712–6. 25 years and future perspectives. Neuroscience 1992;49:497–528.
[28] Smith AD, Winkler H. Purification and properties of an acidic [37] Woody RW. Circular dichroism. Methods Enzymol 1995;246:34–71.




















