
Hoe om vir jouself en u gesin tuis te sorg soos ons na die piek ...
Upload
kalasalingamCategory
view
1download
0
Plant Molecular Biology 29: 125-133, 1995. © 1995 Kluwer Academic Publishers. Printed in Belgium. 125
Rice scutellum induces Agrobacterium tumefaciens vir genes and T-strand generation
K. Vijayachandra, K. Palanichelvam and K. Veluthambi * Department of Plant Molecular Biology, School of Biotechnology, Madurai Kamaraj University, Madurai 625 021, India (* author for correspondence)
Received 2 September 1994; accepted in revised form 26 June 1995
Key words: Agrobacterium tumefaciens, plant transformation, rice, scutellum, T-strands, vir genes
Abstract
For successful transformation of a plant by Agrobacterium tumefaciens it is essential that the explant used in cocultivation has the ability to induce Agrobacterium tumour-inducing (Ti) plasmid virulence (vir) genes. Here we report a significant variation in different tissues of Indica rice (Oryza sativa L. cv. Co43) in their ability to induce Agrobacterium tumefaciens vir genes and T-strand generation, using explants preincubated in liquid Murashige and Skoog (MS) medium. An analysis of rice leaf segments revealed that they neither induced vir genes nor inhibited vir gene induction. Of different parts of rice plants of different ages analysed only scutellum from four-day old rice seedlings induced vir genes and generation of T-strands. We observed that the physical presence of preincubated scutella is required for vir gene induction. Conditioned medium from which preincubated scutella were removed did not induce the vir genes. Scutellum-derived calli, cultured for 25 days on medium containing 2,4-D, also induced virE to an appreciable level. These results suggest that scutellum and scutellum-derived calli may be the most susceptible tissues of rice for Agrobacterium-mediated transformation.
Introduction
Agrobacterium tumefaciens transforms plants by transferring a part of its DNA called transferred DNA (T-DNA) from its tumour-inducing (Ti) plasmid to the plant genome. The virulence (vir) region of the Ti plasmid codes for functions re- quired for processing and transfer of T-DNA [12, 27]. Induction of vir genes by plant signal molecules [25] leads to the generation of three different T-DNA transfer intermediates: double- stranded circular intermediates called T-circles [ 13], single-stranded linear intermediates called T-strands [28, 31 ] and double-stranded linear in- termediates [30].
Compared to dicotyledonous plants, mono- cotyledonous plants are less susceptible to Agrobacterium-mediated transformation. Mono- cotyledonous plants like rice [2, 11], maize [6], Gladiolus [8], Asparagus [10] and Dioscorea [22] have been transformed by Agrobacterium. Yet, unlike Agrobacterium-mediated transformation of dicotyledonous plants, transformation of mono- cotyledonous plants has not become a routine laboratory technique because of lower efficiency of transformation. Recalcitrance of monocotyle- donous plants to Agrobacterium-mediated trans- formation may be because of a block in one or more of the following steps involved in transfor- mation: chemotaxis of Agrobacterium towards

126
wounded plant cells [20], binding of Agrobacte- rium to plant cells [4], induction of vir genes by plant signal molecules [ 25 ], generation of T-DNA transfer intermediates [ 13, 28, 30, 31], transfer of T-DNA transfer intermediates to plant cells and integration of T-DNA into the plant genome [23 ].
There are very few reports of analysis of vir inducers in monocotyledonous plants. Usami et al. [29] have shown that the transition region between shoot and root of wheat and oats pro- duces high molecular weight vir inducer(s). GC-MS analysis of extracts from stem or leaf section-conditioned media of plant species be- longing to different flowering plant families, in- cluding some of the monocotyledonous plant families [24] showed the presence of vir-inducing phenolics. Of these monocotyledonous plants, Asparagus (Liliaceae) produces vanillin and me- thyl vanillate while Dioscorea (Dioscoreaceae), Zingiber (Zingiberaceae), Zea and Lolium (Poaceae) produce syringic acid, ferulic acid, si- napic acid and vanillic acid. Messens et al. [17] found that Triticum monococcum suspension culture, recalcitrant to Agrobacterium-mediated transformation, produces a potent vir inducer, ethyl ferulate. Xu etal. [32] analysed extracts from different parts of rice plants and found that extracts from leaves of panicle-differentiating plants induce vir genes better than the extracts from all the other parts analysed. Apart from vir inducers, monocotyledonous plants are known to contain vir inhibitors. Sahi et al. [21] found that homogenates of maize contain 2,4-dihydroxy-7- methoxy-2H-1,4-benzoxazin-3 (4H)-one which inhibits the growth of Agrobacterium and vir gene induction.
We have standardised a simple leaf segment system to analyse the ability of plants to induce vir genes and generation of T-DNA transfer in- termediates (Vijayachandra et al., manuscript in preparation). Using this system, we assayed the ability of rice plants to induce vir genes. Of vari- ous parts of rice tested, scutellum from four-day old rice seedlings was the only part that induced vir genes and generation ofT-strands. Scutellum- derived calli cultured on N6 medium with 2,4-D, reported to be highly susceptible to Agrobacte-
rium-mediated transformation by Hiei et al. [ 11 ], also induced virE to an appreciable level.
Materials and methods
Plant materials
Tobacco (Nicotiana tabacum L. cv. Wisconsin 38) leaves were taken from plants propagated axeni- cally on BGS medium which contains MS salts [19], 0.4 mg/1 thiamine hydrochloride, 100 mg/l myoinositol, 0.001 mg/1 folic acid, 0.057/~M IAA, 0.14/~M kinetin, 3~o (w/v) sucrose and 0.8~o (w/v) agar. Seeds of Indica rice (Oryza sativa L. cv. Co43) were obtained from Tamilnadu Agri- cultural University, Coimbatore, India. Rice seeds were dehusked and surface-sterilized by the following treatments: (1) a wash with 70~o etha- nol (v/v) for 2 min followed by a wash with sterile water; (2) a wash with 4~o (active chlorine) so- dium hypochlorite with a drop of Tween 80 for 15 min followed by a wash with sterile water; (3) a wash with 0.1 ~o mercuric chloride for 5 rain followed by five washes with sterile water. Seeds were germinated on a medium with Murashige and Skoog (MS) salts [19], 3~o sucrose (w/v) and 0.9~o agar in a tissue culture room set with 16 h light and 8 h dark photoperiod at 25 °C. Explants were taken from seedlings grown for different time periods and preincubated in the in- duction medium. The induction medium is a modified MS medium containing MS salts, 3~o sucrose, 4 #M benzylaminopurine, 0.5/~M naph- thaleneacetic acid, 0.1 mg/l nicotinic acid, 1 mg/l thiamine hydrochloride, 10mg/l myoinositol and 12.5mM phosphate buffer (K2HPO4- NaH2PO4), pH 5.6. Scutella were taken from four-day old seedlings, when the size of scutellum reaches the maximum. Endosperm was removed from four-day old seedlings and the scutella were excised by separating them carefully from the por- tions containing shoot, root and the embryonic axis.
Two media, 2N6 and MS9D, were used for getting scutella from seeds germinated on me- dium containing 2,4-D and for raising scutellum-

derived calli. The 2N6 medium prepared accord- ing to Hiei etal. [11] contains N6 salts and vitamins, 3~o sucrose, 9 ~ M 2,4-D, 0 .9~ casamino acids and 0 .8~ agar, pH 5.8. MS9D medium contains MS salts and vitamins, 3~o su- crose, 9/~M 2,4-D, 0.9~o casamino acids and 0.8~o agar, pH 5.8. Seeds were germinated on these media in dark. Scutella were excised after 5 days and kept for preincubation in induction medium for 72h. After 21 days of growth, scutellum-derived calli were subcultured on the same medium after cutting into 4 to 6 pieces and kept in dark for four days. Healthy parts from these calli were cut approximately into 2 mm seg- ments and kept for 72 h of preincubation in in- duction medium.
Bacterial strains and plasmids
Agrobacterium tumefaciens strain A348 harbour- ing a cosmid clone pSM358 (virE-lacZ) and strain A348 (pTi234) which is a homogenote for virB- lacZ were used to analyse vir gene induction [26]. A. tumefaciens strain A348 with a cosmid clone pVK224 (with extra copies of virG, C, D and E) was used to analyse T-strand generation [31]. A. tumefaciens strain A136 which is an avirulent derivative of strain C58 and strain A348 with the cosmid pVK224 were used to analyse Vir pro- teins. An Escherichia coli strain harbouring plas- mid pDG2 was used as a source of Hind III frag- ment 1 of pTiA6 which was used as a probe in T-strand analysis.
Induction of Agrobacterium
Agrobacterium was grown to 1 OD at 600 nm in AB medium [16] at 28 °C and 200 rpm shaking. The culture was centrifuged for 15min at 5000 × g. The pellet was resuspended in the same volume of induction medium. To 15 ml of induc- tion medium containing 60/~M acetosyringone (AS) or preincubated plant parts, 5 ml of bacte- rial cells were added. Explants from aseptically grown rice plants were cut in Petri plates with
127
15 ml medium and 25 explants each were trans- ferred after 30 min to 100 ml Erlenmeyer flasks with 15 ml MS medium. These flasks were kept in the tissue culture room without shaking for indicated time periods, after which the bacteria were added and kept on a shaker for induction. The standard induction condition used was 120 rpm shaking at 25 °C for 24 h with an initial bacterial density of 0.25 OD at 600 nm. After 24 h induction the cells were pelleted and taken for fl-galactosidase assay [18], for total DNA isola- tion [ 30] or for [ 35S ]methionine incorporation to analyse proteins.
Protein analysis
To radiolabel the Vir proteins, 4/~Ci of [35S]me- thionine (specific activity 400 Ci/mmol supplied by BRIT, Bombay) was added to 20 ml of the induced Agrobacterium culture and kept shaking at 120 rpm for 30 min at 25 °C. Cells were pel- leted and washed with 1 ml of ice-cold unlabelled methionine (1 mM). The cell pellet was resus- pended in 150/~1 of ice-cold sterile distilled water and an equal volume of 2 x lysis buffer [14] was added. Samples were boiled for 5 min in a boil- ing water bath and centrifuged for 10 min in a microfuge. Supernatants were transferred to fresh tubes and trichloroacetic acid-precipitable counts were measured. Samples with equal radioactivity (25 000 cpm)were analysed on 8 to 16 ~o gradient SDS-polyacrylamide gels [ 14]. Gels were stained with Coomassie brilliant blue R-250 and sub- jected to fluorography followed by autoradiogra- phy [ 1].
T-strand analysis
Agrobacterium D N A concentration was estimated fluorometrically using Hoechst dye 33258 in a Hoefer fluorometer as per the instructions of the manufacturer. One ktg of each D N A sample was fractionated by electrophoresis through a 0.7~o agarose gel in Tris/boric acid/EDTA buffer. Non- denaturing Southern transfer was performed as

128
described earlier [31]. Radiolabelled Hind III fragment 1 of pTiA6 was used as probe to detect T-strands.
Results
Induction ofvir genes
We analysed the vir inducibility of rice plants using a simple leaf segment system standardised by us for analysing the ability of plants to induce vir genes (Vijayachandra et al., manuscript in prepa- ration). Induction of vir genes was monitored using Agrobacterium strains harbouring cosmid clones with vir-lacZ fusions. Level of fl-galactosidase activity in the induced bacterial cells is a measure of expression of the vir genes. Rice leaf segments preincubated for different time periods were assayed for their ability to induce virE and virB genes (Table 1). Induction of vir genes was very low with rice leaf segments. The maximum level of induction was achieved with 72 h preincubated rice leaf segments and it was only 5 ~o of the level of induction achieved with tobacco leaf segments. These results show that
Table 1. Int roduct ion of Agrobacterium vir genes by rice leaf
segments .
Strain Preincubat ion M ean fl-galactosidase period (h) units + s tandard error l
A348(pSM358) 2 24 74 + 4 48 151 + 17 72 223 + 30 96 156 + 21
A348(pTi234) 3 24 11+ 1 48 24 + 1 72 4 1 + 1 96 36 _+ 1
Values are m e a n s o f six assays . 2 Strain A348(pSM358) is meroploid for virE-lacZ fusion.
Control values for this strain: M S medium, 66 + 3; 60 # M AS, 5115 + 152; tobacco leaf segments pre- incubated for 48 h, 4424+ 177.
3 Strain A348(pTi234) is a homogeno te for virB-lacZ. Control values for this strain: M S medium, 6 + 1; 60/~M AS 208 + 8; tobacco leaf segments pre- incubated for 48 h, 303 + 19.
rice leaves do not produce appreciable levels of vir inducers. We assayed whether rice leaves pro- duce inhibitor s of vir gene induction. A S-mediated virE induction was performed in the absence and in the presence of rice leaf segments. Rice leaf segments did not inhibit AS-mediated virE induc- tion (Table 2).
Various parts of rice plants of different ages were analysed for their ability to induce the virE gene (Table 3). The level of induction achieved with 72 h preincubated rice roots, leaf bases and leaf segments of different ages was very low. Only scutellum from four-day old rice seedlings prein- cubated for 72 h induced virE to a high level (ca. 40 ~o of induction by tobacco). We have ob- served that vir induction with freshly cut tobacco leaf segments was very low and a 48 to 72 h pre- incubation of leaf segments was required to achieve the maximum level of vir induction (Vi- j ayachandra et al., manuscript in preparation). To find out whether pre-incubation in MS medium was required for rice scutellum to induce vir genes, virE induction was performed with freshly excised scutella and 72 h preincubated scutella (Table 4). Freshly excised scutella did not induce virE, whereas 72 h pre-incubated scutella induced virE very well. We analysed the scutellum-conditioned medium (medium in which scutella were pre- incubated for 72 h and were removed at the end
Table 2. Effect of rice leaf segments on AS-media ted induc- tion o f Agrobacterium virE gene. 1,2
Plant leaf segments / AS Mean fl-galactosidase pre- incubat ion period (h) (/~M) units + s tandard error 3
- - 1 8 2 + 45 - 60 1899 _+ 366 Tobacco/40 - 4182 + 80 Tobacco/40 60 3840 + 97
Rice/40 - 47 + 4 Rice/40 60 1749 + 89 Rice/64 - 188 + 37 Rice/64 60 1623 + 105
1 Induct ion was done with A. tumefaciens strain A348(pSM358) .
2 Induct ion condit ions: 100 rpm shaking at 20 to 25 °C for 24 h.
3 Values are m e a n s o f six assays.
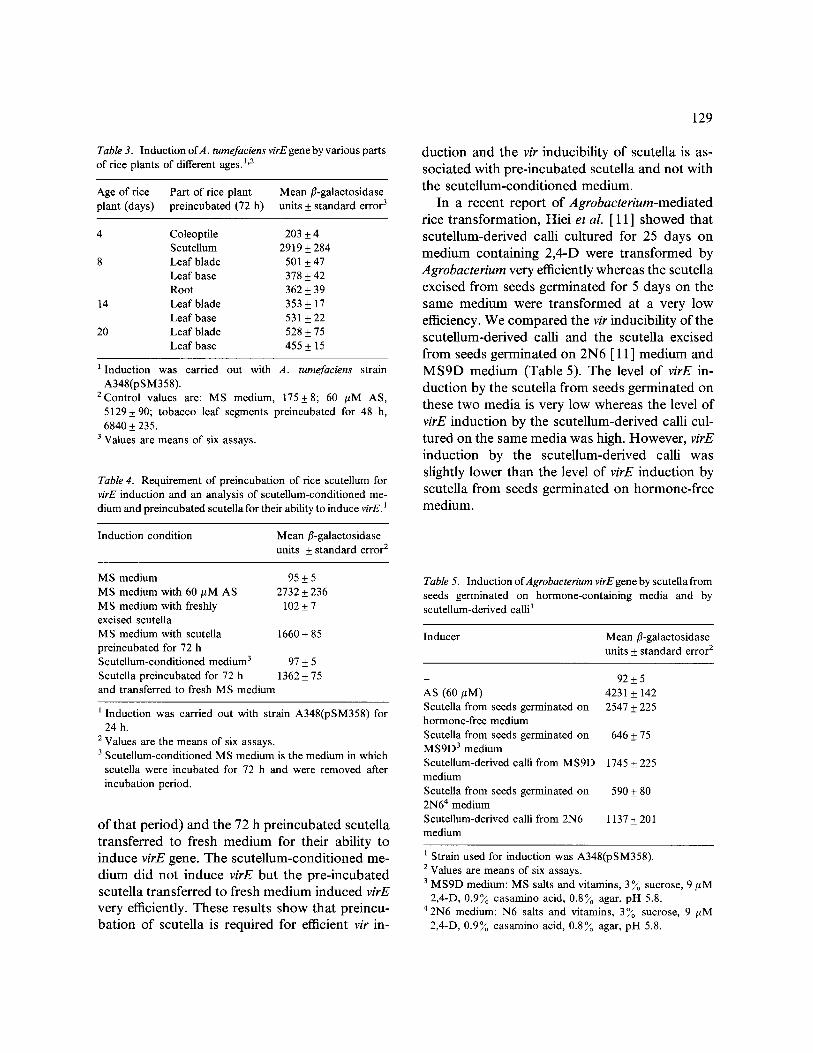
Table 3. Induction ofA. tumefaciens virE gene by various parts of rice plants of different ages. 1'2
Age of rice Part of rice plant Mean fl-galactosidase plant (days) preincubated (72 h) units + standard error 3
4 Coleoptile 203 + 4 Scutellum 2919 + 284
8 Leaf blade 501 + 47 Leaf base 378 + 42 Root 362 + 39
14 Leaf blade 353 + 17 Leaf base 531 + 22
20 Leaf blade 528 + 75 Leaf base 455 + 15
l Induction was carried out with A. tumefaciens strain A348(pSM358).
2Control values are: MS medium, 175 + 8; 60 /~M AS, 5129+ 90; tobacco leaf segments preincubated for 48 h, 6840 + 235.
3 Values are means of six assays.
Table 4. Requirement of preincubation of rice scutellum for virE induction and an analysis of scutellum-conditioned me- dium and preincubated scutella for their ability to induce virE. 1
Induction condition Mean fl-galactosidase units + standard error 2
MS medium 95 + 5 MS medium with 60/~M AS 2732 + 236 MS medium with freshly 102 + 7 excised scutella MS medium with scutella 1660 _+ 85 preincubated for 72 h Scutellum-conditioned medium 3 97 _+ 5 Scutella preincubated for 72 h 1362 + 75 and transferred to fresh MS medium
i Induction was carried out with strain A348(pSM358) for 24 h.
2 Values are the means of six assays. 3 Scutellum-conditioned MS medium is the medium in which
scutella were incubated for 72 h and were removed after incubation period.
of that period) and the 72 h preincubated scutella transferred to fresh medium for their ability to induce virE gene. The scutellum-conditioned me- dium did not induce virE but the pre-incubated scutella transferred to fresh medium induced virE very efficiently. These results show that preincu- bation of scutella is required for efficient vir in-
129
duction and the vir inducibility of scutella is as- sociated with pre-incubated scutella and not with the scutellum-conditioned medium.
In a recent report of Agrobacterium-mediated rice transformation, Hiei et al. [11] showed that scutellum-derived calli cultured for 25 days on medium containing 2,4-D were transformed by Agrobacterium very efficiently whereas the scuteUa excised from seeds germinated for 5 days on the same medium were transformed at a very low efficiency. We compared the vir inducibility of the scutellum-derived calli and the scutella excised from seeds germinated on 2N6 [ 11 ] medium and MS9D medium (Table 5). The level of virE in- duction by the scutella from seeds germinated on these two media is very low whereas the level of virE induction by the scutellum-derived calli cul- tured on the same media was high. However, virE induction by the scutellum-derived calli was slightly lower than the level of virE induction by scutella from seeds germinated on hormone-free medium.
Table5. Induction of Agrobacterium virE gene by scutella from seeds germinated on hormone-containing media and by scutellum-derived calli 1
Inducer Mean fl-galactosidase units __ standard error 2
- 92+5 AS (60/~M) 4231 + 142 Scutella from seeds germinated on 2547 _+ 225 hormone-free medium Scutella from seeds germinated on 646 _+ 75 MS9D 3 medium Scutellum-derived calli from MS9D 1745 +_ 225 medium Scutella from seeds germinated on 590 + 80 2N64 medium Scutellum-derived calli from 2N6 1137 +_ 201 medium
I Strain used for induction was A348(pSM358). 2 Values are means of six assays. 3 MS9D medium: MS salts and vitamins, 3~o sucrose, 9 ~tM
2,4-D, 0.9~o casamino acid, 0.8~o agar, pH 5.8. 42N6 medium: N6 salts and vitamins, 3~o sucrose, 9 /~M
2,4-D, 0.9~o casamino acid, 0.8~o agar, pH 5.8.

130
Induction of vir proteins
Radiolabelled proteins from uninduced A. tume- faciens strain A136 and strain A348(pVK224) and bacteria induced with AS, tobacco leaf segments, rice leaf segments, rice leaf bases, rice roots or rice scutella were analysed on SDS-polyacryla- mide gels (Fig. 1). Proteins of molecular masses 82, 69, 65, 59, 57, 30, 27, 23, 16 and 12 kDa were induced by tobacco leaf segments and AS. Of the different parts of rice analysed, only scutella from four-day old seedlings induced Vir protein syn- thesis. The amount of Vir proteins synthesised in the presence of rice scutella was much lower than the amount of Vir proteins synthesized in the
presence of AS or tobacco leaf segments. Rice scutella induced the synthesis of 69, 65, 57, 27, 23 and 16 kDa proteins. The 82, 59, 30 and 12 kDa proteins which were induced by AS and tobacco leaf segments were not induced by rice scutellum. Additional experiments would have to be done to understand the basis and significance of differen- tial pattern of Agrobacterium protein synthesis in response to tobacco and rice. Molecular weights of these proteins matched with those of many known Vir proteins [5 ]. These proteins were not induced in strain A136 which lacks a Ti plasmid (data not shown).
Fig. 1. Analysis of Vir proteins induced in Agrobacterium by rice. [35S]Methionine-labelled protein samples from unin- duced and induced Agrobacterium strain A348 (pVK224) were analysed by SDS-PAGE. Protein samples from uninduced bacteria (lane 1) and bacteria treated with 60/~M AS (lane 2), tobacco leaf segments (lane 3), rice scutella (lane 4), rice co- leoptiles (lane 5), rice leaf segments (lane 6) or rice roots (lane 7) were analysed. Tobacco leaf segments were pre- incubated for 48 h and rice explants were pre-incubated for 72 h. Equal amount of radioactivity (25 000 cpm) was loaded in each lane.
Fig. 2. Analysls otT-strands generated mAgrobacterium upon induction by rice. DNA samples from uninduced and induced A. tumefaciens strain A348 (pVK224) were subjected to non- denaturing Southern analysis using Hind III fragment 1 of pTiA6 as probe. DNA samples from control bacteria (lane 1) and bacteria induced with AS (lane 2), tobacco leaf segments (lane 3), rice coleoptiles (lane4), rice roots (lane 5), rice scutella (lane 6), or rice leaf segments (lane 7) were used for analysis. Tobacco leaf segments were pre-incubated for 48 h and rice explants were pre-incubated for 72 h. Period of ex- posure for the lanes 4 to 7 was twice more than the period of exposure for the lanes 1 to 3.

131
Generation of T-strands
To detect T-strands in A. tumefaciens strain A348 (pVK224), bacteria were induced with AS, to- bacco leaf segments or different parts of rice plants. DNA samples from control and induced bacteria were subjected to non-denaturing South- ern analysis. Radiolabelled Hind III fragment 1 of pTiA6 was used as probe. Since this fragment includes regions of homology to all three parts of T-DNA (TL, TC and TR) it can detect all the six T-strands of pTiA6 [31 ]. Signals corresponding to four T-strands (TL, TR, TL + TC, TR + TC) were detected in DNA from bacteria induced with AS and tobacco leaf segments (Fig. 2). T-strands were not detected in bacteria induced with rice roots, leaf bases or leaf segments. Two of the T-strands, TL and TL + TC which are generally produced in larger quantities, were detected in DNA from bacteria induced with rice scutella. Therefore, the scutellum is the only part of rice that induces generation of T-strands in Agrobac- terium. However, the amount of T-strands gen- erated upon induction by rice scutellum was very low compared to the level of T-strands generated by tobacco leaf segments.
Discussion
We have analysed rice plants for their ability to induce or inhibit vir genes of Agrobacterium. Rice leaf segments were compared with tobacco leaf segments for their ability to induce vir genes. The level Of vir induction by rice leaf segments was very low compared to the level of induction by tobacco leaf segments (Table 1). It is known that corn produces inhibitors of vir induction and Agrobacterium growth [21] and the presence of inhibitors of vir induction can be a problem in Agrobacterium-mediated transformation of mono- cotyledonous plants. Our results show that rice leaf segments did not inhibit AS-mediated vir in- duction. Therefore, rice leaf segments do not pro- duce vir inhibitors. We also observed that rice leaf segments did not inhibit the growth of Agrobac- terium (data not shown).
Since we could not detect vir induction by rice leaves, we analysed other parts office for their vir inducibility. Grimsley et al. [9] have shown that meristematic tissues of maize, at or close to the apex are more susceptible to agroinfection com- pared to other non-meristematic tissues. Graves and Goldman [7] have reported that inoculation of maize seedlings in the scutellar region with wild-type Agrobacterium lead to the production of opines. Variations found in different tissues in their susceptibility to Agrobacterium-mediated transformation may be because of differences in their ability to induce vir genes. Our analysis of different parts of rice plants of different ages re- vealed that scutellum from four-day old seedlings is the only part that induced vir genes of Agro- bacterium quite efficiently (Table 3).
Freshly excised scutellum did not induce virE whereas scutellum pre-incubated for 72 h induced virE very well. It is known that pre-incubation of explants before cocultivation with Agrobacterium improves transformation efficiency in several plants [ 3]. We have observed that vir inducibility as well as transformation efficiency of tobacco were improved upon pre-incubation (Sunilkumar et aL, unpublished results). Since vir inducibility of scutellum increases upon pre-incubation, pre- incubation is likely to help in the transformation of rice scutellum. The inducibility of vir genes was found to be associated with the pre-incubated scutella and not with the scutellum-conditioned medium (Table 4). From these results, we suggest that either a physical association between scutel- lum and Agrobacterium is required for vir induc- tion or highly labile compounds that are continu- ously secreted by scutellum induce the vir genes.
Recently, Hiei et al. [ 11 ] reported that the scutellum-defived callus of rice was transformed efficiently by Agrobacterium. They found that the scutellum-derived calli maintained on medium containing 2,4-D for 25 days were transformed very efficiently, but the scutella excised from seed- lings after 5 days of germination on medium con- taining 2,4-D were transformed at a very low ef- ficiency. We compared vir inducibility of these explants cultured in conditions similar to those described by Hiei et al. [ 11 ] for rice transforma-

132
tion. We found that the scutellum-derived calli induced virE to an appreciable level but the level of induction of virE by the scutella excised from the seedlings germinated on medium containing 2,4-D was much lower. There is a good correla- tion between the vir inducibility of these two tis- sues (our results) and susceptibility of these tis- sues to Agrobacterium-mediated transformation (results of Hiei et al.). The scutellum-derived calli which induced virE more efficiently in our study were also found to be transformed more efficiently by Agrobaeterium [ 11]. Though the scutellum- derived calli induced virE, the level of induction was slightly lower than the level of induction achieved by using scutella from seeds germinated on hormone-free medium. Earlier, Li et al. [ 15] found that presence of 2,4-D in rice germination medium decreased the efficiency of Agrobacte- rium-mediated transformation. Germination of seeds on hormone-containing medium decreases the ability of the explants to induce vir genes and also decreases the transformation efficiency. Our vir induction results and the transformation re- sults of Hiei etal. show that culturing of scutellum-derived calli for 25 days on a medium containing 2,4-D restored vir inducibility and helped in improving transformation efficiency of the scutellar region.
This is the first report showing a high magni- tude of variation among different parts of a plant in the ability to induce vir genes. Xu et al. [32] showed that extracts of leaves from panicle- differentiating rice plants induced vir genes better than extracts from many other parts of rice plants. But the level of vir induction by the leaves from panicle-differentiating rice plants in that report was quite low compared to the level of induction by scutellum that we are reporting here. Usami et al. [29] reported that in three days-old wheat seedlings, only the extract of the transition region between shoot and root contained vir inducer(s), but extracts of shoot or root did not contain vir inducers. It is likely that scutella of other mono- cotyledonous plants also produce vir inducers. The vir inducers detected by Usami et al. [29] in the transition region of wheat seedlings may be because of the presence of scutellum in the tran-
sition region. In these two reports, plant extracts were used to analyse vir induction. We used plant parts for induction under conditions which are physiologically similar to those used in Agrobac- terium-mediated transformation. The ability of rice scutellum and the calli derived from scutel- lum to induce Agrobacterium vir genes indicates that they may be the right tissues for use in Agrobacterium-mediated transformation of rice.
Acknowledgements
The authors thank the Department of Biotech- nology, Govt. of India, for financial support. K. V. received a Senior Research Fellowship from the Council of Scientific and Industrial Research, Govt. of India, and K. P. received a Senior Re- search Fellowship from the University Grants Commission, Govt. of India. We thank Dr Stan- ton Gelvin, Purdue University, USA, for provid- ing the bacterial strains and the Bioinformatics Centre of Madurai Kamaraj University for the help we received.
References
1. Bonner WM, Laskey RA: A film detection method for tritium labelled proteins and nucleic acids in polyacryla- mide gels. Eur J Biochem 46:83-88 (1974).
2. Chan MT, Chang HH, Ho SL, Tong WF, Yu SM: Agro- bacterium-mediated production of transgenic rice plants expressing a chimeric a-amylase promoter/fl-glucuron- idase gene. Plant Mol Biol 22:491-506 (1993).
3. De Bondt A, Eggermont K, Druart P, De Vii M, Go- deris I, Vanderleyden J, Broekaert WF: Agrobacterium- mediated transformation of apple (Malus x domestica Borkh.): an assessment of factors affecting gene transfer efficiency during early transformation steps. Plant Cell Rep 13:587-593 (1994).
4. Douglas CJ, Halperin W, Nester EW: Agrobacterium tumefaciens mutants affected in attachment to plant cells. J Bact 152:1265-1275 (1982).
5. Engstrom P, Zambryski P, Van Montagu M, Stachel SE: Characterization of Agrobacterium tumefaciens virulence proteins induced by the plant factor acetosyringone. J Mol Biol 197:635-645 (1987).
6. Gould J, Devey M, Hasegawa O., Ulian EC, Peterson G, Smith RH: Transformation of Zea mays L. using Agro- bacterium tumefaciens and the shoot apex. Plant Physiol 95:426-434 (1991).

7. Graves ACF, Goldman SL: The transformation of Zea mays seedlings with Agrobacterium tumefaciens. Plant Mol Biol 7 :43-50 (1986).
8. Graves ACF, Goldman SL: Agrobacteriurn tumefaciens- mediated transformation of the monocot genus Gladiolus: detection of expression of T-DNA encoded genes. J B act 169:1745-1746 (1987).
9. Grimsley NH, Ramos C, Hein T, Hohn B: Meristematic tissues of maize plants are most susceptible to agroinfec- tion with maize streak virus. Bio/technology 6:185-189 (1988).
10. Hernalsteens JP, Thia-Toong L, Schell J, Van Mon- tagu M: An Agrobacterium-transformed cell culture from the monocot Asparagus officinalis. EMBO J 3 3039-3041 (1984).
11. Hiei Y, Ohta S, Komari T, Kumashiro T: Efficient trans- formation of rice (Oryza sativa L.) mediated by Agrobac- terium and sequence analysis of the boundaries of the T-DNA. Plant J 6:271-282 (1994).
12. lyer VN, Klee H J, Nester EW: Units of genetic expres- sion in the virulence region of a plant tumour-inducing plasmid of Agrobacterium tumefaciens. Mol Gen Genet 188:418-424 (1982).
13. Koukolikova-Nicola Z, Shillito RD, Hohn B, Wang K, Van Montagu M, Zambryski P: Involvement of circular intermediates in the transfer of T-DNA from Agrobacte- rium tumefaciens to plant cells. Nature 313:191-196 (1985).
14. Laemmli UK: Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685 (1970).
15. Li X-Q, Liu C-N, Ritchie SW, Peng J, Gelvin SB, Hodges TK: Factors influencing Agrobacterium-mediated transient expression of gusA in rice. Plant Mol Biol 20: 1037-1048 (1992).
16. Lichtenstein C, Draper J: Genetic engineering of plants. In: Glover DM (ed) DNA Cloning: A Practical Ap- proach, vol II, pp. 67-119. IRL Press, Oxford (1985).
17. Messens E, Dekeyser R, Stachel SE: A non-transform- able Triticum monococcum monocotyledonous culture produces the potent Agrobacterium vir-inducing com- pound ethyl ferulate. Proc Natl Acad Sci USA 87: 4368- 4372 (1990).
18. Miller JH: Experiments in Molecular Genetics. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1972).
19. MurashigeT, Skoog F: A revised method for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473-497 (1962).
133
20. Parke D, Ornston LN, Nester EW: Chemotaxis to plant phenolic inducers of virulence genes is constitutively ex- pressed in the absence of the Ti plasmid in Agrobacterium tumefaciens. J Bact 169:5336-5338 (1987).
21. Sahi SV, Chilton MD, Chilton WS: Corn metabolites af- fect growth and virulence of Agrobacterium tumefaciens. Proc Natl Acad Sci USA 87:3879-3883 (1990).
22. Schafer W, Gorz A, Kahl G: T-DNA integration and ex- pression in a monocot crop plant after induction of Agro- bacterium. Nature 327:529-532 (1987).
23. Simpson RA, O'Hara PJ, Kwok W, Montoya AL, Lich- tenstein C, Gordon MP, Nester EW: DNA from the A6S/2 crown gall tumor contains scrambled Ti-plasmid sequences near its junctions with plant DNA. Cell 29: 1005-1014 (1982).
24. Spencer PA, Towers GHN: Restricted occurrence of ac- etophenone signal compounds. Phytochemistry 30: 2933- 2937 (1991).
25. Stachel SE, Messens E, Van Montagu M, Zambryski P: Identification of signal molecules produced by wounded plant cells that activate T-DNA transfer in Agrobacterium tumefaciens. Nature 318:624-629 (1985).
26. Stachel SE, An G, Flores C, Nester EW: A Tn3 lacZ transposon for the random generation of fl-galactosidase gene fusions: application to the analysis of gene expres- sion in Agrobacterium. EMBO J 4:891-898 (1985).
27. Stachel SE, Nester EW: The genetic and transcriptional organization of the vir region of the A6 Ti plasmid of Agrobacterium tumefaciens. EMBO J 5:1445-1454 (1986).
28. Stachel SE, Timmerman B, Zambryski P: Generation of single-stranded T-DNA molecules during the initial stages of T-DNA transfer from Agrobacterium tumefaciens to plant cells. Nature 322:706-712 (1986).
29. Usami S, Okamoto S, Takebe I, Machida Y: Factor in- ducing Agrobacterium tumefaciens vir gene expression is present in monocotyledonous plants. Proc Natl Acad Sci USA 85:3748-3752 (1988).
30. Veluthambi K, Jayaswal RK, Gelvin SB: Virulence ge- nes A, G and D mediate the double-stranded border cleavage of TDNA from the Agrobacterium Ti plasmid. Proc Natl Acad Sci USA 84:1881-1885 (1987).
31. VeluthambiK, Ream W, Gelvin SB: Virulence genes, borders and overdrive generate single-stranded T-DNA molecules from the A6 Ti plasmid ofAgrobacteriurn tume- faciens. J Bact 170:1523-1532 (1988).
32. Xu Y, Bu W, Li B: Metabolic factors capable of inducing Agrobacterium vir gene expression are present in rice (Oryza sativa L.).Plant Cell Rep 12:160-164 (1993).
Copyright © 2022 FDOKUMEN




















