
Vocalizations of the sei whale Balaenoptera borealis off the Hawaiian Islands

Mass spectrometric investigation of the neuropeptide complement
and release in the pericardial organs of the crab, Cancer borealis
Lingjun Li,*,�,�Wayne P. Kelley,�,1 Cyrus P. Billimoria,� Andrew E. Christie,§,2 Stefan R. Pulver,�Jonathan V. Sweedler� and Eve Marder�
*School of Pharmacy & Department of Chemistry, University of Wisconsin, Madison, Wisconsin, USA
�Department of Chemistry and the Beckman Institute, University of Illinois, Urbana, Illinois, USA
�Department of Biology, Volen Center, Brandeis University, Waltham, Massachusetts, USA
§Department of Neuroscience, University of Pennsylvania School of Medicine, Philadelphia, Pennsylvania, USA
Abstract
The crustacean stomatogastric ganglion (STG) is modulated
by both locally released neuroactive compounds and circula-
ting hormones. This study presents mass spectrometric
characterization of the complement of peptide hormones
present in one of the major neurosecretory structures, the
pericardial organs (POs), and the detection of neurohormones
released from the POs. Direct peptide profiling of Cancer
borealis PO tissues using matrix-assisted laser desorption/
ionization (MALDI) time-of-flight (TOF) mass spectrometry
(MS) revealed many previously identified peptides, including
proctolin, red pigment concentrating hormone (RPCH), crus-
tacean cardioactive peptide (CCAP), several orcokinins, and
SDRNFLRFamide. This technique also detected corazonin, a
well-known insect hormone, in the POs for the first time.
However, most mass spectral peaks did not correspond to
previously known peptides. To characterize and identify these
novel peptides, we performed MALDI postsource decay (PSD)
and electrospray ionization (ESI) MS/MS de novo sequencing
of peptides fractionated from PO extracts. We characterized a
truncated form of previously identified TNRNFLRFamide,
NRNFLRFamide. In addition, we sequenced five other novel
peptides sharing a common C-terminus of RYamide from the
PO tissue extracts. High K+ depolarization of isolated POs
released many peptides present in this tissue, including sev-
eral of the novel peptides sequenced in the current study.
Keywords: Cancer borealis, MALDI MS, neuropeptides,
neurosecretion, pericardial organs, postsource decay peptide
sequencing.
J. Neurochem. (2003) 87, 642–656.
The crustacean stomatogastric ganglion (STG) is one of the
leading systems for studying the neural basis of motor pattern
generation and the modulatory effects of neuroactive
substances at the cellular and network levels (Marder and
Hooper 1985; Harris-Warrick et al. 1992; Marder et al.
1995; Marder and Calabrese 1996; Skiebe 2001; Nusbaum
and Beenhakker 2002). Previous studies have shown that the
STG is modulated by both neuroactive agents released
locally from input axons and circulating hormones delivered
via the hemolymph (Marder 1987; Christie et al. 1995). A
large number of studies have documented that a diverse
assortment of small molecule transmitters, amines and
Received April 30, 2003; revised manuscript received July 16, 2003;
accepted July 16, 2003.
Address correspondence and reprint requests to Dr Lingjun Li, School
of Pharmacy, University of Wisconsin, 777 Highland Avenue, Madison,
WI 53705–2222, USA. E-mail: [email protected] address: Pharmaceutical Development, GlaxoSmithKline, King
of Prussia, PA 19406, USA.2Current address: Department of Biology, University of Washington,
Seattle, WA 98195–1800, USA.
Abbreviations used: ACN, acetonitrile; AST, allatostatin; CabTRP,
Cancer borealis tachykinin-related peptide; CCAP, crustacean cardio-
active peptide; CHH, crustacean hyperglycemic hormone; CID, colli-
sional induced dissociation; DHB, 2,5-dihydroxybenzoic acid; DG
neuron, dorsal gastric neuron; dgn, dorsal gastric nerve; ESI, electro-
spray ionization; IC neuron, inferior cardiac neuron; lvn, lateral ven-
tricular nerve; LP neuron, lateral pyloric neuron; MALDI, matrix-
assisted laser desorption/ionization; MS, mass spectrometry; mvn, medial
ventricular nerve; PD neuron, pyloric dilator neuron; pdn, pyloric dilator
nerve; PO, pericardial organ; PSD, postsource decay; PY neuron, pyloric
neuron; pyn, pyloric nerve; RPCH, red pigment concentrating hormone;
RP-HPLC, reverse phase high performance liquid chromatography;
STG, stomatogastric ganglion; STNS, stomatogastric nervous system;
TFA, trifluoroacetic acid; TOF, time-of-flight; VD neuron, ventricular
dilator neuron.
Journal of Neurochemistry, 2003, 87, 642–656 doi:10.1046/j.1471-4159.2003.02031.x
642 � 2003 International Society for Neurochemistry, J. Neurochem. (2003) 87, 642–656

neuropeptides are present in the STG neuropil. Several recent
reports have shown that many of the same neuromodulatory
compounds are also present in the neurohemal organs of
crustaceans (Christie et al. 1995; Skiebe 1999; Skiebe et al.
1999; Pulver and Marder 2002). One such neurohemal
structure, the pericardial organ (PO), has long been known to
be an important source of circulating hormones. Moreover,
several studies have shown that many of the hormones
present in the POs can modulate the neural circuits in the
STG, which is situated in the ophthalmic artery anterior to
the heart (Alexandrowicz and Carlisle 1953; Stangier et al.
1986; Stangier et al. 1988; Keller 1992; Christie and
Nusbaum 1995). The current study focuses on mass spectr-
ometric characterization of neuropeptides present in and
released from the POs of the crab Cancer borealis.
Most of our knowledge concerning the peptide content of
the stomatogastric nervous system (STNS) and the POs is
based on immunocytochemistry. This technique, while a
powerful first step in determining modulator presence, leaves
the amino acid sequence identity of the native peptides
unknown. Clearly, unambiguous determination of the actual
peptide structure with accurate molecular weight and amino
acid sequence information is essential for studies of the
physiological functions of these molecules. Matrix-assisted
laser desorption/ionization mass spectrometry (MALDI MS)
is a highly accurate method that can be used to directly
profile peptides from complex biological samples such as
tissue sections or single cells with minimal sample prepar-
ation (Jimenez et al. 1994; Li et al. 1999b; Li et al. 2000a;
Li et al. 2000b; Predel 2001). Its application to peptide
identification has recently been extended to the crustacean
STNS in the identification and characterization of multiple
members of the orcokinin peptide family (Li et al. 2002;
Skiebe et al. 2002). In the current study, we aim to determine
the complement of peptides present in the POs of the crab, C.
borealis. To overcome the challenges of the high chemical
complexity of these neuropeptides, and the almost complete
lack of genomic information, we employed front-end multi-
stage microseparation to simplify cellular matrices, and
performed de novo mass spectrometric sequencing to obtain
the primary structures of several previously unknown
peptides. To explore the putative hormonal role of these
peptides, we also performed the first mass spectrometric
assay of peptide release from the POs using high K+-induced
depolarizations.
Materials and methods
Animals and dissection
Jonah’s crabs, C. borealis were obtained from Commercial Lobster
(Boston, MA, USA) and maintained without food in artificial
seawater tanks at 10–12�C. Animals were cold-anesthetized by
packing in ice for 15–30 min prior to dissection. The POs were
dissected by removing the carapace covering the heart and
subsequently removing the longitudinal body muscles, hyperdermis,
and heart. The animal was then pinned ventral side up in a Sylgard-
lined dissection dish to expose the pericardial cavity. The POs were
identified visually as an iridescent web of nerves branching over the
muscles surrounding the pericardial cavity and dissected free. All
dissection was carried out in chilled (approximately 4�C) physio-logical saline (composition in mM: NaCl, 440; KCl, 11; MgCl2, 26;
CaCl2, 13; Trizma base, 11; maleic acid, 5; pH 7.45).
Cellular sample preparation for MALDI MS analysis
Small pieces of tissues were dissected and prepared for MALDI MS
analysis. Physiological saline was replaced with an aqueous MALDI
matrix solution, 10 mg/mL of 2,5-dihydroxybenzoic acid (DHB)
(ICN Pharmaceuticals, Costa Mesa, CA, USA), to remove the
extracellular salts associated with the tissue sample (Garden et al.
1996). Tungsten needles and fine forceps were used to dissect and
transfer small pieces of tissue onto a MALDI sample plate
containing 0.5 lL of either regular aqueous DHB (10 mg/mL)
matrix solution or concentrated DHB (50 mg/mL) in acetone/water
(4 : 1) mixed solvent. Once on the sample plate, the tissue was
smashed with dissection tools and allowed to dry at ambient
temperature and then subjected to MALDI MS analysis.
Microbore reverse phase (RP)-HPLC of homogenates
The extraction and first stage separation procedure were performed
as reported previously (Li et al. 2002). Peptides were initially
extracted from 26 POs using acidified acetone (1 : 40 : 6, concen-
trated HCl/acetone/H2O) as described previously (Floyd et al.
1999). Briefly, samples were homogenized in a microhomogenizer
(Jencons Scientific Ltd, UK), and the supernatant drawn off and
centrifuged (Baxter Biofuge 15, Mcgraw Park, IL, USA). This
process was repeated several times, water was added and then the
extract was concentrated under a stream of nitrogen to approxi-
mately 300 lL.Separations were performed utilizing a microbore HPLC (Magic
2002, Michrom BioResources, Auburn, CA, USA). For the first stage
separation, an aliquot of the extract was injected onto a reverse phase
1.0 · 150 mm C-18 column (Reliasil) with a 5-lm particle size and
30 nm pore size. The column was equilibrated with solvent A at a
programmed temperature of 35�C. An aliquot of the aqueous extractwas injected onto the column at a constant flow rate of 50 lL/min anda gradient developed from 5 to 98% of solvent B in 34 min. Solvent A
consisted of 2% acetonitrile (ACN), 98% H2O and 0.1% trifluoro-
acetic acid (TFA) (v/v). Solvent B consisted of 95% ACN, 5% H2O
and 0.1% TFA (v/v). Sample peaks were detected via absorbance at
214 and 280 nm wavelengths and the eluent collected by a small
volume fraction collection system (Gilson FC 203B, Middleton, WI,
USA). To identify peptides of interest, each fraction was screened
using MALDI MS; approximately 0.25 lL of each fraction (approxi-mately 20–200 lL total) was deposited on aMALDIMS sample platefollowed by the same volume of an a-cyano-4-hydroxycinnamic acidmatrix (10 mg/mL in 6 : 3 : 1 ACN/H2O/3% TFA) (Aldrich,
Milwaukee, WI, USA). Thus more than 95% of each fraction was
available for further assays.
Second stage microbore HPLC separation
Fractions assayed by MALDI time-of-flight (TOF) MS that
contained insufficient intensity or interfering peaks from coelution
Neuropeptides in C. borealis pericardial organs 643
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 87, 642–656

were further purified by a second stage HPLC. Second stage
separations were effected by changing the column to a
1.0 · 150 mm Vydac C-18 MS column (218MS5115, Grace Vydac,
Hesperia, CA, USA) consisting of 5 lm particles with a 30-nm pore
size, changing the solvent additives, slowing the relative flow rate
and by flattening the gradient profile. Separations of aliquots from
the combined first stage fractions were performed both with on-line
electrospray ionization (ESI)-MS detection and off-line fraction
collection. Both procedures utilized identical solvents and methods.
Fractions were collected manually on a small volume fraction
collection system.
The selected fractions from the first-stage analysis were com-
bined and concentrated to an aqueous medium using a commercial
Speedvac system (Thermo-Savant, Holbrook, NY, USA). An aliquot
of the sample was then loaded onto a peptide trap inline with the
injection port of the microbore HPLC instrument. For HPLC-ESI-
MS experiment a 5-lL injection of the concentrate was used and theremainder, approximately 50 lL, was saved for the second stageseparation and fraction collection. Each aliquot was injected onto
the column at a uniform flow rate of 50 lL/min. The mobile phaseconsisted of solvent A: 95% H2O (Burdick and Jackson, Muskegon,
MI, USA), 5% ACN, 0.1% acetic acid (v/v) and 0.02% TFA (v/v)
(Sigma-Aldrich, Milwaukee, WI, USA) and B: 10% H2O, 90%
2 : 2 : 1 ACN, 0.1% acetic acid (v/v) and 0.014% TFA (v/v). A
gradient was developed from 5 to 15% solvent B in 5 min and 15–
45% B in an additional 30 min.
MALDI MS
MALDI mass spectra were obtained using a Voyager DE STR
(Applied Biosystems, Framingham, MA, USA) TOF mass spectro-
meter equipped with delayed ion extraction. A pulsed nitrogen laser
(337 nm) was used as desorption/ionization source, and positive-ion
mass spectra were acquired using both linear and reflectron mode.
Each representative mass spectrum shown is the smoothed average
of 128–256 laser pulses. Mass calibration was performed externally
using a mixture of synthetic peptide standards (PE Biosystems,
Framingham, MA, USA). Mass accuracy was typically better than
0.01% (Li et al. 1998).
PSD analysis
Semi-purified microbore LC fractions containing a peptide of
interest, or 2 pmol of synthetic peptide standards (synthesized by
either the Biotechnology Center at the University of Illinois or the
Protein Chemistry Laboratory at the University of Pennsylvania,
School of Medicine), were subjected to PSD analysis. For MALDI
PSD analysis, the matrix a-cyano-4-hydroxycinnamic acid (10 mg/mL in 6 : 3 : 1 ACN/water/3% TFA) was used. In these experi-
ments, the total acceleration voltage was 20 kV, grid voltage set at
75%, guide wire voltage set at 0.03%, and a delay time of 75 ns
used. By the use of timed ion selector, different precursor ions were
selected from a mixture of peptides and subjected to fragmentation.
Under these experimental conditions, the mass accuracy of the
precursor ion was within 30 p.p.m., and the average error on the
mass assignment of the PSD ions was less than 0.3 Da. Spectra were
obtained by accumulating data from 100 to 256 laser shots. To
obtain complete PSD spectra, a series of reflectron TOF spectral
segments were acquired, each optimized to focus fragment ions
within different m/z ranges (Kaufmann et al. 1993). Each segment
was stitched together using the Biospectrometry Workstation
software to generate a composite PSD spectrum.
On-line ESI MS/MS analysis
For on-line MS detection, the column was connected directly to the
atmospheric pressure inlet port of an LCQ Deca, ESI-ion trap mass
spectrometer (Thermo-Finnigan, San Jose, CA, USA). The transfer
volume to the MS detector was calculated to be similar to that of the
absorbance detector resulting in retention times that are similar to
the fraction collection assays. The MS tune method employed a
spray voltage of 4.3 kV, a capillary temperature of 220�C, capillaryvoltage of 21 Vand a tube lens offset of 10 V. By default, automatic
gain was used to control injection of ions into the trap. The Xcalibur
instrument setup software (Thermo-Finnigan, San Jose, CA, USA)
employed a modified triple play data dependent acquisition control.
Briefly, a full scan MS was followed by a zoomscan and then MS2
of the largest peak in the full scan. Dynamic exclusion was utilized
in order to include high resolution and MS2 information on
secondary peaks within the scan.
Nanospray ESI MS/MS
To perform collisional-induced dissociation (CID) experiments for
sequence information while utilizing minimal sample, the ion trap
instrument was fitted with a static nanospray source (Thermo-
Finnigan, San Jose, CA, USA). An aliquot of approximately 5 lLwas taken from a fraction from the second stage separation
corresponding to the mass of interest, concentrated via Speedvac
and reconstituted in 10 lL of 50/50 (v/v) CH3OH and H2O with
0.1% formic acid. The sample was loaded into a tapered, platinum
coated, borosilicate nanospray emitter, PicoTip� (New Objective,
Woburn, MA, USA) and a spray voltage of 1.5 kV was applied. The
instrument capillary temperature was set at 200�C with an applied
potential of 39 V. The optimized spray voltage and ion optic settings
were adjusted using the instrument’s auto tune function. Once a full
scan was observed in real time, MS2 and MS3 experiments were
directed by manual input of the m/z-values.
Tandem MS sequencing experiments used a mass isolation width
of 1.5 Da for the precursor ion and 1.0 Da for resulting MS2
fragment ions to be retained in the trap. An activation Q (ion
instability parameter) value of 0.25 at an activation time of 30 ms
was utilized. The normalized collisional (RF) energy was set at 35%
for both MS2 and MS3 experiments. Spectra were acquired for
approximately 2 min resulting in 164 scans. Sequence verification
was facilitated using the online Protein Prospector, MS-Product
program from the University of California, San Francisco.
MALDI assay for peptides released from the POs
To depolarize the POs, we employed a 10-fold higher concentration
of K+ (110 mM) than that in normal saline to the isolated pair of POs
from each animal (N ¼ 3). A cocktail of peptidase inhibitors
including amastatin (20 lM), leupeptin (10 lM), antipain-dihydro-chloride (71 lM), bestatin (130 lM), phosphoramidon (37 lM), andaprotinin (0.3 lM) (Roche Molecular Biochemicals, Germany) wasused throughout the experiment. All of the preparations were kept
on ice (�4�C) for the duration of the experiment. To determine if thedetected peptides were released in a depolarization and Ca2+-
dependent manner, we repeated the experiment without high
K+ depolarization in normal saline as well as in high K+/low Ca2+
644 L. Li et al.
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 87, 642–656

(1.3 mM Ca2+ and 13 mM Mn2+) saline. Each preparation was
incubated for 10 min in the experimental condition with two washes
in normal saline between each condition to remove peptides
resulting from previous experimental protocol.
We sampled 10 lL of releasate from each experimental condi-
tion, desalted and concentrated the releasate with ZipTip pipette tips
(Millipore, Bellerica, MA, USA) packed with C18 reverse phase
media prior to spotting onto the MALDI sample plate. Briefly, the
ZipTip was wet with 50% ACN in Milli-Q water, equilibrated with
0.1% TFA in Milli-Q water, loaded with sample containing 0.1%
TFA by aspirating and dispensing the sample 10 times, and then
washed with 5% methanol in 0.1% TFA/water, followed by elution
directly onto a MALDI MS sample plate with 2 lL DHB (50 mg/
mL) in 50% ACN with 0.1% TFA.
Electrophysiology
The stomach was removed as previously described. The STNS was
isolated from the stomach and pinned onto a Sylgard (Dow Corning,
Midland, MI, USA) coated dish. The STG and the stomatogastric
nerve (stn) were desheathed. The STG was isolated from anterior
ganglia inputs by placing isotonic sucrose (750 mM) containing
10)6 M tetrodotoxin in a Vaseline well built around the stn.
Extracellular recordings were made from pyloric and gastric nerves
using stainless steel pin electrodes within saline-filled Vaseline
wells. Signals were amplified by an AM-Systems 1700 differential
amplifier (Carlsborg, WA, USA) and recorded using an Axon
Instruments (Foster City, CA, USA) data interface board. Spike time
data were extracted using scripts written by Dirk Bucher in Spike2
(Cambridge Electronic Design, Cambridge, UK). Two-minute
epochs of data were used in the analysis of peptide actions. The
two minutes preceding the application of peptide and the eighth to
tenth minute of application were analyzed.
Results
Direct MALDI MS profiling of peptides
in the pericardial organs reveals a multitude
of peptides present
As shown in Fig. 1, direct MALDI MS profiling of a small
piece of freshly isolated PO from C. borealis revealed the
presence of a number of previously identified peptides
including proctolin, crustacean cardioactive peptide (CCAP),
SDRNFLRFa, and Ala13-orcokinin, as well as many other
unknown peptides. Figure 1(b) shows the mass spectrum
acquired in the high mass region. A number of peaks were
detected in the mass range spanning from 2000 to 9650 Da. A
multitude ofCHHs have been previously characterized in sinus
gland (SG) neurosecretory system in the eyestalk and POs of
various species of crustacea (Keller 1992; Dircksen et al.
2001), with molecular weights ranging from 8420 to 8634 Da.
The peak atm/z 8561.70, however, does not correspond to any
of the previously identified CHHs. The detection of this peak
and a few other peaks around 8000 and 9000 Da in the mass
spectrum suggested the possible presence of CHH peptides in
the crab, C. borealis. Another interesting peak that might be
related to the CHH precursor was the mass spectral peak atm/z
4071.68, whose molecular weight was in close agreement to
that of CHH-precursor related peptide (CPRP, with 0.002%
mass measurement error) of the shore crab, Carcinus maenas
(Dircksen et al. 2001). However, without additional informa-
tion from sequencing analysis, conclusive assignment of these
mass spectral peaks was not possible.
Because peptides are distributed differentially throughout
the POs, direct tissue profiling only provides a snapshot of the
peptides present in a specific region of this neurohemal organ.
Due to the heterogeneous distribution of modulators in this
tissue, a given peptide was not always detected in the MALDI
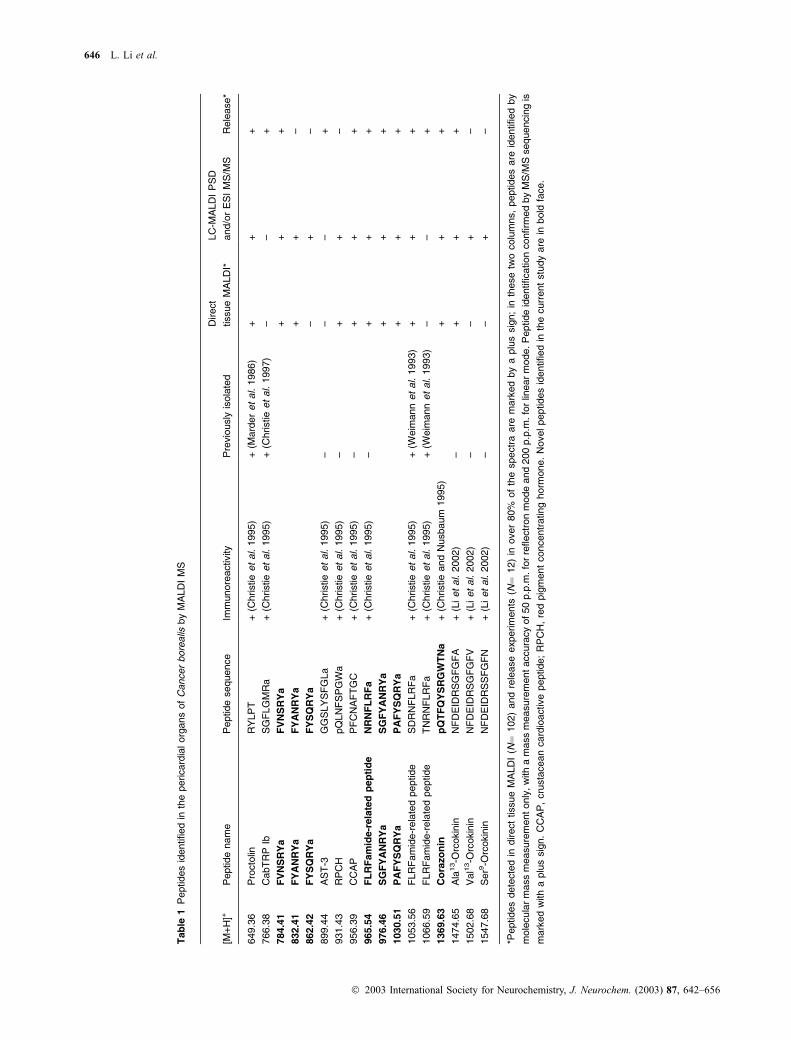
mass spectra of individual PO tissue samples. In Table 1, we
consider a peptide to be present (+) in the POs only if we
observed the peptide signals in over 80% of the total spectra
generated (N ¼ 102). This cut-off value is determined based
on the occurrence of known peptides present in the POs and
the reproducibility of our MS detection.
Off-line coupling microbore LC fractionation
of PO tissue extract with MALDI MS detection
for peptide sequencing
To obtain a comprehensive description of peptides present in
the PO tissue and simplify the complex mixture resulting
from the tissue extract, we performed multiple stages of
microbore LC separation prior to MALDI MS analysis.
(a)
(b)
Fig. 1 Representative MALDI mass spectrum of a small piece of
freshly isolated pericardial organ from C. borealis. (a). The mass
region of 625–1525 is displayed (b). The mass range from 2000 to
9650 is displayed. Signals correspond to protonated molecular ions,
[M + H]+, where M is the molecular weight of each peptide. Several
previously identified peptides are labeled with asterisks, and repre-
sented in bold face.
Neuropeptides in C. borealis pericardial organs 645
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 87, 642–656
Tab
le1
Peptides
identified
inth
epericard
ialorg
ans
of
Cancer
bore
alis
by
MA
LD
IM
S
[M+
H]+
Peptide
nam
eP
eptide
sequence
Imm
unore
activity
Pre
vio
usly
isola
ted
Direct
tissue
MA
LD
I*
LC
-MA
LD
IP
SD
and/o
rE
SI
MS
/MS
Rele
ase*
649.3
6P
rocto
linR
YLP
T+
(Christie
et
al.
1995)
+(M
ard
er
et
al.
1986)
++
+
766.3
8C
abT
RP
IbS
GF
LG
MR
a+
(Christie
et
al.
1995)
+(C
hristie
et
al.
1997)
––
+
784.4
1F
VN
SR
Ya
FV
NS
RY
a+
++
832.4
1F
YA
NR
Ya
FY
AN
RY
a+
+–
862.4
2F
YS
QR
Ya
FY
SQ
RY
a–
+–
899.4
4A
ST
-3G
GS
LY
SF
GLa
+(C
hristie
et
al.
1995)
––
–+
931.4
3R
PC
HpQ
LN
FS
PG
Wa
+(C
hristie
et
al.
1995)
–+
+–
956.3
9C
CA
PP
FC
NA
FT
GC
+(C
hristie
et
al.
1995)
–+
++
965.5
4F
LR
Fam
ide-r
ela
ted
pep
tid
eN
RN
FL
RF
a+
(Christie
et
al.
1995)
–+
++
976.4
6S
GF
YA
NR
Ya
SG
FY
AN
RY
a+
++
1030.5
1P
AF
YS
QR
Ya
PA
FY
SQ
RY
a+
++
1053.5
6F
LR
Fam
ide-r
ela
ted
peptide
SD
RN
FLR
Fa
+(C
hristie
et
al.
1995)
+(W
eim
ann
et
al.
1993)
++
+
1066.5
9F
LR
Fam
ide-r
ela
ted
peptide
TN
RN
FLR
Fa
+(C
hristie
et
al.
1995)
+(W
eim
ann
et
al.
1993)
––
+
1369.6
3C
ora
zo
nin
pQ
TF
QY
SR
GW
TN
a+
(Christie
and
Nusbaum
1995)
++
+
1474.6
5A
la13-O
rcokin
inN
FD
EID
RS
GF
GF
A+
(Liet
al.
2002)
–+
++
1502.6
8V
al1
3-O
rcokin
inN
FD
EID
RS
GF
GF
V+
(Liet
al.
2002)
––
+–
1547.6
8S
er9
-Orc
okin
inN
FD
EID
RS
SF
GF
N+
(Liet
al.
2002)
––
+–
*Peptides
dete
cte
din
direct
tissue
MA
LD
I(N
¼102)
and
rele
ase
experim
ents
(N¼
12)
inover
80%
of
the
spectr
aare
mark
ed
by
aplu
ssig
n;
inth
ese
two
colu
mns,
peptides
are
identified
by
mole
cula
rm
ass
measure
ment
only
,w
ith
am
ass
measure
ment
accura
cy
of50
p.p
.m.fo
rre
flectr
on
mode
and
200
p.p
.m.fo
rlin
ear
mode.P
eptide
identification
confirm
ed
by
MS
/MS
sequencin
gis
mark
ed
with
aplu
ssig
n.
CC
AP
,cru
sta
cean
card
ioactive
peptide;
RP
CH
,re
dpig
ment
concentr
ating
horm
one.
Novelpeptides
identified
inth
ecurr
ent
stu
dy
are
inbold
face.
646 L. Li et al.
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 87, 642–656

A chromatogram of the first stage LC separation is depicted
in Fig. 2 (center). Several representative MALDI survey
spectra of corresponding LC fractions are shown surrounding
the LC chromatogram. In many ways MALDI-TOF MS is an
orthogonal separation technique in that it provides a further
dynamic range and resolution to the separation of the crude
material. As shown in the displayed MALDI spectra of
various LC fractions in Fig. 2, each LC fraction still
contained numerous compounds, illustrating the high com-
plexity of the crude material. With the precursor ion selection
capability, MALDI postsource decay (PSD) analysis can be
performed on semipurified LC fractions to obtain peptide
sequence information. As an example, Fig. 3 shows several
MALDI PSD fragmentation spectra obtained from a single
LC fraction, with those selected precursor ions labeled with
dots. Detailed analyses of PSD fragmentation spectra of
precursor ions at m/z 965.543, 976.464, 1030.51, and
1369.60 are described below; analysis of the fragmentation
pattern of the precursor ion at m/z 1474.6 confirms the
identity of an orcokinin peptide reported previously (Li et al.
2002).
MALDI-PSD analysis of putative corazonin peptide
As shown in Fig. 1, among the numerous peaks detected in
the crab PO tissue, the mass spectral peak at m/z 1369.62
matches the calculated protonated molecular weight of
corazonin (Mr ¼ 1369.63), an insect hormone that is
conserved in all but one species examined (Veenstra 1989,
1991). To determine if the mass spectral peak at m/z 1369.62
has an identical amino acid sequence to that of authentic
corazonin, we performed PSD analysis.
Figure 4 shows the peptide sequence of corazonin (Fig.
4a) and PSD spectra obtained from the putative corazonin
containing LC fraction (Fig. 4b, upper trace) and the
synthetic corazonin standard (Fig. 4b, lower trace). The
identity of authentic corazonin in the PO extract is strongly
supported by the virtually identical fragmentation pattern
obtained between the putative and the synthetic corazonin
PSD spectra. The complete fragmentation observed from the
precursor ion at m/z 1369.63 in the PO extract also allowed
de novo sequencing of the peptide. Thus, we deduced the
amino acid sequence of the putative corazonin peak using the
mass difference between consecutive b- or y-type ions.
The sequence is identical to authentic corazonin-
qTFQYRGWTN-amide.
To confirm the sequence of corazonin, we entered the
derived sequence into the MS-Product software program
developed by the UCSF Mass Spectrometry Facility (http://
prospector.ucsf.edu). The software calculates the possible
fragment ions resulting from PSD processes. The fragment
ions detected in the PSD spectra match the predicted
fragment ions and several additional internal fragment ions
are labeled (Fig. 4b). The measured molecular weight of the
precursor ion and the mass of the C-terminal fragment ions
Fig. 2 An illustration of MALDI MS tracking of microbore HPLC
separation of crude tissue extract of pericardial organs from C. bore-
alis. A chromatogram from the first stage HPLC separation of the
extract is shown in the center. Detection was achieved by absorbance
at a wavelength of 214 nm. Several MALDI MS survey spectra of
corresponding LC fractions are displayed surrounding the LC chro-
matogram.
Fig. 3 MALDI PSD sequencing of multiple novel peptides from a
semipurified microbore LC fraction of C. borealis pericardial organ
extract. The spectrum in the center is a MALDI mass spectrum of a
single LC fraction, with gray dots on the selected peaks indicating the
precursor ions being isolated for MALDI PSD fragmentation. The
surrounding traces are PSD composite fragmentation spectra of var-
ious precursor ions selected.
Neuropeptides in C. borealis pericardial organs 647
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 87, 642–656

confirm the amidated C-terminus and a pGlu-modified
N-terminus in the peptide. This result confirms that authentic
corazonin is present in the POs of C. borealis. Authentic
corazonin has recently been shown to be a powerful
modulator of the neural circuits present in the STG of
C. borealis (A.E. Christie, unpublished observation).
A new member of the FMRFamide peptide family
Figure 5(a) shows the MALDI PSD fragmentation spectrum
of a precursor ion at m/z 965.54. Using the same sequencing
strategy outlined above, we determined the amino acid
sequence to be NRNFLRFamide. This represents a truncated
form of the TNRNFLRFamide peptide that was previously
isolated and sequenced from the STNS of the crab C. borealis
(Weimann et al. 1993). The derived amino acid sequence is
shown at the top of the spectrum. The sequence interpretation
was performed from both the N- (b-ions) and C-termini
(y-ions). The PSD analysis of the synthetic peptide standard
NRNFLRFamide produced an identical fragmentation pat-
tern (data not shown), further substantiating the proposed
sequence.
The similarity between this peptide and those previously
shown to be physiologically active on STG motor patterns
(Weimann et al. 1993) caused us to ask whether NRNFLRF-
amide would mimic the actions of the previously identified
extended FLRFamides. Therefore, we carried out a series of
electrophysiological experiments. Figure 5(b) shows that
bath application of 10)6 M synthetic NRNFLRFamide can
activate both pyloric and gastric rhythms in preparations in
which neither rhythm is being expressed (N ¼ 4). Shown on
the left is a schematic drawing illustrating the extracellular
recording sites and experimental conditions used to collect
the data shown on the right. The STG was isolated from
anterior ganglia inputs by placing sucrose and 10)6 M
tetrodotoxin in a Vaseline well on the stn, as indicated in
Fig. 5(b). Extracellular recording electrodes were placed on
the nerves containing the axons of motor neurons of the
STNS. The lateral ventricular nerve (lvn) carries the axons of
the lateral pyloric (LP) neuron, the pyloric (PY) neurons, and
pyloric dilator (PD) neurons, all active in the pyloric rhythm.
The pyloric nerve (pyn) and the pyloric dilator nerve (pdn)
contain axons of the PY and PD neurons, respectively. The
median ventricular nerve (mvn) carries the axons of the
ventricular dilator (VD) and inferior cardiac (IC) neurons.
The dorsal gastric nerve (dgn) carries the axon of the dorsal
gastric (DG) motor neuron, an important component of the
gastric mill network. Under control conditions (Fig. 5b, right
upper panel), the mvn and pdn were silent, and the other
nerve recordings showed some sporadic activity. Application
of 10)6 M NRNFLRFamide (Fig. 5b, right lower panel)
activated the DG neuron in four of four experiments. In all
four experiments the DG neuron was silent in control saline.
In three of the four experiments, NRNFLRFamide induced
DG bursting. In these three experiments NRNFLRFamide
increased the mean number of spikes per burst from 0 to
105 ± 16.7 (standard error of the mean, p < 0.05), increased
the mean burst duration from 0 to 7.91 ± 0.90 s (p < 0.02)
(a)
(b)
Fig. 4 MALDI PSD analysis of the putative corazonin peptide. (a) The
sequence of corazonin, with the N-terminal on the left, and the
C-terminal on the right. Single letter amino acid abbreviations are
used, with q (pGlu), T (Thr), F (Phe), Q (Gln), Y (Tyr), S (Ser), R (Arg),
G (Gly), W (Trp), and N (Asn). Observed b-type (bottom) and y-type
(top) ion pairs are indicated by arrows. (b) MALDI PSD fragmentation
ion spectra of both the ion at m/z 1369.63 from the LC fraction of the
PO extract from C. borealis (upper trace) and the synthetic corazonin
standard (lower trace). As shown in the figure, the immonium ions
observed in the low mass region of the spectrum (as highlighted in the
inset) indicates the presence of amino acids including Ser (60), Arg
(129, 70, 87, 112), Thr (74), pGlu (84), Asn (87), Gln (101), Phe (120),
Tyr (136), and Trp (159). Mass signal pairs (mb + my ¼ mprecursor + 1)
at m/z 112.1/1259.4, 213.2/1158.3, 360.4/1011.1, 488.5/883.0, 651.7/
719.8, 738.8/632.7, 895.0/476.5, 952.0/419.5, 1138.2/233.2, and
1239.3/132.1 are assigned as paired b-type and y-type ions. The
b-type ions are determined based on the detection of their corres-
ponding a-type ions (loss of CO, yielding a 28-Da lower mass ion). The
fragment ion labels used are based on the established nomenclature
(Roepstorff and Fohlman 1984). N-Terminal ion series such as a/b-
ions and their loss of neutrals, C-terminal ion series such as y-ions,
and several internal fragment ions as well as immonium ions are
labeled only in the upper trace spectrum. The masses of the fragment
ion signals labeled with asterisks in the lower trace are identical to
fragment ions detected in the upper trace.
648 L. Li et al.
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 87, 642–656

with a mean cycle period of 16.0 s. In the fourth experiment
NRNFLRFamide induced tonic firing of the DG neuron.
NRNFLRFamide applications significantly increased the
mean duty cycle (fraction of the time the neuron is active
over one entire period of activity) of the LP neuron from
0.12 ± 0.05–0.23 ± 0.07 (p < 0.05). These effects are simi-
lar to those previously reported for SDRNFLRFamide and
TNRNFLRFamide (Weimann et al. 1993).
De novo sequencing of RYamide peptides
Because of the lack of genomic sequence information, partial
sequence-specific fragmentation is not sufficient for peptide
identification. Thus, only high quality PSD fragmentation
spectra resulted in confident peptide identification. Several
PSD analyses generated complete sequence specific frag-
mentation that allowed de novo sequencing of previously
unknown peptides. Figure 6 shows the PSD fragmentation
spectrum of a precursor ion at m/z 976.46. The determined
amino acid sequence was SGFYANRY-amide. We performed
the PSD analysis of the synthetic peptide of the above
sequence which showed identical fragmentation, thus con-
firming the identity of the proposed amino acid sequence
(data not shown).
Another unknown peak at m/z 1030.51 in the LC fraction
was also subjected to PSD fragmentation analysis as shown
in Fig. 7(a). Using the same procedure outlined above, we
obtained a partial sequence of P/A-FYSQRY-amide, with
(a)
(b)
Fig. 5 Characterization of NRNFLRFamide. (a) MALDI PSD frag-
mentation spectrum of a precursor ion at m/z 965.54. The derived
amino acid sequence is shown at the top of the spectrum. The frag-
ment ions are labeled using the established nomenclature. The amino
acid is deduced from both directions using the complete N- and C-
terminal ion series. Detected b/y ion pairs include 115.1/851.5, 271.3/
695.4, 385.2/581.4, 532.3/434.3, 645.3/321.2, 801.4/165.1. Immonium
ions indicating the presence of Asn (87, 70), Arg (70, 87, 112, and
129), Phe (120), and Leu (86) are detected and labeled as single-letter
amino acid abbreviations in Fig. 5(a). Single letter amino acid abbre-
viations are used, with N (Asn), R (Arg), F (Phe), and L (Leu). (b)
Actions of NRNFLRFamide on the motor patterns of the stomato-
gastric ganglion (STG) of the crab, C. borealis. Shown on the left is a
schematic diagram of the experimental configuration. The STG was
isolated by placing sucrose in a Vaseline well around the stomato-
gastric nerve (stn). Shown on the right are simultaneous extracellular
recordings from the pyloric nerve (pyn), the medial ventricular nerve
(mvn), the pyloric dilator nerve (pdn), the lateral ventricular nerve (lvn),
and the dorsal gastric nerve (dgn). The upper panel shows the
recordings in control saline, and the lower panel shows the initiation
and activation of pyloric and gastric mill rhythms upon the application
of 10)6M NRNFLRFamide.
Fig. 6 MALDI PSD de novo sequencing of an octapeptide (m/z
976.46) fractionated from C. borealis PO extract. The derived amino
acid sequence is shown at the top of the spectrum. In the low mass
region, immonium ions indicative of the presence of Gly (30), Ala (44),
Ser (60), Asn (70, 87), Arg (70, 87 112), Phe (120), and Tyr (136) were
detected in the spectrum. These ions are labeled using single letter
code, with S (Ser), G (Gly), F (Phe), Y (Tyr), A (Ala), N (Asn), R (Arg).
Starting from the high mass end, using the formula [M + H]+ )18
(H2O) –X (where X ¼ each of the 20 amino acid residue masses), the
highest b-type ion is determined to be 796.4, with y1 ion at 181.1, thus
indicating that the C-terminal residue is Tyr, with an amidation modi-
fication. Because the b-type ions were generally accompanied by their
corresponding a-type ions in MALDI PSD fragmentation process, the
b/y ion pairs were identified as following: 796.4/181.1, 640.3/337.2,
526.2/451.2, 455.2/522.3, 292.1/685.3, 145.1/832.4, 88.1/889.4. The
amino acid sequence was then deduced from the mass difference
between consecutive b- or y-ions. Several internal fragment ions are
also labeled in the spectrum.
Neuropeptides in C. borealis pericardial organs 649
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 87, 642–656

ambiguity of the order of the first two amino acids at the
N-terminus. To obtain complementary sequence specific
fragmentation information to allow the unambiguous deter-
mination of the N-terminal sequence, and to confirm the
proposed sequence, we performed ESI MS/MS analysis. We
pooled several fractions containing the peak at m/z 1030 from
the first stage separation and performed a second stage HPLC
separation (see Materials and methods). A repeat of the
separation on a much smaller aliquot was performed with
ESI MS detection inline. This allowed the assignment of m/z-
values for each of the fractions and rapid identification of the
fraction from the larger scale separation that contained the
peak at m/z 1030.5. Figure 7(b) illustrates the ion chroma-
togram resulting from the HPLC-ESI MS separation (inset)
and the resulting full scan MS of the peak eluting at
11.3 min. High resolution, zoomscan spectra for the two
major components confirm that the peak at m/z 1030.5 was
singly charged and the peak at m/z 515.82 was the doubly
charged species.
We used nanospray ionization (NSI) mass spectrometry to
elucidate the amino acid sequence of the peak at m/z 1030.5,
detected in the HPLC fraction. A MS2 collisional spectrum of
the singly charged ion at 1030.5 is presented in Fig. 7(c). The
tandem MS spectrum corroborated the proposed peptide
sequence for the peak at m/z 1030, PAFYSQRY-amide.
Compared to the MALDI PSD data, the detection of y7 ion at
m/z 933.5 allowed the unambiguous assignment of the
N-terminal sequence as PA. MS3 experiments on the b7fragment were also performed (not shown) and gave similar
information on the b6-b3 and y2-y5 fragments. The NSI-MS2
together with the MALDI PSD spectra and identified
immonium ions provide substantive evidence for the pro-
posed sequence.
Interestingly, we also identified three other novel peptides
containing RYamide at their C-terminus using the same MS-
based sequencing and spectral interpretation procedure as
described above. The derived amino acid sequences are
FVNSRYamide, FYANRYamide, and FYSQRYamide. The
(a)
(b)
(c)
Fig. 7 Mass spectrometric characterization of a novel peptide at m/z
1030.51. (a) MALDI PSD fragmentation ion spectrum of precursor ion
at m/z 1030.51. The derived amino acid sequence is shown at the top
of the spectrum. The detected b/y ion pairs are shown on the peptide
sequence, with b-ions at the bottom and y-ions on the top. The frag-
ment ions in the spectrum are labeled using the established nomen-
clature. The immonium ions are labeled with single-letter amino acid
code, with P (Pro), A (Ala), F (Phe), Y (Tyr), S (Ser), Q (Gln), and R
(Arg). Note the ambiguity at the N-terminal sequence. (b) MS spec-
trum from the HPLC-ESI MS assay corresponding to the peak from the
ion chromatogram (inset) eluting at 11.3 min. The singly and doubly
charged species of the m/z 1030 peak are labeled. (c) MS/MS spec-
trum of the singly charged m/z 1030.5 peak. Loss of H2O from the
precursor was observable at m/z 1012.4. The singly charged b7 ion
corresponding to the loss of RY-NH2 was readily visible at m/z 850.3.
Mass spectral peaks at m/z 833.2, 832.3 and 822.3 were assigned as
b7 ion with loss of NH3, H2O (not labeled) and CO (a7 ion). The low
mass cut-off of the ion trap for this parent ion precluded inspection of
the b2 and a2 ions; however, the corresponding y6 ion can be seen at
m/z 862.3. The y6-NH3 ion was also visible. The remaining b and
related ions were readily identifiable; a3/b3 (m/z 287.9 and 316.0),
a4/b4 (m/z 450.9, 479.0), a5 (not labeled), b5-H2O and b5 (m/z 538.1,
548.1, 566.1), and b6-NH3 (not labeled), b6-H2O and b6 (m/z 676.1,
677.2, 694.3). Ions of singly charged y-type fragments likewise can be
identified; y5 and y5-NH3 (m/z 715.3, 698.4), y4 (m/z 552.3), y3 (m/z
465.3), y2 and y2-NH3 (m/z 337.2, 320.2). The tandem MS spectrum
contains the fragmentation ion information (most notably, the y7 ion at
933.5) that supports the proposed sequence of PAFYSQRY-amide.
650 L. Li et al.
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 87, 642–656

sequences of these new peptides are summarized in bold face
in Table 1. We have tested the physiological actions of some
of the RYamide peptides on the STG network; so far no
apparent actions have been found. Study of their actions on
other target tissues in C. borealis is ongoing.
Peptide release from the POs
To investigate the putative hormonal roles of the peptides
present in the POs, we performed mass spectrometric assay
for the peptides released upon high K+ depolarization
(N ¼ 3). Figure 8 shows representative data from one
experiment. Figure 8(a) shows the MALDI mass spectrum
of the releasate collected in normal saline prior to high K+
depolarization. Here, a few peaks were detected, presumably
due to constitutive release. Once the preparation was
transferred to the high K+ saline solution, a dramatic increase
in the number of peptide peaks was seen (Fig. 8b). Several of
these peaks correspond to known peptides including proct-
olin, Cancer borealis tachykinin-related peptide Ib (Cab-
TRP1b), CCAP, SDRNFLRFamide, and Ala13-orcokinin. In
addition, we also detected many of the peptides identified for
the first time in this report, including corazonin, NRNFLRF-
amide, FVNSRYamide, SGFYANRYamide, and PA-
FYSQRYamide, indicating that these new peptides are
released by high K+ depolarizations. Figure 8(c) shows the
result from high K+ depolarization in low Ca2+, high Mn2+
saline (reversible calcium channel blocker); minimal peptide
release was observed, suggesting a Ca2+-dependence of
release. Finally, after several washes with normal Ca2+ saline,
we applied a second high K+ depolarization in normal saline.
Many of the same peptides were released, with slightly
reduced signal intensity (Fig. 8d). The last column in Table 1
summarizes the peptides detected in total of 12 release
experiments. If a peptide peak was detected in over 80% of
the spectra, we consider a peptide to be released (+) from the
POs in response to high K+ depolarization. Due to the low
concentrations of peptides being released, peptide analysis
was performed in linear mode only, with average mass
measurement accuracy at 200 p.p.m.
Discussion
The POs are major neurosecretory structures that can release
amines and peptides into the hemolymph and elicit a variety
of physiological actions. Berlind (1976) demonstrated that
extracts of the POs of crabs injected into intact animals
caused an increase in the frequency of scaphognathite
beating. Neuromodulators found in decapod POs also
modulate the amplitude and frequency of heart beat
(Alexandrowicz and Carlisle 1953). Additionally, it has been
shown that neuroactive substances in the POs altered
properties of cardiac ganglion (Cooke and Hartline 1975),
neuromuscular junctions, and muscle contractibility (Kravitz
et al. 1980; Lingle 1981; Mercier et al. 1990; Meyrand and
Marder 1991; Worden et al. 1995; Jorge-Rivera and Marder
1996; Jorge-Rivera et al. 1998). Furthermore, neurohor-
mones found in the POs, are capable of exerting a wide range
of modulatory effects on the neural circuits in the STG
(Nusbaum and Beenhakker 2002). For all these reasons,
considerable efforts have been made to determine the
composition of the neuroactive substances present in the
POs in various decapod species. Using immunocytochemical
and biochemical techniques, a wide array of neuromodulatory
(a)
(b)
(c)
(d)
Fig. 8 MALDI MS profiling of releasate from C. borealis PO. (a)
MALDI mass spectrum of control sample in normal saline before high
K+ depolarization. Baseline activity is observed with a few peaks
labeled. (b) MALDI mass spectrum of releasate collected from high K+
depolarization. Numerous peptide peaks are detected, with those
labeled with dots being previously known peptides or newly
sequenced peptides in the current study. The inset lists all the pep-
tides identified based on molecular weight measurement. (c) MALDI
profile collected from preparation in low Ca2+, high Mn2+ (reversible
calcium channel blocker) saline with high K+ depolarization. Minimal
peptide release was observed. (d) MALDI mass spectrum of releasate
collected from a second high K+ depolarization after switching the
preparation back in normal saline. Most of the same peptides seen in
(b) were detected. The identified peaks are listed in the inset.
Neuropeptides in C. borealis pericardial organs 651
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 87, 642–656

substances have been identified in the POs in both adult
animals and during embryonic and larval development
(Keller 1992; Christie et al. 1995; Pulver and Marder
2002). In addition to various amines such as serotonin
(Beltz and Kravitz 1983), dopamine (Siwicki et al. 1987),
and octopamine (Evans et al. 1976), many peptides such as
proctolin, FLRFamide-like, CCAP, and orcokinin peptides
have been found in the POs of numerous decapod crustacean
species (Keller 1992; Christie et al. 1995; Pulver and Marder
2002). For example, proctolin is present in the POs of the
lobster, Homarus americanus (Schwarz et al. 1984), the
crayfish, Procamburus clarkii (Siwicki and Bishop 1986),
and the shore crab Carcinus maenas (Stangier et al. 1986).
FMRFamide-like peptides are localized in the POs of the
lobster H. americanus (Kobierski et al. 1987; Trimmer et al.
1987), the crayfish P. clarkii (Mercier et al. 1993), and
several crab species (Krajniak 1991; Christie et al. 1995).
Moreover, allatostatin-like peptides (Skiebe 1999), orcokinin
family peptides (Li et al. 2002; Skiebe et al. 2002; Skiebe
2003), crustacean hyperglycemic hormone (Keller 1992;
Dircksen et al. 2001), and CCAP (Stangier et al. 1988;
Christie et al. 1995; Skiebe et al. 1999) have also been
previously reported to be present in the POs of several
decapod species.
MS-based peptide identification strategy
The current study represents the first mass spectrometric
investigation of the neuromodulatory complement of the POs
in the crab C. borealis. As summarized in Table 1, many
previously identified peptides were detected in direct tissue
MALDI analysis. Only four of these peptides were previ-
ously isolated and sequenced in C. borealis using conven-
tional biochemical techniques that involved multiple steps of
purification of a large pool of tissue samples followed by
Edman degradation (Marder et al. 1986; Weimann et al.
1993; Christie et al. 1997). Other peptides were previously
identified based only on immunoreactivities (Christie et al.
1995; Skiebe 2001; Li et al. 2002). While useful for tissue
localization, immunocytochemistry suffers from limitations
including cross-reactivity with structurally similar peptides
thus preventing the unequivocal identification of a specific
peptide, and the small number of peptides that can be
analyzed simultaneously. As the next step after determining
the identity of a neuromodulator in a particular neuronal
circuit is often determining its physiological role using
exogenous application of synthetic peptides, knowing the
exact chemical structure of the putative hormone is important
information not provided by immunocytochemistry.
MALDI-based peptide identification allows simultaneous
detection of a full spectrum of peptides present at significant
concentrations directly from tissue samples with high mass
accuracy. As shown in the current study, direct tissue
MALDI and PSD sequencing analysis of the LC fractions
resulting from the PO tissue extract confirmed the structures
of several known peptides, such as proctolin, RPCH, CCAP,
extended FLRFamide-related peptides, and orcokinins. In the
case of peptide families, MALDI MS analysis allows
unambiguous identification of the actual forms and different
members of a peptide family. For example, several forms of
the extended FLRFamide-related peptides (including a novel
truncated form of the FLRFamide peptide) were identified in
the POs to substantiate the positive FLRFamide immunore-
activity documented previously. Similarly, Ala13-orcokinin
was detected in the PO tissue samples by MALDI MS, and
two additional forms of orcokinins (Val13- and Ser9-orcoki-
nins) were detected in LC fractions from pooled PO tissue
extract, indicating a possible differential expression of
different forms of orcokinins in the PO, as the latter two
forms of orcokinins were not observed in MALDI spectra of
the tissue samples surveyed. These two forms of orcokinins
are likely expressed in the different regions of the PO tissue
from that of Ala13-orcokinin, or at much lower level to be
detected in individual organs by MS. As we have previously
demonstrated simultaneous detection of five different forms
of orcokinins, including Ala13-, Val13-, and Asn13-orcokinins
in the PO tissue from H. americanus (Li et al. 2002), it is
unlikely that the absence of Val13- and Ser9-orcokinins is due
to the difference of ionization efficiency or analyte suppres-
sion of Ala13-orcokinin. Thus, it is advantageous to use the
combination of direct tissue profiling and HPLC fraction-
ation of pooled tissue extract to generate a more complete
characterization of peptides present in the POs.
As demonstrated in Fig. 1, there are many mass spectral
peaks that do not correspond to previously identified
peptides. To characterize several of these new peptides, we
performed de novo MALDI PSD sequencing of a number of
unknown peaks fractionated from tissue extracts. To simplify
the complex mixture resulting from the PO tissue extract and
also concentrate the peaks, microbore LC separation was
performed prior to MALDI MS analysis. While on-line LC
coupling with ESI MS/MS has been the preferred method for
large-scale peptide identification from protein digests and
other complex mixtures, the off-line coupling MALDI PSD
approach has some advantages, including the ability to select
many more peaks for sequencing without time constraints.
This is particularly useful for analyzing complex tissue
extracts, where many peptides coelute and low abundance
peptides are often missed in LC ESI-MS/MS analysis due to
the preferential selection and identification of high-intensity
peaks in the elution time window. As evident from Fig. 3,
multiple PSD sequencing analyses were performed on
several precursor ions from individual LC fractions, yielding
enhanced chemical information from the limited amount of
samples. Because no genomic sequence information is
available for C. borealis, only peptides producing complete
fragmentation allowed the derivation of full amino acid
sequence. While all of the HPLC fractions were analyzed by
mass spectrometry, many LC fractions contained larger
652 L. Li et al.
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 87, 642–656

peptides (precursor mass > 2000 Da) that fragment less
efficiently, which makes it difficult to generate complete
sequence information. Furthermore, several peptides were
eluted in a few consecutive LC fractions, so we were only
able to fully sequence seven new peptides based on PSD
fragmentation analysis, but many more peptides generated
partial sequence-specific fragmentation that did not yield
complete peptide identification. We have also demonstrated
the use of combination of MALDI-PSD and nanoESI CID
fragmentation techniques to generate complementary
sequence-specific fragmentation information to allow the
complete characterization of the peak at m/z 1030.51. While
CID generates more efficient fragmentation, the low mass
cut-off limitation associated with the ion trap mass analyzer
prevents the effective observation of low mass regions. In
contrast, PSD produces abundant immonium ions in the low
mass region that are indicative of amino acid compositions,
which are especially useful for de novo sequencing of
unknown peptides.
FMRFamide-like peptides
FMRFamide-like peptides are perhaps the most widely
distributed neuropeptides in the animal kingdom. Since their
first discovery in mollusks (Price and Greenberg 1977), a
large number of related peptides have been chemically
characterized in many different phyla (De Loof and Schoofs
1990; Krajniak 1991; Mercier et al. 1991; Mercier et al.
1993; Schoofs et al. 1997; Li et al. 1999a; Sithigorngul
et al. 2001; Baggerman et al. 2002). Two extended forms of
FLRFamide, TNRNFLRFamide and SDRNFLRFamide were
previously purified and sequenced in the lobster H. americ-
anus (Trimmer et al. 1987) and the crab C. borealis
(Weimann et al. 1993). Here we report a new member of
the extended FLRFamide peptide, NRNFLRFamide in the
C. borealis based on de novo sequencing analysis of the PO
extract. This peptide was previously isolated in the POs from
crayfish, P. clarkii (Mercier et al. 1993). This result is
consistent with previous observation that additional FMRF-
amide-like immunoreactive HPLC fractions were detected in
the crab nervous system (Marder et al. 1987; Weimann et al.
1993). Physiological experiments demonstrated that
NRNFLRFamide elicits effects on the pyloric and gastric
mill rhythms similar to those seen with TNRNFLRFamide
and SDRNFLRFamide. This may suggest that only the
sequence of RNFLRFamide is important for receptor recog-
nition and binding. It is also interesting to note that
NRNFLRFamide is often the most intense peak in the mass
spectra of the PO tissue samples, suggesting a differential
expression of these peptides in the POs and the STNS, and a
potential neurohormonal role of the peptide.
Corazonin
While corazonin has been reported in a number of insect
species (Veenstra 1989, 1991, 1994; Schoofs et al. 2000;
Hansen et al. 2001), the occurrence of corazonin in crusta-
cean species is documented for the first time in the current
study. Consistent with the highly conserved amino acid
sequence of this peptide throughout arthropod species, the
de novo sequencing of the putative corazonin peak in
C. borealis PO extract revealed the authentic form of this
peptide is also found in C. borealis. With the availability of
multiple corazonin gene and precursor sequences of several
insect species, it was possible to use a multiple sequence
alignment procedure to locate the highly conserved
sequence. We then employed this template sequence as a
guide to search for putative corazonin in crustacean species.
Despite the highly conserved peptide structure, the functions
of corazonin appear to be species specific and even tissue
specific. For example, this peptide is highly effective at
stimulating the activity of the heart and hyperneural muscle,
whereas other visceral muscles are completely insensitive to
this peptide (Predel and Eckert 2000). The corazonin
myostimulatory effect is mainly restricted to the American
cockroach, Periplaneta americana (Predel and Eckert 2000;
Predel et al. 2001), while in locust His7-corazonin induces
body color pigmentation (Tawfik et al. 1999; Schoofs et al.
2000). Electrophysiological experiments have demonstrated
strong modulatory effects on the pyloric rhythm of the
isolated STG upon application of synthetic corazonin (A.E.
Christie, unpublished observation), which adds to the wide
spectrum of the functions of the corazonin peptide.
CHH and other larger peptides in the PO
Although the major focus of the current study was charac-
terizing peptides in the lower mass range, several peaks were
detected in the mass range from 2000 to 9650 Da. One
notable peptide detected in the MALDI profiling of the POs
is a peak at 8562 Da, a possible candidate for CHH in
C. borealis. CHH are involved in various physiological
processes including regulation of blood glucose and lipids.
Since the first identification of a SG-CHH in the shore crab
Carcinus maenas, more than 20 SG-derived CHHs have been
isolated and identified in various species of crustacea (Keller
1992). While the primary source of CHHs is in the SG in the
eyestalk, multiple forms of novel CHH-like peptides were
recently reported in the POs from the shore crab, C. maenas
(Dircksen et al. 2001). Given the molecular masses of these
previously identified CHHs, it is possible that the peak at
8562 Da might be the CHH present in C. borealis with
several amino acid substitutions. Due to the inefficiency of
MS/MS fragmentation for peptides larger than 2500 Da, the
MS-based sequence analysis of the intact CHH peptide was
not possible; therefore, the conclusive peptide assignment
can not be made without additional confirmation. The
detection of a mass spectral peak at m/z 4072, which
corresponds to a putative peptide encoded by the CHH
precursor, supported the presence of CHH in the POs of
C. borealis. The identical molecular weight (and likely
Neuropeptides in C. borealis pericardial organs 653
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 87, 642–656

sequence) of CPRP in C. borealis to that of CPRP from
C. maenas supports earlier observation that PO-CHH and
SG-CHH share an identical N-terminal sequence (positions
1–40), but differ considerably in the remaining sequence.
RYamides and peptide release
Finally, we sequenced several new peptides sharing a
common C-terminal, RYamides. While the physiological
effects of these newly sequenced peptides are not known, the
fact that these peptides are C-terminally amidated and
released upon high K+ depolarization strongly suggests
neurohormonal roles for these peptides. Several of these new
RYamide peptides are coreleased with NRNFLRFamide,
corazonin, proctolin, CCAP, and Ala13-orcokinin by high K+
depolarization in a Ca2+-dependent manner. This is also the
first demonstration of assaying peptide release from
C. borealis POs using mass spectrometric techniques. While
some variability was observed between different preparations
and the first and second high K+ depolarizations, several
peptides including proctolin, CabTRP1b, FVNSRYamide,
NRNFLRFamide, SGFYANRYamide, PAFYQSRYamide,
and Ala13-ocrokinin were always released. It is interesting
to note that CabTRP1b was consistently detected in releasate,
yet no CabTRP1a was detected. Previous studies on the two
tachykinin-related peptides showed that CabTRP1a is 20
times more abundant and 500 times more potent than
CabTRP1b (Christie et al. 1997). These results led to the
speculation that CabTRP1b is a breakdown product of
CabTRP1a. However, the detection of CabTRP1b in relea-
sates in the current study suggests that this shorter form of
the peptide may be directly cleaved from the precursor
protein and could serve a physiological role in C. borealis.
In summary, the combination of both direct tissue MALDI
profiling and MS-based sequencing allows comprehensive
characterization of the peptide complement in a nervous
system at higher throughput with greater chemical details.
The large-scale mass spectrometric investigation of neuro-
peptides and hormones in the pericardial organs of C. borealis
revealed much greater diversity and complexity of the
peptide messengers than had been previously demonstrated
by immunocytochemical and electrophysiological approa-
ches. We confirmed many of the previously known peptides
and unambiguously identified different chemical forms of the
peptide families. Furthermore, we have fully sequenced and
identified several new peptides. However, it is evident from
the current study that many more peptides remain unchar-
acterized. It is worth noting that members of several well-
known peptide families visualized in immunocytochemical
studies were not observed. This is likely due to amino acid
variations from the authentic forms found in other species,
post-translational modifications and perhaps cross-immuno-
reactivity and/or insufficient sensitivity of the current MS
methods. Future work will aim to characterize these peptides
by employing the combination of immunoaffinity and
MS-based sequencing approaches. Furthermore, the de novo
sequencing methodology will be coupled with database
searching via homology from related species whose genomic
sequences are available. Such peptidomic approaches prom-
ise to significantly accelerate the discovery of new peptides
and yield a complete description of the peptide signaling
molecules involved in the crustacean nervous system and to
further increase our understanding of peptide functions at the
network level.
Acknowledgements
This work was supported by National Institute of Neurological
Disorder and Stroke grants NS17813 (EM) and NS31609 (JVS). We
thank Dr Michael Nusbaum (University of Pennsylvania School of
Medicine) for the gift of synthetic corazonin.
References
Alexandrowicz J. S. and Carlisle D. B. (1953) Some experiments on the
function of the pericardial organs in Crustacea. J. Mar. Biol. Assoc.
UK 32, 175–192.
Baggerman G., Cerstiaens A., De Loof A. and Schoofs L. (2002) Pep-
tidomics of the larval Drosophila melanogaster central nervous
system. J. Biol. Chem. 277, 40368–40374.
Beltz B. S. and Kravitz E. A. (1983) Mapping of serotonin-like im-
munoreactivity in the lobster nervous system. J. Neurosci. 3,
585–602.
Berlind A. (1976) Neurohemal organ extracts effect the ventilation
oscillator in crustaceans. J. Exp. Zool. 195, 165–170.
Christie A. E. and Nusbaum M. P. (1995) Distribution and effects of
corazonin-like and allatotropin-like peptides in the crab stomato-
gastric nervous system. Soc. Neurosci. Abstr. 21, 629.
Christie A. E., Skiebe P. and Marder E. (1995) Matrix of neuromodu-
lators in neurosecretory structures of the crab, Cancer borealis.
J. Exp. Biol. 198, 2431–2439.
Christie A. E., Lundquist T., Nassel D. R. and Nusbaum M. P. (1997)
Two novel tachykinin-related peptides from the nervous system of
the crab Cancer borealis. J. Exp. Biol. 200, 2279–2294.
Cooke I. M. and Hartline D. K. (1975) Neurohormonal alteration of
integrative properties of the cardiac ganglion of the lobster
Homarus americanus. J. Exp. Biol. 63, 33–52.
De Loof A. and Schoofs L. (1990) Homologies between the amino acid
sequences of some vertebrate peptide hormones and peptides iso-
lated from invertebrate sources. Comp. Biochem. Physiol. B 95,
459–468.
Dircksen H., Bocking D., Heyn U., Mandel C., Chung J. S., Baggerman
G., Verhaert P., Daufeldt S., Plosch T., Jaros P. P. et al. (2001)
Crustacean hyperglycaemic hormone (CHH)-like peptides and
CHH-precursor-related peptides from pericardial organ neurose-
cretory cells in the shore crab, Carcinus maenas, are putatively
spliced and modified products of multiple genes. Biochem. J. 356,
159–170.
Evans P. D., Kravitz E. A., Talamo B. R. and Wallace B. G. (1976) The
association of octopamine with specific neurones along lobster
nerve trunks. J. Physiol. 262, 51–70.
Floyd P. D., Li L., Moroz T. P. and Sweedler J. V. (1999) Characteri-
zation of peptides from Aplysia using microbore liquid
chromatography with matrix-assisted laser desorption/ionization
time-of-flight mass spectrometry guided purification. J. Chroma-
togr. A 830, 105–113.
654 L. Li et al.
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 87, 642–656

Garden R. W., Moroz L. L., Moroz T. P., Shippy S. A. and Sweedler J. V.
(1996) Excess salt removal with matrix rinsing: direct peptide
profiling of neurons from marine invertebrates using matrix-
assisted laser desorption/ionization time-of-flight mass spectrome-
try. J. Mass Spectrom. 31, 1126–1130.
Hansen I. A., Sehnal F., Meyer S. R. and Scheller K. (2001) Corazonin
gene expression in the waxmoth Galleria mellonella. Insect Mol
Biol. 10, 341–346.
Harris-Warrick R. M., Marder E., Selverston A. I. and Moulins M.
(1992) Dynamic Biological Networks. The Stomatogastric Nervous
System, p. 328. MIT Press, Cambridge.
Jimenez C. R., van Veelen P. A., Li K. W., Wildering W. C., Geraerts W.
P., Tjaden U. R. and van der Greef J. (1994) Neuropeptide
expression and processing as revealed by direct matrix-assisted
laser desorption ionization mass spectrometry of single neurons.
J. Neurochem. 62, 404–407.
Jorge-Rivera J. C. and Marder E. (1996) TNRNFLRFamide and
SDRNFLRFamide modulate muscles of the stomatogastric system
of the crab Cancer borealis. J. Comp. Physiol. A 179, 741–751.
Jorge-Rivera J. C., Sen K., Birmingham J. T., Abbott L. F. and Marder E.
(1998) Temporal dynamics of convergent modulation at a crusta-
cean neuromuscular junction. J. Neurophysiol. 80, 2559–2570.
Kaufmann R., Spengler B. and Lutzenkirchen F. (1993) Mass spectro-
metric sequencing of linear peptides by product-ion analysis in a
reflectron time-of-flight mass spectrometer using matrix-assisted
laser desorption ionization. Rapid Commun. Mass Spectrom. 7,
902–910.
Keller R. (1992) Crustacean neuropeptides: structures, functions and
comparative aspects. Experientia 48, 439–448.
Kobierski L. A., Beltz B. S., Trimmer B. A. and Kravitz E. A. (1987)
FMRFamidelike peptides of Homarus americanus: distribution,
immunocytochemical mapping, and ultrastructural localization in
terminal varicosities. J. Comp. Neurol. 266, 1–15.
Krajniak K. G. (1991) The identification and structure-activity relations
of a cardioactive FMRFamide-related peptide from the blue crab
Callinectes sapidus. Peptides 12, 1295–1302.
Kravitz E. A., Glusman S., Harris-Warrick R. M., Livingstone M. S.,
Schwarz T. and Goy M. F. (1980) Amines and a peptide as neu-
rohormones in lobsters: actions on neuromuscular preparations and
preliminary behavioural studies. J. Exp. Biol. 89, 159–175.
Li L., Moroz T. P., Garden R. W., Floyd P. D., Weiss K. R. and Sweedler
J. V. (1998) Mass spectrometric survey of interganglionically
transported peptides in Aplysia. Peptides 19, 1425–1433.
Li C., Nelson L. S., Kim K., Nathoo A. and Hart A. C. (1999a) Neu-
ropeptide gene families in the nematode Caenorhabditis elegans.
Ann. NY Acad. Sci. 897, 239–252.
Li L., Garden R. W., Romanova E. V. and Sweedler J. V. (1999b) In
situ sequencing of peptides from biological tissues and single
cells using MALDI-PSD/CID analysis. Anal. Chem. 71, 5451–
5458.
Li L., Garden R. W. and Sweedler J. V. (2000a) Single-cell MALDI: a
new tool for direct peptide profiling. Trends Biotechnol. 18, 151–
160.
Li L., Romanova E. V., Rubakhin S. S., Alexeeva V., Weiss K. R., Vilim
F. S. and Sweedler J. V. (2000b) Peptide profiling of cells with
multiple gene products: combining immunochemistry and MALDI
mass spectrometry with on-plate microextraction. Anal. Chem. 72,
3867–3874.
Li L., Pulver S. R., Kelley W. P., Thirumalai V., Sweedler J. V. and
Marder E. (2002) Orcokinin peptides in developing and adult
crustacean stomatogastric nervous systems and pericardial organs.
J. Comp. Neurol. 444, 227–244.
Lingle C. (1981) The modulatory action of dopamine on crustacean
foregut neuromuscular preparations. J. Exp. Biol. 94, 285–299.
Marder E. (1987) Neurotransmitters and neuromodulators, in The
Crustacean Stomatogastric Nervous System. A Model for the Study
of Central Nervous Systems. (Selverston, A. I. and Moulins, M.,
eds), pp. 263–300. Springer-Verlag, New York.
Marder E. and Hooper S. L. (1985) Neurotransmitter modulation of the
stomatogastric ganglion of decapod crustaceans, in Model Neural
Networks and Behavior (Selverston, A. I., ed.), pp. 319–337.
Plenum Press, New York.
Marder E. and Calabrese R. L. (1996) Principles of rhythmic motor
pattern generation. Physiol. Rev. 76, 687–717.
Marder E., Christie A. E. and Kilman V. L. (1995) Functional organ-
ization of cotransmission systems: lessons from small nervous
systems. Invert. Neurosci. 1, 105–112.
Marder E., Hooper S. L. and Siwicki K. K. (1986) Modulatory action
and distribution of the neuropeptide proctolin in the crustacean
stomatogastric nervous system. J. Comp. Neurol. 243, 454–467.
Marder E., Calabrese R. L., Nusbaum M. P. and Trimmer B. A. (1987)
Distribution and partial characterization of FMRFamide-like pep-
tides in the stomatogastric nervous systems of the rock crab,
Cancer borealis, and the spiny lobster, Panulirus interruptus.
J. Comp. Neurol. 259, 150–163.
Mercier A. J., Orchard I. and TeBrugge V. (1991) FMRFamide-like
immunoreactivity in the crayfish nervous system. J. Exp. Biol. 156,
519–538.
Mercier A. J., Schiebe M. and Atwood H. L. (1990) Pericardial peptides
enhance synaptic transmission and tension in phasic extensor
muscles of crayfish. Neurosci. Lett. 111, 92–98.
Mercier A. J., Orchard I., TeBrugge V. and Skerrett M. (1993) Isolation
of two FMRFamide-related peptides from crayfish pericardial
organs. Peptides 14, 137–143.
Meyrand P. and Marder E. (1991) Matching neural and muscle oscilla-
tors: control by FMRFamide-like peptides. J. Neurosci. 11, 1150–
1161.
Nusbaum M. P. and Beenhakker M. P. (2002) A small-systems approach
to motor pattern generation. Nature 417, 343–350.
Predel R. (2001) Peptidergic neurohemal system of an insect: mass
spectrometric morphology. J. Comp Neurol. 436, 363–375.
Predel R. and Eckert M. (2000) Neurosecretion: peptidergic systems in
insects. Naturwissenschaften 87, 343–350.
Predel R., Nachman R. J. and Gade G. (2001) Myostimulatory neuro-
peptides in cockroaches: structures, distribution, pharmacological
activities, and mimetic analogs. J. Insect. Physiol. 47, 311–324.
Price D. A. and Greenberg M. J. (1977) Purification and characterization
of a cardioexcitatory neuropeptide from the central ganglia of a
bivalve mollusc. Prep. Biochem. 7, 261–281.
Pulver S. R. and Marder E. (2002) Neuromodulatory complement of the
pericardial organs in the embryonic lobster, Homarus americanus.
J. Comp. Neurol. 451, 79–90.
Roepstorff P. and Fohlman J. (1984) Proposal for a common nomen-
clature for sequence ions in mass spectra of peptides. Biomed.
Mass Spectrom. 11, 601.
Schoofs L., Veelaert D., Vanden Broeck J. and De Loof A. (1997)
Peptides in the locusts, Locusta migratoria and Schistocerca gre-
garia. Peptides 18, 145–156.
Schoofs L., BaggermanG., Veelaert D., BreuerM., Tanaka S. andDe Loof
A. (2000) The pigmentotropic hormone [His(7)]-corazonin, absent
in a Locusta migratoria albino strain, occurs in an albino strain of
Schistocerca gregaria. Mol. Cell Endocrinol. 168, 101–109.
Schwarz T. L., Lee G. M., Siwicki K. K., Standaert D. G. and Kravitz
E. A. (1984) Proctolin in the lobster: the distribution, release, and
chemical characterization of a likely neurohormone. J. Neurosci. 4,
1300–1311.
Sithigorngul P., Saraithongkum W., Longyant S., Panchan N.,
Sithigorngul W. and Petsom A. (2001) Three more novel FMRF-
Neuropeptides in C. borealis pericardial organs 655
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 87, 642–656

amide-like neuropeptide sequences from the eyestalk of the giant
freshwater prawn Macrobrachium rosenbergii. Peptides 22, 191–
197.
Siwicki K. K. and Bishop C. A. (1986) Mapping of proctolinlike im-
munoreactivity in the nervous systems of lobster and crayfish.
J. Comp. Neurol. 243, 435–453.
Siwicki K. K., Beltz B. S. and Kravitz E. A. (1987) Proctolin in iden-
tified serotonergic, dopaminergic, and cholinergic neurons in the
lobster, Homarus americanus. J. Neurosci. 7, 522–532.
Skiebe P. (1999) Allatostatin-like immunoreactivity within the stoma-
togastric nervous system and the pericardial organs of the crab
Cancer pagurus, the lobster Homarus americanus, and the crayfish
Cherax destructor and Procambarus clarkii. J. Comp. Neurol. 403,
85–105.
Skiebe P. (2001) Neuropeptides are ubiquitous chemical mediators:
using the stomatogastric nervous system as a model system. J. Exp.
Biol. 204, 2035–2048.
Skiebe P. (2003) Neuropeptides in the crayfish stomatogastric nervous
system. Microsc. Res. Tech. 60, 302–312.
Skiebe P., Dietel C. and Schmidt M. (1999) Immunocytochemical
localization of FLRFamide-, proctolin-, and CCAP- like peptides
in the stomatogastric nervous system and neurohemal structures of
the crayfish, Cherax destructor. J. Comp. Neurol. 414, 511–532.
Skiebe P., Dreger M., Meseke M., Evers J. F. and Hucho F. (2002)
Identification of orcokinins in single neurons in the stomatogastric
nervous system of the crayfish, Cherax destructor. J. Comp. Neu-
rol. 444, 245–259.
Stangier J., Dircksen H. and Keller R. (1986) Identification and
immunocytochemical localization of proctolin in pericardial organs
of the shore crab, Carcinus maenas. Peptides 7, 67–72.
Stangier J., Hilbich C., Dircksen H. and Keller R. (1988) Distribution of
a novel cardioactive peptide (CCAP) from pericardial organs of the
shore crab, Carcinus maenas. Peptides 9, 795–800.
Tawfik A. I., Tanaka S., De Loof A., Schoofs L., Baggerman G.,
Waelkens E., Derua R., Milner Y., Yerushalmi Y. and Pener M. P.
(1999) Identification of the gregarization-associated dark-pigmen-
totropin in locusts through an albino mutant. Proc. Natl Acad. Sci.
USA 96, 7083–7087.
Trimmer B. A., Kobierski L. A. and Kravitz E. A. (1987) Purification
and characterization of FMRFamide-like immunoreactivity sub-
stances from the lobster nervous system: isolation and sequence
analysis of two closely related peptides. J. Comp. Neurol. 266, 16–
26.
Veenstra J. A. (1989) Isolation and structure of corazonin, a cardio-
active peptide from the American cockroach. FEBS Lett. 250,
231–234.
Veenstra J. A. (1991) Presence of corazonin in three insect species, and
isolation and identification of [His7]corazonin from Schistocerca
americana. Peptides 12, 1285–1289.
Veenstra J. A. (1994) Isolation and structure of the Drosophila corazonin
gene. Biochem. Biophys. Res. Commun. 204, 292–296.
Weimann J. M., Marder E., Evans B. and Calabrese R. L. (1993) The
effects of SDRNFLRFamide and TNRNFLRFamide on the motor
patterns of the stomatogastric ganglion of the crab Cancer borealis.
J. Exp. Biol. 181, 1–26.
Worden M. K., Kravitz E. A. and Goy M. F. (1995) Peptide F1, an
N-terminally extended analog of FMRFamide, enhances contractile
activity in multiple target tissues in lobster. J. Exp. Biol. 198, 97–
108.
656 L. Li et al.
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 87, 642–656
Copyright © 2022 FDOKUMEN




















