
LONG-WAVELENGTH UVA RADIATION INDUCES OXIDATIVE STRESS, CYTOSKELETAL DAMAGE and HEMOLYSIS
9
Photochemistry and Photobioiogy, Vol. 57, No. 6, pp. 1018-1026, 1993 Printed in the United States. All rights reserved 003 1-8655/93 $05.00+0.00 0 L993 American Society for Photobiology LONG-WAVELENGTH WA RADIATION INDUCES OXIDATIVE STRESS, CYTOSKELETAL DAMAGE AND HEMOLYSIS* DIANNE E. GODARt, DELMA P. THOMAS, SHARON A. MILLER and WAH LEE Center for Devices and Radiological Health, Food and Drug Administration, Rockville, MD 20857, USA (Received 3 August 1992; accepted 2 November 1992) Abstract- We investigatedthe ability of the different wavelength regions of UV radiation, UVA (320-400 nm), UVB (290-320 nm) and UVC (200-290 nm), to induce hemolysis. Sheep erythrocytes were exposed to radiation from either a UVAl (>340 nm) sunlamp, a UVB sunlamp, or a UVC germicidal lamp. The doses used for the three wavelength regions were approximately equilethal to the survival of L5 178Y murine lymphoma cells. Following exposure, negligible hemolysis was observed in the UVB- and UVC-irradiated erythrocytes, whereas a decrease in the relative cell number (RCN), indicative of hemolysis, was observed in the UVAl-exposed samples. The decrease in RCN was dependent on dose (0-1625 kJ/m2),time (0-78 h postirradiation) and cell density (106-107 cells/mL). Hemolysis decreased with increasing concentration of glutathione, hemoglobin or cell number, while the presence of pyruvate drasticallyenhanced it. Because scanning spectroscopy (200-700 nm) showed that hemoproteins and nicotin- amide adenine dinucleotides were oxidized, cytoplasmic oxidative stress was implicated in the lytic mechanism. Further evidence of oxidation was obtained from electron micrographs, which revealed the formation of Heinz bodies near the plasma membrane. The data demonstrate that exposure of erythrocytes to UVAl, but not UVB or UVC, radiation causes oxidation of cytoplasmic components, which results in cytoskeletal damage and hemolysis. INTRODUCTION UV radiation (UVR)$ of shorter wavelengths, UVB (290- 320 nm) and UVC (200-290 nm), have been the most ex- tensively investigated wavelength regions due to the direct effects on genetic material.’ Long-wavelength UVR, or UVA (320-400 nm), was thought to be rather innocuous to genetic material when compared to either UVC or UVB radiation on a per quantum basis. However, when equilethal doses (fluence required to kill 37% of the target population) were used for comparison, UVA photons were discovered to be about an order of magnitude or more effective in causing oxidative damage to DNA, DNA-to-protein cross-links and single strand breaks, than UVB or UVC photons.2 Over the course of time, many findings have demonstrated that a va- riety of cytotoxic cellular effects do occur from exposure of both prokaryotic and eukaryotic cells to UVA radiation. Exposure of prokaryotic cells to UVA radiation results in a variety of deleterious effects to different cellular compo- nents, which may be responsible for the cellular inactivation originally observed by H~llaender.~ DNA damage results from prokaryotic exposure to UVA radiation and causes sin- *Dedicated to Jack Johnson (father of D.E.G.), deceased 17 May +To whom correspondence should be addressed. *Abbreviations: CALC, calculated; GSH, reduced glutathione; HEPES, N-[2-hydroxyethyl] piperazine-K-12-ethane sulfonic acid]; HPLC, high-performanceliquid chromatography; IR, infrared; NADH, reduced nicotinamide adenine dinucleotide; NADPH, reduced nicotinamide adenine dinucleotide phosphate; OD, optical den- sity; PBS, phosphate-buffered saline; PYR, pyruvate; RBC, red blood cell; RCN, relative cell number (i.e. the number of cells in the exposed sample normalized by the number of cells in the sham-exposed sample); STD, standard;TBA, thiobarbituricacid; TEM, transmission electron microscopy; UVA, ultraviolet-A (320-400 nm); UVAI, ultraviolet-A1 (340-400 nm); UVB, ultraviolet-B (290-320 nm); UVC, ultraviolet-C (200-290 nm); UVR, ultraviolet radiation. 1992. gle strand cyclobutane-type pyrimidine and pyrimidine 6-4 adducts.’ In addition to DNA other cel- lular components, such as lipids,8 cell membrane^,'.^.'^ RNA,“-j3 DNA repair enzymes, transport and metabolic systemsI4 are all damaged by UVA radiation. In addition to cytoplasmic components, nucleated eu- karyotes have organelles that can be damaged by exposure to UVA radiation. It is well established that exposure of eukaryotic cells to UVA radiation results in cellular inacti- vation through oxidative stress.I5 UVA-induced oxidative damage has been reported to occur to lipids,r619 the coen- zymes reduced nicotinamide adenine dinucleotide (NADH) and reduced nicotinamide adenine dinucleotide phosphate (NADPH),2u.Z’ tryptophanzzand DNA.’.2 On the level of or- ganelles, nucleated mammalian cells were reported to have damaged cellular structures after exposure to UVA radiation; such as, the microtubules of the cyto~keleton,~~ the plasma the nuclear membrane and the rough en- doplasmic r etic~lum.~~,~~ Furthermore, UVAl radiation (340- 400 nm) has been reported to induce cell lysis in some mam- malian nucleated cell types as well as in anucleated cells, i.e. erythrocytes.26 The simplicity of the mammalian red blood cell (RBC) structural and biochemical system renders RBC as conve- nient models for investigating anuclear effects as well as bi- ological mechanisms in eukaryotes. Exposure of RBC to bi- ologically relevant doses of UVAl radiation results in hemolysi~,~~~~~ as is also the case for biologically and envi- ronmentally irrelevant doses of UVC.z8-3’ Slight hemolysis was previously observed in UVA-irradiated RBC samples in the absence of photo~ensitizer,~~.~~ but no further investiga- tions of this phenomenon have been reported. Nevertheless, UVAI-induced anuclear damage, which may lead to he- molysis, has been, until recently, basically overlooked. Thus, the mechanism of UVA 1 -induced hemolysis is not currently understood, though photosensitized mechanisms have been investigated at great length.33 1018
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of LONG-WAVELENGTH UVA RADIATION INDUCES OXIDATIVE STRESS, CYTOSKELETAL DAMAGE and HEMOLYSIS

Photochemistry and Photobioiogy, Vol. 57, No. 6, pp. 1018-1026, 1993 Printed in the United States. All rights reserved
003 1-8655/93 $05.00+0.00 0 L993 American Society for Photobiology
LONG-WAVELENGTH W A RADIATION INDUCES OXIDATIVE STRESS, CYTOSKELETAL DAMAGE AND HEMOLYSIS*
DIANNE E. GODARt, DELMA P. THOMAS, SHARON A. MILLER and WAH LEE Center for Devices and Radiological Health, Food and Drug Administration, Rockville, MD 20857, USA
(Received 3 August 1992; accepted 2 November 1992)
Abstract- We investigated the ability of the different wavelength regions of UV radiation, UVA (320-400 nm), UVB (290-320 nm) and UVC (200-290 nm), to induce hemolysis. Sheep erythrocytes were exposed to radiation from either a UVAl (>340 nm) sunlamp, a UVB sunlamp, or a UVC germicidal lamp. The doses used for the three wavelength regions were approximately equilethal to the survival of L5 178Y murine lymphoma cells. Following exposure, negligible hemolysis was observed in the UVB- and UVC-irradiated erythrocytes, whereas a decrease in the relative cell number (RCN), indicative of hemolysis, was observed in the UVAl-exposed samples. The decrease in RCN was dependent on dose (0-1625 kJ/m2), time (0-78 h postirradiation) and cell density (106-107 cells/mL). Hemolysis decreased with increasing concentration of glutathione, hemoglobin or cell number, while the presence of pyruvate drastically enhanced it. Because scanning spectroscopy (200-700 nm) showed that hemoproteins and nicotin- amide adenine dinucleotides were oxidized, cytoplasmic oxidative stress was implicated in the lytic mechanism. Further evidence of oxidation was obtained from electron micrographs, which revealed the formation of Heinz bodies near the plasma membrane. The data demonstrate that exposure of erythrocytes to UVAl, but not UVB or UVC, radiation causes oxidation of cytoplasmic components, which results in cytoskeletal damage and hemolysis.
INTRODUCTION
UV radiation (UVR)$ of shorter wavelengths, UVB (290- 320 nm) and UVC (200-290 nm), have been the most ex- tensively investigated wavelength regions due to the direct effects on genetic material.’ Long-wavelength UVR, or UVA (320-400 nm), was thought to be rather innocuous to genetic material when compared to either UVC or UVB radiation on a per quantum basis. However, when equilethal doses (fluence required to kill 37% of the target population) were used for comparison, UVA photons were discovered to be about an order of magnitude or more effective in causing oxidative damage to DNA, DNA-to-protein cross-links and single strand breaks, than UVB or UVC photons.2 Over the course of time, many findings have demonstrated that a va- riety of cytotoxic cellular effects do occur from exposure of both prokaryotic and eukaryotic cells to UVA radiation.
Exposure of prokaryotic cells to UVA radiation results in a variety of deleterious effects to different cellular compo- nents, which may be responsible for the cellular inactivation originally observed by H ~ l l a e n d e r . ~ DNA damage results from prokaryotic exposure to UVA radiation and causes sin-
*Dedicated to Jack Johnson (father of D.E.G.), deceased 17 May
+To whom correspondence should be addressed. *Abbreviations: CALC, calculated; GSH, reduced glutathione; HEPES,
N-[2-hydroxyethyl] piperazine-K-12-ethane sulfonic acid]; HPLC, high-performance liquid chromatography; IR, infrared; NADH, reduced nicotinamide adenine dinucleotide; NADPH, reduced nicotinamide adenine dinucleotide phosphate; OD, optical den- sity; PBS, phosphate-buffered saline; PYR, pyruvate; RBC, red blood cell; RCN, relative cell number (i.e. the number of cells in the exposed sample normalized by the number of cells in the sham-exposed sample); STD, standard; TBA, thiobarbituric acid; TEM, transmission electron microscopy; UVA, ultraviolet-A (320-400 nm); UVAI, ultraviolet-A1 (340-400 nm); UVB, ultraviolet-B (290-320 nm); UVC, ultraviolet-C (200-290 nm); UVR, ultraviolet radiation.
1992.
gle strand cyclobutane-type pyrimidine and pyrimidine 6-4 adducts.’ In addition to DNA other cel- lular components, such as lipids,8 cell membrane^,'.^.'^ RNA,“-j3 DNA repair enzymes, transport and metabolic systemsI4 are all damaged by UVA radiation.
In addition to cytoplasmic components, nucleated eu- karyotes have organelles that can be damaged by exposure to UVA radiation. It is well established that exposure of eukaryotic cells to UVA radiation results in cellular inacti- vation through oxidative stress.I5 UVA-induced oxidative damage has been reported to occur to lipids,r619 the coen- zymes reduced nicotinamide adenine dinucleotide (NADH) and reduced nicotinamide adenine dinucleotide phosphate (NADPH),2u.Z’ tryptophanzz and DNA.’.2 On the level of or- ganelles, nucleated mammalian cells were reported to have damaged cellular structures after exposure to UVA radiation; such as, the microtubules of the cy to~ke le ton ,~~ the plasma
the nuclear membrane and the rough en- doplasmic r e t i c ~ l u m . ~ ~ , ~ ~ Furthermore, UVAl radiation (340- 400 nm) has been reported to induce cell lysis in some mam- malian nucleated cell types as well as in anucleated cells, i.e. erythrocytes.26
The simplicity of the mammalian red blood cell (RBC) structural and biochemical system renders RBC as conve- nient models for investigating anuclear effects as well as bi- ological mechanisms in eukaryotes. Exposure of RBC to bi- ologically relevant doses of UVAl radiation results in h e m o l y s i ~ , ~ ~ ~ ~ ~ as is also the case for biologically and envi- ronmentally irrelevant doses of UVC.z8-3’ Slight hemolysis was previously observed in UVA-irradiated RBC samples in the absence of p h o t o ~ e n s i t i z e r , ~ ~ . ~ ~ but no further investiga- tions of this phenomenon have been reported. Nevertheless, UVAI-induced anuclear damage, which may lead to he- molysis, has been, until recently, basically overlooked. Thus, the mechanism of UVA 1 -induced hemolysis is not currently understood, though photosensitized mechanisms have been investigated at great length.33
1018

UVA 1 -induced hemolysis 1019
We investigated the effects of RBC exposure to U V A l radiation, because a significant portion of this wavelength region can penetrate to the dermal layer of the skid4 and may expose circulating blood. In this paper, we present re- sults on the biological effectiveness of U V A l radiation as a cytoplasmic oxidative agent resulting in cytoskeletal damage and hemolysis of sheep RBC in vitro. We first compared the relative photohemolytic effectiveness of the three wavelength regions of U V R on RBC and then investigated the UVAI- induced hemolytic mechanism. The data reveal that expo- sure of RBC t o biologically relevant doses (equilethal to L5 178Y cells26) of UVA1, but not UVB or UVC, radiation causes oxidation of cytoplasmic components that interact adversely with the cytoskeleton resulting in Heinz body for- mations and consequently hemolysis.
MATERIALS AND METHODS
Radiation sources
UVAI. UVASUN 2000 or 3000 (Mutzhas International AG, Switzerland) sunlamps, emitting wavelengths > 340 nm with emis- sion maximum at about 365 nm were used at a distance of 4 1 2 3 or 70-72 cm, respectively. UVA 1 irradiance at the sample level was approximately 260 W/m2 for both lamps.
UVB. FS20 (Westinghouse) sunlamp, emission peak at 313 nm, was equipped with a U-340 filter (Hoya Optics, Fremont, CA) to reduce UVA, UVC and visible light output. UVB irradiance was approximately 0.48 W/m2 at 29 cm (total irradiance 0.8 1 W/m2).
UVC. G15T8 (General Electric) germicidal lamp was equipped with a 254 nm narrow bandpass filter (Microcoatings, Westford, MA) having a bandwidth of 10 nm and a diameter of 1 cm. UVC irradiance was approximately 0.06 W/m2 at 33 cm.
Visible + inji-ared (IR) components. UVASUN lamps were used for exposures to wavelengths ,400 nm at the distances used for UVAl exposures. To ensure that the visible + IR components of the UVASUN lamp emission spectrum had no effect on hemolysis, the exposures were performed as follows. The samples were covered with a UF3 filter (400 nm cutoff and 90% transmission of radiation 1400 nm; Rohm and Haas, Bristol, PA) to remove the UVA 1 emis- sion and irradiated for 1.1 x the time used for the 1200 kJ/m2 UVAl -irradiated samples, to account for the transmission properties of the filter. This exposure resulted in a total dose of visible + IR equivalent to the amount received by the UVAl-irradiated samples.
The instruments used for routine dosimetric measurements were: for UVAI, a 160B digital multimeter (Keithley, Cleveland, OH) equipped with a long-wave UV 5-22 1 detector (Ultraviolet Products Inc., San Gabriel, CA); and for UVB or UVC, a 40A Optometer equipped with a broadband UV detector (United Detector Tech., Santa Monica, CA).
Detectors were calibrated with an NM-9 DH double-grating spec- troradiometer (EG&G Gamma Scientific, San Diego, CA).
Cells Sheep RBC (Advanced Biotechnologies Inc., Columbia, MD) were
stored in Alsever’s solution at 4°C and diluted in Dulbecco’s phos- phate-buffered saline (PBS) containing Ca2+ and Mg2+ prior to use (Whittaker Bioproducts Inc., Walkersville, MD or Advanced Bio- technologies Inc.) because the transmission through PBS is >99% for all wavelengths between 230 and 800 311n.j~ Sterile conditions were maintained throughout the experiments.
Exposure conditions Irradiation of the RBC samples (5 mL) was done in uncovered 60
mm polystyrene culture dishes (Costar Corp., Cambridge, MA) at room temperature. Sample temperature was checked with a model 46 TUC Tele-thermometer (Yellow Springs Instrument Co. Inc., Yellow Springs, OH): temperature was controlled using a cooling plate connected to a circulating water bath.
In order to compare the hemolytic effectiveness of the different
wavelength regions of UVR to RBC, doses that were equilethal to the survival of L5 178Y murine lymphoma cells (yielding a survival of approximately lo%, as determined by the clonogenicity assay) were used? UVAI, I200 kJ/m2; UVB, 390 J/m2; UVC, 17 J/m2. For the UVAl dose-response relationships, the dose (0-1625 kJ/ m2), observation times (0-78 h) and density of RBC (1.&-1.2 x 106 or 1.2-1.3 x lo7 cells/mL) were varied. When biochemicals were used in the experiments, stock solutions (10 x or more) were prepared in PBS and sterile filtered. PBS was added to the samples that did not receive any biochemicals, so that the final volume and density of the cells were initially equal in all samples.
When the cell density was increased, hemolysis decreased. To determine if release of hemoglobin during exposure by damaged cells might be responsible for the observed decrease in hemolysis, by absorbing UVA 1 radiation, increasing concentrations of twice-crys- tallized sheep hemoglobin were added prior to exposure (Sigma Chemical Co., St. Louis, MO).
To investigate the possibility that oxidative stress might be re- sponsible for hemolysis, approximately 1 O6 RBC/mL were exposed to 1000 kJ/m2 of UVAl radiation in PBS containing the reducing agent glutathione (GSH; Gibco Laboratories, Life Technologies Inc., Grand Island, NY).
Glycolytic biochemicals were added to investigate the role gly- colysis might have in the hemolytic mechanism. Approximately 3 x I O6 RBC/mL were exposed to 1000 kJ/m2 of UVA 1 radiation in PBS containing either 0.33 mM sodium pyruvate (PYR), 0.33 mM lactic acid or 5.6 mM D-glucose (Sigma Chemical Co.). In order to determine whether the oxidative stress responsible for hemolysis was internal or external, the hemolytic accelerator and glycolytic metabolite PYR was added in combination with the antioxidant GSH (10 mM).
For absorption scans of the RBC lysates immediately following exposure, 5 mL samples of 0.7-1.2 x 10’ RBC/mL were either sham-exposed or exposed to 1000 kJ/m2 of UVAl radiation in PBS.
The lipid peroxidation determinations were performed immedi- ately following exposure: two 5 mL samples of approximately lo7 RBC/mL were sham-exposed or exposed to 500, 1000 or 1500 kJ/ m2 of UVAl radiation in the absence or presence of PYR (0.33 mM).
The loss of water due to evaporation during UVASUN 2000 ex- posures were corrected mathematically. The UVASUN 3000 lamp did not cause any appreciable difference in evaporation between the exposed and sham-exposed samples during the exposure times em- ployed in this study (< 75 min).
Postexposure treatment and observations
To assure good recovery (>go%) of the cells following exposure, the samples were diluted 1:1 with medium, either FIOP,j6 i.e. Fi- scher’s medium for leukemic cells of mice (Gibco Laboratories) with 0.12 mM pluronic F68 (BASF Wyandotte Corp., Parsippany, NJ), or RPMI-1640 with L-glutamine and 25 mM N-[2-hydroxyethyl] piperazine-N’-[2-ethane sulfonic acid] (HEPES; Gibco Laborato- ries), supplemented with 10°/o serum, either equine (Hyclone Lab- oratories Inc., Logan, UT), or fetal bovine (Advanced Biotechnol- ogies Inc.). After addition of medium, the cells were incubated at 37°C (5% C02, 95Yo air and 95% humidity) for 30 min to release the cells from the dishes. The cells were then removed from the dishes by gentle pipetting to minimize membrane damage. These samples were placed in T-25 flasks (Coming Glass Works, Corning, NY) and incubated at 37”C, as described above.
Absorption scans (200-700 nm) were done by collecting the cells (without addition of medium) immediately following exposure, cen- trifuging at 200-250 g at room temperature for 10 min and lysing the cell pellet with 1 mL of high-performance liquid chromatography (HPLC) grade water (J. T. Baker Inc., Phillipsburg, NJ). These lysates were scanned with a model DU-7 Beckman spectrophotometer (Beckman Instruments Inc., Columbia, MD). The background was established by prescanning HPLC grade water from 200 to 700 nm.
Lipid peroxidation was determined using a slightly modified thio- barbituric acid (TBA) procedure” after the appropriate samples were pooled, centrifuged at 225 g, aspirated and resuspended in 0.25 mL PBS. Briefly, 0.25 mL samples were acidified with 0.5 mL of 10% (wthol) trichloroacetic acid and reacted with 1.0 mL of 0.67% (wt/ vol) TBA at 100°C for 15 min in lightly stoppered 5 mL borosilicate
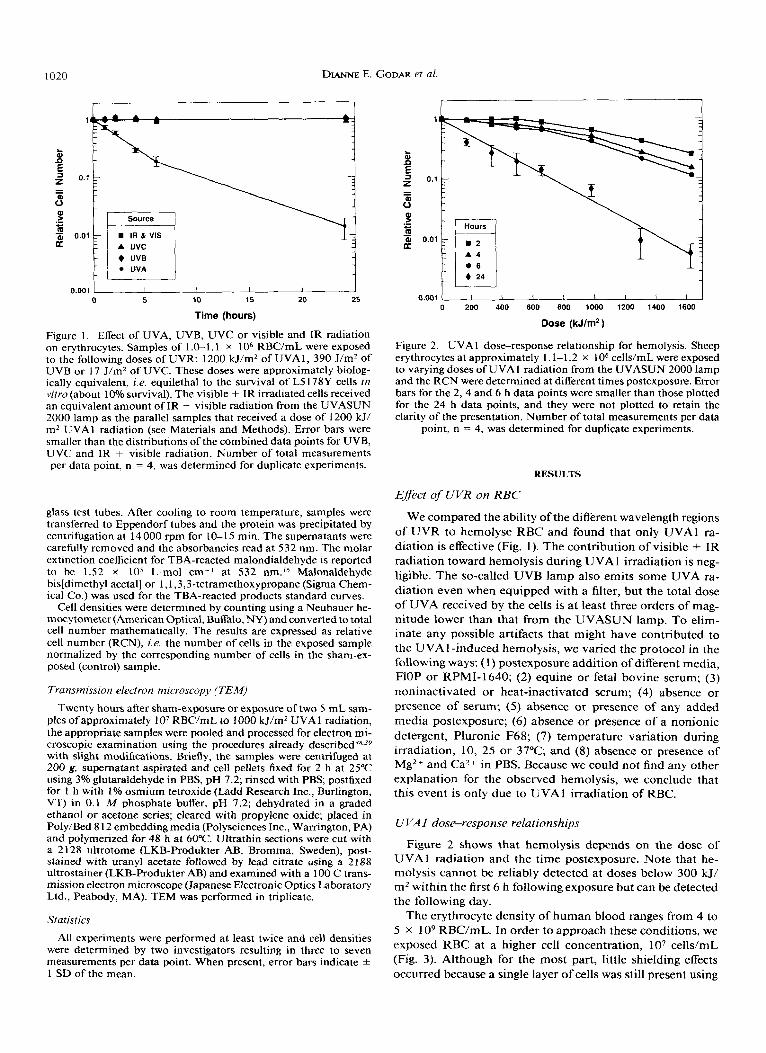
1020 DIANNE E . GODAR et al.
r - I
L \ i
0.1 =-
0.01 =- Source
IR 8 VIS
UVA
0.001 I I I I 1 I 0 5 10 15 20 25
Time [hours)
Figure 1. Effect of UVA, UVB, UVC or visible and IR radiation on erythrocytes. Samples of 1.0-1.1 x lo6 RBC/mL were exposed to the following doses of UVR: 1200 kJ/m2 of UVA 1, 390 J/m2 of UVB or 17 J/m2 of UVC. These doses were approximately biolog- ically equivalent, i.e. equilethal to the survival of L5178Y cells in vitro (about 10% survival). The visible + IR irradiated cells received an equivalent amount of IR + visible radiation from the UVASUN 2000 lamp as the parallel samples that received a dose of 1200 W/ m2 UVAl radiation (see Materials and Methods). Error bars were smaller than the distributions of the combined data points for UVB, UVC and IR + visible radiation. Number of total measurements per data point, n = 4, was determined for duplicate experiments.
glass test tubes. After cooling to room temperature, samples were transferred to Eppendorf tubes and the protein was precipitated by centrifugation at 14 000 rpm for 10-1 5 min. The supernatants were carefully removed and the absorbancies read at 532 nm. The molar extinction coefficient for TBA-reacted malondialdehyde is reported to be 1.52 x los L.mol cm-' at 532 nrn.l6 Malonaldehyde bis[dimethyl acetal] or 1,1,3,3-tetramethoxypropane (Sigma Chem- ical Co.) was used for the TBA-reacted products standard curves.
Cell densities were determined by counting using a Neubauer he- mocytometer (American Optical, Buffalo, NY) and converted to total cell number mathematically. The results are expressed as relative cell number (RCN), i.e. the number of cells in the exposed sample normalized by the corresponding number of cells in the sham-ex- posed (control) sample.
Transmission electron microscopy (TEM)
Twenty hours after sham-exposure or exposure of two 5 mL sam- ples of approximately lo7 RBC/mL to 1000 H/m2 UVAl radiation, the appropriate samples were pooled and processed for electron mi- croscopic examination using the procedures already d e ~ c r i b e d ~ ~ . ~ ~ with slight modifications. Briefly, the samples were centrifuged at 200 g, supernatant aspirated and cell pellets fixed for 2 h at 25'C using 3% glutaraldehyde in PBS, pH 7.2; rinsed with PBS; postfixed for 1 h with 1% osmium tetroxide (Ladd Research Inc., Burlington, VT) in 0.1 A4 phosphate buffer, pH 7.2; dehydrated in a graded ethanol or acetone series; cleared with propylene oxide; placed in Poly/Bed 8 12 embedding media (Polysciences Inc., Warrington, PA) and polymerized for 48 h at 60°C. Ultrathin sections were cut with a 2 128 ultrotome (LKB-Produkter AB, Bromma, Sweden), post- stained with uranyl acetate followed by lead citrate using a 2188 ultrostainer (LKB-Produkter AB) and examined with a 100 C trans- mission electron microscope (Japanese Electronic Optics Laboratory Ltd., Peabody, MA). TEM was performed in triplicate.
Statistics
All experiments were performed at least twice and cell densities were determined by two investigators resulting in three to seven measurements per data point. When present, error bars indicate k 1 SD of the mean.
4 0.001
0 200 400 600 800 1000 1200 1400 1600
Dose (kJ/m2)
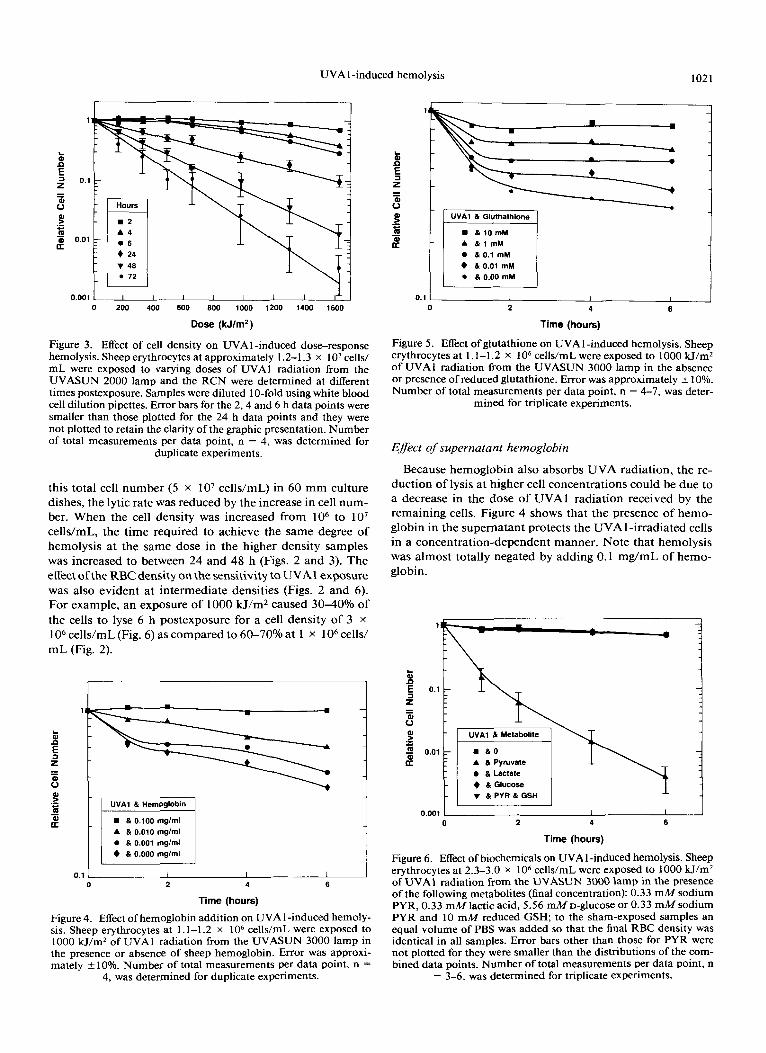
Figure 2. UVA 1 dose-response relationship for hemolysis. Sheep erythrocytes at approximately 1.1-1.2 x lo6 cells/mL were exposed to varying doses of UVA 1 radiation from the UVASUN 2000 lamp and the RCN were determined at different times postexposure. Error bars for the 2, 4 and 6 h data points were smaller than those plotted for the 24 h data points, and they were not plotted to retain the clarity of the presentation. Number of total measurements per data
point, n = 4, was determined for duplicate experiments.
RESULTS
Effect of UVR on RBC
We compared the ability of the different wavelength regions of U V R t o hemolyse RBC and found that only UVAl ra- diation is effective (Fig. 1). T h e contribution of visible + I R radiation toward hemolysis during UVA 1 irradiation is neg- ligible. T h e so-called U V B lamp also emits some UVA ra- diation even when equipped with a filter, but the total dose of UVA received by the cells is a t least three orders of mag- nitude lower than that from the UVASUN lamp. To elim- inate any possible artifacts that might have contributed t o the UVA 1 -induced hemolysis, we varied the protocol in the following ways: (1) postexposure addition of different media, HOP or RPMI-1640; (2) equine or fetal bovine serum; (3) noninactivated or heat-inactivated serum; (4) absence or presence of serum; (5) absence or presence of any added media postexposure; (6) absence or presence of a nonionic detergent, Pluronic F68; (7) temperature variation during irradiation, 10, 25 or 37°C; and (8) absence or presence of Mg2+ and Ca2+ in PBS. Because we could not find any other explanation for the observed hemolysis, we conclude that this event is only due to UVAl irradiation of RBC.
U VAI dose-response relationships
Figure 2 shows that hemolysis depends o n the dose of U V A l radiation and the time postexposure. Note that he- molysis cannot be reliably detected at doses below 300 kJ/ m2 within the first 6 h following exposure but can be detected the following day.
The erythrocyte density of human blood ranges from 4 to 5 x 1 O9 RBC/mL. In order t o approach these conditions, we exposed RBC at a higher cell concentration, lo7 cells/mL (Fig. 3). Although for the most part, little shielding effects occurred because a single layer of cells was still present using
UVA 1 -induce d hemolysis 1021
ii n f z
0.001 I I I I I I I 0 200 400 600 800 1000 1200 1400 1600
Dose (kJ/m2)
Figure 3. Effect of cell density on UVAl -induced dose-response hemolysis. Sheep erythrocytes at approximately 1.2-1.3 x lo7 cells/ mL were exposed to varying doses of UVAl radiation from the UVASUN 2000 lamp and the RCN were determined at different times postexposure. Samples were diluted 10-fold using white blood cell dilution pipettes. Error bars for the 2,4 and 6 h data points were smaller than those plotted for the 24 h data points and they were not plotted to retain the clarity of the graphic presentation. Number of total measurements per data point, n = 4, was determined for
duplicate experiments.
this total cell number (5 x lo7 cells/mL) in 60 mrn culture dishes, the lytic rate was reduced by the increase in cell num- ber. When the cell density was increased from lo6 to lo7 cells/mL, the time required to achieve the same degree of hemolysis at the same dose in the higher density samples was increased to between 24 and 48 h (Figs. 2 and 3). The effect ofthe RBC density on the sensitivity to UVAl exposure was also evident at intermediate densities (Figs. 2 and 6). For example, an exposure of 1000 kJ/m2 caused 30-40% of the cells to lyse 6 h postexposure for a cell density of 3 x 1 O6 cells/mL (Fig. 6) as compared to 60-70°/o at 1 x 1 O6 cells/ mL (Fig. 2).
1
A & 0.010 mglml 0 & 0.001 mglml + & 0.000 mglml
~ ~~
0 2 4 6
Time (hours)
Figure 4. Effect of hemoglobin addition on UVAl -induced hemoly- sis. Sheep erythrocytes at 1.1-1.2 x lo6 cells/mL were exposed to 1000 kJ/m2 of UVAl radiation from the UVASUN 3000 lamp in the presence or absence of sheep hemoglobin. Error was approxi- mately k 10%. Number of total measurements per data point, n =
4, was determined for duplicate experiments.
8 n f z
1
w - I UVA1 & Gluthathione 1
; Z y F y I , , + & 0.01 mM a & 0.00 mM
0.1 0 2 4 6
Time (hours)
Figure 5. Effect of glutathione on UVAI-induced hemolysis. Sheep erythrocytes at 1.1-1.2 x lo6 cells/mL were exposed to 1000 W/mZ of UVAl radiation from the UVASUN 3000 lamp in the absence or presence of reduced glutathione. Error was approximately k 10%. Number of total measurements per data point, n = 4-7, was deter-
mined for triplicate experiments.
Efect of supernatant hemoglobin
Because hemoglobin also absorbs UVA radiation, the re- duction of lysis at higher cell concentrations could be due to a decrease in the dose of UVAl radiation received by the remaining cells. Figure 4 shows that the presence of hemo- globin in the supernatant protects the UVA 1 -irradiated cells in a concentration-dependent manner. Note that hemolysis was almost totally negated by adding 0.1 mg/mL of hemo- globin.
-
UVAl & Metabolite
a A & Pyruvate 0 (I Lactate 4 &Glucose T &PVR&GSH
0.001 1 1 I I 0 2 4 6
Time (hours)
Figure 6. Effect of biochemicals on UVAI-induced hemolysis. Sheep erythrocytes at 2.3-3.0 x lo6 cells/mL were exposed to 1000 kJ/m2 of UVAl radiation from the UVASUN 3000 lamp in the presence of the following metabolites (final concentration): 0.33 mM sodium PYR, 0.33 mM lactic acid, 5.56 mM D-glucose or 0.33 mM sodium PYR and 10 mM reduced GSH; to the sham-exposed samples an equal volume of PBS was added so that the final RBC density was identical in all samples. Error bars other than those for PYR were not plotted for they were smaller than the distributions of the com- bined data points. Number of total measurements per data point, n
= 3-6, was determined for triplicate experiments.
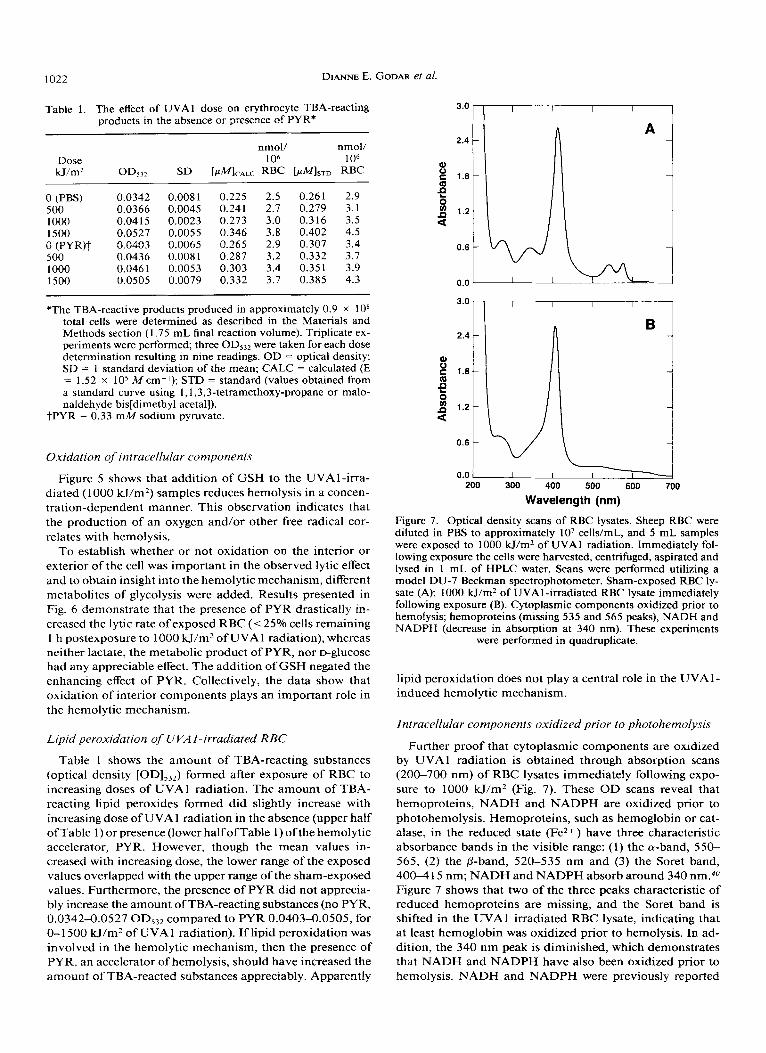
DIANNE E. GODAR et a/ . 1022
2.4
Q 0
m c 1.8
c 1 1.2 U
0.6
0.0
Table 1. The effect of UVA 1 dose on erythrocyte TBA-reacting products in the absence or presence of PYR*
-
-
-
-
-
0 (PBS) 500 1000 1500
500 I000 1500
0 (PYR)t
0 B -
-
-
-
I I I I -
0.0342 0.0366 0.04 15 0.0527 0.0403 0.0436 0.046 1 0.0505
0.008 1 0.0045 0.0023 0.0055 0.0065 0.008 1 0.0053 0.0079
0.225 0.24 1 0.273 0.346 0.265 0.287 0.303 0.332
2.5 2.7 3.0 3.8 2.9 3.2 3.4 3.7
0.26 1 0.279 0.316 0.402 0.307 0.332 0.35 1 0.385
2.9 3.1 3.5 4.5 3.4 3.7 3.9 4.3
*The TBA-reactive products produced in approximately 0.9 x lo8 total cells were determined as described in the Materials and Methods section (1.75 mL final reaction volume). Triplicate ex- periments were performed; three OD53Z were taken for each dose determination resulting in nine readings. OD = optical density; SD = 1 standard deviation of the mean; CALC = calculated (E = 1.52 x lo5 M cm-I); STD = standard (values obtained from a standard curve using 1,1,3,3-tetramethoxy-propane or malo- naldehyde bis[dimethyl acetal]).
tPYR = 0.33 mM sodium pyruvate.
Oxidation of intracellular components
Figure 5 shows that addition of GSH to the UVAl-irra- diated (1 000 kl/m2) samples reduces hemolysis in a concen- tration-dependent manner. This observation indicates that the production of an oxygen and/or other free radical cor- relates with hemolysis.
To establish whether or not oxidation on the interior or exterior of the cell was important in the observed lytic effect and to obtain insight into the hemolytic mechanism, different metabolites of glycolysis were added. Results presented in Fig. 6 demonstrate that the presence of PYR drastically in- creased the lytic rate of exposed RBC ((25% cells remaining I h postexposure to 1000 W/m2 of UVAl radiation), whereas neither lactate, the metabolic product of PYR, nor D-glucose had any appreciable effect. The addition of GSH negated the enhancing effect of PYR. Collectively, the data show that oxidation of interior components plays an important role in the hemolytic mechanism.
Lipid peroxidation of UVAl-irradiated RBC
Table 1 shows the amount of TBA-reacting substances (optical density [OD],,,) formed after exposure of RBC to increasing doses of UVAl radiation. The amount of TBA- reacting lipid peroxides formed did slightly increase with increasing dose of UVA 1 radiation in the absence (upper half ofTable 1) or presence (lower half ofTable 1) ofthe hemolytic accelerator, PYR. However, though the mean values in- creased with increasing dose, the lower range of the exposed values overlapped with the upper range of the sham-exposed values. Furthermore, the presence of PYR did not apprecia- bly increase the amount of TBA-reacting substances (no PYR, 0.0342-0.0527 OD,32 compared to PYR 0.0403-0.0505, for 0-1 500 kJ/m2 of UVAl radiation). If lipid peroxidation was involved in the hemolytic mechanism, then the presence of PYR, an accelerator of hemolysis, should have increased the amount of TBA-reacted substances aDDreCiabh. ADDarentlY
3.0
2.4
Q 0
m E 1.8
c 1 1.2 U
0.6
0.0
Wavelength (nm)
Figure 7. Optical density scans of RBC lysates. Sheep RBC were diluted in PBS to approximately lo7 cells/mL, and 5 mL samples were exposed to 1000 H/m2 of UVA 1 radiation. Immediately fol- lowing exposure the cells were harvested, centrifuged, aspirated and lysed in 1 mL of HPLC water. Scans were performed utilizing a model DU-7 Beckman spectrophotometer. Sham-exposed RBC ly- sate (A); 1000 H/m2 of UVAI-irradiated RBC lysate immediately following exposure (B). Cytoplasmic components oxidized prior to hemolysis; hemoproteins (missing 535 and 565 peaks), NADH and NADPH (decrease in absorption at 340 nm). These experiments
were performed in quadruplicate.
lipid peroxidation does not play a central role in the UVA 1 - induced hemolytic mechanism.
Intracellular components oxidized prior to pholohemolysis
Further proof that cytoplasmic components are oxidized by UVAl radiation is obtained through absorption scans (200-700 nm) of RBC lysates immediately following expo- sure to 1000 H / m 2 (Fig, 7). These O D scans reveal that hemoproteins, NADH and NADPH are oxidized prior to photohemolysis. Hemoproteins, such as hemoglobin or cat- alase, in the reduced state (Fez+ ) have three characteristic absorbance bands in the visible range: (1) the a-band, 550- 565, (2) the @-band, 520-535 nm and (3) the Soret band, 400-4 15 nm; NADH and NADPH absorb around 340 nm.#O Figure 7 shows that two of the three peaks characteristic of reduced hemoproteins are missing, and the Soret band is shifted in the UVAl irradiated RBC lysate, indicating that at least hemoglobin was oxidized prior to hemolysis. In ad- dition, the 340 nm peak is diminished, which demonstrates that NADH and NADPH have also been oxidized prior to hemolvsis. NADH and NADPH were Dreviouslv reDorted
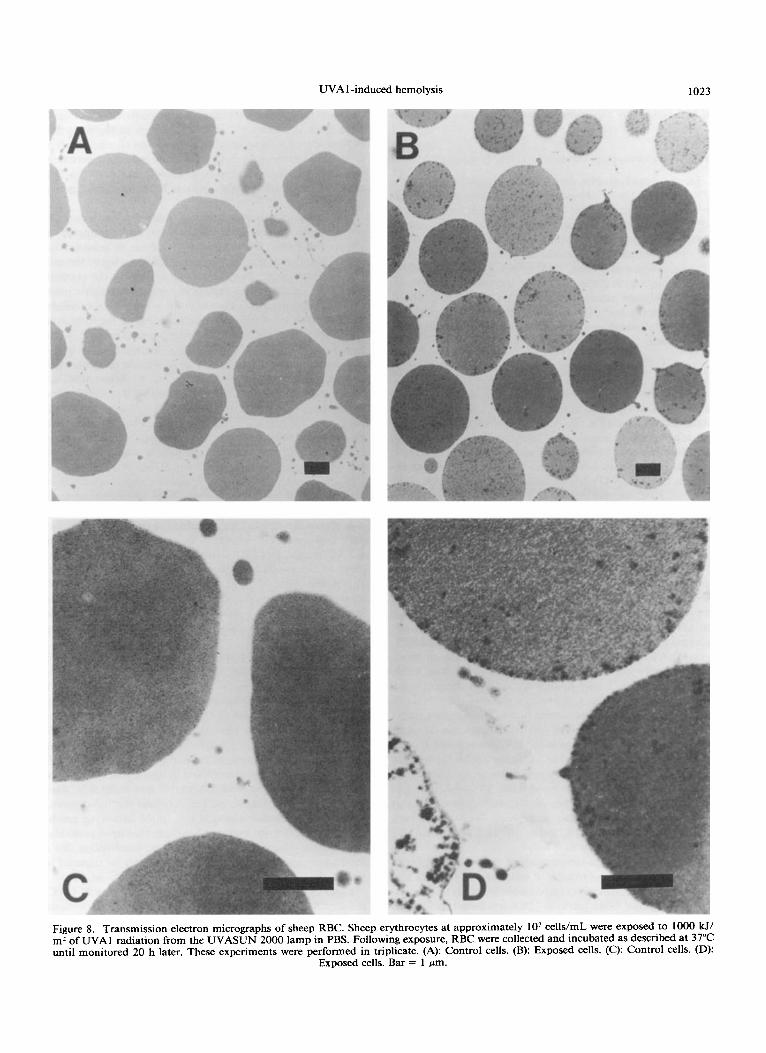
UVA 1 -induced hemolysis 1023
Figure 8. Transmission electron micrographs of sheep RBC. Sheep erythrocytes at approximately lo7 celldrnl were exposed to 1000 W, m2 of UVAl radiation from the UVASUN 2000 lamp in PBS. Following exposure, RBC were collected and incubated as described at 37°C until monitored 20 h later. These experiments were performed in triplicate. (A): Control cells. (B): Exposed cells. (C): Control cells. (D)
Exnosed cells. Bar = 1 vm.

1024 DIANNE E. GODAR et a[.
to be oxidized by exposure of these coenzymes in aqueous solution to UVA
Heinz body formations induced by UVAI irradiation of RBC
Electron micrographs demonstrate Heinz bodies in the UVA 1 exposed cells 20 h postexposure to a dose of 1000 W/ m2 (Fig. 8). The fact that Heinz bodies can be formed in RBC through oxidation and denaturation of intracellular components, particularly hemoglobin and certain glycolytic enzymes, treated with chemical oxidants such as phenylhy- drazine, is well e~tablished."~'~ We have also observed Heinz body formations in phenylhydrazine-treated human RBC us- ing transmission electron microscopy (unpublished obser- vations). Heinz body formations give further evidence that cytoplasmic oxidative stress is the pivotal reason for pho- tohemolysis.
DISCUSSION
The results show that when biologically equivalent doses are utilized, UVA 1, but not UVB or UVC, radiation causes hemolysis in vitro (Fig. 1). A variety of experiments ruled out other possible causes of hemolysis (results not shown, see Materials and Methods). Therefore, the observed lytic effect is due only to UVAl irradiation of RBC. Slight he- molysis induced by UVA irradiation (sources contained some UVB) of RBC was previously n ~ t e d . ~ ~ , ~ ~ The previously re- ported mechanistic studies of Cook28-31 on UVC-induced hemolysis used extremely high doses (>200 J/m*). These doses are not biologically relevant; 17 J/m2 reduced the sur- vival of L5 178Y-S cells (the UV-resistant strain) to about 6%. In addition, doses above 200 J/mZ are not suited for hemolytic comparison between the three wavelength ranges of UVR because they are at least two orders of magnitude too low for UVA, and are an order of magnitude too high for UVC, radiation. But when the criteria of biologically relevant doses are met, based on survival of nucleated cells, only UVAl radiation is found to be hemolytically effective.
We observe UVAl -induced hemolysis to be dose, time, cell-density (Figs. 2 and 3) and hemoglobin-concentration dependent (Fig. 4). The reduction in the amount of lysis by the higher number of cells present during irradiation appears to be due to the increased release of hemoglobin. Because hemoglobin absorbs photons in the UVA wavelength region, the reduced lytic rate observed at higher cell densities may be primarily attributable to the proportionate increase in the release of hemoglobin by the lysed RBC. The dose, time and cell-density dependence of hemolysis could explain why this phenomenon went previously undetected. We can barely de- tect hemolysis below 300 kJ/mz at lo7 or lo6 RBC/mL for postexposure times prior to 6 h.
The fact that GSH, a reducing agent that does not absorb UVA1, can protect the irradiated cells (Fig. 5) suggests that a free radical mechanism is responsible for the lytic process. Our finding that GSH protects UVA-irradiated anucleated cells agrees with previous reports that GSH plays a major role in protecting nucleated cells from inactivation by singlet oxygen species generated by UVA Free rad- ical reactions have been implicated in UV-mediated cuta- neous damage, which is thought to be associated with lipid
peroxide formation.46 Mechanisms for peroxidation of lipids have been disc~ssed.~' GSH is a known endogenous free radical scavenger and antioxidant and can also be involved in the reduction of hydrogen peroxide and lipid peroxides through the enzymatic action of GSH peroxidase. So, it is possible that GSH actually protects RBC from lipid perox- idation.
To determine whether interior or exterior oxidation was important in the hemolytic mechanism, we investigated the effect of different metabolites of glycolysis in combination with GSH. One might conclude that GSH protected the cells (Fig, 5) from exterior oxidation, but increasing the extracel- Mar GSH concentration results in a somewhat higher intra- cellular concentration. To maintain a normal intracellular level of GSH, the RBC is partly dependent on the hydrolysis of extracellular GSH by the membrane-bound enzyme, y-glu- tamyl transpeptidase.q8 Experiments with metabolites of gly- colysis demonstrate that oxidation of interior components is important in the lytic mechanism (Fig. 6). When RBC are irradiated with UVAl in the presence of PYR, extensive and rapid hemolysis occurs, but when GSH was also present the effect of PYR was completely negated. These observations show that oxidation of cytoplasmic components rather than exterior membrane components play a major role in the UVAl -induced hemolytic mechanism, because PYR is a cy- toplasmic metabolite of glycolysis. The apparent enhanced hemolytic effect of PYR may have been indirectly due to its enzymatic product, lactate. However, no enhanced hemolysis was observed when lactate was added to the supernatant, nor with D-glucose (Fig. 6). Only the presence of PYR during UVAl irradiation of RBC produced the observed enhance- ment of hemolysis. We have previously suggested a mech- anism involving PYR in UVA 1 -induced h e m o l y s i ~ . ~ ~ Al- though interior or cytoplasmic oxidation was found to be involved in the UVA I -induced hemolytic mechanism, a role for lipid peroxidation could not be entirely ruled out based on these findings.
Because the role of lipid peroxidation in UVAl-induced hemolysis was uncertain, the formation of lipid peroxides after UVAl irradiation of RBC was determined. Table 1 shows the amount of lipid peroxides formed after exposing RBC to increasing doses of UVAl radiation in the absence or presence ofPYR. We have already demonstrated that PYR accelerates hemolysis, so it follows that more TBA-reacting substances should have been formed in the presence of PYR if lipid peroxidation was involved in hemolysis. Because the nmol of TBA-reacted products per 1 O6 remaining RBC does not appreciably increase, we can conclude that lipid perox- idation does not play a central role in the hemolytic mech- anism. However, we may not conclude that lipid peroxida- tion has no role at all in the hemolytic mechanism, for a minimum amount of peroxidized lipids may be required to render the plasma membrane structurally unstable.
Lipid peroxidation was thought to be the cause of UVC- induced hemolysis, because both hemolysis and the amount of peroxidized lipids correlated with increasing dose of UVC radiation.l6 However, these independent observations do not constitute proof of a mechanism. Regardless, studies such as these are not really suitable for making comparisons between the different wavelength regions' ability to cause Dhotohemo-

UVA 1 -induced hemolysis 1025
lysis and photoperoxidation of lipids, because they were based on equivalent doses (0-24 kJ/m2) rather than on equilethally based doses. Note that for UVC and UVB radiations this dose range is well beyond biological relevance. Our results are in agreement with Punnonen et all9 in that their data also showed no significant increase in TBA reactive products with increasing UVA dose, though they did find an increase in conjugated dienes. Careful analysis of the data from Ma- tuso et al l8 shows that lipid peroxidation is probably not involved in the UVA-hemolytic mechanism, for they did not obtain any oxidized lipids in the absence of photosensitizer using doses up to 2500 H/m2 of UVA radiation, and we have shown that hemolysis occurs at lower doses. However, lipid peroxidation may still play some role in the UVA 1 -induced hemolytic mechanism, though our data strongly suggest that it is not the primary cause of hemolysis.
Our results demonstrate that exposure of about lo7 RBC/ mL to UVAl radiation results in hemoglobin, NADH and NADPH oxidation as demonstrated by the absorption spec- tral scans (200-700 nm) of RBC lysates immediately follow- ing exposure to UVAl radiation (Fig. 7). Oxidation of these intracellular components is revealed by the decrease or dis- appearance of absorption peaks at 340 (NADH and NADPH), 535 and 565 nm p he mop rote in^).^^ Other researchers have reported that NADH and NADPH are involved in oxygen- dependent UVA-initiated photosensitization reactions2’: chemical characterization shows that photooxidation of NADH and NADPH occurs even in the absence of oxygen.20 Collectively, the data indicate that intracellular oxidation plays a significant role in the UVA 1 -induced hemolytic mechanism.
Further support for intracellular oxidation as the cause of UVA 1 -induced hemolysis is found by Heinz body forma- tions near the plasma membranes of irradiated RBC (Fig. 8) . Heinz bodies are known to form through oxidation of intra- cellular components, such as hemoglobin, GSH and certain glycolytic enzymes. Heinz bodies are RBC inclusions com- prised of oxidizedldenatured hemoglobins (hemichromes) and glycolytic enzymes that are irreversibly bound to the cyto- ~keleton.~’ It is thought that Heinz bodies may cause he- molysis by resulting in an ionic imbalance through band 3, the transmembrane anion transporter protein, since the cy- toskeleton is attached, in part, to the membrane through band 3.42 In addition, band 3 has been implicated in the light- photosensitized hemolytic m e c h a n i ~ m . ~ ~ . ’ ~ Our data strongly suggest that intracellular oxidation of GSH, hemoglobin and glycolytic enzymes occurs in UVA 1 -irradiated cells resulting in cytoskeletal damage as demonstrated by the formation of Heinz bodies and subsequent hemolysis. Using D,o survival values (1 0% remaining target population) to compare the relevant biological effects of UVA, UVB and UVC radiations on the cytoskeletal components of human white cells, Za- mansky and C ~ O U ~ ~ observed that UVA radiation caused damage to the microtubules, whereas UVA did not cause disruption of the intermediate filaments.” Thus, cytoskeletal damage may occur as a result of UVA irradiation to a variety of cell types (red and white); however, the mechanism and type of damage incurred may vary. RBC do not have mi- crotubules or intermediate filaments, but do have UVA l - induced membrane-associated cytoskeletal damage.
SUMMARY
Our data show that (1) only UVAl radiation can induce hemolysis when biologically equivalent doses of the three UV wavelength regions are employed; (2) UVAl -induced hemolysis is dose, cell density and time dependent; (3) hemolysis is also dependent on the concentration of free hemoglobin in the supernatant; (4) a free radical process correlates with hemolysis; (5) lipid peroxidation does not strongly correlate with hemolysis; (6) intracellular oxidation of hemoglobin, NADH, NADPH and probably GSH is caused by UVAl irradiation of RBC and results in (7) cytoskeletal damage, as shown by (8) Heinz body formations, which even- tually result in (9) hemolysis.
Acknowledgements-The authors thank Drs. Mays L. Swicord and Janusz Z. Beer for helpful scientific discussions, Harry Bushar for statistical consultation and Larry W. Cress for reviewing this manu- script. We also thank Jerome I. Levine for superior literature searches and Amparo G. Strickland for some technical assistance. This re- search was conducted in part while one of the authors (D.E.G.) held a National Research Council Research Associateship at the Center for Devices and Radiological Health, Food and Drug Administra- tion. The National Research Council Research Associateship pro- gram for the Food and Drug Administration is under contract no. (FDA) 223-88-6076.
REFERENCES
1. Passchier, W. F. and B. F. M. Bosnjakovic (eds.) (1987) Hu- man Exposure to Ultraviolet Radiation. Excerpta Medica, Arn- sterdam.
2. Peak, J. G. and M. J. Peak (1 99 1) comparison of initial yields of DNA-to-protein crosslinks and single-strand breaks induced in cultured human cells by far- and near-ultraviolet light, blue light and X-rays. Mutat. Res. 246, 187-191.
3. Hollaender, A. (1 943) Effect of long ultraviolet and short vis- ible radiation (35004900 A) on Escherichia coli. J. Bacteriol. 46, 531-541.
4. Tyrrell, R. M., R. B. Webb and R. D. Ley (1974) Induction of single-strand breaks (alkali-labile bonds) in bacterial and phage DNA by near UV (365 nm) radiation. Photochem. Photobiol.
5. Peak, M. J. and J. G. Peak (1 982) Single-strand breaks induced in Bacillus subtilus DNA by ultraviolet light: action spectrum and properties. Photochem. Photobiol. 35, 675-680.
6. Tyrrell, R. M. (1973) Induction of pyrimidine dimers in bac- terial DNA by 365 nm radiation. Photochem. Photobiol. 17,69- 73.
7. Peak, M. J., J. G. Peak, M. P. Moehringand R. B. Webb (1984) Ultraviolet action spectra for DNA dimer induction, lethality and mutagenesis in Escherichia coli with emphasis on the UVB region. Photochem. Photobiol. 40, 6 13-620.
8. Chamberlain, J. and S. H. Moss (1987) Lipid peroxidation and other membrane damage produced in Escherichia coli K1060 by near-UV radiation and deuterium oxide. Photochem. Pho- tobiol. 45, 625-630.
9. Jagger, J. (1981) Near-UV radiation effects on microorgan- isms. Photochem. Photobiol. 34, 76 1-768.
10. Jagger, J. (1985) Solar-UV Action on Living Cells. Praeger, New York.
1. Jagger, J. (1972) Growth delay and photoprotection induced by near-ultraviolet light. In Research Progress in Organic, Bi- ological and Medicinal Chemistry, Vol. 3 (Edited by U. Gallo and L. Santamaria), Part 1, pp. 383401. Elsevier, New York.
2. Ramabhadran, T. V. (1975) Effects of near-ultraviolet radia- tions (3 13405 nm) on DNA, RNA, and protein synthesis in E. coli B/r: implications for growth delay. Photochem. Photobiol.
3. Ramabhadran, T. V. and J. Jagger (1976) In vivo induction
20, 395-398.
22, 117-123.

DIANNE E. GODAR et a1 1026
of 4-thiouridine-cytidine adducts in t-RNA of E. coli B/r by near-ultraviolet radiation. Photochem. Photobiol. 23, 3 15-32 1.
14. Webb, R. B. (1977) Lethal and mutagenic effects of near- ultraviolet radiation. In Photochemistry and Photobiology Re- views, Vol. 2 (Edited by K.C. Smith), pp. 169-262. Plenum, New York.
15. Tyrrell, R. M. and S. M. Keyse (1990) The interaction ofUVA radiation with cultured cells. J. Photochem. Phofobiol. 4B, 349- 361.
16. Roshchupkin, D. I., A. B. Pelenitsyn, A. Ya. Potapenko, V. V. Talitsky and Yu. A. Vladimirov ( I 975) Study of the effects of ultraviolet light on biomembranes-IV. The effect of oxygen and lipid photoperoxidation in rat erythrocytes and liposomes. Photochem. Photobiol. 21, 63-69.
17. Hayakawa, K. and I. Matsuo (1989) Alfoqualone phototox- icity-photosensitized lipid peroxidative potency and its biolog- ical significance. Tokai J . Exp. Clin. Med. 14, 15-24.
18. Matsuo, I., H. Fujita, K. Hayakawa and M. Ohkido (1986) Lipid peroxidative potency of photosensitized thiazide diuretics. J. Invest. Dermatol. 87, 637-641.
19. Punnonen, K., A. Puntala and M. Ahotupa (1991) Effects of ultraviolet A and B irradiation on lipid peroxidation and activity of the antioxidant enzymes in keratinocytes in culture. Photo- derm. Photoimmunol. Photomed. 8, 3-6.
20. Czochralska, B., W. Kamczynski, G. Bartosz and D. Shugar (1984) Oxidation of excited-state NADH and NAD dimer in aqueous medium involvement of 02- as a mediator in the pres- ence of oxygen. Biochim. Biophys. Acta 801, 403-409.
21. Cunningham, M. L., J. S. Johnson, S. M. Giovanazzi and M. J. Peak (1 985) Photosensitized production of superoxide anion by monochromatic ( 2 9 e 4 0 5 nm) ultraviolet irradiation of NADH and NADPH coenzymes. Photochem. Photobiol. 42, 125-128.
22. McCormick, J. P., J. R. Fischer and J. P. Pachlatko (1976) Characterization of a cell-lethal product from the photooxida- tion of tryptophan: hydrogen peroxide. Science 191, 468-469.
23. Zamansky, G. B. and I.-N. Chou (1987) Environmental wave- lengths of ultraviolet light induce cytoskeletal damage. J. In- vest. Dermatol. 89, 603-606.
24. Wennersten, G. (1 977) Photodynamic aspects of some metal complexes. Acta Dermato-venereol (Stockholm) 57, 5 19-524.
25. Thomas, D. P. and D. E. Godar (1 99 1) Ultrastructural changes in murine lymphoma cells exposed to ultraviolet-A radiation. In Proceedings, Forty-ninth Annual Meeting, Electron Micros- copy Society ofAmerica (Edited by G. W. Bailey and E. L. Hall), pp. 104-105. San Francisco Press, San Francisco.
26. Godar, D. E. and J. Z. Beer (1992) UVAl-induced anuclear damage in mammalian cells. In Biological Responses to Ultra- violet A Radiation (Edited by F. Urbach), pp. 65-73. Valdenmar Publishing Co., Overland Park, KS.
27. Ciodar, D. E. and M. L. Swicord (1992) Studies on the rnech- anisrn of UVA I -induced hemolysis. In Biologic Effects ofLight (Edited by M. F. Hollick and A. Kligman), pp. 349-353. Walter de Gruyter, Berlin.
28. Cook, J. S. (1956) Some characteristics ofhemolysis by ultra- violet light. J. Cell. Comp. Physiol. 47, 55-84.
29. Cook, J. S. (1 96 1) Hemolysis by ultraviolet radiation. In Prog- ress in Photobiology (Edited by B. C. Christensen and B. Buch- man), pp. 453-457. Elsevier, Amsterdam.
30. Cook, J. S. (1 965) The quantitative interrelationships between ion fluxes, cell swelling and radiation dose in ultraviolet he- molysis. J. Gen. Physiol. 48, 719-734.
31. Cook, J. S. (1974) Photopathology of the erythrocyte mem- brane. In Pathological Aspects of Cell Membranes (Edited by B.
F. Trump and A. Arstila), pp. 199-213. Academic Press, New York.
32. Meffert, V. H., D. Metz and N. Sonnichhsen (1980) Stabili- sierung der Erythrozytenmembran gegenuber UVA-Bestrahlung durch 8-MOP. Dermatol. Monatsschr. 166, 160-1 6 1.
33. Girotti, A. W. (1983) Mechanisms ofphotosensitization. Pho- tochem. Photobiol. 38, 745-75 1.
34. Jacobson, E. D., L. E. Bockstahler, R. P. Felten, K. B. Hellman, V. M. Hitchins, A. Lamanna, J. I. Levine, C. D. Lytle, S. M. Sykes and M. Waxler (1987) Safety measures in optical ra- diation treatment. In Photomedicine, Vol. 111 (Edited by E. Ben- Hur and I. Rosenthal), pp. 158-1 59. CRC Press, Boca Raton.
35. Jacobson, E. D., K. Krell and M. J. Dempsey (1981) The wavelength dependence of ultraviolet light-induced cell killing and mutagenesis in L5 178Y mouse lymphoma cells. Photochem.
36. Clive, D. and J. F. S. Spector (1975) Laboratory procedure for assessing specific locus mutations at the TK locus in cultured L5178Y mouse lymphoma cells. Mutat. Res. 31, 17-29.
37. Stacey, N. H., L. R. Cantilena and C. D. Klaassen (1980) Cad- mium toxicity and lipid peroxidation in isolated rat hepatocytes. Toxicol. Appl. Pharmacol. 53, 470-480.
38. Hayat, M. A. (1972) Basic Electron Microscopy Techniques. Van Nostrand Reinhold, New York.
39. Hayat, M. A. (1986) Basic Techniques for Transmission Elec- tron Microscopy. Academic Press, San Diego.
40. Scott, T. and M. Eagleson (eds.) (1988) Concise Encyclopedia Biochemistry. Walter de Gruyter, New York.
41. Jandl, J. H. (1987) Blood. Little, Brown, Boston. 42. Low, P. S. (1989) Interaction of native and denatured he-
moglobins with band 3: consequences for erythrocyte structure and function. In Red Blood Cell Membranes: Structure, Func- tion, Clinical Implications, Vol. 1 I (Edited by P. Agre and J. C. Parker), pp. 237-260. Marcel Dekker, New York.
43. Tyrrell, R. M. and M. Pidoux (1986) Endogenous glutathione protects human skin fibroblasts against the cytotoxic action of UVB, UVA and near-visible radiations. Photochem. Photobiol.
44. Tyrrell, R. M. and M. Pidoux (1988) Correlation between endogenous glutathione content and sensitivity of cultured hu- man skin cells to radiation at defined wavelengths in the solar ultraviolet range. Photochem. Photobiol. 47, 405-4 12.
45. Tyrrell, R. M. and M. Pidoux (1989) Singlet oxygen involve- ment in the inactivation of cultured human fibroblasts by UVA (334 nm, 365 nm) and near-visible (405 nm) radiations. Pho- tochem. Photobiol. 49, 407-4 12.
46. Black, H. S. (1987) Potential involvement of free radical re- actions in ultraviolet light mediated cutaneous damage. Pho- tochem. Photobiol. 46, 2 13-23 1.
47. Girotti, A. W. (1985) Mechanisms of lipid peroxidation. J. Free Rad. Biol. Med. 1 , 87-95.
48. Laperche, Y., G. Guellaen, R. Barouki and J. Hanoune (1990) Biosynthesis and regulation of gamma-glutamyl transpeptidase. In Glutathione: Metabolism and Physiological Functions (Edited by J. Vina), pp. 80-8 1. CRC Press, Boca Raton.
49. Pooler, J. P. (1986) A new hypothesis for the target in pho- tohemolysis: dimers of band 3 protein. Photochem. Photobiol.
50. Pooler, J. P. and A. W. Girotti (1986) Photohemolysis of human erythrocytes labeled in band 3 with eosin-isothiocyanate. Photochem. Photobiol. 44, 495-499.
5 1. Zamansky, G. B. and I.-N. Chou (1 990) Disruption of keratin intermediate filaments by ultraviolet radiation in cultured hu- man keratinocytes. Photochem. Photobiol. 52, 903-906.
Photobiol. 33, 257-260.
44, 561-564.
43, 263-266.




















