
Lipoteichoic acid from Lactobacillus plantarum elicits both the production of Interleukin23p19 and...
10
Lipoteichoic acid from Lactobacillus plantarum elicits both the production of Interleukin-23p19 and suppression of pathogen-mediated Interleukin-10 inTHP-1cells Han Geun Kim 1 , Min Geun Gim 1 , Joo Yun Kim 1 , Hyun Jin Hwang 2 , Min Seok Ham 2 , Jung Min Lee 1 , Thomas Hartung 3 , Jung Woo Park 4 , Seung Hyun Han 5 & Dae Kyun Chung 1,2 1 School of Biotechnology and Institute of Life Science and Resources, Kyung Hee University, Suwon, Korea; 2 RNA Inc., #308 College of Industry, Kyung Hee University, Suwon, Korea; 3 EU Joint Research Centre, European Centre for the Validation of Alternative Methods (ECVAM), Ispra, Italy; 4 Department of Biological Sciences, University of Ulsan, Ulsan, Korea; and 5 Humoral Immunology Section International Vaccine Institute, SNU Research Park, Seoul, Korea Correspondence: Dae Kyun Chung, School of Biotechnology and Institute of Life Science and Resources, Kyung Hee University, Suwon, 449-701, Korea. Tel.: 182 31 201 2465; fax: 182 31 202 8333; e-mail: [email protected] Received 26 January 2006; revised 08 September 2006; accepted 09 October 2006. First published online 21 December 2006. DOI:10.1111/j.1574-695X.2006.00175.x Editor: Artur Ulmer Keywords Lactobacillus plantarum ; Lipoteichoic acid; Interleukin-23; Interleukin-10. Abstract In this study, the stimulatory effects of different lactic acid bacteria strains, and their subcellular fractions, on the THP-1 cell line were evaluated. Lactobacillus plantarum was found in particular to induce high levels of IL-23p19 mRNA, but it moderately induced TNF-a production. IL-10 production was not entirely affected by L. plantarum stimulation. When subcellular fractions of L. plantarum were used to treat THP-1 cells, IL-23p19 mRNA expression was enhanced in a dose- responsive manner, specifically by lipoteichoic acid (LTA). The cotreatment of THP-1 cells by both L. plantarum and Staphylococcus aureus LTA resulted in decreased IL-10 production when compared with cells treated by S. aureus LTA alone. Taken together, these data suggest that LTA isolated from L. plantarum elicits stimulatory effects upon the expression of IL-23p19 and inhibitory effects on pathogen-mediated IL-10 production. Introduction Lactic acid bacteria (LAB) are known for their health- promoting effects such as the nonspecific enhancement of the immune system, protection against intestinal infection, the reduction of serum cholesterol level, and their antiox- idant properties (Meydani & Ha, 2000; Von der Weid et al., 2001; Matsuguchi et al., 2003). Several strains of LAB also have effects upon the production of cytokines, such as IL-12, IL-10, TNF-a, TGF-b, IL-8 and RANTES, and upon cell proliferation in human intestinal epithelial cells (Hessle et al., 1999; Kato & Yokokura, 1999; Wallace et al., 2003). Interleukin (IL)-23 is a heterodimeric cytokine formed by complexes of p19 (Oppmann et al., 2000) and a subunit of IL-12, p40 (Trinchieri, 2003). IL-23 has biological functions that are similar to IL-12, such as interferon-g production from CD41 T cells, antitumor effects and antimetastatic activity. Some previous reports also suggest that IL-23 induces a more robust and sustained cytotoxic T lympho- cyte and Th1 immune response than IL-12 (Belladonna et al., 2002; Lo et al., 2003; Trinchieri, 2003; Ha et al., 2004). Furthermore, IL-23N220L, an N-glycosylation mutant of this protein that shows reduced expression of excess p40 but has no inhibitory effects upon IL-23 levels, has been proposed as an effective adjuvant of a DNA vaccine for antigen-specific T cell immunity (Ha et al., 2004). However, our current understanding of the biology of IL-23 is still considerably less than IL-12, which to-date has been studied more extensively. The human cytokine IL-10 is secreted from macrophages by stimulation of several endogenous and exogenous factors such as endotoxin, TNF-a, catecholamines and cAMP-ele- vating drugs (Meisel et al., 1996; Platzer et al., 2000; Riese et al., 2000; Asadullah et al., 2003). The principal immuno- logical role of IL-10 is the regulation of the Th1/Th2 balance and this molecule promotes the development of the Th2 cytokine pattern by inhibiting IFN-gamma production of T lymphocytes, via the suppression of IL-12 synthesis in accessory cells (Asadullah et al., 2003). Moreover, IL-10 represses both proinflamatory cytokine production and the antigen-presenting capacity of macrophages and dendritic cells (De Waal Malefyt et al., 1991). IL-10 overexpression is FEMS Immunol Med Microbiol 49 (2007) 205–214 c 2006 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Lipoteichoic acid from Lactobacillus plantarum elicits both the production of Interleukin23p19 and...

Lipoteichoic acid fromLactobacillusplantarum elicits boththeproductionof Interleukin-23p19and suppressionofpathogen-mediated Interleukin-10 inTHP-1cellsHan Geun Kim1, Min Geun Gim1, Joo Yun Kim1, Hyun Jin Hwang2, Min Seok Ham2, Jung Min Lee1,Thomas Hartung3, Jung Woo Park4, Seung Hyun Han5 & Dae Kyun Chung1,2
1School of Biotechnology and Institute of Life Science and Resources, Kyung Hee University, Suwon, Korea; 2RNA Inc., #308 College of Industry, Kyung
Hee University, Suwon, Korea; 3EU Joint Research Centre, European Centre for the Validation of Alternative Methods (ECVAM), Ispra, Italy;4Department of Biological Sciences, University of Ulsan, Ulsan, Korea; and 5Humoral Immunology Section International Vaccine Institute,
SNU Research Park, Seoul, Korea
Correspondence: Dae Kyun Chung, School
of Biotechnology and Institute of Life Science
and Resources, Kyung Hee University, Suwon,
449-701, Korea. Tel.: 182 31 201 2465; fax:
182 31 202 8333;
e-mail: [email protected]
Received 26 January 2006; revised 08
September 2006; accepted 09 October 2006.
First published online 21 December 2006.
DOI:10.1111/j.1574-695X.2006.00175.x
Editor: Artur Ulmer
Keywords
Lactobacillus plantarum ; Lipoteichoic acid;
Interleukin-23; Interleukin-10.
Abstract
In this study, the stimulatory effects of different lactic acid bacteria strains, and
their subcellular fractions, on the THP-1 cell line were evaluated. Lactobacillus
plantarum was found in particular to induce high levels of IL-23p19 mRNA, but it
moderately induced TNF-a production. IL-10 production was not entirely affected
by L. plantarum stimulation. When subcellular fractions of L. plantarum were
used to treat THP-1 cells, IL-23p19 mRNA expression was enhanced in a dose-
responsive manner, specifically by lipoteichoic acid (LTA). The cotreatment of
THP-1 cells by both L. plantarum and Staphylococcus aureus LTA resulted in
decreased IL-10 production when compared with cells treated by S. aureus LTA
alone. Taken together, these data suggest that LTA isolated from L. plantarum
elicits stimulatory effects upon the expression of IL-23p19 and inhibitory effects
on pathogen-mediated IL-10 production.
Introduction
Lactic acid bacteria (LAB) are known for their health-
promoting effects such as the nonspecific enhancement of
the immune system, protection against intestinal infection,
the reduction of serum cholesterol level, and their antiox-
idant properties (Meydani & Ha, 2000; Von der Weid et al.,
2001; Matsuguchi et al., 2003). Several strains of LAB also
have effects upon the production of cytokines, such as IL-12,
IL-10, TNF-a, TGF-b, IL-8 and RANTES, and upon cell
proliferation in human intestinal epithelial cells (Hessle
et al., 1999; Kato & Yokokura, 1999; Wallace et al., 2003).
Interleukin (IL)-23 is a heterodimeric cytokine formed by
complexes of p19 (Oppmann et al., 2000) and a subunit of
IL-12, p40 (Trinchieri, 2003). IL-23 has biological functions
that are similar to IL-12, such as interferon-g production
from CD41 T cells, antitumor effects and antimetastatic
activity. Some previous reports also suggest that IL-23
induces a more robust and sustained cytotoxic T lympho-
cyte and Th1 immune response than IL-12 (Belladonna
et al., 2002; Lo et al., 2003; Trinchieri, 2003; Ha et al., 2004).
Furthermore, IL-23N220L, an N-glycosylation mutant of
this protein that shows reduced expression of excess p40 but
has no inhibitory effects upon IL-23 levels, has been
proposed as an effective adjuvant of a DNA vaccine for
antigen-specific T cell immunity (Ha et al., 2004). However,
our current understanding of the biology of IL-23 is still
considerably less than IL-12, which to-date has been studied
more extensively.
The human cytokine IL-10 is secreted from macrophages
by stimulation of several endogenous and exogenous factors
such as endotoxin, TNF-a, catecholamines and cAMP-ele-
vating drugs (Meisel et al., 1996; Platzer et al., 2000; Riese
et al., 2000; Asadullah et al., 2003). The principal immuno-
logical role of IL-10 is the regulation of the Th1/Th2 balance
and this molecule promotes the development of the Th2
cytokine pattern by inhibiting IFN-gamma production of T
lymphocytes, via the suppression of IL-12 synthesis in
accessory cells (Asadullah et al., 2003). Moreover, IL-10
represses both proinflamatory cytokine production and the
antigen-presenting capacity of macrophages and dendritic
cells (De Waal Malefyt et al., 1991). IL-10 overexpression is
FEMS Immunol Med Microbiol 49 (2007) 205–214 c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved

also considered to be responsible for immune deviation
towards Th2, which is a hallmark of atopic disorders such as
atopic dermatitis and allergic asthma (Asadullah et al., 2003).
The toll-like receptors (TLRs) are a family of vertebrate
receptors that are homologous to the drosophila toll recep-
tors (Dabbagh & Lewis, 2003). The associated signaling
pathways that are initialized by TLRs, from the recognition
of conserved products of microorganisms, cause maturation
of antigen-presenting cells (APC), which then generate pro-
inflammatory cytokines such as IL-12, IL-23 and IL-27, and
promote Th1 cell proliferation (Dabbagh & Lewis, 2003;
Trinchieri, 2003; Ha et al., 2004).
In our current study, we examined the effects of LTA,
which is one of subfractions from Lactobacillus plantarum,
on the production of IL-23p19 and IL-10 mRNAs. Our
results indicate that L. plantarum LTA has different induc-
tion and suppression effects upon a number of cytokine
productions when compared with LTA isolated from the
pathogenic bacterium Staphylococcus aureus.
Materials and methods
Cell line and reagents
The human monocyte-like cell line THP-1 was obtained
from the Korean Cell Line Bank (KCLB, Korea) and main-
tained in RPMI 1640 with 10% FBS (JBI, Korea) at 37 1C in
5% CO2. The THP-1 cells were placed in 12-well plates, and
they were stimulated with heatkilled bacteria or subfractions
from L. plantarum for 4 h or 24 h. Mouse monoclonal anti-
human TLR2 (TL2.1; IgG2a) and mouse monoclonal anti-
human TLR4 (HTA 125; IgG2a) antibodies were purchased
from Santa Cruz Biotechnology Inc., and mouse monoclo-
nal anti-human CD14 (clone #134620; IgG1) was purchased
from R&D Systems. Anti-human TLR1 (GD2.F4; IgG1), and
all isotype-matched control antibodies (IgG1 and IgG2a)
were purchased from eBioscience.
Bacterial strains
The bacterial strains used for the stimulation of THP-1 cells
were Lactobacillus bulgaricus (KCTC 3188), Lactobacillus
casei ssp. casei (KCTC 3109), Lactococcus lactis ssp. cremoris
(ML 4), Enterococcus faecalis (KCTC 3195), Lactobacillus
helveticus (KCTC 3545), Leuconostoc mesenteroides (KCTC
3100), Staphylococcus aureus (KCTC 1621), L. plantarum
(KCTC 10887BP) and Escherichia coli DH5a. L. bulgaricus,
L. casei, L. helveticus, Leu. mesenteroides and L. plantarum
were cultured with MRS broth at 37 1C for 18 h, and
Leuconostoc cremoris and Enterococcus faecalis were cultured
with M17 broth containing 5% glucose at 30 1C for 18 h.
Staphylococcus aureus was amplified with BHI broth
containing 0.5% glucose and 0.5% cystein at 37 1C for 24 h,
and Escherichia coli DH5a was cultured with LB broth
at 37 1C for 18 h.
Preparation of subcellular fractions
Bacterial subcellular fractions were prepared by the method
of Roman Dziarski et al. with minor modifications (Ro-
senthal & Dziarski, 1994). Briefly, bacteria were harvested in
exponential growth phase at an OD (600 nm) of 0.6 and
quickly chilled in an ice-ethanol bath until the temperature
dropped to 4 1C. Pellets were suspended in saline, and the
suspension was boiled at 100 1C for 20 min. Heatkilled
bacterial cells were suspended with ice cold water and mixed
with 0.1 mm glass beads until total disruption of cells was
achieved. Unbroken cells were sedimented by centrifugation
(1500 g, 4 1C) for 10 min, and then the supernatant was
prepared as crude extract. To collect cell walls from the
supernatant, the crude extract was centrifuged for 30 min at
6500 g at 4 1C followed by washing with water three times.
The precipitated cell walls were incubated with RNase
(100 mg mL�1) and DNase (50mg mL�1) for 18 h, and then
trypsin (200mg mL�1) was added and incubated for another
18 h at 37 1C. The cell walls were sedimented by centrifuga-
tion at 6500 g and 4 1C for 30 min followed by washing with
water four times, and the purified cell walls were lyophilized.
One gram of cell walls was incubated with 5% trichloroace-
tic acid (TCA) for 18 h at 22 1C, and sedimented by
centrifugation at 6500 g for 30 min. Teichoic acid (TA) was
recovered from the supernatants by TCA extraction with
ethyl ether. At the same time, the sediment from TCA
extraction was washed three times with water and three
times with acetone, and dried to make peptidoglycan
powder. Genomic DNA was isolated from L. plantarum with
a genomic DNA prep kit (Promega corporation), and then
purified by phenol extraction. Briefly, prepared genomic
DNA was incubated with 100mg mL�1 RNase for 1 h at 37 1C
water bath, and then 100mg mL�1 of proteinase K was added
and incubated for another 1 h in 50 1C water bath. DNA
samples were extracted with equal volumes of (25 : 24 : 1)
phenol:chloroform:isoamyl alcohol. Extracted DNA was
then precipitated with 0.75 volume of room-temperature
isopropanol for 10 min followed by washing with 70%
ethanol. After resuspension of the pellet with distilled
endotoxin-free water, the removal of contamination was
confirmed by agarose gel electrophoresis. Highly purified
LTAs from L. plantarum (pLTA) and S. aureus (aLTA) were
prepared by butanol extraction followed by purification
using octyl-sepharose and DEAE–sepharose chromatogra-
phy as described previously (Morath et al., 2001).
RNA preparation and real-time PCR
Total RNA was extracted from THP-1 cells with a guanidium
thiocyanate-acid phenol-chloroform method. cDNA was
FEMS Immunol Med Microbiol 49 (2007) 205–214c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
206 H.G. Kim et al.

synthesized with Improm-IITM reverse transcription system
(Promega corporation) according to the manufacturer’s
instructions. To quantify the cytokine mRNA, real-time
PCR amplification was carried out using the ABI prism
7000 sequence detection system (Applied BioSystems, USA),
and the PCR products were detected using SYBR Green. The
primers used for cytokine detection were as follows: IL-23
p19 forward, 50-GCTGAACAGAGAGAATCAGG-30; IL-
23p19 reverse, 50-CTTTTTGCCT-TCTTCTCAAA-30; IL-10
forward, 50-ATGTTCTTTGGGGAGCCAA-30; IL-10 reverse,
50-GTAGCAGTTAGGAAGCCCAA-30; TNF-a forward, 50-
CTCTTCTGGCTCAAAAAGA-GAATT-30; TNF-a reverse,
50-AGGCCCCAGTTTGAATTCTT-30. Cytokine expressions
were normalized using glyceraldehydes-3-phospate dehy-
drogenase (GAPDH).
Establishment of dominant-negative MyD88transfectants and Transient transfection
Dominant negative (dn) MyD88 was manufactured by the
method of Masuo Hosokawa et al. with minor modifications
(Wang et al., 2002). cDNA for dnMyD88 (amino acids
156–296) that contains toll/interleukin-1 receptor domain
was amplified from mRNA obtained from THP-1 cells by
reverse transcription polymerase chain reaction (RT-PCR)
and cloned into pCMV-Tag 2A (Stratagene, USA). THP-1
cells were transfected with the expression vector with
WelFec-QTM transfection reagent (JBI, Korea) according to
the manufacturer’s instructions. At 36 h after the transfec-
tion, cells were used for further analysis. PCR primers used
for RT-PCR were as follows: dnMyD88 forward, 50-GCTAG-
CATGCCTGAGCGTTTCG-30; dnMyD88 reverse, 50-
GGATCCTCAGGGCAGGGACAA-30.
ELISA assay
After stimulation, cell supernatants were removed and
assayed for cytokine production by standard sandwich
ELISA. The TNF-a ELISA was performed using monoclonal
mouse IgG1, clone #28401 for capture and the biotinylated
human TNF-a specific polyclonal goat IgG followed by
streptavidin HRP for detection. ELISA was developed using
o-phenylenediamine as a substrate, and OD was determined
at a wavelength of 450 nm using a 550-nm reference
wavelength. All values were interpolated from either a log-
log or a four-parameter fit of a curve generated from
appropriate standards.
Statistical analysis
All experiments were performed at least three times. The
data shown are representative results as the means� the SD
of triplicate experiments. An unpaired t test was used to
determine the significance between two groups of the data.
Results
The preparations of major subcellular fractions
When studies are performed with gram-positive bacterial
subcellular fractions, it is important to exclude any potential
contaminants, especially endotoxin. This is because human
monocytes are highly responsive to endotoxin. Therefore, we
removed all endotoxin contaminants from prepared L. plan-
tarum subcellular fractions. The LTA preparation was purified
with octyl-sepharose followed by DEAE-sepharose chromato-
graphy, and then a Limulus Amebocyte Lysate (LAL) test and
silver staining were performed to confirm the absence of
endotoxin contamination (Fig. 1a). Prepared peptidoglycan
was repurified with polymyxin B-agarose, and then THP-1
cells were treated with repurified peptidoglycan in the pre-
sence or absence of 500mg mL�1 polymyxin B for 4 h followed
by examination of TNF-a production (Fig. 1b). The genomic
DNA prepared from L. plantarum was treated with phenol
extraction following RNase treatment. Purity of DNA fraction
was confirmed by agarose gel electrophoresis (Fig. 1c), and the
examination of TNF-a production after the stimulation of
THP-1 cells in the presence or absence of polymyxin B (Fig.
1b). The purity of all subcellular fractions from L. plantarum
was tested with a LAL test kit (Cambrex Bio Science, Walkers-
ville). Major fractions such as LTA, peptidoglycan, and DNA
showed high purity, as we expected (Table 1). Especially, LTA
from L. plantarum contains 0.0285 endotoxin unit per mL
(EU mL�1), which means no endotoxin contamination in the
sample. LTA from S. aureus used in this experiment also
showed high purity.
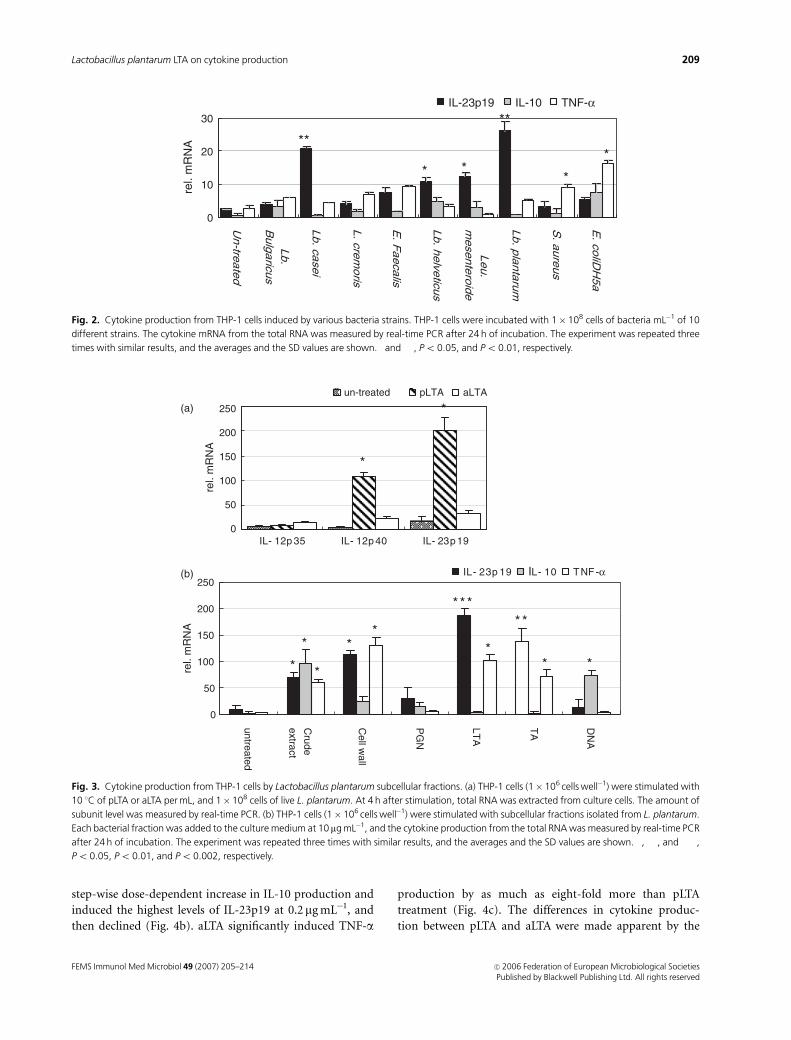
Analysis of cytokine production using variousbacterial strains in THP-1 cells
Nine different heat-killed bacterial strains were examined
for their ability to induce cytokine production in THP-1
cells (Fig. 2). Lactobacillus casei and L. plantarum signifi-
cantly induced IL-23p19 mRNA, whereas Enterococcus fae-
calis, Escherichia coli DH5a and S. aureus induced high level
of TNF-a transcription. Some LAB, such as L. bulgaricus, L.
helveticus and Leu. mesenteroides, as well as Escherichia coli
DH5a, induced IL-10 production in THP-1 cells. In parti-
cular, L. plantarum was a highly potent inducer of IL-23p19
mRNA, but it did not induce IL-10 production in THP-1
cells. Hence, we selected L. plantarum and examined its
ability to induce cytokine production in more detail.
LTA isolated from L. plantarum shows the highestIL-23p19 mRNA induction activity in THP-1 cells
IL-23 is a heterodimeric cytokine composed of p19 and p40,
one of a subunit of IL-12. To examine the inducing activity
of those subunits, we quantified the mRNA levels of the
FEMS Immunol Med Microbiol 49 (2007) 205–214 c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
207Lactobacillus plantarum LTA on cytokine production

IL-12 and IL-23 subunits in THP-1 cells exposed to LTA
isolated from either L. plantarum (pLTA) or S. aureus
(aLTA). Transcript levels of the p35, p40 and p19 subunits
were then measured by real-time PCR. Each of these subunit
mRNA levels were found to be higher in pLTA treated cells
than in aLTA treated cells, and IL-23p19 mRNA production
was observed to be the most highly inducible species when
compared with the other subunits (Fig. 3a). Therefore, we
chose a p19 subunit for the following experiment as well as
TNF-a and IL-10.
In order to determine the components of L. plantarum
that had cytokine-inducing activities, THP-1 cells were
stimulated with subcellular fractions of L. plantarum. Lacto-
bacillus plantarum extracts were separated into six fractions:
crude extract, cell wall, peptidoglycan , lipoteichoic acid
(LTA), teichoic acid (TA) and DNA. Each of these fractions
was then added to the THP-1 culture medium at a concen-
tration of 10 mg mL�1 and the cytokine mRNA levels were
measured by real time PCR after 24 h of incubation.
Although the cell wall, LTA and TA fractions were each
found to significantly increase IL-23p19 mRNA production,
the LTA fraction showed the highest IL-23p19-inducing
activity (Fig. 3b). In contrast, peptidoglycan and DNA
showed only slight induction of IL-23p19 mRNA. IL-10
mRNA was found to be remarkably induced by crude extract
and DNA. From our six L. plantarum subfractions, all except
DNA and peptidoglycan significantly increased TNF-a pro-
duction in THP-1 cells. DNA and peptidoglycan were again
found to be inconsiderable inducers, in this case of TNF-aproduction.
pLTA, highly purified from L. plantarum inducedIL-23p19 and TNF-a production, but not IL-10production
To examine the effects of pLTA on cytokine production in
more detail, THP-1 cells were stimulated with pLTA as
a dose- or time-dependent manner. aLTA isolated from
S. aureus was used as a control to compare the differences
between pLTA and aLTA for cytokine production. Treatment
of THP-1 cells with pLTA resulted in a dose-dependent
induction in IL-23p19 transcription. In contrast, expression
of IL-10 was almost entirely unaffected by pLTA treatment
(Fig. 4a). Compared with pLTA treatment, aLTA displayed a
1 2 3
Lane 1: High molecular weightmarker (Lambda DNA-HindIIIdigested molecular marker).
Lane 2: DNA preparation is both degraded as well as contaminated with RNA.
Lane 3: High molecular weightgenomic NDA freeof RNAcontamination after RNasetreating followed by phenolextraction.
BSA ng well−1
2 20 200 2000Size
marker pLTA aLTA pTA pDNA
untreatment Polymyxin B 0.5 mg/ml
8
7
6
5
4
3
2
1
0LTA TA DNA PGN
TN
F-α
ng/m
l
TN
F-α
ng/m
l
200
150
50
0
100
LPS
(a)
(b)
(c)
Fig. 1. The confirmation of the purity of the major subcellular fractions.
(a) 20mg of pLTA, aLTA, pTA and pDNA were subjected to 12% sodium
dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) followed
by silver staining. Bovine serum albumin (BSA) was used as a protein
standard. (b) THP-1 cells were incubated with 10 mg mL�1 of LTA, TA,
DNA. Peptidoglycan, and 0.5mg mL�1 lipopolysaccharide in the presence
or absence of polymyxin B (0.5 mg mL�1) for 4 h, and the TNF-a produc-
tion was examined by ELISA. (c) Lactobacillus plantarum genomic DNA
was repurified by phenol extraction to remove RNA or protein contam-
ination, and separated by agarose gel electrophoresis.
Table 1. LAL test to confirm the contamination of endotoxin
Subfractions pPGN pCX pCW pTA pDNA pLTA aLTA
OD 405 nm 0.0173 0.272 0.2375 0.2667 0.0249 0.0205 0.027
EU mL�1 0.0216 0.567 0.4932 0.1852 0.0379 0.0285 0.042
LAL, Limulus Amebocyte Lysate; pPGN, peptidoglycan isolated from Lactobacillus plantarum; pCX, crude extract fraction of L. plantarum; pCW, cell wall
fraction of L. plantarum; pTA, teichoic acid isolated from L. plantarum; pDNA, genomic DNA fraction of L. plantarum; pLTA, lipoteichoic acid isolated
from L. plantarum; aLTA, lipoteichoic acid isolated from S. aureus.
FEMS Immunol Med Microbiol 49 (2007) 205–214c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
208 H.G. Kim et al.
step-wise dose-dependent increase in IL-10 production and
induced the highest levels of IL-23p19 at 0.2mg mL�1, and
then declined (Fig. 4b). aLTA significantly induced TNF-a
production by as much as eight-fold more than pLTA
treatment (Fig. 4c). The differences in cytokine produc-
tion between pLTA and aLTA were made apparent by the
0
10
20
30
Un-treated
Lb.B
ulgaricus
Lb. casei
L. cremoris
E. F
aecalis
Lb. helveticus
Leu.m
esenteroide
Lb. plantarum
S. aureus
E. coliD
H5a
rel.
mR
NA
IL-23p19 IL-10 TNF-α
****
*** *
Fig. 2. Cytokine production from THP-1 cells induced by various bacteria strains. THP-1 cells were incubated with 1� 108 cells of bacteria mL�1 of 10
different strains. The cytokine mRNA from the total RNA was measured by real-time PCR after 24 h of incubation. The experiment was repeated three
times with similar results, and the averages and the SD values are shown. �and ��, Po 0.05, and Po 0.01, respectively.
0
50
100
150
200
250
untreated
Crude
extract
Cell w
all
PG
N
LTA
TA
DN
Are
l. m
RN
A
0
50
100
150
200
250
rel.
mR
NA
IL- 23p 19 IL- 10 TNF-α
*
*
*
**
** *
*****
IL- 12p 35 IL- 12p 40 IL- 23p19
un-treated pLTA aLTA
*
*
(b)
(a)
Fig. 3. Cytokine production from THP-1 cells by Lactobacillus plantarum subcellular fractions. (a) THP-1 cells (1� 106 cells well�1) were stimulated with
10 1C of pLTA or aLTA per mL, and 1�108 cells of live L. plantarum. At 4 h after stimulation, total RNA was extracted from culture cells. The amount of
subunit level was measured by real-time PCR. (b) THP-1 cells (1� 106 cells well�1) were stimulated with subcellular fractions isolated from L. plantarum.
Each bacterial fraction was added to the culture medium at 10mg mL�1, and the cytokine production from the total RNA was measured by real-time PCR
after 24 h of incubation. The experiment was repeated three times with similar results, and the averages and the SD values are shown. �, ��, and ���,
Po 0.05, Po 0.01, and Po 0.002, respectively.
FEMS Immunol Med Microbiol 49 (2007) 205–214 c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
209Lactobacillus plantarum LTA on cytokine production

time-course treatment of LTAs. IL-23p19 production was
more responsive to pLTA than to aLTA (Fig. 5a), whereas
IL-10 and TNF-a production were more responsive to aLTA
than to pLTA (Fig. 5b and c). Interestingly, pLTA did not
affect to IL-10 production. These results suggest that pLTA
may induce a Th1-type response.
pLTA inhibited aLTA-induced IL-10 production,but showed a synergistic effect on IL-23p19
To examine the effects of pLTA on the aLTA-mediated
cytokine production, THP-1 cells were stimulated with
pLTA and/or aLTA, and then real-time PCR or ELISA was
performed in order to determine cytokine production.
Compared with 10 mg mL�1 of pLTA or aLTA treatment
alone, IL-23p19 production was induced after cotreatment
with 10 mg mL�1 aLTA and increased dose of pLTA (Fig. 6a).
The cotreatment of 10 mg mL�1 pLTA and increased dose of
aLTA did not affect TNF-a production compared with aLTA
treatment alone (Fig. 6b), but IL-10 production was down-
regulated by pLTA-aLTA cotreatment. In particular, the
pretreatment of 10 mg mL�1 pLTA for 24 h followed by aLTA
restimulation at the indicated concentrations significantly
down-regulated IL-10 production (Po 0.05) compared
with aLTA treatment alone (Fig. 6c). These results suggested
that highly purified LTA from LAB might have the ability to
induce Th1 type response by inhibition of pathogen-
induced Th2-type response associated with atopic disorder.
Regulation of cytokine production in THP-1 cellsstimulated by bacterial cells and LTA from eitherL. plantarum or S. aureus
To examine whether bacterial cells can stimulate the expres-
sion of TLRs, we stimulated THP-1 cells with various
0
50
100
150
200
250
0 0.2 2 5 10
0 0.2 2 5 10
Lb. plantarum LTA µg mL−1
LTA µg mL−1
IL- 23p19 IL- 10
0
20
40
60
80
100
S. aureus
rel.
mR
NA
/GA
PD
Hre
l. m
RN
A/G
AP
DH
IL- 23p19 IL- 10
0
50
100
150
200
250
0 µgmL−10.2 µgmL−12 µgmL−1 5 µgmL−110 µgmL−1
TN
F-α
ng
mL−1
pLTA aLTA
(a)
(b)
(c)
Fig. 4. Cytokine production from THP-1 cells stimulated with pLTA or
aLTA. THP-1 cells were stimulated with pLTA (a) or aLTA (b) as indicated
concentration for 4 h. IL-23p19 and IL-10 mRNA production was
determined by real-time PCR from total RNA. (c) TNF-a production was
measured by ELISA from THP-1 cells stimulated with pLTA or aLTA as
indicated concentration for 4 h. The experiment was repeated three
times with similar results, and the averages and the SD values are shown.
0
50
100
150
200
250
0 h 1 h 4 h 6 h 12 h 24 h
0 h 1 h 4 h 6 h 12 h 24 h
0 h 1 h 4 h 6 h 12 h 24 h
IL-2
3p19
rel
. mR
NA
/GA
PD
H
pLTA 10 aLTA 10
0
50
100
150
200
250
TN
F-
α ng
mL−1
pLTA 10 aLTA 10
0
100
200
300
400
500
600
700
IL-1
0 p
g m
L−1
pLTA 10 aLTA 10
(a)
(b)
(c)
Fig. 5. Time course of cytokine induction by pLTA or aLTA treatment. (a)
THP-1 cells were stimulated with 10 mg mL�1 pLTA or aLTA for indicated
time points. The amount of IL-23p19 mRNA was determined by real-time
PCR from total RNA. TNF-a (b) and IL-10 (c) production from THP-1 cells
stimulated with 10 mg mL�1 of pLTA or aLTA for indicated time points was
examined by ELISA. Data (mean � SD) of three independent experi-
ments are presented.
FEMS Immunol Med Microbiol 49 (2007) 205–214c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
210 H.G. Kim et al.

bacteria and measured the mRNA levels of TLR1 through
TLR10. All bacterial strains used in this experiment induced
TLR2 expression (Fig. 7a); however, there were slight
differences between strains (Fig. 7b). To examine whether
TLRs mediate LAB-induced cytokine production, we per-
formed neutralization experiments with anti-TLRs or anti-
CD14 antibodies. THP-1 cells were incubated with anti-
TLR1, anti-TLR2, anti-TLR4 or anti-CD14 for 30 min
before stimulation of heat-killed L. plantarum for 24 h.
Results demonstrated that preincubation with anti-TLR1,
anti-TLR2 and anti-CD14 antibodies inhibited production
of cytokines, including IL-23p19, TNF-a and IL-10 (Fig. 7c),
indicating that L. plantarum induced cytokine production
through TLR1-TLR2 signaling including CD14.
To investigate if pLTA and aLTA have different require-
ments for TLR stimulation, we examined the need for TLR1,
TLR2, TLR4 and CD14. Treatment with antibodies to TLR1,
TLR2, or CD14 reduced the TNF-a production by THP-1
cells in response to pLTA (Fig. 8a). Similarly, antibodies to
TLR1, TLR2 or CD14 significantly reduced the aLTA-induced
TNF-a production (Fig. 8b). Anti-TLR2 and anti-CD14
inhibited production of IL-23p19 in response to pTLA
treatment, and IL-10 production, was also inhibited by anti-
TLR2 or anti-CD14 in THP-1 cells stimulated by aLTA (Fig.
8c). To show the involvement of the TLR2 signaling pathway
in more detail, dnMyD88-expressing vector was transiently
transfected into THP-1 cells. At 36 h following transfection,
the expression of dnMyD88 was confirmed by Western
blotting, and these cells showed no increase in either IL-
23p19 or IL-10 production in response to pLTA or aLTA,
respectively (Fig. 8d). These results suggested that both types
of LTA share several additional receptor molecules required
for the stimulation of THP-1 cells.
Discussion
LTA is a cell wall component of gram-positive bacteria. LTA
is considered to be analogous to the lipopolysaccharide of
gram-negative bacteria, and shares many of its biochemical
and physiological properties (Ginsburg, 2002). (Morath
et al.(2001) has reported that the pro-inflammatory prop-
erty of LTA is dependent upon its D-alanine content . LTA
from L. plantarum Dlt-mutant containing 1.1% D-Ala sub-
stitutions showed an increased ability to stimulate IL-10 and
a decreased ability to induce IL-12 in comparison to wild-
type LTA (41% of D-Ala substitution). This resulted in the
induction of Th2 type response (Grangette et al., 2005).
These results emphasize the importance of LTA composition
in the proinflammatory or anti-inflammatory properties of
gram-positive bacteria. As shown in Fig. 3a, the peptidogly-
can fraction of L. plantarum only moderately increased the
cytokine production in THP-1 cells, which is consistent with
0
50
100
150
200
250
0 µg mL 0.01 0.1µg mL 1µg mL 10 µg mL
TN
F-a
ng
mL
aLTA onlypLTA 10µg mL +aLTA cotreatmentpLTA10 µg mL (-20 h pretreatment->aLTA 6 h stimulation
0
50
100
150
200
250
300
350
400
0.01 0.1µg mL 1µg mL 10 µg mL
IL-1
0 pg
mL
aLTA only
pLTA10+aLTA cotreatmentpLTA10(-24 h prestimulation)->aLTA 6 h stimulation
*
0 µg mL
µg mL
µg mL
(b)
(a)
(c)
0
400
800
1200
1600
un
-tre
ate
d
aL
TA
10
aL
TA
10
+p
LT
A0
.2
aL
TA
10
+p
LT
A2
aL
TA
10
+p
LT
A5
aL
TA
10
+p
LT
A1
0
aL
TA
10
+p
LT
A2
0
pL
TA
10
rel.
mR
NA
/GA
PD
H IL- 23p19 pf
pf
pfpf
Fig. 6. Synergistic or inhibitory effect of cytokine production in THP-1
cells by pLTA and aLTA cotreatment. THP-1 cells were costimulated with
10 mg mL�1 aLTA and 0.2–20 mg mL�1 pLTA for 4 h, and the amount of IL-
23p19 mRNA was examined by real-time PCR from total RNA The
statistical significances were represented by the comparison between
pLTA treatment only and cotreatment of pLTA and aLTA (a). THP-1 cells
were preincubated with 10mg mL�1 pLTA or medium for 24 h, and
restimulated with 0.01–10 mg mL�1 aLTA for 4 h. THP-1 cells were also
costimulated with 10 mg mL�1 pLTA and 0.01–10 mg mL�1 aLTA for 4 h.
TNF-a (b) and IL-10 (c) production was determined from culture super-
natants by ELISA. The experiment was repeated three times, and data are
represented as means� SDs. �, Po 0.05.
FEMS Immunol Med Microbiol 49 (2007) 205–214 c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
211Lactobacillus plantarum LTA on cytokine production

another recent report showing that peptidoglycan is recog-
nized by a nucleotide-binding oligomerization domain
(NOD) but not TLR2 (Travassos et al., 2004). Peptidoglycan
was recognized by the LRR region of NOD, and the Rip2
adaptor molecule is associated with the CARD domain of
NOD, resulting in the activation of NF-kB (Chin et al., 2002;
Kobayashi et al., 2002; Inohara & Nunez, 2003). Bacterial
unmethylated DNA (CpG ODN) binds to TLR9 and initi-
ates signal transduction cascades in a MyD88–dependent
manner. Recently, CpG ODN was used as vaccine adjuvant
(Ulevitch, 2004). Our experiments using subcellular frac-
tions, including LTA, peptidoglycan, DNA from L. plantar-
um, demonstrate that IL-23p19 and TNF-a were
significantly induced by LTA. Although all subcellular frac-
tions were able induced cytokine production, IL-10 produc-
tion was entirely unaffected by LTA (Fig. 3b). These results
suggest that pLTA has a potent Th1-inducing activity.
IL-23 is a heterodimeric cytokine composed of p19 and
p40, one of subunits of IL-12 (Trinchieri, 2003). Many
reports have characterized the role of the IL-12 subunits,
p35 and p40, but very few studies have investigated p19.
Recent studies have shown that IL-23 and IL-12 have similar
biological functions (Ha et al., 2004), and our experiments
showed that THP-1 cells stimulated with whole cell bacteria
or LTAs induced higher levels of IL-23p19 than other
subunits such as IL-12p35 and IL-12p40 (Fig. 3a). Hence,
0
50
100
150
200
250
Un-
trea
ted
S. c
rem
oris
E. F
aeca
lis
Leu.
m
esen
tero
ide
Lb. p
lant
arum
S. a
ureu
s
TLR
2 m
RN
A/G
AP
DH
0
5
10
15
20
25
30
35
rel.m
RN
A
IL-23p19 TNF-α IL-10
L. plantarum − + + + + + + +
IgG1 Antibody − − αTLR1 αTLR2 αTLR4 αCD14 IgG2a
**
*****
**
*****
***
**
**
GAPDH
TLR 1
TLR 2
TLR 3
TLR 4
TLR 5
TLR 6
TLR 7
TLR 8
TLR 9
TLR 10
(a)
(c)
(b)
Fig. 7. THP-1 cells stimulated with bacteria-induced cytokine production through the TLR2 signaling. THP-1 cells were stimulated with
1� 108 cells mL�1 of bacteria for 24 h, and then cDNAs were synthesized with extracted total RNA. The amount of TLR expression was examined with
reverse transcriptase-polymerase chain reaction (RT-PCR) using specific primers for TLRs (a) and quantitative real-time PCR (b). The primers used in real-
time PCR are: TLR2 forward, 50 ACC CTA GGG GAA ACA TCT CT 30; TLR2 reverse, 50 AGC TCT GTA GAT CTG AAG CAT C 30. (c) THP-1 cells were
pretreated with 10mg mL�1 of anti-TLR1, anti-TLR2, anti-TLR4 and anti-CD14 antibodies for 30 min before 1� 108 cells mL�1 of Lactobacillus plantarum
treatment for 24 h. The level of IL-23p19, TNF-a and IL-10 was determined by real-time PCR. Data are expressed as means� SDs. �� and ���, Po 0.01
and Po 0.001, respectively. Statistical significance between groups was evaluated by ANOVA.
FEMS Immunol Med Microbiol 49 (2007) 205–214c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
212 H.G. Kim et al.

we chose p19 as one of the target cytokines in the present
study in addition to TNF-a, a Th1-inducing proinflamma-
tory cytokine. In addition, we considered the production of
IL-10 because it is a known factor associated with atopic
disorder and an inhibitor for Th1 type response (Asadullah
et al., 2003). In contrast, TNF-a induces Th1 responses, but
the overproduction of TNF-a can induce systemic autoim-
mune disease. Although an appropriate production of IL-10
and TNF-a is essential to the activation of the immune
system, excessive production of these cytokines cause severe
immune diseases. Therefore, the immune regulation with
LAB or their cell wall components is considerable because
they induce moderate cytokine production and inhibit
pathogen-induced cytokine overproduction.
In general, cytokine expression is dependent upon TLR
signaling pathways (Schroder et al., 2003). However, the
mechanism of the association of IL-23p19 production with
TLRs has not previously been reported. In our present study,
LTA from L. plantarum (pLTA) induced less cytokine
production than LTA from S. aureus (aLTA). Compared
with pLTA-induced cytokine production, aLTA-induced IL-
10 and TNF-a production were elevated as much as two-fold
and 9.7-fold, respectively (Figs 4 and 5). Like aLTA, the
signaling transduction following pLTA treatment required
TLR1, TLR2, CD14, and MyD88 recruitment (Fig. 8). These
results suggest that the diversity of cytokine production was
not occurred in signaling pathway. These differences may be
due to structure differences between both LTAs, but we
believe a detailed experiment is necessary to verify this
hypothesis.
In conclusion, L. plantarum seems to be involved in IL-23
mRNA production in THP-1 cells. In particular, pLTA
isolated from L. plantarum was a potent inducer of IL-
23p19 mRNA. In addition, the upregulation of IL-10
production by aLTA is decreased by cotreatment with pLTA.
Both pLTA-mediated IL-23p19 mRNA production and
aLTA-mediated IL-10 production are dependent on TLR1-
TLR2-CD14 and MyD88. Our current findings may indicate
that highly purified LTA from L. plantarum is a good
candidate for therapeutic use as it may reduce IL-10-
mediated atopic dermatitis and may also induce proinflam-
matory cytokines such as IL-23 and TNF-a through the
TLR2 signaling pathway.
(a)
(b)
(c)
(d)
Fig. 8. pLTA- and aLTA-induced cytokine production through TLR1, TLR2
and CD14 dependent signaling. THP-1 cells were treated with anti-TLR1,
anti-TLR2, anti-TLR4, anti-TLR6 or anti-CD14 for 30 min before stimula-
tion with 10mg mL�1 pLTA (a) and 5 mg mL�1 aLTA (b) for 4 h. TNF-aproduction was examined by ELISA. (c) THP-1 cells were pretreated with
anti-TLR2 or anti-CD14 for 30 min, and then stimulated with 10mg mL�1
pLTA or aLTA for 4 h. (d) THP-1 cells were transiently transfected with
dominant negative (dn) MyD88, and stimulated with 10 mg mL�1 pLTA or
aLTA for 4 h. IL-23p19 and IL-10 mRNA production were determined by
real-time PCR from total RNA. Data are expressed as the mean � SDs,
and are representative of three separate experiments. �� and ���,
Po 0.01 and Po 0.001, respectively. Statistical significance between
groups was evaluated by ANOVA.
FEMS Immunol Med Microbiol 49 (2007) 205–214 c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
213Lactobacillus plantarum LTA on cytokine production

Acknowledgements
This work was supported by the Korea Research Foundation
Grant funded by the Korean Government (MOEHRD)
(KRF-2004-041-F00076), Republic of Korea.
References
Asadullah K., Sterry W. & Volk HD. (2003) Interleukin-10
therapy-Review of a new approach. Pharmacol Rev 55:
241–269.
Belladonna ML., Renauld JC. & Puccetti P. (2002) IL-23 and IL-
12 have overlapping, but distinct, effects on murine dendritic
cells. J Immunol 168: 5448–5454.
Chin AI., Dempsey PW., Bruhn K., Miller JF., Xu Y. & Cheng G.
(2002) Involvement of receptor-interacting protein 2 in innate
and adaptive immune response. Nature 416: 190–194.
Dabbagh K. & Lewis DB. (2003) Toll-like receptors and T-helper-
1/T-helper-2 responses, Curr. Opin Infect Dis 16: 199–204.
De Waal Malefyt R., Abrams J., Bennett B., Figdor CG. & De Vries
JE. (1991) Interleukin 10 (IL-10) inhibits cytokine synthesis by
human monocytes: an autoregulatory role of IL-10 produced
by monocytes. J Exp Med 174: 1209–1220.
Ginsburg I. (2002) Role of lipoteichoic acid in infection and
inflammation. Lancet Infect Dis 2: 171–179.
Grangette C., Nutten S., Palumbo E., Morath S., Hermann C.,
Dewulf J., Pot B., Hartung T., Hols P. & Mercenier A. (2005)
Enhanced anti-inflammatory capacity of a Lactobacillus
plantarum mutant synthesizing modified teichoic acids. PNAS
102: 10321–10326.
Ha SJ., Kim DJ., Baek KH., Yun YD. & Sung YC. (2004) IL-23
induces stronger sustained CTL and Th1 immune responses
than IL-12 in Hepatitis C virus envelope protein 2 DNA
immunization. J Immunol 172: 525–531.
Hessle C., Hanson LA. & Wold AE. (1999) Lactobacilli from
human gastrointestinal mucosa are strong stimulators of IL-12
production. Clin Exp Immunol 116: 276–282.
Inohara N. & Nunez G. (2003) Nods: intracellular proteins
involved in inflammation and apoptosis. Nat Rev Immunol 3:
371–382.
Kato IK. & Yokokura T. (1999) Lactic acid bacterium potently
induces the production of interleukin-12 and interferon-
gamma by mouse splenocytes. Int J Immunopharmacol 21:
121–131.
Kobayashi K., Inohara N., Hernandez LD., Galan JE., Nunez G.,
Janeway CA., Medzhitov R. & Flavell RA. (2002) Rick/Rip2/
CARDIAK mediates signalling for receptors of the innate and
adaptive immune systems. Nature 416: 194–199.
Lo CH., Lee SC., Wu PY. et al. (2003) Antitumor and
antimetastatic activity of IL-23. J Immunol 171: 600–607.
Matsuguchi T., Takagi A., Matsuzaki T., Nagaoka M., Ishikawa K.,
Yokokura T. & Yoshikai Y. (2003) Lipoteichoic acids from
Lactobacillus strains elicit strong tumor necrosis factor alpha-
inducing activities in macrophages through Toll-like receptor
2. Clin Diagn Lab Immun 10: 259–266.
Meisel C., Vogt K., Platzer C., Randow F., Liebenthal C. & Volk
HD. (1996) Differential regulation of monocytic tumor
necrosis factor-alpha and interleukin-10 expression. Eur J
Immunol 26: 1580–1586.
Meydani SN. & Ha WK. (2000) Immunologic effects of yogurt.
Am J Clin Nutr 71: 861–872.
Morath S., Geyer A. & Hartung T. (2001) Structure-Function
Relationship of Cytokine Induction by Lipoteichoic Acid from
Staphylococcus aureus. J Exp Med 193: 393–397.
Oppmann B., Lesley R., Blom B., Timans JC., Xu Y., Hunte B.,
Vega F., Yu N., Wang J. & Singh K. (2000) Novel p19 protein
engages IL-12p40 to form a cytokine, IL-23, with biological
activities similar as well as distinct from IL-12. Immunity 13:
715–725.
Platzer C., Docke WD., Volk HD. & Prosch S. (2000)
Catecholamines trigger IL-10 release in acute systemic stress
reaction by direct stimulation of its promoter/enhancer
activity in monocytic cells. J Neuroimmunol 44: 31–38.
Riese U., Brenner S., Docke WD., Prosch S., Reinke P., Oppert M.,
Volk HD. & Platzer C. (2000) Catecholamines induce IL-10
release in patients suffering from acute myocardial infarction
by transactivating its promoter in monocytic but not in T cells.
Mol Cell Biochem 212: 45–50.
Rosenthal RS. & Dziarski R. (1994) Isolation of peptidoglycan
and soluble peptidoglycan fragments. Methods Enzymol 235:
253–285.
Schroder NW., Morath S., Alexander C., Hamann L., Hartung T.,
Zahringer U., Gobel UB., Weber JR. & Schumann RR. (2003)
Lipoteichoic acid (LTA) of Streptococcus pneumoniae and
Staphylococcus aureus activates immune cells via Toll-like
receptor (TLR)-2, Lipopolysaccharide-binding protein (LBP),
and CD14, whereas TLR4 and MD-2 are not involved. J Biol
Chem 278: 15587–15594.
Travassos LH., Girardin SE., Philpott DJ., Blanot D., Nahori MA.,
Werts G. & Boneca IG. (2004) Toll-like receptor 2-dependent
bacterial sensing does not occur via peptidoglycan recognition.
EMBO Rep 5: 1–7.
Trinchieri G. (2003) Interleukin-12 and the regulation of innate
resistance and adaptive immunity. Nat Rev Immunol 3: 133–146.
Ulevitch RJ. (2004) Therapeutics targeting the innate immune
system. Nat Rev Immunol 4: 512–520.
Von der Weid T, Bulliard C & Schiffrin EJ (2001) Induction by a
Lactic acid bacterium of a population of CD41 T cells with
low proliferative capacity that produce transforming growth
factor b and Interleukin-10. Clin Diagn Lab Immun 8:
695–701.
Wallace TD, Bradley S, Buckley ND & Green-Johnson JM (2003)
Interactions of lactic acid bacteria with human intestinal
epithelial cells: effects on cytokine production. J Food Protect
66: 466–472.
Wang J, Kobayashi M, Han M, Choi S, Takano M, Hashino S,
Tanaka J, Kondoh T, Kawamura KI & Hosokawa M (2002)
MyD88 is involved in the signaling pathway for Taxol-induced
apoptosis and TNF-a expression in human myelomonocytic
cells. Br J Haematol 118: 638–645.
FEMS Immunol Med Microbiol 49 (2007) 205–214c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
214 H.G. Kim et al.




















