
Lipase-Catalyzed Acidolysis of Egg-Yolk Phosphatidylcholine ...
Upload
independentCategory
view
0download
0
This article was downloaded by: [Universiti Putra Malaysia]On: 19 March 2012, At: 17:58Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
British Poultry SciencePublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/cbps20
Egg production, faecal pH and microbial population,small intestine morphology, and plasma and yolkcholesterol in laying hens given liquid metabolitesproduced by Lactobacillus plantarum strainsD.W. Choe a , Dr T.C. Loh a b , H.L. Foo c d , M. Hair-Bejo e & Q.S. Awis aa Department of Animal Science, Faculty of Agriculture, University Putra Malaysiab Institute of Tropical Agriculture, University Putra Malaysiac Department of Bioprocess Technology, Faculty of Biotechnology and BiomolecularScience, University Putra Malaysiad Institute of Bioscience, University Putra Malaysiae Department of Pathology and Microbiology, Faculty of Veterinary Medicine, UniversityPutra Malaysia, Selangor, Malaysia
Available online: 12 Mar 2012
To cite this article: D.W. Choe, Dr T.C. Loh, H.L. Foo, M. Hair-Bejo & Q.S. Awis (2012): Egg production, faecal pH andmicrobial population, small intestine morphology, and plasma and yolk cholesterol in laying hens given liquid metabolitesproduced by Lactobacillus plantarum strains, British Poultry Science, 53:1, 106-115
To link to this article: http://dx.doi.org/10.1080/00071668.2012.659653
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form toanyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss, actions,claims, proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly orindirectly in connection with or arising out of the use of this material.

British Poultry Science Volume 53, Number 1 (February 2012), pp. 106—115
Egg production, faecal pH and microbial population, small intestinemorphology, and plasma and yolk cholesterol in laying hens given liquidmetabolites produced by Lactobacillus plantarum strains
D.W. CHOE, T.C. LOH1, H.L. FOO2,3, M. HAIR-BEJO4AND Q.S. AWIS
Department of Animal Science, Faculty of Agriculture, University Putra Malaysia, 1Institute of TropicalAgriculture, University Putra Malaysia, 2Department of Bioprocess Technology, Faculty of Biotechnology andBiomolecular Science, University Putra Malaysia, 3Institute of Bioscience, University Putra Malaysia, and4Department of Pathology and Microbiology, Faculty of Veterinary Medicine, University Putra Malaysia,Selangor, Malaysia
Abstract 1. Various dosages of metabolite combinations of the Lactobacillus plantarum RI11, RG14and RG11 strains (COM456) were used to study the egg production, faecal microflora population, faecalpH, small intestine morphology, and plasma and egg yolk cholesterol in laying hens.2. A total of 500 Lohmann Brown hens were raised from 19 weeks to 31 weeks of age. The birds wererandomly divided into 5 groups and fed on various treatment diets: (i) basal diet withoutsupplementation of metabolites (control); (ii) basal diet supplemented with 0�3% COM456 metabolites;(iii) basal diet supplemented with 0�6% COM456 metabolites; (iv) basal diet supplemented with 0�9%COM456 metabolites; and (v) basal diet supplemented with 1�2% COM456 metabolites.3. The inclusion of 0�6% liquid metabolite combinations, produced from three L. plantarum strains,demonstrated the best effect in improving the hens’ egg production, faecal lactic acid bacteriapopulation, and small intestine villus height, and reducing faecal pH and Enterobacteriaceae population,and plasma and yolk cholesterol concentrations.4. The metabolites from locally isolated L. plantarum are a possible alternative feed additive in poultryproduction.
INTRODUCTION
Over the past two decades, the use of antimicro-bials as growth promoters in the livestock indus-try has become widespread, and this has resultedin common problems such as development ofmultidrug resistant pathogenic bacteria (Sorumand Sunde, 2001). Furthermore, excessive use ofantimicrobials may result in the accumulation ofantimicrobial residues in animal products, whichcan pose a threat to consumers (Mikkelson andJensen, 2000). As a result, many countrieshave started to limit and prohibit the usage ofantimicrobials and opt for naturally derivedfeed additives which exert effects similar to
those of antimicrobials. Currently, the advance-ment of biotechnology has led to thedevelopment of various types of natural feedadditives. These additives include probiotics andprebiotics (Marcel, 2000), enzymes (Parker et al.,2007) and fermented products (Loh et al., 2003,2007).
Lactic acid bacteria (LAB) have been inves-tigated as a source of beneficial microbial feedadditives to replace antimicrobials in the agricul-tural sector. However, there are some limitationsinvolved in the use of probiotics, as severalstringent criteria need to be met before choosingappropriate bacterial strains. These criteriainclude biosafety aspects; ease of microbial
Correspondence to: Dr Loh Teck Chwen, Department of Animal Science, Faculty of Agriculture, University Putra Malaysia, 43300 Serdang, Selangor,
Malaysia. E-mail: [email protected]
Accepted for publication 28th July 2011.
ISSN 0007–1668(print)/ISSN 1466–1799 (online)/12/010106—10 � 2012 British Poultry Science Ltdhttp://dx.doi.org/10.1080/00071668.2012.659653
Dow
nloa
ded
by [
Uni
vers
iti P
utra
Mal
aysi
a] a
t 17:
58 1
9 M
arch
201
2

production, handling and storage; route of pro-biotic administration; stability; and survival of theprobiotic in the host and tolerance for bile(Reque et al., 2003). In addition, a recent studyindicated that even probiotics such as LAB haveacquired resistance to antimicrobials that arecommonly used in the human and animal healthsciences (Shalini and Rameshwar, 2005). Thesituation is further complicated by the fact thatthese acquired resistance traits can be transferredeasily over species and genus borders by con-jugative plasmids and transposons (Wang andLee, 1997). The possible risk of probiotics assupplements for humans and animals should benoted and evaluated, despite their intensive usein recent years. Thus, metabolites produced fromLAB have been suggested as feed additives inanimal feeding (Loh et al., 2009b, 2010; Thanhet al., 2009).
LAB produce metabolites comprisingorganic acids, bacteriocins and ethanol, whichare able to control and inhibit the growth ofintestinal pathogenic bacteria (Foo et al., 2003;Thanh et al., 2010). Compared with live microor-ganisms, LAB metabolites are relatively morestable in terms of handling and storage (Lohet al., 2010) and are less likely to transferantimicrobial resistance traits to other bacteria.Improvements in growth performance, faecallactic acid bacteria and villus height wereobserved when metabolite combinations wereadded to the feed of broilers (Thanh et al., 2009;Loh et al., 2010) and pigs (Thu et al., 2010).Similar effects were also observed in rats whenthe metabolite was added to the drinking water(Foo et al., 2003; Loh et al., 2008). However, thereis limited information regarding the effect ofthese metabolites on the production perfor-mance and egg quality of laying hens. Thus, thepresent study was designed to investigate theeffect of adding combined liquid metabolitesproduced from three L. plantarum strains(RG14, RG11 and RI11) into the diet of layinghens on their egg production, faecal microflorapopulation, faecal pH, small intestine morphol-ogy, and plasma and egg yolk cholesterol.
MATERIALS AND METHODS
Bacteria source and preparation ofmetabolite
Three strains of L. plantarum (RG14, RG11 andRI11) were used in this study. These strains wereobtained from this centre’s own collection(Department of Biotechnology, University PutraMalaysia, Malaysia). Stock cultures of thesestrains were prepared by inoculating L. plantarum(2%, v/v) into MRS broth (Merck) and incubatingat 30�C anaerobically for 24 h before glycerol was
added to a final concentration of 20% (v/v). Thestock cultures were kept at �20�C until required.Before each use, the stock cultures were revivedtwice by transferring into 10 ml MRS broth, andincubated overnight at 30�C anaerobically. Platespreading was then conducted for the revivedcultures, followed by 48 h of incubation. A singlecolony of L. plantarum was picked and inoculatedinto 10 ml MRS broth and incubated anaerobi-cally for 24 h, followed by re-subculturing into10 ml MRS broth and incubating anaerobicallyfor another 24 h. Finally, 2% (v/v) of overnightculture was inoculated into 1 l MRS broth andincubated overnight at 30�C. The metaboliteswere collected by separating the bacterial cells bycentrifugation at 10000 g for 15 min. The metab-olites were then kept at 4�C until further usage,as described by Foo et al. (2003) and Loh et al.(2010). A study conducted by Thu et al. (2011)showed that the metabolites consisted mainly ofbacteriocins (1600 AU/ml), and organic acidssuch as lactic acid (3�43 g/l) and acetic acid(1�18 g/l).
Experimental animals and diets
A total of 500 Lohmann Brown layers, aged 19weeks, were used in this study. They wererandomly assigned to 5 numerically equalgroups, with 50 replicates per treatment, twobirds per cage with a size of 55� 50� 45 cm3
(width�depth�height). The feeding periodlasted 12 weeks, commencing when the layerswere 19 weeks of age and ending when they were31 weeks of age, with the addition of one weekfor feed adaptation. Water and basal diet(Table 1) were supplied ad libitum. The dietarytreatments consisted of: (i) basal diet withoutsupplementation of metabolites (control); (ii)basal diet supplemented with 0�3% COM456metabolites; (iii) basal diet supplemented with0�6% COM456 metabolites; (iv) basal dietsupplemented with 0�9% COM456 metabolites;and (v) basal diet supplemented with 1�2%COM456 metabolites. The metabolite combina-tions used in this study were prepared and storedseparately and were mixed at equal ratios beforebeing added to the feed.
Laying performance
Feed intake was measured by subtracting thebalance of feed from the quantity originallysupplied to the laying hens. Eggs from individualcages were collected daily and weighed. Thehen/day egg production and feed efficiency werecalculated as the rate of production per hen per dand feed intake/egg mass.
DIETARY LACTOBACILLUS METABOLITES IN LAYING HENS 107
Dow
nloa
ded
by [
Uni
vers
iti P
utra
Mal
aysi
a] a
t 17:
58 1
9 M
arch
201
2
Faecal lactic acid bacteria (LAB) andEnterobacteriaceae count (ENT)
Fresh faecal droppings were collected asepticallyonce weekly in the morning, using a sterile plasticbag from every cage, and stored in the chillerprior to laboratory analysis. The faecal LAB andENT population was determined using themethod described by Foo et al. (2003) and Lohet al. (2010). Faecal samples were diluted insterile peptone water to 10% (w/v) and left atroom temperature for an hour prior to tenfoldserial dilutions (v/v). Enumerations of LAB werecarried out on Lactobacillus agar DE Man,ROGOSA and SHARPE (MRS-agar, Merck,KgaA, Darmstadt, Germany). The plates wereincubated anaerobically at 30�C for 48 h. ENTwere spread and enumerated on Eosin-methy-lene-blue lactose sucrose agar plates (EMB-agar,Merck, KgaA, Darmstadt, Germany) and incu-bated aerobically for 24 h at 37�C. The number ofcolony forming units (CFU) was expressed as thebase 10 logarithm of CFU (log10 CFU) per gram.All samples were repeated in triplicate. TheLAB/ENT ratio was calculated and presentedaccording to the average of LAB and ENT overthe experimental period.
Faecal pH
One gram of fresh faecal sample was mixedhomogeneously with 9 ml of deionised distilledwater in a sterile tube. The pH was measured
using a Mettler-Toledo pH meter with a glasselectrode (Mettler-Toledo, England). The meterwas calibrated using a buffer solution (Merck,KGaA, Darmstadt, Germany) at pH 4 and 7.
Small intestine morphology
At the end of the experiment, the birds werehumanely killed by the gas stunning methodfollowed by ventral cutting of the neck, whichsevered both carotid arteries resulting in rapiddeath. Different sections of the small intestinewere obtained for morphometric analysis. Theprocedure was a modified method from Hair-Bejo (1990), as described by Thanh et al. (2009).Segments of about 5 cm in length were removedfrom the small intestine (duodenum, jejunumand ileum) at the following locations: (i) themiddle part of the duodenal loop, (ii) midwaybetween the endpoint of the duodenal loop andMeckel’s diverticulum (jejunum) and (iii) midwaybetween the Meckel’s diverticulum and the ileo-caecal junction. Removed segments were flushedwith a 10% neutral buffered formalin solutionand were then used for morphometric analysis.For morphometric analysis, segments were fixedin a 10% neutral buffered formalin solutionovernight. Intestinal samples were then excised,dehydrated in a tissue processing machine (LeicaMicrosystem K. K. Tokyo, Japan) and embeddedin paraffin wax. Sections of 4 mm were cut fromeach sample, fixed onto slides, stained withhaematoxylin and eosin, mounted and examined
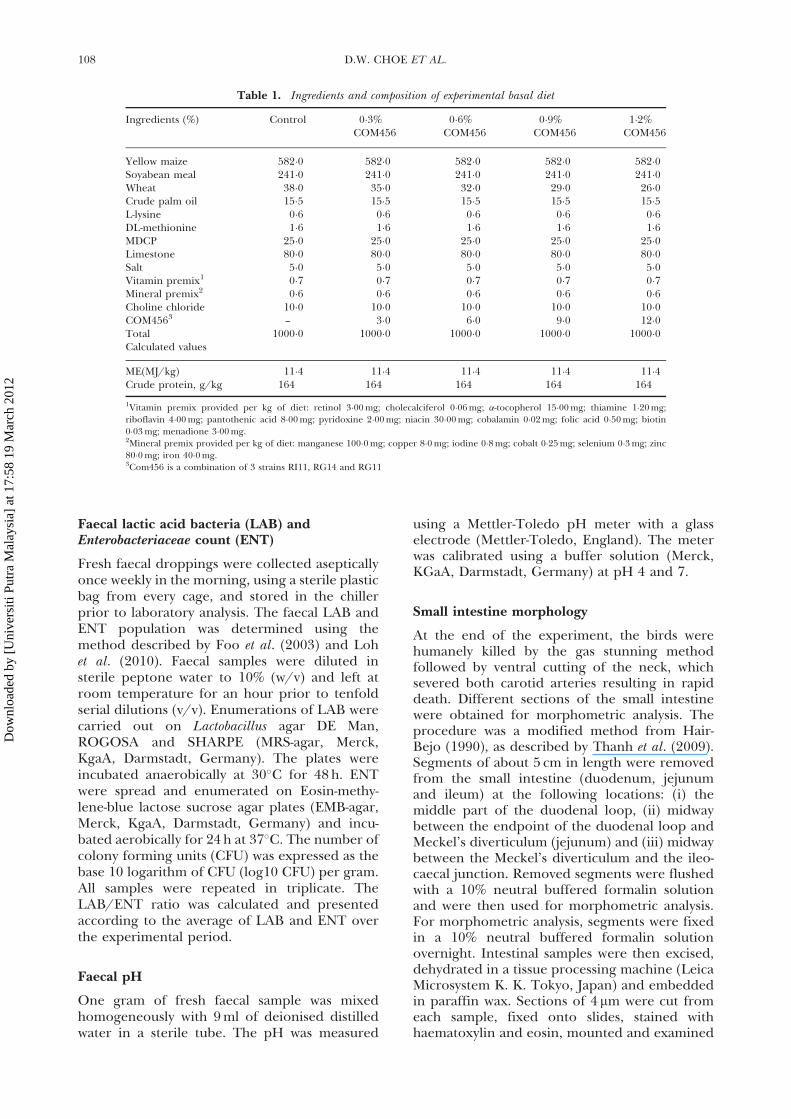
Table 1. Ingredients and composition of experimental basal diet
Ingredients (%) Control 0�3% 0�6% 0�9% 1�2%COM456 COM456 COM456 COM456
Yellow maize 582�0 582�0 582�0 582�0 582�0Soyabean meal 241�0 241�0 241�0 241�0 241�0Wheat 38�0 35�0 32�0 29�0 26�0Crude palm oil 15�5 15�5 15�5 15�5 15�5L-lysine 0�6 0�6 0�6 0�6 0�6DL-methionine 1�6 1�6 1�6 1�6 1�6MDCP 25�0 25�0 25�0 25�0 25�0Limestone 80�0 80�0 80�0 80�0 80�0Salt 5�0 5�0 5�0 5�0 5�0Vitamin premix1 0�7 0�7 0�7 0�7 0�7Mineral premix2 0�6 0�6 0�6 0�6 0�6Choline chloride 10�0 10�0 10�0 10�0 10�0COM4563 — 3�0 6�0 9�0 12�0Total 1000�0 1000�0 1000�0 1000�0 1000�0Calculated values
ME(MJ/kg) 11�4 11�4 11�4 11�4 11�4Crude protein, g/kg 164 164 164 164 164
1Vitamin premix provided per kg of diet: retinol 3�00 mg; cholecalciferol 0�06 mg; �-tocopherol 15�00 mg; thiamine 1�20 mg;
riboflavin 4�00 mg; pantothenic acid 8�00 mg; pyridoxine 2�00 mg; niacin 30�00 mg; cobalamin 0�02 mg; folic acid 0�50 mg; biotin
0�03 mg; menadione 3�00 mg.2Mineral premix provided per kg of diet: manganese 100�0 mg; copper 8�0 mg; iodine 0�8 mg; cobalt 0�25 mg; selenium 0�3 mg; zinc
80�0 mg; iron 40�0 mg.3Com456 is a combination of 3 strains RI11, RG14 and RG11
108 D.W. CHOE ET AL.
Dow
nloa
ded
by [
Uni
vers
iti P
utra
Mal
aysi
a] a
t 17:
58 1
9 M
arch
201
2

under the light microscope. The morphometricvariables included villus height (from the tip ofthe villi to the villus-crypt junction) and cryptdepth (depth of the invagination between adja-cent villi). The villus height and crypt depth weremeasured and recorded using an image analyser.Only villi sectioned from their base to the topwith a single epithelial layer at their tip, andcrypts with a visible lumen along their entiredepth, were considered for quantitative evalua-tion (Czernichow et al., 1996).
Plasma and yolk cholesterol
Blood samples were collected by jugular veinpuncture into tubes containing EDTA(Vacutainer, USA). Plasma was obtained aftercentrifuging the blood at 3000 g for 10 min. Totalplasma cholesterol concentrations were deter-mined through the Enzymatic Endpoint Methodusing a commercial diagnostic kit (Randox, UK)as described by Loh et al. (2002, 2003). Thedetermination of yolk cholesterol was performedusing a direct saponification gas chromato-graphic method (Folch et al., 1957).Approximately 1�0 g egg yolk was saponifiedwith 3 ml of 33% KOH and 95% methanol in a70�C waterbath for 90 min. After saponification,10 ml of hexane and 3 ml of water were added,followed by vigorous shaking for 10 min. Aninternal standard (5�- cholestan) was used. Thesolution was extracted with hexane and analysedby gas chromatography (HP 6890, USA)equipped with a fused silica (30 m� 0�32 mm�0�20 mm) capillary column (Loh et al., 2009).
Statistical analysis
The data were analysed using the GLM proce-dure by Statistical Analysis System (SAS, 1998).Orthogonal polynomial contrasts were used to
test the linear or quadratic nature of the responseto incremental concentrations of the liquidmetabolites. Only the results of linear and qua-dratic contrasts were reported in the presentpaper, because no significant cubic effects weredetected. The differences among treatments,when significant, were ordered using Tukey’stest (Kuehl, 1994). The results of statisticalanalysis were presented as mean� standarderror of the mean (SEM).
RESULTS
Laying performance
The egg production performance for variousdietary treatments is presented in Table 2. Therewas no significant difference (P > 0�05) among thetreatment groups in overall feed intake, eggweight, egg mass and feed conversion efficiency.However, the treatment group supplementedwith 0�6% COM456 metabolites had significantlyhigher (P < 0�05) hen/day egg production thanthe other groups.
Faecal pH
The faecal pH for various treatment groups isshown in Table 3. There was no significantdifference (P > 0�05) in faecal pH among thetreatment groups at the first week of sampling(21 weeks of age). However, the treatmentsupplemented with 1�2% COM456 metabolitesstarted to show a significant reduction (P < 0�05)in faecal pH, compared with the other groups,from 23 weeks of age onward. The treatmentgroup supplemented with 0�9% COM456 metab-olites started to exhibit a significant effect inreduced faecal pH at 27 weeks of age, whereasreduction (P < 0�05) of pH in the treatment groupsupplemented with 0�6% COM456 metaboliteswas observed at 29 weeks of age.
Table 2. Feed intake, hen/day egg production, egg weight, egg mass and feed conversion efficiency following treatments supplementedwith various concentrations of metabolites from COM456
Dietary treatments1
Control 0�3% COM456 0�6% COM456 0�9% COM456 1�2% COM456 SEM Linear Quad
Hen/d egg production (%) 82�83b 83�15b 85�45a 82�47b 81�98b 0�65 NS *Feed intake (g/hen/d) 83�51 84�23 84�48 82�97 82�35 1�03 NS NSEgg weight (g/egg) 54�88 55�32 55�41 54�73 54�90 0�83 NS NSEgg mass (g/hen/d) 45�49 46�08 47�21 45�20 45�18 0�98 NS NSFeed conversion efficiency 1�83 1�82 1�80 1�83 1�82 0�03 NS NS
1Com456 is a combination of 3 strains RI11, RG14 and RG112a-bMeans in the same row not sharing a common superscript are significantly different (P < 0�05).3SEM: standard error of the means (pooled).4Linear or quadratic response estimated using orthogonal polynomial contrasts (NS: non-significant; * P < 0�05).5Data are means of 50 cages of 2 hens per cage.
DIETARY LACTOBACILLUS METABOLITES IN LAYING HENS 109
Dow
nloa
ded
by [
Uni
vers
iti P
utra
Mal
aysi
a] a
t 17:
58 1
9 M
arch
201
2

Faecal lactic acid bacteria andEnterobacteriaceae count
Faecal LAB and ENT counts are presented inTable 4. There was no significant difference(P > 0�05) in LAB count among the dietary treat-ments at 21 and 23 weeks of age. However, thetreatment groups supplemented with 0�9% and1�2% COM456 metabolites started to show sig-nificantly higher (P < 0�05) LAB counts from 25weeks of age onward. The treatment groupsupplemented with 0�6% COM456 metaboliteshad a significant increase (P < 0�05) in faecal LABcounts from 27 weeks of age onwards. Incontrast, a significant reduction (P < 0�05) offaecal ENT (Table 5) was observed in the treat-ment groups supplemented with 1�2% COM456metabolites from 27 weeks of age onward,whereas 0�6% and 0�9% COM456 exhibitedsimilar effects at 29 weeks of age comparedwith the control group. In general, the treatmentgroup supplemented with a 0�3% combination ofCOM456 metabolites did not exhibit any desir-able effect, as compared with the control group.The mean ENT/LAB ratio is presented in
Table 6. There was no significant difference(P > 0�05) in LAB/ENT ratio between the controlgroup and 0�3% COM456. However, 0�9% and1�2% of COM456 showed a significantly higherLAB/ENT ratio, followed by 0�6% COM456(P < 0�05) as compared with the control group.
Plasma and yolk cholesterol
The plasma and yolk cholesterol compositionsare presented in Tables 7 and 8. At the end of theexperiment, all treatment groups supplementedwith COM456 metabolites had significant reduc-tions (P < 0�05) in plasma and yolk cholesterol.The lowest concentrations (P < 0�05) of plasmacholesterol were observed in the groups with0�9% and 1�2% COM456 metabolites, while 0�6%COM456 metabolites induced the lowest concen-tration (P < 0�05) of yolk cholesterol.
Small intestinal villus height and crypt depth
The intestinal villus height and crypt depth forthe control group and birds treated with differ-ent concentrations of metabolite combinations
Table 4. Faecal LAB counts following treatments supplemented with various concentrations of metabolites from COM456
Dietary treatments1
Control 0�3% COM456 0�6% COM456 0�9% Com456 1�2% COM456 SEM Linear Quad
LAB counts (log CFU/g)Week21 6�06 6�12 6�18 6�20 6�11 0�04 NS NS23 5�98 6�06 6�12 6�11 6�04 0�03 NS NS25 6�08b 6�13b 6�21b 6�47a 6�47a 0�06 * NS27 5�96c 6�01c 6�33b 6�96a 6�88a 0�05 * NS29 5�78c 5�84c 6�88b 6�90b 7�22a 0�07 * NS
1Com456 is a combination of 3 strains RI11, RG14 and RG11.2a-cMeans in the same row not sharing a common superscript are significantly different (P < 0�05).3SEM: standard error of the means (pooled).4Linear or quadratic response estimated using orthogonal polynomial contrasts (NS: non-significant; * P < 0�05).5Data are means of 50 cages of 2 hens per cage.
Table 3. Faecal pH following treatments supplemented with various concentrations of metabolites from COM456
Dietary treatments1
Control 0�3% COM456 0�6% COM456 0�9% Com456 1�2% COM456 SEM Linear Quad
pHWeek21 7�00 6�97 6�98 6�95 6�95 0�01 NS NS23 7�00a 6�99a 6�98a 6�98a 6�86b 0�02 * NS25 6�98a 6�98a 6�97a 6�96a 6�86b 0�02 * *27 6�96a 6�95a 6�95a 6�79b 6�78c 0�04 * NS29 6�97a 6�95a 6�91b 6�82c 6�83c 0�03 * NS
1Com456 is a combination of 3 strains RI11, RG14 and RG11.2a-cMeans in the same row not sharing a common superscript are significantly different (P < 0�05).3SEM: standard error of the means (pooled).4Linear or quadratic response estimated using orthogonal polynomial contrasts (NS: non-significant; * P < 0�05).5Data are means of 50 cages of 2 hens per cage.
110 D.W. CHOE ET AL.
Dow
nloa
ded
by [
Uni
vers
iti P
utra
Mal
aysi
a] a
t 17:
58 1
9 M
arch
201
2

Table 5. Faecal ENT counts following treatments supplemented with various concentrations of metabolites from COM456
Dietary treatments1
Control 0�3% COM456 0�6% COM456 0�9% Com456 1�2% COM456 SEM Linear Quad
ENT counts (log CFU/g)Week21 7�65 7�68 7�57 7�67 7�77 0�03 NS NS23 7�59 7�60 7�62 7�51 7�36 0�05 NS NS25 7�40 7�30 7�45 7�07 7�11 0�07 NS NS27 7�03a 6�93a 6�48ab 6�45ab 6�18c 0�07 * NS29 6�63a 6�69a 6�24b 5�35c 5�83c 0�11 * NS
1Com456 is a combination of 3 strains RI11, RG14 and RG11.2a-cMeans in the same row not sharing a common superscript are significantly different (P < 0�05).3SEM: standard error of the means (pooled).
Table 8. Egg yolk cholesterol following treatments supplemented with various concentrations of metabolites from COM456 at 31weeks of age
Dietary treatments1
Control 0�3% COM456 0�6% COM456 0�9% COM456 1�2% COM456 SEM Linear Quad
Yolk weight (g) 16�33 16�32 16�31 16�30 16�34 0�05 NS NSYolk cholesterol (mg/100 g) 12�73a 12�17b 11�36c 11�96b 12�13b 0�17 * *Yolk cholesterol (mg/yolk) 209�62a 199�71b 187�31d 195�74c 199�43b 2�28 * *
1Com456 is a combination of 3 strains RI11, RG14 and RG11.2a-dMeans in the same row not sharing a common superscript are significantly different (P < 0�05).3SEM: standard error of the means (pooled).4Linear or quadratic response estimated using orthogonal polynomial contrasts (NS: non-significant; * P < 0�05).5Data are means of 50 cages of 2 hens per cage.
Table 7. Plasma cholesterol following treatments supplemented with various concentrations of metabolites from COM456 at 31 weeksof age
Dietary treatments1
Control 0�3% COM456 0�6% COM456 0�9% COM456 1�2% COM456 SEM Linear Quad
Plasma(mg/dl) 186�28a 168�12b 152�62c 128�56d 125�42d 1�52 * NS
1Com456 is a combination of 3 strains RI11, RG14 and RG11.2a-dMeans in the same row not sharing a common superscript are significantly different (P < 0�05).3SEM: standard error of the means (pooled).4Linear or quadratic response estimated using orthogonal polynomial contrasts (NS: non-significant; * P < 0�05).5Data are means of 50 cages of 2 hens per cage.
Table 6. Mean LAB/ENT ratio following treatments supplemented with various concentrations of metabolites from COM456
Dietary treatments1
Control 0�3% COM456 0�6% COM456 0�9% COM456 1�2% COM456 SEM Linear Quad
LAB/ENT ratio 0�81c 0�82c 0�90b 0�98a 0�98a 0�02 * NS
1Com456 is a combination of 3 strains RI11, RG14 and RG11.2a-cMeans in the same row not sharing a common superscript are significantly different (P < 0�05).3SEM: standard error of the means (pooled).4Linear or quadratic response estimated using orthogonal polynomial contrasts (NS: non-significant; * P < 0�05).5Data are means of 50 cages of 2 hens per cage.
DIETARY LACTOBACILLUS METABOLITES IN LAYING HENS 111
Dow
nloa
ded
by [
Uni
vers
iti P
utra
Mal
aysi
a] a
t 17:
58 1
9 M
arch
201
2

of COM456 are presented in Table 9. All of thebirds supplemented with metabolite combina-tions of COM456 had higher (P < 0�05) duodenalvilli, compared with the control group. Thehighest duodenal villus heights were observedin 0�6% COM456 (P < 0�05), followed by the birdsfed with 0�9%, 0�3% and 1�2% COM456 metab-olites. The birds treated with 0�6%, 0�9% and1�2% COM456 metabolites had higher (P < 0�05)jejunum and ileum villus heights, as comparedwith the control and 0�3% COM 456 groups.
The birds supplemented with the 0�6% and0�9% COM456 metabolites had the greatest(P < 0�05) duodenal crypt depths, followed by1�2% COM456, compared with the controlgroup. For jejunal crypts, the treatment groupssupplemented with 0�6% and 0�9% COM456 hadthe greatest crypt depths (P < 0�05). Lastly, great-est (P < 0�05) ileal crypt depths were observed inthe treatment group supplemented with 0�6%and 0�9% COM456, followed by 1�2%. In general,birds treated with 0�3% COM456 had a similareffect as those in the control group.
DISCUSSION
The feed intake, egg weight, egg mass and FCRwere not significantly different among the treat-ment groups. These results were in agreementwith a study conducted by Loh et al. (2007),which indicated similar attributes, in terms of eggproduction, when fermented products wereadded to the diets of laying hens. However,higher overall hen/day egg production wasobserved in the treatment group supplementedwith 0�6% of COM456, compared with othergroups. Various studies had demonstrated apositive influence on growth performance whenmetabolites are added to the feed or drinking
water (Foo et al., 2003; Loh et al., 2003; Thanhet al., 2009).
The present study demonstrated the effect ofmetabolite combinations in reducing (P < 0�05)faecal ENT and increasing (P� 0�05) LAB counts.A few in vitro and in vivo studies have shown thatLAB produced a range of metabolites withbactericidal effect on undesirable bacteria, suchas bacteriocins and organic acids (Forestier et al.,2001; Foo et al., 2003). The mechanism of actionof bacteriocins is very complex. Most of thebacteriocins produced by LAB are less effectiveagainst gram-negative bacteria (Skytta et al.,1993). However, recent studies have shown thatcertain LAB bacteriocins possess a positive anti-microbial effect toward gram-negative bacteria(Ivanova et al., 1998; Suma et al., 1998). Theorganic acids present in the metabolites acted insuch a way that, when the acid passes through thebacterial cell wall, it disperses into the internalenvironment and reduces the pH (Beal et al.,2002). This causes the cessation of enzymaticprocesses and the collapse of the bacterial protonmotive force. Thus, the growth of the bacteria isinhibited, as its remaining energy is fully utilisedto maintain the internal pH for survival and notfor other metabolic processes. Daeschel andPenner (1992) indicated a synergistic effect oforganic acid with other antimicrobial com-pounds, such as hydrogen peroxide, in which ithelps in potentiating the antimicrobial effect ofother components against pathogenic bacteria.
Laying hen diets supplemented with metab-olites had higher populations of faecal LAB,compared with the control group. The acidifyingeffect of the organic acids (acetate, butyrate andpropionate) in the metabolites provides a goodenvironment for indigenous LAB to grow andflourish. Fuller (1977) reported that LAB aremuch better adapted to an acidic environment
Table 9. Villi height and crypt depth following treatments supplemented with various concentrations of metabolites from COM456
Dietary treatments1
Control 0�3% COM456 0�6% COM456 0�9% COM456 1�2% COM456 SEM Linear Quad
Villi height (mm)Duodenal 1025�62d 1067�88c 1198�69a 1096�39b 1058�20c 7�44 * *Jejunal 642�60c 650�49c 699�56a 708�72a 685�00b 5�96 * *Ileal 476�15c 476�69c 518�24a 527�49a 498�02b 5�70 * *
Crypt depth (mm)Duodenal 116�80b 115�50b 134�03a 129�52a 126�36a 1�64 * *Jejunal 90�06b 89�56b 100�03a 99�55a 91�24b 0�96 * *Ileal 85�98c 86�06c 98�36a 90�00b 89�65b 0�93 * *
1Com456 is a combination of 3 strains RI11, RG14 and RG11.2a-dMeans in the same row not sharing a common superscript are significantly different (P < 0�05).3SEM: standard error of the means (pooled).4Linear or quadratic response estimated using orthogonal polynomial contrasts (NS: non-significant; * P < 0�05).5Data are means of 50 cages of 2 hens per cage.
112 D.W. CHOE ET AL.
Dow
nloa
ded
by [
Uni
vers
iti P
utra
Mal
aysi
a] a
t 17:
58 1
9 M
arch
201
2

produced by lactic acid bacteria than are unde-sirable pathogens, such as Salmonella and E. coli.Therefore, the better-developing population ofLAB would suppress other bacteria in the intes-tinal system via competitive exclusion. In addi-tion, metabolites produced by LAB consist ofantimicrobial compounds, which inhibit the via-bility of ENT. The negative correlation betweenENT and LAB has proven that increased LABcount in treated groups could reduce the ENTpopulation (Loh et al., 2003; Loh et al., 2009a).
The pH of the faeces was similar in alltreatment groups for the first three weeks of theexperiment. However, a linear reduction offaecal pH was observed after that time point.The reduced pH in the gut could be due to theacidifying effect of organic acids, which arepresent in the metabolites. The acidic environ-ment would favour the growth of LAB, whichcould subsequently cause further production oforganic acids and antimicrobial compounds,such as bacteriocins. LAB ferment hexoses intolactic acid, acetic acid, formic acid and ethanolwhen there is a limitation to glucose supply (Fooet al., 2003).
In the present study, plasma cholesteroldecreased linearly with increasing concentrationof metabolite when compared with the controlgroups. This reduction could be due to the effectof metabolite supplementation, which provides abetter internal environment for LAB to grow andthrive in the gut. LAB has been reported to beable to utilise cholesterol and deconjugate bilesalt in the small intestine (Jin et al., 1998), thusmaking it less available for absorption into theblood circulation. Deconjugated bile salt isexcreted as part of the faecal material, whichstimulates synthesis of new bile salts for lipidmetabolism. Cholesterol in the blood circulationwill be utilised, as it is a necessary precursor forthe production of new bile salt (Chowdhury et al.,2005). This phenomenon could be the reason forthe reduction of plasma cholesterol in thepresent study.
The egg yolk cholesterol concentration wassignificantly reduced when metabolites wereadded to the feed. This result is probably dueto the bile-salt deconjugating activity of LAB inthe intestinal system, which reduces the availabil-ity of cholesterol to be incorporated into lipo-protein. Cholesterol in the blood circulation istransported by very low density lipoproteinsacross the ovarian membranes and depositedinto the developing yolk through the oocytevitellogenesis receptor (Elkin et al., 2009).Therefore, one of the major excretion pathwaysof cholesterol is through the production of eggyolk. Reduced availability of cholesterol in the
plasma would affect the concentration of choles-terol in the yolk. This possibility is in agreementwith the current findings, in which the plasmaand egg yolk cholesterol concentrations werelower in metabolite groups than in the controlgroup.
Villi are important structures in the smallintestine, involved mainly in nutrient absorption.Therefore, an increased villus height wouldincrease the surface area for nutrient absorption.However, the growth of villi is greatly dependenton the regeneration capability of enterocytesfrom the crypts of Lieberkuhn (Kirk, 2000). Anyinfectious or noninfectious processes that causeinflammation and damage to the intestinalmucosa may lead to the shortening of villi,which subsequently impairs digestive and absorp-tive activities. Bacteriocins, organic acids andother antimicrobial compounds present in themetabolites are able to alleviate undesirablepathogens which may cause subclinical infections(Miles et al., 2006). As a result, added metabolitesmay provide a better environment for the growthand development of the villi and crypts ofLieberkuhn, as found in those treatment groupssupplemented with metabolite combinations.Thanh et al. (2009) and Loh et al. (2010) reportedsimilar observations when metabolites wereadded to the diet of broiler chickens.
In conclusion, the supplementation of 0�6%COM456 metabolites gave the best result inimproving the hen/day egg production, andshowed positive effects such as increased faecalLAB and reduced ENT count, reduced faecal pH,reduced plasma and yolk cholesterol, andincreased small intestine villus height. This find-ing indicates that metabolites from locally iso-lated L. plantarum are a possible alternative feedadditive in poultry production.
ACKNOWLEDGEMENTS
The authors would like to express sincere gratitudeto Lay Hong Berhad for providing the facilities tocarry out the study, to S. Patmanaban Saminathanand the farm workers for their kind involvementand valuable dedication throughout the course ofthe feeding trial and, finally, to Goh Yong Meng,UPM, Malaysia, for assistance in the statisticalanalysis of data. This study was supported by agrant from Malaysia Technology DevelopmentCentre (MTDC).
REFERENCES
BEAL, J.D., NIVEN, S.J., CAMPBELL, A. & BROOKE, P.H. (2002)The effect of temperature on growth and persistent ofSalmonella in fermented liquid pig feed. InternationalJournal of Food Microbiology, 79: 99–104.
DIETARY LACTOBACILLUS METABOLITES IN LAYING HENS 113
Dow
nloa
ded
by [
Uni
vers
iti P
utra
Mal
aysi
a] a
t 17:
58 1
9 M
arch
201
2

CHOWDHURY, S.R., SARKER, D.K., CHOWDHURY, S.D.,SMITH, T.K., ROY, P.K. & WAHID, M.A. (2005) Effects ondietary tamarind on cholesterol metabolism in laying hens.Poultry Science, 84: 56–60.
CZERNICHOW, B., GALLUSER, M., CUI, S.Q., GOSSE, F. & RAUL, F.(1996) Comparison of enteral and parenteraladministration of medium chained triglycerides onintestinal mucosa in adult rats. Nutrition Research,16(5): 797–804.
DAESCHEL, M.A. & PENNER, M.H. (1992) Hydrogen peroxide,lactoperoxidase systems and reuterin, in: RAY, B. &DAESCHEL, M. (Eds) Food Preservatives of Microbial Origin,pp. 155—176 (Boca Raton, FL, CRC Press).
ELKIN, R.G., YAN, Z.H., CHONG, Y., DONKIN, S.S.,BUHMAN, K.K., STORY, J.A., TUREK, J.J., PORTER JNR, R.E.,ANDERSON, M., HOMAN, R. & NEWTON, R.S. (1999) Select3-Hydroxyl-3 - Methylglutaryl-Coenzyme A Reductaseinhibitors vary in their ability to reduce egg yolkcholesterol levels in laying hens through alteration ofhepatic cholesterol biosynthesis and plasma VLDL com-position. Journal of Nutrition, 129: 1010–1019.
FOLCH, J., LEES, M. & STANLEY, G.H. (1957) A simple methodfor the isolation and purification of total lipids fromanimal tissues. Journal of Biological Chemistry, 226: 497–509.
FOO, H.L., LOH, T.C., LAW, F.L., LIM, Y.Z., KUFLI, C.N. &RUSUL, G. (2003) Effects of feeding Lactobacillusplantarum I-UL4 isolated form Malaysian Tempeh ongrowth performance, faecal flora and lactic acid bacteriaand plasma cholesterol concentrations in postweaning rats. Journal of Food Science and Biotechnology,12: 403–408.
FORESTIER, C., DE CHAMPS, C., VATOUX, C. & JOLY, B. (2001)Probiotics activity of Lactobacillus casei rhamnous: in-vitroadherence to intestinal cells and antimicrobial activity.Research in Microbiology, 152: 167–173.
FULLER, R. (1977) The importance of Lactobacilli in main-taining normal microflora balance in the crop. BritishPoultry Science, 18: 85–94.
HAIR-BEJO, M. (1990) Gastrointestinal response to copperexcess: studies on copper (and zinc) loaded rats. Ph.D.Thesis. Department of Veterinary Pathology, University ofLiverpool, United Kingdom.
IVANOVA, I., MITEVA, V., STEFANOVA, T.S., PANTEV, A.,BUDAKOV, I., DANAVO, S., MONCHEVA, P., NIKOLOVA, I.,DOUSSET, X. & BOYAVAL, P. (1998) Characterizationof a bacteriocin produced by Streptococcus thermophi-lus 81. International Journal of Food Microbiology,42: 147–158.
JIN, L.Z., HO, Y.W., ABDULLAH, N. & JALALUDDIN, S. (1998)Growth performance, intestinal microbialpopulations, and serum cholesterol of broilers fed dietscontaining Lactobacillus cultures. Poultry Science,77: 1259–1265.
KIRK, C.K. (2000) Comparative Avian Nutrition (Wallingford,UK, CAB International).
KUEHL, R.O. (1994) Design of Experiments: Statistical Principlesof Research Design and Analysis, 2nd ed. (Brooks, CA,Duxbury Press).
LOH, T.C., FOO, H.L., TAN, B.K. & JELAN, Z.A. (2002) Effectsof palm kernel cake on growth performance and bloodlipids in rats. Asian-Australasian Journal of Animal Science,15: 1165–1169.
LOH, T.C., FOO, H.L., TAN, S.H., GOH, Y.M.,SHUKRIYAH, M.H. & KUFLI, C.N. (2003) Effects offermented products on performance, faecal pH, enter-obacteriaceae and lactic acid bacteria counts andinterrelationships and plasma cholesterol concentrationin rats. Journal of Animal Feed and Sciences,12(3): 625–636.
LOH, T.C., LAW, F.L., FOO, H.L., GOH, Y.M. & ZULKIFLI, I.(2007) Effects of feeding a fermented product on eggproduction, fecal microflora and fecal pH in laying hens.Journal of Animal Feed and Sciences, 16: 452–462.
LOH, T.C., LEE, T.M., FOO, H.L., LAW, F.L. & RAJION, M.A.(2008) Growth performance and faecal microflora of ratsoffered with metabolites from lactic acid bacteria indrinking water. Journal of Applied Animal Research,34: 61–64.
LOH, T.C., CHONG, S.W., FOO, H.L. & LAW, F.L. (2009a)Effects on growth performance, faecal microflora andplasma cholesterol after supplementation of spray driedmetabolite to post weaning rats. Czech Journal of AnimalSciences, 54(1): 10–16.
LOH, T.C., LAW, F.L., FOO, H.L., GOH, Y.M. & ZULKIFLI, I.(2009b) Effects of feeding fermented fish on egg choles-terol content in hens. Journal of Animal Sciences, 80: 27 – 33.
LOH, T.C., THANH, N.T., FOO, H.L., BEJO, M.H. & KASIM, A.(2010) Feeding of different levels of metabolite combina-tions produced by Lactobacillus plantarum on growthperformance, faecal lactic acid bacteria andEnterobacteriaceae count, volatile fatty acids and villiheight in broilers. Journal of Animal Sciences, 81: 205–214.
MARCEL, B.R. (2000) Prebiotics and probiotics: are theyfunctional foods? American Journal of Clinical Nutrition,71(6): 1282–1287.
MIKKELSEN, L.L. & JENSEN, B.B. (2000) Effects of fermentedliquid feed on activity and composition of the micro-biodata in the guts of pigs. Pig News Information, 21: 59–66.
MILES, R.D., BUTCHER, G.D., HENRY, P.R. & LITTELL, R.C.(2006) Effect of antibiotic growth promoters on broilerperformance, intestinal growth parameters and quantita-tive morphology. Poultry Science, 85: 476–485.
PARKER, J., OVIEDO-RONDON, E.O., CLACK, B.A., CLEMENTE-HERNANDEZ, S., OSBORNE, J., REMUS, J.C., KETTUNEN, H.,MAKIVUOKKO, H. & PIERSON, E.M. (2007) Enzymes as feedadditives to aid in response against Eimeria spp. incoccidian vaccinated broilers fed corn-soybean mealdiets with different protein levels. Poultry Science,86: 643–653.
REQUE, E.F., PANDEY, A., FRANCO, S.G. & SOCCOL, G.R. (2003)Isolation, identification and physiological study ofLactobacillus fermentum LPB for use as probiotics inchickens. Brazillian Journal of Microbiology, 31: 303–307.
SHALINI, M. & RAMESHWAR, S. (2005) Antibiotic resistant infood lactic acid bacteria. International Journal of FoodMicrobiology, 105: 281–295.
SKYTTA, E., HAIKARA, A. & MATTILDA-SANDHOLM, T. (1993)Production and characterization of antibacterialcompounds produced by Pediococcus damnosusand P. pentosaceus. Journal of Applied Bacteriology,74: 134–142.
SORUM, H. & SUNDE, M. (2001) Resistence to antibiotics inthe normal flora of animals. Veterinary Resources,32: 227–241.
STATISTICAL ANALYSIS SYSTEM (SAS) (1998) SAS User Guide:Statistic SAS Institute Inc., North Carolina, USA.
SUMA, K., MISRA, M.C. & VADARAJ, M.C. (1998) PlantaricinLP84, a broad spectrum heat stable bacteriocin ofLactobacillus plantarum NCIM 2084 produced in asimple glucose broth medium. International Journal ofFood Microbiology, 40: 17–25.
THANH, N.T., LOH, T.C., FOO, H.L., BEJO, M.H. &AZHAR, K. (2009) Effects of feeding metabolitecombinations produced by Lactobacillus plantarumon growth performance, faecal microflora population,small intestine villus height and faecal volatile fattyacids in broilers. British Poultry Science,50(3): 298–306.
114 D.W. CHOE ET AL.
Dow
nloa
ded
by [
Uni
vers
iti P
utra
Mal
aysi
a] a
t 17:
58 1
9 M
arch
201
2

THANH, N.T., LOH, T.C., FOO, H.L., BEJO, M.H. & AZHAR, K.(2010) Inhibitory activity of metabolites produced bystrains of Lactobacillus plantarum isolated formMalaysian fermented food. International Journal ofProbiotics and Prebiotics, 5(1): 37–44.
THU, T.V., LOH, T.C., FOO, H.L., HALIMANTUN, Y. & BEJO, M.H.(2010) Effects of liquid metabolites combinations byLactobacillus plantarum on growth performance, faecescharacteristics, intestinal morphology and diarrhea
incidence in postweaning piglets. Journal of TropicalAnimal Health and Production, 43(1): 69–75.
THU, T.V., FOO, H.L., LOH, T.C. & BEJO, M.H. (2011)Inhibitory activity and organic acid concentration ofmetabolite combinations produced by various strains ofLactobacillus plantarum. African Journal of Biotechnology(Accepted).
WANG, T.T. & LEE, B.H. (1997) Plasmids in Lactobacillus.Critical Review of Biotechnology, 17: 227–272.
DIETARY LACTOBACILLUS METABOLITES IN LAYING HENS 115
Dow
nloa
ded
by [
Uni
vers
iti P
utra
Mal
aysi
a] a
t 17:
58 1
9 M
arch
201
2
Copyright © 2022 FDOKUMEN




















