
The Oral Poetics of Professional Wrestling, or Laying the ...

This article was downloaded by: [117.211.89.164]On: 12 October 2014, At: 07:57Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Avian PathologyPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/cavp20
Experimental infection of egg‐layinghens with Salmonella enteritidis phagetype 4P. A. Barrow a & Margaret A. Lovell aa ARFC Institute for Animal Health, Houghton Laboratory ,Houghton, Huntingdon, Cambridgeshire, PE17 2DA, EnglandPublished online: 12 Nov 2007.
To cite this article: P. A. Barrow & Margaret A. Lovell (1991) Experimental infection ofegg‐laying hens with Salmonella enteritidis phage type 4, Avian Pathology, 20:2, 335-348, DOI:10.1080/03079459108418769
To link to this article: http://dx.doi.org/10.1080/03079459108418769
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to orarising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Avian Pathology (1991) 20, 335-348
Experimental infection of egg-laying hens withSalmonella enteritidis phage type 4
P. A. BARROW & MARGARET A. LOVELL
ARFC Institute for Animal Health, Houghton Laboratory, Houghton, Huntingdon,Cambridgeshire PE17 2DA, England
SUMMARY
Chickens were inoculated intravenously with 105 or 106 organisms. Heavy infection of theovaries occurred and some infection persisted in this organ for several weeks. Most of theovarian infections were confined to the interstitial tissues and not to the yolk contained in thelarge follicles. Infections of the ovary did not result from contamination from infected air sacs.None of 810 eggs laid contained S. enteritidis.
Chickens infected orally gave similar results to those following intravenous inoculation althoughthe number of isolations obtained from the caeca and cloaca were higher. S. enteritidis wasisolated from two of 633 eggs in which the contents only were cultured and from 36 of 614 eggsin which both shell and contents were cultured.
The serum IgG response to oral inoculation was monitored by an ELISA using a whole cellsonicate or lipopolysaccharide antigen. High titres of IgG were detected for 27 weeks afterinfection when the experiment was terminated.
The experiments suggest that most infected eggs laid by 5. enteritidis-infected hens are surface-contaminated and do not result from infected ovaries.
INTRODUCTION
There have been large increases in several countries over the last few years in the numberof cases of human food-poisoning caused by Salmonella enteritidis in which eggs havebeen a major source of infection (St. Louis et al., 1988; Report, 1989). In the UK phagetype 4 (PT4) strains are implicated more frequently than other phage types.
Analysis of outbreaks of human infection have revealed that intact shell eggs and foodprepared from raw eggs are the main source of infection (Coyle et al., 1988). Theorganism has been isolated from both the contents and the shells of such eggs (Mawer etal., 1989; Humphrey et al, 1989; Report, 1989). Hopper & Mawer (1988) investigated acommercial layer flock which had been the source of eggs implicated in an outbreak offood-poisoning. Although mortality in the flock was at a normal level, S. enteritidis wasisolated from 13 of 50 chickens which died and six of these showed ovarian infection. Theorganism was also isolated from a number of other tissues. Bygrave & Gallagher (1989)and Chart et al. (1989) have also isolated PT4 strains from the tissues of commercialbirds. The invasiveness of PT4 strains has been demonstrated experimentally in broilers
Received 22 June 1990; accepted 17 July 1990.
335
Dow
nloa
ded
by [
117.
211.
89.1
64]
at 0
7:57
12
Oct
ober
201
4

336 P. A. BARROW & MARGARET A. LOVELL
(Hinton et al, 1989; Barrow, 1990) and in layers (Humphrey et al, 1989; Timoney et al,1989). Because of the extensive involvement of 5. entérinais PT4 in mortality in youngbroilers and its isolation from broiler-breeders a number of authors have suggested thatthe organism can be transmitted vertically (Lister, 1988; Hopper & Mawer, 1988;Bygrave & Gallagher 1989). The ability of 5. enteritidis to localise in the ovary of thefowl has been recognised for some time (Faddoul & Fellows, 1966; Snoeyenbos et al,1969).
Although Humphrey et al (1989) did not detect IgG or IgM antibodies in theirexperimentally infected birds, Chart et al. (1989) were able to detect a strong serologicalresponse by ELISA. Detection of agglutinating antibodies by using the related S.pullorum stained antigen has been found to be of little use in detecting infected chickens(Hopper & Mawer, 1988; Cooper et al, 1989). The aim of the present experiments was totry to establish a model infection of S. enteritidis PT4 in laying hens in which the ovarycould become infected and in which the incidence of contaminated eggs could be studied.An infection model was essential for further studies on immune protection to be carriedout. Chickens were infected orally or intravenously, the latter to ensure that some degreeof ovarian infection occurred. In the experiment in which chickens were infected orallythe birds were bled regularly and the serological response to infection was monitored byELISA.
MATERIALS AND METHODS
Chickens
Commercial laying hens which had not been screened for Salmonella were obtained froma local producer. In one experiment they were obtained at 40 weeks of age and were usedtwo weeks after being housed at the Institute. In the second and third experiments 16-week-old chickens were obtained which were used at 24 weeks of age, several weeks aftercoming into lay. All chickens were housed individually in cages and were given layers'pellets (Labsure, Manea, Cambridgeshire, England) and water ad libitum.
On arrival, and immediately prior to an experiment starting, cloacal swabs were takenfrom each bird and cultured individually in selenite broth (CM395, Oxoid) for 18 h at37°C before being plated on to Brilliant Green agar (CM263 Oxoid) and Desoxycholateagar (CM163, Oxoid). Chickens in the third experiment were bled simultaneously and theserum tested in an ELISA using S. enteritidis antigens.
Eggs were collected on a daily basis from each bird from three weeks before inoculationuntil each experiment was terminated.
Bacteria and inoculation
A strain of S. enteritidis phage type 4, P125589, used throughout the experiments, waskindly supplied by Dr B. Rowe, Central Public Health Laboratory, London. It wasoriginally isolated from a case of human food-poisoning. For ease of enumeration aspontaneous nalidixic acid resistant mutant of this strain, was used. The strain wasmaintained on a Dorset's egg slope at 4°C.
Bacterial cultures were made in 10 ml nutrient broth (Oxoid, CM67) incubated at37°C for 24 h in a shaking water bath (100 strokes/min). These cultures containedapproximately 109 cfu/ml.
Chickens were inoculated intravenously with a decimal dilution of a broth culture
Dow
nloa
ded
by [
117.
211.
89.1
64]
at 0
7:57
12
Oct
ober
201
4

S. ENTERITIDIS IN LAYING HENS 337
containing either 105 or 106 cfu in 0.1 ml or orally with 0.3 ml of an undiluted culture,containing approximately 3 X 10s organisms in this volume, directly into the crop.
Experimental plan
Experiment 1
Fifty 40-week-old chickens, in-lay, were inoculated intravenously with 105 cfu. Tenchickens were killed weekly for 5 weeks post-inoculation and post-mortem examinationswere carried out after soaking the surface of the ventral side of the chicken in industrialalcohol. Organs were sampled for bacteriological analysis in the order: ovary, oviduct(with or without egg), myocardium, spleen, liver, caecum, cloaca. The oviduct, caeca andcloaca were swabbed. Complete ovaries were removed and samples of tissue were takenfrom the remaining organs. All laid eggs were cultured, examination being made for S.entérinais in the egg contents (see below).
Experiment 2
Ten 24-week-old chickens, in-lay, were inoculated intravenously with 106 organisms.Five chickens were killed at one and two weeks post-inoculation and post-mortemexaminations were again carried out as described above. Organs were sampled in theorder: ovary (yolk was removed after the ovary was taken out and cultured separatelyfrom the interstitial tissue), air sac, oviduct, myocardium, spleen, liver, caeca and cloaca.Air sac samples were taken as swabs. Laid eggs were cultured, examination for S.entérinais in the contents and from the entire egg being made on alternate days.
Experiment 3
Fifty-five 24-week-old chickens, in-lay, were inoculated orally with 3 x 108 organisms.Eight chickens were killed at one, two, three, five and seven weeks post-inoculation andpost-mortem examinations were carried out. Organs were removed or sampled in the orderdescribed in Expt 2. Caecal samples were taken for bacterial enumeration. Cloacal swabswere taken on a weekly basis and were examined for S. entérinais as described below.Five of the remaining chickens were bled from the wing vein at frequent intervals and theserum examined for Salmonella-sptci&c IgG by ELISA. Eggs were cultured as describedfor Expt 2.
Bacteriological analysis
All samples taken were plated directly and after overnight incubation at 37°C in selenitebroth onto Brilliant Green agar containing sodium nalidixate, 20 /ig/ml and novobiocin, 1fig/ml. Complete ovaries, interstitial ovarian tissue or yolk, released from the ovules withthe tip of a pair of sterile scissors, were placed directly into selenite broth. Once in theselenite the complete ovaries were crudely cut up with scissors. Eggs removed asepticallyfrom oviducts were also broken with the shells into selenite broth.
Swabs from air sacs, oviducts, caeca and cloaca were placed in 2 ml selenite broth. Thecloacal swabs were plated out using the standard method for semi-quantitative enumera-tion devised by Smith & Tucker (1975). They and the remaining swabs were thencultured overnight at 37°C.
Dow
nloa
ded
by [
117.
211.
89.1
64]
at 0
7:57
12
Oct
ober
201
4

338 P. A. BARROW & MARGARET A. LOVELL
The viable numbers of 5. entérinais organisms in tissue samples (liver, spleen,myocardium and caecal contents) were estimated using the methods of Miles et al.(1938). Decimal dilutions of homogenized tissue were made in selenite broth, and platedout and then incubated further at 37°C.
Laid eggs were cultured in two ways. The first method was to culture the contents.Eggs were candled and then soaked in a 1% solution of Diversol (chlorinated trisodiumphosphate; Diversey Ltd., Northampton, England) for 30 min before being washed for 5min in running tap water. They were air-dried and a small hole was made in the shell overthe air space. A sterile loop was inserted into the egg and the contents were homogenizedin situ for 1 min. The loop was removed and a piece of adhesive tape placed over thehole. The eggs were incubated at 37°C overnight before being plated onto the antibiotic-containing Brilliant Green agar. In the second method the whole egg was cultured. Theeggs were placed into sterile honey jars, broken and the contents mixed by shaking thejars. These were incubated at 37°C and plated out as described above.
A preliminary experiment was carried out in which eggs were experimentally inocu-lated via the albumen with 101 S. entérinais organisms using a 25-gauge needle. Eggsinoculated in this way and processed by the above methods yielded profuse growth of S.entérinais on Brilliant Green agar. This therefore indicated that the methods used wouldbe sufficiently sensitive to allow very small numbers of Salmonella organisms to bedetected.
Serological analysis
Sera obtained from chickens in experiment 3 maintained for 27 weeks post-inoculationwere analysed by an indirect ELISA. The method used was essentially that described byHassan et al. (1990). Essentially, the wells of an ELISA plate (Dynatech Laboratories)were coated with a solution of the chosen antigen (see below) in 50 HIM carbonate bufferpH 9.6. After washing the plate in phosphate buffered saline containing 0.1% Tween 20this was followed, in turn, by the test sera, then rabbit anti-chicken IgG alkalinephosphatase conjugate (Sigma) followed by the substrate, p-nitrophenyl phosphate indiethanolamine buffer pH 9.8. The plate was washed in between each stage. The S.entérinais antigens used, a whole bacterial cell sonicate and lipopolysaccharide (LPS)from strain 125589, were prepared as described by those authors. The LPS appeared to bepure when checked by polyacrylamide gel electrophoresis.
RESULTS
Experiment 1
The 40-week-old chickens in experiment 1 were well into lay at the time of inocu-lation and egg production, expressed as the number of eggs laid/the number of chickensper unit time, had stabilised at around 80% (Table 1). Within two weeks of infectionthe productivity had fallen to 69.5% and it did not return to 80% until four weeksafter inoculation. Whether this was a result of 5. entérinais infection or simply fromhandling and inoculation is unclear. In total, 801 eggs were laid during the courseof the experiment but S. entérinais was not isolated from the contents of any of theseeggs.
During the experiment one chicken died and a post-mortem examination revealedpericarditis typical of that associated with S. entérinais infection and hepatomegaly.
Dow
nloa
ded
by [
117.
211.
89.1
64]
at 0
7:57
12
Oct
ober
201
4

S. ENTERITIDIS IN LAYING HENS 339
Table 1. Egg production before and after infection withS. enteritidis phage type 4. Number of eggs produced/
number of chickens (%)
Weekspost-inoculation
- 2- 1
0+ 1+ 2+3+4+ 5
Expt 1(Inoculation
iv)
74.582.582.076.969.577.387.080.0
Expt 3(Inoculation
orally)
87.093.192.889.295.592.590.594.0
S. enteritidis was isolated from the liver but not from the heart. It was also isolated fromthe interstitial tissue of the ovary but there were no follicles present.
A second chicken had diarrhoea during the experiment. At slaughter at the third week,this bird was thin and had a congested liver. The ovary contained some inspissated,misshapen follicles. S. enteritidis was isolated from these and from the liver, spleen, heart,caecum, ovary and oviduct. All the remaining birds stayed healthy thoughout theexperiment. Three abnormal ovaries were found and S. enteritidis was isolated from twoof them. These two and a normal ovary for comparison, are shown in Figure 1. Thefollicles were misshapen and the contents of several of them were coagulated. Smallnecrotic lesions in the spleen, typical of Salmonella infection, were seen in 34 of 46 birds
Figure 1. Two abnormal ovaries (above) and a normal ovary for comparison (below) taken fromS. enteritidis infected birds. Note inspissated and misshapen follicles in abnormal ovaries.
Dow
nloa
ded
by [
117.
211.
89.1
64]
at 0
7:57
12
Oct
ober
201
4
340 P. A. BARROW & MARGARET A. LOVELL
examined. They were present in all 10 birds in the first week and had declined infrequency to 4/10 birds in the fifth week.
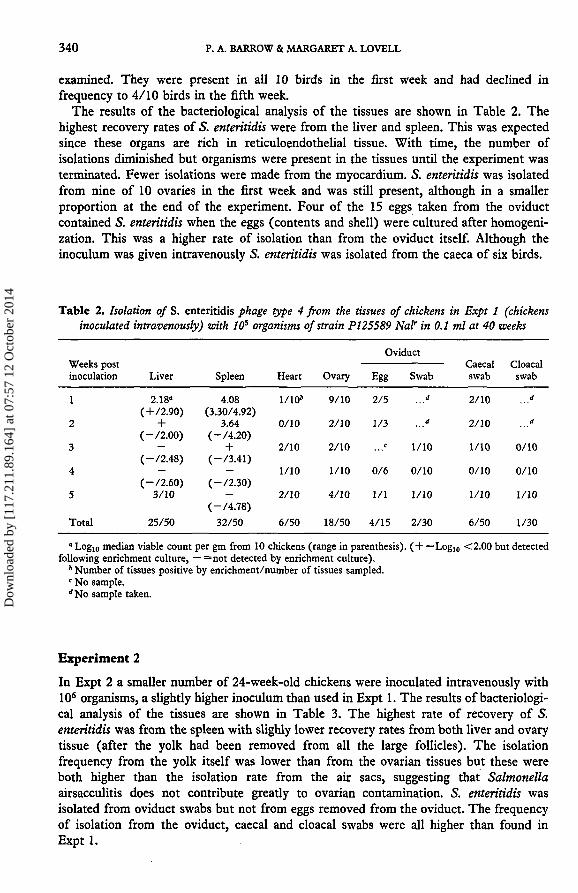
The results of the bacteriological analysis of the tissues are shown in Table 2. Thehighest recovery rates of 5. enteritidis were from the liver and spleen. This was expectedsince these organs are rich in reticuloendothelial tissue. With time, the number ofisolations diminished but organisms were present in the tissues until the experiment wasterminated. Fewer isolations were made from the myocardium. S. enteritidis was isolatedfrom nine of 10 ovaries in the first week and was still present, although in a smallerproportion at the end of the experiment. Four of the 15 eggs taken from the oviductcontained S. enteritidis when the eggs (contents and shell) were cultured after homogeni-zation. This was a higher rate of isolation than from the oviduct itself. Although theinoculum was given intravenously S. enteritidis was isolated from the caeca of six birds.
Table 2. Isolation of S. enteritidis phage type 4 from the tissues of chickens in Expt 1 (chickensinoculated intravenously) with 10s organisms of strain PI25589 Nal' in 0.1 ml at 40 weeks
Weeks postinoculation
1
2
3
4
5
Total
Liver
2.18°( + /2.90)+
(-/2.00)—
(-/2.48)—
(-/2.60)3/10
25/50
Spleen
4.08(3.30/4.92)
3.64(-/4.20)+
(-/3.41)—
(-/2.30)—
(-/4.78)32/50
Heart
1/10»
0/10
2/10
1/10
2/10
6/50
Ovary
9/10
2/10
2/10
1/10
4/10
18/50
Oviduct
Egg
2/5
1/3
c
0/6
1/1
4/15
Swab
d
d
1/10
0/10
1/10
2/30
Caecalswab
2/10
2/10
1/10
0/10
1/10
6/50
Cloacalswab
d
d
0/10
0/10
1/10
1/30
" Log10 median viable count per gm from 10 chickens (range in parenthesis). ( + =Log10 <2.00 but detectedfollowing enrichment culture, — =not detected by enrichment culture).
* Number of tissues positive by enrichment/number of tissues sampled.c No sample.''No sample taken.
Experiment 2
In Expt 2 a smaller number of 24-week-old chickens were inoculated intravenously with106 organisms, a slightly higher inoculum than used in Expt 1. The results of bacteriologi-cal analysis of the tissues are shown in Table 3. The highest rate of recovery of 5.enteritidis was from the spleen with slighly lower recovery rates from both liver and ovarytissue (after the yolk had been removed from all the large follicles). The isolationfrequency from the yolk itself was lower than from the ovarian tissues but these wereboth higher than the isolation rate from the air sacs, suggesting that Salmonellaairsacculitis does not contribute greatly to ovarian contamination. 5. enteritidis wasisolated from oviduct swabs but not from eggs removed from the oviduct. The frequencyof isolation from the oviduct, caecal and cloacal swabs were all higher than found inExpt 1.
Dow
nloa
ded
by [
117.
211.
89.1
64]
at 0
7:57
12
Oct
ober
201
4
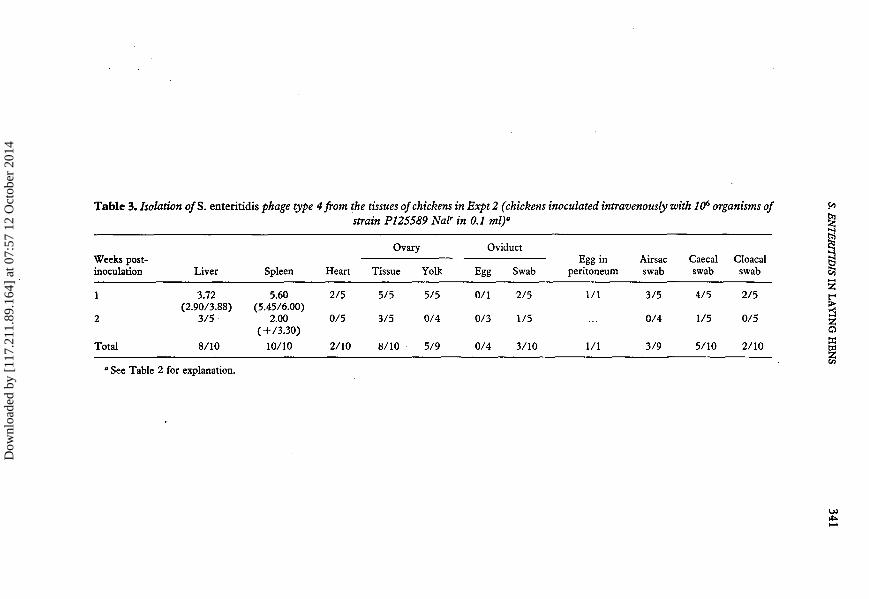
Table 3. Isolation of S. enteritidis phage type 4 from the tissues of chickens in Expt 2 (chickens inoculated intravenously with 106 organisms ofstrain PI25589 Nalr in 0.1 ml)"
Weeks post-inoculation
1
2
Total
Liver
3.72(2.90/3.88)
3/5
8/10
Spleen
5.60(5.45/6.00)
2.00(+/3.30)
10/10
Heart
2/5
0/5
2/10
Ovary
Tissue
5/5
3/5
8/10
Yolk
5/5
0/4
5/9
Oviduct
Egg
0/1
0/3
0/4
Swab
2/5
1/5
3/10
Egg inperitoneum
1/1
1/1
Airsacswab
3/5
0/4
3/9
Caecalswab
4/5
1/5
5/10
Cloacalswab
2/5
0/5
2/10
" See Table 2 for explanation.
IE1zoo
Dow
nloa
ded
by [
117.
211.
89.1
64]
at 0
7:57
12
Oct
ober
201
4

342 P. A. BARROW & MARGARET A. LOVELL
Experiment 3
Oral infection of 24-week-old chickens with S. enteritidis appeared to have little or noeffect on egg production compared with that resulting from intravenous inoculation(Table 1). In total 1247 eggs were laid. The two methods of examination (see Methodssection) were used on alternate days. S. enteritidis was isolated from 36 of 614 eggs inwhich the contents and shell were examined. The organism was isolated from two of 633eggs in which the contents only were examined. Most of these contaminated eggs weredetected within the first two weeks post-inoculation (Table 4).
Table 4. Isolation of S. enteritidis phage type 4 fromlaid eggs from chickens in Experiment 3 (chickens
inoculated orally)
Samplingdays post-inoculation
1-23-45-67-89-10
11-1213-1415-1617-1819-2021-48Total
Egg contents
1/50°1/310/560/410/270/550/260/350/300/300/2522/633
Egg contents and shell
5/404/34
12/535/285/442/361/382/320/290/300/250
36/614
" Number of eggs infected/number of eggs cultured.
The chickens remained healthy throughout the experiment. On post-mortem examina-tion spleen lesions were found in a total of 12 of 40 chickens. None was seen in the firstweek followed by 7, 1, 4 and 0 at 2,3,5 and 7 weeks post-inoculation respectively. Liverlesions were seen in two birds in the first week.
The results of bacteriological analysis of the tissues taken during the seven weeks ofthe experiment are shown in Table 5. Unlike the results following intravenous inoculationnot all chickens infected orally contained S. enteritidis organisms in their spleens oneweek later. The number of isolations from the liver and spleen reduced with time untilnone was isolated at 7 weeks post-inoculation. Again very few isolations were made fromthe myocardium. S. enteritidis was isolated frequently from the ovarian interstitial tissuesbut not from yolk separated from these tissues. Only one isolation was made from an airsac, in the first week post-inoculation. The number of isolations from the caecal andcloacal swabs was much higher than from the birds inoculated intraveneously, reflectingthe different route of inoculation.
Sera were obtained weekly for up to 27 weeks from five chickens in Expt 3. IgG titresdetected by ELISA using S. enteritidis sonicate and lipopolysaccharide antigens arerepresented in Figure 2 as the geometric mean titres with the standard deviation. By twoweeks post-inoculation titres detected by the sonicate increased markedly and stayed highfor a further month or so. Although these titres declined for several weeks after this timethey rose again and stayed high until the chickens were killed 27 weeks post-inoculation.The mean titres were all higher post-inoculation than before inoculation. The titres
Dow
nloa
ded
by [
117.
211.
89.1
64]
at 0
7:57
12
Oct
ober
201
4
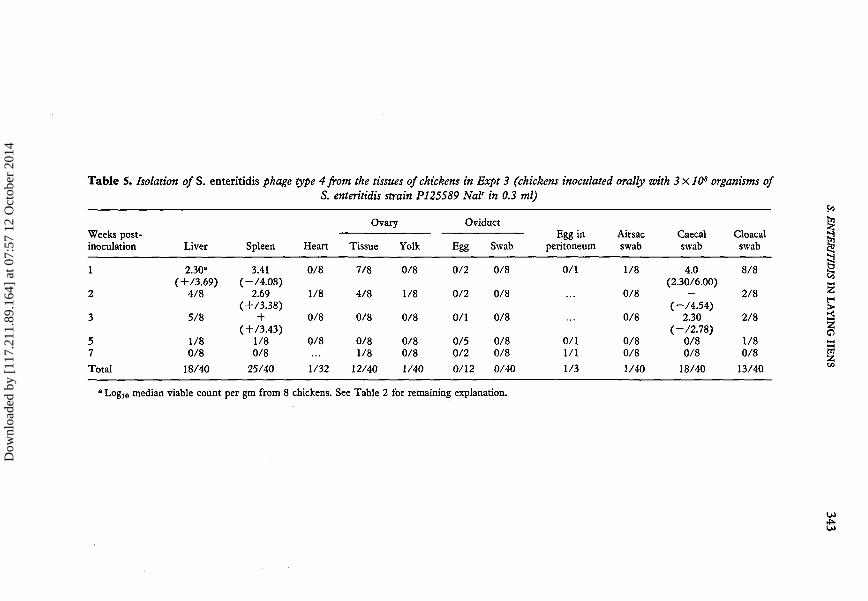
Table 5. Isolation of S. enteritidis phage type 4 from the tissues of chickens in Expt 3 (chickens inoculated orally with 3xlOs organisms ofS. enteritidis strain P125S89 Nalr in 0.3 ml)
Weeks post-inoculation
1
2
3
57
Total
Liver
2.30«(+/3.69)
4/8
5/8
1/80/8
18/40
Spleen
3.41(-/4.08)
2.69(+/3.38)
+( + /3.43)
1/80/8
25/40
Heart
0/8
1/8
0/8
0/8
1/32
Ovary
Tissue
7/8
4/8
0/8
0/81/8
12/40
Yolk
0/8
1/8
0/8
0/80/8
1/40
Oviduct
Egg
0/2
0/2
0/1
0/50/2
0/12
Swab
0/8
0/8
0/8
0/80/8
0/40
Egg inperitoneum
0/1
0/11/1
1/3
Airsacswab
1/8
0/8
0/8
0/80/8
1/40
Caecalswab
4.0(2.30/6.00)
—(-/4.54)
2.30(-/2.78)
0/80/8
18/40
Cloacalswab
8/8
2/8
2/8
1/80/8
13/40
I3
a
median viable count per gm from 8 chickens. See Table 2 for remaining explanation.
Dow
nloa
ded
by [
117.
211.
89.1
64]
at 0
7:57
12
Oct
ober
201
4

344 P. A. BARROW & MARGARET A. LOVELL
detected by the lipopolysaccharide antigen were initially lower than those detected by thesonicate indicating greater specificity. However, within two weeks of inoculation themean titres were higher than those detected by the sonicate and with one exception (23weeks post-inoculation) they remained higher. The standard deviations of the meansusing lipopolysaccharide antigen were greater than those using the sonicate.
9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27
Time (weeks) post inoculation
Figure 2. Serum IgG response to S. enteritidis phage type 4 infection in hens in experiment 3. IgGdetected by sonicate (•) and by lipopolysaccharide (o). All chickens infected orally with 10s
organisms at 24 weeks of age. Figure shows mean of 5 samples ± standard deviation.
DISCUSSION
Laid eggs were more frequently infected on the egg surface than in the contents of theegg. On only two occasions, in Expt 3, were egg contents infected. This was in layinghens infected at 24 weeks of age whereas none was found in the 40-week-old chickensinoculated intraveneously in Expt 1. A similar experience has been reported by Hum-phrey et al. (1989) where S. enteritidis-contaminated egg contents were recovered fromexperimentally infected 18- but not 52-week-old hens. The reason for this is unclear,since intravenous inoculation produces an initial haematogenous spread of infection to anumber of organs before bacteria are cleared from the blood by the reticulo-endothelialsystem. The degree of tissue contamination one week after infection of both 24- and 40-week-old hens was similar, suggesting that the differences, if any, may be physiological.However, the significance of the difference betweeen frequencies of 0/810 and 2/633 isquestionable.
Although the method employed for egg culture did not involve culturing egg shells andtheir associated membranes separately, the differences in contamination rates for eggcontents and complete homogenized eggs were sufficiently great to suggest clearly thatthe egg surface was the main site of contamination. A number of eggs removed from theoviduct or peritoneal cavity of chickens in Expt 1 were also infected, despite the fact thatnone of the contents of the laid eggs was infected. Thus it is likely that under the variousconditions and routes by which chickens might become infected by S. enteritidis phage
Dow
nloa
ded
by [
117.
211.
89.1
64]
at 0
7:57
12
Oct
ober
201
4

S. ENTERITIDIS IN LAYING HENS 345
type 4, eggs are more likely to become contaminated during passage through the cloacathan as a result of ovarian infection. Oviduct infection appeared to be the result ofhaematogenous spread resulting from the intravenous route of inoculation since it doesnot occur following oral infection. Neither did it appear to result necessarily in eggshaving S. entérinais contaminated contents (Expt 1). This is in contrast to the study ofTimoney et al. (1989) who suggested that infected eggs arose either from infected ovariesor from passage through an infected oviduct. Faecal excretion of Salmonella organismswas not considered to be a major source of infection although this was persistent.
Salmonella organisms on the surface of shell eggs are relatively easily removed eitherduring boiling and some may be removed when the egg is cracked. However, it is wellknown that these organisms can penetrate through the shell (Stokes et al., 1956; Board etal., 1964). S. entérinais organisms can multiply rapidly in the yolk at an appropriatetemperature (Humphrey, 1990). Thus it is unclear whether the large number of surface-contaminated eggs or the much small number of eggs which contain S. entérinais in theircontents would be the major source of infection for man. It is likely that both contributeto infection.
Apart from the liver and spleen, which contain phagocytic cells and therefore clearSalmonella organisms from the blood, S. entérinais also localised in the ovary. The reasonfor this is unclear. S. entérinais phage type 4 is highly virulent for young chickens(Barrow, 1991) and one of the major components of virulence is invasiveness followingoral inoculation (Hinton et al., 1989; Barrow, 1991). However, this microbial character-istic alone does not explain ovarian localization, compared with, for example, themyocardium. The extensive permeability of the vascular endothelia observed in the ovary(Griffin et al., 1984) may contribute to localisation at this site. In most cases in thepresent experiment, S. entérinais was isolated more frequently from the interstitial tissueof the ovary rather than from yolk from the follicles. Thus the infected tissue was eithernot associated with follicle production or perhaps infected follicles do not develop intoeggs. However, the experiments lasted 7 weeks at most. Perhaps prolonging the experi-ments would have increased our chance of producing infected follicles, since every ovarycontains many dozens of ovules.
5. entérinais has previously been isolated from ovaries (Snoeyenbos et al., 1969). Othergroups have found that other Salmonella serotypes, with the exception of 5. pullorum-S.gallinarum can also result in ovarian infections under experimental conditions (Gibbons& Moore, 1946; Gauger & Greaves, 1946; Mundt & Tugwell, 1958; Gordon & Tucker,1965; Forsythe et al., 1967) in some cases following intravenous inoculation. All theseworkers report that the incidence of shell contamination was much higher than that ofSa/mone/Za-contaminated contents. However, discussions over the main site of infectionare of little value unless some control can be exercised over the main route ofcontamination.
If, despite the greater number of surface-contaminated eggs, eggs resulting frominfected ovaries are the major source of infection for man then control either byvaccination or by serological testing followed by slaughter is feasible. Contamination byfaecal material would make control by vaccination more difficult, in view of the generallypoor reductions in faecal excretion by vaccinating chickens against Salmonella (Barrow,1989). However, S. enteritidis is probably sufficiently invasive for a serological test to beuseful to detect infected chickens. The ELISA is a sensitive assay which has already beenused to detect S. enteritidis and S. typhimurium infection in chickens (Barrow et al., 1989;Cooper et al., 1989; Hassan et al, 1990). Serum IgG titres detected by LPS and sonicateantigen increased following infection and stayed at high levels during the experiment.
Dow
nloa
ded
by [
117.
211.
89.1
64]
at 0
7:57
12
Oct
ober
201
4

346 P. A. BARROW & MARGARET A. LOVELL
This suggests that if ELISA are used to detect S. entérinais infection it must be borne inmind that higher titres will reflect that infection has occurred at some time in the pasteven though the birds might not be excreting Salmonella organisms at the time ofsampling. The LPS antigen was more specific than the sonicate. Although this has alsobeen found to be the case for S. typhimurium the sonicate for the serotype alsodemonstrates considerable specificity (Hassan et al., 1990). Such ELISA tests, made morespecific by including additional antigens such as flagella antigens into the assay, could bein conjunction with bacteriological methods to improve our understanding and control ofS. entérinais infections.
ACKNOWLEDGEMENT
The authors wish to acknowledge Mrs K. Murphy for excellent technical assistance andMiss V. Peters and Mr P. Roderick for assistance in different ways.
REFERENCES
BARROW, P.A. (1989). Salmonellosis—prospects for microbiological control in poultry, Avian Pathology, 18, 557-561.BARROW, P.A. (1991). Experimental infection of chickens with Salmonella enteritidis, Avian Pathology, 20, 145-153.BARROW, P.A., HASSAN, J.O., MOCKETT, A.P.A. & MCLEOD, S. (1989). Detection of salmonella infection by ELISA,
Veterinary Record, 125, 586.BOARD, R.G. (1964). The growth of Gram-negative bacteria in the hens egg, Journal of Applied Bacteriology, 27,
350-364.BYGRAVE, A.C. & GALLAGHER, J. (1989). Transmission of Salmonella enteritidis in poultry, Veterinary Record, 125, 571.CHART, H., BASKERVILLE, A., HUMPHREY, T.J. & ROWE, B. (1990). Serological response of chickens to Salmonella
enteritidis infections, Epidemiology and Infection, 104, 63-72.COOPER, G.L., NICHOLAS, R.A. & BRACEWELL, C.D. (1989). Serological and bacteriological investigations from flocks
naturally infected with Salmonella enteritidis, Veterinary Record, 125, 567-572.COYLE, E.F., PALMER, S.R., RIEEIRO, C.D., JONES, H.I., HOWARD, A.J., WARD, L. & ROWE, B. (1988). Salmonella
enteritidis phage type 4 infection: association with hens eggs, Lancet, II, 1295-1297.FADDOUL, G.P. & FELLOWS, G.W. (1966). A five year study of the incidence of Salmonellae in avian species, Avian
Diseases, 10, 296-304.FORSYTHE, R.H., Ross, W.J. & AYRES, J.C. (1967). Salmonellae recovery following gastrointestinal and ovarian
inoculation in the domestic fowl, Poultry Science, 46, 849-855.GAUGER, H.C. & GREAVES, R.E. (1946). Isolation of Salmonella typhimurium from the faeces of turkeys, Poultry
Science, 25, 232-235.GIBBONS, N.E. & MOORE, R.L. (1946). A note on aritifically infected fowl as carriers of Salmonella, Poultry Science, 25,
115-118.GORDON, R.F. & TUCKER, J.F. (1965). The epizootiology of Salmonella menston infection of fowls and the effects of
feeding poultry food artificially infected with Salmonella, British Poultry Science, 6, 251-264.GRIFFIN, H.D., PERRY, M.M. & GILBERT, A.B. (1984). Yolk formation, in: FREEMAN, B.M. (ed.), Physiology and
Biochemistry of the Domestic Fowl, pp. 345-378. Academic Press.HASSAN, J.O., BARROW, P.A., MOCKETT, A.P.A. & MCLEOD, S. (1990). Antibody response to experimental Salmonella
typhimurium infection in chickens measured by ELISA, Veterinary Record, 126, 519-522.HINTON, M.H., PEARSON, G.R., THRELFALL, E.J., ROWE, R., WOODWARD, M. & WRAY, C. (1989). Experimental
Salmonella enteritidis infection in chickens, Veterinary Record, 124, 223.HOPPER, S.A. & MAWER, S. (1988). Salmonella enteritidis in a commercial layer flock, Veterinary Record, 123, 351.HUMPHREY, T.J. (1990). The growth of Salmonella in the contents of intact shell eggs: the influence of storage
temperature, Veterinary Record, 126, 292.HUMPHREY, T.J., BASKERVILLE, A., CHART, H. & ROWE, B. (1989). Infection of egg laying hens with Salmonella
enteritidis PT4 by oral inoculation, Veterinary Record, 125, 531-532.HUMPHREY, T.J., CRUIKSHANK, J.C. & ROWE, B. (1989). Salmonella enteritidis phage type 4 and hens eggs, Lancet, i,
281.LISTER, S.A. (1988). Salmonella etertitidis infection in broilers and broiler breeders, Veterinary Record, 123, 350.MAWER, S.L., SPAIN, G.E. & ROWE, B. (1989). Salmonella enteritidis phage type 4 and hens eggs, Lancet, i, 280-281.MILES, A.A., MISRA, S.S. & IRWIN, J.O. (1938). The estimation of the bactericidal power of the blood, Journal of
Hygiene, Cambridge, 38, 732-749.MUNDT, J.O. & TUGWELL, R.L. (1958). The relationship of the chicken egg to selected paratyphoids, Poultry Science,
37, 415-420.
Dow
nloa
ded
by [
117.
211.
89.1
64]
at 0
7:57
12
Oct
ober
201
4

S. ENTERITIDIS IN LAYING HENS 347
REPORT (1989). Salmonella in eggs, Report and Proceedings of the House of Commons Agricultural Committee. London:Her Majesty's Stationery Office.
SMITH, H.W. & TUCKER, J.F. (1975). The effect of antibiotic therapy on the faecal excretion of Salmonellatyphimurium by experimentally infected chickens, Journal of Hygiene, Cambridge, 75, 275-292.
SNOEYENBOS, G.H., SMYSER, C.F. & VAN ROEKEL, H. (1989). Salmonella infections of the ovary and peritoneum of thechicken, Avian Diseases, 13, 668-670.
ST. LOUIS, M.E., MORSE, D.L., POTTER, M.E., DEMELFI, T.M., GUZEWICH, J.J., TAUXE, R.V., BLAKE, P.A. & THESALMONELLA ENTERITIDIS WORKING GROUP (1989). The emergence of grade A eggs as a major source ofSalmonella enteritidis infections, Journal of the American Medical Association, 259, 2103-2107.
STOKES, J.L., OSBORNE, W.W. & BAYNE, H.G. (1956). Penetration and growth of Salmonellae in shell eggs, Journal ofFood Science, 21, 510-518.
TIMONEY, J.F., SHIVAPRASAD, H.L., BAKER, R.C. & ROWE, B. (1989). Egg transmission after infection of hens withSalmonella enteritidis phage type 4, Veterinary Record, 125, 600-601.
RESUME
Infection expérimentale de pondeuses avec Salmonella enteritidis
Des poules ont été inoculées par voie intraveineuse avec 105 ou 106 organismes de S. enteritidis. Uneinfection importante des ovaires est survenue et a persisté pendant plusieurs semaines. La plupart desinfections ovariennes était localisée aux tissus interstitiels et non dans le vitellus contenu dans de largesfollicules. L'infection de l'ovaire ne résulte pas d'une contamination des sacs aériens. Aucun des 810oeufs pondus n'a révélé la présence de S. enteritidis.
Les poulets infectés oralement ont donné des résultats similaires à ceux inoculés par voie intraveineusebien que la nombre d'isolements positifs à partir du caecum et du cloaque ait été plus élevé. S. enteritidisa été isolé de deux des 633 oeufs pour lesquels le contenu seulement a été mis en culture et de 36 des 614oeufs pour lesquels à la fois la coquille et le contenu ont été mis en culture. La réponse sérique enanticorps IgG suivant l'inoculation orale a été recherchée par la technique ELISA utilisant la celluleentière ultrasonnée ou l'antigène lipo-polysaccharide. Des titres élevés d'IgG ont été décelés pendant 27semaines après l'infection correspondant à la fin de l'essai.
Ces expériences suggèrent que la plupart des oeufs pondus, infectés par S. enteritidis, sont le résultatd'une contamination de surface plutôt que d'une contamination ovarienne.
ZUSAMMENFASSUNG
Experimentelle Infektion legender Hennen mit Salmonella enteritidis vomPhagen-Typ 4
Hühner wurden intravenös mit 105 oder 106 Keimen von S. enteritidis inokuliert. Es kam zu einerschweren Infektion der Eierstöcke, die in einigen Fällen mehrere Wochen lang persistierte. Die meistenEierstocksinfektionen waren auf die interstitiellen Gewebe beschränkt und erstreckten sich nicht auf denDotter in den großen Follikeln. Infektionen des Eierstocks waren nicht die Folge einer Kontaminationdurch infizierte Luftsäcke. Kein der 810 gelegten Eier enthielt S. enteritidis.
Oral infizierte Küken lieferten ähnliche Resultate wie die nach intravenöser Infektion, jedoch war dieAnzahl der Erregerisolierungen aus den Blinddärmen und Kloaken höher. S. enteritidis wurde aus zweivon 633 Eiern isoliert, bei denen nur vom Inhalt Kulturen angelegt wurden, und von 36 von 614 Eiern,bei denen sowohl die Schale als auch der Inhalt kultiviert wurden.
Die IgG-Reaktionen im Serum nach oraler Inokulation wurden mit einem ELISA verfolgt, bei demUltraschall-Präparationen ganzer Zellen oder Lipopolysaccharid als Antigen verwendet wurden. HoheIgG-Titer wurden nach der Infektion 27 Wochen lang, bis der Versuch beendet wurde, nachgewiesen.
Die Versuchsergebnisse lassen darauf schließen, daß die meisten der von S. enteritidis-infiziertenHennen gelegten infizierten Eier nur auf der Oberfläche und nicht infolge infizierter Eierstöckekontaminiert sind.
Dow
nloa
ded
by [
117.
211.
89.1
64]
at 0
7:57
12
Oct
ober
201
4

348 P. A. BARROW
RESUMEN
Infeccion experimental de gallinas pondedoras con el fago tipo 4 de Salmonellaenteritidis
Se inocularon pollos intravenosamente con 105 o 10* microorganismos. Se produjo una intensa infecciónen los ovarios, persistiendo la infección en este órgano durante varias semanas. La mayoría de lasinfecciones ováricas estaban confinadas a los tejidos intersticiales y no al vitelo contenido en los folículosgrandes. Les infecciones ováricas no eran resultado de la contaminación de los sacos aéreos infectados.Ninguno de los 810 huevos puestos contenía S. enteritidis.
Los pollos infectados por vía oral nostraron resultados similares a los apreciados tras la inoculaciónintravenosa aunque el número de aislamientos obtenidos del ciego y de la cloaca fueron más elevados. Seaisló S. enteritidis de dos de 633 huevos en los que únicamente el contenido fue cultivado y de 36 sobre614 huevos en los que tanto la cáscara como el contenido fueron cultivados.
Le respuesta de IgG sérica a la inoculación oral fue estudiada mediante ELISA empleando célulascompletas sonicadas o antígeno lipopolisacárido. Se detectaran titulos elevados de IgG durante 27semanas tras la infección, período de duración del experimento.
Estos resultados sugieren que la mayoría de los heuvos infectados puestos por aves infectadas por S.enteritidis están contaminados en la superficie en vez de ser el resultado de una infección ovárica.
Dow
nloa
ded
by [
117.
211.
89.1
64]
at 0
7:57
12
Oct
ober
201
4
Copyright © 2022 FDOKUMEN




















