
PROTEOLYTIC ACTIVITY OF LACTOBACILLUS SAKEI, LACTOBACILLUS FARCIMINIS AND LACTOBACILLUS PLANTARUM ON...
18
PROTEOLYTIC ACTIVITY OF LACTOBACILLUS SAKEI, LACTOBACILLUS FARCIMINIS AND LACTOBACILLUS PLANTARUM ON SARCOPLASMIC PROTEINS OF PORK LEAN ANNA LISA BASSO', GIANLUCA PICAMELL@, RAFFAELE COPPOLAf3, and ALDO DI LUCCIA2* ' PATRIZIO TREMONTE~, SALVATORE SPAGNA wss04 'Istitutoper il Sistema Produzione Animale in Ambiente Mediterraneo (ISPAAM) CNR Via Argine I085 801 4 7 Napoli, Italy 21stituto di Scienze dell 'Alimentazione(ISA)-CNR Via Roma 52 A/C 83100 Avellino, Italy 'Dipartimento di Scienze e Tecnologie Agro-Alimentari Ambientali e Microbiologiche Universit2degli Studi del Molise Via De Sanctis 861 00 Campobasso, Italy 'Dipartimento di Scienza degli Alimenti Universita degli Studi di Napoli Federico II Parco Gussone 80055 Portici (Napoli), Italy 'Dipartimento di Produzione Animale Universitbdegli Studi di Bari Via Amendola I6YA 70126 Bari, Italy Received for Publication April 30,2003 Accepted for Publication October 17,2003 ABSTRACT The aim of this snu;iy was to assess the proteolytic activity of Lactobacillus sakei (DSM 6333). L. plantarum (B21). and to a lesser extent, L. farciminis (DSM 201 84) on meat sarcoplasmic proteins. The protein composition was assayed by sodium dodecyl sulfate-polyaciylamide gel electrophoresis (SDS- PAGE) and capillary electrophoresis after incubation of meat extract 'Author to whom correspondence should be add&. Prof. Aldo Di Luccia, Institute of Food Science - National Research Council (CNR), Via Roma 52 N C , 83100 Avellino, Italy. TEL: +39 (0)825 2991 1 1 ; FAX +39 (OM25 781 585; EMAIL [email protected] Journal of Food Biochemistry 28 (2004) 195-212. AN Rights Reserwd. OCopyrighi 2004 by Food & Nutrition Press, Inc.. Trumbull, Connecticut. 195
Transcript of PROTEOLYTIC ACTIVITY OF LACTOBACILLUS SAKEI, LACTOBACILLUS FARCIMINIS AND LACTOBACILLUS PLANTARUM ON...

PROTEOLYTIC ACTIVITY OF LACTOBACILLUS SAKEI, LACTOBACILL US FARCIMINIS AND LACTOBACILLUS
PLANTARUM ON SARCOPLASMIC PROTEINS OF PORK LEAN
ANNA LISA BASSO', GIANLUCA PICAMELL@, RAFFAELE COPPOLAf3,
and ALDO DI LUCCIA2* ' PATRIZIO TREMONTE~, SALVATORE SPAGNA w s s 0 4
'Istituto per il Sistema Produzione Animale in Ambiente Mediterraneo (ISPAAM) CNR Via Argine I085
801 4 7 Napoli, Italy
21stituto di Scienze dell 'Alimentazione (ISA)-CNR Via Roma 52 A/C
83100 Avellino, Italy
'Dipartimento di Scienze e Tecnologie Agro-Alimentari Ambientali e Microbiologiche
Universit2 degli Studi del Molise Via De Sanctis
861 00 Campobasso, Italy
'Dipartimento di Scienza degli Alimenti Universita degli Studi di Napoli Federico II
Parco Gussone 80055 Portici (Napoli), Italy
'Dipartimento di Produzione Animale Universitb degli Studi di Bari
Via Amendola I6YA 70126 Bari, Italy
Received for Publication April 30,2003 Accepted for Publication October 17,2003
ABSTRACT
The aim of this snu;iy was to assess the proteolytic activity of Lactobacillus sakei (DSM 6333). L. plantarum (B21). and to a lesser extent, L. farciminis (DSM 201 84) on meat sarcoplasmic proteins. The protein composition was assayed by sodium dodecyl sulfate-polyaciylamide gel electrophoresis (SDS- PAGE) and capillary electrophoresis after incubation of meat extract
'Author to whom correspondence should be add&. Prof. Aldo Di Luccia, Institute of Food Science - National Research Council (CNR), Via Roma 52 NC, 83100 Avellino, Italy. TEL: +39 (0)825 2991 1 1 ; FAX +39 (OM25 781 585; EMAIL [email protected]
Journal of Food Biochemistry 28 (2004) 195-212. AN Rights Reserwd. OCopyrighi 2004 by Food & Nutrition Press, Inc.. Trumbull, Connecticut. 195

196 A.L. BASSO ETAL.
inoculated with bacteria. AN strains showed proteolytic activity: a band about 94 kDa disappeared in samples inoculated with L. farcimins and L. plantarum and strongly decreased in those inoculated with L. sakei. The intensity of the bands with a molecular weight between 94 and 38 kDa decreased in all samples. Capillary electrophoresis analysis ascertained the disappearance of thefiactions corresponding to 8.64 and 8.66 min retention time in all samples. The bands corresponding to 94 kDa and 38 kDa were, respectively, identified as glycogen phosphorylase muscle isoform and glyceraldehyde 3-phosphate dehydrogenase, by in s ib digestion of protein gel bands and peptide map analysis using Matrix Assisted Laser Desorption/Ionization - Time of Flight Mass Spectrometry (MALDI-TOF MS).
INTRODUCTION
In manufacturing fermented sausages, the tenderized meat is minced and blended with other ingredients and a fermentation process takes place with the onset of bacterial activities. Amongst the natural microflora, Lactic Acid Bacteria (LAB) play an important role: they ferment sugars to acid, lower pH and improving the texture of the product and stability against the proliferation of food pathogens and produce certain aromatic compounds (Kelly et al. 1996; Coppola et al. 1998; Klaenhammer 1998; Luongo et al. 2001).
Many microorganisms are present during the overall process. L. sakei, L. curvatus and L. plantarum (Hammes et al. 1990; Coppola et al. 1998), are the most frequently found LAB in such products. The presence of L. casei, L. farciminis and L. alimentarim is also described (Samelis et al. 1994; Hammes et al. 1990; Coppola et al. 1998). Their role in protein degradation is still not fully understood. Hierro et al. (1999) found that myofibrillar proteins are hydrolysed by endogenous muscle proteases. Other experimental works using antibiotics or inhibitors of proteases confirmed that bacterial peptidases are involved almost exclusively in the degradation of peptides deriving from hydrolysis by endogenous enzyme activity (Molly et al. 1997). Nevertheless, the addition of a serine protease extracted fiom a L. paracasei subsp. paracasei strain accelerates sausage ripening (Hagen et al. 1996). Moreover, recent studies have demonstrated that the most commonly found Lactobacillm species in dry fermented meats, e.g., L. sakei, L. curvatus, L. plantarum and L. casei, are able to hydrolyse myofibrillar and sarcoplasmic muscle proteins in vitro (Fadda et al. 1999a, b; Sanz and Toldrii 2001). Proteolysis observed after incubation of the protein extract with the whole cells suggested the existence of enzymes bound to the cell wall, as it is typical for LAB investigated to date (Kunji et al. 1996; Fad& et al. 1999b).

PROTEOLYSIS OF SARCOPLASMIC PROTEINS I97
The hydrophylic peptides derived from the hydrolybc activity of some species on the water-soluble proteins play an important role in determining the aroma (Fadda et al. 1999b). Many peptidases have been isolated from lactobacilli utilized in meat fermentation. A tripeptidase, a broad-specificity aminopeptidase, a dipeptidase, an X-prolyl-dipeptidyl peptidase, an arginine aminopeptidase have been purified from L. sukei (Monte1 et al. 1995; Sanz et al. 1998,1999; Sanz and TolM 1997% b); a prolinase has been characterized from L. curvatus (Magboul and McSweeney 1997,1998).
The aim of the present work was two-fold to detect possible specificity of LAB species in the hydrolysis of meat sarcoplasmic proteins and to contribute to the knowledge of the roles played by endogenous tissue enzymes or by microorganisms. Our final goal was to identify which proteins are preferentially degraded by bacterial proteases. Hence, we compared the hydrolyhc capacity of three species of LAB (L. saki DSM 6333, L. farciminis DSM 20184 and L. pluntumm B21) on sarcoplasmic proteins h m pig muscle by means of SDS- PAGE and Free-phase Capillary Zone Electrophoresis (CZE).
MATERIALS AND METHODS
Preparation of Srrcoplasmic Proteins
Muscle samples were taken from Longissirnus dorsi immediateiy after exsanguination and stored at - 80C until the time of protein extraction.
In the first step, all samples were cleaned by removing fat and connective tissue from muscle tissue by using scalpels. Fitly grams of the product were resuspended in 50 mL of milli Q water and homogenized (homogenizer, PBI International, Italy) for 5 min at OC in an ice bath. The homogenized tissue was centrifuged at 1,200 g for 30 min at 4C (centrifuge, Minifuge RF Sepatech Heraeus Instr., Hanau, D). The collected supernatants were centrifuged again at 15000 g for 15 min at OC (centrifuge, Sorvall RMC 14, DuPont, Newtown, CT). The obtained Supernatant was then filtered (filters, 0.45 pm, Millipore, Ireland) and the protein concentration of the filtrate was determined by the method of Bradford (1976) using bovine serum albumin as standard.
Four hundred microliters of the filtrate were concentrated in a Microcon 3 mim-concentrator (Amicon, Beverly, MA) by centrifugation at 10,000 g for 20 min at 1OC.

198 A.L. BASSO ETAL.
Bacterial Strains and Growth Conditions
The following strains were used L. sakei DSM 6333, L. furciminis DSM 20184 and L. pIanturum B21 (Ciafardini et ul. 1994). Cells were grown in MRS broth at 30C and stored in 15% glycerol at a temperature of -8OC. To determine proteolytic activity on sarcoplasmic proteins, the preparation was inoculated with bacteria cultures incubated for 14 h at 30C.
Proteolytic Activity on Sarcoplasmic Proteins
The cells were centrifuged at 3,000 g for 15 min at 4C (Minihge RF Sepatech Heraeus), washed in 0.9% NaCl solution and resuspended in the same diluent. The suspension was added to the sarcoplasmic protein extract (filtered through 0.2 pm filters, Millipore) in such a way to obtain a final optical density of 0.1 at 600 nm. The mixture was subsequently incubated for 48 h at 30C; extract without cells was incubated as a control.
Polyacrylamide Gel Electrophoresis (SDS-PAGE)
The proteolytic actvity on sarcoplasmic proteins was evaluated by electrophoresis at alkaline pH in the presence of sodium dodecyl sulphate (SDS) (Laemmli 1970). The hctionated proteins were revealed through use of Comassie brilliant blue R250. The estimated molecular weight was obtained by using standard proteins having molecular masses ranging from 94 to 14 kDa.
Free-Phase Capillary Zone Electrophoresis (CZE)
The extracts after bacteria-mediated proteolysis were analysed by Capillary Zone Electrophoresis using an instrument mod. P/ACE 2000 equipped with software GOLD (Beckman htr . , Fullerton, CA). Fifty nL of samples were injected in a uncoated silica capillary of 75 pm x 50 cm with high pressure for 1 s. Assays were carried out at a constant temperature of 25C using phosphate buffer of 40 mM pH 2.5 at a steady 12 kV for 20-25 min.
In-Gel Digestion of Excised Protein Spots
Manually excised spots from Coomassie stained gels of aqueous meat extracts, before and after incubation, were destained, reduced and alkylated by carboxymethylation, and then digested in situ by trypsin (Sigma Chemical Co., St. Louis, MO), overnight at 37C, according to the procedure by Shevchenko et al. (1996).

PROTEOLYSIS OF SARCOPLASMIC PROTEINS 199
Peptides resulting from digestion were extracted in 40 pL of a acetonitrileK% formic acid 111 (v/v) solution and than concentrated by vacuum centrikgation for mass spectrometric analysis.
All steps of the excision procedure, destaining, reductiodalkylation and peptide extraction were carried out under a laminar flow hood to avoid contamination by keratins.
Peptide-Mass Mapping by MALDI-TOF MS
Mass spectrometry experiments were carried out on a PerSeptive Biosystems (Framingham, MA) Voyager DE-Pro instrument equipped with a N, laser (3 3 7 nm, 3 nsec pulse width).
One microliter of peptide mixture h m each digested spot was loaded on a stainless steel plate together with 1 pL of matrix (a-ciano-4-hydroxycic acid) prepared by dissolving 10 mg in 1 mL of aqueous 50% acetonitrile containing 0.1% TFA.
Typically, 250 laser pulses were acquired for each spectrum and the mass spectra were acquired in the reflector mode with DE; the accelerating voltage was 20 kV.
External mass calibration was performed with low-mass peptide standards.
Protein Identification
The measured molecular mass of tryptic peptides were used as inputs to search the NCBI and Swiss-Prot databases.
All searches were carried out by using the PROTEIN PROSPECTOR/MS- FIT software developed at the University of California, San Francisco (http://prospector.ucsf.edu) and the searches were restricted to mammalian species proteins.
The allowed protein molecular mass range was chosen by referring to electrophoretic MW standards or fi-om 10,OOO to 200,000 Da. Isoelectric points were allowed to range from 3.0 to 10.0, and oxidation of Met was included as a possible side reaction.
Up to two missed tryptic cleavages were considered, and a conservative mass accuracy o f f 0.4 Da was used for all tryptic-mass searches.

200 A.L. BASSO ETAL.
RESULTS AND DISCUSSION
The electrophoretic patterns of sarcoplasmic proteins demonstrated the proteolyhc capability of L. farciminis and confiied that of L. sakei and L. plantarum as previously reported (Fadda et al. 1999 a, b).
In the first step of our investigation, the aqueous extract of the muscle proteins was inoculated with three Lactobacillus strains (L. sakei DSM 6333, L. farciminis DSM 20184 and L. plantarum B21). This allowed us to establish which sarcoplasmic components are more prone to lactobacilli proteolytic activity. The three species investigated, including L. plantarum B21, isolated h m a nonmeat food, exhibited the same substrate specificity. This may point to the presence of a cell-wall protease, common to LAB or at least to the Lactobacillus species. After only 48 h, electrophoretic analysis on the sarcoplasmic proteins were carried out to identify which components of this hction were most susceptible to hydrolyhc action. Figure 1 shows the SDS- PAGE electrophoretic patterns relative to the separation of sarcoplasmic proteins fiom pork lean before and after incubation. Lane 2 shows aqueous extract of meat proteins before incubation, while lane 3 shows the same aqueous protein extract after incubation at 30C for 48 h, but without any inoculation. In lanes 5 and 6, corresponding to aqueous extract incubation with L. plantarum and L. farciminis, respectively, the total disappearance of the protein band at an estimated molecular weight of 94 kDa can be observed. Lane 4, relating to incubation with L. sakei shows a vary faint band. The band of about 38 kDa decreased sharply in intensity for all three strains. The two bands with molecular weight slightly higher than 45 kDa, indicated as “x” and “y” in Fig. 1, are not well separated; however, it is worth of note that the intensity of the band marked “x” strongly decreased in all three strain patterns whereas the band marked “y” decreased only in the pattern of L. plantamm B2 1. In this case the poor resolution of the SDS-PAGE gel has made difficult to cut the bands “x” and “y” for their exact identification. These two bands could correspond to phosphoglucose isomerase (49 m a ) and enolase (46 kDa) as identified by Martin et al. (2001).
The differences observed in proteolytic activity of our strains (L. farciminis DSM 20184, L. plantarum B21 and L. sakei DSM 6333) highlight the influence of microbial composition on meat product characteristics. On increasing the incubation time to 96h the disappearance of proteins with molecular weights of 97,45,43,37 and 26 kDa in samples inoculated with L. sakei and L. plantarum has been reported by Fadda et al. (1999a, b). In agreement with these latter results, we have ascertained the disappearance of four sarcoplasmic proteins (94, 38 kDa and two bands hgher than 45 ma). Analysis by capillary electrophoresis confirmed that the strains use the same proteins in the first
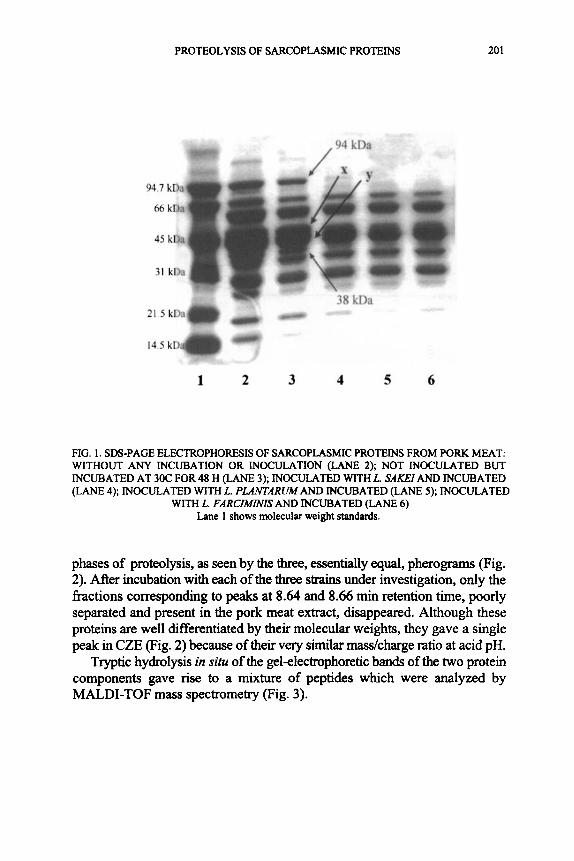
PROTEOLYSIS OF SARCOPLASMIC PROTEINS 20 1
FIG. 1. SDS-PAGE ELECTROPHORESIS OF SARCOPLASMIC PROTEINS FROM PORK MEAT: WITHOUT ANY INCUBATION OR INOCULATION (LANE 2); NOT INOCULATED BUT INCUBATED AT 30C FOR 48 H (LANE 3); INOCULATED WITH L. SAKEI AND INCUBATED (LANE 4); INOCULATED WITH L. PLANTARUM AND INCUBATED (LANE 5); INOCULATED
WITH L. FARCZMINIS AND INCUBATED (LANE 6) Lane 1 shows molecular weight standards.
phases of proteolysis, as seen by the h e , essentially equal, pherograms (Fig. 2). After incubation with each of the three strains under investigation, only the fractions corresponding to peaks at 8.64 and 8.66 min retention time, poorly separated and present in the pork meat extract, disappeared. Although these proteins are well differentiated by their molecular weights, they gave a single peak in CZE (Fig. 2) because of their very similar masdcharge ratio at acid pH.
Tryptic hydrolysis in situ of the gelelectrophoretic bands of the two protein components gave rise to a mixture of peptides which were analyzed by MALDI-TOF mass spectrometry (Fig. 3).

202
1
A.L. BASSO ET AL.
1 x U L YVL
h 16.1 s
h s.
.- E +-
0002
00'01
' 000
.- B - 000t
00'01
.OO'O

PROTEOLYSIS OF SARCOPLASMIC PROTEINS 203
.- E 4-
00'02
00'01
00'0
0
E 4-
WOZ
M'OI
00-0
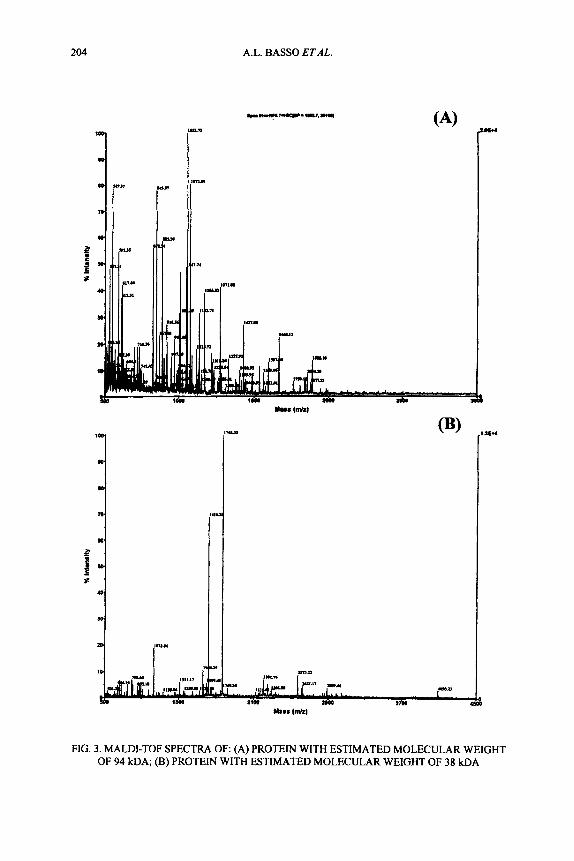
204 A.L. BASSO ETAL.
J YUI
FIG. 3. MALDI-TOF SPECTRA OF: (A) PROTEIN WITH ESTIMATED MOLECULAR WEIGHT OF 94 kDA; (B) PROTEIN WITH ESTIMATED MOLECULAR WEIGHT OF 38 kDA

PROTEOLYSIS OF SARCOPLASMIC PROTEINS 205
The peptide mixtures derived from tryptic digest were used in NCBI database research using Protein Prospector software developed at the University of California, San Francisco. As a result, the protein with an estimated weight of 94 kDa was identified as glycogen phosphorylase-muscle isoform (myophosphorylase) (PHS2 - P.A.N. Swiss-Prot: P11217), an enzymatic isoform of muscle glycolysis, and the protein having an estimated molecular weight of 38 kDa as glyceraldehyde 3- phosphate dehydrogenase (G3PD - P.A.N. Swiss-Prot: P00355) from pig, an isofom of muscle glycolysis as well. Since neither the primary structure, nor the cDNA sequence from pig are included in the database, the rnyophosphorylase was identified by alignment with the sequence of the human homologous protein.
Table 1 shows the molecular mass of the single peptides compared to theoretical figures, whereby a high degree of matching can be observed. This, therefore, ensures an accurate identification of myophosphorylase, which has, for the human homologous protein, a molecular weight of 97027.06.
Table 2 reports the analogous results obtained for the protein with an estimated molecular weight of 38 kDa, which was identified as glyceraldehyde 3- phosphate dehydrogenase, in this case from pig, since the primary sequence is present in the database (molecular weight 35704.85 Da).
By identifylng these proteins we were able to verify the masdcharge ratios responsible for electrophoretic mobility, resulting in 759.6 for G3PD and 735.5 for PHS2. In fact, the slight difference in electrophoretic mobility deriving h m these similar ratios was not detectable in our experimental conditions.
PHS2 and G3PD are cytoplasmatic enzymes: the first catalyses hydrolysis of glycosidic residues h m the nonreducing end of glycogen molecules and the second catalyses oxidation of phospho-glyceraldehyde to acyl-phosphate during glycolysis. These enzymes are present in liver and muscle. The latter is the main ingredient used in the preparation of sausages, which at the end of ripening are particularly rich in valine, methionine and alanine (Dierick et al. 1974). Leucine and alanine residues are present at high levels in PHS2 and G3PD, as they make up 17% and 15% of the total amino acids of PHS2 and G3PD, respectively. Previous works showed that when sarcoplasmic proteins from pork lean are incubated with bacterial cell extracts or with whole cells, a strong increase in alanine, leucine and glutamic acid is observed (Verplaetse 1994; Fadda et al. 1999b; S a m and ToldrA 2001). Moreover, all proteolytx enzymes purified fiom L. saki , the bacterial species most responsible for meat fermentation, hydrolyze especially leucine- and alanine-containing substrates ( S a m et al. 1999; Sanz and Toldri 1997a, b; Molly et al. 1997).

TAB
LE I
.
(AV
ERA
GE
MA
SS):
9706
2.06
; Mw
(MO
NO
ISO
TOPI
C M
ASS
): 97
030.
62
N 0
OI
GLY
CO
GEN
PH
OSP
HO
RY
LASE
-MU
SCLE
ISO
FORM
(PH
SZ - P
.A.N
. SWISS-PROT P
1121
7 FO
R H
UM
AN
). TH
EORE
TICA
L PI
: 6.5
7 ; M
W
mea
sure
d m
/z (
Mm
549.
38
550.
39
59 1.
24
61 1.
32
617.
32
627.
33
704.
27
822.
40
845.
33
846.
41 (*
) 88
5.33
9 91
6.45
94
5.41
94
7.44
1001
.49
1007
.43 (
*)
1047
.56
1053
.54 (
*)
1072
.52 (
*)
1085
.60
994.
53 (*
)
teor
ical
m/z
(M
H3
549.
3 1
550.
27
59 1.
29
61 1.
29
I 62
7.38
70
4.35
82
2.45
84
5.46
84
6.45
I
9 16.
48
945.
46
947.
47
994.
58
1001
.56
1007
.47
1047
.59
1053
.57
1072
.54
I
AM
(%
)
+ 0.
013
+ 0.
022
- 0.0
08
+ 0.
005
I - 0
.008
- 0
.01 1
- 0
.006
- 0
.015
- 0
.005
1
- 0.0
03
- 0.0
05
- 0.0
03
-0.0
05
- 0.0
06
- 0.0
04
- 0.0
03
- 0.0
04
-0.0
02
I
posi
tion
in
iden
tifie
d pr
otei
n 41
1-41
4 3 1
-34
797-
800
8 12-
816
I 73
6.74
0 71
6-72
1 8 1
7-82
3 57
1-57
6 36
5-37
1 I
27 1-
278
610-
61 8
236-
243
35-4
2 5 7
0-57
6 3 1
7-32
4
788-
796
643-
650
824-
832
I
miss
ed
clav
age
0 0 0 0 I 0 0 0 1 0 I 0 0 0 0 2 1 1 0 0 I
pept
ide
sequ
ence
FLN
R
NFN
R
EWTR
FS
SDR
no
mat
ch
IPEL
R
VED
VD
K
TIA
QY
AR
IH
EYK
R
AW
DV
TVR
no
mat
ch
NLA
ENIS
R
AA
PGY
HM
AK
N
NV
VN
TMR
H
LHFT
LVK
R
IHEY
KR
FG
CR
DPV
R; C
ys 3
19
carb
oxym
ethy
late
d V
SALY
KN
PR
VIF
LEN
Y R
EIW
GV
EPSR
no
mat
ch
?
!-
h
Y L
P

11 17
.56 (
*)
1 166
.62
1218
.67
1229
.70
1380
.66 (
*)
1414
.69
1426
.79 (
*)
1442
.72
1558
.80
1593
.88
1623
.85
1662
.89 (
*)
1757
.92
1837
.92
1840
.95
1875
.97
1886
.93 (
*)
1889
.03
1992
.02 (
*)
1994
.05
2044
.15
2184
.13
2307
.35
2384
.32
2448
.29
11 17
.59
I 12
18.6
5 12
29.6
7 13
80.6
5 14
14.7
1 14
26.7
8 14
42.6
9 I
1593
.80
1623
.88
I 17
57.9
4
1837
.97
1840
.92
1875
.97
1886
.90
1889
.02
1991
.99
1994
.02
2044
.09
2184
.11
2307
.15
2384
.20
2448
.19(
*)
- 0.0
03
I +
0.00
2 +
0.00
2 +
0.00
1 - 0
.001
+
0.00
1 +
0.00
2 I
+ 0.
005
- 0.0
02
1 - 0
.001
- 0.0
03
+ 0.
002
0.00
0 +
0.00
2 +0.0005
+ 0.
001
+ 0.
001
+ 0.
003
+ 0,
001
+ 0.
009
+ 0.
005
+ 0.
004
415-
425
I 42
9-43
9 18
-29
761-
771
8 12-
823
400-
4 10
279-
290
I 78
4-79
6
556-
569
I 57
7-59
0
522-
537
279-
293
8 17-
832
726-
740
18-3
4 44
0-45
8
45-6
1 55
3-56
9 46
7-48
3 2 1
6-23
5 75
5-77
3 65
7-68
1
1 I 1 0 0 1 0 0 I 2 0 I 0
VA
AA
FPG
DV
DR
no
mat
ch
MSL
VEE
GA
VK
G
LAG
VEN
VTE
LK
DIV
NM
LMH
HD
R
FSSD
RTI
AQ
YA
R
HLQ
IIY
EIN
QR
V
LY
PND
NFF
EGK
no
mat
ch
z ca
rbox
ymet
hyla
ted
5 C
QEK
VSA
LYK
NPR
; Cys
784
3
VH
INPN
SLFD
DIQ
VK
E !2 E
TIA
QY
AR
EIW
GV
EPSR
E
GY
NA
QEY
YD
RIP
ELR
ii
INM
AH
LCIA
GSH
AV
NG
GV
AR
3
no m
atch
Q
LLN
CLH
VIT
LYN
R C
ys 5
81
carb
oxym
ethy
late
d LL
SFV
DD
EAFI
RD
VA
K
VLY
PND
NFF
EGK
ELR
v) > F E
GLA
GV
ENV
TELK
KN
FNR
Cys
446
ca
rbox
ymet
hyla
ted
NV
ATP
RD
Y Y
FALA
HTV
R
EY K
VH
INPN
SLFD
IQV
K
TIFK
DFY
ELEP
HK
QN
K
WV
DTQ
VV
LAM
PYD
TPV
PGY
R
2 v)
QPD
LFK
DIV
NM
LMH
HD
RFK
N
VIP
AA
DLS
EQIS
TAG
TEA
SGTG
NM
K
s (*
) Pea
ks u
sed
for d
atab
ase s
earc
h

TAB
LE 2
.
THEO
RET
ICA
L PI
: 8.5
2; M
w (A
VER
AG
E M
ASS
): 35
704.
85; M
w (M
ON
OIS
OTO
PIC
MA
SS):
3568
2.23
G
LYC
ERA
LDEH
YD
E 3-
PHO
SPH
ATE
DEH
YD
RO
GEN
ASE
SW
ISS-
PRO
T: (G
3P P
IG) P
.A.N
.: PO0355
mea
sure
d m
/z
teor
ical
m/z
AM
po
sitio
n in
m
isse
d se
quen
ce
(Ma
(MH
3 (%
) id
entif
ied
clav
age
643.
44
643.
41
-+ 0
.005
64
-69
0 LV
lNG
K
795.
48 (*
) 79
5.42
+
0.00
7 22
5-23
1 0
LTG
MA
FR
802.
43
802.
55
- 0.0
1 19
2-19
7 1
LWRD
GR
805.
57
805.
43
+ 0.
017
3-10
0
VG
VN
GFG
R 89
3.14
I
I I
I no
mat
ch
909.
45 (
*)
909.
52
- 0.0
08
105-
1 14
1 A
GA
HLK
GG
AK
97
7.70
(*)
977.
54
+ 0.0
16
70-7
7 0
AIT
IFQ
ER
1032
.59
1031
.64
- 0.00
5 3-
13
1 V
KV
GV
NG
FGR
1062
.60
1062
.63
- 0.0
03
105-
115
2 A
GA
HLK
GG
AK
R 13
58.8
8 13
58.6
8 +
0.01
5 32
1-33
2 1
VV
DLM
VH
MA
SKE
1558
.24
(*)
1557
.79
-+ 0.0
29
232-
245
0 V
PTPN
VSV
VD
LTC
R;
1615
.48
(*)
1615
.88
- 0.0
25
70-8
3 1
AIT
IFQ
ERD
PAN
IK
1646
.36
1645
.91
-+ 0
.027
21
7-23
1 1
VIP
ELN
GK
LTG
MA
FR
1763
.45
(*)
1763
.80
- 0.0
20
307.
320
0 LI
SWY
DN
EFG
YSN
R 18
21.4
8 18
2 1.8
7 - 0
.02
1 14
3-1.
59
0 IV
SN A
SCTT
NCL
APL
AK
;
Dro
tein
Cys
244
car
boxy
met
hyla
ted
Cys
149
, 153
carb
oxym
ethy
late
d 22
01.7
3 (*
) I
I 1
I no
mat
ch
2248
.62
(*)
2249
.04
- 0.0
19
84- 1
04
0 W
GD
AG
ATW
VES
TGV
FTTM
EK
2333
.86
2334
.19
- 0.0
14
225-
245
1 LT
GM
AFR
VPT
PNV
S WD
LT
CR
;
2618
.19
2618
.38
- 0.0
07
160-
1 83
0 V
IHD
HFG
IVEG
LMTT
VH
AIT
ATQ
K
2858
.47
2858
.50
- 0.0
01
232-
256
2 VPTPNVVSVVDLTCRLEKPAKYDDJ
78-1
04
0 DPANIKWGDAGATYVVES
TGVF
ITM
4088
.84
+ 0.
034
269-
306
0 G
ILG
Y TE
DQ
VV
SCD
FNSD
THSS
TFD
Cys
244
car
boxy
met
hyla
ted
Cys
244
car
boxy
met
hyla
ted
2887
.46
(*)
2887
.38
+ 0.
003
4090
.22
GA
GIA
L N
DH
FVK
; C
ys 2
81 c
arbo
xym
ethy
late
d
(*) P
eaks
used
for d
atab
ase
sear
ch
h) 0
00
? r hy b h

PROTEOLYSIS OF SARCOPLASMIC PROTEINS 209
This could explain the quick rate of complete hydrolysis of the two enzymes by bacteria-generated proteases during the first 48 h. Taking into account the previously reported protease activity of LAB during cheese ripening, with particular reference to a,,-, 0- and K- caseins (Kunji et al. 1996; Tsakalidou et al. 1999), it is possible to reconfii the hypothesis described above in our work. h o n g LAB used in cheese making, two classes of cell-wall proteases have been identified, both being serine-proteases: type PI and type P, which do not have high specificity for breakdown sites (Kunji et al. 1996). However, it has been established that the preferred sites for p-casein are L165-S166, Q17SK176, Y143-Ql4-4 (Tsakalidou et al. 1999). The oligo-peptides &riving from this fvst hydrolysis are substrates for subsequent action of bacterial endo- and exo-peptidase, some of which have also been found in L. sakei and L. curvatus (Christensen ei al. 1999).
As described for Lactococcus lactis, the expression of cell-wall proteases is regulated by the presence of specific peptides (Marugg et al. 1995) together with intracellular amino-peptidase activities of PepN and PepXP (Meijer et al. 1996). This could explain the negligible variation of the total amount of amino acids in controlled fermentation experiments, as described by Hierro et al. (1999). It is indeed possible that bacterial proteolyt~c enzymes are not active in the presence of preformed substrates, such as low molecular weight peptides generated by hydrolybc activity of endogenous proteases.
Based on these considerations, we identified on the primary structures of PHS2 and G3PD the probable sites for LAB proteolytic enzymes studied thus far. By analyzing the primary structure of PHS2 and G3PD, we can observe that the sites susceptible to attack by LAB proteases, QK, SG and LG, are present in both proteins (Table 3). The existence of a number of sites that are specific for dipeptidases and amino-peptidases from L. sakei, could explain the rapid rate of disappearance of these proteins according to S a m and Told13 (2002).
In conclusion, results obtained show that: (1) LAB are able to hydrolyze prefrnntially some of the proteins which constitute the sarcoplasmic fraction; (2) in the early phases of ripening, at least one enzyme characterized by a high substrate specificity is present in the three species of Lactobacillus studied in this w o k (3) capillary electrophoresis is a reliable analytical tool to detect the proteolybc action of various bacterial strains, as previously shown (Martin et al. 2001). In a relatively simple and accurate way, this technique pennits the monitoring of the pathways of proteolybc action occurring during sausage ripening.

210 A.L. BASSO ETAL
TABLE. 3. THEORETICAL CLEAVAGE SITES IN THE PRIMARY STRUCTURE OF G3PD AND PHS2
Enzyme I P,- P,,-type. X-PDAP* tripeptidase* aminopeptidase* dipeptidase*
G3PD
PHS2
proteinases
' PI- PI,,-type proteinases from lactic acid bacteria (Kunji er nl. 1996) *peptidases from L. snkei (Sanz and Toldri 1997a, 2001; Sanz er 01. 1998; Monte1 ef nl. 1995)
ACKNOWLEDGMENT
The research has been founded by project PRIN 2002 [AttivitA enzimatiche espresse dalle componenti microbiche specifiche implicate nella maturazione dei salumi]. The authors thank Mr. D. Balzarano for his technical assistance. They also thank Prof. Antonio Malorni, head of the Center of Mass Spectrometry of National Research Council (CNR) - Avellino (Italy), where mass spectrometric analysis were carried out.
REFERENCES
BRADFORD, M.M. 1976. A rapid and sensitive method for the quantitation of micrograms quantities of protein utilising the principle of protein-dye binding. Anal. Biochem. 72,248-254.
CHRISTENSEN, J.E., DUDLEY, E.G., PEDERSON, J.A. and STEELE, J.L. 1999. Peptidases and amino acid catabolism in lactic acid bacteria. Antonie van Leeuwenhoek 76,217-246.
CIAFARDINI, G., MARSILIO, V., LANZA, B. and POZZI, N. 1994. Hydrolysis of oleuropein by Lactobacillus plantarum strains associated with olive fermentation. Appl. Environ. Microbiol. 60,4142-4147.
COPPOLA, R., GIAGNACOVO, B., IORJZZO, M. and GRAZIA, L. 1998. Characterization of lactobacilli involved in the ripening of soppressata molisana, a typical southern Italy fermented sausage. Food Microbiol. 15,347-353.
DIERICK, N., VANDECKERKOVE, P. and DEMEYER, D. 1974. Changes in non protein nitrogen compounds during dry sausage ripening. J. Food Sci. 39, 301-304.

PROTEOLYSIS OF SARCOPLASMIC PROTEINS 21 1
FADDA, S., SANZ, Y., VIGNOLO, G., ARISTOY, M.-C., OLIVER, G. and T O L D a , F. 1999a. Hydrolysis of pork muscle sarcoplasmic proteins by Lactobacillus curvatus and Lactobacillus sakei. Appl. Environ. Microbiol. 65, 578-584.
FADDA, S., SANZ, Y., VIGNOLO, G., ARISTOY, M-.C., OLIVER, G. and T O L D a , F. 1999b. Characterization of muscle sarcoplasmic and myofibrillar protein hydrolysis caused by Lactobacillus plantarum. Appl. Environ. Microbiol. 65, 3540- 3546.
HAGEN, B.F., BERDAGUE, J.-L., HOLCK, A.L., NES, H. and BLOM, H. 1996. Bacterial proteinase reduces maturation time of dry fermented sausages. J. Food Sci. 61, 1024-1029.
HAMMES, W.P., BANTLEON, A. and MIN, S. 1990. Lactic acid bacteria in meat fermentation FEMS. Microbiol. Rev. 87, 165-174.
HIERRO, E., DE LA HOLZ, L. and ORDOmZ, J A. 1999. Contribution of the microbial and meat endogenous enzymes to the free amino acid and amine contents of dry fermented sausages. J. Agric. Food Chem. 47, 1156-1 161.
KELLY, W.J., ASMUNDSON, R.V. and HUANG, C.M. 1996. Isolation and characterization of bacteriocin-producing lactic acid bacteria from ready-to-eat food products. Intern. J. Food Microbiol. 33,209-218.
KLAENHAMMER, T.R. 1988. Bacteriocins of lactic acid bacteria. Biochirme
KUNJI, E.R.S., MIERAU, I., HAGTING, A., POOLMAN, B. and KONINGS, W.N. 1996. The proteolytic systems of lactic acid bacteria. Antonie van Leeuwenhoek 70,187-22 1.
LAEMMLI, U.K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T,,. Nature 227,680-685.
LUONGO, D., GIAGNACOVO, B., FIUME, I., IORIZZO, M. and COPPOLA, R. 2001. Volatile Compounds in “soppressata molisana” style salami fermented by Lactobacillus sakei. Ital. J. Food Sci. 13, 19-28.
MAGBOUL, A. and MCSWEENEY, P.L.H. 1997. Study of peptidase activities of Lactobacillus curvatus DPC2024. Irish J. Agnc. Food Res. 36,264-275.
MAGBOUL, A. and MCSWEENEY, P.L.H. 1998. Purification and characterisation of an aminopeptidase fiom Lactobacillus curvatus DPC2024. Aus. J. Dairy Technol. 53, 114.
MARTIN, A., CORDOBA, J.J., RODRIGUEZ, M.M., ”EZ, F. and ASENSIO, M.A. 2001. Evaluation of microbial proteolysis in meat products by capillary electrophoresis. J. Appl. Microbiol. 90, 163- 17 1.
MARUGG, J.D., MEIJER, W., VAN KRANENBURG, R., LAVERMAN, P., BRUINENBERG, P.G. and DE VOS, W.M. 1995. Medium-dependent regulation of proteinase gene expression in Lactococcus lactis: control of transcroption by specific dipeptides. J. Bacteriol. 1 77,2982-2989.
70(3), 337-349.

212 A.L. BASSO ETAL.
MEIJER, W., MARUGO, J.D. and HUGENHOLTZ, J. 1996. Regulation of proteolytic enzyme activity in Lactococcus lactis. Appl. Environ. Microbiol.
MOLLY, K., DEMEYER, D., JOHANSSON, G., RAEMAKERS, M., GHISTELINCK, M. and GEENEN, I. 1997. The importance of meat enzymes in ripening and flavour generation in dry fermented sausages. First results of a European project. Food Chem. 59,539-545.
MONTEL, M.C., SERONIE, M.P., TALON, R. and HEBRAUD, M. 1995. Purification and characterization of a dipeptidase from Lactobacillus sake. Appl. Environ. Microbiol. 61(2), 837-839.
SAMELIS, J., MAUROGENAKIS, F. and METAXOPOULOS, J. 1994. Characterisation of lactic acid bacteria isolated from naturally fermented Greek dry salami. Intern. J. Food Microbiol. 23, 179-196.
T O L D a , F. 1999. Hydrolyhc action of Lactobacillus casei CRL 705 on pork muscle sarcoplasmic and myofibrillar proteins. J. Agric. Food Chem. 47,3441- 3448.
SANZ, Y., MULHOLLAND, F. and T O L D a , F. 1998. Purification and characterization of a tripeptidase from Lactobacillus sakei. J. Agric. Food Chem 46,349-353.
SANZ, Y. and T O L D a , F. 1997a. Purification and charactzerization of an aminopeptidase from Lactobacillus sake. J. Agric. Food Chem. 45, 1552- 1558.
SANZ, Y. and T O L D a , F. 1997b. Activities of aminopeptidases from Lactobacillus sake in models of curing ingredients and processing conditions for dry sausage. J. Food Sci. 62, 121 1-1234.
SANZ, Y. andTOLDa, F. 2001. Purification and charactzerization of an X- prolyl-dipeptidyl peptidase from Lactobacillus saki. Appl. Environ. Microbiol.
SANZ, Y. and T O L D a , F. 2002. Purification and characterization of an arginine aminopeptidase from Lactobacillus saki. Appl. Environ. Microbiol. 68, 1980- 1987.
SHEVCHENKO, A., WOLM, M., VORM, 0. and MA", M. 1996. Mass spectrometric sequencing of proteins Silver-stained polyacrylamide gels. Anal. Chem. 68, 850-858.
TSAKALIDOU, E., ANASTASIOU, R., VANDENBERGHE, I., VAN BEEUMEN, J. and KALANTZOPOULOS, G. 1999. Cell-wall-bound proteinase of Lactobacillus delbrueckii subsp. lactis ACA-DC 178: characterization and specificity for @-casein. Appl. Environ. Microbiol. 65,
VERPLAETSE, A. 1994. Influence of raw meat properties and processing technology on aroma quality of raw fermented meat products. In Roc. 40th Int. Congr. Meat Sci. Technol. pp. 45-65, The Hague, The Netherlands.
62, 156-161.
SANZ, Y., FADDA, S., VIGNOLO, G., ARISTOY, M.-C., OLIVER, G. and
67, 1815-1820.
2035-2040.




















