
Pleiotropic effect of the proton pump inhibitor esomeprazole ...

c
tínez
al step of,H),
nt.he wild
producedtate
.ffect
Research in Microbiology 156 (2005) 641–649
www.elsevier.com/locate/resmi
Pleiotropic effects of lactate dehydrogenase inactivationin Lactobacillus casei
Rosa Viana, María Jesús Yebra, José Luis Galán, Vicente Monedero, Gaspar Pérez-Mar∗
Biotechnology Department, Instituto de Agroquímica y Tecnología de Alimentos, IATA-CSIC, P.O. Box 73, 46100 Burjassot, Valencia, Spain
Received 6 August 2004; accepted 1 February 2005
Available online 22 April 2005
Abstract
In lactic acid bacteria, conversion of pyruvic to lactic acid through the activity of lactate dehydrogenase (Ldh) constitutes the finthe homofermentative pathway.Lactobacillus casei has two characterized genes encoding Ldh activities. TheldhL gene codes for an L-Ldhwhich specifically catalyzes the formation ofL-lactate, whereas thehicD gene codes for aD-hydroxyisocaproate dehydrogenase (HicDwhich catalyzes the conversion of pyruvate intoD-lactate. InL. casei cells fermenting glucose, a mixture ofL-/D-lactate with a 97:3% ratiowas formed. Inactivation ofhicD led to undetectableD-lactate levels after glucose fermentation, whileL-lactate levels remained constaInactivation ofldhL did not abolish the production ofL-lactate, but the lactate final concentration decreased about 25% compared to ttype, suggesting the presence of at least a second L-Ldh. Moreover, part of the pyruvate flux was rerouted and half of the lactatewas in theD-isomer form.ldhL inactivation inL. casei showed additional interesting effects. First, the glycolytic flux from pyruvate to lacwas redirected and other fermentation products, including acetate, acetoin, pyruvate, ethanol, diacetyl, mannitol and CO2, were producedSecond, a lack of carbon catabolite repression of lactose metabolism andN -acetyl-glucosaminidase activity was observed. This second ecould be partly avoided by growing the cells under aeration, since NADH oxidases could account for NAD+ regeneration. 2005 Elsevier SAS. All rights reserved.
Keywords: Lactobacillus; Lactate dehydrogenase;D-Hydroxyisocaproate dehydrogenase
riar-heionH
re-
es.mes39],dh)
to
se
ibitw-truc-an
ies
ants
in
nce
1. Introduction
The fermentative metabolism of lactic acid bacte(LAB) is characterized by the glycolytic breakdown of cabohydrates leading to the production of lactic acid [20]. Tlate step in this pathway is distinguished by the conversof pyruvate into lactate, a reaction which oxidizes the NADformed during glycolysis, thus maintaining the cellulardox balance. In LAB, both lactate isomers,L-lactate andD-lactate, can be formed by two isomer-specific enzymL-lactate dehydrogenases (L-Ldh) are a group of enzysharing homologies in bacteria, plants and animals [while D-lactate is produced by a group of enzymes (D-Lstructurally unrelated to L-Ldh and showing homologies
* Corresponding author.
E-mail address: [email protected] (G. Pérez-Martínez).0923-2508/$ – see front matter 2005 Elsevier SAS. All rights reserved.doi:10.1016/j.resmic.2005.02.011
short-chain fatty acids dehydrogenases (D-2-ketoacid dehy-drogenases) [23,40].D-hydroxyisocaproate dehydrogena(HicDH) is a second enzyme responsible forD-lactate syn-thesis. HicDHs share homologies with D-Ldhs but exhgreater affinity for 2-ketoacids than for pyruvate [2]. Hoever, the specificity of these enzymes resides in a few stural features, and conversion of D-Ldh into a D-HicDH cbe achieved by single amino acid changes [42].
In LAB, the presence of one type of Ldh or another varbetween species. InLactobacillus plantarum, genes for anL-Ldh and a D-Ldh have been characterized and muthave been obtained [8].L. plantarum produces a mixtureof 50% D-, 50% L-lactate but a double knock-out strainldhL andldhD is still able to synthesize lactic acid. InLac-tobacillus sakei, disruption of the singleldhL abolished theformation of both lactate isomers, suggesting the preseof racemase activity responsible for conversion ofL-lactate
into D-lactate [32]. InLactococcus lactis, disruption ofldhL
642 R. Viana et al. / Research in Microbiology 156 (2005) 641–649
ned1
on-bo-trucnen-mere of
rna-hisrod-the
u-ucerainabo
eem-ta-ruitdhentuc-ndnd
nmntr-of
umas
e
cia)
io-lated-
tore-thein-ingtedtion6
n-nt-mlas
p ap-d asd ond-N
r an-e-
zed
NAg
the
m
icendM) of7d
a-tant
ncepec-0bes
resulted in low accumulation of lactate that was explaiby the activation of a crypticldh gene as a result of an IS98element specific insertion [3].
The D-lactate isomer has been considered to be a nphysiological isomer, as it cannot be produced or metalized by mammals. Thus, efforts have been made to consLAB strains where only theL isomer is produced, which cabe useful for obtaining non-racemic lactate through fermtation, suitable for pharmaceutical biodegradable polysynthesis and food applications [1,24,25]. In the absencLdh activity, pyruvate can be metabolized by several altetive pathways which account for NADH regeneration. Tis an interesting feature, as most of the alternative end pucts obtained through these pathways are important indevelopment of flavor in fermented foods [5]. Also, by mtation of Ldh and expression of other activities able to redpyruvate (i.e., alanine dehydrogenase), engineered stcan be constructed to produce different added-value metlites [14].
L. casei is a lactic acid bacterium participating in thproduction of fermented dairy products and it has beenployed for the biosynthesis of lactic acid through fermention of many substrates, such as whey ultrafiltrate or fjuice [35,38]. In this species, the genes coding for an L-L[21] and a HicDH [27] have been described. In the preswork, the importance of these genes in lactic acid prodtion in L. casei has been studied via their inactivation athe impact of their mutations on end-product formation agrowth behavior.
2. Materials and methods
2.1. Strains and growth conditions
L. casei wild type (BL23) and ldhL (BL176), hicD(BL198) andccpA (BL190) [7] mutant strains were growin MRS medium (Oxoid) or MRS fermentation mediu(Adsa-Micro) supplemented with 0.5% (w/v) of differecarbohydrates at 37◦C under static conditions. For expeiments requiring aeration, cells were cultured in 25 mlmedium in 500 ml Erlenmeyer flasks at 37◦C and agitatedat 200 rpm.Escherichia coli DH5α was used in all cloningexperiments and was grown in Luria–Bertani (LB) mediat 37◦C under agitation. When required, erythromycin wadded to theL. casei growth medium at 5 µg/ml final con-centration. Ampicillin was used at 100 µg/ml for E. coli.
2.2. Construction of strains
Internal fragments ofL. casei ldhL andhicD genes wereamplified by PCR usingL. casei chromosomal DNA and tholigonucleotide pairs 5′-GACGCGATTGACTTAAGCAAC/5′-GTAAACGGACAGTGGCAGAAC and 5′-TTCATT-GCAGACAACGCC/5′-CGAGCCGTGTTGATCACA, re-
spectively. The PCR products were isolated, made bluntt
s-
and phosphorylated by using the SureClone kit (Pharmaand cloned into plasmid pRV300 [26] digested withSmaI,giving plasmids pVBldh and pVBhic, respectively.L. caseicells were electroporated [37] with these plasmids in a BRad Genepulser apparatus. Electroporated cells were pon MRS agar plates with 5 µg/ml erythromycin and incubated at 37◦C. Colonies developed on the plates after twothree days of incubation. Single cross-over integrationssulted in strains with a 62 and 102 amino acid deletion atC-terminus of Ldh and HicDH, respectively. The correcttegration of the plasmids was verified by Southern blottand PCR with appropriate oligonucleotides on DNA isolafrom several clones. Clones carrying the correct integraat theldhL andhicD loci were identified and named BL17and BL198, respectively.
2.3. RNA isolation and northern blotting
L. casei strains were grown in 10 ml of MRS fermetation medium supplemented with 0.5% (w/v) of differesugars to OD550 nmof 0.8. Cells were collected by centrifugation, washed with 50 mM EDTA and resuspended in 1of Trizol (Gibco BRL). One gram of 0.1-mm glass beads wadded and the cells were broken by shaking in a Fastpreparatus (Biospec), three times for 30 s. RNA was isolaterecommended by the manufacturers of Trizol, separatea formaldehyde–agarose gel and transferred to Hybonmembranes (Amersham). TheldhL and hicD RNA probesused in the northern blots were synthesized using eitheinternal ldhL or a hicD fragment cloned in pRV300 (plasmids pVBldh and pVBhic). As these plasmids are pBluScriptII SK derivatives, antisense RNA could be synthesiin vitro from linearized plasmids with T3 or T7 RNA poly-merase using the reagents from the Roche digoxigenin–Rlabelling kit. Hybridization (55◦C), washing and staininwere performed as recommended by the supplier usingCDP-star chemiluminescent substrate.
2.4. 13C-NMR analysis
L. casei cells were grown in MRS fermentation mediusupplemented with 0.5% glucose to an OD550 nm of 0.8.Cells were collected by centrifugation and washed twwith 10 mM potassium phosphate buffer (pH 6.8) aMgCl2 1 mM. The cell pellet was suspended in 50 mpotassium phosphate buffer (pH 6.8) to 12 mg (dry masscells per ml. The cell suspension was thermostatted at 3◦Cfor 5 min and [1-13C]glucose (Sigma–Aldrich) was addeto a final concentration of 10 mM. After 30 min incubtion at 37◦C the cells were centrifuged and the supernawas kept at−20◦C until NMR measurements.13C-NMRspectra were recorded at 75.47 MHz using a Bruker AvaDPX-300 spectrometer. Acquisition parameters were: stral width, 18 kHz; data size, 32 K; repetition delay, 1 s; 3◦pulse angle (5.5 µs); number of scans, 20 000. NMR tu
with a 5 mm diameter were used and 10% (vol/vol)2H2O
R. Viana et al. / Research in Microbiology 156 (2005) 641–649 643
-ar-fol-pedgaslett–con
kit
n,.8),ffernd
ellhos-
asithreded-ea-rH
wasin
yderud
nns.ibed
fferM
oftheOK,xi-the
n-
Cl.d.5)glassfor
n-ara-ro-and
ainstti-tase
or-acider-ctorormct
tud-t to
nted
umiletio
wondi-ium].le
-ildo--
lac-d inpe
f a76
not
was added to provide a lock signal. The13C chemical shiftswere relative to dioxane, designated at 67.19 ppm.
2.5. Metabolite determination
The concentration ofL- and D-lactate in the growthmedium was measured by using theD/L-lactic acid measurement kit from Roche. For the determination of volatile cbon metabolite formation, a resting cell system was usedlowed by concentration in a purge and trap system equipwith a Vocarb 3000 trap (Supelco) and analyzed bychromatography and mass spectroscopy using a HewPackard 7695 chromatograph as described [4]. Acetatecentrations were determined with the acetate detectionfrom Roche.
2.6. Enzymatic activities
N -acetyl-glucosaminidase and phospho-β-galactosidasewere measured in permeabilizedL. casei cells. For per-meabilization, cells were collected as for RNA isolatiowashed with 50 mM potassium phosphate buffer (pH 6and resuspended in one tenth volume of the same buFifty microliters of toluene:acetone (1:9) were added acells were vortexed at full speed for 5 min. Ten µl of csuspension were added to 250 µl of 10 mM potassium pphate buffer (pH 6.8), 1 mM MgCl2, 5 mM p-nitrophenyl-N -acetyl-β-D-glucosaminide (Sigma) and the mixture wincubated at 37◦C for 10 min. The reaction was stopped w250 µl of 5% Na2CO3 and the absorbance was measuat 420 nm. Phospho-β-galactosidase activity was measuras above using 5 mMp-nitrophenyl-β-D-galactopyranoside6-phosphate (Sigma) as substrate. Ldh activity was msured at 25◦C in 500 µl of 100 mM triethanolamine buffe(pH 6.8), 5 mM fructose-1,6-bisphosphate, 0.15 mM NADand 10–30 µg of cellular extract proteins. The reactionstarted by addition of pyruvate to 5 mM and changesthe absorbance at 340 nm were registered. Glyceraldeh3-phosphate (GAP) dehydrogenase was measured in cextracts ofL. casei strains grown in MRS fermentatiomedium with glucose under static or aeration conditioGAP dehydrogenase activity was determined as descrpreviously [11]. The activity was measured at 25◦C and thereaction mixture contained 125 mM triethanolamine bu(pH 7.2), 1 mM NAD+, 5 mM sodium arsenate and 5 mcysteine. The reactions were initiated by the addition2 mM GAP. Cellular extracts were obtained by shakingcells in a Bead-beater apparatus (Biospec, Bartlesville,USA) with 0.1 mm glass beads (four cycles of 30 s at mamal speed). Protein concentrations were determined withBio-Rad dye-binding assay.
2.7. Western blot
L. casei cells were grown in MRS medium (50 ml) co
taining 0.5% glucose to an OD550 nm of 0.6. The pH of-
.
-e
the culture was lowered to 4.5 by adding concentrated HCells were collected by centrifugation at 4◦C, resuspendein 1 ml of cold 20 mM potassium acetate buffer (pH 4and broken in a Bead-beater apparatus (Biospec) usingbeads. Cell-free extracts were obtained by centrifugation10 min at 14 000g and 4◦C and were separated on nodenaturing 15% polyacrylamide gels, which enabled seption of phosphorylated from non-phosphorylated HPr. Pteins were electroblotted onto nitrocellulose membranesHPr was detected using a rabbit antiserum raised agB. subtilis HPr (kindly supplied by J. Deutscher) as first anbody and anti-rabbit IgG conjugated to alkaline phosphaas second antibody.
3. Results
3.1. Inactivation of ldhL and hicD in L. casei
ldhL and hicD genes have been isolated fromL. casei(acc. nos. M76708 and A14931, respectively) [21,27]. Inder to determine the contribution of these genes to lacticproduction in this bacterium, DNA corresponding to intnal fragments of both genes was cloned in the suicide vepRV300 and the resulting plasmids were used to transfthe L. casei wild-type strain. Two integrants in the correloci, BL176 (ldhL::pRV300) and BL198 (hicD::pRV300),were obtained and the impact of the mutations was sied by measuring lactate production on cultures broughlate stationary growth phase in MRS medium supplemewith 0.5% glucose. Under such conditions,L. casei wildtype accumulated 31.4 mM lactate in the growth medi(residualL-lactate concentration in MRS was 3 mM, whno D-lactate could be detected), with a 97:3 molar raof L- to D-isomers (Table 1). The ratio between the tlactate isomers changed as function of the growth cotions; for instance, acetate, present in the MRS med(60 mM), can be an inhibitor of HicDH activity [17,18The BL198 (hicD::pRV300) mutant showed no detectabD-lactate production, although theL-lactate final concentration reached values similar to total lactate in the wtype. TheldhL mutation had a major effect on lactate prduction. In strain BL176 (ldhL::pRV300), total lactate production decreased by 25% and an increase in theD-lactateconcentration was observed (Table 1). These changes intate production had an impact on the final pH reachethe growth medium, which was around 4.5 for the wild tyandhicD mutant, while it was 5.2 for theldhL mutant. Thedecrease inL-lactate produced was the consequence oremarkable decrease in L-Ldh activity detected in BL1(ldhL::pRV300) compared to the wild type or thehicD mu-tant (Table 1). Mutation of thehicD gene in BL198 hadno obvious effect on growth, but disruption ofldhL ledto an elevated doubling time in all sugars tested (data
shown).
644 R. Viana et al. / Research in Microbiology 156 (2005) 641–649
Table 1Effect of ldhL andhicD inactivation on glucose fermentation products inL. casei
Strain Ldh activitya L-Lactateb D-Lactateb pH Ethanolc Acetatec Acetaldehydec Diacetylc
Wild type 962.2± 68.1 30.6± 2.7 0.8± 0.5 4.5± 0.1 29.1± 2.6 4.1± 0.2 3.5± 0.2 2.4± 0.1hicD 960.7± 27.1 35.1± 1.2 < 0.02 4.5± 0.1 ND ND ND NDldhL 59.9± 19.6 12.4± 1.3 11.2± 0.4 5.2± 0.2 1217.8± 26.3 12.3± 0.5 5.3± 0.2 41.2± 2.2
ND: not determined.a Activity is given in nmol/min mg protein.b Lactate concentration is given in mM.c Concentrations are given in µM and were determined in a cell resting system (see Section 2).
-log
is,m
eralthen
. 1).
onpho-thaomrin,stinnt
tionissisuldnd
ton-nd
gheroin,
ion,
i-ldetyl.
in-byd di-d).
atechl bycanb-le
gesn-
eyP)
Fig. 1. Northern blot analysis ofldhL transcription inL. casei. RNA wasextracted from cells grown in MRS plus 0.5% of different sugars to midphase and probed with a DIG-labeledldhL probe. Strains are: BL23 (wildtype) and BL190 (ccpA). G, glucose; L, lactose; R, ribose.
3.2. ldhL transcriptional analysis
According to sequence data, bothldhL and hicD fromL. casei are monocistronic genes. In order to prove thnorthern blots were performed with RNA isolated froL. casei and usingldhL andhicD probes. Northern blots withahicD probe did not show any detectable signal after sevtrials in both the wild-type andldhL mutant strains (data noshown), suggesting that the gene is poorly expressed. WanldhL probe was used, a 1-kb signal was detected (FigThis size is in good agreement with the size ofldhL, indicat-ing that inL. casei ldhL is monocistronic. TheldhL signalwas clearly stronger in RNA isolated from cells grownglucose or lactose, which are transported by the phosenolpyruvate:sugar phosphotransferase system (PTS),in the non-PTS sugar ribose. RNA was also isolated frstrain BL190, mutated in theccpA gene encoding the majocarbon catabolite regulator protein (CcpA) [7]. In this straldhL messenger levels decreased in PTS sugars, suggethat ldhL in L. casei could be regulated by CcpA-dependeactivation.
3.3. Effect of ldhL inactivation on levelsof fermentation end products
Lactate dehydrogenase is implicated in the re-oxidaof NADH formed during glycolysis. In the absence of thenzymatic activity, the pyruvate produced during glycolymust be metabolized by alternative pathways, which coalso oxidize NADH. We investigated the fermentation eproducts produced by resting cells ofL. casei wild type and
different mutants by NMR. As described in Fig. 2, undern
g
Table 2GAP dehydrogenase activities in crude extracts ofL. casei strains grownunder static or aeration conditions
Strain Activity (nmol/min mg protein)a Aeration/static ratioStatic Aeration
Wild type 2.040± 41 2.296± 67 1.13ldhL 1.004±106 1.146±170 1.14hicD 1.747± 28 1.955±135 1.12
a Enzyme activities are means± standard deviations.
these conditions,L. casei wild type carried out an almosexclusive homolactic conversion of glucose with the ccomitant production of lactic acid, although pyruvic aacetic acid were also detected in small amounts. TheldhLmutant produced, in addition to these compounds, a hiproportion of other fermentation products such us acetmannitol and ethanol (Fig. 2A). ThehicD mutant showed thesame pattern of fermentation as the wild type. In additan L. casei ccpA mutant, which showed anldhL transcrip-tion lower than that of the wild type, still displayed a simlar 13C-NMR metabolite profile. The NMR technique counot be used to detect volatile compounds such us diacTherefore, wild typeL. casei and BL176 (ldhL::pRV300)strains were grown in a resting cell medium [4] containg glucose, and different metabolites were quantifiedgas chromatography. Acetate, acetaldehyde, ethanol anacetyl production in theldhL mutant was 3, 1.5, 42 an17-fold higher than in the wild type, respectively (Table 1
The accumulation of high amounts of ethanol in theldhLstrain indicates that a significant amount of the pyruvpool is channeled toward formation of acetyl-CoA, whiis ultimately transformed into acetaldehyde and ethanothe alcohol dehydrogenase enzyme (Adh). This activitybe of crucial importance in NADH regeneration in the asence of Ldh, as it oxidizes two moles of NADH per moof acetyl-CoA. Indeed, Adh activity in theldhL strain was7.5-fold greater than in the wild type (2.4 ± 0.3 and 17.9±1.3 nmol/min mg protein for wild-type andldhL strains,respectively). This would indicate that metabolic chancaused by anldhL mutation can induce the expression of ezymatic activities responsible for NADH oxidation. The kenzymatic activity of glyceraldehyde-3-phosphate (GA
dehydrogenase, an important flux-controlling enzyme [11],
R. Viana et al. / Research in Microbiology 156 (2005) 641–649 645
was
Fig. 2.13C nuclear magnetic resonance spectra of end products from cell suspensions of (A)L. casei BL23 (wild type), (B)L. casei BL176 (ldhL::pRV300),(C) L. casei BL190 (ccpA) and (D)L. casei BL198 (hicD::pRV300), incubated with [1-13C]-glucose. Spectra are presented from 10 to 70 ppm. Dioxane used as reference signal at 67.19 ppm.de-).
iteellsr is
upionow-thAc-e-ild
me
htg
oseasetthexic
was also measured in the mutants, displaying a two-foldcrease in theldhL strain compared to the wild type (Table 2
3.4. Pleiotropic effect of the ldhL mutationin catabolite repression
TheL. casei wild-type strain displays a strong catabolrepression (CR) effect upon lactose utilization. When cgrow on a mixture of glucose and lactose, the first sugautilized, followed by an arrest in growth which lasts forto 15 h [44]. After that, growth is resumed by consumptof the second sugar, as shown in previous works [44]. Hever, the BL176 strain did not display this diauxic growbehavior and no arrest in growth was observed (Fig. 3A).cordingly, while phospho-β-galactosidase, the enzyme rsponsible for lactose utilization, was repressed in the wtype grown on glucose plus lactose (0.6 nmol/min mg dryweight), it was fully expressed in BL176 under the saconditions (12.6 nmol/min mg dry weight).
One of the consequences ofldhL inactivation could be theinability of the cells to efficiently re-oxidate NADH througreduction of pyruvate to lactate. However, O2-dependenNADH oxidation could constitute a direct form for testin
whether re-oxidation of NADH by alternative pathways re-Fig. 3. Typical growth curves ofL. casei wild type and anldhL mutant inMRS containing 0.1% glucose plus 0.5% lactose at 37◦C: (A) growth understatic conditions; (B) growth under aeration.
stores CR in theL. casei ldhL mutant. Growth ofL. caseiwas measured in MRS culture medium containing glucplus lactose under vigorous agitation, which would increthe amount of dissolved O2 and thus promote O2-dependenNADH oxidase activity [33]. Fig. 3B shows the growcurves of theL. casei wild type andldhL mutants under thesconditions. The wild-type strain also showed normal diau
growth (about 15-h lag) also under aeration conditions. In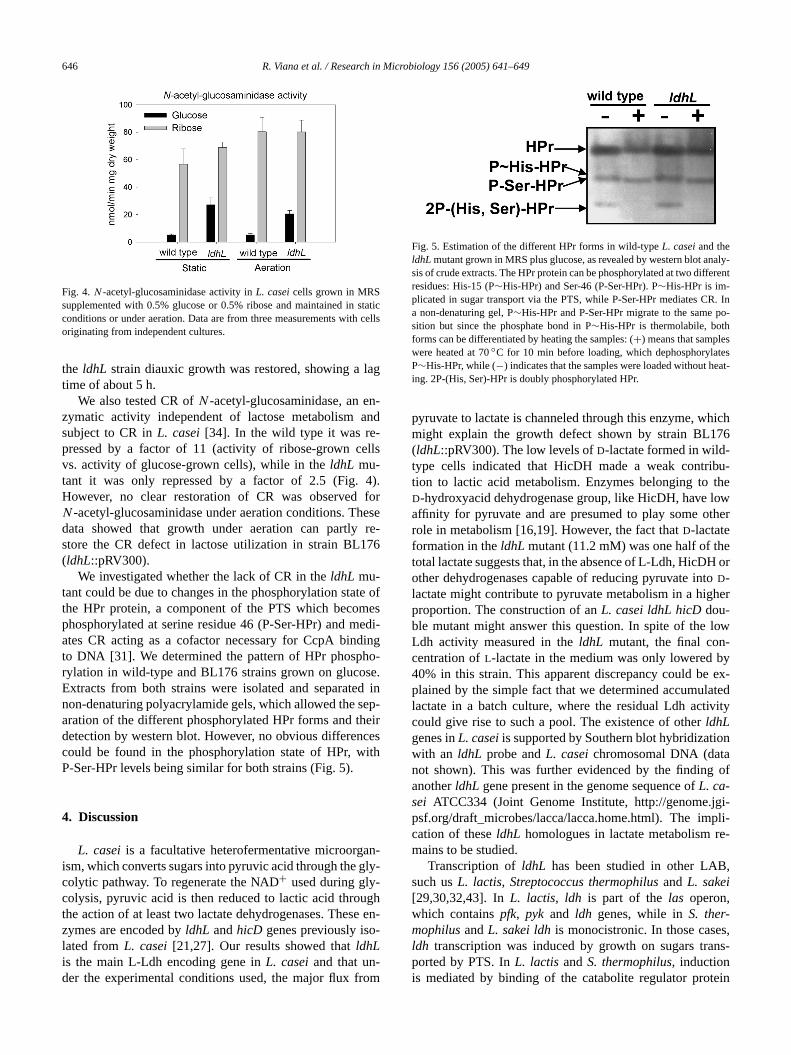
646 R. Viana et al. / Research in Microbiology 156 (2005) 641–649
taticcells
lag
n-nd
lls
4).foresere-
76
te ofes
edi-dingo-e.d inep-eircesith
n-ly-
ghe en-
aly-erent
R. Ino-
steseat-
hich76
u-theowther
eor
oher
low-byex-ted
vity
n
of
gi-pli-e-
,
s,ns-
Fig. 4. N -acetyl-glucosaminidase activity inL. casei cells grown in MRSsupplemented with 0.5% glucose or 0.5% ribose and maintained in sconditions or under aeration. Data are from three measurements withoriginating from independent cultures.
the ldhL strain diauxic growth was restored, showing atime of about 5 h.
We also tested CR ofN -acetyl-glucosaminidase, an ezymatic activity independent of lactose metabolism asubject to CR inL. casei [34]. In the wild type it was re-pressed by a factor of 11 (activity of ribose-grown cevs. activity of glucose-grown cells), while in theldhL mu-tant it was only repressed by a factor of 2.5 (Fig.However, no clear restoration of CR was observedN -acetyl-glucosaminidase under aeration conditions. Thdata showed that growth under aeration can partlystore the CR defect in lactose utilization in strain BL1(ldhL::pRV300).
We investigated whether the lack of CR in theldhL mu-tant could be due to changes in the phosphorylation stathe HPr protein, a component of the PTS which becomphosphorylated at serine residue 46 (P-Ser-HPr) and mates CR acting as a cofactor necessary for CcpA binto DNA [31]. We determined the pattern of HPr phosphrylation in wild-type and BL176 strains grown on glucosExtracts from both strains were isolated and separatenon-denaturing polyacrylamide gels, which allowed the saration of the different phosphorylated HPr forms and thdetection by western blot. However, no obvious differencould be found in the phosphorylation state of HPr, wP-Ser-HPr levels being similar for both strains (Fig. 5).
4. Discussion
L. casei is a facultative heterofermentative microorgaism, which converts sugars into pyruvic acid through the gcolytic pathway. To regenerate the NAD+ used during gly-colysis, pyruvic acid is then reduced to lactic acid throuthe action of at least two lactate dehydrogenases. Theszymes are encoded byldhL andhicD genes previously isolated fromL. casei [21,27]. Our results showed thatldhLis the main L-Ldh encoding gene inL. casei and that un-
der the experimental conditions used, the major flux from-
Fig. 5. Estimation of the different HPr forms in wild-typeL. casei and theldhL mutant grown in MRS plus glucose, as revealed by western blot ansis of crude extracts. The HPr protein can be phosphorylated at two diffresidues: His-15 (P∼His-HPr) and Ser-46 (P-Ser-HPr). P∼His-HPr is im-plicated in sugar transport via the PTS, while P-Ser-HPr mediates Ca non-denaturing gel, P∼His-HPr and P-Ser-HPr migrate to the same psition but since the phosphate bond in P∼His-HPr is thermolabile, bothforms can be differentiated by heating the samples: (+) means that samplewere heated at 70◦C for 10 min before loading, which dephosphorylaP∼His-HPr, while (−) indicates that the samples were loaded without hing. 2P-(His, Ser)-HPr is doubly phosphorylated HPr.
pyruvate to lactate is channeled through this enzyme, wmight explain the growth defect shown by strain BL1(ldhL::pRV300). The low levels ofD-lactate formed in wild-type cells indicated that HicDH made a weak contribtion to lactic acid metabolism. Enzymes belonging toD-hydroxyacid dehydrogenase group, like HicDH, have laffinity for pyruvate and are presumed to play some orole in metabolism [16,19]. However, the fact thatD-lactateformation in theldhL mutant (11.2 mM) was one half of thtotal lactate suggests that, in the absence of L-Ldh, HicDHother dehydrogenases capable of reducing pyruvate intD-lactate might contribute to pyruvate metabolism in a higproportion. The construction of anL. casei ldhL hicD dou-ble mutant might answer this question. In spite of theLdh activity measured in theldhL mutant, the final concentration ofL-lactate in the medium was only lowered40% in this strain. This apparent discrepancy could beplained by the simple fact that we determined accumulalactate in a batch culture, where the residual Ldh acticould give rise to such a pool. The existence of otherldhLgenes inL. casei is supported by Southern blot hybridizatiowith an ldhL probe andL. casei chromosomal DNA (datanot shown). This was further evidenced by the findinganotherldhL gene present in the genome sequence ofL. ca-sei ATCC334 (Joint Genome Institute, http://genome.jpsf.org/draft_microbes/lacca/lacca.home.html). The imcation of theseldhL homologues in lactate metabolism rmains to be studied.
Transcription ofldhL has been studied in other LABsuch usL. lactis, Streptococcus thermophilus and L. sakei[29,30,32,43]. InL. lactis, ldh is part of thelas operon,which containspfk, pyk and ldh genes, while inS. ther-mophilus andL. sakei ldh is monocistronic. In those caseldh transcription was induced by growth on sugars traported by PTS. InL. lactis andS. thermophilus, induction
is mediated by binding of the catabolite regulator protein
R. Viana et al. / Research in Microbiology 156 (2005) 641–649 647
ting, as
thaten
the
lev-
ticnd
elarr-of
sted
part
trob-to
e of
andtha
eledasearac
liq-at ins-,
bothay
ex-
ted
1-Pose-
sed
-
n-
Apro-.rinecpA
tedingse,lase,n of
ofvelse,test ofher-P ore-nt inges
smtiv-es
butge-ight-as,
% is
ingR.
t inds
ject
va,
2
ook,e-
CcpA to the promoter [28,43]. Transcription ofldhL in anL. casei ccpA mutant decreased in PTS sugars, suggesthat it could be regulated by CcpA-dependent activationit is in L. lactis and S. thermophilus. The fact that readilyutilized sugars (PTS sugars) promote the expression ofldhLseems to be a characteristic of LAB. It seems, therefore,these bacteria need to accommodate the levels of thiszyme to ensure efficient metabolism of PTS sugars. Atsame time, Ldh-L fromL. casei is allosterically activatedby fructose-1,6-bisphosphate [10], whose intracellularels increase upon PTS sugar utilization [41].
In L. lactis, the lack ofldh transcriptional activation ina ccpA mutant has the effect of a shift from homolacto mixed acid fermentation, with production of acetate aethanol [28]. Transcription analysis in theL. casei ccpA mu-tant showed thatldhL transcription was lower than in thwild type. However, both strains still displayed a simi13C-NMR metabolite profile with almost exclusive convesion of glucose into lactic acid, although small amountspyruvic and acetic acid were also detected. This suggethat low transcription ofldhL in L. casei did not significantlyaffect the general balance of metabolites during the lastof glycolysis.
Inactivation of ldhL in L. casei, although not sufficiento produce a dramatic decrease in lactate production, pably produced a misbalance in NADH levels, leadingactivation of other catabolic balancing pathways capablNAD+ regeneration. In addition to lactate, theldhL mutantproduced pyruvate, acetate, acetoin, ethanol, mannitoldiacetyl. The presence of these metabolites suggestedin the absence of the main Ldh, pyruvate could be channmainly via α-acetolactate synthase, pyruvate-formate lyand pyruvate oxidase pathways. These pathways are chterized by oxidation of NADH and by the formation of CO2at different enzymatic steps. Indeed, theL. casei ldhL mu-tant was able to produce gas (probably CO2) from glucosefermentation, as evidenced when using Durhan bells inuid medium (data not shown). These data suggested ththeL. casei ldhL strain, pyruvate metabolism is rerouted esentially as it is in other LAB impaired in Ldh [15]. Alsothe presence of pyruvate as a product of fermentation,in wild-type andldhL strains, indicated that under our assconditions, the fermentative metabolism ofL. casei cannotaccommodate all pyruvate produced, which is partlycreted, as reported for anldh mutant ofLactobacillus fer-mentum [1]. Production of mannitol has also been reporfor ldh mutants ofL. lactis, L. plantarum and L. fermen-tum [1,9,36] as a result of the presence of a mannitol-dehydrogenase. This enzyme is able to transform fruct6-P into mannitol-1-P linked to regeneration of NAD+. Theconversion of mannitol-1-P into mannitol has been propoto occur via mediation by phosphatase activity of the EIIMtl
mannitol transporter inL. plantarum [9], and by a hypothetical mannitol-1-P phosphatase inL. lactis [12].
In addition to the redistribution of the pattern of ferme
tation products, theldhL mutation led to an apparent lack-
t,
-
of CR. In L. casei, CR is mediated by binding of the Ccpregulator protein to specific sequences present in themoter of catabolite-regulated genes (cre sequences) [31]The HPr protein of PTS, when phosphorylated at the se46 residue (P-Ser-HPr), acts as cofactor necessary for Cbinding to DNA. P-Ser-HPr is formed when the elevaconcentration of glycolytic intermediates produced durutilization of a readily metabolizable sugar (i.e., glucofructose) activates the enzyme HPr kinase/phosphorywhich catalyzes phosphorylation and dephosphorylatioHPr at Ser-46 [6]. Analysis of the phosphorylation stateHPr indicated that no obvious decrease in P-Ser-HPr leoccurred in theldhL strain compared to the wild type. Hencthe likely changes in the pools of glycolytic intermediagenerated by an Ldh-L deficiency did not affect this parthe CR signal transduction pathway. It is known that otcofactors such as fructose-1,6-bisphosphate, glucose-6NADP may influence CcpA binding to DNA [13,22]. Therfore, it can also be suggested that the CR defect presetheL. casei ldhL mutant might be a consequence of chanin a cofactor required by CcpA.
It was conceivable that the modification in metaboliproduced by an Ldh-L deficiency also affected the acity or expression of glycolytic or other metabolic enzymrequiring cofactors. We showed that anldhL mutation influ-enced not only the final balance of excreted metabolitesalso the expression of key enzymes, like GAP dehydronase. In addition, this enzyme is strongly inhibited by a hNADH/NAD+ ratio [11]. Some LAB have O2-dependenNADH oxidases able to carry out NAD+ regeneration; however, they may play a secondary role in cell metabolismin L. bulgaricus, only 2–3% of NADH formed during gly-colysis is oxidized by these enzymes, whereas 97–98oxidized by Ldh [33]. We showed that growth of theL. ca-sei ldhL mutant under intense aeration in conditions favorthe activity of NADH oxidases led to partial recovery of CIt seems, therefore, that restoration of NAD+ levels was suf-ficient to revert some of the pleiotropic effects of anldhLmutation.
Acknowledgements
We thank Dr. Isabel Solana for her technical supporNMR experiments. This project was supported by funfrom the Spanish Ministerio de Educación y Ciencia (ProBIO2001-01616).
References
[1] J. Aarnikunnas, N. Von Weymarn, K. Ronnholm, M. Leisola, A. PalMetabolic engineering ofLactobacillus fermentum for production ofmannitol and pureL-lactic acid or pyruvate, Biotechnol. Bioeng. 8(2003) 653–663.
[2] N. Bernard, K. Johnsen, T. Ferain, D. Garmyn, P. Hols, J.J. HolbrJ. Delcour, NAD(+)-dependentD-2-hydroxyisocaproate dehydrognase ofLactobacillus delbrueckii subsp.bulgaricus. Gene cloning and
enzyme characterization, Eur. J. Biochem. 224 (1994) 439–446.
648 R. Viana et al. / Research in Microbiology 156 (2005) 641–649
ink,dap-
lin-
togr.
04)
ez-nal
sion,
ez,
ng
yn,
ance
)
44
on-on
man,
ge-. 70
op-trol
oltz,
ring,
ro,, G.c en-als,
-riol.
thef
.
on
ro-
ge-e
nie
e ofon.
orci.
vo--ge-
i-
ts.
e--
ns,
.M.
8)
e-
ofc-ctate
ez,e re-
c,
–
f hy-
pl.
ssione-
rd,en
.eida,
. J.
[3] R.S. Bongers, M.H. Hoefnagel, M.J. Starrenburg, M.A. SiemerJ.G. Arends, J. Hugenholtz, M. Kleerebezem, IS981-mediated ative evolution recovers lactate production byldhB transcription activa-tion in a lactate dehydrogenase-deficient strain ofLactococcus lactis,J. Bacteriol. 185 (2003) 4499–4507.
[4] P. Dauneau, G. Pérez-Martínez, Fractional factorial and multipleear regression to optimize extraction of volatiles from aLactobacillusplantarum bacterial suspension using purge and trap, J. ChromaA. 775 (1997) 225–230.
[5] W.M. de Vos, J. Hugenholtz, Engineering metabolic highways inLac-tococci and other lactic acid bacteria, Trends. Biotechnol. 22 (2072–79.
[6] V. Dossonnet, V. Monedero, M. Zagorec, A. Galinier, G. PérMartínez, J. Deutscher, Phosphorylation of HPr by the bifunctioHPr Kinase/P-ser-HPr phosphatase fromLactobacillus casei controlscatabolite repression and inducer exclusion but not inducer expulJ. Bacteriol. 182 (2000) 2582–2590.
[7] C.D. Esteban, K. Mahr, V. Monedero, W. Hillen, G. Pérez-MartínF. Titgemeyer, Complementation of a�ccpA mutant ofLactobacilluscasei with CcpA mutants affected in the DNA- and cofactor-bindidomains, Microbiology 150 (2004) 613–620.
[8] T. Ferain, J.N. Hobbs Jr., J. Richardson, N. Bernard, D. GarmP. Hols, N.E. Allen, J. Delcour, Knockout of the twoldh genes hasa major impact on peptidoglycan precursor synthesis inLactobacillusplantarum, J. Bacteriol. 178 (1996) 5431–5437.
[9] T. Ferain, A.N. Schanck, J. Delcour, 13C nuclear magnetic resonanalysis of glucose and citrate end products in anldhL–ldhD double-knockout strain ofLactobacillus plantarum, J. Bacteriol. 178 (19967311–7315.
[10] E.I. Garvie, Bacterial lactate dehydrogenases, Microbiol. Rev.(1980) 106–139.
[11] C. Garrigues, P. Loubiere, N.D. Lindley, M. Cocaign-Bousquet, Ctrol of the shift from homolactic acid to mixed-acid fermentatiin Lactococcus lactis: Predominant role of the NADH/NAD+ ratio,J. Bacteriol. 179 (1997) 5282–5287.
[12] P. Gaspar, A.R. Neves, A. Ramos, M.J. Gasson, C.A. ShearH. Santos, EngineeringLactococcus lactis for production of mannitol:High yields from food-grade strains deficient in lactate dehydronase and the mannitol transport system, Appl. Environ. Microbiol(2004) 1466–1474.
[13] R. Gosseringer, E. Kuster, A. Galinier, J. Deutscher, W. Hillen, Coerative and non-cooperative DNA binding modes of catabolite conprotein CcpA fromBacillus megaterium result from sensing two dif-ferent signals, J. Mol. Biol. 266 (1997) 665–676.
[14] P. Hols, M. Kleerebezem, A.N. Schanck, T. Ferain, J. HugenhJ. Delcour, W.M. de Vos, Conversion ofLactococcus lactis from ho-molactic to homoalanine fermentation through metabolic engineeNat. Biotechnol. 17 (1999) 588–592.
[15] J. Hugenholtz, W. Sybesma, M.N. Groot, W. Wisselink, V. LadeK. Burgess, D. van Sinderen, J.C. Piard, G. Eggink, E.J. SmidSavoy, F. Sesma, T. Jansen, P. Hols, M. Kleerebezem, Metaboligineering of lactic acid bacteria for the production of nutraceuticAntonie Van Leeuwenhoek 82 (2002) 217–235.
[16] W. Hummel, H. Schütte, M.-R. Kula,D-2-Hydroxyisocaproate dehydrogenase fromLactobacillus casei: A new enzyme suitable fostereospecific reduction of 2-ketocarboxylic acids, Appl. MicrobBiotechnol. 21 (1985) 7–15.
[17] T. Iino, T. Uchimura, K. Komagata, The effect of sodium acetate ongrowth yield, the production ofL- andD-lactic acid, and the activity osome enzymes of the glycolytic pathway ofLactobacillus sakei NRIC1071(T) andLactobacillus plantarum NRIC 1067(T), J. Gen. ApplMicrobiol. 48 (2002) 91–102.
[18] T. Iino, T. Uchimura, K. Komagata, The effect of sodium acetatethe activity ofL- andD-lactate dehydrogenases inLactobacillus sakeiNRIC 1071(T) and other lactic acid bacteria, J. Gen. Appl. Mic
biol. 49 (2003) 51–58.[19] H.K.W. Kallwass, Potential of R-2-hydroxyisocaproate dehydronase fromLactobacillus casei for stereospecific reductions, EnzymMicrob. Technol. 14 (1992) 28–35.
[20] O. Kandler, Carbohydrate metabolism in lactic acid bacteria, AntoVan Leeuwenhoek 49 (1983) 209–224.
[21] S.F. Kim, S.J. Baek, M.Y. Pack, Cloning and nucleotide sequencthe Lactobacillus casei lactate dehydrogenase gene, Appl. EnvirMicrobiol. 57 (1991) 2413–2417.
[22] J.H. Kim, M.I. Voskuil, G.H. Chambliss, NADP, corepressor fthe Bacillus catabolite control protein CcpA, Proc. Natl. Acad. SUSA 95 (1998) 9590–9595.
[23] S. Kochhar, P.E. Hunziker, P. Leong-Morgenthaler, H. Hottinger, Elutionary relationship of NAD(+)-dependentD-lactate dehydrogenase: Comparison of primary structure of 2-hydroxy acid dehydronases, Biochem. Biophys. Res. Comm. 184 (1992) 60–66.
[24] K. Kyla-Nikkila, M. Hujanen, M. Leisola, A. Palva, Metabolic engneering ofLactobacillus helveticus CNRZ32 for production of pureL-(+)-lactic acid, Appl. Environ. Microbiol. 66 (2000) 3835–3841.
[25] L. Lapierre, J.E. Germond, A. Ott, M. Delley, B. Mollet,D-Lactate de-hydrogenase gene (ldhD) inactivation and resulting metabolic effecin the Lactobacillus johnsonii strains La1 and N312, Appl. EnvironMicrobiol. 65 (1999) 4002–4007.
[26] L. Leloup, S.D. Ehrlich, M. Zagorec, F. Morel-Deville, Singlcrossover integration in theLactobacillus sake chromosome and insertional inactivation of theptsI and lacL genes, Appl. Environ. Mi-crobiol. 63 (1997) 2117–2123.
[27] H.P. Lerch, H. Blocker, H. Kallwass, J. Hoppe, H. Tsai, J. ColliCloning, sequencing and expression inEscherichia coli of theD-2-hydroxyisocaproate dehydrogenase gene ofLactobacillus casei,Gene 78 (1989) 47–57.
[28] E.J. Luesink, R.E. van Herpen, B.P. Grossiord, O.P. Kuipers, Wde Vos, Transcriptional activation of the glycolyticlas operon andcatabolite repression of thegal operonin Lactococcus lactis are medi-ated by the catabolite control protein CcpA, Mol. Microbiol. 30 (199789–798.
[29] R.M. Llanos, A.J. Hillier, B.E. Davidson, Cloning, nucleotide squence, expression, and chromosomal location ofldh, the gene en-codingL-(+)-lactate dehydrogenase, fromLactococcus lactis, J. Bac-teriol. 174 (1992) 6956–6964.
[30] R.M. Llanos, C.J. Harris, A.J. Hillier, B.E. Davidson, Identificationa novel operon inLactococcus lactis encoding three enzymes for latic acid synthesis: Phosphofructokinase, pyruvate kinase, and ladehydrogenase, J. Bacteriol. 175 (1993) 2541–2551.
[31] K. Mahr, C.D. Esteban, W. Hillen, F. Titgemeyer, G. Pérez-MartínCross communication between components of carbon catabolitpression ofLactobacillus casei andBacillus megaterium, J. Mol. Mi-crobiol. Biotechnol. 4 (2002) 489–494.
[32] C. Malleret, R. Lauret, S.D. Ehrlich, F. Morel-Deville, M. ZagoreDisruption of the soleldhL gene inLactobacillus sakei prevents theproduction of bothL- andD-lactate, Microbiology 144 (1998) 33273333.
[33] C. Marty-Teysset, F. de la Torre, J. Garel, Increased production odrogen peroxide byLactobacillus delbrueckii subsp.bulgaricus uponaeration: Involvement of an NADH oxidase in oxidative stress, ApEnviron. Microbiol. 66 (2000) 262–267.
[34] V. Monedero, M.J. Gosalbes, G. Pérez-Martínez, Catabolite reprein Lactobacillus casei ATCC 393 is mediated by CcpA, J. Bactriol. 179 (1997) 6657–6664.
[35] N. Nancib, A. Nancib, A. Boudjelal, C. Benslimane, F. BlanchaJ. Boudrant, The effect of supplementation by different nitrogsources on the production of lactic acid from date juice byLactobacil-lus casei subsp.rhamnosus, Bioresour. Technol. 78 (2001) 149–153
[36] A.R. Neves, A. Ramos, C. Shearman, M.J. Gasson, J.S. AlmH. Santos, Metabolic characterization ofLactococcus lactis defi-cient in lactate dehydrogenase using in vivo 13C-NMR, Eur
Biochem. 267 (2000) 3859–3868.
R. Viana et al. / Research in Microbiology 156 (2005) 641–649 649
.M.to-lusppl.
cid-
ro-NA
these-
)
m-
Y.chi,a
id re-
Vos,gly-
ndent982–
the
on,
[37] M. Posno, R.J. Leer, N. van Luijk, M.J.F. van Giezen, P.T.HHeuvelmans, B.C. Lokman, P.H. Pouwels, Incompatibility of lacbacillus vectors with replicons derived from small cryptic lactobacilplasmids and segregational instability of the introduced vectors, AEnviron. Microbiol. 57 (1991) 1822–1828.
[38] A. Senthuran, V. Senthuran, R. Hatti-Kaul, B. Mattiasson, Lactic aproduction by immobilizedLactobacillus casei in recycle batch reactor: A step towards optimization, J. Biotechnol. 73 (1999) 61–70.
[39] D.W. Stock, J.M. Quattro, G.S. Whitt, D.A. Powers, Lactate dehydgenase (LDH) gene duplication during chordate evolution: The cDsequence of the LDH of the tunicateStyela plicata, Mol. Biol. Evol. 14(1997) 1273–1284.
[40] H. Taguchi, T. Ohta,D-lactate dehydrogenase is a member ofD-isomer-specific 2-hydroxyacid dehydrogenase family: Cloning,quencing, and expression inEscherichia coli of theD-lactate dehydro-genase gene ofLactobacillus plantarum, J. Biol. Chem. 266 (199112588–12594.
[41] F. Titgemeyer, W. Hillen, Global control of sugar metabolism: A grapositive solution, Antonie Van Leeuwenhoek 82 (2002) 59–71.
[42] C. Tokuda, Y. Ishikura, M. Shigematsu, H. Mutoh, S. Tsuzuki,Nakahira, Y. Tamura, T. Shinoda, K. Arai, O. Takahashi, H. TaguConversion ofLactobacillus pentosus D-lactate dehydrogenase toD-hydroxyisocaproate dehydrogenase through a single amino acplacement, J. Bacteriol. 185 (2003) 5023–5026.
[43] P.T. van den Bogaard, M. Kleerebezem, O.P. Kuipers, W.M. deControl of lactose transport, beta-galactosidase activity, andcolysis by CcpA inStreptococcus thermophilus: Evidence for car-bon catabolite repression by a non-phosphoenolpyruvate-depephosphotransferase system sugar, J. Bacteriol. 182 (2000) 55989.
[44] A. Veyrat, V. Monedero, G. Pérez-Martínez, Glucose transport byphosphoenolpyruvate:mannose phosphotransferase system inLacto-bacillus casei ATCC 393 and its role in carbon catabolite repressiMicrobiology 140 (1994) 1141–1149.
Copyright © 2022 FDOKUMEN




















