
Leuchtende Tiere: Chemie und biologische Bedeutung
10
Klaus H. Hoffmann: Leuchtende Tiere : Chemie und biologische Bedeutung franzosische Physiologe Raphael Dubois aus Leuchtkafern zwei Stoffe isoliert, die im Rea- genzglas vereinigt in Gegenwart von Sauer- stoff aufleuchteten. Die hitzestabile Kompo- nente bezeichnete er als Luciferin, die hitze- labile als Luciferase. Mit dem Nachweis von Luciferin und Luciferase brachte Dubois die Erzeugung von Licht durch Tiere und Pflan- zen mit anderen biologischen Vorgangen in Zusammenhang, welche durch Enzyme kata- lysiert werden; er erkannte, dafi das Aussen- den von Licht auf bestimmten biochemischen Reaktionen beruht [8]. Wahrend die chemischen Grundlagen dieser enzymatischen Reaktionen in den letzten Jahren weitgehend aufgeklart werden konn- ten, sind iiber den biologischen Zweck des Phanomens Vermutungen immer noch haufi- ger als gesichertes Wissen. 2. Verbreitung der Biolumineszenz im Tierreich Das biologische Leuchten hat im Tierreich eine ganz erstaunliche Verbreitung, von den Einzellern bis hin zu den Knochenfischen (Osteichthyes). Nur die Amphibien, Repti- lien, Vogel und Saugetiere haben keine For- men mit Leuchtvermogen hervorgebracht. Die Verbreitung iiber die anderen Tierklas- sen erscheint ziemlich regellos (Tabelle I). Die meisten leuchtenden Tiere leben im Meer. So zeigen in Wassertiefen von mehr als 700 m bis zu 90% aller Organismen Biolumi- neszenz. Leuchtende Einzeller wurden noch in Tiefen von 7200 m nachgewiesen. Im Siifi- wasser tritt nur eine einzige Leuchtform auf, die neuseelandische Lungenschnecke Latia nerttoides, die zudem vom Land her ins Sufi- wasser eingewandert ist. Auf dem Festland finden wir einige Hundert- und Tausendfiifi- ler, Gurtelwiirmer, Schnecken und viele In- sekten mit Leuchtvermogen. Wahrend die meisten Tiere selbstleuchtend sind (primares Leuchten), besteht bei einigen Cephalopoden (Kopffufiler) und Fischen eine merkwurdige 1. Einleitung Das Phanomen, dafi lebende Organismen sichtbares Licht ausstrahlen, bezeichnet man als Biolumineszenz. Da diese Lichterschei- nungen nicht mit Temperaturanderungen verbunden sind, werden sie auch als ,,kaltes Leuchten" beschrieben. Bereits 1886 hat der Biologze in wnserer Zeit / 11. Jahrg. 1981 / Nr. 4 0 Verlag Chemie GmbH, 0-6940 Weinhetm, 1981 004J-20JX/81/0408-0097 $ 02.>0/0 97
-
Upload
uni-bayreuth -
Category
Documents
-
view
1 -
download
0
Transcript of Leuchtende Tiere: Chemie und biologische Bedeutung
Klaus H. Hoffmann: Leuchtende Tiere : Chemie und biologische Bedeutung
franzosische Physiologe Raphael Dubois aus Leuchtkafern zwei Stoffe isoliert, die im Rea- genzglas vereinigt in Gegenwart von Sauer- stoff aufleuchteten. Die hitzestabile Kompo- nente bezeichnete er als Luciferin, die hitze- labile als Luciferase. Mit dem Nachweis von Luciferin und Luciferase brachte Dubois die Erzeugung von Licht durch Tiere und Pflan- zen mit anderen biologischen Vorgangen in Zusammenhang, welche durch Enzyme kata- lysiert werden; er erkannte, dafi das Aussen- den von Licht auf bestimmten biochemischen Reaktionen beruht [8].
Wahrend die chemischen Grundlagen dieser enzymatischen Reaktionen in den letzten Jahren weitgehend aufgeklart werden konn- ten, sind iiber den biologischen Zweck des Phanomens Vermutungen immer noch haufi- ger als gesichertes Wissen.
2 . Verbreitung der Biolumineszenz im Tierreich
Das biologische Leuchten hat im Tierreich eine ganz erstaunliche Verbreitung, von den Einzellern bis hin zu den Knochenfischen (Osteichthyes). Nur die Amphibien, Repti- lien, Vogel und Saugetiere haben keine For- men mit Leuchtvermogen hervorgebracht. Die Verbreitung iiber die anderen Tierklas- sen erscheint ziemlich regellos (Tabelle I).
Die meisten leuchtenden Tiere leben im Meer. So zeigen in Wassertiefen von mehr als 700 m bis zu 90% aller Organismen Biolumi- neszenz. Leuchtende Einzeller wurden noch in Tiefen von 7200 m nachgewiesen. Im Siifi- wasser tritt nur eine einzige Leuchtform auf, die neuseelandische Lungenschnecke Latia nerttoides, die zudem vom Land her ins Sufi- wasser eingewandert ist. Auf dem Festland finden wir einige Hundert- und Tausendfiifi- ler, Gurtelwiirmer, Schnecken und viele In- sekten mit Leuchtvermogen. Wahrend die meisten Tiere selbstleuchtend sind (primares Leuchten), besteht bei einigen Cephalopoden (Kopffufiler) und Fischen eine merkwurdige
1. Einleitung
Das Phanomen, dafi lebende Organismen sichtbares Licht ausstrahlen, bezeichnet man als Biolumineszenz. Da diese Lichterschei- nungen nicht mit Temperaturanderungen verbunden sind, werden sie auch als ,,kaltes Leuchten" beschrieben. Bereits 1886 hat der
Biologze in wnserer Zeit / 11. Jahrg. 1981 / Nr. 4 0 Verlag Chemie GmbH, 0-6940 Weinhetm, 1981 004J-20JX/81/0408-0097 $ 02.>0/0
97

Tabelle I. Verbreitung der Biolumineszenz im Tierreich (System nach H. Wurmbach, Lehrbuch der Zoologie, Gustav Fischer, Stut tgar t 1962). UKr. =Unterkreis, Kr. = Kreis.
UKr. Actinophora Kr. Porifera Kr. Cnidaria
Kr. Crenophord
Kr. Nemertini Kr. Mollusca
Kr. Annelida
Kr. Arthropod2
Kr. Heniichordata Kr. Echinodermata
UKr. Tunicatd
UKr. Vertebrard
Klasse: Radiolaria Calcarea Hydrozoa Scyphozoa Anthozoa Tentaculata Atentaculata Enopla Gastropoda Bivalvia Cephalopoda (aufler Octopoda) Polychaeta Oligochaeta Pycnogonida Crustacea Diplopoda Chilopoda Insecta (Collembola, Hemiptera, Coleoptera, Diptera) Enteropneusta Crinoidea O p hiuroidea Asteroidea Holothurioidea Larvacea Thaliacea Chondrichthyes Osteichthyes
Synibiose mit leuchtenden Bakterien (sekun- dares Leuchten oder Fremdleuchten).
3 . Biochemie des Leuchtens
Obwohl es verschiedene Typen von Leuchr- enzymen (Luciferasen) gibt, scheint allen Biolumineszenzerscheinungen derselbe fun- damentale Reaktionsmechanismus zugrunde zu liegen: eine durch Enzyme katalysierte Oxidation unter Verbrauch von Leuchtstof- fen (Luciferine oder ein langkettiger aliphati- scher Aldehyd). Die Leuchtreaktionen laufen uberwiegend intrazellular ab, nur selten wer- den die Leuchtstoffe und Luciferasen aus Se- kretdrusen in das umgebende Medium abge- geben (extrazellulares Leuchten). Die Ab- strahlung von ,,kaltem Licht" im sichtbaren Wellenlangenbereich beruht physikoche- misch auf der direkten Umwandlung von chemischer Energie in elektronische Anre- gungsenergie; Valenzelektronen von minde- stens einer an der Reaktion beteiligten Atom- oder Molekulart werden in einen angeregten
Zustand uberfuhrt (P':., angeregter Singulett- oder Triplettzustand)'> [4]. Die Anregungs- energie kann bei Ruckkehr der betreffenden Valenzelektronen in den Grundzustand (So) in Form von sichtbarem Licht ausgestrahlt werden (Abbildung 1). Die Reaktionen sind irreversibel und extrem exergonisch. Die Biolumineszenz ist von den Erscheinungen der Fluoreszenz und der Phosphoreszenz da- durch abgegrenzt, daf3 in diesen beiden Fal- len das Molekul erst durch Absorption von Licht in den angeregten Zustand hoherer Energie gehoben wird.
Da das Spektrum des ausgestrahlten Lichtes leuchtender Tiere uberwiegend im Wellen- langenbereich von 400-600 nm liegt, mug nach der bekannten Formel E = h . c/h bei der Reaktion eine Energie von 188-293 kJ (45 bis 70 kcal) pro Formelumsatz in moglichst einem Reaktionsschritt umgesetzt werden
'>s. bei Abbildung 1.
[4]. Dies stellt insofern eine Besonderheit dar, als in biologischen Systemen die Ener- gieiibertragung gewohnlich in Schritten von 3 3 4 2 kJ (8-10 kcal) erfolgt, entsprechend et- wa der Energie, die bei der Hydrolyse von 1 Mol ATP freigesetzt wird. Eine Quantenaus- beute von 1 wird bei der Leuchtreaktion dann erreicht, wenn jedes reagierende Mole- kul ein Lichtquant hervorbringt. Tatsachlich erfolgt die Lichtemission in der Biolumines- zenz mit hohen Quantenausbeuten. Bei fast allen bekannten Biolumineszenzreaktionen handelt es sich um eine ,,Abbau"-Chemilu- mineszenz. Beim Abbau der Leuchtstoffe treten Peroxide oder Dioxane als Zwischen- produkte auf, an denen der Elektronentrans- fer stattfindet [6]. N u r das Leuchten der Bak- terien weist einen vollig anderen Reaktions- mechanismus auf.
Die Leuchtreaktion bei Bakterien
Leuchtende Bakterien sind Dauerleuchter und kommen frei oder als Symbionten vor. Haufig leuchtet nur die symbiontische Form einer Art, nicht aber die freilebende. Die wichtigsten leuchtenden Gattungen sind Photobacterium, Beneckea und Vibrio. Ob- wohl die verschiedenen Arten unterschiedli- che, z. T. membrangebundene Luciferasen besitzen, ist die Leuchtreaktion doch uberall die gleiche. Die Reaktion benotigt reduzierte Flavinmononucleotide (FMNHZ = Ribofla- vin-5-phosphat reduz.) und einen langketti- gen aliphatischen Aldehyd zur Energiezu- fuhr (Abbildung 2). Nach der Oxidation durch molekularen Sauerstoff emittiert das enzymgebundene angeregte Riboflavin-5- phosphat beim Obergang in den Grundzu- stand und der Freisetzung der Luciferase Licht von I,,, = 490 nm. Die Quantenaus- beute betragt hier nur 0,l-0,3. Die Leistung vieler Leuchtbakterien liegt bei 103-106 Pho- tonen pro Sekunde und Zelle; das bedeutet, dai3 etwa 20% des verbrauchten Sauerstoffs fur die Leuchtreaktion aufgewendet werden.
Alle anderen bekannten Leuchtreaktionen benotigen Luciferine als Leuchtstoff. Dabei ist das Leuchten stets intermittierend, die Leuchtdauer kann allerdings sehr unter- schiedlich sein (einige Millisekunden bis mehrere Minuten).
An ausgewahlten Beispielen sollen verschie- dene Mechanismen von Luciferin/Lucifera- se-Reaktionen naher erlautert werden. Er- ganzende Daten dazu finden sich in Abbil- dung 2.
98 Biologie in unserer Zeit / 11. Jahrg. 1981 / Nr. 4

Biologic in unsevev Zeit / 11. Jahrg. 1981 / Nv. 4 99

Luciferin vom Coelenteraten-Typ
Das Coelenteraten-Typ-Luciferin (Abbil- dung 3) findet man in 5 Tierkreisen, den Cni- daria (Anthozoa und Hydrozoa), Ctenopho- ra, Mollusca, Arthropoda (Crustacea) und Chordata (Pisces). Der bekannteste Vertreter unter den leuchtenden Anthozoen ist das Seestiefmutterchen, Renilla reniformis. Auf einen taktilen oder elektrischen Reiz hin lau- fen konzentrische Leuchtwellen (h,,, = 509 nm) mit einer Geschwindigkeit von etwa 8 cm/sec (bei 21°C) uber den Tierstock. Das Leuchten ist intrazellular. Aus den Tieren wurden vier Proteine isoliert, die an der Leuchtreaktion beteiligt sind: eine monome- re Luciferase mit einem Molekulargewicht von 35 000, eine Luciferin-Sulfokinase, ein luciferinbindendes Protein mit einem Mole- kulargewicht von 16 500 und zwei Ca'O-Bin- dungsstellen, sowie schliefllich ein dimeres Grun-Fluoreszenz-Protein (GFP) mit einem Molekulargewicht von 55 000. Der von Cor- mier und Mitarbeitern (1974) [3] im in vitro- Versuch gefundene Reaktionsablauf ist in Abbildung 2 wiedergegeben. Die hohe Pho- tonenausbeute bei A,,, = 509 nm wird erst durch die Gegenwart des GFP ermoglicht, in Abwesenheit des GFP erhalt man nur 1/10 der Photonenausbeute mit h,,, = 480 nm. In vivo sind das proteingebundene Luciferin, die Luciferase und das GFP durch eine Mem- bran von den Ca2m-Ionen getrennt, d.h. in Vesikel eingeschlossen, die man als Lumiso- men bezeichnet. Den CaZm-Ionen kommt ei- ne steuernde Funktion zu, vergleichbar der bei der Aktivierung der Wirbeltierskelett- muskulatur (vgl. BIUZ 11/1, 7-14, 1981).
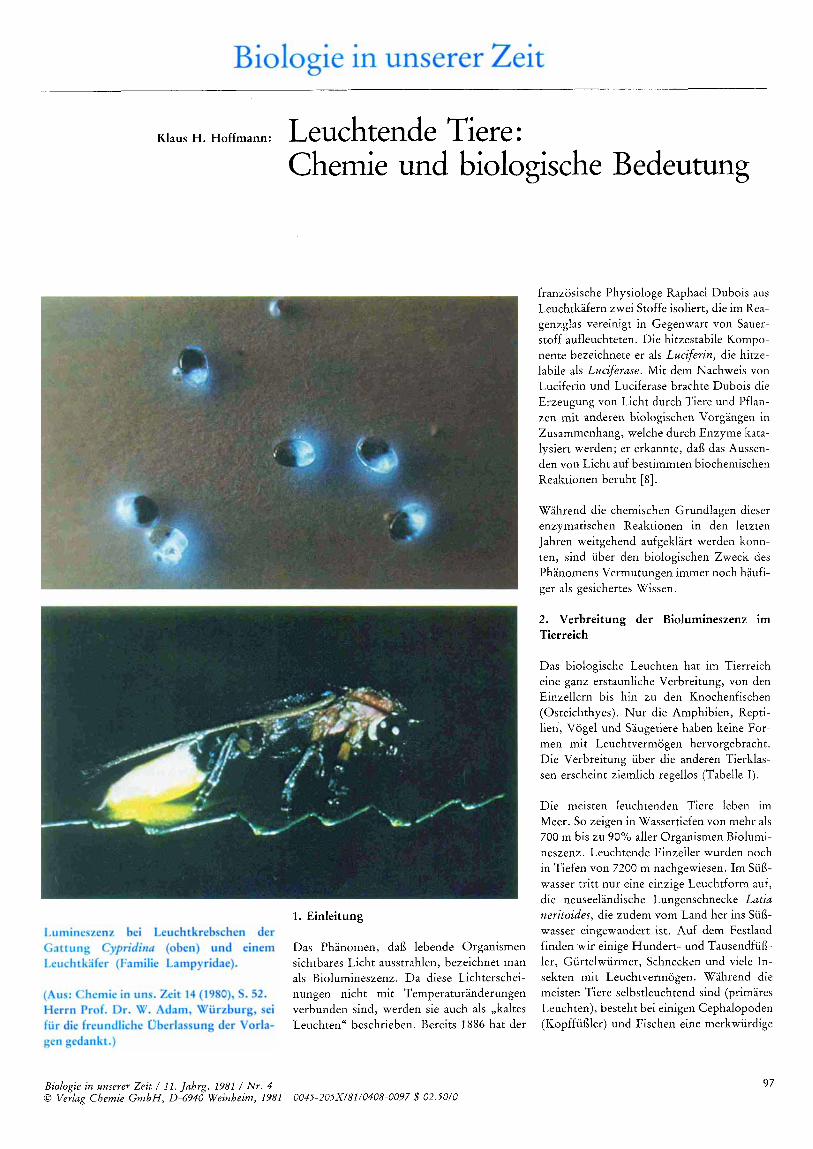
Einen ahnlichen Leuchtreaktionsmechanis- mus wie bei Renilla findet man bei den Leuchtkrebschen der Gattung Cypridina (Ostracoda, Cypridinidae, s . Abbildung S. 97); Substrate und Enzyme sind zwischen beiden Typen (Abbildung 3) aber nicht aus- tauschbar.
Im Gegensatz zu den Anthozoen benotigen Hyrozoen und Ctenophoren keinen freien Sauerstoff fur die Leuchtreaktion. Die Funk- tion des Bindungsproteins und der Luciferase werden von einem Photoprotein wahrge- nommen. In Abwesenheit von CaZe-Ionen wirkt das Photoprotein als luciferinbinden- des Protein, in Anwesenheit von Ca2*-Ionen als Luciferase. Die Bindung von Ca2" indu- ziert eine Konformationsanderung am Pro- tein und damit eine Funktionsanderung. Der Ablauf der Leuchtreaktion ist in Abbildung 2
aufgezeichnet, Substrate und Endprodukte sind die gleichen wie bei den Anthozoen. Der Wert h,,, liegt fur die in vitro-Reaktion bei 469 nm. Aus der Hydrozoe Aquorea wurde als Photoprotein das Aquorin isoliert, ein monomeres Protein mit einem Moleku- largewicht von 20000 und 3 Ca2e-Bindungs- stellen pro Molekul. Die meisten Hydrozo- en, nicht aber die Ctenophoren, enthalten zusatzlich ein Griin-Fluoreszenz-Protein, das in vivo zu einer spektralen Verschiebung nach 490-516 nm fuhrt.
Chemie der Leuchtkafer-Lumineszenz
Bei Leuchtkafern (Abb. auf S. 97) lauft die Lumineszenz stets intrazellular ab. Die Bio- chemie des Leuchtens ist fur alle echten Leucht- kafer aus der Familie Lampyridae die gleiche (Abb. 2 ) . Das Leuchtkafer-Luciferin (LH,) (Abb. 3) und eine Luciferase (alle Leuchtka- ferluciferasen sind dimere Proteine mit einem Molekulargewicht von ca. 50000 pro Unter- einheit) bilden rnit Mg . ATP einen Enzym- Luciferin- AMP-Komplex und anorganisches Pyrophosphat. Der Enzym-Luciferin-AMP- Komplex wird durch Sauerstoff oxidiert, wo- bei ein Enzymprodukt-Komplex in den an- geregten Zustand ubergeht und COZ freige- setzt wird. Der Oxidationsprozei3 ist in Abbildung 4 wiedergegeben. Radioaktivmar- kierungsversuche mit gleichzeitiger Verwen- dung von Sauerstoffisotopen "0 und "0 ha- ben gezeigt, dafl wohl kein Sauerstoffatom des gebildeten CO' aus dem molekularen Sauerstoff stammt, mindestens eines kommt aus dem Wasser [6]. Der angeregte Komplex zerfallt beim Obergang in den Grundzu- stand, wobei Licht im Wellenlangenbereich von 490-630 nm emittiert wird. Unterschie- de in der Wellenlange der Lichtemission be- ruhen auf unterschiedlichen Luciferasen in den verschiedenen Leuchtkaferarten. Aber auch innerhalb einer Art konnen zwei ver- schiedene Luciferasen vorhanden sein. So leuchtet der Automobilkafer (Pyrophorus at- lanticus) in zwei verschiedenen Farben, was auf Unterschieden in der Struktur der Lucife- rasen beruht [8]. Zusatzlich entstehen Unter- schiede im Lichtemissionsspektrum durch Veranderungen des pH-Wertes in den Leuchtzellen. Der pH-Wert der Leuchtzel- len beeinflufit auch die Quantenausbeute. Im schwach alkalischen Bereich ist die Quanten- ausbeute bei Leuchtkafern nahezu 1.
In Neuseeland und Australien treten hohlen- bewohnende leuchtende Larven von Pilz- mucken (Mycetophilidae) (Arachnocampa
luminosa und A. richardsae) in grol3er Zahl auf. Das Maximum ihrer Lichtemission liegt bei 488 nm, also vie1 weiter im blauen Be- reich als bei Leuchtkafern. Die Leuchtreak- tion wird aber ahnlich der bei Leuchtkafern durch ATP, Mg2' und Sauerstoff stimuliert. Das Luciferin der Pilzmucken ist noch nicht identifiziert [5].
Chemie der Dinofiagellaten-Lumineszenz
Obwohl nicht alle Dinoflagellaten zu den tie- rischen Organismen gezahlt werden konnen, wollen wir ihr Leuchten hier besprechen. Di- noflagellaten treten besonders in tropischen Gewassern in groi3en Mengen auf (lo4-lo7 IndividuedLiter) und verursachen einen Teil des sogenannten ,, Meeresleuchtens ", die ,,Milchsee" und eine Form der ,,red tide". Je- des Einzelindividuum sendet nach Stimula- tion, z. B. durch Wasserbewegung, Lichtblit- ze (h,,, = 470 nm) sehr kurzer Dauer (Milli- sekunden-Bereich) aus. Oft werden die Lichtblitze durch ein Dauergluhen uberla- gert. Bei fast allen Arten weist das Leucht- vermogen eine circadiane Rhythmik auf. Die Dinoflagellaten-Luciferase hat ein Moleku- largewicht von 130000. Eine Erhohung auf ca. 400000 im basischen pH-Bereich beruht auf einer Anlagerung anderer Zellproteine. Das Enzym wird dadurch inaktiviert. Eine Erniedrigung des pH-Wertes auf pH 6,7 setzt nicht nur die Luciferase wieder frei, sondern auch das an ein Bindungsprotein ge- bundene Luciferin und macht somit erst den fur die Leuchtreaktion notwendigen Oxida- tionsprozefi moglich. Eine Modellvorstel- lung uber die pH-Wert-Aktivierung ist in Abbildung 5 wiedergegeben. Dieses Model1 setzt die Anlagerung von Luciferase und Bin- dungsprotein an eine intrazellulare Membran voraus. Tatsachlich wurden aus den Einzel- lern besondere Zellorganellen (Scintillons) extrahiert, die eine Sedimentationskonstante von 10000 S aufweisen (vergleiche Riboso- men 70 bzw. 80 S), was einem Partikelge- wicht von etwa lo9 entspricht.
4. Morphologische und physiologische An- passungen im Dienste des Leuchtens
Bildung und Funktion von Leuchtorganen
Extrazellulares Leuchten setzt das Vorhan- densein von Sekretdrusen voraus, aus denen die Leuchtreaktionsstoffe auf einen Reiz hin ins umgebende Medium abgegeben werden. Nervenfasern, die an die sekretorischen Zel- len heranziehen, konnten 2.B. durch Druck
100 Btologte in unserer Zett / 11. Jahrg. 1981 1 Nr. 4
gereizt werden. Die Sekretion kann aber auch auf einen Hormon-Reiz hin erfolgen.
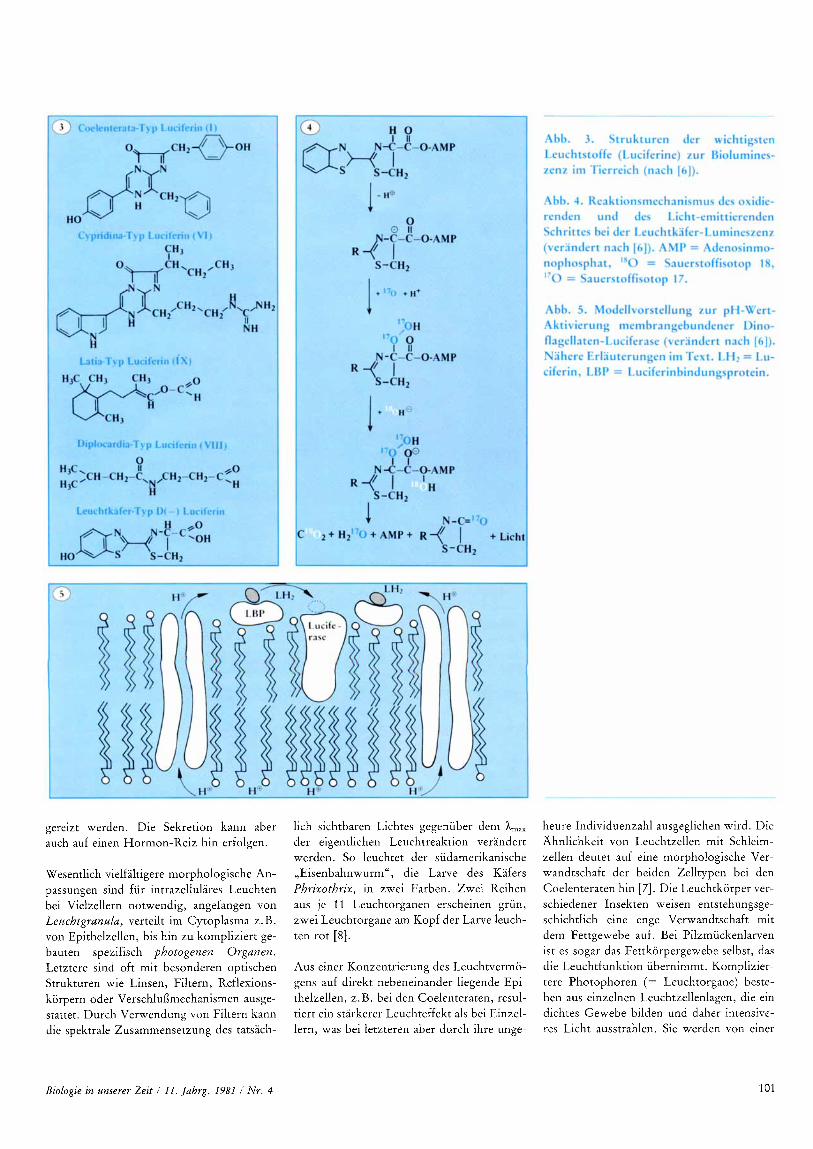
Wesentlich vielfaltigere morphologische An- passungen sind fur intrazellulares Leuchten bei Vielzellern notwendig, angefangen von Leuchtgranulu, verteilt im Cytoplasma z. B. von Epithelzellen, bis hin zu kompliziert ge- bauten spezifisch photogenen Organen. Letztere sind oft mit besonderen optischen Strukturen wie Linsen, Filtern, Reflexions- korpern oder Verschlufimechanismen ausge- stattet. Durch Verwendung von Filtern kann die spektrale Zusammensetzung des tatsach-
lich sichtbaren Lichtes gegenuber dem A,,, der eigentlichen Leuchtreaktion verandert werden. So leuchtet der siidamerikanische ,,Eisenbahnwurm", die Larve des Kafers Phrixothrix, in zwei Farben. Zwei Reihen aus je 11 Leuchtorganen erscheinen grun, zwei Leuchtorgane am Kopf der Larve leuch- ten rot [8].
Aus einer Konzentrierung des Leuchtvermo- gens auf direkt nebeneinander liegende Epi- thelzellen, z. B. bei den Coelenteraten, resul- tiert ein starkerer Leuchteffekt als bei Einzel- lern, was bei letzteren aber durch ihre unge-
heure Individuenzahl ausgeglichen wird. Die Ahnlichkeit von Leuchtzellen mit Schleim- zellen deutet auf eine morphologische Ver- wandtschaft der beiden Zelltypen bei den Coelenteraten hin [ 7 ] . Die Leuchtkorper ver- schiedener Insekten weisen entstehungsge- schichtlich eine enge Verwandtschaft mit dem Fettgewebe auf. Bei Pilzmuckenlarven ist es sogar das Fettkorpergewebe selbst, das die Leuchtfunktion ubernimmt. Komplizier- tere Photophoren (= Leuchtorgane) beste- hen aus einzelnen Leuchtzellenlagen, die ein dichtes Gewebe bilden und daher intensive- res Licht ausstrahlen. Sie werden von einer
Biologie in unserer Zeit / 1 1 . jahrg. 1981 / Nr. 4 101
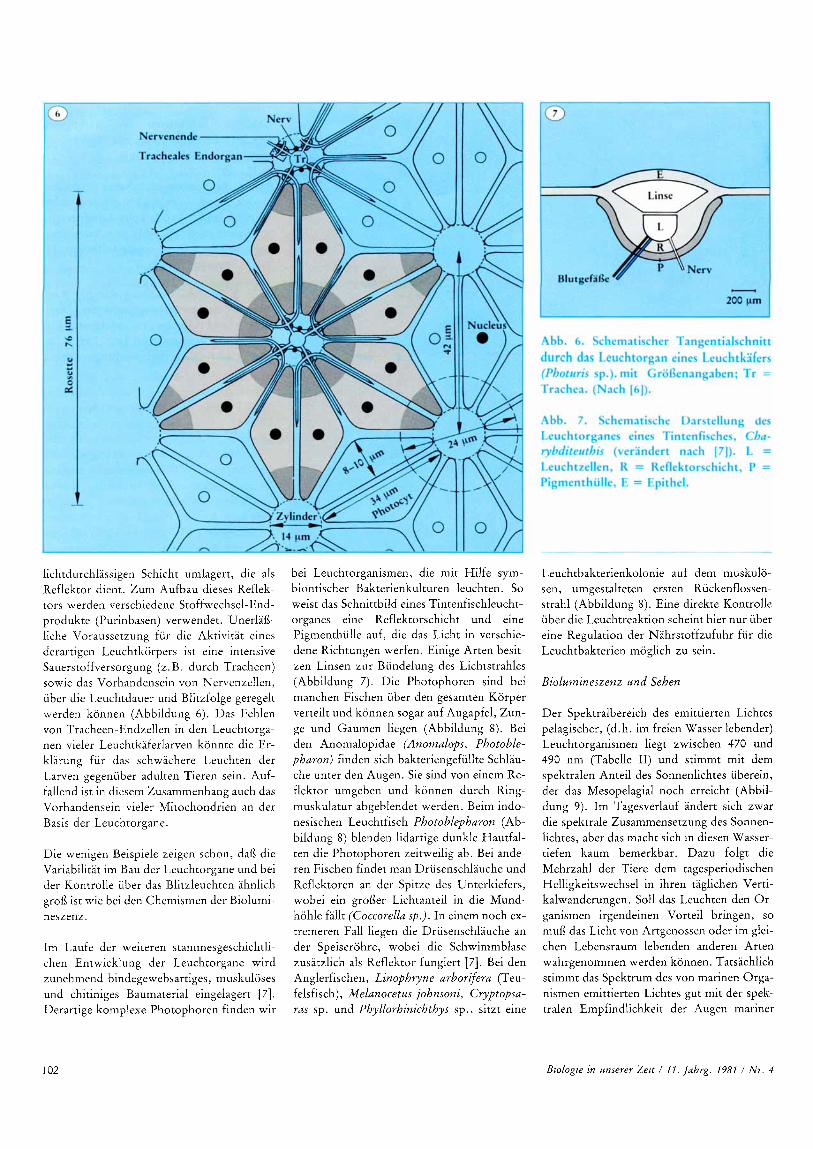
lichtdurchlassigen Schicht umlagert, die als Reflektor dient. Zum Aufbau dieses Reflek- tors werden verschiedene Stoffwechsel-End- produkte (Purinbasen) verwendet. Unerlai3- liche Voraussetzung fur die Aktivitat eines derartigen Leuchtkorpers ist eine intensive Sauerstoffversorgung (z. B. durch Tracheen) sowie das Vorhandensein von Nervenzellen, uber die Leuchtdauer und Blitzfolge geregelt werden konnen (Abbildung 6 ) . Das Fehlen von Tracheen-Endzellen in den Leuchtorga- nen vieler Leuchtkaferlarven konnte die Er- klarung fur das schwachere Leuchten der Larven gegenuber adulten Tieren sein. Auf- fallend ist in diesem Zusammenhang auch das Vorhandensein vieler Mitochondrien an der Basis der Leuchtorgane
Die wenigen Beispiele zeigen schon, dai3 die Variabilitat im Bau der Leuchtorgane und bei der Kontrolle iiber das Blitzleuchten ahnlich groi3 ist wie bei den Chemismen der Biolumi- neszenz.
Im Laufe der weiteren stammesgeschichtli- chen Entwicklung der Leuchtorgane wird zunehmend bindegewebsartiges, muskuloses und chitiniges Baumaterial eingelagert [7]. Derartige komplexe Photophoren finden wir
bei Leuchtorganismen, die mit Hilfe sym- biontischer Bakterienkulturen leuchten. So weist das Schnittbild eines Tintenfischleucht- organes eine Reflektorschicht und eine Pigmenthulle auf, die das Licht in verschie- dene Richtungen werfen. Einige Arten besit- Zen Linsen zur Bundelung des Lichtstrahles (Abbildung 7). Die Photophoren sind bei manchen Fischen iiber den gesamten Korper verteilt und konnen sogar auf Augapfel, Zun- ge und Gaumen liegen (Abbildung 8). Bei den Anomalopidae (Anornulops, Photoble- pharon) finden sich bakteriengefullte Schlau- che unter den Augen. Sie sind von einem Re- flektor umgeben und konnen durch Ring- muskulatur abgeblendet werden. Beim indo- nesischen Leuchtfisch Photoblepharon (Ab- bildung 8 ) blenden lidartige dunkle Hautfal- ten die Photophoren zeitweilig ab. Bei ande- ren Fischen findet man Drusenschlauche und Reflektoren an der Spitze des Unterkiefers, wobei ein groaer Lichtanteil in die Mund- hohle fallt (Coccorella sp.). In einem noch ex- tremeren Fall liegen die Drusenschlauche an der Speiserohre, wobei die Schwimmblase zusatzlich als Reflektor fungiert [7]. Bei den Anglerfischen, Linophryne arborifera (Teu- felsfisch), Melanocetus johnsoni, Cryptopsa- ras sp. und Phyllorhinichthys sp., sitzt eine
Leuchtbakterienkolonie auf dem muskulo- sen, umgestalteten ersten Ruckenflossen- strahl (Abbildung 8). Eine direkte Kontrolle uber die Leuchtreaktion scheint hier nur iiber eine Regulation der Nahrstoffzufuhr fur die Leuchtbakterien moglich zu sein.
Biolurnineszenz und Sehen
Der Spektralbereich des emittierten Lichtes pelagischer, (d. h. im freien Wasser lebender) Leuchtorganismen liegt zwischen 470 und 490 nm (Tabelle 11) und stimmt mit dem spektralen Anteil des Sonnenlichtes uberein, der das Mesopelagial noch erreicht (Abbil- dung 9). Im Tagesverlauf andert sich zwar die spektrale Zusammensetzung des Sonnen- lichtes, aber das macht sich in diesen Wasser- tiefen kaum bemerkbar. D a m folgt die Mehrzahl der Tiere dem tagesperiodischen Helligkeitswechsel in ihren taglichen Verti- kalwanderungen. Soil das Leuchten den Or- ganismen irgendeinen Vorteil bringen, so mui3 das Licht von Artgenossen oder im glei- chen Lebensraum lebenden anderen Arten wahrgenommen werden konnen. Tatsachlich stimmt das Spektrum des von marinen Orga- nismen emittierten Lichtes gut mit der spek- tralen Empfindlichkeit der Augen mariner
102 Biologie in unserer Zeit / 11. Jahrg. 1981 I Nr. 4

Organismen uberein (Tabelle 11). L)ie maxi- male Absorption der Sehfarbstoffe liegt z. B. in den Augen von Tintenfischen (mit Rho- dopsinen) bei 493 nm, bei den meisten Ku- stenfischen (Porphyropsine) bei 500 nm und bei Tiefseefischen (Chrysopsine) bei 475490 nm. Terrestrische Insekten weisen drei Maxi- ma in ihrer spektralen Empfindlichkeit auf, 340-365 nm, 410450 nm und 510-567 nm. Auch hier stimmt die spektrale Zusammen- setzung des ausgesandten Lichtes (540-570 nm) mit einem Maximum der spektralen Empfindlichkeit ihrer optischen Sinnesorga- ne uberein. Bei der Lumineszenzstrahlung handelt es sich um inkoharentes Licht, d. h. bei der Ausbreitung gehen erhebliche Men- gen durch Absorption und Streuung verlo- ren. Theoretisch genugt das Auftreffen eines Lichtquants zur Erregung einer Photorezep- torzelle, tatsachlich werden aber z.B. fur die
103

Erregung eines Stabchens im Wirbeltierauge wenigstens 10 Lichtquanten benotigt. Trans- parente Linsen, groi3e Okulardurchmesser, groi3e Pupillen und besondere Anpassungen im Aufbau der Retina fiihren besonders bei Tiefseefischen zu einer hohen Empfindlich- keit der Lichtsinnesorgane. Damit erreichen Leuchtorganismen im Wasser Sehweiten (d. h. Entfernungen, aus denen sich die Tiere einer Art noch gegenseitig erkennen) von etwa 50 cm bei kleinen Euphausiaceen, iiber 10-40 m bei
verschiedenen Tiefseefischen, bis zu 140 m bei der Rippenqualle Beroe. Die optischen Sinnesorgane von Leuchtkafern sind stets vom Typ des Superpositionsauges, d. h. die Lichtausnutzung ist maximalisiert; sie weisen besonders gut entwickelte, doppelschichtige Rhabdomeren auf.
5 . Biologische Funkt ion des Leuchtens
Da wir nur wenige sichere Aussagen uber die
Tabelle IIa. Spektrale Emission u n d Strahlungsintensitaten tierischer Leuchtorgane (nach [6]).
Tierart h max (nm) Lichtflui3 in 1 m Entfernung pW/cm2 Rezeptoroberflache
Dinoflagellata Gonyaulax
Hydromedusa Halistaura
Ctenophora Mnemiopsis
Crustacea Acanthephyra
Insecta Photuris Pyrophorus
Tunicata Pyrosoma
Pisces Photoblepharon
470480 2 x J/Blitz
465 1 o - ~ pW/cm’
490 3 ~ 1 0 - ’ - 6 x l O - ~
biologische Bedeutung des tierischen Leuch- tens machen konnen, ist die Zahl der aufge- stellten Theorien entsprechend groi3. Grund- satzlich sol1 das Leuchtvermogen bei Bakte- rien eine temperaturabhangige Teilerschei- nung der Zellatmung sein [7]. Bei Mutanten leuchtender Bakterien geht diese Leuchtfa- higkeit oft verloren.
Hell leuchtende Photophoren in der Nahe der optischen Sinnesorgane und auf der Ven- tralseite nutzen eine Anzahl von Tintenfi- schen und Fischen als Suchlichter beim Auf- spuren von Beute. Ventrale Leuchtorgane mit schwachem ,,Dauerlicht“ konnen zur Tarnung der Silhouette dienen (2.B. Argyro- pelecus, Abbildung 8). Da Licht wahrend der Tagesstunden im Wasser von oben nach un- ten gerichtet ist, sehen Tiere von unten be- trachtet gegen den hellen Hintergrund schwarz aus. Viele Fische jagen ihre Beute aber gerade von unten her, was an den auf- warts gerichteten Augen zu erkennen ist. Ventrale Photophoren verhelfen zusammen mit der Pigmentierung, der Reflexionsfahig- keit der Epidermis und der Transparenz be- stimmter Korperstellen zu einer Tarnung der Silhouette.
Naheliegend ist es, dem tierischen Leuchten eine Signalbedeutung zuzuschreiben.
456463 8 x 10-6
552 1-7 x 10-6 538-540 2 x 1 0 - ’ ~ 7 x lo-’
480,500 8 x 10-’-4 x
490 4 x 10-4
Tabelle I1 b. Maximale Photosensitivitat der optischen Sinnesorgane leuchtender Tiere (nach [ h ] ) .
Tierart Tintenfische Euphauside Krebse Dekapode Krebse Tiefseefische Oberflachenfische Kus tenfische Brackwasserfische Insekten
Wellenlangenbereich h (nm) 470-577 460-480 470-516 478-492 486-499 497-500 504-512 340-365, 410-450, 510-567
Signalwirkung bei marinen Organismen
Die Leuchtorgane der Bohrmuschel Pholas ebenso wie die Schwanzorgane der Laternen- fische (Myctophidae) senden kurze, helle Lichtblitze aus, die einen Rauber blenden und so dem Beuteobjekt zur Flucht verhel- fen. Einige Tintenfische, Copepoden und Ostracoden stoi3en bei Gefahr eine Leucht- wolke in das Wasser aus, verbunden mit ei- ner Fluchtreaktion. Bei einigen Laternenfi- schen besitzt nur das Mannchen ein caudales Leuchtorgan. Vom Mannchen ausgeschickte Lichtblitze lenken einen Rauber vom im Hinblick auf die Reproduktion ,,wertvolle- ren“ Weibchen ab. Leuchtsignale konnen als Warnsignale gegeniiber einem potentiellen Rauber dienen, z.B. vor der Ungeniei3bar- keit einer Qualle. Andere Arten nutzen diese Erscheinung aus, indem sie dieses Leuchtmu- ster nachahmen (Mimikri), um selbst ge- schiitzt zu sein. Die Anziehungskraft kleiner Lichtquellen mit geringer Intensitat nutzen einige Fische mit schlechten Schwimmeigen- schaften und wenig ausgebildeten Sinnesor- ganen zur Koderung von Nahrung aus, so 2.B. die schon erwahnten Anglerfische. Im
104 Biologie in unserer Zeit / 11. Jahrg. 1981 / Nr. 4
freien Wasser der Tiefsee gibt es kaum Mog- lichkeiten zur Abgrenzung eines Territo- riunis. Licht kann zur Schaffung einer noti- gen Distanz zwischen Individuen z.B. bei der Futtersuche oder zur Abgrenzung einer Population helfen (Ausschlufl-Prinzip). Vie- len Fischen, Tintenfischen und decapoden Krebsen verhilft die Lumineszenz uber ein intraspezifisches Erkennungsmuster auch bei der semipermanenten Bildung von Wander- oder Brutschwirnien [ 111.
Besonders deutlich wird die funktionelle Be- deutung der Bioluniineszenz, wenn das Leuchten in das generelle Verhaltensmuster eines Tieres eingebaut ist, etwa bei Signalen Zuni gegenseitigen Auffinden der Geschlech- ter (Sexualkommunikation). Ein diesbezug- lich bemerkenswertes Verhaltensmuster zeigt der marine Borstenwurm Odontosyllus eno- pla: Das Weibchen konimt aus einer Tiefe von 5 4 in an die Wasseroberflache und schwimmt im Kreis umher, wobei es Licht- blitze definierter Dauer aussendet. Etwa 25 Seltunden spater beginnt das Mannchen, ebenfalls blinkend, auf diesen Kreis zuzu- schwimmen. Sobald die Mitte des leuchten- den Kreiscs erreicht ist, stofit das Weibchen die Eier aus, und sie werden vom Mannchen
besamt. Das erstaunliche ist, daR die Weib- chen nur am 3 . Tag nach Vollniond und stets 54+5 Minuten nach Sonnenuntergang an die Oberflache kommen.
Sexualkommunikation bei Leuchtkafern
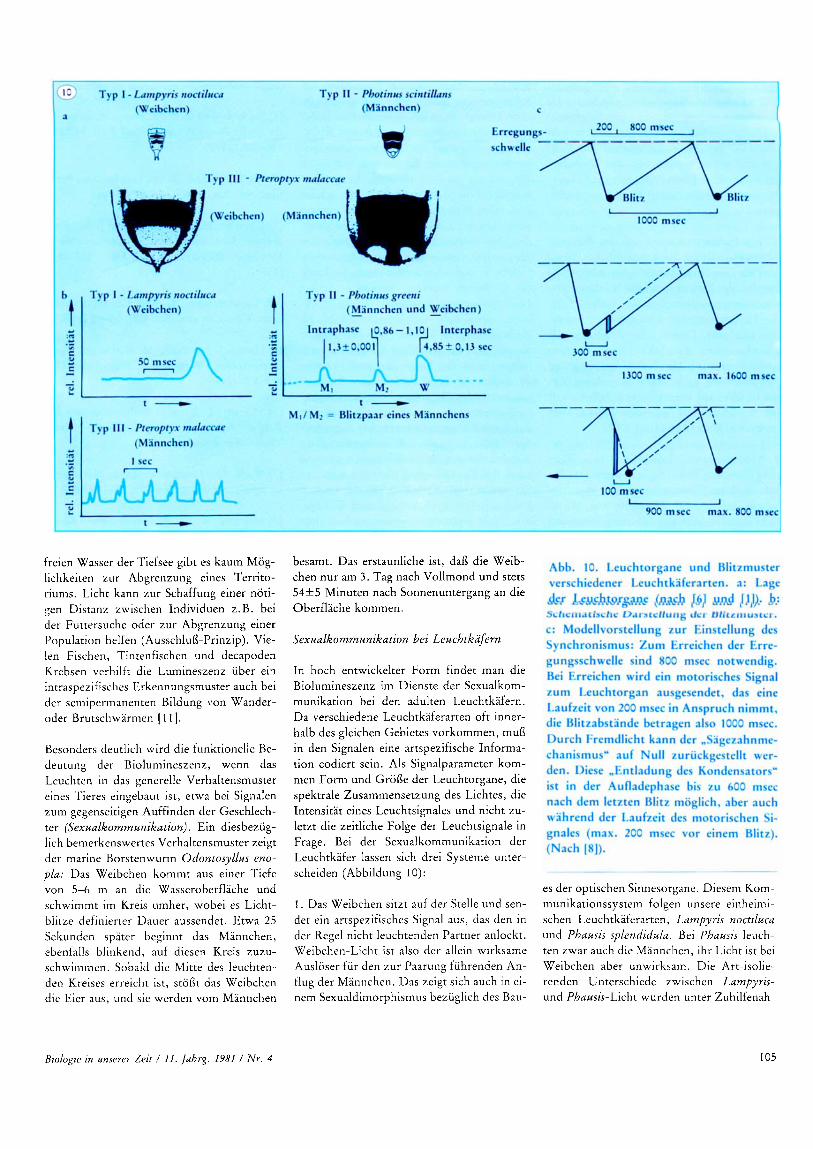
In hoch entwickelter Form findet man die Biolumineszenz im Dienste der Sexualkom- munikation bei den adulten Leuchtkafern. Da verschiedene Leuchtkaferarten oft inner- halb des gleichen Gebietes vorkommen, mufl in den Signalen eine artspezifische Informa- tion codiert sein. Als Signalparameter kom- men Form und Grofle der Leuchtorgane, die spektrale Zusammensetzung des Lichtes, die Intensitat eines Leuchtsignales und nicht zu- letzt die zeitliche Folge der Leuchtsignale in Frage. Bei der Sexualkommunikation der Leuchtkafer lassen sich drei Systeme unter- scheiden (Abbildung 10):
1. Das Weibchen sitzt auf der Stelle und sen- det ein artspezifisches Signal aus, das den in der Regel nicht leuchtenden Partner anlockt. Weibchen-Licht ist also der allein wirksame Ausloser fur den zur Paarung fuhrenden An- flug der Mannchen. Das zeigt sich auch in ei- nem Sexualdimorphismus bezuglich des Bau-
es der optischen Sinnesorgane. Diesem Koni- munikationssystem folgen unsere einheinii- schen Leuchtkaferarten, Lampyris noctiluca und Pbausis splendidula. Bci Phausis leucli- ten zwar auch die Miinnchen, ihr Licht ist bei Weibchen aber unwirltsam. Die Art-isolie- renden Unterschiede zwischen Lampyris- und Phausis-Licht wurden unter Zuhilfenah-
Biologic in unserer Zett / 11. jahrg. 1981 / Nr. 4 105

me von Leuchtattrappenversuchen bestatigt [lo]. Lampyris-Mannchen verwechseln das Leuchtmuster von Phausis- und Lampyris- Weibchen nicht, Verwechslungen kommen allerdings bei Phausis-Mannchen vor, art- fremde Partner werden von den Weibchen gefressen. Eine Oberproduktion an Phausis- Mannchen scheint diesen Mangel auszuglei- chen.
2. Ein Partner, in der Regel das fliegende Mannchen, sendet ein artspezifisches Signal aus, das aus Einzelblitzen oder Blitzpaaren besteht, der Partner antwortet wiederum mit einem artspezifischen Signal (Abbildung lob); der Hochzeitswerber wird auf die Antwort hin angelockt. Dieses Schema gilt fur die amerikanischen Leuchtkaferarten Photuris und Photinus [ 11. Die Signalerkennung und Art-Isolierung zwischen verschiedenen Ar- ten beruhen auf artspezifischen Zeitinterval- len zwischen dem mannlichen und dem weiblichen Blitz (Abbildung lob).
3. Sudostasiatische Leuchtkafer (u. a. Gat- tung Pteroptyx) haben Signalsysteme entwik- kelt, in denen Elemente aus beiden Systemen kombiniert sind [2]. Hunderte von Mann- chen sitzen im Schwarm auf einem Baum und zeigen rhythmischen Blitzsynchronismus. So senden P. cribellata Mannchen (25°C) alle 1000 rfr 20 msec einen Blitz von 150 msec Dauer aus. Das Problem liegt hier in der not- wendigen exakten Zeitmessung durch das Zentralnervensystem. Bisher konnte zwar weder ein Schrittmacher noch ein Oszillator im Gehirn der Tiere eindeutig identifiziert werden, indirekt wurde ihre Existenz aber durch Verhaltensversuche bestatigt, die in Abbildung 1Oc erlautert sind. Wahrend ein Einzeltier bei diesen Versuchen schnell in den Synchronismus einfallt, braucht eine Ko- lonie 10-15 Zyklen. Dies entspricht durchaus dem langsam einsetzenden Synchronismus der Mannchen auf einem Baum in der Abenddammerung. Jedes Individuum profi- tiert vom synchronen Blitzen des Nachbarn, gemeinsames Leuchten lockt die Weibchen eher an ihre Gruppe. Das Weibchen ist durch diese Fahigkeit gegen mutante Mannchen, die auger Phase blitzen, geschutzt. Jedes in- takte Mannchen behalt seine individuelle Auswahlbarkeit und hilft gleichzeitig den Nachbarn bei der Partnerfindung.
Biolumineszenz ist in praktisch allen Tier- stammen und allen Lebensraumen entwik-
kelt, vergleichbar etwa der Entstehung von Sehpigmenten. Sicher liefen die ursprungli- chen Systeme chemisch wesentlich einfacher ab als viele Leuchtreaktionen heute. Wahr- scheinlich entstand die Biolumineszenz, be- vor Sauerstoff in der Atmosphare vorhanden war und ehe auf der Erdoberflache die Pho- tosynthese in Gang kam [8]. Mit groger Wahrscheinlichkeit diente die Biolumines- zenz dann den tierischen Organismen als ein Entg~tungsmechanismus, als beim Auftreten der ersten photosynthetischen Organismen Sauerstoff in die Atmosphare kam. Im Laufe der Evolution wurden zunehmend hellere Leuchtkoper mit besseren Kontrollmoglich- keiten entwickelt, wobei unklar bleibt, ob die Verteilung der Leuchtorgane eine primare Konsequenz der zufalligen Leuchtorganent- stehung oder die Folge adaptiver Prozesse ist. Sicher verhalf die Fahigkeit zur Biolumi- neszenz vielen Organismen bei der Besiede- lung extremer Biotope wie der Tiefsee oder der dichten Vegetation tropischer Regenwal- der. Das Beispiel der siidostasiatischen Leuchtkafer mit ihreni Blitzsynchronismus zeigt, daf3 diese Fahigkeit nicht nur einzelnen Mannchen oder Weibchen Vorteile bringen kann, sondern auch einer ganzen Kolonie. Ein solch eindeutig kooperatives Verhalten ist allerdings wieder weniger fur eine naturli- che Selektion geeignet. So bleiben noch viele Evolutions- und Anpassungsfragen bei der Biolumineszenz offen.
7. Schlui3wort
Nicht unerwahnt sol1 zum Schluf3 bleiben, dai3 sich der Mensch gerade in den letzten Jahren die Fahigkeit tierischer Organismen zur Biolumineszenz praktisch zum Nutzen macht. Das Licht, das bei der enzymatischen Oxidation von Luciferinen ausgestrahlt wird, reicht ja tatsachlich aus, um in der Dunkel- heit Landkarten oder Zeitung zu lesen.
Verschiedene Biolumineszenzreaktionen wer- den in einer Reihe von analytischen Ver- fahren zur Bestimmung kleinster Mengen an ATP, Sauerstoff, NAD(P), CaZ@-Ionen oder ATP- bzw. NAD(P)-abhangiger Enzymakti- vitaten herangezogen (Luminometrie).
Zur Einarbeitung enipfohlenr Literatur:
[ I ] Buck, J., and E. Buck: Photic signaling in the firefly Photinus greeni. Biol. Bull. 142(2), 195-205 (1972).
[2] Buck, J., and E. Buck: Synchronous kire- flies. Sci. Am. 234(5), 74-85 (1976).
[3] Cormier, M. J . , K. Hori, and J . M. An- derson: Bioluminescence in Coelenterates. Biochim. Biophys. Acta 346, 137-164 ( 1 974).
[4] Gundermann, K.-D.: Chernilurnines- zenz. Chem. in uns. Zeit 4(2), 55-60 (1970).
[5] Henry, J. P., and A. M. Michelson: Bio- luminescence. Photochem. Photobiol. 27, 855-858 (1978).
[6] Herring, P. J. (ed.): Bioluminescence in Action. Academic Press, London - New York - San Francisco 1978.
[7] Korn, H.: Leuchtende Tiere. Naturwis- senschaft und Medizin 6(27), 34-44 (1969).
[8] McElroy, W. D.: Biolumineszenz - Che- mie und biologische Bedeutung. Umsch. Wiss. Techn. 69(15), 472-474 (1969).
[9] Schramm, E. : Bioluminescence: Funda- mental and practical aspects. Arch. Int. Phy- siol. Biochim. 81, 561-605 (1973).
[ lo] Schwalb, H. H.: Beitrage zur Biologie der einheimischen Lampyriden Lampyris nocttluca und Phausis splendidula, und expe- rimentelle Analysen ihres Beutefang- und Sexualverhaltens. Zool. Jahrb. Abt. Syst. Oekol. Geogr. Tiere 88, 399-550 (1961).
[11] Tett, P. B., and M. G. Kelly: Marine bioluminescence. Oceanogr. Mar. Biol. 11, 89-173 (1973).
Lebenslauf siehe Biologie in unserer Zeit 9/1, S. 26 (1979); seit 1.10.1978 ist der Verfasser Professor fur Allgemeine Zoologie an der Universitat Ulm.
6. Evolution der Biolumineszenz Anschrift: Prof. Dr . K. H. Hoffmann, Allg. Zoologie (Biologie I) der Universitat, Oberer Eselsberg, D-7900 Ulm.
106 Btologte in unserer Zett / 11. Jahrg. 1981 / Nr. 4




















