Leishmanicidal activity assessment of olive tree extracts
7
Phytomedicine 20 (2013) 275–281 Contents lists available at SciVerse ScienceDirect Phytomedicine j ourna l ho mepage: www.elsevier.de/phymed Leishmanicidal activity assessment of olive tree extracts Joannis D. Kyriazis a , Nektarios Aligiannis b , Panagiotis Polychronopoulos b , Alexios-Leandros Skaltsounis b , Eleni Dotsika a,∗ a Laboratory of Cellular Immunology, Hellenic Pasteur Institute, 127 Vas. Sofias Avenue, 11521 Athens, Greece b Department of Pharmacognosy and Natural Products Chemistry, Faculty of Pharmacy, University of Athens, Panepistimiopolis Zografou, 15771 Athens, Greece a r t i c l e i n f o Keywords: Natural products Oleuropein Hydroxytyrosol Visceral leishmaniasis a b s t r a c t Leishmaniasis, a protozoan parasitic disease that remains a major worldwide health problem with high endemicity in developing countries, is prevalent around the Mediterranean basin. High cost, systemic toxicity, and diminished efficacy due to development of parasite resistance are the serious drawbacks of current treatment options. Thus, identifying new, effective, and safer anti-leishmanial drug(s) is of paramount importance. Here we tested the anti-promastigote and anti-amastigote activity of five natural products, including oleuropein and hydroxytyrosol, present in olive tree leaves and olive mill wastewater. These products are recognized as low-cost starting materials rich in bioactive compounds, particularly biophenols. Oleuropein and hydroxytyrosol exhibited the best inhibitory effect among the natural products tested in both stationary and middle logarithmic phase promastigotes of L. infantum, L. donovani, and L. major. Similarly, oleuropein and hydroxytyrosol demonstrated the highest selectivity index ratio against L. donovani amastigotes that parasitize J774A.1 macrophages. Moreover, oleuropein was tested in vivo in an experimental visceral leishmaniasis model. L. donovani-infected BALB/c mice received intraperitoneal oleuropein a total of 14 times at intervals of every other day. Three days after treatment termination, the spleen parasitic burden was reduced >80%. Of interest, this effect of oleu- ropein persisted and was even enhanced 6 weeks after the termination of the treatment, as determined by parasite depletion of >95% in liver and spleen. These findings contribute to the potential develop- ment of natural products as effective drugs against parasites of the Leishmania genus, with low cost and diminished cytotoxicity. © 2012 Elsevier GmbH. All rights reserved. Introduction Leishmaniasis is a vector-borne protozoan disease caused by species of the genus Leishmania. Leishmania spp. are responsible for parasitic diseases with a wide range of symptoms and consid- erable impact on public health worldwide. Leishmaniasis is still one of the world’s most neglected diseases, accounting for 1.5–2 mil- lion new cases and 50,000 deaths attributable to the visceral form annually (World Health Organization, Leishmaniasis Control home page). The persistence of and increase in leishmaniasis is mainly due to factors such as the AIDS epidemic, climatic changes, inter- national travel, migration flows, lack of effective vaccines, problems with vector control, and development of drug resistance (Dujardin et al., 2008). Treatment and control of leishmaniasis involve a limited num- ber of drugs with various adverse reactions, effectiveness, and long ∗ Corresponding author. Tel.: +30 210 6478 828; fax: +30 210 6478 828. E-mail address: [email protected] (E. Dotsika). duration of treatments. So far, chemotherapy includes pentavalent antimonials, numerous forms of amphotericin B, paromomycin (aminosidine), pentamidine isethionate, hexadecylphospho- choline (HePC; miltefosine), and azoles such as ketoconazole, fluconazole, and itraconazole (World Health Organization, Leish- maniasis Control home page). Despite the numerous treatments, none is fully satisfactory because of the toxicity of the treatment itself, a tendency to host relapse, and the development of resistant parasite strains (Kappagoda et al., 2011). All of these disadvantages are assisted by the fact that many people worldwide have no access to conventional pharmacological treatments (den Boer et al., 2011). These patients prefer to focus their efforts for therapy on folk remedies (alternative treatments) that consist basically of natural products that are considered non-toxic, but their safety and effectiveness require evaluation (Sen and Chatterjee, 2011). Screening natural products for potential use in the therapy of leishmaniasis is essential to satisfy the urgent need for alternative treatments (Rocha et al., 2005). Olives and olive oil are considered the cornerstone of the Mediterranean diet and are recognized for their overall health 0944-7113/$ – see front matter © 2012 Elsevier GmbH. All rights reserved. http://dx.doi.org/10.1016/j.phymed.2012.11.013
Transcript of Leishmanicidal activity assessment of olive tree extracts

L
JAa
b
a
KNOHV
I
sfeolapdnwe
b
0h
Phytomedicine 20 (2013) 275– 281
Contents lists available at SciVerse ScienceDirect
Phytomedicine
j ourna l ho mepage: www.elsev ier .de /phymed
eishmanicidal activity assessment of olive tree extracts
oannis D. Kyriazisa, Nektarios Aligiannisb, Panagiotis Polychronopoulosb,lexios-Leandros Skaltsounisb, Eleni Dotsikaa,∗
Laboratory of Cellular Immunology, Hellenic Pasteur Institute, 127 Vas. Sofias Avenue, 11521 Athens, GreeceDepartment of Pharmacognosy and Natural Products Chemistry, Faculty of Pharmacy, University of Athens, Panepistimiopolis Zografou, 15771 Athens, Greece
r t i c l e i n f o
eywords:atural productsleuropeinydroxytyrosolisceral leishmaniasis
a b s t r a c t
Leishmaniasis, a protozoan parasitic disease that remains a major worldwide health problem with highendemicity in developing countries, is prevalent around the Mediterranean basin. High cost, systemictoxicity, and diminished efficacy due to development of parasite resistance are the serious drawbacksof current treatment options. Thus, identifying new, effective, and safer anti-leishmanial drug(s) isof paramount importance. Here we tested the anti-promastigote and anti-amastigote activity of fivenatural products, including oleuropein and hydroxytyrosol, present in olive tree leaves and olive millwastewater. These products are recognized as low-cost starting materials rich in bioactive compounds,particularly biophenols. Oleuropein and hydroxytyrosol exhibited the best inhibitory effect among thenatural products tested in both stationary and middle logarithmic phase promastigotes of L. infantum,L. donovani, and L. major. Similarly, oleuropein and hydroxytyrosol demonstrated the highest selectivityindex ratio against L. donovani amastigotes that parasitize J774A.1 macrophages. Moreover, oleuropeinwas tested in vivo in an experimental visceral leishmaniasis model. L. donovani-infected BALB/c mice
received intraperitoneal oleuropein a total of 14 times at intervals of every other day. Three days aftertreatment termination, the spleen parasitic burden was reduced >80%. Of interest, this effect of oleu-ropein persisted and was even enhanced 6 weeks after the termination of the treatment, as determinedby parasite depletion of >95% in liver and spleen. These findings contribute to the potential develop-ment of natural products as effective drugs against parasites of the Leishmania genus, with low cost anddiminished cytotoxicity.© 2012 Elsevier GmbH. All rights reserved.
ntroduction
Leishmaniasis is a vector-borne protozoan disease caused bypecies of the genus Leishmania. Leishmania spp. are responsibleor parasitic diseases with a wide range of symptoms and consid-rable impact on public health worldwide. Leishmaniasis is still onef the world’s most neglected diseases, accounting for 1.5–2 mil-ion new cases and 50,000 deaths attributable to the visceral formnnually (World Health Organization, Leishmaniasis Control homeage). The persistence of and increase in leishmaniasis is mainlyue to factors such as the AIDS epidemic, climatic changes, inter-ational travel, migration flows, lack of effective vaccines, problemsith vector control, and development of drug resistance (Dujardin
t al., 2008).Treatment and control of leishmaniasis involve a limited num-
er of drugs with various adverse reactions, effectiveness, and long
∗ Corresponding author. Tel.: +30 210 6478 828; fax: +30 210 6478 828.E-mail address: [email protected] (E. Dotsika).
944-7113/$ – see front matter © 2012 Elsevier GmbH. All rights reserved.ttp://dx.doi.org/10.1016/j.phymed.2012.11.013
duration of treatments. So far, chemotherapy includes pentavalentantimonials, numerous forms of amphotericin B, paromomycin(aminosidine), pentamidine isethionate, hexadecylphospho-choline (HePC; miltefosine), and azoles such as ketoconazole,fluconazole, and itraconazole (World Health Organization, Leish-maniasis Control home page). Despite the numerous treatments,none is fully satisfactory because of the toxicity of the treatmentitself, a tendency to host relapse, and the development of resistantparasite strains (Kappagoda et al., 2011). All of these disadvantagesare assisted by the fact that many people worldwide have noaccess to conventional pharmacological treatments (den Boeret al., 2011). These patients prefer to focus their efforts for therapyon folk remedies (alternative treatments) that consist basically ofnatural products that are considered non-toxic, but their safetyand effectiveness require evaluation (Sen and Chatterjee, 2011).Screening natural products for potential use in the therapy of
leishmaniasis is essential to satisfy the urgent need for alternativetreatments (Rocha et al., 2005).Olives and olive oil are considered the cornerstone of theMediterranean diet and are recognized for their overall health

2 medicine 20 (2013) 275– 281
bapteltp
oatgiedrcr(ecwdarG2w(a
sOtmFv
M
C
ksr(dom(ti9oc
(le(afT
76 J.D. Kyriazis et al. / Phyto
enefits (Cicerale et al., 2009). Olive tree (Olea europaea) leavesnd olive mill wastewater (OMWW), which is a high-polluting by-roduct of olive oil production, are regarded as low-cost sourceshat are rich in phenolic compounds with strong antioxidant prop-rties (Cardinali et al., 2010). The majority of the compounds, ateast 30, are present in olive oil, and recent data have suggestedhat many of the constituents may have more health benefits thanreviously thought (Cicerale et al., 2009).
Oleuropein obtained from olive leaves and hydroxytyrosolbtained from OMWW have multiple biological properties (Tucknd Hayball, 2002; Waterman and Lockwood, 2007). Oleuropein,he main ingredient of olives and olive leaves, is a secoiridoidlucoside and belongs to the polyphenol group. Hydroxytyrosols basically found in olives and in olive oil in the form of itslenolic acid ester oleuropein, and especially after hydrolysisuring degradation, in its plain form. The major activity of oleu-opein and hydroxytyrosol is to scavenge free radicals to protectells from oxidation. Especially, the anti-oxidant ability of oleu-opein may slow ageing and reduce the risk of coronary diseaseAndreadou et al., 2006; Katsiki et al., 2007). Moreover, oleuropeinnhances the response of macrophages to bacterial lipopolysac-haride via an increase in inducible nitric oxide synthase activityhereas oleuropein by itself does not enhance nitric oxide pro-uction by macrophages (Visioli et al., 1998). Finally, oleuropeinnd hydroxytyrosol have been reported to have antitumor, antivi-al, antibacterial, and antiparasitic action (Bisignano et al., 1999;ranados-Principal et al., 2010; Jiang et al., 2008; Lee-Huang et al.,007a,b; Tuck and Hayball, 2002; Waterman and Lockwood, 2007)hereas, in the year 2000, oleuropein was claimed in a U.S. patent
Fredrickson, Num: 6,117,844) to have potent antiviral activitygainst numerous viral species.
In this study, we evaluated five natural products, two pure con-tituents and three crude extracts, derived from olive leaves andMWW, as potent in vitro and in vivo anti-leishmanial agents. For
his purpose, they were tested against promastigotes of three Leish-ania species and against L. donovani intracellular amastigotes.
inally, oleuropein was also tested in vivo against experimentalisceral leishmaniasis caused by L. donovani parasites.
aterials and methods
rude extracts and pure compounds
Air-dried and pulverized leaves (5 kg) from Olea europaea varoroneiki collected in Crete (Greece) were extracted by mechanicaltirring for 12 h with acetone (2× 2.5 l). The extract was evapo-ated completely and washed with a mixture of CH2Cl2/MeOH:98/23 l). The insoluble material (360 g) was separated and dried, pro-ucing a yellow powder containing 60% oleuropein. Ten gramsf this yellow powder was subjected to countercurrent chro-atography, using a fast centrifugal partition chromatograph
FCPC) apparatus (Kromaton). The system of solvents used inhis procedure was EtOAc/EtOH/H2O:10/1/10 (6 l). The capac-ty of the column was 1 l, the rapidity of the rotation was00 rpm, and the flow rate was 15 ml/min. A total of 5.0 g ofleuropein (purity 90%) was isolated by the above-mentioned pro-ess.
Olive leaf water extraction (OLWE) and olive leaf decoctionOLD) were derived from the same collection of Olea europaeaeaves. Olive leaf powder was extracted using the method of accel-rated solvent extraction (ASE) performed on an ASE 300 system
Dionex, USA). The extraction was conducted with deionized watert 80 ◦C and the OLWE was finally lyophilized. For the other portion,resh leaves were blended using a general-purpose electric blender.he blended material was added to water and boiled for 3 min.Fig. 1. Molecular formulas of (a) hydroxytyrosol and (b) oleuropein.
Then, the hot mixture was filtered through a filter paper, and thefiltrate was extracted and evaporated to dryness under reducedpressure to yield a solid residue.
Hydroxytyrosol was extracted from OMWW resulting fromolive oil production using olive fruits of Olea europaea var koroneiki.During the process, the material was filtered using filter paper.Then filtered wastewater was applied to Amberlite XAD-4 resinand the column eluted with 96% ethanol. Then ethanol was evap-orated and a phenolic fraction was received. The final step aimedat the purification of hydroxytyrosol. The above polyphenolic frac-tion was submitted to countercurrent chromatography using theFCPC apparatus (Kromaton). The system of solvents used in thisprocedure was c-hexane/EtOAc/MeOH/H2O:4/6/4/6 (Agalias et al.,2007). The capacity of the column was 1 l, the rapidity of therotation was 900 rpm, and the flow rate was 15 ml/min. Hydrox-ytyrosol (purity > 90%) was isolated using the above-mentionedprocess.
The purity of oleuropein, hydrotyrosol (Fig. 1) and olive leafdecoction was established by high performance liquid chro-matography (HPLC) (Fig. 2) and nuclear magnetic resonance(NMR). Significant was, the presence of oleuropein and hydroxy-tyrosol alongside with other unidentified compounds in the oliveleaves decoction. Prior to use, pure oleuropein, pure hydroxyty-rosol, and the oleuropein 60% mixture were diluted in distilledwater while the other two crude extracts, OLWE and OLD, werediluted in dimethyl sulfoxide (DMSO). All extracts were Mil-lipore filtered with a 0.45 �m filter and stored at 4 ◦C untiluse.
Parasites
Three reference Leishmania strains were used in this study:Leishmania infantum (zymodeme MON-1, strain MCAN/PT/98/IMT 244), Leishmania donovani (zymodeme MON-2, strainMHOM/IN/1996/THAK35), and Leishmania major (zymodemeLV39, strain MRHO/SU/59/P). The promastigotes were culturedin complete medium consisting of RPMI-1640 (with low contentof phenol red) (Biochrom AG, Berlin, Germany) supplementedwith 2 mM l-glutamine, 10 mM HEPES, 24 mM NaHCO3, 100 U/mlpenicillin, 100 �g/ml streptomycin, and 10% (v/v) heat-inactivatedfetal bovine serum (FBS; Gibco, Paisley, UK) at 26 ◦C. Cultureswere estimated daily for parasite growth, and promastigotes werecollected at the middle of the logarithmic or at the beginning ofthe stationary phase.
Cell cultures
Macrophages of the J774A.1 murine macrophage cell line fromATCC (American Type Culture Collection, Rockville, MD, USA) were
cultured in complete medium consisted of RPMI-1640 mediumsupplemented with 2 mM l-glutamine, 10 mM HEPES, 24 mMNaHCO3, 100 U/ml penicillin, 100 �g/ml streptomycin, and 10%heat-inactivated FBS, at 37 ◦C in a 5% CO2 environment. Subcultures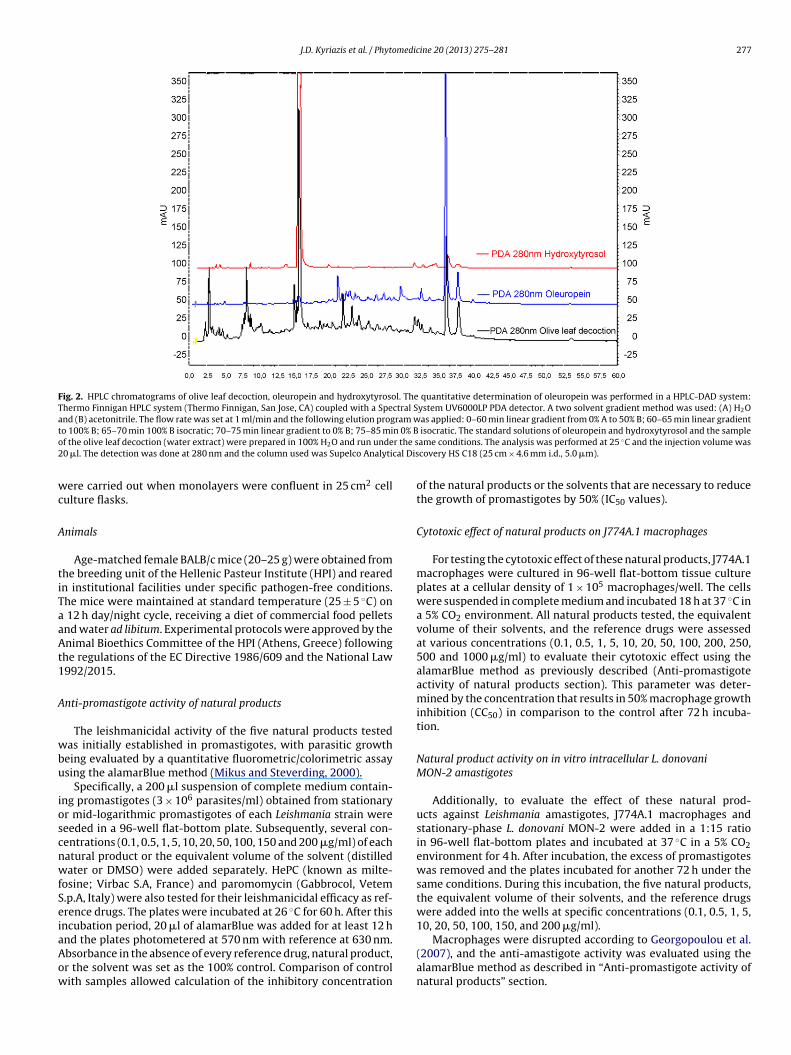
J.D. Kyriazis et al. / Phytomedicine 20 (2013) 275– 281 277
Fig. 2. HPLC chromatograms of olive leaf decoction, oleuropein and hydroxytyrosol. The quantitative determination of oleuropein was performed in a HPLC-DAD system:Thermo Finnigan HPLC system (Thermo Finnigan, San Jose, CA) coupled with a Spectral System UV6000LP PDA detector. A two solvent gradient method was used: (A) H2Oand (B) acetonitrile. The flow rate was set at 1 ml/min and the following elution program was applied: 0–60 min linear gradient from 0% A to 50% B; 60–65 min linear gradientt n 0% Bo r the s2 cal Dis
wc
A
tiTaaAt1
A
wbu
ioscnwfSeiaAow
10, 20, 50, 100, 150, and 200 �g/ml).
o 100% B; 65–70 min 100% B isocratic; 70–75 min linear gradient to 0% B; 75–85 mif the olive leaf decoction (water extract) were prepared in 100% H2O and run unde0 �l. The detection was done at 280 nm and the column used was Supelco Analyti
ere carried out when monolayers were confluent in 25 cm2 cellulture flasks.
nimals
Age-matched female BALB/c mice (20–25 g) were obtained fromhe breeding unit of the Hellenic Pasteur Institute (HPI) and rearedn institutional facilities under specific pathogen-free conditions.he mice were maintained at standard temperature (25 ± 5 ◦C) on
12 h day/night cycle, receiving a diet of commercial food pelletsnd water ad libitum. Experimental protocols were approved by thenimal Bioethics Committee of the HPI (Athens, Greece) following
he regulations of the EC Directive 1986/609 and the National Law992/2015.
nti-promastigote activity of natural products
The leishmanicidal activity of the five natural products testedas initially established in promastigotes, with parasitic growth
eing evaluated by a quantitative fluorometric/colorimetric assaysing the alamarBlue method (Mikus and Steverding, 2000).
Specifically, a 200 �l suspension of complete medium contain-ng promastigotes (3 × 106 parasites/ml) obtained from stationaryr mid-logarithmic promastigotes of each Leishmania strain wereeeded in a 96-well flat-bottom plate. Subsequently, several con-entrations (0.1, 0.5, 1, 5, 10, 20, 50, 100, 150 and 200 �g/ml) of eachatural product or the equivalent volume of the solvent (distilledater or DMSO) were added separately. HePC (known as milte-
osine; Virbac S.A, France) and paromomycin (Gabbrocol, Vetem.p.A, Italy) were also tested for their leishmanicidal efficacy as ref-rence drugs. The plates were incubated at 26 ◦C for 60 h. After thisncubation period, 20 �l of alamarBlue was added for at least 12 h
nd the plates photometered at 570 nm with reference at 630 nm.bsorbance in the absence of every reference drug, natural product,r the solvent was set as the 100% control. Comparison of controlith samples allowed calculation of the inhibitory concentrationisocratic. The standard solutions of oleuropein and hydroxytyrosol and the sampleame conditions. The analysis was performed at 25 ◦C and the injection volume wascovery HS C18 (25 cm × 4.6 mm i.d., 5.0 �m).
of the natural products or the solvents that are necessary to reducethe growth of promastigotes by 50% (IC50 values).
Cytotoxic effect of natural products on J774A.1 macrophages
For testing the cytotoxic effect of these natural products, J774A.1macrophages were cultured in 96-well flat-bottom tissue cultureplates at a cellular density of 1 × 105 macrophages/well. The cellswere suspended in complete medium and incubated 18 h at 37 ◦C ina 5% CO2 environment. All natural products tested, the equivalentvolume of their solvents, and the reference drugs were assessedat various concentrations (0.1, 0.5, 1, 5, 10, 20, 50, 100, 200, 250,500 and 1000 �g/ml) to evaluate their cytotoxic effect using thealamarBlue method as previously described (Anti-promastigoteactivity of natural products section). This parameter was deter-mined by the concentration that results in 50% macrophage growthinhibition (CC50) in comparison to the control after 72 h incuba-tion.
Natural product activity on in vitro intracellular L. donovaniMON-2 amastigotes
Additionally, to evaluate the effect of these natural prod-ucts against Leishmania amastigotes, J774A.1 macrophages andstationary-phase L. donovani MON-2 were added in a 1:15 ratioin 96-well flat-bottom plates and incubated at 37 ◦C in a 5% CO2environment for 4 h. After incubation, the excess of promastigoteswas removed and the plates incubated for another 72 h under thesame conditions. During this incubation, the five natural products,the equivalent volume of their solvents, and the reference drugswere added into the wells at specific concentrations (0.1, 0.5, 1, 5,
Macrophages were disrupted according to Georgopoulou et al.(2007), and the anti-amastigote activity was evaluated using thealamarBlue method as described in “Anti-promastigote activity ofnatural products” section.

278 J.D. Kyriazis et al. / Phytomedicine 20 (2013) 275– 281
Table 1IC50 (�g/ml) of olive tree natural products vs Leishmania promastigotes.
L. infantum MON-1 L. donovani MON-2 L. major LV-39
IC50 promastigote IC50 promastigote IC50 promastigote
Sa Lb Sa Lb Sa Lb
Natural productOleuropein 145 ± 21 191.9 ± 19.3 77.2 ± 10.5 69.4 ± 6.3 141 ± 23 –Oleuropein 60% 144 ± 32 – 84 ± 13.5 – 151.8 ± 40.4 –Hydroxytyrosol 32.9 ± 0.1 71.8 ± 12.6 60.5 ± 7.2 75.5 ± 0.6 50.1 ± 4.8 105 ± 13OLDc 113 ± 25.5 – 60 ± 9.5 – – –OLWEd 179.1 ± 35.5 – 76 ± 19 127.9 ± 3.3 – –
DrugParomomycin 0.42 ± 0.16 0.72 ± 0.06 3.11 ± 0.3 1.94 ± 0.09 0.11 ± 0.05 0.35 ± 0.05HePC 3.02 ± 0.21 3.75 ± 0.74 0.35 ± 0.02 3.27 ± 0.09 0.94 ± 0.54 4.99 ± 1.03
–, no leishmanicidal activity detected.a Stationary-phase promastigotes.
Om
8pat5ffrlm(
S
aVimAtgw
R
E
mtpaaop6mayp7
extracts, oleuropein and hydroxytyrosol, exhibited the highest SIratios, with values of 3.24 and 4.65 at 110 ± 32 and 38.7 ± 3 �g/ml,respectively, for inhibiting 50% of the L. donovani amastigotegrowth. The oleuropein 60% mixture and OLD showed relatively
Table 2IC50 and CC50 (�g/ml) of olive tree natural products vs. Leishmania intracellularamastigotes in J774A.1 macrophages.
L. donovani MON-2 J774A.1 macrophages SIa
IC50 amastigote CC50
Natural productOleuropein 110 ± 32 356 ± 23 3.24Oleuropein 60% 242 ± 118 355 ± 41 1.47Hydroxytyrosol 38.7 ± 3 180 ± 16 4.65OLDb 98 ± 30 149 ± 23 1.52OLWEc 203 ± 57 196 ± 11 0.71
DrugParomomycin 1.2 ± 0.85 275 ± 53 229.2
b Logarithmic-phase promastigotes.c Olive leaf decoction.d Olive leaf water extraction.
leuropein leishmanicidal activity in L. donovani-infected BALB/cice
Stationary-phase L. donovani promastigotes were used to infect–10-week-old female BALB/c mice by administration of 1.5 × 107
romastigotes per animal intravenously. Mice were randomlyssigned to five groups 15 days post infection. Animals in the firsthree groups received for 28 days, every other day, 45, 15 and
mg/kg of body weight (b.w.) of pure oleuropein, respectively. Theourth group received 4 mg/kg b.w. HePC by oral gavage every dayor 28 consecutive days. Finally, mice infected with L. donovani andeceiving no treatment served as positive control. The spleen andiver parasitic load was determined at 3 days and 6 weeks post treat-
ent using the limited dilution method as previously describedCarrion et al., 2006).
tatistical analysis
In vitro leishmanicidal activity and cytotoxicity were expresseds the IC50 and CC50 by the linear regression analysis, respectively.alues are mean ± S.E. from at least three independent experiments
n duplicate. Differences between means from the in vivo experi-ent were analyzed for significance using one-way ANOVA. WhenNOVA indicated significant differences, a post-hoc analysis using
he Gabriel test was conducted for multiple comparisons amongroups. All data are presented as mean values ± S.E. Differencesere considered significant at a 0.05 level of confidence.
esults
ffect of natural product activity on promastigote growth
Any anti-leishmania effect was first determined using the ala-arBlue method. Pure oleuropein extract showed an effect similar
o that of the oleuropein 60% mixture in all three stationary-hase Leishmania strains tested (Table 1). However, their activitygainst L. donovani promastigotes was markedly higher (77.2 ± 10.5nd 84 ± 13.5 �g/ml, respectively). On the other hand, only pureleuropein inhibited the growth of L. infantum and L. donovaniromastigotes obtained from logarithmic phase (191.9 ± 19.3 and9.4 ± 6.3 �g/ml, respectively). Hydroxytyrosol also inhibited pro-astigote growth in the stationary and logarithmic phases in
ll three Leishmania strains tested. As Table 1 shows, hydrox-tyrosol demonstrated leishmanicidal activity similar to that ofure oleuropein against L. donovani promastigotes (60.5 ± 7.2 and5.5 ± 0.6 �g/ml vs. 77.2 ± 10.5 and 69.4 ± 6.3 �g/ml, respectively).
Of interest, hydroxytyrosol exerted the highest leishmanicidaleffect against stationary and logarithmic L. major promastigotesamong all natural products tested in this study (50.1 ± 4.8 and105 ± 13 �g/ml, respectively).
OLD exhibited antiparasitic activity only against L. infan-tum and L. donovani promastigotes obtained from stationaryphase (113 ± 25.5 and 60 ± 9.5 �g/ml, respectively). OLWE had thesame leishmanicidal effect as OLD against stationary L. infantumand L. donovani promastigotes (179.1 ± 35.5 and 76 ± 19 �g/ml,respectively) but also inhibited logarithmic phase L. donovani pro-mastigotes (127 ± 3.3 �g/ml). Overall, pure oleuropein and purehydroxytyrosol were the most potent natural extracts tested; pureoleuropein was effective against L. donovani MON-2 promastigotesand hydroxytyrosol was more effective against L. infantum MON-1. As expected, paromomycin and HePC drastically reduced thepromastigote growth of all three Leishmania strains during bothcultivation phases.
Effect of natural product activity on L. donovani amastigotegrowth and cytotoxic effects on J774A.1 macrophages
To investigate whether these natural products exert activityagainst the intracellular form of the parasite, we calculated theselectivity index (SI) ratio (CC50 for macrophage/IC50 for amastig-otes) for every natural product tested (Table 2). The two pure
HePC 0.48 ± 0.23 143 ± 43 297.9
a Selectivity index ratio (CC50 macrophage/IC50 amastigote).b Olive leaf decoction.c Olive leaf water extraction.

medic
lucaS
Ed
ieHpa1mt
nc
Ftd(wr
J.D. Kyriazis et al. / Phyto
ow selective activity against intracellular amastigotes, with SI val-es at 1.47 and 1.52, respectively. In addition, OLWE showed greaterytotoxicity against J774A.1 macrophages than an ability to inhibitmastigote growth, with an SI ratio of 0.71. Paromomycin and HePCI values were 229.2 and 297.9, respectively.
ffect of natural product activity on the parasitic burden of L.onovani-infected BALB/c mice
Three doses of oleuropein (45, 15 and 5 mg/kg b.w.) were givenntraperitoneally to L. donovani-infected BALB/c mice for 28 days,very second day. A control group of infected mice received oralePC (4 mg/kg b.w.) for 28 consecutive days. During the treatmenteriod, the mice were weighed as an indirect indicator of their over-ll health. Their weight had a non-significant increase ranging from0% to 16% during the treatment period (data not shown). Experi-ental mice were sacrificed 3 days or 6 weeks post treatment and
heir spleens and livers dissected to determine their parasitic load.Data obtained from mice sacrificed 3 days after treatment termi-
ation showed that all oleuropein doses (45, 15 and 5 mg/kg b.w.)aused a significant reduction in parasitic load in the spleen, up
ig. 3. Spleen (a) and liver (b) parasitic burden, 3 days after the termination ofreatment. Oleuropein (Ole) was administered intraperitoneally in three indepen-ent groups (n = 8) at different doses (45, 15 and 5 mg/kg of body weight). HePCmiltefosine) as reference drug was administered by oral gavage at 4 mg/kg of bodyeight (n = 10). Positive control (+++) represents the untreated group (n = 10). The
esults are expressed as ×103 parasites/g of tissue ± S.E.
ine 20 (2013) 275– 281 279
to 79.5% (p = 0.000), 86.8% (p = 0.000), and 57.2% (p = 0.001) com-pared with the positive untreated control, respectively (Fig. 3(a)).The parasitic burden in the liver of these mice was also significantlyreduced by 58.1% (p = 0.003), 62% (p = 0.001), and 65.7% (p = 0.001),respectively (Fig. 3(b)). HePC, as the reference drug, exhibited a sig-nificant reduction of 98.8% (p = 0.000) and 98.2% (p = 0.000) in theparasitic load of both spleen and liver tissues compared with thepositive untreated control.
To determine whether treatment with oleuropein resulted inpersistent control of visceral leishmaniasis, mice were sacrificed6 weeks after treatment completion and their parasitic burdenin spleen and liver assessed. We found that all three doses ofoleuropein (45, 15 and 5 mg/kg b.w.) drastically diminished thespleen parasitic burden by 99.7% (p = 0.000), 99.8% (p = 0.000),and 96.9% (p = 0.000), respectively (Fig. 4(a)). Similarly, micetreated with HePC also had a significantly reduced parasitic loadin both spleen and liver, by 99.9% (p = 0.000) and 99.7% (p = 0.011),respectively (Fig. 4(b)). 45 and 15 mg/kg b.w. oleuropein still had
a leishmanicidal effect on liver parasitic burden, displaying asignificant reduction of 99.6% (p = 0.011). In contrast, 6 weeks aftertreatment termination, mice treated with 5 mg/kg b.w. oleuropeinFig. 4. Spleen (a) and liver (b) parasitic burden, 6 weeks after treatment termi-nation. Oleuropein (Ole) was administered intraperitoneally in three independentgroups (n = 4) at different doses (45, 15 and 5 mg/kg of body weight). HePC (miltefo-sine) as reference drug was administered by oral gavage at 4 mg/kg of body weight(n = 4). Positive control (+++) represents the untreated group (n = 4). The results areexpressed as ×103 parasites/g of tissue ± S.E.

2 medic
ds
D
irtllapbic
iLwdpgantgvtwmsact
timtwtaaaawoRmpaet(acetitauapa
80 J.D. Kyriazis et al. / Phyto
id not sustain control over the multiplication of the parasite andhowed a recovered parasitic load in the liver.
iscussion
Here we demonstrate for the first time that biophenols presentn olive leaves and OMWW, such as oleuropein and hydroxyty-osol exhibit leishmanicidal activity in vitro. Moreover, we showedhat the non-toxic secoiridoid oleuropein can abate spleen andiver parasitic burden in BALB/c mice with experimental visceraleishmaniasis. So far, the only survey that has highlighted thentiparasitic activity of oleuropein concerns its effect against Toxo-lasma gondii in vitro by inhibiting proliferation of Madin–Darbyovine kidney cells infected with tachyzoites and in vivo reduc-
ng by inhibiting 55.4% the expansion of parasites in the peritonealavity of infected mice (Jiang et al., 2008).
Oleuropein and hydroxytyrosol, as the only pure extracts testedn this study, showed antiparasitic activity against L. infantum and. donovani, which are responsible for visceral leishmaniasis, asell as against L. major, which causes the cutaneous form of theisease. Another flavonoid in olive oil, luteolin, also exhibits anti-arasitic activity against L. donovani by inhibiting promastigote cellrowth and inducing drastic morphological changes of the par-site as evidenced by loss of cellular integrity and degranulateduclei of the promastgotes (Mittra et al., 2000). We showed herehat oleuropein and hydroxytyrosol inhibited promastigote cellrowth from both logarithmic and stationary phases of L. dono-ani and L. infantum cultures. Hydroxytyrosol was also capableo abort L. major logarithmic and stationary phase promastigotes,hile oleuropein inhibited only the growth of stationary phase L.ajor promastigotes. Reduced concentrations of the active sub-
tances in the crude extracts tested (oleuropein 60% mixture, OLD,nd OLWE) in combination with the admixtures reduced the effi-acy of these extracts, resulting in decreased activity against thesehree Leishmania strains.
Nevertheless, testing the activity of the natural products againsthe intracellular amastigote forms of the Leishmania parasites ismportant since they survive and multiply within mammalian
acrophages. Because L. donovani was the most sensitive amonghe three species examined, the efficacy of the five natural productsas tested against the amastigote form of L. donovani protozoans
hat parasitize J774A.1 macrophages. Based on SI ratios, oleuropeinnd hydroxytyrosol demonstrated the highest anti-amastigotectivity, taking into account the cytotoxicity that they exhibitedgainst uninfected macrophages. Moreover, both pure oleuropeinnd the oleuropein 60% mixture showed identical cytotoxicity,hereas their leishmanicidal activity against L. donovani amastig-
tes differed significantly, being two-fold greater with oleuropein.egarding cytotoxicity, pure oleuropein was less toxic to mam-alian cells when compared with the standard drugs although
aromomycin and HePC exhibited powerful activity against themastigote forms of the parasite. In addition to luteolin, whichxhibits anti-amastigote activity (Mittra et al., 2000), other bioac-ive compounds in olive oil and OMWW such as verbascoside,+)-taxifolin, o-coumaric acid, p-coumaric acid, caffeic acid, feruliccid, cinnamic acid, and gallic acid also exert in vitro leishmani-idal activity against axenic L. donovani amastigotes (Tasdemirt al., 2006). In these cases, natural products come into direct con-act with axenic amastigote parasites, circumventing the difficultynherent in having to enter a host cell to be active. However, inhis study, we found that pure oleuropein and hydroxytyrosol have
direct effect on promastigotes (through programmed cell death;
npublished results) and also block proliferation of intracellularmastigotes. The latter indicates either the ability of these two bio-henols to enter into macrophages and remain active directly or toctuate the host cell and exert indirect leishmanicidal activity.ine 20 (2013) 275– 281
Luteolin, which is more active than oleuropein and hydroxy-tyrosol in vitro, seems to have little effect in L. donovani-infectedBALB/c mice (Tasdemir et al., 2006). On the other hand, luteolincan diminish the spleen parasitic load of golden hamsters infectedby the same Leishmania species (Mittra et al., 2000). In our study,oleuropein in an in vivo experimental model not only managed toblock the spread of organ parasitosis but also was able to maintainan enhanced durable control of the visceral form of the disease. Allthree doses of pure oleuropein almost depleted the parasitic loadof the spleen 42 days after treatment termination, demonstratingan effect similar to the standard drug HePC used in clinical practiceagainst human and canine visceral leishmaniasis. Analogous deple-tion of parasites occurred in the liver with 45 and 15 mg/kg b.w.of oleuropein and HePC, while in contrast oleuropein at a doseof 5 mg/kg b.w. failed to inhibit the progression of the disease.An explanation for this finding may be the faster metabolism ofoleuropein in the liver during detoxification relative to its persis-tence in the plasma, which is correlated with spleen absorptionbecause of the close relationship of the spleen with blood circula-tion (Serra et al., 2012). Sustained oleuropein levels in plasma mayexert an anti-inflammatory effect via IL-1b decrease since has beenreported that in human whole blood cultures stimulated with LPS,oleuropein inhibited the production of IL-1b by 80% (Miles et al.,2005). The capacity of oleuropein to reduce IL-1 molecules suggestsa possible switching of the immunological response toward the Th-1 type. This later hypothesis is supported by the presence of IL-12and IL-18 in spleen cell culture of IL-1 knockout BALB/c mice thatselectively induce IFN-� production that determines resistance tovisceral leishmaniasis (Voronov et al., 2010).
It is important to investigate the exact mechanism by whicholeuropein inhibits visceral leishmaniasis caused by L. donovani.The low cost of oleuropein and its high efficacy are importantbecause of the great need for new antiparasitic compounds acces-sible for low-income patients. Of note, oleuropein might constitutean ideal chemotherapy for patients who are co-infected with vis-ceral leishmaniasis and HIV because oleuropein can inhibit bothviral entry and integration (Lee-Huang et al., 2007a,b). Finally,oleuropein could serve as a forerunner for the rational design ofsynthetic analogs with higher in vitro and in vivo activities.
Conflict of interest
The authors declare no financial or commercial conflict of inter-est.
Acknowledgments
This work was funded by an internal grant from the HPI. I.D.K.received scholarships from the General Secretariat of Research andTechnology, Ministry of Development, Competitiveness, Infrastruc-ture, Transport and Networks.
We thank Dr. K. Soteriadou for the introduction of our labora-tories, helpful discussions and for providing us L. donovani MON-2strain. Also, we thank Dr. E. Karagouni for critical discussions.
References
Agalias, A., Magiatis, P., Skaltsounis, A.L., Mikros, E., Tsarbopoulos, A., Gikas, E.,Spanos, I., Manios, T., 2007. A new process for the management of olive oil millwaste water and recovery of natural antioxidants. Journal of Agricultural andFood Chemistry 55, 2671–2676.
Andreadou, I., Iliodromitis, E.K., Mikros, E., Constantinou, M., Agalias, A., Magiatis,P., Skaltsounis, A.L., Kamber, E., Tsantili-Kakoulidou, A., Kremastinos, D.T.,2006. The olive constituent oleuropein exhibits anti-ischemic, antioxidative,and hypolipidemic effects in anesthetized rabbits. Journal of Nutrition 136,2213–2219.

medic
B
C
C
C
d
D
G
G
J
K
K
L
L
J.D. Kyriazis et al. / Phyto
isignano, G., Tomaino, A., Lo Cascio, R., Crisafi, G., Uccella, N., Saija, A., 1999. Onthe in-vitro antimicrobial activity of oleuropein and hydroxytyrosol. Journal ofPharmacy and Pharmacology 51, 971–974.
ardinali, A., Cicco, N., Linsalata, V., Minervini, F., Pati, S., Pieralice, M., Tursi, N., Lat-tanzio, V., 2010. Biological activity of high molecular weight phenolics from olivemill wastewater. Journal of Agricultural and Food Chemistry 58, 8585–8590.
arrion, J., Nieto, A., Iborra, S., Iniesta, V., Soto, M., Folgueira, C., Abanades, D.R.,Requena, J.M., Alonso, C., 2006. Immunohistological features of visceral leish-maniasis in BALB/c mice. Parasite Immunology 28, 173–183.
icerale, S., Conlan, X.A., Sinclair, A.J., Keast, R.S., 2009. Chemistry and health of oliveoil phenolics. Critical Reviews in Food Science and Nutrition 49, 218–236.
en Boer, M., Argaw, D., Jannin, J., Alvar, J., 2011. Leishmaniasis impact and treatmentaccess. Clinical Microbiology and Infection 17, 1471–1477.
ujardin, J.C., Campino, L., Canavate, C., Dedet, J.P., Gradoni, L., Soteriadou, K., Maz-eris, A., Ozbel, Y., Boelaert, M., 2008. Spread of vector-borne diseases and neglectof Leishmaniasis, Europe. Emerging Infectious Diseases 14, 1013–1018.
eorgopoulou, K., Smirlis, D., Bisti, S., Xingi, E., Skaltsounis, L., Soteriadou, K., 2007.In vitro activity of 10-deacetylbaccatin III against Leishmania donovani pro-mastigotes and intracellular amastigotes. Planta Medica 73, 1081–1088.
ranados-Principal, S., Quiles, J.L., Ramirez-Tortosa, C.L., Sanchez-Rovira, P.,Ramirez-Tortosa, M.C., 2010. Hydroxytyrosol: from laboratory investigations tofuture clinical trials. Nutrition Reviews 68, 191–206.
iang, J.H., Jin, C.M., Kim, Y.C., Kim, H.S., Park, W.C., Park, H., 2008. Anti-toxoplasmosiseffects of oleuropein isolated from Fraxinus rhychophylla. Biological and Phar-maceutical Bulletin 31, 2273–2276.
appagoda, S., Singh, U., Blackburn, B.G., 2011. Antiparasitic therapy. Mayo ClinicProceedings 86, 561–583.
atsiki, M., Chondrogianni, N., Chinou, I., Rivett, A.J., Gonos, E.S., 2007. The oliveconstituent oleuropein exhibits proteasome stimulatory properties in vitroand confers life span extension of human embryonic fibroblasts. RejuvenationResearch 10, 157–172.
ee-Huang, S., Huang, P.L., Zhang, D., Lee, J.W., Bao, J., Sun, Y., Chang, Y.T., Zhang, J.,
2007a. Discovery of small-molecule HIV-1 fusion and integrase inhibitors oleu-ropein and hydroxytyrosol: Part I. Fusion [corrected] inhibition. Biochemical andBiophysical Research Communications 354, 872–878.ee-Huang, S., Huang, P.L., Zhang, D., Lee, J.W., Bao, J., Sun, Y., Chang, Y.T., Zhang,J., 2007b. Discovery of small-molecule HIV-1 fusion and integrase inhibitors
ine 20 (2013) 275– 281 281
oleuropein and hydroxytyrosol: Part II. Integrase inhibition. Biochemical andBiophysical Research Communications 354, 879–884.
Mikus, J., Steverding, D., 2000. A simple colorimetric method to screen drug cytotox-icity against Leishmania using the dye Alamar Blue. Parasitology International48, 265–269.
Miles, E.A., Zoubouli, P., Calder, P.C., 2005. Differential anti-inflammatory effects ofphenolic compounds from extra virgin olive oil identified in human whole bloodcultures. Nutrition 21, 389–394.
Mittra, B., Saha, A., Chowdhury, A.R., Pal, C., Mandal, S., Mukhopadhyay, S., Bandyo-padhyay, S., Majumder, H.K., 2000. Luteolin, an abundant dietary component is apotent anti-leishmanial agent that acts by inducing topoisomerase II-mediatedkinetoplast DNA cleavage leading to apoptosis. Molecular Medicine 6, 527–541.
Rocha, L.G., Almeida, J.R., Macedo, R.O., Barbosa-Filho, J.M., 2005. A review of naturalproducts with antileishmanial activity. Phytomedicine 12, 514–535.
Sen, R., Chatterjee, M., 2011. Plant derived therapeutics for the treatment of leish-maniasis. Phytomedicine 18, 1056–1069.
Serra, A., Rubio, L., Borras, X., Macia, A., Romero, M.P., Motilva, M.J., 2012. Distributionof olive oil phenolic compounds in rat tissues after administration of a phenolicextract from olive cake. Molecular Nutrition & Food Research 56, 486–496.
Tasdemir, D., Kaiser, M., Brun, R., Yardley, V., Schmidt, T.J., Tosun, F., Ruedi, P.,2006. Antitrypanosomal and antileishmanial activities of flavonoids and theiranalogues: in vitro, in vivo, structure–activity relationship, and quantitativestructure–activity relationship studies. Antimicrobial Agents and Chemother-apy 50, 1352–1364.
Tuck, K.L., Hayball, P.J., 2002. Major phenolic compounds in olive oil: metabolismand health effects. The Journal of Nutritional Biochemistry 13, 636–644.
Visioli, F., Bellosta, S., Galli, C., 1998. Oleuropein, the bitter principle of olives,enhances nitric oxide production by mouse macrophages. Life Sciences 62,541–546.
Voronov, E., Dotan, S., Gayvoronsky, L., White, R.M., Cohen, I., Krelin, Y., Benchetrit, F.,Elkabets, M., Huszar, M., El-On, J., Apte, R.N., 2010. IL-1-induced inflammationpromotes development of leishmaniasis in susceptible BALB/c mice. Interna-
tional Immunology 22, 245–257.Waterman, E., Lockwood, B., 2007. Active components and clinical applications ofolive oil. Alternative Medicine Review 12, 331–342.
World Health Organization. Leishmaniasis Control home page: http://whqlibdoc.who.int/trs/WHO TRS 949 eng.pdf