
Distribution, frequency and diversity of Bacillus thuringiensis in olive tree environments in Spain
7
System. App!. Microbio!. 20, 652-658 (1997) © Gustav Fischer Verlag Distribution, frequency and diversity of Bacillus thuringiensis in olive tree environments in Spain YOLANDA BEL, FRANCISCO GRANERO, TRINIDAD M. ALBEROLA, MARIA J. MARTINEZ-SEBASTIAN and JUAN FERRE Departament de Genetica, Universitat de Valencia, Burjassot (Valencia), Spain Received April 17, 1997 Summary Bacillus thuringiensis was isolated from samples collected from olive tree related habitats (olive groves, olive storage facilities and oil mills) of different olive producing regions in Spain. This bacterium was found in a high percentage (92%) of samples. From 72 samples analysed, 2244 sporulating colonies were selected and observed under phase contrast microscopy. From these, 414 colonies (18%) were clas- sified as B. thuringiensis based on the production of parasporal crystals. A great variability of spore, crystal size and morphology was observed. The results indicate that olive groves are as rich as soil olive mills and olive storage facilities regarding the B. thuringiensis population. Analysis by SDS-PAGE of the proteins in the parasporal crystal showed a considerable heterogeneity be- tween B. thuringiensis strains. This was especially relevant in samples obtained from oil mills and stor- age centres, indicating the high polymorphism in this species. Strains isolated from the same sample showing the same SDS-PAGE profile were not considered differ- ent, and just one of them was selected for further tests. A total of 188 out of the 414 B. thuringiensis iso- lates were shown to be different strains and tested for their toxicity against adult animals of the olive fly Bactrocera oleae. Approximately 40% of the strains caused similar or higher mortality than the dipteran active strain B. thuringiensis israelensis (45% mortality). Most strains (71 %) producing a toxicity high- er than 65% came from olive mills and olive storage facilities. Three of these strains exhibited a mortali- ty higher than 70% in our test conditions. Key words: B. thuringiensis - Olive environments - Crystal proteins - Bactrocera oleae - Insecticidal activity - Diptera Introduction Bacillus thuringiensis is a common Gram-positive soil microorganism widely distributed throughout the world. It has been isolated from soil, mud, plants, insects and other habitats, but the usual habitat of this bacterium, as well as the need of an association with insects, remain undetermined (MARTIN and TRAVERS, 1989; SMITH and COUCHE, 1991; ISHII and OHBA, 1993; MEADOWS, 1993). The main interest in this bacterium is that, when sporulation occurs, it produces crystalline protein inclu- sions which may be toxic to several species of insect pests. These inclusions consist of Cry and Cyt proteins, in different comhinations. Until now, 56 different Cry proteins and 4 Cyt proteins have been described (CRICK- MORE, 1996). Because of the advantages in the use of biopesticides as opposed to chemical insecticides, the use of B. thuringiensis based insecticides is increasing: sever- al strains are already being used to biocontrol pest in- sects in the orders Lepidoptera (e.g. Heliathis, Spad aptera, Pieris), Coleoptera (e.g. Leptinatarsa) and Diptera (e.g. Aedes, Culex). But the search for new strains with increased toxicity against a particular pest or pests, and strains with new insect specificities is continu- ing (CHILCOT and WIGLEY, 1994). The goal of this research was to study the occurrence of B. thuringiensis strains in olive tree related areas to de- termine their distribution and genetic diversities in these habitats, and to test the possible correlation, in terms of toxicity, hetween the isolated strains and one of the most important olive pests, the olive fly Bactrocera aleae. B. aleae (Gmelin), depending on several factors (as the climate), produces a 10 to 70% of damage in the olive harvest of the southern European countries, Spain
Transcript of Distribution, frequency and diversity of Bacillus thuringiensis in olive tree environments in Spain

System. App!. Microbio!. 20, 652-658 (1997) © Gustav Fischer Verlag
Distribution, frequency and diversity of Bacillus thuringiensis in olive tree environments in Spain
YOLANDA BEL, FRANCISCO GRANERO, TRINIDAD M. ALBEROLA, MARIA J. MARTINEZ-SEBASTIAN and JUAN FERRE
Departament de Genetica, Universitat de Valencia, Burjassot (Valencia), Spain
Received April 17, 1997
Summary
Bacillus thuringiensis was isolated from samples collected from olive tree related habitats (olive groves, olive storage facilities and oil mills) of different olive producing regions in Spain. This bacterium was found in a high percentage (92%) of samples. From 72 samples analysed, 2244 sporulating colonies were selected and observed under phase contrast microscopy. From these, 414 colonies (18%) were classified as B. thuringiensis based on the production of parasporal crystals. A great variability of spore, crystal size and morphology was observed. The results indicate that olive groves are as rich as soil olive mills and olive storage facilities regarding the B. thuringiensis population. Analysis by SDS-PAGE of the proteins in the parasporal crystal showed a considerable heterogeneity between B. thuringiensis strains. This was especially relevant in samples obtained from oil mills and storage centres, indicating the high polymorphism in this species. Strains isolated from the same sample showing the same SDS-PAGE profile were not considered different, and just one of them was selected for further tests. A total of 188 out of the 414 B. thuringiensis isolates were shown to be different strains and tested for their toxicity against adult animals of the olive fly Bactrocera oleae. Approximately 40% of the strains caused similar or higher mortality than the dipteran active strain B. thuringiensis israelensis (45% mortality). Most strains (71 %) producing a toxicity higher than 65% came from olive mills and olive storage facilities. Three of these strains exhibited a mortality higher than 70% in our test conditions.
Key words: B. thuringiensis - Olive environments - Crystal proteins - Bactrocera oleae - Insecticidal activity - Diptera
Introduction
Bacillus thuringiensis is a common Gram-positive soil microorganism widely distributed throughout the world. It has been isolated from soil, mud, plants, insects and other habitats, but the usual habitat of this bacterium, as well as the need of an association with insects, remain undetermined (MARTIN and TRAVERS, 1989; SMITH and COUCHE, 1991; ISHII and OHBA, 1993; MEADOWS, 1993).
The main interest in this bacterium is that, when sporulation occurs, it produces crystalline protein inclusions which may be toxic to several species of insect pests. These inclusions consist of Cry and Cyt proteins, in different comhinations. Until now, 56 different Cry proteins and 4 Cyt proteins have been described (CRICKMORE, 1996). Because of the advantages in the use of biopesticides as opposed to chemical insecticides, the use of B. thuringiensis based insecticides is increasing: sever-
al strains are already being used to biocontrol pest insects in the orders Lepidoptera (e.g. Heliathis, Spadaptera, Pieris), Coleoptera (e.g. Leptinatarsa) and Diptera (e.g. Aedes, Culex). But the search for new strains with increased toxicity against a particular pest or pests, and strains with new insect specificities is continuing (CHILCOT and WIGLEY, 1994).
The goal of this research was to study the occurrence of B. thuringiensis strains in olive tree related areas to determine their distribution and genetic diversities in these habitats, and to test the possible correlation, in terms of toxicity, hetween the isolated strains and one of the most important olive pests, the olive fly Bactrocera aleae.
B. aleae (Gmelin), depending on several factors (as the climate), produces a 10 to 70% of damage in the olive harvest of the southern European countries, Spain

among them, that are the major olive oil producers in the world (KARAMANLIDOU et aI., 1991). The female olive fly lays a single egg under the skin of non mature olives. Larvae emerge after two or three days and feed by forming tunnels (approximately 15 mm long) and destroying a fourth or a fifth of the olive's interior. Fruits fall prematurely and their oil is of low quality and higher acidity (DE ANDRES, 1991; DOMINGUEZ, 1993). The method of controlling this pest has been to spray with chemical pesticides, a strategy that kills the natural enemies of the fly and that can lead to the rapid development of insecticide resistance. Therefore, the discovery of novel biological pesticides to control this pest, as the ones based on the bacterium B. thuringiensis, is highly desirable.
Materials and methods
Sample collection: Sampled habitats consisted of olive grove soils, olive-oil producing mills or olive storage facilities . The samples collected included soils, dust olive or leaf residues and water coming from olive washing places. Soil samples consisted of 1 to 10 g of material collected with a clean gardening scraper after removing the soil top layer. Dust samples were obtained by scooping directly the floor of the olive storage places or by scraping as described for soil samples. Water and olive and leaf residues were collected directly from the olive washing places and olive residue piles respectively. In all cases, samples were collected into sterile plastic tubes or bags and stored at -20°C until processed.
B. thuringiensis cultures: B. thuringiensis was cultured at 30°C on CCY agar plates (13 mM KHzP04, 26 mM KzHP04,
0.002% [w/vj L-glutamine, 0.1% [w/v] casein hydrolysate, 0.1 % [w/v] bacto casitone, 0.04% bacto yeast extract, 0.6% [w/v] glycerol, 0.05 M ZnClz, 0.5 M MgClz, 0.01 M MnCI2,
0.2 M Cactz, 0.05 M FeCI}) (STEwARTet aI., 1981 ).
B. thuringiensis isolation: B. thuringiensis isolation was performed as described previously (OHBA and AIZAWA, 1986 ) with some modifications: approximately 1 g of either soil or dust was suspended in 10 ml of sterile 0.05 M phosphate buffer pH 7.0 or distilled water, and mixed vigorously on a mechanical shaker for 10 min. The resultant suspension was heated at 70°C for 15 min, mixed again for 10 seconds and heated for 15 more min (water samples were subjected to the heat shock directly, with no addition of buffer). Several aliquots of the suspensions (2, 5 and 10 pi ) were spread on the surface of CCY agar plates with a sterile glass rod, and incubated at 30°C for 2 to 3 days. Approximately 30 to 50 colonies with a Bacilluslike aspect were observed in each sample by phase-contrast microscopy. When a colony with parasporal crystal was found, it was identified as B. thuringiensis. These isolates were plated again for single colony purification on CCY agar plates and stored in sterile liquid Nutrient Broth containing 50% glycerol, at -20°C.
Protein profiling: B. thuringiensis isolates were cultured and allowed to sporulate in CCY medium. When sporulation was completed and crystals and spores were released (as checked by phase contrast microscopy), two loopfulls of the colony material were removed from the plate and transferred to sterile test tubes containing 1 ml ice-cold 0.5 M NaCI. Resuspended sam-
B. thuringiensis in olive habitats 653
pies were centrifuged for 5 min at 13,OOOxg and supernatants were discarded. Pellets were resuspended in 140 pi of 1 % SDS-0.01 % ~-mercaptoethanol, and boiled for 10 min to solubilize crystals. Samples were then centrifuged at 13,800xg for 10 min and supernatants removed to be used for electrophoresis by mixing the sample with sample buffer (0.15 M Tris/HCI pH 8.8, 3.75 mM EDTA, 0.75 M sucrose, 0.075 % bromophenol blue, 2.5% SDS and 7.4 mM dithiothreitol) in equal volumes. The mixture was boiled for 10 min. Eletrophoresis was carried out in 10% polyacrylamide gels, at 35 rnA.
Protein bands were stained with the Protein Assay Reagent from Bio-Rad (Bio-Rad Laboratories GmbH, Munchen) for 45 min, and destained with a solution of 25 % methanol 10% acetic acid.
Preparation of isolates for bioassays: The B. thuringiensis isolates were grown in CCY agar medium. When sporulation was accomplished, two loopfulls of bacteria were transferred to 0.5 ml of sterile water and mixed to have a homogeneous suspension. Tubes were then heated at 70°C for 30 min. After cooling, the total content of the tube was transferred into 50 ml of liquid CCY medium pre-warmed at 30°C. The culture was incubated at 30 °C in an orbital shaker at 250 rpm until more than 90% of the cells released the spores and crystals into the medium (as checked by phase-contrast microscopy). The suspension was then centrifuged at 9,700xg for 10 min at 4 0C. The pellet was resuspended in 25 ml ice-cold distilled water, and centrifuged at 9,700xg for 10 min. This washing procedure was performed twice. After the last centrifugation, the final pellet was resuspended in about 4 ml ice-cold distilled water. After quantification of the optical density of the sample (by measuring absorbance at 600 nm), the spores and crystals mixture was stored at -20°C.
Toxicity assays of Bactrocera oleae: B. oleae pupae were supplied to our laboratory by the "Demokritos" National Research Centre in Athens, Greece. Pupae were kept at 25°C until adults emerged. Newly emerged fasting flies (from 0 to 22 h) were used for bioassays.
Bioassays were performed in 500 ml plastic boxes with a transparent cover. In each box, a pinch of solid brewer's yeast (yeast hydrolysate enzymatic from ICN Biochemicals, Aurora, Ohio) was disposed under a small piece of mosquito plastic net, in order to provide a source of proteins. The adequate concentration of spore and crystal suspensions (a 2 fold more concentrated suspension that the one that gives an absorbance of 1.2 at 600 nm) with 30% sucrose was prepared and supplied to the flies by putting 200 pi of the suspension in a pipette yellow tip and pricking it in the wall of the boxes in such a way that flies drink from the tip. Sucrose (30%) in water was used as control.
For testing the activity of the B. thuringiensis preparations, 40 flies were used per sample. Flies were distributed in four boxes (5 males and 5 females per box). Mortality was scored after 6 days (tips containing spores and crystals and sucrose were changed by new ones once during the course of the bioassay, at the 3rd day). The B. thuringiensis strains that caused a mortality of 60% or higher were bioassayed at least twice. In the case of samples giving a high mortality, the presence of ~-exotoxin was also tested by bioassaying the supernatants that had been obtained during the preparation of the crystals and spores mixture, after sterilization at 121°C for 10 min.
In all cases, mortality of the strains tested were corrected by the Abbot's formula (ABBOT, 1925) with respect to the control mortality.
654 Y. BEL et a1.
Results
B. thuringiensis isolation
The occurrence of B. thuringiensis in olive storage warehouses, mills and olive groves has been studied.
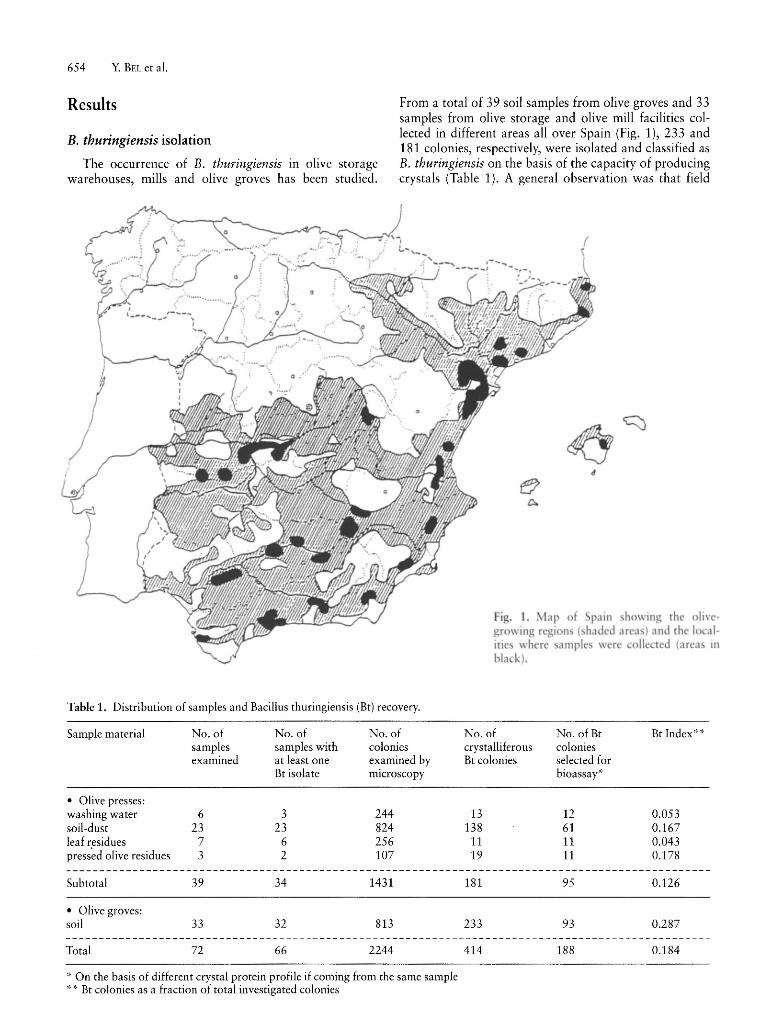
From a total of 39 soil samples from olive groves and 33 samples from olive storage and olive mill facilities collected in different areas all over Spain (Fig. 1), 233 and 181 colonies, respectively, were isolated and classified as B. thuringiensis on the basis of the capacity of producing crystals (Table 1). A general observation was that field
Table 1. Distribution of samples and Bacillus thuringiensis (Bt) recovery.
Sample material No. of No. of No. of samples samples with colonies examined at least one examined by
Bt isolate microscopy
• Olive presses: washing water 6 3 244 soil-dust 23 23 824 leaf residues 7 6 256 press~d olive residues 3 2 107
i . 1. Map of pain howing the olivegrO\ ing region (haded area) and the 10 alitie where amples were collected (area in black).
No. of No.ofBt Bt Index"* crystalliferous colonies Bt colonies selected for
bioassay*
13 12 0.053 138 61 0.167
11 11 0.043 19 11 0.178
-------------------------------------------------------------------------------------------------Subtotal 39 34 1431
• Olive groves: soil 33 32 813
Total 72 66 2244
'f On the basis of different crystal protein profile if coming from the same sample 'f * Bt colonies as a fraction of total investigated colonies
181 95 0.126
233 93 0.287
414 188 0.184
soil samples were poorer than samples from olive presses regarding the amount of sporulating colonies obtained.
B. thuringiensis was found in almost 100% of the dust and soil samples analysed (only one sample from an olive grove did not contain B. thuringiensis). On the contrary, 50% of the water samples did not contain any B. thuringiensis colony; olive washing water samples, leaf residues and pressed olive residue samples were quite irregular regarding the number of sporulating bacteria obtained from the extraction, ranging from zero to hundreds of colonies in the same conditions.
Olive grove samples gave the highest Bt index (No. of B. thuringiensis colonies found with respect to the total number of colonies examined in the sample), 1.6 times higher than the index obtained from samples coming from olive facilities (0.287 versus 0.126). The Bt index varied by a factor of 4 within the samples collected from the different habitats in the olive mills .
Crystal protein profiles
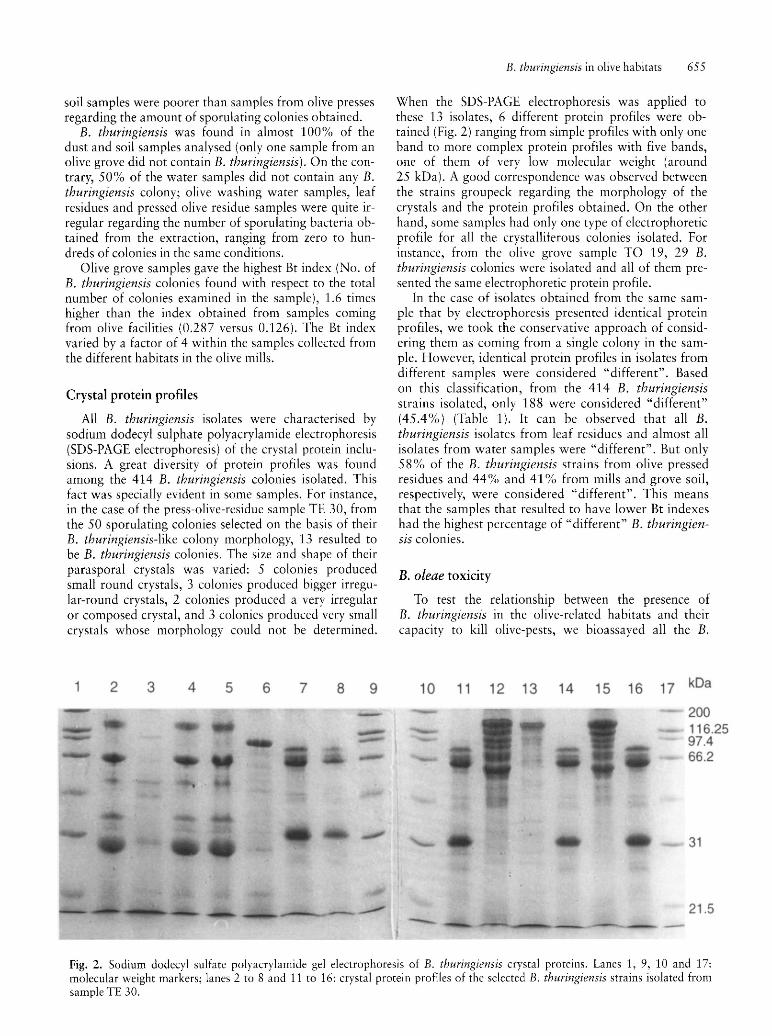
All B. thuringiensis isolates were characterised by sodium dodecyl sulphate polyacrylamide electrophoresis (SDS-PAGE electrophoresis) of the crystal protein inclusions. A great diversity of protein profiles was found among the 414 B. thuringiensis colonies isolated. This fact was specially evident in some samples. For instance, in the case of the press-olive-residue sample TE 30, from the 50 sporulating colonies selected on the basis of their B. thuringiensis-like colony morphology, 13 resulted to be B. thuringiensis colonies. The size and shape of their parasporal crystals was varied: 5 colonies produced small round crystals, 3 colonies produced bigger irregular-round crystals, 2 colonies produced a very irregular or composed crystal, and 3 colonies produced very small crystals whose morphology could not be determined.
2 3 4 5 6 7 8 9
B. thuringiensis in olive habitats 655
When the SDS-PAGE electrophoresis was applied to these 13 isolates, 6 different protein profiles were obtained (Fig. 2) ranging from simple profiles with only one band to more complex protein profiles with five bands, one of them of very low molecular weight (around 25 kDa). A good correspondence was observed between the strains groupeck regarding the morphology of the crystals and the protein profiles obtained. On the other hand, some samples had only one type of electrophoretic profile for all the crystalliferous colonies isolated. For instance, from the olive grove sample TO 19, 29 B. thuringiensis colonies were isolated and all of them presented the same electrophoretic protein profile.
In the case of isolates obtained from the same sample that by electrophoresis presented identical protein profiles, we took the conservative approach of considering them as coming from a single colony in the sample. However, identical protein profiles in isolates from different samples were considered "different". Based on this classification, from the 414 B. thuringiensis strains isolated, only 188 were considered "different" (45.4%) (Table 1). It can be observed that all B. thuringiensis isolates from leaf residues and almost all isolates from water samples were "different". But only 58% of the B. thuringiensis strains from olive pressed residues and 44 % and 41 % from mills and grove soil, respectively, were considered "different". This means that the samples that resulted to have lower Bt indexes had the highest percentage of "different" B. thuringiensis colonies.
B. oleae toxicity
To test the relationship between the presence of B. thuringiensis in the olive-related habitats and their capacity to kill olive-pests, we bioassayed all the B.
10 11 12 13 14 15 16 17 kDa
200 116.25 97.4 66.2
31
21.5
Fig. 2. Sodium dodecyl sulfate polyacrylamide gel electrophoresis of B. thuringiensis crystal proteins. Lanes 1, 9, 10 and 17: molecular weight markers; lanes 2 to 8 and 11 to 16: crystal protein profiles of the selected B. thuringiensis strains isolated from sample TE 30.
656 Y. BEL et al.
50
40
>-!:: 30
C'O -lI... 0 20 ~
~ 0
10
0 0
""A
.... --:::: ........... .-_-e'*""''' ... " ..........
.."," -..¥ ....- _... .------.. -; ........... _-........... -
....... :: ........ ..... . ' I r •••• .' ,:;. .. ,
2 3 4 5 6 7 8 9 10
Days
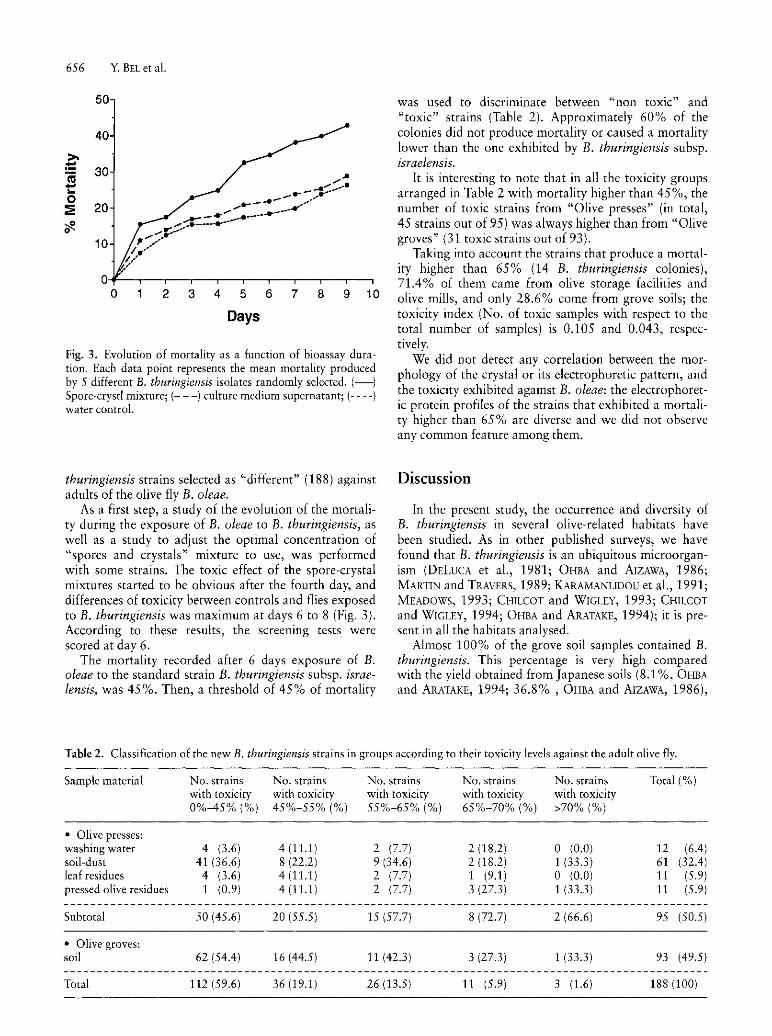
Fig. 3. Evolution of mortality as a function of bioassay duration. Each data point represents the mean mortality produced by 5 different B. thuringiensis isolates randomly selected. (-) Spore-cryst! mixture; (- - -) culture medium supernatant; (- - --) water control.
thuringiensis strains selected as "different" (188) against adults of the olive fly B. oleae.
As a first step, a study of the evolution of the mortality during the exposure of B. oleae to B. thuringiensis, as well as a study to adjust the optimal concentration of "spores and crystals" mixture to use, was performed with some strains. The toxic effect of the spore-crystal mixtures started to be obvious after the fourth day, and differences of toxicity between controls and flies exposed to B. thuringiensis was maximum at days 6 to 8 (Fig. 3). According to these results, the screening tests were scored at day 6.
The mortality recorded after 6 days exposure of B. oleae to the standard strain B. thuringiensis subsp. israelensis, was 45%. Then, a threshold of 45% of mortality
was used to discriminate between "non toxic" and "toxic" strains (Table 2). Approximately 60% of the colonies did not produce mortality or caused a mortality lower than the one exhibited by B. thuringiensis subsp. israelensis.
It is interesting to note that in all the toxicity groups arranged in Table 2 with mortality higher than 45%, the number of toxic strains from "Olive presses" (in total, 45 strains out of 95) was always higher than from "Olive groves" (31 toxic strains out of 9 3) .
Taking into account the strains that produce a mortality higher than 65% (14 B. thuringiensis colonies), 71.4% of them came from olive storage facilities and olive mills, and only 28.6% come from grove soils; the toxicity index (No. of toxic samples with respect to the total number of samples) is 0.105 and 0.043, respectively.
We did not detect any correlation between the morphology of the crystal or its electrophoretic pattern, and the toxicity exhibited against B. oleae: the electrophoretic protein profiles of the strains that exhibited a mortality higher than 65% are diverse and we did not observe any common feature among them.
Discussion
In the present study, the occurrence and diversity of B. thuringiensis in several olive-related habitats have been studied. As in other published surveys, we have found that B. thuringiensis is an ubiquitous microorganism (DELuCA et aI., 1981; OHBA and AIZAWA, 1986; MARTIN and TRAVERS, 1989; KARAMANLIDOU et aI., 1991; MEADOWS, 1993; CHILCOT and WIGLEY, 1993; CHILCOT and WIGLEY, 1994; OHBA and ARATAKE, 1994); it is present in all the habitats analysed.
Almost 100% of the grove soil samples contained B. thuringiensis. This percentage is very high compared with the yield obtained from Japanese soils (8.1 %, OHBA and ARATAKE, 1994; 36.8% , OHBA and AIZAWA, 1986),
Table 2. Classification of the new B. thuringiensis strains in groups according to their toxicity levels against the adult olive fly.
Sample material No. strains No. strains No. strains No. strains No. strains Total (%) with toxicity with toxicity with toxicity with toxicity with toxicity 0%-45% (%) 45%-55% (%) 55%-65% (%) 65%-70% (%) >70% (%)
• Olive presses: washing water 4 (3.6) 4 (11.1) 2 (7.7) 2 (18.2) 0 (0.0) 12 (6.4) soil-dust 41 (36.6) 8 (22.2) 9 (34.6) 2 (18.2) 1 (33.3) 61 (32.4) leaf residues 4 (3.6) 4 (11.1) 2 (7.7) 1 (9.1) 0 (0.0) 11 (5.9) pressed olive residues 1 (0.9) 4 (11.1) 2 (7.7) 3 (27.3) 1 (33.3) 11 (5.9) -------------------------------------------------------------------------------------------------Subtotal 50 (45.6) 20 (55.5) 15 (57.7) 8 (72.7) 2 (66.6) 95 (50.5)
• Olive groves: soil 62 (54.4) 16 (44.5) 11 (42.3) 3 (27.3) 1 (33.3) 93 (49.5) -------------------------------------------------------------------------------------------------Total 112 (59.6) 36 (19.1) 26 (13.5) 11 (5.9) 3 (1.6) 188 (100)

soils from Chile (20.4%, VASQUEZ et aI., 1995), soils from Indonesia (45%, HASTOWO et aI., 1992) or soils from New Zealand (70.0%, CHILCOT and WIGLEY, 1993). Probably this difference is due to the exhaustive screening done in the present study, in which a minimum of 30 to 50 sporulating colonies per sample were observed by phase contrast microscopy. A detailed survey, as well as the type of sample analysed, are the most important factors for an efficient B. thuringiensis isolation. Using a non-selective spore-forming medium, we have obtained percentages comparable to those of other authors using acetate-selection systems (55.6 to 94.3% depending on the soil analysed (MARTIN and TRAVERS, 1989)). Regarding dust samples, we have found B. thuringiensis in 100% of the samples, in contrast to the 55% found by DELuCA et al. (1982) in settled-dusts of grain elevators or the 78 % found by MEADOWS et al. (1992) for dusts in animal feed mills.
Taking all the samples as a whole, B. thuringiensis was present in the 91.7% of them, olive rinsing water being the poorest medium with respect to the B. thuringiensis content (50%), probably as a consequence of the lower solid content compared to the rest of the samples.
The Bt indexes are considered as measure of the success in isolating B. thuringiensis. These indexes, for soil samples, have been reported to range from values lower than 0.060 (DELUCCA et aI., 1981; OHBA and ARATAKE, 1994; OHBA and AIZAWA, 1986; HASTOWO et aI., 1992), to 0.5 (CHILCOT and WIGLEY, 1993), ore even 0.85 when acetate selection is used (MARTIN and TRAVERS, 1989). Our Bt index value (0.287) falls within this range and is in good agreement with reported values specific for agricultural and horticultural indexes (0.25 and 0.22, respectively) (CHILCOT and WIGLEY, 1993).
The Bt indexes we have obtained for samples from olive presses and olive mills are lower, ranging from 0.053 (olive washing water) to 0.178 (pressed olive residues). If we compare these values with the ones obtained from B. thuringiensis isolation programs performed in other "closed areas", such as an animal feed mill (0.67) or a sericulture farm (0.045), it seems that the frequency of the bacterium is not regular and depends mainly on the type of material sampled.
In summary, among all the habitats studied in this report, olive washing water and olive tree leave residues had the poorest B. thuringiensis contents. On the contrary, the highest proportion of B. thuringiensis colonies was found in soils from olive groves and was also high in soil or dust from olive storage facilities or oil mills. Then it seems that B. thuringiensis is better maintained or protected in the soil. One reason for this may be that when samples of soils and dust are collected, the surface is always rejected and the material of the sample belongs to a soil layer about at least 5 cm under the surface, a level where UV light damage is not possible and temperature is more stable. Furthermore, it has been demonstrated that B. thuringiensis can grow in soils (TRINDADE et aI., 1996) and it has been repeatedly suggested that the soil can act as a reservoir of spores and that is the normal
B. thuringiensis in olive habitats 657
habitat of B. thuringiensis (AKIBA, 1986; MARTIN and TRAVERS, 1989; MEADOWS, 1993; OHBA and ARATAKE, 1994; LERECLUS, 1996). Consequently, one should find, for instance, a higher amount of B. thuringiensis colonies but less diversity in a single sample of field soil than in a water sample. We have observed this effect in the present work: after applying SDS-PAGE electrophoresis analysis to the crystals of the B. thuringiensis colonies isolated, we have found that the possibility of finding the same electrophoretic profile for all or part of the B. thuringiensis isolates of a single field soil sample was higher than in the samples of other origins.
To assess the possible relationship existing between the insects and the B. thuringiensis populations that coexist in the same habitats, we have tested the toxicity of the 188 selected strains against the olive fly B. oleae. We selected the adult flies for the bioassays in spite of the fact that mortality produced by B. thuringiensis in adult stages is very low compared to the mortality rates obtained when larvae are used (KARAMANLIDOU et aI., 1991). This selection was based on the fact that if a strain of B. thuringiensis was to be used in the future to spray olive trees, the adult fly would be the target, not the larva.
Approximately 40% of the strains (76 from 188) caused a mortality similar or higher than the B. thuringiensis subsp. israelensis (45%). The toxic B. thuringiensis isolates came preferably from the samples obtained from olive storage facilities and olive mills; almost three fourths of the strains that gave toxicity rates higher than 65% came from these habitats. Olive groves are open habitats where the possibility of finding non-relatedolive-pest-B. thuringiensis strains is higher than in olive storage warehouses and olive mills, much more closed habitats, where the outside apportions consist on olives (some containing larvae of B. oleae) accompained of olive related material. Then, a slight association can be deduced from out data. This agrees with other studies performed in tobacco storage habitats (KAELIN et aI., 1994) in which results suggested that B. thuringiensis may be associated with the tobacco beetle Lasioderma serricone, and studies in silkworm-breeding environments (OHBA and ARATAKE, 1994) where the majority of the isolates from silkworm litters were toxic to these insect species (in contrast to what they found in soils). It would be interesting to test in the future the toxicity of our strain collection against other olive pests such as the lepidopteran Prays oleae.
Protein profiles of the crystals of the most active strains were analysed by SDS-PAGE electrophoresis and compared in order to look for a common characteristic band that could be associated with the mortality. ISHII and OHBA (1993) analysed some environmental isolates that showed larvicidal activity specific for dipterans as mosquitoes; the toxic B. thuringiensis strains belonged to several serotypes but had some common features, as for example that parasporal inclusions of most of them contained high molecular proteins (130-140 kDa) and/or medium MW proteins (65 kDa or 72-90 kDa), but all of them had the 2S-29 kDa cytolytic proteins. Our most

658 Y. BEL et a!.
toxic strains did not share characteristic bands and low molecular weight proteins of the cytolytic type were found in just one of the three most toxic strains. As an example of the variability found in this study, the six groups in which the isolates from the sample TE 30 can be assigned to, show in all cases medium toxicity (ranging from 46.5 to 57.4) but very different protein profiles, some of them including low molecular proteins bands. This fact corroborates that the crystal protein profile does not always indicate the toxicity of the isolate (CHILCOT and WIGLEY, 1994) and further tests are necessary to relate protein bands and the toxicity.
Acknowledgements We thank all the farmers and olive millers, as well as the
Spanish olive related organisations that have collaborated in this work by allowing us to collect the samples. We also acknowledge to Dr. S. KOLIAIS his comments on the manuscript.
This study was funded by the European Communities, Program AIR 3-CT93-0868, by the Spanish Ministerio de Educaci6n y Ciencia (Project Number AGF93-1321-CE) and by the Spanish Ministerio de Educaci6n y Ciencia and the British Council, through a Cooperative Project, Number HB94-229.
References
ABBOTT, W. S.: A method of computing the effectiveness of insecticide. J. Econ. Entomo!' 18,265-267 (1925).
AKIBA, Y.: Microbial ecology of Bacillus thuringiensis. IV. Germination of Bacillus thuringiensis spores in the soi!' App!. Ent. Zoo!. 21, 76-80 (1986).
CRICKMORE, N., ZEIGLER, R. D., FEITELSON, J., SCHNEPF, E., LAMBERT, B., LERECLUS, D., GAWRON-BuRKE, c., and DEAN, D. H.: "Bacillus thuringiensis toxin gene nomenclature". httpllwww.susx.ac.uk:80IUsers/bafn6/btlindex.htlm. Last updated July 1996.
CHILCOTT, C. N., and WIGLEY, P. J.: Isolation and toxicity of Bacillus thuringiensis from soil and insect habitats in New Zealand. J. Invertebr. Patho!' 61,244-247 (1993).
CHILCOTT, C. N., and WIGLEY, P. J.: Opportunities for finding new Bacillus thuringiensis strains. Agric. Ecosyst. Environ. 49,51-57 (1994).
DE ANDRES, E: Capitulo IV: Parasitos del reino animal, pp. 175-428. In: Enfermedades y plagas del olivo (RIQUELME and VARGAS, eds.) 2nd. ed., Jaen 1991.
DELUCA, A. J., PALMGREN, M. S., CiEGLER, A.: Bacillus thuringiensis in grain elevator dusts. Can. J. Microbio!. 28, 452-456 (1982).
DELUCCA, A. J., SIMONSON, J. G., LARSON, A. D.: Bacillus thuringiensis distribution in soils of the United States. Can. J. Microbio!. 27, 865-870 (1981).
DOMINGUEZ, E: Dipteros: La mosca del olivo Dacus aleae Rossi, pp. 679-685. In: Plagas y enfermedades de las plantas cultivadas (MUNDI-PRENSA, eds.) 9th ed., Madrid 1993.
HASTOWO, S., LAY, B. W., and OHBAR, M.: Naturally occurring Bacillus thuringiensis in Indonesia. J. App!. Bacteria!' 73, 108-113 (1992).
ISHII, T., OHBA, M.: Diversity of Bacillus thuringiensis environmental isolates showing larvicidal activity specific for mosquitoes. J. Gen. Microbio!. 139,2849-2854 (1993).
KAELIN, P., MOREL, P., GADANI, E: Isolation of Bacillus thuringiensis from stored tobacco and Lasiaderma serricone (E). App!. Environ. Microbio!. 60, 19-25 (1994).
KAJI, D. A., ROSATO, Y. B., CANHOS, V. P., PRIEST, E G.: Characterization by polyacrylamide gel electrophoresis of whole-cell proteins of some Bacillus thuringiensis subsp. israelensis strains isolated in Brazi!' System. App!. Microbio!. 17, 104-107 (1994).
KARAMANLIDOU, G., LAMBROPOULOS, A. E, KOLIAIS, S. 1., MANOUSIS, T., ELLAR, D., KASTRITSIS, c.: Toxicity of Bacillus thuringiensis to laboratory populations of the olive fruit fly (Dacus aleae). App!. Environ. Microbio!. 57; 2277-2282 (1991).
LERECLUS, D.: Views on the ecology and the virulence of Bacillus thuringiensis. Abstracts. 29th. Annual Meeting and IIIrd Internation Colloquium on Bacillus thuringiensis. C6rdoba, Spain (1996).
MARTIN, P. A. W., TRAVERS, R. S.:Worldwide abundance and distribution of Bacillus thuringiensis isolates. App!. Environ. Microbio!. 55,2437-2442 (1989).
MEADOWS, M. P., ELLIS, D. J., BUTT, J. JARRET, P., BURGES, H. D.: Distribution, frequency, and diversity of Bacillus thuringiensis in an animal feed mill. App!. Environ. Microbio!. 58, 1344-1350 (1992).
MEADOWS, M. P.: Bacillus thuringiensis in the environment: ecology and risk assesment, pp. 193-220. In: Bacillus thuringiensis, an environmental biopesticide: theory and practice (P. E ENTWISTLE, J. S. CORY, M. J. BAILEY, S. HIGGS, eds.), John Wiley & Sons, 1993.
OHBA, M., AIZAWA, K.: Distribution of Bacillus thuringiensis in soils of Japan. J. Invertebr. Patho!' 47,277-282 (1986).
OHBA, M., ARATAKE, Y.: Comparative study of the frequency and flagelar serotype flora of Bacillus thuringiensis in soils and silkworm-breeding environments. J. App!. Bact. 76, 203-209 (1994).
SMITH, R. A., and COUCHE, G. A.: The phylloplane as a source of Bacillus thuringiensis variants. App!. Environ. Microbio!. 5~311-315(1991).
STEWART, G. S. A. B., JOHNSTONE, K., HAGELBERG, E., ELLAR, D. J.: Commitment of bacterial spores to germinate. Biochem. J. 198,101-106 (1981).
SUN, M., Luo, X., DAI, J., Qu, K., LIU, Z., Yu, L., CHEN, Y., Yu, Z.: Evaluation of Bacillus thuringiensis and Bacillus sphaericus strains from chinese soils toxic to mosquito larvae. J. Invertebr. Patho!' 68, 74-77 (1996).
TRINDADE, G. E, VILAS BOAS, L., LERECLUS, D., ARANTES, O. N.: Bacillus thuringiensis conjugation in environmental conditions. Abstracts. 29th. Annual Meeting and IIIrd International Colloquium on Bacillus thuringiensis. C6rdoba, Spain (1996).
VASQUEZ, M., PARRA, c., HUBERT, E., ESPINOZA, P., THEODULOZ, c., MEZA-BASSO, L.: Specificity and insecticidal activity of chilean strains of Bacillus thuringiensis. J. Invertebr. Patho!' 66, 143-148 (1995).
Corresponding author: Dr. J. FERRE, Dep. Genetica, C/Dr. MOLlNER, 50, 46100 - Burjassot (Valencia), Espana; Te!.: 34-6-3864506; Fax: 34-6-386 43 79; e-mail: [email protected]




















