
Cloning and Characterization of Manduca Sexta and Plutella Xylostella Midgut Aminopeptidase N...
14
Eur. J. Biochem. 248,748-761 (1997) 0 FEBS 1997 Cloning and characterization of Manduca sexta and Plutella xy lostella midgut aminopeptidase N enzymes related to Bacillus thuringiensis toxin-binding proteins Peter DENOLF'.L, Koen HENDRICKX', Jose VAN DAMME', Stefan JANSENS I, Marnix PEFEROEN I, Danny DEGHEELE' and Jeroen VAN RIE' I Plant Genetic Systems, Gcnt, Belgium ' Vakgroep Gewasbescherming.Faculteit Landbouwkundige en Toegepaste Biologische Wetenschappen, Universiteit Gent, Gent, Belgium ' ZoBlogisch Instituut, Leuven, Belgium ' Vakgroep Biochemie, Faculteit Geneeskunde, Universiteit Gent, Gent. Belgium (Received 6 June 1997) ~ EJB 97 081 1/2 We report the purification, cloning and characterization of an aminopeptidase N from the midgut epithelium of Manduca sexta that binds Cry1 Ab5, an insecticidal crystal protein [ICP] from Bacillus fhuringiensis. Sequence information derived from this M. sexta aminopeptidase N was used for the clon- ing of an aminopeptidase N from the midgut brush-border membrane of Plutrlla xylostella, an insect species of which some populations acquired resistance against Cry1 Ab5. Affinity chromatography on a CrylAb5 matrix was used to isolate a 220-kDa glycoprotein from the larval midgut of the lepidopteran M. sexta. On ligand blots the purified 120-kDa protein discriminates between the lepidopteran-specific Cry1Ab5 and the coleopteran-specific Cry3A d-endotoxin. Internal amino acid sequences from the 120- kDa protein were used for the design of degenerate oligonucleotides. From a nested PCR with M. sexta midgut cDNA as template, a DNA fragment was obtained which shows similarity to prokaryotic and eukaryotic aminopeptidase N genes. This PCR fragment was used to screen cDNA libraries of larval midguts from M. sexta and P. xylostella. From the M. sextu midgut cDNA library a 2973-bp nucleotide sequence was cloned. The ORF of the sequence encodes a 942-residue aminopeptidase N (M. sexta Apn2) containing two hydrophobic regions. The NH,-terminal hydrophobic region corresponds to a secretory signal sequence and the COOH-terminal hydrophobic region is typical of glycosylphosphatidylinositol (glycosyl-PtdTns)-anchored proteins. Low-stringency hybridization of the tl xylostella midgut cDNA li- brary with M. sexta up2 probes enabled the isolation of a 3118-bp sequence with an ORF encoding a 946-residue preproprotein. This aminopeptidase N (P xylostella Apnl) displays 61 % amino acid identity to M. sexta Apn2 and contains a COOH-terminal signal peptide for glycosyl-PtdIns anchor addition. Both M. sexta Apn2 and P. .xy'lostella Apnl contain four Cys residues, which are highly conserved among eukaryotic aminopeptidase N molecules. Treatment of Sf9 cells expressing the P xylostellu apnl gene with PtdIns-specific phospholipase C demonstrated that tl xylostella Apnl is attached to the insect cell tnembrane by a glycosyl-Ptdlns anchor. Keywords: Mundiica sexta ; Plutella xylostella ; aminopeptidase N ; Bacillus thuringiensis ; insecticidal- crystal-protein-binding protein. The interaction with high-affinity binding sites in the midgut epithelium of susceptible insect species is a tnajor determinant for the selectivity of the insecticidal crystal proteins (ICP) or 6- endotoxins of Bacillus thuringiensis. The importance of midgut Correspmzdence 10 P. Denolf. Plant Genetic Systems, J. Plateaustraat 22, B-9000 Gent. Belgium. Fux: +32 9 224 06 94. E-mail: [email protected] Ahhwviufion.s. BBMV, brush-border membrane vesicles; PtdIns, phosphatidylinositol: ICP. insecticidal crystal proteins; Apn, aminopep- tidase N; PLC. phospholipase C ; AcNPV. Autogruphu culifilrnicrr nuclear polyhedrosis virus. Enzymes. Trypsin (EC 3.4.21.4) : pliosphatidylinositol phospholipase C (EC 3.1.4.10); glycopeptide N-glycosidase (EC 3.5.1.52); aminopepti- dase N (EC 3.4.11.2) Nore. The nucleotide sequence data reported in this paper have been submitted to the EMBL, GenBank and DDBJ nucleotide sequence data- bases and are available under the accession numbers X97877 (M. .wxrci qJf12) and X97878 (i? ~~Jlo.vtellr1 apnl). target sites was demonstrated by performing in vitro binding studies with insect midgut brush-border-membrane vesicles (BBMV) and 12'I-labeled [ 1-71, ["S]methionine-labeled IS] or biotinylated ICP [9, lo]. Toxin-binding proteins have also been identified by ligand blotting (toxin overlay assays [I 1 - 19]), by incubation of larval midgut histological sections with &endotox- ins [6, 7, 20-221 and by binding experiments using the surface- plasmon-resonance technique [23 -251. In general, the results of these experiments have demonstrated the presence of one or more ICP-binding proteins in the midgut brush-border meni- brane of susceptible insect larvae. Different ICP can either com- pete for the same binding site or recognize distinct sites. It is expected that the application of different ICP types that bind to distinct sites in the insect midgut may be very useful in retarding ICP-resistance development of target insect species 1261. ICP binding in the insect midgut appears to be a two-step process characterized by an initial reversible recognition of the target molecule followed by an irreversible association of the
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Cloning and Characterization of Manduca Sexta and Plutella Xylostella Midgut Aminopeptidase N...

Eur. J. Biochem. 248,748-761 (1997) 0 FEBS 1997
Cloning and characterization of Manduca sexta and Plutella xy lostella midgut aminopeptidase N enzymes related to Bacillus thuringiensis toxin-binding proteins Peter DENOLF'.L, Koen HENDRICKX', Jose VAN DAMME', Stefan JANSENS I, Marnix PEFEROEN I ,
Danny DEGHEELE' and Jeroen VAN RIE' I Plant Genetic Systems, Gcnt, Belgium ' Vakgroep Gewasbescherming. Faculteit Landbouwkundige en Toegepaste Biologische Wetenschappen, Universiteit Gent, Gent, Belgium ' ZoBlogisch Instituut, Leuven, Belgium ' Vakgroep Biochemie, Faculteit Geneeskunde, Universiteit Gent, Gent. Belgium
(Received 6 June 1997) ~ EJB 97 081 1/2
We report the purification, cloning and characterization of an aminopeptidase N from the midgut epithelium of Manduca sexta that binds Cry1 Ab5, an insecticidal crystal protein [ICP] from Bacillus fhuringiensis. Sequence information derived from this M. sexta aminopeptidase N was used for the clon- ing of an aminopeptidase N from the midgut brush-border membrane of Plutrlla xylostella, an insect species of which some populations acquired resistance against Cry1 Ab5. Affinity chromatography on a CrylAb5 matrix was used to isolate a 220-kDa glycoprotein from the larval midgut of the lepidopteran M. sexta. On ligand blots the purified 120-kDa protein discriminates between the lepidopteran-specific Cry1 Ab5 and the coleopteran-specific Cry3A d-endotoxin. Internal amino acid sequences from the 120- kDa protein were used for the design of degenerate oligonucleotides. From a nested PCR with M. sexta midgut cDNA as template, a DNA fragment was obtained which shows similarity to prokaryotic and eukaryotic aminopeptidase N genes. This PCR fragment was used to screen cDNA libraries of larval midguts from M. sexta and P. xylostella. From the M. sextu midgut cDNA library a 2973-bp nucleotide sequence was cloned. The ORF of the sequence encodes a 942-residue aminopeptidase N (M. sexta Apn2) containing two hydrophobic regions. The NH,-terminal hydrophobic region corresponds to a secretory signal sequence and the COOH-terminal hydrophobic region is typical of glycosylphosphatidylinositol (glycosyl-PtdTns)-anchored proteins. Low-stringency hybridization of the tl xylostella midgut cDNA li- brary with M. sexta u p 2 probes enabled the isolation of a 3118-bp sequence with an ORF encoding a 946-residue preproprotein. This aminopeptidase N ( P xylostella Apnl) displays 61 % amino acid identity to M . sexta Apn2 and contains a COOH-terminal signal peptide for glycosyl-PtdIns anchor addition. Both M. sexta Apn2 and P. .xy'lostella Apnl contain four Cys residues, which are highly conserved among eukaryotic aminopeptidase N molecules. Treatment of Sf9 cells expressing the P xylostellu apnl gene with PtdIns-specific phospholipase C demonstrated that tl xylostella Apnl is attached to the insect cell tnembrane by a glycosyl-Ptdlns anchor.
Keywords: Mundiica sexta ; Plutella xylostella ; aminopeptidase N ; Bacillus thuringiensis ; insecticidal- crystal-protein-binding protein.
The interaction with high-affinity binding sites in the midgut epithelium of susceptible insect species is a tnajor determinant for the selectivity of the insecticidal crystal proteins (ICP) or 6- endotoxins of Bacillus thuringiensis. The importance of midgut
Correspmzdence 1 0 P. Denolf. Plant Genetic Systems, J. Plateaustraat 22, B-9000 Gent. Belgium.
Fux: +32 9 224 06 94. E-mail: [email protected] Ahhwviufion.s. BBMV, brush-border membrane vesicles; PtdIns,
phosphatidylinositol: ICP. insecticidal crystal proteins; Apn, aminopep- tidase N ; PLC. phospholipase C ; AcNPV. Autogruphu culifilrnicrr nuclear polyhedrosis virus.
Enzymes. Trypsin (EC 3.4.21.4) : pliosphatidylinositol phospholipase C (EC 3.1.4.10); glycopeptide N-glycosidase (EC 3.5.1.52); aminopepti- dase N (EC 3.4.11.2)
Nore. The nucleotide sequence data reported in this paper have been submitted to the EMBL, GenBank and DDBJ nucleotide sequence data- bases and are available under the accession numbers X97877 (M. .wxrc i qJf12) and X97878 (i? ~~Jlo.vtellr1 a p n l ) .
target sites was demonstrated by performing in vitro binding studies with insect midgut brush-border-membrane vesicles (BBMV) and 12'I-labeled [ 1-71, ["S]methionine-labeled IS ] or biotinylated ICP [9, lo]. Toxin-binding proteins have also been identified by ligand blotting (toxin overlay assays [I 1 - 19]), by incubation of larval midgut histological sections with &endotox- ins [6, 7, 20-221 and by binding experiments using the surface- plasmon-resonance technique [23 -251. In general, the results of these experiments have demonstrated the presence of one or more ICP-binding proteins in the midgut brush-border meni- brane of susceptible insect larvae. Different ICP can either com- pete for the same binding site or recognize distinct sites. It is expected that the application of different ICP types that bind to distinct sites in the insect midgut may be very useful in retarding ICP-resistance development of target insect species 1261.
ICP binding in the insect midgut appears to be a two-step process characterized by an initial reversible recognition of the target molecule followed by an irreversible association of the

Denolf et al. (ELM J. Bloc hem 248) 749
toxin with the insect midgut membrane 11, 3, 4, 27-29]. For several ICP, amino acids or 1CP domains that participate in the initial and irreversible binding events [30-401, or that enable the membrane insertion of the S-endotoxin [41-471, have been identified. The results from these mutagenesis studies are ex- pected to provide useful information for the design of new ICP with improved binding and insertional capacities which may en- sure the efficacy of B. thuringiensis ICP.
To understand how ICP-binding-protein characteristics con- tribute to high-affinity binding, ICP-binding molecules have been purified and cloned from different insect species. A Cryl Ab5-binding protein from Munducu sextu shares similarity with cadherins [17, 481, whereas i n M. sextu and Heliothis virescens glycosylphosphatidylinositol (glycosyl-Ptd1ns)-an- chored aminopeptidase N serves as a CrylAcl receptor [49- 521. Moreover, purification and initial characterization of the Cryl Acl-binding protein from the midgut epithelium of Ly- rnuntrin dispar [ 53, 541 and the CrylC-binding protein from M. sextu 1551 gives additional evidence that midgut aminopeptidase N is a key determinant for the binding of B. thuringiensis ICP in the larval midgut.
This article describes the purification, cloning and character- ization of a 120-kDa Cryl Ab5-binding aminopeptidase N from M. s u m , termed M. sexta Apn2. By screening with probes de- rived from this aminopeptidase N we were also able to clone and characterize an aminopeptidase N from the midgut epithelium of P. xylostella, termed I? xylostella Apnl .
EXPERIMENTAL PROCEDURES
Purification and activation of B. thuringiensis ICP. CryIAb5, CrylBa, CrylCax (named Bt15 in [4], but belong- ing to the CrylCa class of B. thuringierzsis ICP) and [Alal641Cry9Cal were expressed as recombinant proteins in E.scherichio c d i . The gene for CrylAb5 was cloned from B. thurin~ieizs~s berliner [56]. The gene for CrylBa was cloned from B. thuriiigiensis enfomocidus HD-I 10; its sequence is iden- tical to the sequence described by Brizzard and Whiteley [57]. The crjICax gene was cloned from B. thuringiensis entornoci- dus HD-I10 (Hiifte, H., unpublished data). The gene for Cry9Cal was cloned from B. thuringiensis BTS02618A and Argl64 was replaced by Ala by site-directed mutagenesis to eliminate a trypsin cleavage site at residue 164 [lo]. The Cryl Acl toxin was obtained from B. thuringiensis kurstuki HD73, which produces crystals containing only this ICP [58]. Purification of protoxins and trypsin-resistant toxins was per- formed as described by Hofte et al. [56] and Hofinann et al. [ 2 ] , except for lAlal641Cry9Ca1, which was purified according to the description of Lambert et al. [ 101. The protein concentrations of the purified and activated b-endotoxins were obtained by de- termining A,,,, with a Uvikon model 8 10P spectrophotometer, using an extinction coefficient of 76050M-' . cm I for CrylAb5, 85 100 M-' . cm-' for CrylAcl, 86400 M I . c m - ' for CrylBa, 75650M ' . cm ' for CrylCax and 57893 M-' . cm I for [Alal64]Cry9Cal.
To ensure high-specific-activity labeling, CrylCax and [Alal64]Cry9Cal were subjected to an additional purification step prior to iodination. Both toxins were loaded onto a Super- dex 75 gel-filtration column (Pharmacia) in the presence of 14 mM dithiothreitol. Following filtration dithiothreitol was re- moved by dialysis against four changes of 100 mM NaHCO,, 150 mM NaCl, pH 9. CrylAb5, CrylAcl and [Alal64]Cry9Cal were iodinated by means of chloramine-T [59]. The specific ra- dioactivity for CrylAbS was 0.5 pCi/yg CrylAb5. The lodogen-
Iodination and biotinylation of B. thuringiensis ICP.
labeling method [60] was used for CrylBa and CrylCax. Following the labeling reaction the samples were loaded onto a Biogel P-30 column to remove free iodine and degradation prod- ucts. ICP biotinylation was performed according to the mcthod described by Denolf et al. [21].
Preparation of BBMV. The midguts of fifth instar larvae from M. .sextu and P. xylostellu were dissected and BBMV were prepared by the differential magnesium precipitation method de- scribed by Wolfersberger et al. [61]. The BBMV were suspended in 150 mM mannitol, 2.5 mM EGTA, 8.5 mM Tris/HCI, pH 7.5, frozen i n liquid nitrogen and stored at -80°C. The amount of BBMV proteins was measured using the Bradford method [62] with BSA as the reference protein.
Isolation of a CrylAb5-binding protein from the midgut brush-border membrane of M. sexta. An affinity chromatogra- phy matrix for the isolation of the toxin receptor was prepared by coupling the purified CryIAb5 toxic fragment to CNBr-acti- vated Sepharose 4B (Sigma) according to the manufacturer's in- structions. The concentration of CrylAb5 on the column was 0.2 mg toxin/mg Sepharose 4B. Affinity chromatography was performed on a BioRad Econo chromatography column. BBMV were solubilized by treatment with 1 mM sodium cholate in NaCl/P, (8 mM Na2HP04, 2 m M KH,PO,, 1 5 0 m M NaCl, 50 mM EDTA and 2 mM phenylmethylsulfonyl fluoride, pH 7.4) for 1 h at 4°C. The supernatant obtained after centrifu- gation (100000Xg, 1 h) was applied to the CrylAb5 matrix. The column was washed with 1 mM sodium cholate in NaCI/P, [0.5 ml/min]. Bound proteins were eluted with 50 mM Na2C0,, pH 11. The absorbance of the collected fractions was monitored at 280 nm. Eluted fractions were concentrated on Centricon tubes with an exclusion limit of 10 kDa (Amicon). The Cryl Ab5-binding capacity of the fractions was analyzed by SDYPAGE 1631 and ligand blotting. A 120-kDa Cry I Ab5-bind- ing protein isolated with the Cryl Ab5-affinity matrix was sub- jected to amino-acid-sequence analysis.
Enzymatic treatment of BBMV. To enable an initial bio- chemical characterization of the 120-kDa Cry1 Ab5-binding pro- tein from M. sextu, BBMV were treated with N-glycosidase F and phospholipase C (PLC). BBMV proteins (200 pg) were in- cubated with 1 U N-glycosidase F (Boehringer Mannheim) i n 100 p1 NaCl/P, for 12 h and 24 h at 37 "C. As a control, untreated BBMV were incubated over the same period of time. PLC treat- ment was performed by incubation of BBMV with 1 U Ptdlns- specific PLC from B. cereus (Sigma) in 100 p1 NaCI/P, for 45 min and 90 min at 37°C. Following centrifugation (15 min, 16000 X g) of the enzymatically treated BBMV, the pellets were suspended and assayed for specific binding of 'Z'I-labeled CrylAb5 using the method described by Van Rie et a]. 131. The proteins in the supernatants were subjected to SDS/PAGE [63], transferred to nitrocellulose [64] and probed with Cryl Ab5 (0.1 pg/ml in NaCVP,). Bound CryIAb5 was detected as de- scribed for the ligand-blotting procedure (see below).
Ligand blotting. BBMV were treated with 0.5 G/c SDS, 0.5 5% 2-mercaptoethanol and 0.2% bromophenol blue in 10 mM Tris/ HCI, pH 6.8 for 1 min at 100OC. Samples were electrophoresed on gradient gels ( 5 7i to 10% polyacrylamide) [63]. Separated BBMV proteins were electrotransferred 1641 to nitrocellulose membranes. Following quenching with 0.2 % poly(viny1 pyrroli- done) and 0.25% Tween 20 in NaCl/P,, the blot was washed with NaCUP,, and incubated with the CrylAb5 toxin (0.1 pg/ml in NaCI/P,) for 1 h. Unbound toxin was removed by washing the blot in NaCUP, three times for 10 min. To detect the bound toxin the blot was incubated with the Cryl Ab5-specific mAb 2H7 (12 h, 4°C). The washing step in NaCI/P, was repeated and the blot was incubated with peroxidase-conjugated mouse anti-rab- bit Ig (Prosan; 1 :200) for 1 h. Unbound secondary antibody was

750 Denolf et al. (Eur J. Biochem. 248)
removed by washing the blot in 0.5 M Tris/HCI, pH 7.6 for 15 min. Bound toxin was visualized by incubating the blots with a peroxidase substrate solution (0.01 % 3,3'-diaminobenzidine, 0.003% H202 in 0.5 M Tris/HCI, pH 7.6).
Microsequencing of a CrylAbS-binding protein from M. sexta. Microgram quantities of the affinity-purified Cry1 Ab5- binding protein from the midgut brush-border of M. sextu were separated by SDS/PAGE as described above and electrotrans- ferred to poly(viny1idene difluoride)membranes (Immobilon). After incubation with MeOH for 5 min the membranes and the gels were placed into a blotting sandwich in a BioRad Transblot cell and transferred for 12 h at 3SV. Subsequently, the blot was stained with Naphtol Blue Black (0.1 % in 4 5 % MeOH and 7 % acetic acid) for 30 s. The protein band of interest was excised and quenched in 200 p1 1 % poly(vinylpyrro1idone) (30 kDa) in MeOH for 30 min. The solution was diluted with an equal vol- ume of H,O and incubated for 10 min. The membrane strips were washed four times with 300 p1 H,O and once with 100 mM Tris/HCI, pH 8.5 (buffer A). To perform an in situ membrane digestion the membrane bound proteins were incubated with trypsin (1 pg/ml buffer A) for 4 h at 37°C. Following digestion the membranes were washed once with 100 p1 25% trifluoro- acetic acid and four times with distilled H,O. The peptide solu- tions were separated by reverse-phase HPLC and subjected to Edman degradation with a gas-phase sequencer (model 477A; Applied Biosystems) equipped with an on-line phenyl- thiohydantoin-amino-acid derivative analyzer (model 120) [65].
Oligonucleotide synthesis. Degenerate and unique oligo- nucleotides used as primers in PCR were synthesized using the Gene Assembler Plus (Pharmacia LKB) according to the manu- facturer's instructions.
cDNA synthesis and PCR with degenerate oligonucleo- tides. Total RNA from M. sexta and f? xylostella midguts was isolated according to the methods described by Sambrook et al. [66]. 1 mg total RNA was used for the isolation of mRNA with the polyATract System (Promega). cDNA was synthesized ac- cording to the manual of the Superscript Lambda igt22A Sys- tem (Life Technologies). Degenerate oligonucleotides were de- signed using the amino acid sequence of the internal peptides P4 and P5 of the M . ,sextu Cry1 AbS-binding protein (Fig. 3a). The oli onucleotide mixture derived from P4 was: CACGAC- AT~ATYCA~GT. Using ~ 5 , two mixtures of semi-nested oli- gonucleotides were designed : 1) GTGgTANGGNGTNAC:AA ; and GTNACzAAhTCTTCNCC. Semi-nested PCRs were per- formed with M. sextu midgut cDNA as template. A 296-bp DNA fragment was obtained, which was subcloned into the T-over- hang vector pCEM-T (Promega).
Northern blot analysis. 1.5 pg M. sexto and P xylostella midgut mRNA were separated on a 0.8 o/c formaldehyde-agarose gel and blotted onto a Hybond N' membrane [66]. The mRNA was immobilized by incubation of the membrane in 0.05 M NaOH for 5 min and washed with 2 X NaCI/Cit (20 X NaCI/Cit = 3 M NaCl, 0.3 M sodium citrate, pH 7) for 1 min [671. The membrane-bound mRNA was hybridized with the [E,~ 'P]~CTP labeled 296-bp M. sextci cDNA fragment (see above) as a probe (Rediprime Labeling System. Pharmacia). Following hybridiza- tion the Hybond N ' membrane was washed with 2 X NaCI/Cit + 0.5% SDS at 50°C for 5 inin) and exposed to an X-ray film.
DNA cloning and sequencing. Directional M. sexta and f? xylostellu midgut cDNA libraries were constructed in Rgt22A using the Superscript Lambda System (Life Technologies). The M. sexta cDNA library was screened with the 296-bp PCR frag- ment (see above) as a probe. The hybridization was performed at 65°C for 12 h after which the membranes were washed at 65 "C twice for S min with 2 X NaCI/Cit + 0.5 9% SDS, twice for 10 min with 2 X N a C K i t + 0.1 70 SDS, once for 20 min with
8
0.1 XNaCl/Cit + 0.5% SDS, and once for 20 min with 0.1 X NaCl/Cit + 0.1 % SDS. The insert size of purified recom- binant plaques was evaluated through PCR with forward and reverse primers (Life Technologies) flanking the multiple clon- ing site into igt22A. Two cDNA clones were excised from the phage vector with MluI and NotI. To subclone the insert into SmaIINotI-digested pBlueScript I1 KS ~ (pBSKS- ; Strategene) the MluI-digested M. .sexto cDNA was filled in with Klenow DNA polymerase (Biolabc) prior to the Not1 digestion.
The P. .xylostella cDNA library was screened at low strin- gency with a 296-bp (see above) and a 696-bp fragment (from position 1790 to position 2485 in M. sexta u p 2 ) derived from the cloned M. stxxtu gene. Low-stringency hybridization was per- formed at 60°C for 12 h. Following hybridization the mem- branes containing the recombinant phages were rinsed twice with 2 X NaCl/Cit + 0.5 % SDS for 5 min at 50°C and once for 5 niin with 2 X NaCI/Cit + 0.1 % SDS at 50°C. The DNA from a positive recombinant phage was SulllNotI-digested and sub- cloned into SulIINotT-digested pBSKS . A 1909-bp fragment of the subcloned f? x ) h t e l l u cDNA (from position 781 to position 2689 in P. xylostella upti / ) was used in a high-stringency screen- ing of the F! xylostellu library [hybridization at 65°C for 12 h ; washing at 65°C with 2 X NaCl/Cit + 0.5% SDS (twice for 5 min), 2XNaCUCit + 0.1 % SDS (twice for 10 min), 0.1 X N a C K i t + 0.5 % SDS (20 inin) and 0.1 XNaCl/Cit + 0.1 5% SDS (20 min)]. Following insert-size control by PCR, two cDNA clones were excised from the phage vector, subcloned into Sull/NotI-digested pBSKS - and electroporated into Epicu- rian E. coli SURE cells (Stratagene). Microgram quantities of the cloned genes were obtained with the Wizard Midiprep DNA Purification System (Promega). Sequence analysis of both strands of the cloned genes was performed using an A. L. F. DNA sequencer (Pharmacia).
Expression of I? xylostella Apnl in Sf9 cells. To subclone P. xylostella u p 1 into the SmaI/NotI-digested baculovirus transfer vector pVL1393 (Pharmingen), the gene was SalI di- gested, filled in with Klenow DNA polymerase and NotI di- gested. To generate recombinant Autographs culifornica nuclear polyhedrosis virus (AcNPV) carrying the f? xylostellu apnl gene, 3 X 10" Sf9 cells were cotransfected with 1 pg pVLl393- f? .x.ylo.stella upnl and 0.25 pg BaculoGold DNA (Pharmingen) by the calcium phosphate coprecipitation method. After 4 days, the supernatant of the cotransfection plate was collected and re- combinant viruses were screened by end-point dilution. The in- sert size of recombinant viruses was evaluated by performing a PCR with two oligonucleotides flanking the pVL1393 multiple cloning site. The 5' end of the inserted P. xylostella apnl was sequenced to check its integrity. The transfection supernatant was amplified 4 times to produce a high-titer solution of the recombinant virus. Virus supernatant was stored in the dark at 4°C. Sf9 cells were maintained in suspension culture in TClOO medium (Life Technologies) supplemented with 10% fetal calf serum and 0.05 mg/ml gentamycin at 27°C. For the production of recombinant f? xylostella Apnl, cells were subcultured in monolayer and infected at a multiplicity of infection of at least 1.
Sf9 membrane preparation. Sf9 cell membranes were pre- pared according to the method described by Butkerait et al. [68]. Cells were harvested 84 h post infection, washed twice with 0.9% NaCI, counted and suspended in ice-cold 20 mM Hepes, pH 8, 2 mM MgC12, 1 mM EDTA, 0.1 mM phenylmethyl- sufonyltluoride, 10 pgfml leupeptin and 2 pgiml aprotinin. Following incubation on ice for 5 min, 1 X 10" cells were ho- mogenized by repeated passage through a 25-gauge needle. Homogenates were centrifuged at 100 X g for 5 min, and the resultant supernatant at 16000 X g for 30 min. The final pellet,

Denolf et al. (Eur J. Biochem. 24K) 75 1
A B C 1 2 3
1 2 1 2 1 2
kDa
I70 -
6 1 -
a
kOa
170 -
b Fig. 1. Ligand-blot analyses of immobilized, non-purified and CrylAb5-affinity purified M. sexta BRMV proteins. (a) Nitrocellulose-immobi- lized non-purified and CrylAb5-affinity-purified proteins from the midgut of M . sextu. Lanes 1, 20 pg M. sexta BBMV proteins; lanes 2, CrylAbS- affinity-purified BBMV proteins eluted with SO mM Na,CO,, pH 11. (A) Blot stained with indian ink; (B and C) results from a ligand-blotting assay. In (C) the proteins were incubated with the CrylAbS toxin, whereas in (B) Cryl AbS was omitted to check for potential cross-reaction of the antibodies used. (b) Cry 1 Ab5-affinity-purified proteins from M. sextu. The affinity-purified proteins were concentrated on a filter with a 100-kDa cutoff, separated on a polyacrylamide (5% to 10%) gel and subjected to ligand blotting. Lane 2 was screened for CrylAbS binding. In lane 1, CrylAb5 was substituted by bovine serum albumin. Lane 3 was screened for Cry3A binding using a polyclonal antiserum against the coleopteran active-toxin.
representing the membrane fraction, was suspended in the above RESULTS buffer.
sextu. Sodium-cholate-solubilized BBMV from M. .sexta were wild-type or recombinant AcNPV were harvested 84 h post in- applied to a CrylAb5 matrix. An increase of pH was used to fection and washed with NaCVP,. Samples consisting of lo6 Sf9 elute the binding molecules from the affinity matrix. 50 mM cells were suspended in 100 pl NaClP, and mixed with 100 p1 Na2C0,, pH 11, enabled the partial purification of proteins with substrate solution ( 5 mM leucine-p-nitroanilide in 67 mM molecular masses ranging from 30 kDa to 200 kDa (Fig. 1 a). In
in A,,,,/min was measured over 20 min using a kinetic microtiter 2H7 a 120-kDa protein was identified as a CrylAb5-binding protein (Fig. 1). When using non-purified, solubilized BBMV plate reader (BIO-TEK Instruments).
infection and washed with NaCI/P,. Samples consisting of 10' tein (Fig. The 67-kDa protein that was visualized in the cells were suspended into 100 pl NaCVP, and incubated with ligand blot with the affinity-purified M. .sexfa BBMV proteins 0.5 U PtdIns-specific PLC from B. cereus (Sigma). Following and that was stained when the CrylAb5 was omitted in the li- incubation at 30°C for 10 min and 240 min the cells were centri- gand.blotting procedure represents c r y 1 ~ b 5 leaking from the fuged at 16000xg for 10min. The Pellet was suspended in affinity matrix (Fig. la) . When the pH-11 eluent was screened 100 NaCI/P, and, together with the supernatant, screened for for binding of the coleopteran toxin Cry3A, no binding could be aminopeptidase N activity by the addition of 100 p1 substrate demonstrated (Fig. Ib). (see above). Treatment of M. sexta BBMV with N-glycosidase F or
ICP-binding to SB(R xylostellu Apnl) cell membranes Ptdlns-specific PLC reduced CrylAb5 binding to 5% of the ini- and intact SB(R xylostellu Apnl) cells. The conditions applied tial binding. "'I-Cry1 Ab5 binding to untreated BBMV did not for the binding experiments with '251-labeled ICP and membrane decrease in the same time period (Fig. 2). A ligand-blotting ex- preparations from transfected Sf9 cells were essentially the same periment demonstrated that the supernatant of PLC-treated as described by Van Rie et al. [3]. For each data point 1 pg Sf9 BBMV contained a CryIAb5-binding 120-kDa protein (data not membrane protein was used. In binding experiments with intact shown). Sf9 cells, 2 X 10' cells were transferred to coated cytoslides (Shandon) by centrifugation at 500 rpm for 5 min in a CytosPin Sequencing of the 120 kDa CrylAb5-binding protein. An at- 2 centrifuge (Shandon). The binding of biotinylated I c p was tempt to determine the NH,-terminal amino acid sequence of the evaluated as described previously by Denolf et al. [21]. 120-kDa Cryl Ab5-binding protein failed, probably due to the
Carbohydrate staining of E! xylostella BBMV proteins blocking of the NH,-terminal amino acid. However, by in situ and S B proteins. The digoxygenin glycan-detection kit (Boeh- digestion of the poly(viny1idene &fluoride)-immobilized 120- ringer Mannheim) was used to evaluate the presence of glyco- kDa protein with trypsin and subsequent Edman degradation, proteins in P: xylostellu BBMV and in wild-type and recombi- several internal-peptide sequences were obtained: PI, IPNIN; nant Sf9 cells. P2, TTIYAPAAYATVR; P3, AAPSDLYVALQQALDE ; P4,
Aminopeptidase N assay. Intact Sf9 cells transfected with Purification Of a Cry1Ab5-binding protein from
Na2HPo4, 67 mM KH,P04, pH 7'4) [69i. The change a ligand-blotting experiment with the Cry1AbS-specific mAb
pLc treatment. Infected sf9 were harvested 84 post proteins from M. sexfa, CrylAb5 also bound to a 120-kDa pro-

752 Denolf et al. (ELM J. Biochern. 248)
12 h 24 h
N - glycosidase F treatment
phospholipase C treatment
Fig.2. Specific binding of 1Z51L-CrylAb5 to N-glycosidase F and PtdIns-specific PLC-treated BBMV from M . sexta. The specific bind- ing was calculated relative to the binding of "'I-CrylAb5 to BBMV incubated at 4°C (specific radioactivity for CrylAb5 was 0.5 pCi/pg CrylAbS). Bars represent binding after incubation at 37°C in the ab- sence (*) or presence of N-glycosidase F or PtdIns-specific PLC for the times indicated. In the absence of modifying enzymes, BBMV were incubated for 24 h (glycosidase control) or for YO min (phospholipase control). Standard deviations are indicated with error bars.
NSHEIIHVLNR; PS, SGEEFVTPYHR; P6, (H or E)ESIF- SAGER; and P7, NIAASIAX(S or I)N(S or I)NA(G or M)I(V or M).
cDNA cloning, sequencing and characterization. Using de- generate forward primers derived from peptide P4 and reverse primers derived from peptide PS in PCR with M. .sexta midgut cDNA as template, a 296-bp DNA-fragment was amplified. Alignment of the subcloned and sequenced DNA with the se- quences in the EMBL nucleic acid data bank demonstrated simi- larity of the 296 bp cDNA sequence with rat, rabbit, human and mouse aminopeptidase N coding sequences 170-731. This se- quence was not found in the gene of a CrylAcl-binding protein cloned by Knight et al. [SI]. To distinguish the M. sexta amino- peptidase-N-like genes from each other they are referred to as M. .sexto upnl [51] and M. sexta upn2 (this work).
From a high-stringency screening of the M . sexfa midgut cDNA library with the 296-bp PCR fragment 10 of 200000 screened recombinants were plaque purified. To maximize the chances of obtaining a full-length clone, the insert size of the isolated recombinants was determined by performing a PCR with phage 3.gtl1 forward and reverse primers flanking the insert (Life Technologies). Two subclones with estimated sizes of 2960 bp were retained for sequencing. As a result, a 2973-bp DNA fragment was obtained from the M. sexta midgut cDNA library (Fig. 3 a). This fragment contains an ORF of 2826 bp ( M . sextu apn2) encoding a protein of 942 amino acids ( M . sextu Apn2), which contains all the tryptic peptides derived from the 120-kDa Cryl AbS-binding protein from M. sexta. The bases surrounding the putative translation-start codon at positions 22 -
24 only meet the minimum criteriuin of Kozak for translation initiation, namely the presence of a purine 3 nucleotides up- stream from the start codon [74]. The 126-bp 3'-untranslated region is characterized by the presence of a polyadenylation sig- nal sequence, AATAAA, at position 2954 (Fig. 3 a).
The PC-GENE program was used to analyze the M. sexta Apn2 amino acid sequence. The nascent form of M. sextu Apn2 has a calculated molecular mass of 107535 Da and a PI of 5.58. From a hydropathy plot it was concluded that the protein is char- acterized by the presence of NH,-terminal and COOH-terminal hydrophobic regions, whilst the region in between has an overall hydrophilic character (1751; data not shown). Using the weight- matrix method of von Heijne [76], a signal-sequence cleavage site conforming to the [-3,-11 rule was predicted between Gly15 and Val16 with a score of 10.51. From the alignment of the amino acid region between Tyr2 and Gly1.5 with amino acid sequences in the Swiss Prot data bank we observed significant similarity to the signal peptides of, for example, trypsinogen TIT precursor from rat [77], trypsinogen I precursor from the atlantic salmon [unpublished results] and Ig heavy-chain precursor V region from mouse [78]. Analysis of the COOH-terminal hy- drophobic region between Ser920 and His942 gives strong evi- dence that this is a glycosyl-Ptdhs anchor signal sequence. This region starts with three small amino acids (Ser920 at cu, Gly921 at cuS1, and Ala922 at w+2) which fulfil the criteria for glyco- syl-PtdIns-anchor addition as determined by Kodukula et al. [79]. The glycosyl-PtdIns-anchor signal sequence ends up with a stretch of hydrophobic amino acids (Fig. 3a). The M. sexta Apn2 amino acid sequence in between the outer hydrophobic regions is characterized by the presence of five consensus glyco- sylation sites and by the presence of the zinc signature 180; Figs 3a and 41. Comparison of M. sextu Apn2 with amino acid sequences i n the SwissProt data bank showed significant identity to aminopeptidase N from rabbit (22% identity; [71]), human (20% identity; 172]), rat (19% identity; [70]) and pig (18% identity; [81]), and to human aminopeptidase A (16% identity; [82]). Moreover, M. sextu Apn2 showed 21 % and 23 G/o amino acid identity to the Cryl Acl-binding proteins cloned previously from M. sexta 1511 and H. virescens [52], respectively. For com- parison of these amino acid sequences the method of Myers and Miller [831 was applied. Using this method the Cry1 Acl-binding proteins from hf. sextu and H. virescens displayed 38% amino acid identity. M. .sexta Apn2 contains the sequence QNMIALE, starting at Q275, which was reported to be present in the CrylC- binding protein of M. sextu 1551.
Low-stringency cloning from a f! xylostella midgut cDNA li- brary. From a low-stringency northern hybridization it was con- cluded that the 296-bp PCR fragment of M. sextu upn2 cross- hybridized with a F! xylostella midgut mRNA band of about 3100 bases, which was slightly larger than the M. s e m tran- script (Fig. 5) . This result demonstrated the presence of closely related aminopeptidase-N-like genes i n the midgut of M. sexto and I? xylostella. The 296-bp fragment and a 696-bp fragment (see above) of M. .sextu upn2 were used for a low-stringency screening of the F! xylostella midgut cDNA library. This screen- ing yielded several recombinant plaques, one of which was puri- fied, and the cDNA fragment subcloned into pBSKS-. Partial sequencing of this subclone enabled the design of primers that were used to generate a 1909-bp PCR fragment. This 1909-bp fragment was used as a probe for a high-stringency screening of the F! xylostella midgut cDNA library. Ten recombinant plaques were purified and the cDNA insert size was estimated from a PCR with insert-flanking Agtll forward and reverse primers. Two cDNA inserts of about 3100 bp were subcloned. Nucleic- acid-sequence analysis yielded a 31 18-bp DNA fragment con-

Denolf et a1 ( E M .I Bm/?ern. 248)
A M SEXTA-2
81 108 27 54 QCg tCC gaa tac ata tBC aat ATQ TAT TCA CTA ATA TTC TTG GCT CTC ATC GGA GCG GCG TTT GGT GTC CCA CTG AGC ACC AAC GAA GAC AGC ACT AGA AAC CAA AAC
M Y S L I F L A L I G A A F G V P L S T N E D S T R N Q N
135 162 CTA GCT GCG CTA TAC GTC TTG CCT CAG ACT TCC TAC CCG ACG TTC TAT GAC GTC L A A L Y V L P Q T S Y P T F Y D V
243 270 AGG ATA ATC CCC AAC ATC AAC ATT GAT CAG ATC ACC ATC CAT GCC ATG GCT AT0 R I I P N I N I D Q I T I H A M A M
351 378 TCT GAC TTC ACG CTG GCT ACT GAC GAC ACC CAT CTC TTG ACT ATC AGG CTG ACG S D F T L A T D D T H L L T I R L T
459 486 TAT GCC GAT GAC ATG TTT GGC GTC TAC GTT TCT ACT TAT GAG GAG AAT GGT AGO Y A D D M F G V Y V S T Y E E N G R
567 594 CCC TGC TAC GAT GAG CCG GCT TTA AAA GCC GTT TTC AGA ACC ACC ATC TAT GCC P C Y D E P A L K A V F R T T I Y A
675 702 AAA CCT AAT GAG CCT GGC TAC GTA AAG CAC GAG TTC GAA GAC ACC CTG GTC ATG K P N E P G Y V K H E F E D T L V M
783 810 CAG AAC CCC ATC TAT CCT ATT CCC TTC AGA GTC TAC TCT AGG CCT GGT ACT CAG Q N P I Y P I P F R V Y S R P G T Q
891 918 GAG TAC ACC GAA TTC CCG TAC GCT TTC CCC AAA ATC GAC AAG GCT GCC GTT CCT E Y T E F P Y A F P K I D K A A V P
999 1026 GCC CTT CTC GTA AGG GAA GGT GTG ACG ACA ACG TCG GTG AAG CAG AAC ATT GGC A L L V R E G V T T T S V K Q N I G
1107 1134 ATG TCC TGG ACC TAC ACT TGG CTC AAC GAA GGA TTC GCT AAC TTC TTT GAG AAC M S W T Y T W L N E G F A N F F E N
1215 1242 GCC ATG CAG AAC GTG TTT CCA GTC AGA CGC TGT TCT AAG TGT CAA CCC CAT GAC A M Q N V F P V R R C S K C Q P H D
1323 1350 CCA GAA GTC TGG TTC CGT TAT TCG GAT GTT GCA GCA TTT CAT GAC ACA AGA GAT P E V W F R Y S D V A A F H D T R D
1431 1458 CCC TCA GAT CTC TAC GTT GCC CTC CAD CAA GCC CTC GAT GAG TCC AGT CAC AGA P S D L Y V A L Q Q A L D E S S H R
1539 1566 CCT GTC CTC ACT GTT AGG AGG ACG GCG CCC AAT GCT GAC TCC GTG TTT GTA GCC P V L T V R R T A P N A D S V F V A
1647 1614 CCT GTA AAC TGG GTT ATA TCC TCA AAC GTG AAC TTC AGT GAT ACC AGC CCT CAA P V N W V I S S N V N F S D T S P Q
189 216 CGT CTC TTC ATC GAT CCT GGA TAC ACC GAR GCA TTC CAT GGA AAC GTT TCC ATC R L F I D P G Y T E A F H G N V S I
291 324 AGA ATC GAC TCG ATC AGA GTT GTC AGT GAT GTT AAT CCT AAT GAG GAC CTC TTC R I D S I R V V S D V N P N E D L F
405 432 AGO AAC ATC ACC GCT TTA CAA CCC CAT GTT ATC CAT ATT GAT TAT GTT GCA CAA R N I T A L Q P H V I H I D Y V A O - -. . . . . . .
513 540 ACA GTA AAC CTC GTG ACA TCT CAA CTG CAG CCT ACT TTC GCC CGC CGT GCG TTC T V N L V T S Q L Q P T F A R R A F
621 648 CCA GCG GCT TAC GCC ACA GTC AGA AGC AAC ACT CCT GAA AGG CGT GAT TCC CTC P A A Y A T V R S N T P E R R D S L
729 756 TCC ACC TAC CTC ATT GCC TAC CTG GTC TCC AAC TTT AAC TAC ATC GAG RAT TCC S T Y L I A Y L V S N F N Y I E N S
831 864 AAT ACC GCT GAG TTC GCC CTT GAG TTT GGC CAG CAA AAC ATG ATT GCT CTG GAG N T A E F A L E F G Q Q N M I A L E
945 972 GAC TTT GCT GCT GGT GCC ATG GAG AAC TGG GGA CTG GTT ATT TAC AGA GAG GTA D F A A G A M E N W G L V I Y R E V
1053 1080 AGG ATC ATT TGC CAC GAA AAC ACC CAC ATG TGG TTT GGA AAC GAA GTC GGA CCT R I I C H E N T H M W F G N E V G P
* I
1161 1188 TAC GCT ACT GAT TTT GTC AGG CCG CAG TGG CGC ATG ATG GAT CAG TTT GTG ATC Y A T D P V R P Q W R M M D Q F V I
1269 1296 GCA CCC GGT CTA TAC TCC TTC CCA GAT CAT CGG TAC TTT CAA CGC CGT CGC TTA A P G L Y S F P D H R Y F Q R R R L
1377 1404 TTT CAG GAG AGG TCT GOT CAT CTA CAT CAA AGC CAA CTC TCG CGA CCT GCC GCT F Q E R S G H L H Q S Q L S R P -
1485 1512 ATC CCC AAA CCC ATC TCG ACT ATC ATG ACT GAA TGG AGC ACT CAG GGA GGC TTC I P K P I S T I M T E W S T Q G G F
1593 1620 CAG GAA CGT TAC CTC ACC GAC CGG TCT CTG ACA TCC ACT GAC CGC TGG CAC GTG Q E R Y L T D R S L T S T D R W H V
1701 1728 GCC TGG ATC CTG CCG ACC TTC CCC GCC ACC GCC GTT GAC GTT CCT GGC CTC TCC A W I L F T F P A T A V D V P G L S
1755 1782 AAT GCT GAT TGG TAC ATC TTC AAC AAG CAG CAG ACT GGC TAC TAC CGT GTC AAC N A D W Y I F N K Q Q T G Y Y R V N
1863 1890 ATC ATC CAT GTC CTC AAC CGT GCT CAG ATT GTC GAC GAC GCA TTC AAC TTG GCC I I H V L N R A Q I V D D A F N L A
1971 1998 ATG GAA AAA GAT TAC ATC CCA TGG GCT GCT GCC AAC CCT GCC TTC AAC TAC TTG M E K D Y I P W A A A N P A F N Y L
2079 2106 AAC CTT ACT GCT CCT ATG TTC GAG GAC CTT GGC TTT GAT GTG AAA TCT GGC GAA N L T A P M F E D L G F D V K S G E
~ ....._ ~.
2187 2214 GGC AAC CAG CGG TGC ATC AGC AGG GCC CAG GAA ATC CTG CAA GCT TTC AAG RAT G N Q R C I S R A Q E I L Q A F K N
2295 2322 CTA CGC GCC GGT AAC GTG GAG AAC TTC AAC TTC CTC TGG AAC ATG TAT CTA GGC L R A G N V E N F N F L W N M Y L G
2403 2430 AAT GCT GAA CGT CGT AAC TTC TAT CTG AAC CAA ATC ATC GAC GAT AAC TCT GCA N A E R R N F Y L N Q I I D D N S A
2511 2538 GAR GGC A1G AAC GTC GCT TTG GAC TTT GTC GTT GAA AAC TTC CAC AGA ATA CAA E G M N V A L D F V V E N F H R I Q
2619 2646 CGA CTG ACC ACA AGC GTT CAT AAT GAA AAA ATC GAT GAG CTG GTC AGG CGA CAC R L T T S A H N E K I D E L V R R H
2727 2154 ATC GCA GCC TCC ATC GCT TGG AGC AAT AGC AAT GCA GGC ATC GTC GAA AAT TGG I A A S I A W S N S N A G I V E N W
2835 2862 CTT GTT CTC ATC TCT CTC TTT GTG GCT ATT TTC AAT CAT tag aat tta Ctt att L V L I S L F V A I F N H .
2943 2970 ttt tat att ttt tat act taa tct act aat ttg tgt aaa taa aat att tat aat
1809 1836 TAC GAC GTC GAA AAC TGG GTC GCT CTG GCG CGA GTC CTC AAT RAC TCT CAC GAG Y D V E N W V A L A R V L N N S H E
. . . . . . . . . 1917 1944
AGG AAC GGA CGT CTG CAC TAC AAA AAC GCC TTC GAA ATC TCA AGA TAC TTG GAG R N G R L H Y K N A F E I S R Y L E
2025 2052 GAC ATC GTC CTT AGT GGC GCC AAT TCC TAC AAT CTG TAC AGG TAC TAC CTG CTC D I V L S G A N S Y N L Y R Y Y L L
2133 2160 GAG TTC GTC ACT CCT TAC CAC AGG AAC ATT ATT CTG GAC ATT AAC TGC CGT TTT I ? F V T P Y H R N I I L D I N C R F
2241 2268 AAC CCT AAC CAG CGC CCC AAC CCT GAC ATC CAG ACC TTG GTC TAC TGC TCC AGT N P N Q R P N P D I Q T L V Y C S S
2349 2316 ACG TCC GAC TCC AGC GAG CAG AGC ATC TTG CTT AGT GCC CTG GGA TGC ACT AGC T S D S S E Q S I L L S A L G C T S
2457 2484 GTG CGA GAG CAG GAC AGA CAC TCA ATC GCC GTC TCC GTC ATC AAC TCC AGC CCT V R E Q D R H S I A V S V I N S S P
2565 2592 CCC AGG GTC CAG GCT CTT ACA GGA ACA ACT AAC ATT CTG AAC ACC TTT GCA AGA P R V Q A L T G T T N I L N T F A R
2673 2700 GAG AGC ATC TTC TCA GCC GGA GAG CGT GCG TCC ATC GCC GCT ATT AGA GAG AAT E S I F S A G E R A S I A A I R E N
2181 2808 CTG AAA GAG AAT TAC GGG CCA CCC AGT GGC GCC AAA TCC TTA ACC GCT GGA TTA L K E N Y G P P S G A K S L T A G L
2889 2916 qta att tta tat tta ttt tat aat aat ttt acg ata aaa tat ttt tat act tab
att
753
Fig. 3. Sequences of cDNAs and deduced amino acids of M . sextu upn2 (A) and P. xylostellu upnl (B). The putative translation start sites and the polyadenylation signals are in boldface. 5' and 3' untranslated regions are in lowercase letters while the coding sequences are in capitals. The putative NH,-terminal transport signal peptide and the glycosyl-PtdTns signal peptide are double underlined. The zinc-metallopeptidase signature is irvlicated with asterisks. Peptide sequences obtained from the Cry1 Ab5-affinity-purified M . sexm 120-kDa protein are underlined. Consensus N- linked glycosylation sites are broken underlined. The four Cys residues that are conserved among eukaryotic aminopeptidase N genes are in boldface capitals.

754 Denolf et al. ( E m J . Biockern. 248)
B P.XYLOSTELLA
27 54 81 108 ttt gtg cag tga cgg tgc tgt ttt agg ata ata aat agg gga taa ata ata ata gag A% CGT TTA CTT ATC TGC CTG ACT CTG TTG GGC TTA GTC TGC GGG AAC CCC
M R L L I C L T L L G L V C G N P - 135 162 189 216
CTG CCA GGG GAG TCA TTC CCG ACC TTC TAC GAT GTT CAA CTG TTC TTT GAC CCT GTG CAA CTG ACA GAC AAC AGC ATC GCG CTA CAA AAC ACA TAT GAC AAC TAC GTT V Q L T D N S I A L Q N T Y D N Y V L P G E S F P T F Y D V Q L F F D P
243 270 297 324 GCT ACA CAA GAG ATT GTC CTA CAC GCT ATG GAA ATG GAG ATA CTG TCT ATC CGT GAA TAC GAA GCG TCC TTC AAT GGC ACA GTA GCA ATA AGA GTC GTG CCC AGA ATT
E Y E A S F N G T V A I R V V P R I A T Q E I V L H A M E M E I L S I R
351 378 405 432 TAC ACT TTG GCT ACT GAT GAC ACG CAT CTT CTG AAG ATT CAG TTC ACT AGA GTT GCT TAC TCA GAC TTG CCT AGT GAT GAT AAT TTG AAT GAA h T TTG TTC TCA AGC
A Y S D L P S D D N L N E N L F S S Y T L A T D D T H L L K I Q F T R V
459 486 513 540 TTA GAT GCA CTG CAG CCA ATC ACA GTT GAG ATC TCG TAC TCG GCT CAG TAT GCG L D A L Q P I T V E I S Y S A Q Y A P N M F G V Y V S R Y V E N G A T V
CCC AAC ATG TTT GGA GTC TAC GTG TCC AGA TAT GTG GAG AAT GGA GCT ACT GTG
567 594 621 648 TCC CTG GTG ACA TCC CAG CTG CAG CCC ACG TTC GCT CGC CGC GCG TTC CCG TGC S L V T S Q L Q P T F A R R A F P C Y D E P A L K A V F R T T I Y A P P
TAC GAC GAG CCG GCC CTG AAG GCA GTC TTC CGC ACC ACC ATC TAC GCG CCG CCG
675 702 729 756 GAC CGG CCA GGC TTC ACC AAG CAC GAG TTC CAG GAC ACC CTG GTG ATG TCC AGC GCG TAC AAC GTG GTC GAG ACC AAC ATG CCT TTG AGG ACC GAT AGT TTG AAG TCT
A Y N V V E T N M P L R T D S L K S D R P G F T K H E F Q D T L V M S S
783 810 837 864 CCC ACC TAC GAC AAA TCC ATG AAA GTG TTC TCG AGA CCA GGA ACC CAG AAC ACG TAC CTG CTG GCC TAC CTG GTC TCC AAG TTC GAC TAC ATC TCC AAC GAA AAC AAC
Y L L A Y L V S K F D Y I S N E N N P T Y D K S M K V F S R P G T Q N T
891 918 945 972 ACT GAG TTT CCT TAC GCC TTC CCG AAG ATC GAC AAA GTG GCG GTG CCG GAC TTC GCA GAG TTC GCT TTA GAC TTT GGA CAG AAG AAC ATG GTG GAA CTG GAG AAG TAT
A E F A L D F G Q K N N V E L E K Y T E F P Y A F P K I D K V R V P D F
999 1026 1053 1080 GCC GCT GGT GCC ATG GAG AAC TGG GGA CTG GTT ATT TAC AGA GAA ATA GCG CTG A A G A M E N W G L V I Y R E I A L L V Q E G V T T T S T L Q G I G R I
CTA GTC CAA GAA GGC GTG ACG ACC ACC TCC ACG CTG CAG GGC ATC GGC CGG ATC
1107 1134 1161 1188 ATC TCG CAC GAG AAC ACG CAC CAG TGG TTC GGC AAC GAG GTT GGG CCC GAC TCC TGG ACC TAC ACC TGG CTG AAT GAG GGC TTC GCC AAC TTC TTC GAG AGC TTC GCT I S H E N T H Q W F G N E V G P D S W T Y T W L N E G F A N F F E S F A . .
1215 1242 1269 1296 ACT GAC CTT GTC ClC T(A GAA TGG CGG ATG ATG GAC CAG TTC GTC ATC AAC ATG CAG liAC GTG TTC CAG TCG GAC GCT GTG CTC TCC GTC AAC CCG ATA ACC TTC GAG T D L V L P E W R M M D Q F V I N M Q N V F Q S D A V L S V N P I T F E
1323 1350 1377 1404 TCT GGA TCC GTT RTT CGC ATG ATG CAG CAC TTC TTG ACC CCA GAA ATC TTC CGA GTG CGG ACC CCC TCG CAG ATC CTC GGC ACC TTC AAC TCG GTC GCT TAT CAG AAG
V R T P S Q I L G T F N S V A Y Q K S G S V I R M M Q H F L T P E I F R
1431 1458 1485 1512 AAG AGT CTT GCT CTA TAC ATT TCC ADA ATG TCT CGC AAA GCC GCC AAG CCC ACT GAC CTG TTC GAA GCG ATC CAA GAA GTA GTG GAC GCG TCA GAC CAC AGC ATC CGC K S L A L Y I S R H S R X A A K P T D L F E A I Q E V V D A S D H S I R
1539 1566 1593 1620 TGG CGC CTG TCC ATC ATC ATG AAC CGG TGG ACC CAG CAG GGG GGA TTC CCC GTA GTC ACC GTC AGA CGG TCG GCG CCC TCC GCA CAG TCA TTT GTT ATC ACT CAG CGC W R L S I I M N R W T Q Q G G F P V V T V R R S A P S A Q S F V I T Q R
1647 1674 1701 1728 CGC TTC CTA ACA GAC AGC ACC CAA GAG TCC AAC ACG GTG TGG AAC GTG CCT CTC AAC TGG GTG CTC AGC ACT GAC GTG AAC TTC AAC GAC ACG CGC CCC ATG GCC TGG R F L T D S T Q E S N T V W N V P L N W V L S T D V N F N D T R P M A W
1755 1782 1809 TTG CCG CCC CAG CTC GCG GCC GAG GCG GTC CAG GTC CCT GGA CTC CAG AAC GCT GAG TGG TTC ATT GTC AAT AAG CAA CAG L P P Q L A A E A V Q V P G L Q N A E W F I V N K Q Q
1836 ACT GGT TAC TAC CGC GTA AAC TAC GAC T G Y Y R V N Y D
1863 1890 1917 1944 CCT GAA AAC TGG AGG GCT CTG GCT AAA GTC CTG AAT GAC ACG CAC GAG ATC ATC CAC CTA CTG AAC CGG GCT CAG CTC ATC GAC GAC TCA TIC AAC CTG GCC AGA AAC P E N W R A L A K V L N D T H E I I H L L N R A Q L I D D S F N L A R N
1971 1998 2025 2052 GGA CGT CTT GAC TAC AGC TTG GCC TTC GAC CTG TCC CGG TAC CTG GTA CAG GAG CGC GAC TAC ATC CCG TGG GCA GCC GCC AAC GCC GCC TTC AAC TAT CTC AAC TCC G R L D Y S L A F D L S R Y L V Q E R D Y I P W A A A N A A F N Y L N S
2079 2106 2133 2160 GTG CTG AGC GGC TCT TCT GTA CAC CCC CTG TTC CAG GAA TAC CTT CTC TTC CTC ACC GCG CCG CTT TAC CAG AGG CTT GGC TTC AAC GCC GCT ACC GGC GAA GAG CAT V L S G S S V H P L F Q E Y L L F L T A P L Y Q R L G F N A A T G E E H
2187 2214 2241 2268 GTC ACT CCC TTC CAT CGC AAT ATT ATC CTG AAC ATC AAC TGT CTC CAC GGC AAT GAG GAC TGC GTC AGC ACC GCC GAA ACT CTG CTG CAG AAC TTC AGA GAC AAC CCA V T P F H R N I I L N I N C L H G N E D C V S T A E T L L Q N F R D N P
2295 2322 2349 2376 ACT CAA ACT CTA AAC CCG GAT ATC CAG ACC ACG GTG TTC TGT TCT GGT CTG CGC GGC GGA GAC GTG GAT AAC TTC AAC TTC CTG TGG GCG CGG TAC ACC GCC ACA CAG T Q T L N P D I Q T T V F C S G L R G G D V D N F N F L W A R Y T A T Q
2403 2430 2457 2484 GAC TTC AGT GAA CAG TCC ATC TTG CTC AAC GCT CTG GGA TGC ACG TCT AAT GCC GAT AGG AGG GAC TTC TTA TTC AGC CAA GTT ATT GCA TCA GAT TCC CAA GTG AGA D S S E Q S I L L N A L G C T S N A D R R D F L F S Q V I A S D S Q V R
2511 2538 2565 2592 GAG CAG GAC AGA CAT TCA GTT CTC GTG TCT GCC ATC AAC TCA GGC CCT GAC AAC ATG AAC GCA GCT TTG GAC TTC GTC TTA GAG AAC TTC GCT AAT ATA CAA CCA AAC E Q D R H S V L V S A I N S G P D N M N A A L D F V L E N F A N I Q P N
2619 2646 2673 2700 GTA CAA GGC TTA ACC GGC ACT ACG AAC ATC CTC AAT GCC TTT GCA AGA ACA CTC ACT ACT CAG GAG CAT GCC AAC AAG ATC GAT GAA TTC TCA AAC AAA TAC GCC AAC V Q G L T G T T N I L N A F A R T L T T Q E H A N K I D E F S N K Y A N
2727 GTG TTT ACG GCG GGA GAG ATG GCT TCA V F T A G E M A S
2754 2781 GTG GCG GCC ATC RAA GAG AAC ATC GCC GCT TCC ATA ACG TGG AAC TCT CAA AAC V A A i K E N l A A S I T W N S Q N
2a08 GCT GCT ACG GTC GAG GCC TGG CTG CGA A A T V E A W L R
2835 2862 2889 2916 AAG AAC TTC GGT ACT GAT GGA GCA AGC ACT GTC AGT GCC TCA ATT ACC ATT ATT AT? TCC GCT ATG GTC GCT ATT TAT AI\T ATT CTC taa gtc ata gat tat cgt tta K N F G T D G A S T V S A S I T I I I S A M V A I Y N I L
2943 2970 2997 3024 tit tct aag aat act tta gat gga aac tat tta ttt gaa cat ttg gtg CCt gat gag Cta Ctc ttt att ata aft tgt ttt aaa Lac aga tat tta agc tga aat cat
3051 3078 3105 atc aat att ttg aat caa aga act aga att aag ni t aaa aat ggt aaa gta aat taa gat ctt act taa aaa ata taa tat att ttt aat agt t
Fig. 3. Continued
Denolf et al. (EuI: J . Biochem. 248)
I1 I I
755
I? x,ylostellir Apnl
M. sexto Apnl
H. virrscens ApnI
I l l I
200 -
Fig. 4. Schematic representation of M. sexta Apn2 and I? xylostella Apnl compared with previously cloned CrylAc-binding aminopepti- dase N from M . sexta (M. sex& Apnl) and H. virescens (H. virescens Apnl). These aminopeptidase N isoforms are characterized by the pres- ence of an NH,-terminal hydrophobic domain representing a transport signal peptide and a COOH-terminal hydrophobic domain representing a signal peptide for glycosyl-Ptdlns-anchor addition (grey boxes). A, zinc- inetallopeptidase signature; $, potential N-linked glycosylation sites; 1, conserved cysteine residues.
1 2 kb - - 7.5 - 4.4 - 2.4 - 1.4 -
0.24 -
Fig. 5. Northern blot analysis of M. sexta apn2 and P. xylostella apnl expression. 1 .S pg mRNA isolated from the midgut of M. sextu (lane 1) and I? xylostellu (lane 2) were hybridized with a 296 bp-cDNA fragment from M . sexfu upn2. Molecular size markers are indicated at the left (7.5, 4.4, 2.4, 1.4, 0.24 kb).
taining an ORF of 2838 bp ( P xylostella apnl ; Fig. 3 b). The presence of the purine three bases upstream of the ATG codon at positions 58-60, together with the occurence of two stop codons upstream of and in frame with this codon indicate that it is the translation-start codon. At position 3058 a polyadenylation signal sequence has been identified. The ORF encodes a 946- amino-acid protein (I? xylostella Apnl) with a pl of 4.69 and a calculated molecular mass of 106688 Da. The molecular mass of the P xylostelln CrylAb5-binding protein as observed in a ligand blot assay was 120 kDa (Fig. 6A). The hydropathy plot of P. xylostella Apnl is very similar to that of M. sexta Apn2 (data not shown) and the CrylAcl-binding proteins from the midgut of M. sexta [51] and H. virescens [52]. The NH,-terminal signal peptide that directs the nascent protein to the endoplasmic reticulum has a predicted cleavage site between Gly15 and Asnl6 (score 12.67) 1761. The features of the COOH-terminal hydrophobic sequence of 23 amino acids are typical for a glyco- syl-Ptdlns anchor signal peptide: three sinall amino acids
116 -
97 -
66 -
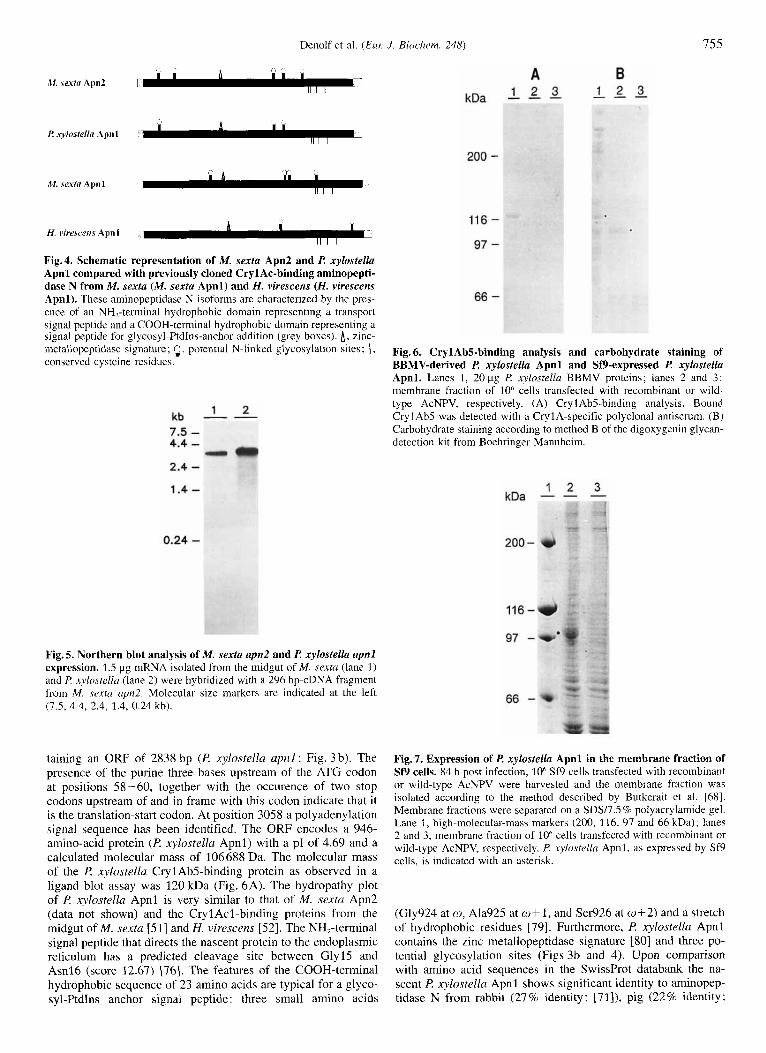
Fig. 6. Cryl Ab5-binding analysis and carbohydrate staining of BBMV-derived I? xylostella Apnl and Sf9-expressed I? xylostella Apnl. Lanes I , 20 pg l? xylostellu BBMV proteins; lanes 2 and 3: membrane fraction of 10" cells transfected with recombinant or wild- type AcNPV, respectively. (A) Cryl Ab5-binding analysis. Bound CrylAbS was detected with a CrylA-specific polyclonal antiserum. (B) Carbohydrate staining according to method B of the digoxygenin glycan- detection kit from Boehringer Mannheim.
1 2 3 kDa - - -
200 -
116 -
97 -
66 -
Fig. 7. Expression of I? xylostella Apnl in the membrane fraction of Sf9 cells. 84 h post infection, 10' Sf9 cells transfected with recombinant or wild-type AcNPV were harvested and the membrane fraction was isolated according to the method described by Butkerdit et al. [68]. Membrane fractions were separated on a SDY7.S % polyacrylamide gel. Lane 1, high-molecular-mass indrkers (200, 116, 97 and 66 kDa); lanes 2 and 3, membrane fraction of 10" cells trensfected with recombinant or wild-type AcNPV, respectively. l? xylostella Apnl, as expressed by Sf9 cells. is indicated with an asterisk.
(Gly924 at w, Ala925 at w + l , and Ser926 at m+2) and a stretch of hydrophobic residues [79]. Furthermore, I? xylostella Apnl contains the zinc metallopeptidase signature 1801 and three po- tential glycosylation sites (Figs 3b and 4). Upon comparison with amino acid sequences in the SwissProt databank the na- scent P xylostella Apnl shows significant identity to aminopep- tidase N from rabbit (27% identity; [71]), pig (22% identity;

756 Denolf et al. (Eur: J. Biochem. 248)
M-SEXTA-2 m S L I F L R L I - - - - G X A F G V P L S T N E D S T ~ Q N ~ - - - - - - - - - - - - - - - - . - - 3 2 P-XYLOSTEL MRLLICLTLL----GLVCGNPVQLTDNSIALQNTYD------------------- 3 2 M-SEXTA-1 FTI--FLGVRLL-QGVLTLSPIPVPE-----EEWRE---FS~----------LR 34 H-VIRESCEN M A A I K L L V L S L A C A C V I A H S ~ I P F A S R T I F L D E R L E G G A F ~ I D ~ ~ I E L S ~ 55
f .
M-SEXTA-2 --LYVLPQTSYFTFYDVRL--FIDffiYTE-------AFHGN 75 P-XYLOSTEL - - N Y V L P G E S F P T F Y D V Q L - - F F D P E Y D - - - - - - - S F N G E 7 5 M-SEXTA-1 D P S Y R L P m R P R H Y A ~ L T P Y F D W P A G V S G L ~ F S F ~ E ~ I Y I S P T Q ~ E 89 H-VIRESCEN ASPYRLE"VPTHYKIL---WIID~HQPVQ-----TYSGNWITLHATQAQWE 1 0 2
f * f . . . . . . . . . . . . . f t. , , I .
M SEXTA-2 STYLIAYLVSNFMYIENSQNPIY?IPfR-VYSRPGTQNTAE--FUEFGQQWIA 279 PIXYLOSTEL S S Y L L A Y L V S K F D Y I S N F X N P T Y D K S M K - V F S R P G M N T A E - E 2 8 3 M SEXTA-1 STYLLAFIVSHYQVISNN"A--RRPFR-IYRR"VGSQGD--WSLEMGEKLLLA 2 9 2 H ~ V I R E S C E N STYLLALIVSEYTSLPATNAA--GEILHEVIARPGAINNGQAVYAQRVGQ=~E 309
* _ - * . * . f * . . . . . . . . . . . * . . . . . . M-SEXTAz2 L E E Y T E F P Y A F P - - - - - K I D X A A V F D F A A G R M E N W G L V l S 3 2 9 P-XYLOSTEL L E K Y T E P P Y A F P - - - - - K I D K V A V P D F A A G A M W G L V L Y Q E G V W S 3 3 3 M SEXTA-1 M ~ A I F Y - Y T M A Q N L D M K Q A A T P D F S A G ~ ~ L L ~ R ~ I L Y D P L N S ~ 3 4 6 HIVIRESCEN M S D H ' I G F D F - Y A Q D P N L I A A I P D F G A G A M E N W G L L T Y S Y 363 . . . . . . . . . . . . . . . . *;** ****.*.*.,,.*. , f
M SFXTA-2 V K Q N I G R I I C H E N T H M W F G N E V G P M S W ~ G F A N P F E N Y A T D F V R P Q M 3 8 4 PIXYLOSTEL T L Q G I G R I I S H E N T H Q W F G N E V G F D S ~ E G F A 388 M-SFXTA-1 YRQRVANIVSHE1AHMWFGNLVTCAWWDNLWL"LGFARFSQl'YLTAWPEGYE 4 0 1 H-VIRESCEN FKQI1AYILSHEIAHMWFGNL"NA~VLWLNEGFARWQYFLTAWE-DLGLA 4 1 7 * , _ f _ , * * ,. * * * * f , ., *.*(I.*.,, , , * , *
M-SEXTA-2 DQFVIA-MQNVFFVRRCSKCQeHD-----APG--LYSFPDHRYFQRRRLPE~~fR 4 3 1 P-XYLOSTEL OQFVIN-KPNVFQSDAVLSPIT-FCIIRTPSQIffiTPNSVAY--QKSGSVIRH- 4 3 8 M-SEXTA-1 I R P I P E Q M V A M P S D S W S ~ ~ ~ P V A V s ~ F ~ I ~ - - ~ G ~ I L R H - 453 H-VIRESCEN T R P I N e O V H A S L L S D S S I Y A P L ~ ~ V G S P R A V S A K F S - - N K G A S I I R H - 4 6 9 . . . . . . . . . . . . . . . M-SEXTA-2 YSDVAAFHDTRDFQBRSGHWQSQLSRPAA-PSDLYVALQQALDESS----HP.I? 4 8 1 P - W S T E L - - - M Q H P L T P E I F R - K S L R L Y I S ~ S ~ ~ L F ~ I Q ~ ~ D - - - - H S I R 4 8 5 M-SEXTA-1 - - -TQHLLSYDTFV-KGLRQYLRARQPDVAEPYHLFSRLDAA 5 0 3 H-VIRESCEN - - - T E H L L G P D V H R - T G L R N Y L K D L A Y K T A Q P I D ~ F T A L B S G S D 5 2 0
M-SEXTA-2 K P I S T I ~ T E W S T Q G G F P V L T R T Z . P ~ ~ ~ S V W A Q E K Y L T D R S L T S - T D R ~ P V 535 P-XYLOSTEL W R L S I I M N P I W T Q Q O G F P W T V R R S A P S A Q S W I ~ R R F L T D S T Q E S - ~ P L 539 M-SEXTA-1 ITIDAYFR?WSEKAGHPLLSV?7mHESGR-MTLVQARWERNTGVSRFP'ZLWHIPI 5 5 7 H-VIRESCEN F D F V K W E S W T E Q P G H W L N V Q I N H Q X Q - M T T T Q R R F D I ~ H ~ Q ~ I I P I 5 7 4
. . . . . . . . . . . . . . . . . . . .
* . . * *..: . . . . . . . . . .
M-SEXTA-2 NWVISSNVNFSDTSPQAWILPTFPATAWVPGLSNiiDWYlFNKQQTGWRVNYDV 590 P-XYLOSTEL N W V L S T D V N F N D T R P M A W L P P Q ~ ~ V Q V ~ L Q N A E W F I D P 594 M-SEXTA-1 ~ G A P D F W L K F S Q ~ T - - - G O S L V L D R G T R G Q E W V I F N G F Y R ~ ~ D N 609 H-VIRESCEN T € T E G R N P S F D N T K P S H I I S - - - K G V ~ I D R G ~ ~ D ~ ~ E N I Q Q ~ N R ~ D D 6 2 6 . . . . . . . . . . . . . . . . . . . . . . . . . . . M-SEXTA-2 E N W V ~ V W N S H - E I I H V L N R A P l V D D A F N L J L R N G R L ~ ~ A F E I S R Y L ~ E K 6 4 4 P-XYLOSTEL E"DTH-EIIHLU4RAQLIDDSFNLRLDYSWDLSRYLVQER 648 M-SEXTA-1 ~ L I T R R L R S A N R ~ I H E L S R S Q I ~ D V F Q L R X S G V M L S Y L R F E D 6 6 4 H-VIRESCEN YTMWLIVLALRGADREKIHEYNRA?IWVFQFARS~~~Q~ILSFLEFET * * 6 8 1
. . . . . . . . . . ............... . . . . . M-SEXTA-2 DYIPWAAANPAFNYLDIVLS-GANSYNLYRYYLLNLTAPMFEDLGFDVKSGEEFV 698 P-XYLOSTEL DYIPWAAANAAFNYLNSVLS-GSSVHPLfQEYLLFQEYLLFLTAPLYQRUFNMmEEW 7 0 2 M-SEXTA-1 A Y A P W L S A I S G F N W V I R R F A K D W L Q T L Q N Q I I G L S E A 719 H-VIRESCEN EYAPWVAAI'lGFNWLRNRLV-GKPQLDELNEKIVQWSSKW4GELTYMPTEGEPFM 7 3 5
. * * * . f .:+.. . . . . . . . . . . .
M-SEXTA-2 RHSIAVSVINSSPEGMNVALDFWENFHRIQPRVQAL'lGTTNlLNTFmRL~SA 8 6 3 P-XYLOSTEL R H S V L V S A I N S G P D N M N A A L D N L E N F R N I Q P N V Q G L T G Q E 8 6 7 M-SEXTA-1 HSSALSSAITSNDVNTMRAFD~ILT)rNVDQI---TRTLGSITS?LNTITSRLLT~ 8 7 9 H-VIRESCEN Y T T A F N T A V S G M ~ L ~ I O R N L Q L V - - - F N A f A S P R T P L S Y I A A R L R T E 8 9 6
I . . . . . . . . . . . . . . .
M-SEXTA-2 ~ E K I D E L V R R K E - - S I F S A G E ~ S I M I R E N I X A S I A W S N S N A G I V ~ K ~ 916 P-XYLOSTEL H R N K T D E F S N X Y A - - N V F T A G E M I \ S V A i l I K ~ I A A S I ~ ~ S Q N ~ T D I ~ R ~ F 920 M-SEXTA.1 QMTQVQTWLDAN--RNTIGAAKWGWGIATSRR-NLQWSANRMSEFLRFFEn2F 931 H-VIRESCEN EWEYQ~TTAIQSALGT"AIYGDSVATYN-SILWVSTTEDSLSTYL?NGN 950
M-SEXTA-2 G - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - P P S G A L L V L I S L F V 9 3 7 p XYLOSTEL G......... ......................... T..ASTSASITIII .- 9 4 0 MISEXTA_l VDDVPSEATIVAPPhETTPSTF--PPTVAPATTPAPGSGNIAALSWSLLVTL 9 8 4 H-VIRESCEN D'JIEPSTS'ITSTTAAPT~II'-QPTITEPSTPTLPELTDS~TSFASLFIISLGaIL 1 0 0 5
M-SEXTA-2 AIFNH 9 4 2 ?-XYLOSTEL A I Y N I L 9 4 6 M-SEXTA-1 AINMVA 590 H-VIKESCW - - H L I - L 1009
Fig. 8. Sequence alignment of midgut aminopeptidase N isoforms from M . sextu (M-SEXTA-1 and M-SEXTA-2), H. virescens (H- VIRESCEN) and E! xylostellu (P-XYLOSTEL). Multiple-sequence alignment was performed according to the method of Higgins and Sharp 1991. *, perfectly conserved residues; *, well conserved residues.
[Sl]), human (20% identity; [72]), rat (20% identity; [70]) and to human aminopeptidase A (18% identity; [821). R xylostellu Apnl shows 61% amino acid identity to M. sexta Apn2 and is similar to the Cryl Acl -binding proteins from M. sextu (24 % identity; [51]) and H. virescens (24% identity; 1521).
Expression of M. sexta Apn2 and ti xylostella Apnl in Sf9 cells. The Sf9 expression of M. sextu Apn2 was unsuccessful. By performing a PCR with primers flanking the pVL1393 clon- ing site, we denionstrated that M. sexfa u p 2 was amplified dur- ing five consecutive virus amplification cycles. However, we observed no sign of infection, nor the protein of interest on a 7.5 % polyacrylamide gel. Moreover, Sf9 cells transfected with the pVL1393IM. sexta apnl construct did not display any aminopeptidase N activity. As a consequence, we did not further evaluate Cryl AbS-binding to transfected Sf9 cells.
In contrast, I? xylostella Apnl was expressed as a protein with an estimated size of 105 kDa (Figs 6 and 7). Using the assay described by Kunz et al. [69] it was demonstrated that Sf9 cells expressing l? xylostella Apnl displayed maximal amino- peptidase N activity 84 h post infection, in contrast to Sf9 cells transfected with the wild-type virus, which did not display any aminopeptidase N activity (data not shown). lncubation with 1 mM amastatin and 1 mM bestatin, reduced the aminopeptidase N activity displayed by Sf9(P. xylostellu Apnl) to 2.67 2 2 . 4 8 % and 11.37 t 3.7696, respectively, compared with the activity dis- played in the absence of these aminopeptidase inhibitors. Upon lysis of recombinant Sf9 cells R xylnstella Apnl was almost completely retained in the cellular debris, indicating that l? xy - lostella Apnl was expressed as a membrane-inserted or mem- brane-anchored protein (data not shown). When Sf9(P xjhstella Apnl) cells were treated with PtdIns-specific PLC only 38.65?3.890/0 of the aminopeptidase N activity was retained in the pellet fraction, showing that I? xylostellu Apnl is attached to Sf9 cells by means of a glycosyl-PtdIns anchor. Homo- logous-competition-binding experiments with 12sI-labeled or biotinylated CrylAb5, CrylAcl , CrylBa, CrylCax and (Alal64)Cry9Cal, and Sf9(l? xylostellu Apnl) membrane frac- tions or intact Sf9(R xylostella Apnl) cells did not reveal any specific ICP binding (data not shown). Using a CrylA-specific polyclonal antiserum to evaluate the binding of CrylAbS in a ligand blot did not demonstrate CryIAbS binding to Sf9(P xy- lostella Apnl) (Fig. 6A). Carbohydrate staining indicated that Sf9(l? xylostella Apnl) was glycosylated, apparently in a dif- ferent way compased to BBMV-derived R xylostelln Apnl (Fig. 6 B).
DISCUSSION In view of the important contribution of high-affinity binding
sites in the midgut of susceptible insect species to the specificity of B. thuringiensis ICP (1-25) much effort is being put into the cloning and characterization of ICP-binding proteins. The majority of ICP-binding proteins characterized belong to the aminopeptidase N family [ 49-55], including the 120-kDa Cryl AbS-binding protein purified from the brush-border mem- brane of M. sextu. Similarly to the cloned CrylAcl receptors from M. sextu (M. sextu Apnl) [Sl] and H. virescens [52] , the 120 kDa CrylAbS-binding protein from M. sexta is an amino- peptidase N (M. sextu Apn2) characterized by the presence of NH,-terminal and COOH-terminal hydrophobic amino acid se- quences (Fig. 3 a). The hydrophobic COOH-terminus is charac- teristic of a glycosyl-Ptdlns anchor, which was confirmed by treatment of BBMV with Ptdlns-specific PLC (Fig. 2). The M. sextu Apn2 amino acid sequence contains five putative N-glyco- sylation sites (Figs 3 a and 4). Glycosylation may partially ac-

Denolf et al. (Eur: J . Riocliern. 248) 757
count for the difference between the calculated 107.5-kDa and the observed 120-kDa molecular masses of M. sexta Apn2. The effect of N-glycosidase F treatment on BBMV indicates that the glycan structures are involved in the specific binding of CryIAbS in the M . sextu midgut (Fig. 2). Whilst monosaccha- rides do not inhibit the binding of CrylAbS to M. sextu BBMV, incubation of sodium-cholate-solubilized BBMV with the lectins concanavalin A and Ulex europaeus agglutinin I interfered with CrylAbS-binding (data not shown). From these results we con- clude that the glycan structure is of the fucosylated type and contains high-mannose oligosaccharides [84]. However, it is un- clear whether the glycan structure is directly or indirectly in- volved in CrylAbS binding either by representing part of the Cryl AbS-binding epitope or by maintaining the structure of the Cryl Ab5-binding protein.
From our ligand blotting with M . sextu BBMV and CrylAbS-affinity-purified BBMV proteins, a 120-kDa protein appears to be responsible for CrylAbS binding (Fig. 1). A 120- kDa protein from M. sextu BBMV displayed CrylAb-binding properties in ligand-blot assays performed by Martinez-Ramirez et al. [8S] with Cryl AbS, and by Chen et al. [46] and Rajamohan et al. [37] with CrylAb9-033. However, these teams also ob- served Cry 1 Ab binding to an approximately 210-kDa protein. Specificity of the CrylAb binding to the 120-kDa and 210-kDa proteins was indicated by competition with non-labeled Cry 1 Ab and the lack of binding of Cry3A 137, 46, 851, an ICP not toxic towards M . sextu. A 210-kDa protein was the only CrylAb- binding protein identified in a ligand-blot experiment by Vadla- mudi et al. [17]. This team purified CrylAbS directly from B. thuringiensis herliner, in contrast to the forementioned teams 137, 46, 851, which used E. coli-expressed ICP. Another team observed binding of E. coli-expressed Cry1 Ab [86] to 210-kDa and 250-kDa proteins from the midgut of M. sexta 1401. The interpretation of the results from the ligand blots is further com- plicated by the results from other types of binding assays. Per- forming binding experiments with BBMV, Van Rie et al. [3] concluded that Cry 1 AbS has only one binding-site population in the midgut epithelium of M. .sexta. Moreover, CrylAal and Cryl Ac 1 competed for the Cry1 AbS-binding site, whereas Cry 1 Aal recognized an additional binding site. From binding experiments using the surface-plasmon-resonance technique, Masson et al. concluded that CrylAa and CryIAb6 bind to a single site of the purified 120-kDa CrylAcl-binding protein from M. sexta, whereas binding by Cryl Acl best fits a two-site model 1251. When comparing BBMV-binding experiments with ligand blotting, Lee and Dean [87] reported difficulties in deter- mining ICP-binding-site relationships in L. dispar. The discrep- ancy between the results from different types of binding experi- ments may result from different experimental conditions. For example, the treatment of BBMV as a function of the binding experiment to be performed might influence ICP binding by changing the accessibility of the membrane proteins. This may be the case in ligand-blotting assays with denatured BBMV [17, 37, 40, 46, 8.5, this work] or in surface-plasmon-resonance ex- periments with sonicated, biotinylated BBMV [2S]. Further- more, ICP binding might change as a function of BBMV-storage conditions. This is suggested from a comparison of the results of binding experiments in which BBMV were used that were prepared according to the method described by Wolfersberger et al. 1611. In ligand blots where CrylAb was found to bind to a 210-kDa protein [37,46, 8.51, BBMV stored in 8 mM Na,HPO,, 2 mM KH,PO,, IS0 mM NaCl, pH7.4, were used, whereas binding to a 120-kDa protein (this work) was observed with BBMV stored in 150 mM mannitol, 2.5 mM EGTA, 8.5 mM Tris/HCl, pH 7.5. The BBMV used in vesicle-binding experi- ments [3] and surface-plasmon-resonance experiments [25] were
stored in the same buffer. Martinez-Ramirez et al. 1851 demon- strated that Cryl AbS binding shifted from the 210-kDa protein to the 120-kDa protein upon incubation of BBMV with increas- ing concentrations of EDTA. ECTA and EDTA share similar physicochemical characteristics and are able to remove periph- eral proteins from membranes or to disturb membrane integrity (881. An alternative explanation for the discrepancy between sets of binding data may be the observed existence of ICP-binding proteins tightly associated with other membrane proteins in de- tergent-solubilized BBMV [24]. The presence of multiple glyco- syl-PtdIns-anchored proteins in such a complex impedes bind- ing-protein purification to homogeneity by Ptdlns-specific-PLC treatment. It cannot be excluded that multiple proteins in the brush-border membrane cooperate to form a functional ICP- binding site. Perhaps high-affinity binding can only be obtained by cooperation of multiple binding proteins each displaying a low binding capacity. Binding experiments with the purified 120-kDa [SO] or 1 IS-kDa proteins from M. sextu 1251 indicated that this BBMV protein alone has a moderate ICP-binding affin- ity. While in ligand-blot assays multiple ICP-binding proteins would yield different bands [37, 40,46, 85, 871, they may appear as one high-affinity binding site in assays on native BBMV [3, 871. It has been demonstrated that rat, human and pig aminopep- tidase N occur as dimers [ 891.
Recently, a 106-kDa aminopeptidase N from the the midgut of M. sexta was identified as a CrylC-binding protein. A V8- protease-resistant 70-kDa peptide from this 106 kDa aminopep- tidase N was characterized by the NH,-terminus QNMIALE. This sequence is present i n the reported M. sextu Cryl AbS-bind- ing protein, starting from Q275 in M. sextu Apn2. Taking into account that M. sextu Apn2 is attached to a glycosyl-PtdIns an- chor via P919, the peptide starting at Q27S and ending at P919 has a calculted molecular mass of 70.95 kDa, which corresponds with the 70-kDa peptide reported by Luo et al. 1551. The 106- kDa CrylC-binding protein has an NH,-terminus that is nearly identical to the NH,-terminus of the 120-kDa Cry 1 Ac-binding protein from M. sextu [SO, 511. As a consequence, the 106 kDa CrylC-binding aminopeptidase N may represent a hybrid iso- form of the 120-kDa Cryl Ac-binding and Cryl AbS-binding pro- teins. Alternatively, the NH,-terminal and internal amino acid sequences reported by Luo et al. [SS] may have been derived from two proteins present in the preparation containing the Cryl C-binding protein.
Use of low-stringency cloning for the isolation of candidate ICP-binding proteins originated from our interest in characteriz- ing the CrylAbS-binding protein from the diamondback moth, f? xylostella. Some populations of the diamondback moth devel- oped resistance against the Cryl AbS b-endotoxin [9, 90-921. Performing a ligand blot we observed the binding of CrylAbS to a 120-kDa protein from the midgut brush-border of l? xylostella (Fig. 6A). The purification of a sufficient amount of this CrylAbS-binding protein from such a small insect as the dia- mondback moth would be very labor intensive. Therefore we used a DNA fragment from the gene encoding the M. sexta Cryl Ab5-binding aminopeptidase N in an effort to clone the Cryl AbS-binding protein from l? xylostellu. The aminopeptidase N isoform cloned from the midgut of CrylAbS-sensitive l? xy- lostella is similar to the CrylAbS-binding protein from M. sextu (Figs 3 and 4) and the Cryl Acl-binding aminopeptidase N from M. sextu [SI] and H. virescens [52] (Fig. 4). However, we do not have any evidence that the cloned f! xylostellu upnl gene encodes the 120-kDa Cryl Ab-binding protein from the midgut of l? xylostella.
In addition to their hydrophobic NH,-terminus and the pres- ence of the HisCluXaaXaaHis signature, which is unique to zinc metallopeptidases [SO], thcse isoforms contain the COOH-termi-

758 Denolf et al. ( E m J. Biochem. 248)
nal hydrophobic region that is characteristic of glycosyl-PtdIns- anchored proteins. Starting from the w site determined by Kodu- kula et al. [79], the glycosyl-PtdIns signal sequence contains 23 amino acids in M. sexta Apn2, I? xylostella Apnl (Fig. 3), and the CrylAcl-binding aminopeptidase N from M. sexta [SI] and H. virescens [S2]. Aminopeptidase N from the larval midgut of B. nzori is also coupled to the cell membranes by means of a glycosyl-PtdIns anchor [93, 941. Glycosyl-PtdIns linkage is a fairly common means of anchoring membrane proteins to mam- malian, yeast and parasitic cells. One of the functions of glyco- syl-PtdIns linkage is apparently to increase protein mobility in the membrane compared with proteins linked by a transmem- brane peptide sequence [95]. This feature may boost the contri- bution of insect aminopeptidase N to hydrolysis of partially di- gested food proteins which takes place within the ectoperitrophic space and at the insect midgut microvillar surface [96]. Differ- ences distinguishing the glycosyl-PtdIns-anchor structures reside in branching substituents at various positions of the glycan core [97]. Since it has been demonstrated that carbohydrates may contribute to ICP binding 111, 12, 141, the glycosyl-PtdIns an- chor may partially account for ICP-binding-site heterogeneity. Moreover, it has been suggested that insects can develop ICP resistance through the elimination of binding sites due to increased levels of endogenous PtdIns-specific PLC [52, 981.
In the CrylAcl-binding proteins from M. sextu and H. virescens the glycosyl-PtdIns signal sequence is preceded by a Ser/Thr-rich region which may provide 0-linked glycosylated residues. M. sexfa Apn2 and I? xylostelta Apnl lack such a po- tential 0-glycosylated stalk. However, they contain consensus N-glycosylation sites. The three potential N-glycosylation sites of I? xylostella Apnl are located at almost identical positions compared with the potential sites in M. sexta Apn2 (Figs 3 and 4). Whether covalently attached carbohydrates contribute to ICP binding in the diamondback moth remains to be determined.
A multiple-sequence alignment according to the method of Higgins and Sharp [991 indicated that the Cry1 Ab5-binding aminopeptidase N from M. s e m i and its isoform from P x.ylos- tella, together with the Cryl Acl-binding proteins from M. sexta and H. viresceizs display 15 % and 45 % amino acid identity and similarity, respectively (Fig. 8). The regions of highest amino acid identity of these four insect aminopeptidases are located in blocks that are conserved among aminopeptidase N molecules from different organisms [loo]. Furthermore, four of the Cys residues present in these aminopeptidase N isoforms occupy a conserved position within a 71 -73-amino-acid stretch starting 239-209 residues before the glycosyl-PtdIns-anchor addition site (Figs 4 and 8). This feature may have implications for the overall protein structure by the formation of disulfide bonds. The four conserved Cys residues are also present in human, rab- bit, rat and pig aminopeptidase N. The multiple-sequence align- ment revealed the presence of four conserved peptides in the M. sexta, P xylostella and H. virescens aminopeptidase N isoforms: A172FPCYDEP (172 refers to the position of the residue in M. sexta Apn2) ; A303GAMENWGL; W360LNEGFA and YsMRV- NYD. From these amino acid sequences degenerate oligonucleo- tides may be designed which can be used for further screening of aminopeptidase N isoforms from the midgut epithelium of insect larvae.
Since P xylostella Apnl, as obtained from the expression with the BaculoGold/Sf9 system, was not able to bind any of the tested B. thuringiensis ICP with high affinity, we do not know whether this aminopeptidase N isoform represents a func- tional ICP-binding protein. From our demonstration that the in vitro-expressed l? xylostella Apnl displays aminopeptidase N activity we know that the structure of the protein, as expressed on the surface of Sf9 cells, resembles the in vivo conformation.
On the other hand, the molecular mass of I? xylostella Apnl as expressed by Sf9 cells was estimated to be about 105 kDa, com- pared with the BBMV-derived I? xvlostella Apnl which mi- grates as a ‘120 kDa-protein upon SDSPAGE. Although Sf9/I? xylostellu Apnl is glycosylated to a certain extent (Fig. 6B), it is possible that differences in the co- or post-translational modi- fications hinder ICP binding by I? xylostella Apnl . In some in- stances only a minor percentage of the proteins expressed with the baculovirus-insect cell system are active, functional or prop- erly processed [101-106]. For example, it has been observed that insect cells, in contrast to mammalian cells, do not add ter- minal sugar residues after trimming high-mannose structures [104]. Another possibility explaining the lack of high-affinity ICP binding to the in vitro-expressed P xylostella Apnl may be the need for binding-protein oligomerization to realize high- affinity binding.
In this report we described the cloning and characterization of a 120-kDa CrylAb5-binding protein from the midgut of M. sexta. This protein represents an aminopeptidase N isoform with very similar structural features to the Cryl Ac-binding proteins from M. sexta and H. virescens. We believe that the 120-kDa Cryl Ab5-binding protein, M. sextu Apn2, represents (part of) a functional ICP receptor from the midgut brush-border membrane of M. sexta. Using the genetic information of the M. sexta CrylAb5-binding protein we were able to clone and characterize an aminopeptidase N isoform from the midgut of I? xylnstella. The identification of P xylostella Apnl as a B. thuringiensis ICP-binding protein will require optimization of the baculovirus/ insect cell system used or expession with other proteins involved in the functional receptor complex. Moreover, we need to evalu- ate the possibility that other aminopeptidase N isoforms from the midgut of I? xylostell~i represent functional ICP-binding pro- teins.
We gratefully acknowledge Dr R. Devos, in whose laboratory the baculovirus work was conducted. We thank I. FachC, Y. Guisez, J. Taver- nier, J. Van Der Heyden and A. Verhee for helpful suggestions through- out the baculovims work, and J. Seurinck for sequence analysis assis- tance. We thank L. Buysse, C. Piens and B. Saey for insect rearing. We acknowledge K. Spruyt and R. Verbanck for the photographic assistance. This work was supported by research grant IWT/OZM/960021 from the Vluurns Instituut voor de bevordering van het Wetenschuppelijk-Techno- logisch onderzock in de ind~rrrie (IWT).
REFERENCES 1. Hofmann. C., Luthy, P., Hutter, R. & Pliska, V. (1988) Binding
of the K-endotoxin from B~icillus thuringiensis to brush-border membrane vesicles of the cabbage butterfly (Pieris hrmsictre), EUK J. Biochern. 173, 85-91.
2. Hofmann, C., Vanderbruggen, H., Hofte, H., Van Rie, J., Jansens, S. & Van Mellaert, H. (1988) Specificity of Bucillus thnrin- giensis d-endotoxins is correlated with the presence of high-af- finity binding sites in the brush-border membrane of target insect midguts, Proc. Nut/ Acud. Sci. USA 85, 7844-7848.
3. Van Rie, J., Jansens, S., Hofte, H., Degheele, D. & Van Mellaert, H. (1989) Specificity of Bricilllis thuringierzsi.s d-endotoxins, Eur: J. Biochem. 186, 239-247.
4. Van Rie, J., Jansens, S., Hofte, H., Degheele, D. & Van Mellaert, H. (1990) Receptors on the brush-border membrane of the insect midgut as determinants of the specificity of Bucillus fhurin- giensis d-endotoxins. Appl. Env. Microbiol. 56, 1378- 1385.
5. Denolf, P., Jansens, S.. Peferoen, M., Degheele, D. & Van Rie, J . (1993) Two different Bacillus thuringiensis d-endotoxin recep- tors in the midgut brush-border membrane of the European corn borer, 0.strinia nubilalis (Hubner) (Lepidoptera: Pyralidae),
6. Escriche, B., Martinez-Ramirez, A. C., Real, M. D., Silva, F. J. & Ferre J. (1994) Occurence of three different binding sites for
Appl. EIIV. Microbial. 59, 1828- 1837.

Denolf et al. (ELK J . Bioclzem. 248) 759
Bacillus fhuringiensis J-endotoxins in the midgut brush-border membrane of the potato tuber moth Phthorimaeu operculella (Zeller), Arch. Insert Biochem. Physiol. 26, 315-327.
7. Estada, U. & FerrC, J. (1994) Binding of insecticidal crystal pro- teins of Bacillus thuringiensis to the midgut brush-border of the cabbage looper, Trichoplusia ni (Hubner) (Lepidoptera: Noctui- dae), and selection for resistance to one of the crystal proteins, Appl. Em]. Microbiol. 60, 3840- 3846.
8. Sanchis, V. Chaufaux, J. & Pauron, D. (1994) A comparison and analysis of the toxicity and receptor binding properties of Buci1- lus thuringiensis CryIC S-endotoxin 011 Spodopreru lirrorulis and Bomhyx mori, FEBS Lett. 353, 259-263.
9. Tang, J. D., Shelton, A. M., Van Rie, J., De Roeck, S., Moar, W. J., Roush, R. T. & Peferoen, M. (1996) Toxicity of Bacillus thu- ringiensis spore and crystal protein to resistant diamondback moth (Plutrllu xylosfella), Appl. Environm. Microbiol. 62, 564- 569.
10. Lambert, B., Buysse, L., Decock, C., Jansens, S., Piens, C., Saey, B., Seurinck, J., Van Audenhove, K., Van Rie, J., Van Vliet, A. & Peferoen, M. (1996) A Bacillus thuringiensis insecticidal crystal protein with a high activity against members of the fan- ily Noctuidae, Appl. Env. Microbiol. 62, 80-86.
11. Knowles, B., Knight, P. & Ellar, D. (1991) N-acetyl galactosainine is part of the receptor in insect gut epithelia that recognizes an insecticidal protein from Bacillus thuringiensis, Proc. R. Sor. Lond. B 245, 31-35.
12. Garczynski, S., Crim, J. & Adang, M. (1991) Identification of pnta- tive insect brush-border membrane-binding molecules specific to Bacillus thuringiensis S-endotoxin by protein blot analysis, Appl. Env. Microhiol. 57, 2816-2820.
13. Oddou, P., Hartmann, H. & Geiser, M. (1991) Identification and charac.terization of Heliothis virescens midgut membrane pro- teins binding Bacillus thuringiensis Gendotoxins, Eur: J. Bin- chein. 202, 673-680.
14. Indrasith, L. & Hori, H. (1992) Isolation and partial characteriza- tion of binding proteins for immobilized delta endotoxin from solubilized brush-border membrane vesicles of the silkworm, Bombyx mori, and the common cutworm, Spodopteru lifura, Comp. Biochenz. Physiol. 102B, 605-610.
15. Oddon, P., Hartmann, H. & Geiser, M. (1993) Iminunologically unrelated Heliothis sp. and Spodoptera sp. midgut membrane- proteins bind Bacillus thuringiensis CryIA(b) S-endotoxin, Eur: J. Biochem. 212, 145-150.
16. Sanchis, V. & Ellar, D. (1993) Identification and partial purification of a Bacillus thuringiensis CryIC S-endotoxin binding protein from Spodoptera littoralis gut membranes, FEBS Lett. 316, 264-268.
17. Vadlamudi, R., Ji, T. & Bulla, L. (1993) A specific binding protein from Munducu sexta for the insecticidal toxin of Bacillus tlzurin- giensis subsp. herliner, J. B id . Chem. 268, 12 334- 12 340.
18. Feldman, F., Dullemans, A. & Waalwijk, C. (1995) Binding of the CryIVD toxin of Bacillus thuringiensis subsp. to larval dipteran midgut proteins, Appl. Env. Microbiol. 61, 2601 -2605.
19. Cowles, E., Yunovitz, H., Charles, J.-F. & Gill, S. (1995) Compari- son of toxin overlay and solid-phase binding assays to identify diverse CrylA(c) toxin-binding proteins in Heliothis virescens midgut, Appl. Eiw. Microbiol. 61, 2738-2744.
20. Bravo, A., Hendrickx, K., Jansens, S. & Peferoen, M. (1992) Im- munohistochemical analysis of specific binding of Bacillus thu- ringiensis insecticidal crystal proteins to lepidopteran and col- eopteran midgut membranes, J . Invertehr: Pathol. 60, 247 -253.
21. Denolf, P., Jansens, S., Van Houdt, S., Peferoen, M., Degheele, D. & Van Rie, J. (1993) Biotinylation of Bacillus rhuringiensis insecticidal crystal proteins, Appl. Env. Microbiol. 59, 1821 - 1827.
22. Ravoahangimalala, 0 . & Charles, J.-F. (1995) In vitro binding of Bacillus thuringiensis var. individual toxins to midgut cells of Anopheles gambiue larvae [Diptera: Culicidae], FEBS Lett. 362, 11 1-1 15.
23. Masson, L., Mazza, A. & Brousseau, R. (1994) Stable immobiliza- tion of lipid vesicles for kinetic studies using surface plasrnon resonance, Anal. Biochem. 218,405-412.
24. Masson, L., Mazza, A., Brousseau, R. & Tabashnik, B. (1995) Ki- netics of Bacillus thuringiensis toxin binding with brush-border
mcinbrane vesicles from susceptible and resistant larvae of Phi- tellu xylostella, J . B id . Chem. 270, l l S87- 1 1 896.
25. Masson, L., Lu, Y.-J., Mazza, A., Brousseau, R. & Adang, M. (1995) The CryIA(c) receptor purified from Manduca sexto dis- plays multiple specificities, J. B id . Chem. 270, 20309 -20315.
26. Roush, R. (1994) Managing pests and their resistance to Bacillus thuringiensis: can transgenic crops be better than sprays'? Rio- control Sci. Technol. 4, 501 -5 16.
27. Wolfersberger, M., Hofinann, C. & Luthy, P. (1986) Interaction of Bacillus thuringiensis S-endotoxin with membrane vesicles isolated from lepidopteran larval midgut, in Bacterial Protein Toxins (Falmagne, P., Alouf, J.: Fehrenbach, F., Jeljaszewicz, J. & Thelestam, M., eds) pp, 237-238, Gustav Fisher Verlag, Stuttgart, Germany.
28. Ihara, H., Kuroda, E., Wadano, A. & Himeno, M. (1993) Specific toxicity of d-endotoxins from Bacillus thuringiensis to Bornbyx mori, Biosci. Biotechnol. Biochem. 57, 200-204.
29. Liang, Y., Patel, S. & Dean, D. (1995) Irreversible binding kinetics of Bacillus thuringiensis CryIA S-endotoxins to Gypsy moth brush-border membrane vesicles is directly correlated to toxicity, J. B id . Chem. 270, 24719-24724.
30. Schnepf, H., Tomczak, K., Ortega, J. & Whiteley, H. (1990) Speci- ficity-determining regions of a lepidopteran-specific insecticidal protein produced by Bacillus thuringiensis, J. B id . Chem. 265,
31. HonCe, G., Convents, D., Van Rie, J., Jansens, S., Peferoen, M. & Visser, B. (1991) The C-terminal domain of the toxic fragment of a Bacillus thuringiensis crystal protein determines receptor binding, Mol. Microbiol. 5, 2799-2806.
32. Ge, A,, Rivers, D., Milne, R. & Dean, D. (1991) Functional do- mains of Bacillus thuringiensis insecticidal crystal proteins, J. Rial. Chenz. 266, 17954-17958.
33. Lee, M., Milne, R., Ge, A. & Dean, D. (1992) Location of a B o m byx mori receptor binding region on a Bacillus thurirzgiensis d- endotoxin, J. Biol. Chem. 267, 311553121,
34. Wu, D. & Aronson, A. (1992) Localized mutagenesis defines re- gions of the Bacillus thuringiensis 6-endotoxin involved in toxicity and specificity, J. B id . Chem. 267, 2311 -2317.
35. Cummings, C. & Ellar, D. (1994) Chemical modification of Bacil- lus fhurirzgiensis activated S-endotoxin and its effect on toxicity and binding to Manduca sexta midgut membranes, Microbiol- og)>, 140, 2737-2747.
36. Lu, H., Rajamohan, F. & Dean, D. (1994) Identification of amino acid residues of Bacillus thuringiensis S-endotoxin CryIAa asso- ciated with membrane binding and toxicity to Bombyx mori, J . Bacteriol. 176, 5554-5559.
37. Rajamohan, F., Alcantara, E., Lee, M., Chen, X., Curtiss, A. & Dean, D. (1995) Single amino acid changes in domain I1 of Bu- cillus thuringiensis CrylAb S-endotoxin affect irreversible bind- ing to Manduca sexta midgut membrane vesicles, J. Bacteriol.
38. Aronson, A,, Wu, D. & Zhang, C. (1995) Mutagenesis of specific- ity and toxicity regions of a Bacillus rhuringiensis protoxin gene, J . Bacteriol. 177, 4059-4065.
39. De Maagd, R., Kwa, M., Van der Klei, H., Yamainoto, T., Schipper, B., Vlak, J., Stiekema, W. & Bosch, D. (1996) Domain 111 suhsti- tution in Bacillus rhuringiensis &endotoxin CrylA(b) results in superior toxicity for Spodoptera exigua and altered membrane protein recognition, Appl. Env. Microhiol. 62, 1537 - 1543.
40. De Maagd, R., Van der Klei, H., Bakker, P., Stiekema, W. & Bosch, D. (1996) Different domains of Bacillus rhuringiensis S-endotox- ins can bind to insect midgut membrane proteins on ligand blots, Appl. Env. Microbiol. 62, 2753-2757.
41. Gazit, E. & Shai, Y. (1993) Structural and functional characteriza- tion of the a5 segment of Bacillus thuringiensis &endotoxin, Biochemistry 32, 3429 -3436.
42. Gazit, E., Bach, D., Kerr, I., Sansom, M., Chejanovski, N. & Shai, Y. (1994) The n5 segment of Bacillus thuringiensis S-endotoxin: in vitro activity, ion channel formation and molecular modelling, Biochem. J. 304, 895-902.
43. Gazit, E. & Shai, Y. (1995) The assembly and organization of the a5 and a7 helices from the pore-forming domain of Bacillus thuringiensis d-endotoxin, J. Biol. Chem. 270, 2571 -2578.
20923 -20930.
177, 2276-2282.

760 Denolf et al. (Eur J. Biochem. 248)
44. Pang, A. (1993) Use of synthetic peptides to probe functional do- mains of il BaciNus tl?z/rif/~ier~.s~.s toxin. J. Inverfeht: Parhoi. 61. 260 - 266.
45. Cummings, C., Armstrong, G., Hodgman, T. & Ellar, D. (1994) Structural and functional studies of a synthetic peptide mimick- ing a proposed membrane inserting region of a Bacillus thurin- gicwis 8-endotoxin, Mol. Memhr Bid . 11, 87-92.
46. Chen, X., Curtis. A,, Alcantara, E. & Dean, D. (1995) Mutations in domain I of Bacillus thuringiensis S-endotoxin CryIAb reduce the irreversible binding of toxin to Manduca sexfa brush-border membrane vesicles, J . Biol. Chem. 270, 6412-6419.
47. Wolfersberger, M., Chen, X. & Dean, D. (1996) Site-directed muta- tions in the third domain of Bacillus thuringiensis rkndotoxin CryIAa affect its ability to increase the permeability of Boml?yx mori midgut brush-border membrane vesicles. Appl. Env. Micro-
48. Vadlamudi, R., Weber, E., Ji, I., Ji, T. & Bulla, L. (1995) Cloning and expression of a receptor for an insecticidal toxin of Bacillus tlzuritzgien.si.s, J , H i d . Cliern. 270, 5490-5494.
49. Knight, P., Crickmore, N. & Ellar, D. J. (1994) The receptor for Buc,il/us fhuringiensis CryIA(c) &endotoxin in the brush-border membrane of the lepidopteran Munduca sexfa is aminopeptidase N, Mol. Microhiol. 11, 429-436.
50. Sangadala, S., Walters, F., English, L. & Adang, M. (1994) A mix- ture of Muiiducrr sexfa aminopeptidase and phosphatase en- hances Bacillus thuringiensis insecticidal CryIA(c) toxin binding and ‘“Rb+-K’ efflux in vitro, J . Biol. Chem. 269, 10088-10092.
51. Knight, P., Knowles, B. & Ellar, D. (1995) Molecular cloning of an insect aminopeptidase N that serves as a receptor for l3ucillu.s thuringien.r.is CryIA(c) toxin, J . Biol. Chern. 270, 17 765 - 17 770.
52. Gill, S., Cowles, E. & Francis, V. (1995) Identification. isolation, and cloning of a Bacillus t11uringiensi.s CryIAc toxin-binding protein from the midgut of the lepidopteran insect Heliothis viresceris, J. Biol. Clwm. 270, 27 277-21 282.
53. Valaitis, A, , Lee, M.-K., Rajamohan, F. & Dean, D. (1995) Brush- border membrane aminopeptidase-N in the midgut of the gypsy moth serves as the receptor for the CryIA[c] d-endotoxin of Ba- cillus thiiringiensis, Insect Biocheni. Molec. B i d . 25. 1143 - 1151.
54. Lee. M., You, T., Young, B., Cotrill, J., Valaitis, A. & Dean, D. (1996) Aminopeptidase N purified from gypsy moth brush-bor- der membrane vesicles is a specific receptor for Bacillus thuriri- girnsi.5 Cry 1Ac toxin, Apyl. Ent! Microhiol. 62, 2845-2849.
55. Luo, K., Lu, Y.-J. & Adang, M. (1996) A 106 kDa form of amino- peptidase is a receptor for Bacillus thuringiensis CrylC &endo- toxin in the brush-border membrane of Munduca . s e y t a , Irzsec,f Biochem. Molec. B i d 26, 783-79 1.
56. Hiifte, H., De Greve, H.. Seurinck, J., Jansens, S., Mahillon. J.. Ampe, C., Vandekerckhove, I., Vanderbruggen, H., Van Mon- tagu, M., Zabeau, M. & Vaeck, M. (1986) Structural and func- tional analysis of a cloned d-endoroxin of Bacillus rhnringierrsis berliner 1715, Eiir J. Biochem. 161, 273-280.
57. Brizzard. B. & Whiteley, H. (1988) Nucleotide sequence of an ad- ditional crystal protein gene cloned from Bacillus tl~iiringiensi.~ subsp. fhuringicvzsis, Nucleic Acids Re.s. 16, 41 68-4169.
58. Kronstad. J. & Whiteley, H. (1986) Three classcs of homologous Bacillus thuringiensi.s crystal protein genes, Gene 43, 29-40.
59. Hunter, W. & Greenwood. F. (1962) Preparation of iodine-131 la- belled human growth hormone of high specific activity, Science 194% 495-496.
60. Fraker, P. & Speck, J. (1978) Protein and cell membrane iodina- tions with il sparingly soluble chloroamide, 1,3,4,6-tetrachloro- 3a,6a-diphenylglycoluril, Biochem. Biophys. Res. Cornmiin. 80, 849- 857.
61. Wolfersberger, M., Luthy. P., Maurer, A , , Parenti. P., Sacchi, V., Giordana, B. & Hanozet, G. (1987) Preparation and partial char- acterization of amino acid transporting brush-border membrane vesicles from the larval midgut of the cabbage butterfly [Pieri.s hrassicae], Conzp. Riochem. Physiol. 86, 30 1 -308.
62. Bradford, M. M. (1976) A rapid and sensitive method for the quan- titation of microgram quantities of protein utilizing the principle of protein-dye binding, Anal. Riochem. 72. 248 - 254.
biol. 62, 279-282.
63. Laemmli, 1J. K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4, Nature 227, 680- 685.
64. Towbin, H., Staehelin, T. & Gordon, J. (1979) Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications, Proc. Nut/ AM^. Sci.
65. Bauw, G., Van Damme, J., Puype, M., Vandekerckhove, J., Gesser, B., Ratz, G., Lauridsen, J. & Celis, J. (1989) Protein-electroblot- ting and -microsequencing strategies in generating protein data bases from two-dimensional gels, Proc. Nafl Acad. Sci. USA 86,
66. Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989) Molecular clon- ing: a laboraror-j manual, 2nd edn, Cold Spring Harbor Labora- tory, Cold Spring Harbor, NY.
67. Beckers, T., Schmidt, P. & Hilgard, P. (1994) Highly sensitive northern hybridization of rare mRNA using a positively charged nylon membrane, Biotechiques 16, 1075- 1078.
68. Rutkerait, P., Zheng, Y., Hallak, H., Graham, T., Miller, H., Burris, K., Molinoff, P. & Manning, D. (1995) Expression of the human 5-hydroxytryptamine,. receptor in Sf9 cells, 1. Bid. Clzen7. 270,
69. Kunz, D., Biihling, F., Hutter, H.-J., Aoyagi, T. & Ansorge, S. (1993) Aminopeptidase N (CD13, EC 3.4.11.2) occurs on the surface of resting and concanavalin A-stimulated lymphocytes, Biol. Clzenr. Hoppe-Seyler 374, 29 1-296.
70. Watt, V. & Yip, C. (1989) Amino acid sequence deduced from a rat kidney cDNA suggests it encodes the Zn-peptidase amino- peptidase N, J . Biol. Chem. 264, 5480-5487.
71. Noren, 0.. Dabelsteen, E., Hoyer, P., Olsen, J., Sjiistrom, H. & Hansen, G. (1989) Onset of transcription of the aminopeptidase N (leukemia antigen CD 13) gene at the crypthillus transition zone during rabbit enterocyte differentiation, FEBS Left. 259,
72. Olsen, J., Cowell, G., Konigshofer, E., Danielsen, E., Moller, J., Lausten, L., Hansen, O., Welinder, K., Engberg, J., Hunziker, W., Spiess, M., Sjostrom. H. & Noren, 0. (1988) Complete amino acid sequence of human intestinal aminopeptidase N as deduced from cloned cDNA, FEBS Left. 238, 307-314.
73. Wu, Q., Lahti, J., Air, G., Burrows, P. & Cooper, M. (1990) Molec- ular cloning of the murine BP-l/6C3 antigen: a member of the Linc-dependent metallopeptidase Family. Proc. Nut/ Acad. Sci.
74. Kozak, M. (1987) An analysis of5’-noncoding sequences from 699 vertebrate messenger RNAs. Nucleic Acid.s Rex 15. 8 125-8148.
75. Kyte, J. & Doolittle, R. (1982) A simple method for displaying the hydropathic character of a protein, J . Mol. B i d . 157, 105-132.
76. von Heijne, G. (1986) A new method for predicting signal se- quence cleavage sites, Nucleic Ac,ids Rex 14, 4683-4690.
77. Fletcher, T., Alhadeff, M., Craik, C. & Largman, C. (1987) Isola- tion and characterization of a cDNA encoding rat cationic tryp- sinogen, Biochemi.stry 26, 3081 -3086.
78. Levy, N., Malipiero, U., Lebecque, S. & Gearhart, P. (1989) Early onset ol somatic mutation in irnniunoglobulin VH genes during the primary immune response, J. E q . Med. 169, 2007-2019.
79. Kodukula, K., Gerber, L., Amthauer, R.. Brink, L. & Udenfriend, S. (1993) Biosynthesis of glycosylphosphatidylinositol (glyco- syl-Ptd1ns)-anchored membrane proteins in intact cells : specific amino acid requirements adjacent to the site of cleavage and glycosyl-PtdIns attachment, J . Cell B i d . 120, 657 - 664.
80. Jongeneel, C., Bouvier, J. & Bairoch, A. (1989) A unique signature identifies a family of zinc-dependent metallopeptidases, FEBS
81. Delmas, B., Gelfi, J., Kut, E., SjiistrGm, H., Noren, 0. & Laude, H. (1994) Determinants essential for the transmissible gastroen- teritis virus-receptor interaction reside within a domain of aminopeptidase-N that is distinct from the enzymatic site, J. Virol. 68, 5216-5224.
82. Nanus, D., Engelstein, D.. Gastl, G., Cluck, L., Vidal. M., Mor- rison, M., Finstad, C., Bander, N. & Albino, A. (1993) Molecular cloning of the human kidney differentiation antigen gpl6O: hu- man aininopeptidase A, Proc. Narl Acad. Sci. USA 90, 7069- 7073.
USA, 76. 4350-4354.
7701 -7705.
18 691 - 18 699.
107- 11 2.
USA, 87, 993-997.
Lett 242, 21 1-214.

Denolr et al. (Eur: J . Biochem. 248) 761
83. Myers, E. & Miller, W. (1988) Optimal alignments in linear space, Compiit. Appl. Bio.yci. 4. 11 - 17.
84. Hendrickx, K. (1992) On the mode of uction of the CryIAb delta- endoroxin ,from the bacterium Bacillus thuringiensis in the mid- gut of Manduca sexta [Insecta: Lepidoptera], PhD thesis, Katho- lieke Universiteit Leuven, Belgium.
85. Martinez-Ramil-cz, A. C., Gonzales-Nebauer, S., Escriche, B. & Real, M. D. (1994) Ligand blot identification of a Munduca sextu midgut binding protein specific to three Bucillus rhurirz- giensis CryIA-type ICPs, Biochem. Biophys. Rex Cornmun. 201. 782-787.
86. Bosch, B., Visser, B. & Stiekema, W. (1994) Analysis of non- active engineered Bacillus thuringiensis crystal proteins, FEMS Microbiol. Lett. 118, 129- 134.
87. Lee, M. & Dean, D. (1996) Inconcistencies in determining Bucillus thuringierisis toxin binding sites relationship by comparing com- petition assays with ligand blotting, Biochem. Biophys. Res. Commm. 220, 575-580.
88. Harris, E. & Angal, S. (1990) Prnreirz pttrificution applications: n practical approach, Oxford University Press, Oxford.
89. Semenza, G. (1986) Anchoring and biosynthesis of stalked brush- border membrane proteins : glycosidases and peptidases of en- terocytes and renal tubuli, Annu. Rev. Cell. Bid. 2, 255-313.
90. Kirsch, K. & Schmutterer, H. (1988) Low efficacy of a Bacillus thuringiensis (Berl.) formulation in controlling the diamondback moth, Plutellu xylovtella (L.), in the Philippines, J . Appl. Ent.
91. Tabashnik, B., Cushing, N., Finson, N. &Johnson, M. (1990) Field development of resistance to Bacillus thuringiensis in diamond- back moth [Lepidoptera: Plutellidae], J . Econ. Entornol. 83,
92. Shelton, A,, Robertson, J., Tang, J., Perez, C., Eigenbrode, S., Preisler, H., Wilsey, W. & Cooley, R. (1993) Resistance of dia- mondback moth to Bacillus thuringiensis subspecies in the field, J. Econ. Entornol. 86, 697-105.
93. Takesue, S., Yokota, K., Miyajima, S., Taguchi, R., Ikezawa, H. & Takesue, Y. (1992) Partial release of aminopeptidase N from lar- val midgut cell membranes of the silkworm, Bombyx mori, by phosphatidylinositol-specific phospholipase C, Comp. Biochem.
94. Tomita, M., Obara, H., Takesue, Y., Tamura, H.-O., Miyajima, S., TdguchI, R. & Ikezawa, H. (1994) Purification of glycosylphos- phatidylinositol-anchoring aminopeptidase N from the plasma membrane of larval midgut epithelial cells of the silkworm, Bombyx mori, Int. J . Biochem. 26, 977-986.
105, 219-255.
1671 - 1676.
Ph~siol. 102B, 7 - 1 1.
95. Udenfriend, S. & Kodukula, K. (1995) How glycosylphosphatidyl- inositol-anchored membrane proteins are made, Annu. Rev. Bio- chem. 64, 563-591.
96. Lee, M. & Anstee, J . (1995) Characterization of midgut exopepti- dase activity from larval Spodopteru littoralis, Insect Biochem. Molec. Biol. 25, 63 -71.
97. Takeda, J. & Kinoshita, T. (1995) Glycosyl-Ptdlns-anchor biosyn- thesis, Trends Biochem. Sci. 20, 367-371.
98. Lu, Y.-J. & Adang, M. (1996) Conversion of Bacillus thiirii7gien.c.i.c CryIAc-binding aminopeptidase to a soluble form by endoge- nous phosphatidylinositol phospholipase C, Iii~ect Binchcm. Mol. Biol. 26, 33-40.
99. Higgins, D. & Sharp, P. (1988) CLUSTAL: a package for perform- ing multiple sequence alignment on a microcomputer, Gene (Amst.) 73, 237-244.
100. Tan, P., van Alen-Boerrigter I . , Poolman, B., Siezen, R., de Vos, W. & Konings, W. (1992) Characterization of the Lucror.orcu.r lacti,s pepN gene encoding aminopeptidase homologous to mam- malian amiopeptidase N, FEBS Lett. 306, 9- 16.
101. Germann, U., Willinghain, M., Pastan, I . & Gottesmdll, M. (1990) Expression of the human multidrug transporter in insect cells by a recombinant baculovirus, Biochemistry 29, 2295 -2303.
102. Kartner, N., Hanrahan, J., Jensen, T., Naismith, A,, Sun, S., Acker- ley, C., Reyes, E., Tsui, L.-C., Rommens, J., Bear, C. & Riordan, J. (1991) Expression of the cystic fibrosis gene in non-epithelial invertebrate cells produces a regulated anion conductance, Cell
103. Fountoulakis, M., Schlaeger, E.-J., Gentz, R., Juranville, J.-F., Manneberg, M., Ozmen, L. & Garotta, G. (1991) Purification and biochemical characterization of a soluble mouse interferon- receptor produced in insect cells, Eur: 1. Biochrm. 198, 441- 450.
104. Fafournoux, P., Ghysdael, J., Sardet, C. & Pouysskgur, J. (1991) Functional expression of the human growth factor activatable Na ' /H ' aritipor-ler (NHE- I ) i r i t~aculovirus-iriftict~~ cells, Bio- chemisrT 30,9510-9515.
105. Rankl, N., Rice, J., Gurganus, T., Barbee, J. & Burns, J. (1994) The production of an active protein kinase C-in insect cells is greatly enhanced by the use of the basic protein promoter, Pro- tein Expr: Purif 5, 346-356.
106. Jarvis, D. & Finn, E. (1995) Biochemical analysis of the N-glyco- sylation pathway i n baculovirus-iiif~~ted lepidopteran insect cells, Virology 212, 500-511.
64, 681-691.




















