
Crithidia deanei: Influence of parasite gp63 homologue on the interaction of endosymbiont-harboring...
9
Crithidia deanei: Influence of parasite gp63 homologue on the interaction of endosymbiont-harboring and aposymbiotic strains with Aedes aegypti midgut Claudia M. d’Avila-Levy a, * , Lı ´via O. Santos a , Fernanda A. Marinho b , Filipe P. Matteoli b , Angela H.C.S. Lopes b , Maria Cristina M. Motta c , Andre ´ L.S. Santos b , Marta H. Branquinha b a Laborato ´ rio de Biologia Molecular e Doenc ¸as Ende ˆmicas, Instituto Oswaldo Cruz (IOC), FIOCRUZ, Rio de Janeiro, RJ 21045-900, Brazil b Instituto de Microbiologia Prof. Paulo de Go ´ es, Centro de Cie ˆncias da Sau ´ de, Bloco I, Universidade Federal do Rio de Janeiro, Cidade Universita ´ ria, Rio de Janeiro, RJ 21941-590, Brazil c Instituto de Biofı ´sica Carlos Chagas Filho, Centro de Cie ˆncias da Sau ´ de, Universidade Federal do Rio de Janeiro, Cidade Universita ´ ria, Rio de Janeiro, RJ 21941-590, Brazil Received 28 May 2007; received in revised form 4 September 2007; accepted 10 September 2007 Available online 15 September 2007 Abstract The present study demonstrates that the endosymbiont of Crithidia deanei influences the expression of surface gp63 molecules. Ultra- structural immunocytochemical analysis shows the presence of the gp63-like protein in the protozoan flagellum and flagellar pocket, either attached to shed membranes or in a free form. This molecule is glycosylphosphatidylinositol (GPI) anchored to the plasma mem- brane as demonstrated by phospholipase C (PLC) treatment and cross-reacting determinant detection by immunoblotting. The gp63 molecule mediates the adhesive process of the protozoan to Aedes aegypti explanted guts, since the binding was reduced by pre-incubat- ing the C. deanei parasites (wild and aposymbiotic strains) with anti-gp63 antibodies, PLC or PLC followed by anti-gp63 antibodies incu- bation. In addition, the number of wild C. deanei bound to A. aegypti explanted guts was twice as that of aposymbiotic parasites. Flow cytometry assays revealed that the reactivity of the wild strain with anti-gp63 antibodies was approximately twice as that of the aposym- biotic strain. We may conclude that higher expression of surface gp63 by the wild strain of C. deanei may positively influence this inter- action, posing a prominent advantage for the endosymbiont-containing trypanosomatids. Ó 2007 Elsevier Inc. All rights reserved. Index Descriptors and Abbreviations: Adhesion; Aedes aegypti; Cellular interaction; Crithidia deanei; Endosymbiont; gp63; Insect; Peptidase; Trypano- somatids 1. Introduction The genus Crithidia comprises single-host parasites of insects that present a barleycorn form, called choanomasti- gote, in their life cycle (McGhee and Cosgrove, 1980). Cri- thidia deanei contains an intracellular symbiotic bacterium, which can be removed by antibiotic treatment, originating an aposymbiotic or cured strain. The comparison between endosymbiont-containing and endosymbiont-free strains of the same species enables a better understanding of how the symbiotic bacterium influences morphological and physi- cal–chemical aspects in the host trypanosomatid (reviewed by De Souza and Motta, 1999). The endosymbiont have a mutualistic relationship with the host protozoan, so that the endosymbiont-harboring trypanosomatids present low nutritional requirements and diminished secretion of prote- olytic enzymes (McGhee and Cosgrove, 1980; De Souza and Motta, 1999; d’Avila-Levy et al., 2001, 2003a, 0014-4894/$ - see front matter Ó 2007 Elsevier Inc. All rights reserved. doi:10.1016/j.exppara.2007.09.007 * Corresponding author. Fax: +55 21 2590 3495. E-mail addresses: davila.levy@ioc.fiocruz.br, [email protected] (C.M. d’Avila-Levy). www.elsevier.com/locate/yexpr Available online at www.sciencedirect.com Experimental Parasitology 118 (2008) 345–353
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Crithidia deanei: Influence of parasite gp63 homologue on the interaction of endosymbiont-harboring...

Available online at www.sciencedirect.com
www.elsevier.com/locate/yexpr
Experimental Parasitology 118 (2008) 345–353
Crithidia deanei: Influence of parasite gp63 homologueon the interaction of endosymbiont-harboring and aposymbiotic
strains with Aedes aegypti midgut
Claudia M. d’Avila-Levy a,*, Lıvia O. Santos a, Fernanda A. Marinho b, Filipe P. Matteoli b,Angela H.C.S. Lopes b, Maria Cristina M. Motta c, Andre L.S. Santos b,
Marta H. Branquinha b
a Laboratorio de Biologia Molecular e Doencas Endemicas, Instituto Oswaldo Cruz (IOC), FIOCRUZ, Rio de Janeiro, RJ 21045-900, Brazilb Instituto de Microbiologia Prof. Paulo de Goes, Centro de Ciencias da Saude, Bloco I, Universidade Federal do Rio de Janeiro,
Cidade Universitaria, Rio de Janeiro, RJ 21941-590, Brazilc Instituto de Biofısica Carlos Chagas Filho, Centro de Ciencias da Saude, Universidade Federal do Rio de Janeiro,
Cidade Universitaria, Rio de Janeiro, RJ 21941-590, Brazil
Received 28 May 2007; received in revised form 4 September 2007; accepted 10 September 2007Available online 15 September 2007
Abstract
The present study demonstrates that the endosymbiont of Crithidia deanei influences the expression of surface gp63 molecules. Ultra-structural immunocytochemical analysis shows the presence of the gp63-like protein in the protozoan flagellum and flagellar pocket,either attached to shed membranes or in a free form. This molecule is glycosylphosphatidylinositol (GPI) anchored to the plasma mem-brane as demonstrated by phospholipase C (PLC) treatment and cross-reacting determinant detection by immunoblotting. The gp63molecule mediates the adhesive process of the protozoan to Aedes aegypti explanted guts, since the binding was reduced by pre-incubat-ing the C. deanei parasites (wild and aposymbiotic strains) with anti-gp63 antibodies, PLC or PLC followed by anti-gp63 antibodies incu-bation. In addition, the number of wild C. deanei bound to A. aegypti explanted guts was twice as that of aposymbiotic parasites. Flowcytometry assays revealed that the reactivity of the wild strain with anti-gp63 antibodies was approximately twice as that of the aposym-biotic strain. We may conclude that higher expression of surface gp63 by the wild strain of C. deanei may positively influence this inter-action, posing a prominent advantage for the endosymbiont-containing trypanosomatids.� 2007 Elsevier Inc. All rights reserved.
Index Descriptors and Abbreviations: Adhesion; Aedes aegypti; Cellular interaction; Crithidia deanei; Endosymbiont; gp63; Insect; Peptidase; Trypano-somatids
1. Introduction
The genus Crithidia comprises single-host parasites ofinsects that present a barleycorn form, called choanomasti-gote, in their life cycle (McGhee and Cosgrove, 1980). Cri-
thidia deanei contains an intracellular symbiotic bacterium,which can be removed by antibiotic treatment, originating
0014-4894/$ - see front matter � 2007 Elsevier Inc. All rights reserved.
doi:10.1016/j.exppara.2007.09.007
* Corresponding author. Fax: +55 21 2590 3495.E-mail addresses: [email protected], [email protected]
(C.M. d’Avila-Levy).
an aposymbiotic or cured strain. The comparison betweenendosymbiont-containing and endosymbiont-free strains ofthe same species enables a better understanding of how thesymbiotic bacterium influences morphological and physi-cal–chemical aspects in the host trypanosomatid (reviewedby De Souza and Motta, 1999). The endosymbiont have amutualistic relationship with the host protozoan, so thatthe endosymbiont-harboring trypanosomatids present lownutritional requirements and diminished secretion of prote-olytic enzymes (McGhee and Cosgrove, 1980; De Souzaand Motta, 1999; d’Avila-Levy et al., 2001, 2003a,

346 C.M. d’Avila-Levy et al. / Experimental Parasitology 118 (2008) 345–353
2005a). Moreover, the symbiont interferes with severalaspects of the host trypanosomatid metabolism (Mundimand Roitman, 1977; Salzman et al., 1985; Frossard et al.,2006), induces several morphological alterations in the try-panosomatid cells (Freymuller and Camargo, 1981; Gade-lha et al., 2005), modulates the glycocalyx composition(Esteves et al., 1982; Oda et al., 1984; Faria-e-Silva et al.,1994; d’Avila-Levy et al., 2005b) and reduces the surfacecharge (Oda et al., 1984). Previous studies showed thatendosymbiont-bearing strains are more efficient in bindingto insect cell lines and midguts, as compared to symbiotic-free strains, which is probably related to differential expres-sion of surface molecules (Fampa et al., 2003; d’Avila-Levyet al., 2005b).
Proteolytic enzymes are implicated with various facetsof host–parasite relationships, including parasite attach-ment, survival and pathogenesis (reviewed by Sajid andMcKerrow, 2002). The leishmanial surface metallopepti-dase, gp63, is a major glycoprotein and is mainly anchoredto the plasma membrane of promastigotes by a glycosyl-phosphatidylinositol (GPI) anchor (reviewed by Yaoet al., 2003). gp63 Molecules on the surface of promastig-otes constitute a significant part of the interface betweenthe invading parasite and the mammalian cell or insect hostat the time of infection. The functions performed by gp63in the interaction of Leishmania with the vertebrate hostsare well characterized, such as extracellular matrix degra-dation, resistance of promastigotes to complement-medi-ated lysis, intracellular amastigote survival withinmacrophage phagolysosomes, and either a direct or indi-rect role in receptor-mediated phagocytosis of Leishmania
(reviewed by Yao et al., 2003). On the other hand, the roleof gp63 in the life cycle of Leishmania during the insect col-onization is poorly explored. Nevertheless, it is known thatgp63 is predominantly expressed on the surface of prom-astigotes residing in the midgut of the phlebotomine sand-fly vector (Grimm et al., 1987; Davies et al., 1990).However, data obtained from gp63 knockouts in Leish-mania major do not support a major role for this metallo-peptidase in parasite survival in the insect (Joshi et al.,2002). Conversely, in Leishmania amazonensis the down-regulation of gp63 adversely affected its early developmentin the neotropical Lutzomyia longipalpis sand fly (Hajmovaet al., 2004). Some insights on the functions performed bygp63 in the insects may be obtained by analyzing gp63homologues in trypanosomatids other than Leishmania(reviewed by Santos et al., 2006b). Homologues of gp63have been described either being released or on the cell sur-face of trypanosomatids belonging to the genera Crithidia
(Jaffe and Dwyer, 2003; d’Avila-Levy et al., 2003a,2006a), Herpetomonas (Elias et al., 2006; Nogueira de Meloet al., 2006; Santos et al., 2006b), Phytomonas (d’Avila-Levy et al., 2006a, 2006b; Santos et al., 2006a, 2006b), Lep-
tomonas (Jaffe and Dwyer, 2003) and Blastocrithidia(d’Avila-Levy et al., 2005a). Moreover, it was recentlyshown by our research group that gp63-like molecules indifferent trypanosomatids, including C. deanei, are relevant
for the flagellate adhesion to the insect gut (reviewed bySantos et al., 2006b).
In the present study we report that the number of wildC. deanei bound to Aedes aegypti explanted guts was twiceas that of aposymbiotic parasites. Also, pre-incubation ofC. deanei (wild and aposymbiotic parasites) with anti-gp63 antibodies, phospholipase C (PLC) or both (PLC fol-lowed by anti-gp63) considerably reduced this adhesion.We also demonstrate here, by flow cytometry analysis, thatanti-gp63 antibody shows a reduced binding rate to the cellsurface of the aposymbiotic strain of C. deanei, as com-pared to the wild strain. Western blotting and flow cytom-etry analyses indicate that these gp63-like molecules areGPI-anchored to the parasite surface, and ultrastructuralanalysis demonstrates the presence of the gp63-like proteinon the protozoan surface, flagellum and in the flagellarpocket, where it was seen either free or attached to shedmembranes.
2. Materials and methods
2.1. Parasites and cultivation
The endosymbiont-containing (ATCC 30255, CT-IOC044) and the aposymbiotic counterpart strain of C. deanei
were provided by Dr. Maria Auxiliadora de Sousa (Cole-cao de Tripanossomatıdeos, FIOCRUZ, Rio de Janeiro,Brazil) and Dr. Maria Cristina Machado Motta, respec-tively. The parasites were cultivated at 28 �C in 3.7% (w/v)brain heart infusion (BHI) medium supplemented with2 mg% (w/v) hemin and 2% (v/v) heat-inactive fetal bovineserum (FBS) in a total volume of 5 ml. The parasites weregrown from 12 to 96 h, harvested by centrifugation at 12 hintervals, washed twice in 0.01 M phosphate buffered saline(PBS) and used for all assays.
2.2. Parasite treatment with PLC
Live parasites (5.0 · 107 cells) were incubated from30 min to 6 h at 37 �C with 0.1 U/ll of Bacillus thuringien-
sis PLC (Sigma Chemical Co., St. Louis, MO, USA)diluted in 50 ll of PBS supplemented with 25% glycerol.Control cells were subjected to the same experimental con-ditions, except for the presence of PLC. Cellular viabilitywas assessed through mobility, Trypan blue cell dye exclu-sion and by monitoring malate dehydrogenase, an intracel-lular enzyme, in the supernatants (d’Avila-Levy et al.,2003b). Supernatants from the reaction mixtures were col-lected by centrifugation (10 min at 1500g), filtered in a0.22-lm membrane (Millipore) and used in the immuno-blotting assays (d’Avila-Levy et al., 2005a).
2.3. Immunoblotting assays
Immunoblottting analysis was performed with thesupernatants from the reaction mixtures of either PLC-treated or control cells (50 lg of protein). The primary

C.M. d’Avila-Levy et al. / Experimental Parasitology 118 (2008) 345–353 347
antibodies used were a rabbit antiserum raised againstrecombinant gp63 (H52) from Leishmania mexicana (Ilget al., 1993) at 1:500 dilution (kindly provided by Dr. PeterOverath, Max-Planck-Institut fur Biologie, AbteilungMembranbiochemie, Germany) or anti-cross-reactingdeterminant (CRD) at 1:400 dilution (kindly provided byDr. Michael A.J. Ferguson and Dr. Maria Lucia S. Guther,University of Dundee, UK). The secondary antibody usedwas horseradish peroxidase-conjugated goat anti-rabbitIgG at 1:25,000 (Pierce, USA). The membranes weredeveloped by chemiluminescence followed by exposure toX-rays films (d’Avila-Levy et al., 2005a).
2.4. Flow cytometry analysis
For fluorescence-activated cell sorter (FACS) analyses,the parasites (1.0 · 106 cells) were fixed for 30 min at roomtemperature in 0.4% (v/v) paraformaldehyde diluted inPBS (100 ll), followed by extensive washing in the samebuffer. The fixed cells maintained their morphologicalintegrity, as verified by light microscopic observation.The cells were then incubated for 1 h at room temperaturewith either a 1:1000 dilution of H52 or rabbit pre-immuneserum (in 100 ll of PBS) (kindly provided by Dr. PeterOverath), and then incubated for an additional hour witha 1:250 dilution (in 100 ll of PBS) of fluorescein isothiocy-anate (FITC)-labeled goat anti-rabbit IgG (Sigma, USA).The cells were then washed three times in PBS and the par-asite-associated fluorescence was excited at 488 nm andquantified on a FACSCalibur (BD Bioscience, San Jose,CA, USA). Non-treated cells, those treated with thepre-immune serum and those treated with the secondaryantibody alone were run in parallel as controls. Eachexperimental population was then mapped by using atwo-parameter histogram of forward-angle light scatterversus side scatter. The mapped population (n = 10,000)was then analyzed for log green fluorescence by using a sin-gle-parameter histogram.
2.5. gp63 Immunofluorescence
The wild strain of C. deanei (1.0 · 106 cells) was adheredto cover slips that had been previously coated with poly-L-lysine (Sigma) for 5 min. Cells were fixed for 5 min in 2%paraformaldehyde diluted in PBS (100 ll), washed twicein the same buffer, and permeabilized for 15 min with1.6% Triton X-100 diluted in 100 ll of PBS. After twowashes in PBS, cells were incubated for 1 h in 100 ll ofPBS containing 1% bovine serum albumin (blocking solu-tion). Cells were washed twice with PBS and incubatedfor 1 h with rabbit anti-gp63 antiserum diluted (1:200) in100 ll of the blocking solution. After two washes in theblocking solution, the cells were incubated for 1 h withAlexa Fluor 488 dye-conjugated goat anti-rabbit antibody(1:800 in 100 ll of PBS) and mounted in glass slides. Con-trol cells were not incubated with the primary antibody.
Samples were observed in a Zeiss LSM S10 meta laser scan-ning confocal microscope.
2.6. gp63 Immunocytochemistry
The wild strain of C. deanei (1.0 · 106 cells) was fixed in100 ll of a solution containing 0.1% glutaraldehyde, 2%formaldehyde in 0.1% cacodylate buffer, pH 7.2. After1 h fixation, cells were washed in PBS, dehydrated in meth-anol, and embedded in Lowicryl K4M resin at �20 �C.Ultrathin sections were collected in nickel grids and incu-bated for 30 min in 50 mM ammonium chloride followedby 30 min incubation in PBS containing 3% bovine serumalbumin, 0.5% teleostean gelatin and 0.02% Tween 20(blocking solution). Then, the grids were incubated for1 h with the rabbit anti-gp63 antibody diluted at 1:50 inthe blocking solution. Grids were washed three times inPBS and subsequently incubated with gold-labeled goatanti-rabbit IgG (10 nm, dilution 1:250) (Sigma, USA) for45 min. Ultrathin sections were stained with uranyl acetateand lead citrate and observed in a Zeiss 900 transmissionelectron microscope.
2.7. Protozoa–insect gut interaction
Adult female mosquitoes (A. aegypti) were reared andmaintained at Laboratorio de Fisiologia e Controle de Vet-ores, Departamento de Entomologia (Instituto OswaldoCruz, Rio de Janeiro, Brazil). Live parasites were previ-ously treated with (i) PLC alone, (ii) PLC followed by incu-bation with anti-gp63 antibody, (iii) directly in the presenceof anti-gp63 antibody, or (iv) rabbit pre-immune serum,both antibodies at 1:1000 dilution, for 1 h at room temper-ature in 100 ll of PBS. After that, parasite viability wasassessed through mobility, Trypan blue cell dye exclusionand by monitoring malate dehydrogenase in the superna-tants (d’Avila-Levy et al., 2003b). The binding of protozoato insect guts was performed as previously described(Fampa et al., 2003). Briefly, trypanosomatids (5.0 · 106
cells in 100 ll) were added to dissected guts (10 per group)and incubated for 1 h at room temperature in PBS (100 ll).After the interaction period, the guts were extensivelywashed with PBS, as judged by examination of the explant-ed guts under an Olympus inverted microscope, and twoguts were transferred to microcentrifuge tubes containing40 ll of PBS and homogenized. The released trypanoso-matids were counted in a Neubauer chamber. The resultsare shown as means ± standard error (SE) of three inde-pendent experiments (Fampa et al., 2003; d’Avila-Levyet al., 2005b, 2006a; reviewed by Santos et al., 2006b).
2.8. Statistical analysis
The experiments were performed in triplicate, in threeindependent experimental sets. The analysis of variancebetween groups was made by means of ANOVA test usingEPI-INFO 6.04 (Database and Statistics Program for
348 C.M. d’Avila-Levy et al. / Experimental Parasitology 118 (2008) 345–353
Public Health) computer software. P values of 0.05 or lesswere considered statistically significant.
3. Results
3.1. Flow cytometry for cell surface gp63
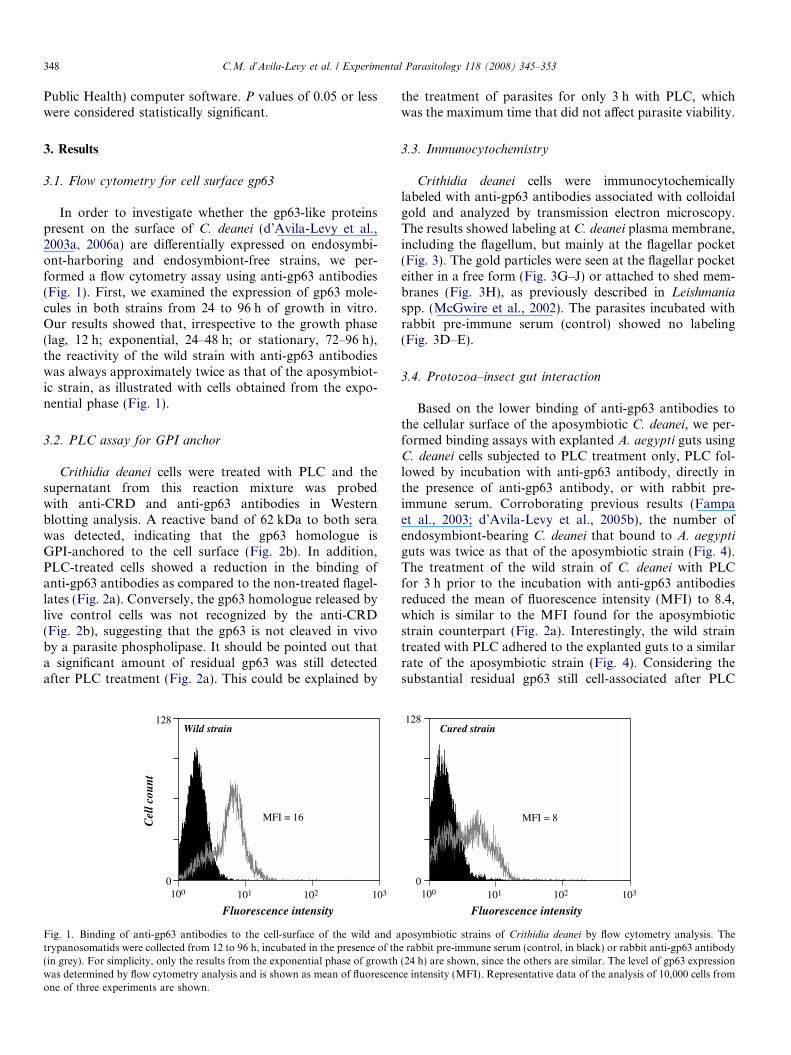
In order to investigate whether the gp63-like proteinspresent on the surface of C. deanei (d’Avila-Levy et al.,2003a, 2006a) are differentially expressed on endosymbi-ont-harboring and endosymbiont-free strains, we per-formed a flow cytometry assay using anti-gp63 antibodies(Fig. 1). First, we examined the expression of gp63 mole-cules in both strains from 24 to 96 h of growth in vitro.Our results showed that, irrespective to the growth phase(lag, 12 h; exponential, 24–48 h; or stationary, 72–96 h),the reactivity of the wild strain with anti-gp63 antibodieswas always approximately twice as that of the aposymbiot-ic strain, as illustrated with cells obtained from the expo-nential phase (Fig. 1).
3.2. PLC assay for GPI anchor
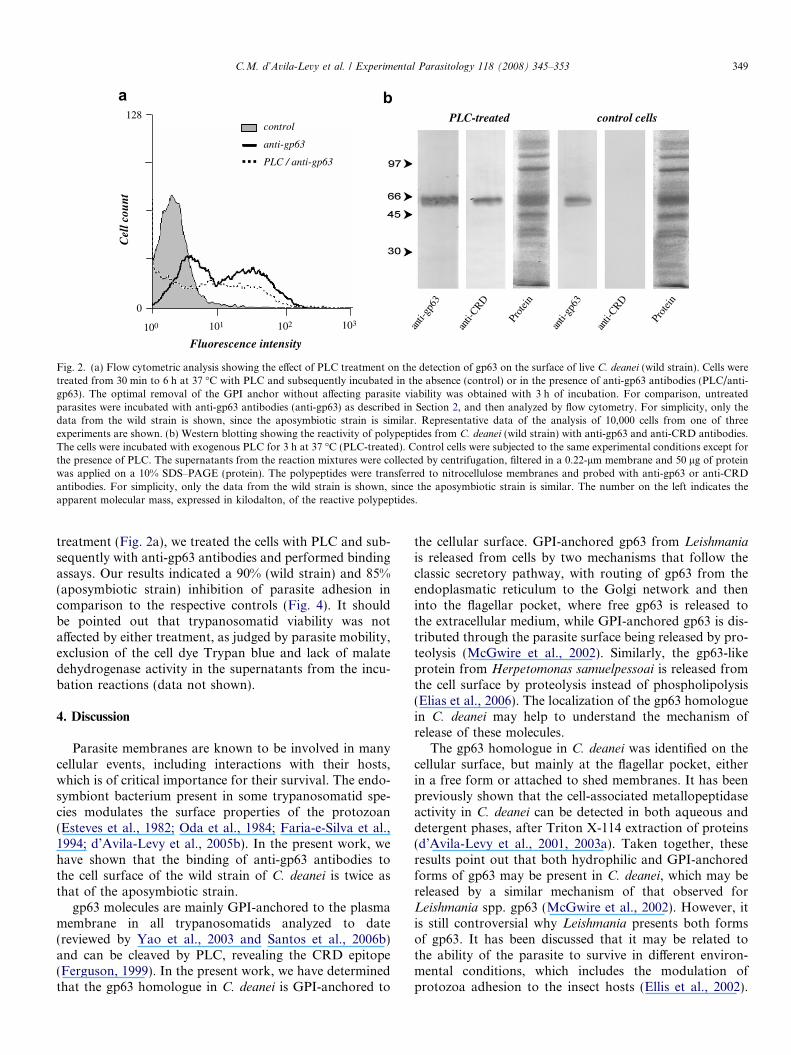
Crithidia deanei cells were treated with PLC and thesupernatant from this reaction mixture was probedwith anti-CRD and anti-gp63 antibodies in Westernblotting analysis. A reactive band of 62 kDa to both serawas detected, indicating that the gp63 homologue isGPI-anchored to the cell surface (Fig. 2b). In addition,PLC-treated cells showed a reduction in the binding ofanti-gp63 antibodies as compared to the non-treated flagel-lates (Fig. 2a). Conversely, the gp63 homologue released bylive control cells was not recognized by the anti-CRD(Fig. 2b), suggesting that the gp63 is not cleaved in vivoby a parasite phospholipase. It should be pointed out thata significant amount of residual gp63 was still detectedafter PLC treatment (Fig. 2a). This could be explained by
103
128
0100 101 102
Wild strain
MFI = 16
Fluorescence intensity
Cel
l cou
nt
Fig. 1. Binding of anti-gp63 antibodies to the cell-surface of the wild and atrypanosomatids were collected from 12 to 96 h, incubated in the presence of th(in grey). For simplicity, only the results from the exponential phase of growthwas determined by flow cytometry analysis and is shown as mean of fluorescenone of three experiments are shown.
the treatment of parasites for only 3 h with PLC, whichwas the maximum time that did not affect parasite viability.
3.3. Immunocytochemistry
Crithidia deanei cells were immunocytochemicallylabeled with anti-gp63 antibodies associated with colloidalgold and analyzed by transmission electron microscopy.The results showed labeling at C. deanei plasma membrane,including the flagellum, but mainly at the flagellar pocket(Fig. 3). The gold particles were seen at the flagellar pocketeither in a free form (Fig. 3G–J) or attached to shed mem-branes (Fig. 3H), as previously described in Leishmania
spp. (McGwire et al., 2002). The parasites incubated withrabbit pre-immune serum (control) showed no labeling(Fig. 3D–E).
3.4. Protozoa–insect gut interaction
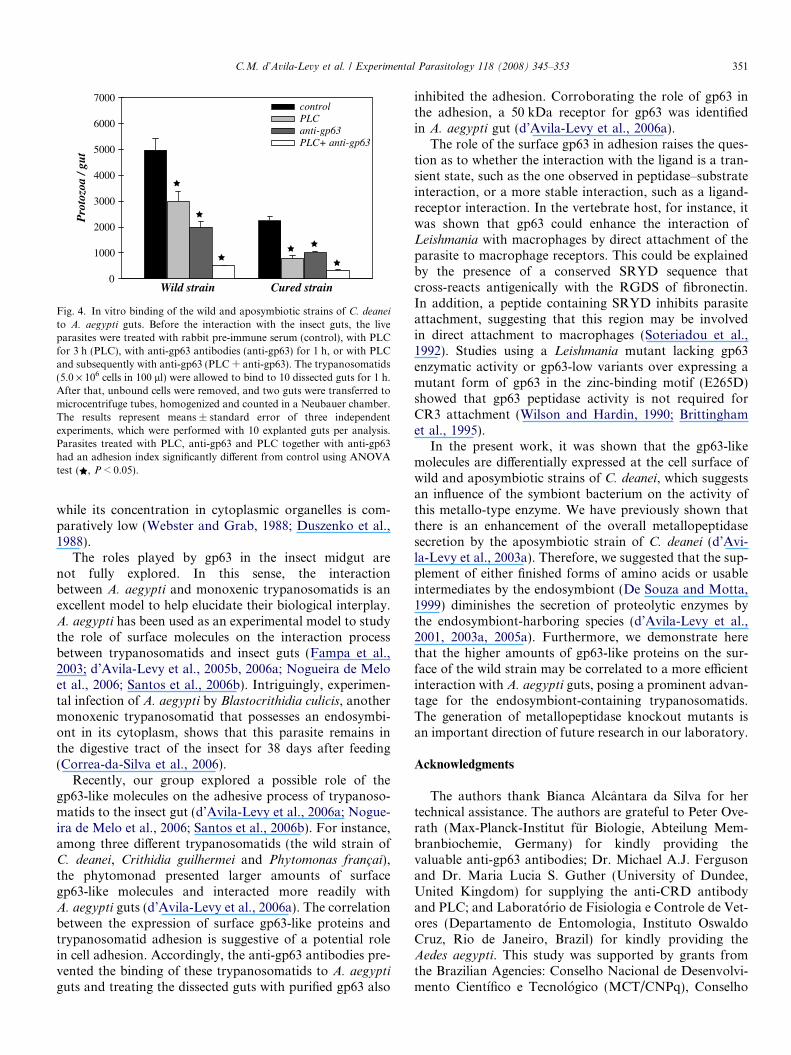
Based on the lower binding of anti-gp63 antibodies tothe cellular surface of the aposymbiotic C. deanei, we per-formed binding assays with explanted A. aegypti guts usingC. deanei cells subjected to PLC treatment only, PLC fol-lowed by incubation with anti-gp63 antibody, directly inthe presence of anti-gp63 antibody, or with rabbit pre-immune serum. Corroborating previous results (Fampaet al., 2003; d’Avila-Levy et al., 2005b), the number ofendosymbiont-bearing C. deanei that bound to A. aegypti
guts was twice as that of the aposymbiotic strain (Fig. 4).The treatment of the wild strain of C. deanei with PLCfor 3 h prior to the incubation with anti-gp63 antibodiesreduced the mean of fluorescence intensity (MFI) to 8.4,which is similar to the MFI found for the aposymbioticstrain counterpart (Fig. 2a). Interestingly, the wild straintreated with PLC adhered to the explanted guts to a similarrate of the aposymbiotic strain (Fig. 4). Considering thesubstantial residual gp63 still cell-associated after PLC
Cured strain
MFI = 8
128
0103100 101 102
Fluorescence intensity
posymbiotic strains of Crithidia deanei by flow cytometry analysis. Thee rabbit pre-immune serum (control, in black) or rabbit anti-gp63 antibody(24 h) are shown, since the others are similar. The level of gp63 expressionce intensity (MFI). Representative data of the analysis of 10,000 cells from
128C
ell c
ount
103
0
100 101 102
control
anti-gp63
PLC / anti-gp63
anti-
gp63
anti-
CRD
anti-
gp63
anti-
CRD
Prot
ein
Prot
ein
Fluorescence intensity
PLC-treated control cells
Fig. 2. (a) Flow cytometric analysis showing the effect of PLC treatment on the detection of gp63 on the surface of live C. deanei (wild strain). Cells weretreated from 30 min to 6 h at 37 �C with PLC and subsequently incubated in the absence (control) or in the presence of anti-gp63 antibodies (PLC/anti-gp63). The optimal removal of the GPI anchor without affecting parasite viability was obtained with 3 h of incubation. For comparison, untreatedparasites were incubated with anti-gp63 antibodies (anti-gp63) as described in Section 2, and then analyzed by flow cytometry. For simplicity, only thedata from the wild strain is shown, since the aposymbiotic strain is similar. Representative data of the analysis of 10,000 cells from one of threeexperiments are shown. (b) Western blotting showing the reactivity of polypeptides from C. deanei (wild strain) with anti-gp63 and anti-CRD antibodies.The cells were incubated with exogenous PLC for 3 h at 37 �C (PLC-treated). Control cells were subjected to the same experimental conditions except forthe presence of PLC. The supernatants from the reaction mixtures were collected by centrifugation, filtered in a 0.22-lm membrane and 50 lg of proteinwas applied on a 10% SDS–PAGE (protein). The polypeptides were transferred to nitrocellulose membranes and probed with anti-gp63 or anti-CRDantibodies. For simplicity, only the data from the wild strain is shown, since the aposymbiotic strain is similar. The number on the left indicates theapparent molecular mass, expressed in kilodalton, of the reactive polypeptides.
C.M. d’Avila-Levy et al. / Experimental Parasitology 118 (2008) 345–353 349
treatment (Fig. 2a), we treated the cells with PLC and sub-sequently with anti-gp63 antibodies and performed bindingassays. Our results indicated a 90% (wild strain) and 85%(aposymbiotic strain) inhibition of parasite adhesion incomparison to the respective controls (Fig. 4). It shouldbe pointed out that trypanosomatid viability was notaffected by either treatment, as judged by parasite mobility,exclusion of the cell dye Trypan blue and lack of malatedehydrogenase activity in the supernatants from the incu-bation reactions (data not shown).
4. Discussion
Parasite membranes are known to be involved in manycellular events, including interactions with their hosts,which is of critical importance for their survival. The endo-symbiont bacterium present in some trypanosomatid spe-cies modulates the surface properties of the protozoan(Esteves et al., 1982; Oda et al., 1984; Faria-e-Silva et al.,1994; d’Avila-Levy et al., 2005b). In the present work, wehave shown that the binding of anti-gp63 antibodies tothe cell surface of the wild strain of C. deanei is twice asthat of the aposymbiotic strain.
gp63 molecules are mainly GPI-anchored to the plasmamembrane in all trypanosomatids analyzed to date(reviewed by Yao et al., 2003 and Santos et al., 2006b)and can be cleaved by PLC, revealing the CRD epitope(Ferguson, 1999). In the present work, we have determinedthat the gp63 homologue in C. deanei is GPI-anchored to
the cellular surface. GPI-anchored gp63 from Leishmania
is released from cells by two mechanisms that follow theclassic secretory pathway, with routing of gp63 from theendoplasmatic reticulum to the Golgi network and theninto the flagellar pocket, where free gp63 is released tothe extracellular medium, while GPI-anchored gp63 is dis-tributed through the parasite surface being released by pro-teolysis (McGwire et al., 2002). Similarly, the gp63-likeprotein from Herpetomonas samuelpessoai is released fromthe cell surface by proteolysis instead of phospholipolysis(Elias et al., 2006). The localization of the gp63 homologuein C. deanei may help to understand the mechanism ofrelease of these molecules.
The gp63 homologue in C. deanei was identified on thecellular surface, but mainly at the flagellar pocket, eitherin a free form or attached to shed membranes. It has beenpreviously shown that the cell-associated metallopeptidaseactivity in C. deanei can be detected in both aqueous anddetergent phases, after Triton X-114 extraction of proteins(d’Avila-Levy et al., 2001, 2003a). Taken together, theseresults point out that both hydrophilic and GPI-anchoredforms of gp63 may be present in C. deanei, which may bereleased by a similar mechanism of that observed forLeishmania spp. gp63 (McGwire et al., 2002). However, itis still controversial why Leishmania presents both formsof gp63. It has been discussed that it may be related tothe ability of the parasite to survive in different environ-mental conditions, which includes the modulation ofprotozoa adhesion to the insect hosts (Ellis et al., 2002).
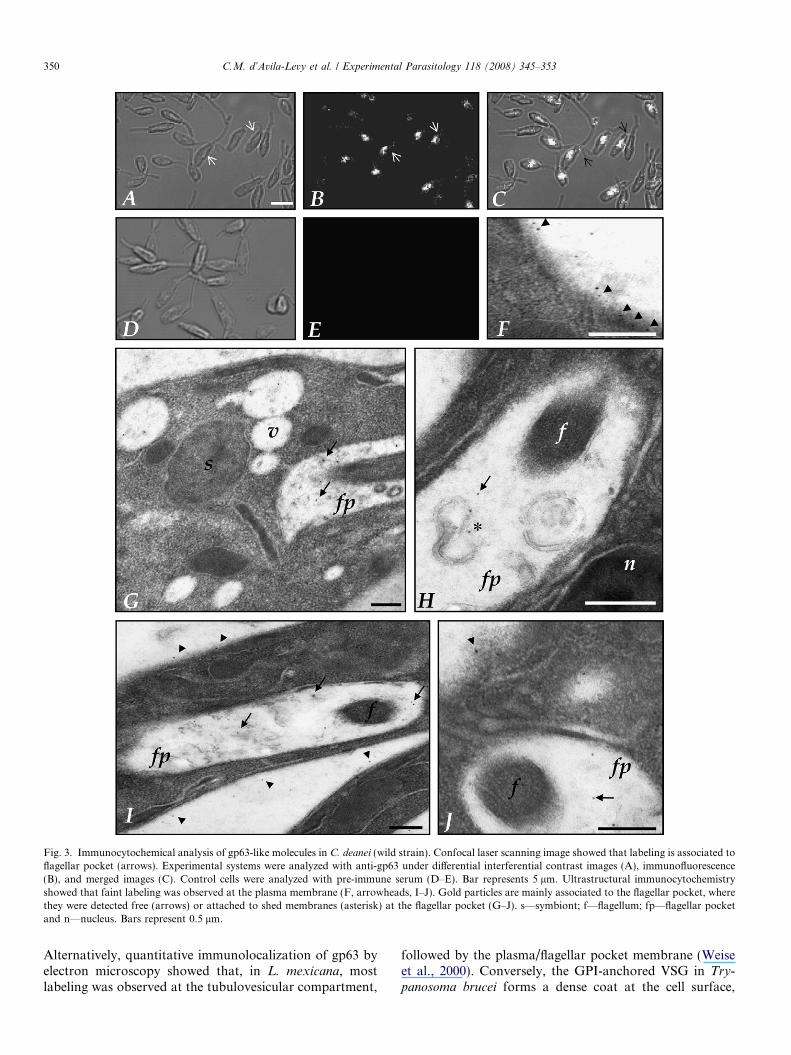
Fig. 3. Immunocytochemical analysis of gp63-like molecules in C. deanei (wild strain). Confocal laser scanning image showed that labeling is associated toflagellar pocket (arrows). Experimental systems were analyzed with anti-gp63 under differential interferential contrast images (A), immunofluorescence(B), and merged images (C). Control cells were analyzed with pre-immune serum (D–E). Bar represents 5 lm. Ultrastructural immunocytochemistryshowed that faint labeling was observed at the plasma membrane (F, arrowheads, I–J). Gold particles are mainly associated to the flagellar pocket, wherethey were detected free (arrows) or attached to shed membranes (asterisk) at the flagellar pocket (G–J). s—symbiont; f—flagellum; fp—flagellar pocketand n—nucleus. Bars represent 0.5 lm.
350 C.M. d’Avila-Levy et al. / Experimental Parasitology 118 (2008) 345–353
Alternatively, quantitative immunolocalization of gp63 byelectron microscopy showed that, in L. mexicana, mostlabeling was observed at the tubulovesicular compartment,
followed by the plasma/flagellar pocket membrane (Weiseet al., 2000). Conversely, the GPI-anchored VSG in Try-
panosoma brucei forms a dense coat at the cell surface,
Pro
tozo
a / g
ut
0
1000
2000
3000
4000
5000
6000
7000controlPLCanti-gp63PLC+ anti-gp63
Wild strain Cured strain
Fig. 4. In vitro binding of the wild and aposymbiotic strains of C. deanei
to A. aegypti guts. Before the interaction with the insect guts, the liveparasites were treated with rabbit pre-immune serum (control), with PLCfor 3 h (PLC), with anti-gp63 antibodies (anti-gp63) for 1 h, or with PLCand subsequently with anti-gp63 (PLC + anti-gp63). The trypanosomatids(5.0 · 106 cells in 100 ll) were allowed to bind to 10 dissected guts for 1 h.After that, unbound cells were removed, and two guts were transferred tomicrocentrifuge tubes, homogenized and counted in a Neubauer chamber.The results represent means ± standard error of three independentexperiments, which were performed with 10 explanted guts per analysis.Parasites treated with PLC, anti-gp63 and PLC together with anti-gp63had an adhesion index significantly different from control using ANOVAtest ( , P < 0.05).
C.M. d’Avila-Levy et al. / Experimental Parasitology 118 (2008) 345–353 351
while its concentration in cytoplasmic organelles is com-paratively low (Webster and Grab, 1988; Duszenko et al.,1988).
The roles played by gp63 in the insect midgut arenot fully explored. In this sense, the interactionbetween A. aegypti and monoxenic trypanosomatids is anexcellent model to help elucidate their biological interplay.A. aegypti has been used as an experimental model to studythe role of surface molecules on the interaction processbetween trypanosomatids and insect guts (Fampa et al.,2003; d’Avila-Levy et al., 2005b, 2006a; Nogueira de Meloet al., 2006; Santos et al., 2006b). Intriguingly, experimen-tal infection of A. aegypti by Blastocrithidia culicis, anothermonoxenic trypanosomatid that possesses an endosymbi-ont in its cytoplasm, shows that this parasite remains inthe digestive tract of the insect for 38 days after feeding(Correa-da-Silva et al., 2006).
Recently, our group explored a possible role of thegp63-like molecules on the adhesive process of trypanoso-matids to the insect gut (d’Avila-Levy et al., 2006a; Nogue-ira de Melo et al., 2006; Santos et al., 2006b). For instance,among three different trypanosomatids (the wild strain ofC. deanei, Crithidia guilhermei and Phytomonas francai),the phytomonad presented larger amounts of surfacegp63-like molecules and interacted more readily withA. aegypti guts (d’Avila-Levy et al., 2006a). The correlationbetween the expression of surface gp63-like proteins andtrypanosomatid adhesion is suggestive of a potential rolein cell adhesion. Accordingly, the anti-gp63 antibodies pre-vented the binding of these trypanosomatids to A. aegypti
guts and treating the dissected guts with purified gp63 also
inhibited the adhesion. Corroborating the role of gp63 inthe adhesion, a 50 kDa receptor for gp63 was identifiedin A. aegypti gut (d’Avila-Levy et al., 2006a).
The role of the surface gp63 in adhesion raises the ques-tion as to whether the interaction with the ligand is a tran-sient state, such as the one observed in peptidase–substrateinteraction, or a more stable interaction, such as a ligand-receptor interaction. In the vertebrate host, for instance, itwas shown that gp63 could enhance the interaction ofLeishmania with macrophages by direct attachment of theparasite to macrophage receptors. This could be explainedby the presence of a conserved SRYD sequence thatcross-reacts antigenically with the RGDS of fibronectin.In addition, a peptide containing SRYD inhibits parasiteattachment, suggesting that this region may be involvedin direct attachment to macrophages (Soteriadou et al.,1992). Studies using a Leishmania mutant lacking gp63enzymatic activity or gp63-low variants over expressing amutant form of gp63 in the zinc-binding motif (E265D)showed that gp63 peptidase activity is not required forCR3 attachment (Wilson and Hardin, 1990; Brittinghamet al., 1995).
In the present work, it was shown that the gp63-likemolecules are differentially expressed at the cell surface ofwild and aposymbiotic strains of C. deanei, which suggestsan influence of the symbiont bacterium on the activity ofthis metallo-type enzyme. We have previously shown thatthere is an enhancement of the overall metallopeptidasesecretion by the aposymbiotic strain of C. deanei (d’Avi-la-Levy et al., 2003a). Therefore, we suggested that the sup-plement of either finished forms of amino acids or usableintermediates by the endosymbiont (De Souza and Motta,1999) diminishes the secretion of proteolytic enzymes bythe endosymbiont-harboring species (d’Avila-Levy et al.,2001, 2003a, 2005a). Furthermore, we demonstrate herethat the higher amounts of gp63-like proteins on the sur-face of the wild strain may be correlated to a more efficientinteraction with A. aegypti guts, posing a prominent advan-tage for the endosymbiont-containing trypanosomatids.The generation of metallopeptidase knockout mutants isan important direction of future research in our laboratory.
Acknowledgments
The authors thank Bianca Alcantara da Silva for hertechnical assistance. The authors are grateful to Peter Ove-rath (Max-Planck-Institut fur Biologie, Abteilung Mem-branbiochemie, Germany) for kindly providing thevaluable anti-gp63 antibodies; Dr. Michael A.J. Fergusonand Dr. Maria Lucia S. Guther (University of Dundee,United Kingdom) for supplying the anti-CRD antibodyand PLC; and Laboratorio de Fisiologia e Controle de Vet-ores (Departamento de Entomologia, Instituto OswaldoCruz, Rio de Janeiro, Brazil) for kindly providing theAedes aegypti. This study was supported by grants fromthe Brazilian Agencies: Conselho Nacional de Desenvolvi-mento Cientıfico e Tecnologico (MCT/CNPq), Conselho

352 C.M. d’Avila-Levy et al. / Experimental Parasitology 118 (2008) 345–353
de Ensino para Graduados e Pesquisa (CEPG/UFRJ),Fundacao de Amparo a Pesquisa do Rio de Janeiro (FA-PERJ), Fundacao Universitaria Jose Bonifacio (FUJB)and FIOCRUZ (Fundacao Oswaldo Cruz).
References
Brittingham, A., Morrison, C.J., Mcmaster, W.R., Mcgwire, B.S., Chang,K.P., Mosser, D.M., 1995. Role of the Leishmania surface proteasegp63 in complement fixation, cell adhesion, and resistance to comple-ment-mediated lysis. Journal of Immunolology 155, 3102–3111.
Correa-da-Silva, M.S., Fampa, P., Lessa, L.P., Dos Reis Silva, E., DosSantos Mallet, J.R., Saraiva, E.M., Motta, M.C., 2006. Colonizationof Aedes aegypti midgut by the endosymbiont-bearing trypanosomatidBlastocrithidia culicis. Parasitology Research 99, 384–391.
Davies, C.R., Cooper, A.M., Peacock, C., Lane, R.P., Blackwell, J.M.,1990. Expression of LPG and GP63 by different developmental stagesof Leishmania major in the sandfly Phlebotomus papatasi. Parasitology101, 337–343.
d’Avila-Levy, C.M., Melo, A.C.N., Vermelho, A.B., Branquinha, M.H.,2001. Differential expression of proteolytic enzymes in endosymbiont-harboring Crithidia species. FEMS Microbiology Letters 202, 73–77.
d’Avila-Levy, C.M., Souza, R.F., Gomes, R.C., Vermelho, A.B., Bran-quinha, M.H., 2003a. A metalloproteinase extracellularly releasedfrom Crithidia deanei. Canadian Journal of Microbiology 49, 625–632.
d’Avila-Levy, C.M., Souza, R.F., Gomes, R.C., Vermelho, A.B., Bran-quinha, M.H., 2003b. A novel extracellular calcium-dependent cysteineproteinase from Crithidia deanei. Archives of Biochemistry andBiophysics 420, 1–8.
d’Avila-Levy, C.M., Araujo, F.M., Vermelho, A.B., Soares, R.M.A.,Santos, A.L.S., Branquinha, M.H., 2005a. Proteolytic expression inBlastocrithidia culicis: influence of the endosymbiont and similaritieswith virulence factors of pathogenic trypanosomatids. Parasitology130, 413–420.
d’Avila-Levy, C.M., Silva, B.A., Hayashi, E.A., Vermelho, A.B., Alviano,C.S., Saraiva, E.M., Branquinha, M.H., Santos, A.L.S., 2005b.Influence of the endosymbiont of Blastocrithidia culicis and Crithidia
deanei on the glycoconjugate expression and on Aedes aegypti
interaction. FEMS Microbiology Letters 252, 279–286.d’Avila-Levy, C.M., Dias, F.A., Melo, A.C.N., Martins, J.L., Lopes,
A.H.C., Santos, A.L.S., Vermelho, A.B., Branquinha, M.H., 2006a.Insights on the role of gp63-like proteins in lower trypanosomatids.FEMS Microbiology Letters 254, 149–156.
d’Avila-Levy, C.M., Santos, L.O., Marinho, F.A., Dias, F.A., Lopes,A.H.C.S., Santos, A.L.S., Branquinha, M.H., 2006b. Gp63-likemolecules in Phytomonas serpens: possible role on the insect interac-tion. Current Microbiology 52, 439–444.
De Souza, W., Motta, M.C.M., 1999. Endosymbiosis in protozoa of theTrypanosomatidae family. FEMS Microbiology Letters 173, 1–8.
Duszenko, M., Ivanov, I.E., Ferguson, M.A.J., Plesken, H., Cross,G.A.M., 1988. Intracellular transport of a variant surface glycoproteinin Trypanosoma brucei. Journal of Cell Biology 106, 77–86.
Elias, C.G.R., Pereira, F.M., Silva, B.A., Alviano, C.S., Soares, R.M.A.,Santos, A.L.S., 2006. Leishmanolysin (gp63 metallopeptidase)-likeactivity extracellularly released by Herpetomonas samuelpessoai. Par-asitology 132, 37–47.
Ellis, M., Sharma, D.K., Hilley, J.D., Coombst, G.H., Mottram, J.C.,2002. Processing and trafficking of Leishmania mexicana GP63. TheJournal of Biological Chemistry 277, 27968–27974.
Esteves, M.J.G., Andrade, A.F.B., Angluster, J., De Souza, W., Mundim,M.H., Roitman, I., Pereira, M.E.A., 1982. Cell surface carbohydratesin Crithidia deanei: influence of the endosymbiont. The EuropeanJournal of Cell Biology 26, 244–248.
Fampa, P., Correa-da-Silva, M.S., Lima, D.C., Oliveira, S.M.P., Motta,M.C.M., Saraiva, E.M.B., 2003. Interaction of insect trypanosomatidswith mosquitoes, sand fly and the respective insect cell lines.International Journal for Parasitology 33, 1019–1026.
Faria-e-Silva, P.M., Fiorini, J.E., Soares, M.J., Alviano, C.S., De Souza,W., Angluster, J., 1994. Membrane-associated polysaccharides com-position, nutritional requirements and cell differentiation in Herpeto-
monas roitmani: influence of the endosymbiont. (Kinetoplastida:Trypanosomatidae). The Journal of Eukaryotic Microbiology 41,55–59.
Ferguson, M.A.J., 1999. The structure, biosynthesis and functions ofglycosylphosphatidylinositol anchors, and the contributions of try-panosome research. Journal of Cell Science 112, 2799–2809.
Freymuller, E., Camargo, E.P., 1981. Ultrastructural differences betweenspecies of trypanosomatids with and without endosymbionts. Journalof Protozoology 28, 175–182.
Frossard, M.L., Seabra, S.H., DaMatta, R.A., de Souza, W., Mello, F.G.,Motta, M.C.M., 2006. An endosymbiont positively modulates orni-thine decarboxylase in host trypanosomatids. Biochemical BiophysicalResearch Communications 343, 443–449.
Gadelha, C., Wickstead, B., De Souza, W., Gull, K., Cunha-e-Silva, N.,2005. Cryptic paraflagellar rod in endosymbiont-containing kineto-plastid protozoa. Eukaryotic Cell 4, 516–524.
Grimm, F., Jenni, L., Bouvier, J., Etges, R.J., Bordier, C., 1987. Thepromastigote surface protease of Leishmania donovani infantum in themidgut of Phlebhotomus perniciosus. Acta Tropica 44, 375–377.
Hajmova, M., Chang, K.P., Kolli, B., Volf, P., 2004. Down-regulation ofgp63 in Leishmania amazonensis reduces its early development inLutzomyia longipalpis. Microbes and Infection 6, 646–649.
Ilg, T., Harbecke, D., Overath, P., 1993. The lysosomal gp63-relatedprotein in Leishmania mexicana amastigotes is a soluble metallopro-teinase with an acidic pH optimum. FEBS Letters 327, 103–107.
Jaffe, C.L., Dwyer, D.M., 2003. Extracellular release of the surfacemetalloprotease, gp63, from Leishmania and insect trypanosomatids.Parasitology Research 91, 229–237.
Joshi, P.B., Kelly, B.L., Kamhawi, S., Sacks, D.L., McMaster, W.R.,2002. Targeted gene deletion in Leishmania major identifies leishman-olysin (GP63) as a virulence factor. Molecular and BiochemicalParasitology 120, 33–40.
McGhee, R.B., Cosgrove, W.B., 1980. Biology and physiology of thelower Trypanosomatidae. Microbiological Reviews 44, 140–173.
McGwire, B.S., O’Connell, W.A., Chang, K.P., Engman, D., 2002.Extracellular release of the glycosylphosphatidylinositol (GPI)-linkedLeishmania surface metalloprotease, gp63, is independent of GPIphospholipolysis. The Journal of Biological Chemistry 277, 8802–8809.
Mundim, M.H., Roitman, I., 1977. Extra nutritional requirements ofartificially aposymbiotic Crithidia deanei. Journal of Protozoology 24,329–331.
Nogueira de Melo, A.C., d’Avila–Levy, C.M., Dias, F.A., Armada,J.L.A., Silva, H.D., Lopes, A.H.C., Santos, A.L.S., Branquinha,M.H., Vermelho, A.B., 2006. Peptidases and gp63-like proteins inHerpetomonas megaseliae: possible involvement in the adhesion to theinvertebrate host. International Journal for Parasitology 36, 415–422.
Oda, L.M., Alviano, C.S., Silva-Filho, F.C., Angluster, J., Roitman, I., DeSouza, W., 1984. Surface anionic groups in symbiont-free strains ofCrithidia deanei. Journal of Protozoology 31, 131–134.
Sajid, M., McKerrow, J.H., 2002. Cysteine proteases of parasiticorganisms. Molecular and Biochemical Parasitology 120, 1–21.
Salzman, T.A., Del, C., Battle, A.M., Angluster, J., De Souza, W., 1985.Heme synthesis in Crithidia deanei: influence of the endosymbiote.International Journal of Biochemisty 17, 1343–1347.
Santos, A.L.S., Branquinha, M.H., d’Avila-Levy, C.M., 2006a. Theubiquitous gp63-like metalloprotease from lower trypanosomatids: inthe search for a function. Anais da Academia Brasileira de Ciencia 78,387–714.
Santos, A.L.S., d’Avila-Levy, C.M., Elias, C.G.R., Vermelho, A.B.,Branquinha, M.H., 2006b. Phytomonas serpens: immunological simi-larities with the human trypanosomatid pathogens. Microbes andInfection 9, 915–921.
Soteriadou, K.P., Remoundos, M.S., Katsikas, M.C., Tzinia, A.K.,Tsikaris, V., Sakarellos, C., Tzartos, S.J., 1992. The Ser-Arg-Tyr-Asp

C.M. d’Avila-Levy et al. / Experimental Parasitology 118 (2008) 345–353 353
region of the major surface glycoprotein of Leishmania mimics theArg-Gly-Asp-Ser cell attachment region of fibronectin. The Journal ofBiological Chemistry 267, 13980–13985.
Webster, P., Grab, D., 1988. Intracellular co-localization of variantsurface glycoprotein and transferring-gold in Trypanosoma brucei. TheJournal of Cell Biology 106, 279–288.
Weise, F., Stierhof, Y., Kuhn, C., Wiese, M., Overath, P., 2000.Distribution of GPI-anchored proteins in the protozoan parasite
Leishmania, based on the improved ultrastructural description usinghigh-pressure frozen cells. Journal of Cell Science 113, 4587–4603.
Wilson, M.E., Hardin, K.K., 1990. The major Leishmania donovani
chagasi surface glycoprotein in tunicamycin-resistant promastigotes.Journal of Immunology 144, 4825–4834.
Yao, C., Donelson, J.E., Wilson, M.E., 2003. The major surface protease(MSP or GP63) of Leishmania sp. biosynthesis, regulation of expres-sion, and function. Molecular and Biochemical Parasitology 132, 1–16.




















