
Molecular identification of a bevy of serine proteinases in Manduca sexta hemolymph
Upload
independentCategory
view
1download
0
Journal of Insect Physiology 56 (2010) 774–779
Midgut and fatbody mitochondrial transhydrogenase activities duringlarval-pupal development of the tobacco hornworm, Manduca sexta
Kurt P. Vandock *, Christopher A. Drummond 1, Stan L. Smith 2, Carmen F. Fioravanti
Department of Biological Sciences, Bowling Green State University, Bowling Green, OH, USA
A R T I C L E I N F O
Article history:
Received 11 November 2009
Received in revised form 21 January 2010
Accepted 22 January 2010
Keywords:
Mitochondrial transhydrogenase
Energy-linked transhydrogenation
Manduca sexta development
Ecdysone 20-monooxygenase
20-hydroxyecdysone
A B S T R A C T
Midgut and fatbody mitochondria from fifth larval instar Manduca sexta display a membrane-
associated transhydrogenase that catalyzes a reversible hydride ion transfer between NADP(H) and
NAD(H). The NADPH-forming activity occurs as a nonenergy- or energy-linked activity with energy
for the latter derived from either electron transport-dependent NADH or succinate utilization, or ATP
hydrolysis by Mg++-dependent ATPase. During the ten-day developmental period preceding the
larval-pupal molt (fifth larval instar), significant peaks in the mitochondrial transhydrogenase
activities of midgut and fatbody tissues were noted and these peaks were coincident with the onset of
wandering behavior and with the fifty-fold increase in ecdysone 20-monooxygenase (E20-M) activity
previously reported for M. sexta midgut. Since E20-M preferentially uses NADPH in catalyzing
ecdysone conversion to the physiologically active molting hormone, 20-hydroxyecdysone, the
physiological and developmental significance of the mitochondrial, NADPH-forming energy-linked
transhydrogenations were made apparent. Moreover, that the increases in all transhydrogenase
activities resulted from de novo enzyme synthesis were indicated by the cycloheximide-dependent
reductions in these activities.
� 2010 Elsevier Ltd. All rights reserved.
Contents lists available at ScienceDirect
Journal of Insect Physiology
journa l homepage: www.e lsev ier .com/ locate / j insphys
1. Introduction
Midgut mitochondria from wandering stage, fifth larvalinstar Manduca sexta exhibit a membrane-associated transhy-drogenase (E.C. 1.6.1.1) that catalyzes the following reversiblereaction:
NADPH þ NADþ $ NADPþ þNADH
(Vandock et al., 2008). The NADH-forming activity and the NADPH-forming activities are termed the NADPH! NAD+ andNADH! NADP+ reactions, respectively. The latter reaction occursas a nonenergy- or energy-linked activity. The reversible M. sexta
mitochondrial transhydrogenase of midgut tissues was character-ized and apparently acts as a transmembrane proton-translocatingsystem (Vandock et al., 2008).
The essential need for conversion of the molting hormone,ecdysone, to its physiologically active form, 20-hydroxyecdysone
* Corresponding author. Department of Biological Sciences, Bowling Green State
University, 217 Life Sciences Building, Bowling Green, OH 43403, USA
Tel.: +1 419 372 2634; fax: +1 419 372 2024.
E-mail address: [email protected] (K.P. Vandock).1 Presently not affiliated with Bowling Green State University.2 Deceased.
0022-1910/$ – see front matter � 2010 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jinsphys.2010.01.008
(20-HE) during M. sexta post-embryonic development is estab-lished (Gilbert et al., 2002). 20-HE formation is catalyzed by acytochrome P450-dependent steroid hydroxylase, viz., ecdysone20-monooxygenase (E20-M; E.C. 1.14.99.22) (Smith et al., 1979;Feyereisen, 2005; Lafont et al., 2005). In M. sexta, E20-M activity ispredominantly associated with the mitochondria of the ecdysonetarget tissues (e.g., midgut, fatbody, and Malphighian tubules) andpreferentially uses NADPH as the reductant (Smith, 1985; Weirich,1997; Gilbert et al., 2002).
On day five of the fifth larval instar, midgut mitochondria ofM. sexta display a fifty-fold increase in E20-M activity (Mitchellet al., 1999). This peak in day five midgut mitochondrial E20-Mactivity coincides with larval-pupal reprogramming, the onset ofwandering stage behavior and expression of the shade genewhich encodes E20-M (Gilbert and Rewitz, 2009). Moreover,E20-M activity in mitochondria from the fatbody of fifthlarval instar M. sexta displays a peak of activity on day four(Mitchell et al., 1999). The latter increase corresponds to asmall peak in hemolymph ecdysteroid titer that serves as thesignal to reprogram larval tissues into pupal tissues (Lafontet al., 2005).
In the mitochondrial E20-M-catalyzed conversion of ecdy-sone to 20-HE, the need for NADPH is apparent and thisreductant could arise from the NADPH-forming M. sexta

K.P. Vandock et al. / Journal of Insect Physiology 56 (2010) 774–779 775
transhydrogenation (Mayer et al., 1978; Vandock et al., 2008).Coupling of this reaction with E20-M would occur as indicatedhere:
Because energy-linkage of the transhydrogenase promotesNADPH accumulation, NADH or succinate oxidation, via theelectron transport system, or hydrolysis of ATP, by themitochondrial Mg++-dependent ATPase (Vandock et al., 2008),would be the preferred NADPH-forming reactions.
The present study centers on an evaluation of mitochondrialtranshydrogenations in midgut and fatbody tissues of M. sexta
during the developmental period preceding the larval-pupal molt(fifth larval instar). Significant peaks in all M. sexta reversibletranshydrogenase activities on day five were noted in both tissuetypes and corresponded with reported increases in E20-M activityduring the fifth larval instar. More specifically, coincidentincreases in the energy-linked NADPH-forming reactions wereconsistent with the need for NADPH in the conversion of ecdysoneto 20-HE.
2. Materials and methods
2.1. Manduca rearing
The animals, representing each day (1–10) of the fifth larvalinstar used for assessments of transhydrogenations, were stagedfrom fourth larval instar head capsule slippage-brown mand-ibles (pharate fifth instar larvae-day zero) and also were selectedon the basis of empirically generated weight curves placing theanimals in the gate II designation (Goodman et al., 1985).Animals were reared on an artificial diet under a non-diapausingphotoperiod (L:D, 16:8) at 268 C and �60% relative humidity(Bell and Joachim, 1976). Wandering stage animals weredefined on the basis of morphological (dorsal vessel exposurewith lateral pink pigmentation) and behavioral (wandering)markers.
2.2. Mitochondrial isolation
M. sexta mitochondria were isolated from last instar larvalmidguts and fatbodies. Larvae were immobilized by immersionin ice water for 20 min at 4 8C. Subsequently, dissection (at 4 8C)of midgut and fatbody tissues were carried out in Lepidopteranringers (Weevers, 1966). Isolated midguts and fatbodies werewashed in Lepidopteran ringers, blotted, and weighed prior tohomogenization. Mitochondria were isolated essentially asdescribed by Vandock et al. (2008). Midguts and fatbodies werehomogenized in mitochondrial medium (9.0 ml/g tissue) con-sisting of 250 mM sucrose, 15 mM ethylenediaminetetracetate(EDTA) and 10 mM trishydroxymethylaminomethane–HCl (Tris–HCl, pH 7.5). Cellular debris was removed by centrifugation at500 � g for 10 min. Mitochondria were isolated from theresulting supernatant fraction at 10,000 � g for 15 min andwashed at 12,000 � g. The resulting pellet was suspended inmitochondrial medium. All steps of the mitochondrial isolationswere performed at 4 8C.
A comparison of enzyme activities of isolated vs. isolated andsonically treated mitochondria revealed that further mitochon-drial disruption did not result in a significant increase in
enzymatic activity as noted previously (Vandock et al., 2008).Therefore, isolated M. sexta mitochondria were used for allassessments.
2.3. Assessments of enzyme activities
Spectrophotometric assessments of the transhydrogenase wereperformed as described by Vandock et al. (2008). Activities weremeasured by following acetylpyridineNAD(P) (AcPyAD(P)) reduc-tion at 375 nm. For assessment of AcPyAD reduction(NADPH! NAD+ reaction), the 1.0 ml assay system containedenzyme and the following in mmol: Tris–HCl (pH 7.5), 100; NADPH,0.24; AcPyAD, 0.60. Where indicated, 25 mM rotenone waspresent. For assessment of AcPyADP reduction (NADH! NADP+
reactions), the 1.0 ml assay system contained enzyme, 10 mgbovine serum albumin (BSA), and the following in mmol: Tris–HCl(pH 7.5), 100; NADH, 0.24; AcPyADP, 0.60.
Additions to the NADH! NADP+ transhydrogenation assaysvaried by reaction. For assessment of the nonenergy-linkedNADPH-forming transhydrogenation the assay contained 25 mMrotenone. For the reaction which coupled NADH oxidation viathe electron transport system, the NADH! NADP+ transhydro-genation was assessed in the presence or absence of 25 mMrotenone and the difference calculated to yield the electrontransport-dependent activity. The NADPH-forming transhydro-genation dependent on succinate oxidation was assessed in thepresence of 25 mM rotenone and 3.0 mM succinate. ATP-dependent activity was measured by addition of 25 mMrotenone, 2.0 mM ATP, and 3.0 mM MgCl2. Rotenone, wasdispensed in ethanolic solution such that the ethanol contentwas 1.9% in the assays. Appropriate controls were evaluatedcontaining 1.9% ethanol alone. All enzyme assessments wereperformed at 25 8C.
2.4. Injections
Animals to be injected were chilled on ice. Injections weregiven via the abdominal prolegs using a 10 ml Hamilton syringeand the injection site ligated with waxed dental floss.Cycloheximide dispensed in 70% ethanol (20, 40 or 60 mg)was injected into animals on day three and total injectionvolumes were 10 ml. Control animals were injected with 10 ml of70% ethanol.
2.5. Protein assessment
Protein content of mitochondrial preparations was determinedby the method of Bradford (1976) using BSA as the standard.
2.6. Statistical analysis and post hoc tests
One Way ANOVAs were carried out to determine differencesbetween means of different days (P < 0.05). Statisticallysignificant ANOVAs were then subjected to Tukey’s PSLD posthoc analysis. P value for difference determinations wereall <0.05 (JMP 7.0, SAS, 2007). Data are represented as themean total activities in micromoles of reducing equivalentstransferred per minute per milligram of wet tissueweight � SEM.
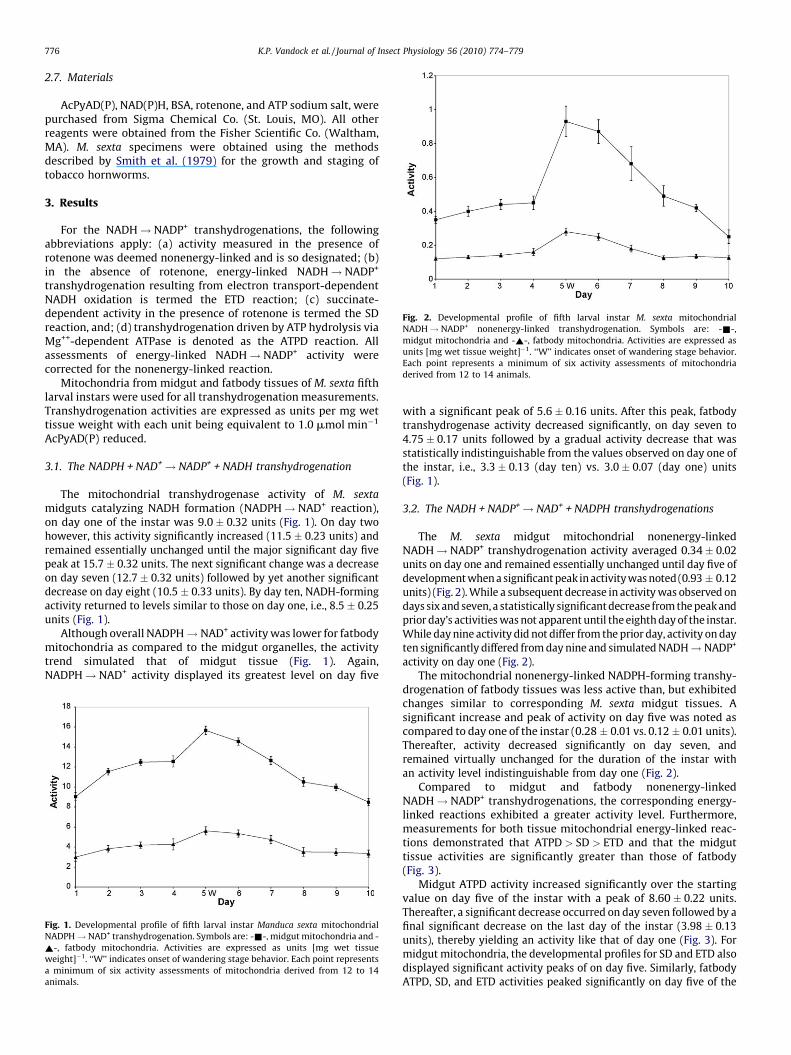
Fig. 2. Developmental profile of fifth larval instar M. sexta mitochondrial
NADH! NADP+ nonenergy-linked transhydrogenation. Symbols are: -&-,
midgut mitochondria and -~-, fatbody mitochondria. Activities are expressed as
units [mg wet tissue weight]�1. ‘‘W’’ indicates onset of wandering stage behavior.
Each point represents a minimum of six activity assessments of mitochondria
derived from 12 to 14 animals.
K.P. Vandock et al. / Journal of Insect Physiology 56 (2010) 774–779776
2.7. Materials
AcPyAD(P), NAD(P)H, BSA, rotenone, and ATP sodium salt, werepurchased from Sigma Chemical Co. (St. Louis, MO). All otherreagents were obtained from the Fisher Scientific Co. (Waltham,MA). M. sexta specimens were obtained using the methodsdescribed by Smith et al. (1979) for the growth and staging oftobacco hornworms.
3. Results
For the NADH! NADP+ transhydrogenations, the followingabbreviations apply: (a) activity measured in the presence ofrotenone was deemed nonenergy-linked and is so designated; (b)in the absence of rotenone, energy-linked NADH! NADP+
transhydrogenation resulting from electron transport-dependentNADH oxidation is termed the ETD reaction; (c) succinate-dependent activity in the presence of rotenone is termed the SDreaction, and; (d) transhydrogenation driven by ATP hydrolysis viaMg++-dependent ATPase is denoted as the ATPD reaction. Allassessments of energy-linked NADH! NADP+ activity werecorrected for the nonenergy-linked reaction.
Mitochondria from midgut and fatbody tissues of M. sexta fifthlarval instars were used for all transhydrogenation measurements.Transhydrogenation activities are expressed as units per mg wettissue weight with each unit being equivalent to 1.0 mmol min�1
AcPyAD(P) reduced.
3.1. The NADPH + NAD+! NADP+ + NADH transhydrogenation
The mitochondrial transhydrogenase activity of M. sexta
midguts catalyzing NADH formation (NADPH! NAD+ reaction),on day one of the instar was 9.0 � 0.32 units (Fig. 1). On day twohowever, this activity significantly increased (11.5 � 0.23 units) andremained essentially unchanged until the major significant day fivepeak at 15.7 � 0.32 units. The next significant change was a decreaseon day seven (12.7 � 0.32 units) followed by yet another significantdecrease on day eight (10.5 � 0.33 units). By day ten, NADH-formingactivity returned to levels similar to those on day one, i.e., 8.5 � 0.25units (Fig. 1).
Although overall NADPH! NAD+ activity was lower for fatbodymitochondria as compared to the midgut organelles, the activitytrend simulated that of midgut tissue (Fig. 1). Again,NADPH! NAD+ activity displayed its greatest level on day five
Fig. 1. Developmental profile of fifth larval instar Manduca sexta mitochondrial
NADPH! NAD+ transhydrogenation. Symbols are: -&-, midgut mitochondria and -
~-, fatbody mitochondria. Activities are expressed as units [mg wet tissue
weight]�1. ‘‘W’’ indicates onset of wandering stage behavior. Each point represents
a minimum of six activity assessments of mitochondria derived from 12 to 14
animals.
with a significant peak of 5.6 � 0.16 units. After this peak, fatbodytranshydrogenase activity decreased significantly, on day seven to4.75 � 0.17 units followed by a gradual activity decrease that wasstatistically indistinguishable from the values observed on day one ofthe instar, i.e., 3.3 � 0.13 (day ten) vs. 3.0 � 0.07 (day one) units(Fig. 1).
3.2. The NADH + NADP+! NAD+ + NADPH transhydrogenations
The M. sexta midgut mitochondrial nonenergy-linkedNADH! NADP+ transhydrogenation activity averaged 0.34 � 0.02units on day one and remained essentially unchanged until day five ofdevelopment when a significant peak in activity was noted (0.93 � 0.12units) (Fig. 2). While a subsequent decrease in activity was observed ondays six and seven, a statistically significant decrease from the peak andprior day’s activities was not apparent until the eighth day of the instar.While day nine activity did not differ from the prior day, activity on dayten significantly differed from day nine and simulated NADH! NADP+
activity on day one (Fig. 2).The mitochondrial nonenergy-linked NADPH-forming transhy-
drogenation of fatbody tissues was less active than, but exhibitedchanges similar to corresponding M. sexta midgut tissues. Asignificant increase and peak of activity on day five was noted ascompared to day one of the instar (0.28 � 0.01 vs. 0.12 � 0.01 units).Thereafter, activity decreased significantly on day seven, andremained virtually unchanged for the duration of the instar withan activity level indistinguishable from day one (Fig. 2).
Compared to midgut and fatbody nonenergy-linkedNADH! NADP+ transhydrogenations, the corresponding energy-linked reactions exhibited a greater activity level. Furthermore,measurements for both tissue mitochondrial energy-linked reac-tions demonstrated that ATPD > SD > ETD and that the midguttissue activities are significantly greater than those of fatbody(Fig. 3).
Midgut ATPD activity increased significantly over the startingvalue on day five of the instar with a peak of 8.60 � 0.22 units.Thereafter, a significant decrease occurred on day seven followed by afinal significant decrease on the last day of the instar (3.98 � 0.13units), thereby yielding an activity like that of day one (Fig. 3). Formidgut mitochondria, the developmental profiles for SD and ETD alsodisplayed significant activity peaks of on day five. Similarly, fatbodyATPD, SD, and ETD activities peaked significantly on day five of the
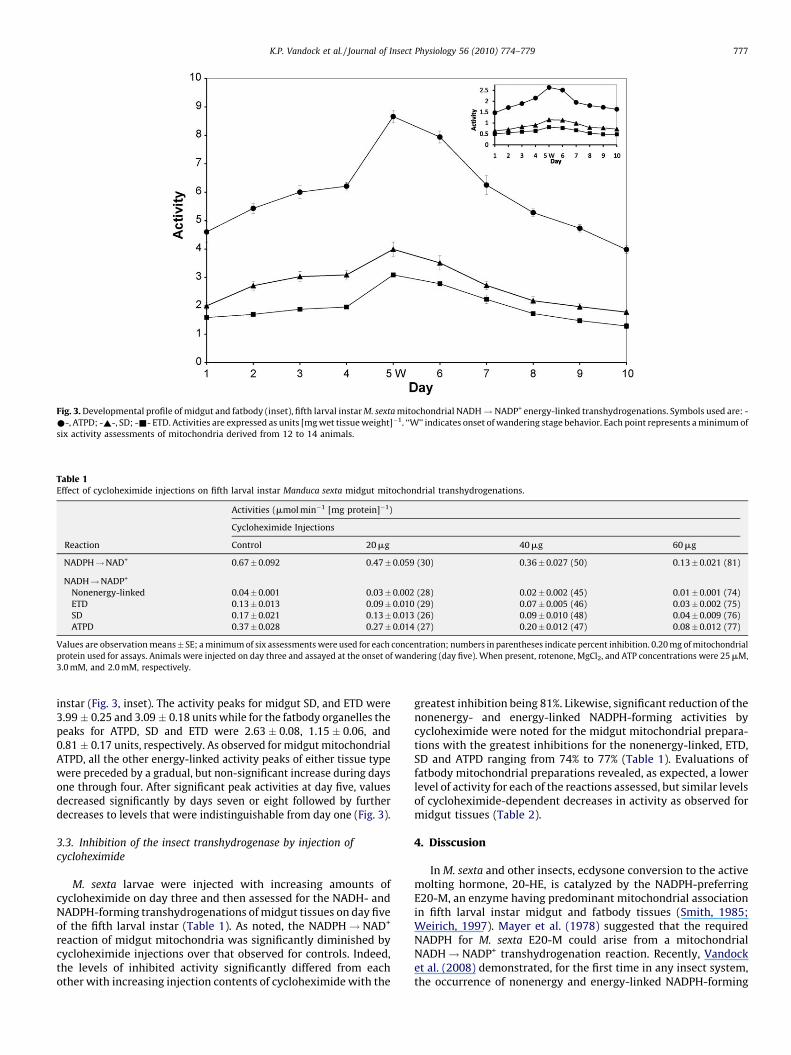
Fig. 3. Developmental profile of midgut and fatbody (inset), fifth larval instar M. sexta mitochondrial NADH! NADP+ energy-linked transhydrogenations. Symbols used are: -
*-, ATPD; -~-, SD; -&- ETD. Activities are expressed as units [mg wet tissue weight]�1. ‘‘W’’ indicates onset of wandering stage behavior. Each point represents a minimum of
six activity assessments of mitochondria derived from 12 to 14 animals.
Table 1Effect of cycloheximide injections on fifth larval instar Manduca sexta midgut mitochondrial transhydrogenations.
Activities (mmol min�1 [mg protein]�1)
Cycloheximide Injections
Reaction Control 20 mg 40 mg 60 mg
NADPH!NAD+ 0.67�0.092 0.47� 0.059 (30) 0.36�0.027 (50) 0.13�0.021 (81)
NADH!NADP+
Nonenergy-linked 0.04�0.001 0.03� 0.002 (28) 0.02�0.002 (45) 0.01�0.001 (74)
ETD 0.13�0.013 0.09� 0.010 (29) 0.07�0.005 (46) 0.03�0.002 (75)
SD 0.17�0.021 0.13� 0.013 (26) 0.09�0.010 (48) 0.04�0.009 (76)
ATPD 0.37�0.028 0.27� 0.014 (27) 0.20�0.012 (47) 0.08�0.012 (77)
Values are observation means� SE; a minimum of six assessments were used for each concentration; numbers in parentheses indicate percent inhibition. 0.20 mg of mitochondrial
protein used for assays. Animals were injected on day three and assayed at the onset of wandering (day five). When present, rotenone, MgCl2, and ATP concentrations were 25 mM,
3.0 mM, and 2.0 mM, respectively.
K.P. Vandock et al. / Journal of Insect Physiology 56 (2010) 774–779 777
instar (Fig. 3, inset). The activity peaks for midgut SD, and ETD were3.99 � 0.25 and 3.09 � 0.18 units while for the fatbody organelles thepeaks for ATPD, SD and ETD were 2.63 � 0.08, 1.15 � 0.06, and0.81 � 0.17 units, respectively. As observed for midgut mitochondrialATPD, all the other energy-linked activity peaks of either tissue typewere preceded by a gradual, but non-significant increase during daysone through four. After significant peak activities at day five, valuesdecreased significantly by days seven or eight followed by furtherdecreases to levels that were indistinguishable from day one (Fig. 3).
3.3. Inhibition of the insect transhydrogenase by injection of
cycloheximide
M. sexta larvae were injected with increasing amounts ofcycloheximide on day three and then assessed for the NADH- andNADPH-forming transhydrogenations of midgut tissues on day fiveof the fifth larval instar (Table 1). As noted, the NADPH! NAD+
reaction of midgut mitochondria was significantly diminished bycycloheximide injections over that observed for controls. Indeed,the levels of inhibited activity significantly differed from eachother with increasing injection contents of cycloheximide with the
greatest inhibition being 81%. Likewise, significant reduction of thenonenergy- and energy-linked NADPH-forming activities bycycloheximide were noted for the midgut mitochondrial prepara-tions with the greatest inhibitions for the nonenergy-linked, ETD,SD and ATPD ranging from 74% to 77% (Table 1). Evaluations offatbody mitochondrial preparations revealed, as expected, a lowerlevel of activity for each of the reactions assessed, but similar levelsof cycloheximide-dependent decreases in activity as observed formidgut tissues (Table 2).
4. Disscusion
In M. sexta and other insects, ecdysone conversion to the activemolting hormone, 20-HE, is catalyzed by the NADPH-preferringE20-M, an enzyme having predominant mitochondrial associationin fifth larval instar midgut and fatbody tissues (Smith, 1985;Weirich, 1997). Mayer et al. (1978) suggested that the requiredNADPH for M. sexta E20-M could arise from a mitochondrialNADH! NADP+ transhydrogenation reaction. Recently, Vandocket al. (2008) demonstrated, for the first time in any insect system,the occurrence of nonenergy and energy-linked NADPH-forming
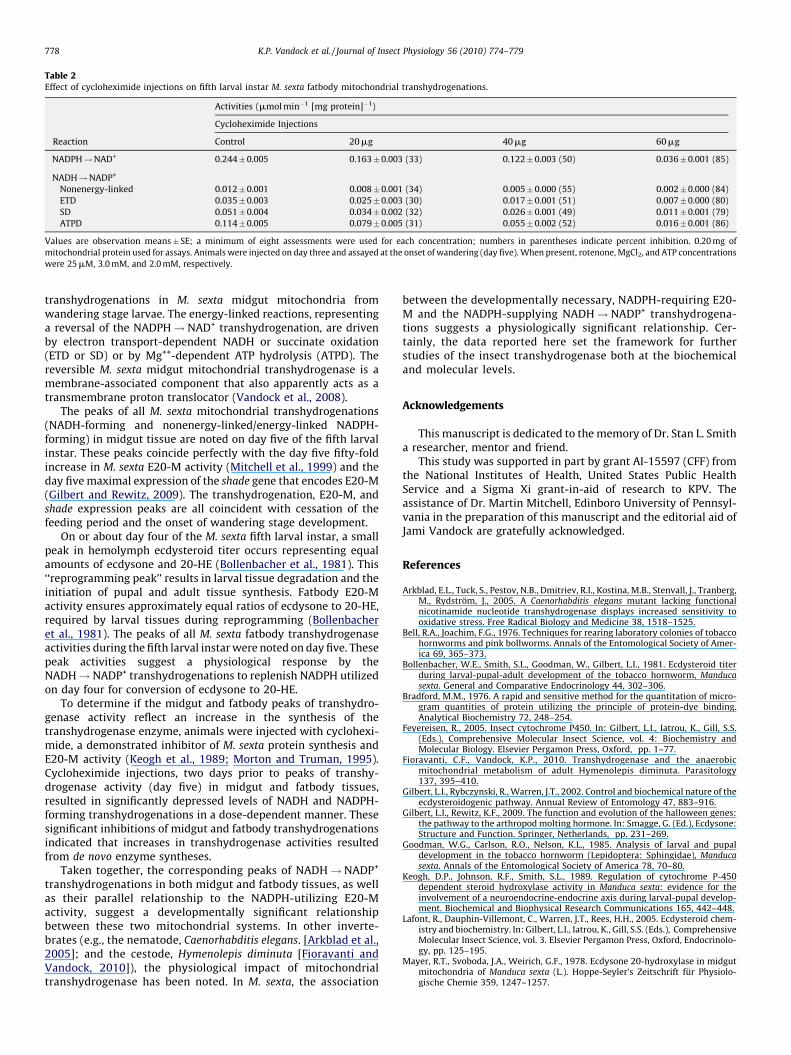
Table 2Effect of cycloheximide injections on fifth larval instar M. sexta fatbody mitochondrial transhydrogenations.
Activities (mmol min�1 [mg protein]�1)
Cycloheximide Injections
Reaction Control 20 mg 40 mg 60 mg
NADPH!NAD+ 0.244�0.005 0.163�0.003 (33) 0.122� 0.003 (50) 0.036�0.001 (85)
NADH!NADP+
Nonenergy-linked 0.012�0.001 0.008�0.001 (34) 0.005� 0.000 (55) 0.002�0.000 (84)
ETD 0.035�0.003 0.025�0.003 (30) 0.017� 0.001 (51) 0.007�0.000 (80)
SD 0.051�0.004 0.034�0.002 (32) 0.026� 0.001 (49) 0.011�0.001 (79)
ATPD 0.114�0.005 0.079�0.005 (31) 0.055� 0.002 (52) 0.016�0.001 (86)
Values are observation means� SE; a minimum of eight assessments were used for each concentration; numbers in parentheses indicate percent inhibition. 0.20 mg of
mitochondrial protein used for assays. Animals were injected on day three and assayed at the onset of wandering (day five). When present, rotenone, MgCl2, and ATP concentrations
were 25 mM, 3.0 mM, and 2.0 mM, respectively.
K.P. Vandock et al. / Journal of Insect Physiology 56 (2010) 774–779778
transhydrogenations in M. sexta midgut mitochondria fromwandering stage larvae. The energy-linked reactions, representinga reversal of the NADPH! NAD+ transhydrogenation, are drivenby electron transport-dependent NADH or succinate oxidation(ETD or SD) or by Mg++-dependent ATP hydrolysis (ATPD). Thereversible M. sexta midgut mitochondrial transhydrogenase is amembrane-associated component that also apparently acts as atransmembrane proton translocator (Vandock et al., 2008).
The peaks of all M. sexta mitochondrial transhydrogenations(NADH-forming and nonenergy-linked/energy-linked NADPH-forming) in midgut tissue are noted on day five of the fifth larvalinstar. These peaks coincide perfectly with the day five fifty-foldincrease in M. sexta E20-M activity (Mitchell et al., 1999) and theday five maximal expression of the shade gene that encodes E20-M(Gilbert and Rewitz, 2009). The transhydrogenation, E20-M, andshade expression peaks are all coincident with cessation of thefeeding period and the onset of wandering stage development.
On or about day four of the M. sexta fifth larval instar, a smallpeak in hemolymph ecdysteroid titer occurs representing equalamounts of ecdysone and 20-HE (Bollenbacher et al., 1981). This‘‘reprogramming peak’’ results in larval tissue degradation and theinitiation of pupal and adult tissue synthesis. Fatbody E20-Mactivity ensures approximately equal ratios of ecdysone to 20-HE,required by larval tissues during reprogramming (Bollenbacheret al., 1981). The peaks of all M. sexta fatbody transhydrogenaseactivities during the fifth larval instar were noted on day five. Thesepeak activities suggest a physiological response by theNADH! NADP+ transhydrogenations to replenish NADPH utilizedon day four for conversion of ecdysone to 20-HE.
To determine if the midgut and fatbody peaks of transhydro-genase activity reflect an increase in the synthesis of thetranshydrogenase enzyme, animals were injected with cyclohexi-mide, a demonstrated inhibitor of M. sexta protein synthesis andE20-M activity (Keogh et al., 1989; Morton and Truman, 1995).Cycloheximide injections, two days prior to peaks of transhy-drogenase activity (day five) in midgut and fatbody tissues,resulted in significantly depressed levels of NADH and NADPH-forming transhydrogenations in a dose-dependent manner. Thesesignificant inhibitions of midgut and fatbody transhydrogenationsindicated that increases in transhydrogenase activities resultedfrom de novo enzyme syntheses.
Taken together, the corresponding peaks of NADH! NADP+
transhydrogenations in both midgut and fatbody tissues, as wellas their parallel relationship to the NADPH-utilizing E20-Mactivity, suggest a developmentally significant relationshipbetween these two mitochondrial systems. In other inverte-brates (e.g., the nematode, Caenorhabditis elegans. [Arkblad et al.,2005]; and the cestode, Hymenolepis diminuta [Fioravanti andVandock, 2010]), the physiological impact of mitochondrialtranshydrogenase has been noted. In M. sexta, the association
between the developmentally necessary, NADPH-requiring E20-M and the NADPH-supplying NADH! NADP+ transhydrogena-tions suggests a physiologically significant relationship. Cer-tainly, the data reported here set the framework for furtherstudies of the insect transhydrogenase both at the biochemicaland molecular levels.
Acknowledgements
This manuscript is dedicated to the memory of Dr. Stan L. Smitha researcher, mentor and friend.
This study was supported in part by grant AI-15597 (CFF) fromthe National Institutes of Health, United States Public HealthService and a Sigma Xi grant-in-aid of research to KPV. Theassistance of Dr. Martin Mitchell, Edinboro University of Pennsyl-vania in the preparation of this manuscript and the editorial aid ofJami Vandock are gratefully acknowledged.
References
Arkblad, E.L., Tuck, S., Pestov, N.B., Dmitriev, R.I., Kostina, M.B., Stenvall, J., Tranberg,M., Rydstrom, J., 2005. A Caenorhabditis elegans mutant lacking functionalnicotinamide nucleotide transhydrogenase displays increased sensitivity tooxidative stress. Free Radical Biology and Medicine 38, 1518–1525.
Bell, R.A., Joachim, F.G., 1976. Techniques for rearing laboratory colonies of tobaccohornworms and pink bollworms. Annals of the Entomological Society of Amer-ica 69, 365–373.
Bollenbacher, W.E., Smith, S.L., Goodman, W., Gilbert, L.I., 1981. Ecdysteroid titerduring larval-pupal-adult development of the tobacco hornworm, Manducasexta. General and Comparative Endocrinology 44, 302–306.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of micro-gram quantities of protein utilizing the principle of protein-dye binding.Analytical Biochemistry 72, 248–254.
Feyereisen, R., 2005. Insect cytochrome P450. In: Gilbert, L.I., Iatrou, K., Gill, S.S.(Eds.), Comprehensive Molecular Insect Science, vol. 4: Biochemistry andMolecular Biology. Elsevier Pergamon Press, Oxford, pp. 1–77.
Fioravanti, C.F., Vandock, K.P., 2010. Transhydrogenase and the anaerobicmitochondrial metabolism of adult Hymenolepis diminuta. Parasitology137, 395–410.
Gilbert, L.I., Rybczynski, R., Warren, J.T., 2002. Control and biochemical nature of theecdysteroidogenic pathway. Annual Review of Entomology 47, 883–916.
Gilbert, L.I., Rewitz, K.F., 2009. The function and evolution of the halloween genes:the pathway to the arthropod molting hormone. In: Smagge, G. (Ed.), Ecdysone:Structure and Function. Springer, Netherlands, pp. 231–269.
Goodman, W.G., Carlson, R.O., Nelson, K.L., 1985. Analysis of larval and pupaldevelopment in the tobacco hornworm (Lepidoptera: Sphingidae), Manducasexta. Annals of the Entomological Society of America 78, 70–80.
Keogh, D.P., Johnson, R.F., Smith, S.L., 1989. Regulation of cytochrome P-450dependent steroid hydroxylase activity in Manduca sexta: evidence for theinvolvement of a neuroendocrine-endocrine axis during larval-pupal develop-ment. Biochemical and Biophysical Research Communications 165, 442–448.
Lafont, R., Dauphin-Villemont, C., Warren, J.T., Rees, H.H., 2005. Ecdysteroid chem-istry and biochemistry. In: Gilbert, L.I., Iatrou, K., Gill, S.S. (Eds.), ComprehensiveMolecular Insect Science, vol. 3. Elsevier Pergamon Press, Oxford, Endocrinolo-gy, pp. 125–195.
Mayer, R.T., Svoboda, J.A., Weirich, G.F., 1978. Ecdysone 20-hydroxylase in midgutmitochondria of Manduca sexta (L.). Hoppe-Seyler’s Zeitschrift fur Physiolo-gische Chemie 359, 1247–1257.

K.P. Vandock et al. / Journal of Insect Physiology 56 (2010) 774–779 779
Mitchell, M.J., Crooks, J.R., Keogh, D.P., Smith, S.L., 1999. Ecdysone 20-monoox-ygenase activity during larval-pupal-adult development of the tobaccohornworm, Manduca sexta. Archives of Insect Biochemistry and Physiology41, 24–32.
Morton, D.B., Truman, J.W., 1995. Effect of cycloheximide on eclosion hormonesensitivity and the developmental appearance of the eclosion hormoneand cGMP regulated phosphoproteins in the CNS of the tobaccohornworm, Manduca sexta. Journal of Receptors and Signal Transduction15, 773–786.
Smith, S.L., Bollenbacher, W.E., Cooper, D.Y., Schleyer, H., Wielgus, J.J., Gilbert, L.I.,1979. Ecdysone 20-monooxygenase: characterization of an insect cytochromeP-450 dependent steroid hydroxylase. Molecular and Cellular Endocrinology15, 111–133.
Smith, S.L., 1985. Regulation of ecdysteroid titer: synthesis. In: Kerkut, G.R., Gilbert,L.I. (Eds.), Comprehensive Insect Physiology, Biochemistry, and Pharmocology.Vol. 7: Endocrinology I. Pergamon Press, Oxford, pp. 295–341.
Vandock, K.P., Smith, S.L., Fioravanti, C.F., 2008. Midgut mitochondrial transhydro-genase in wandering stage larvae of the tobacco hornworm, Manduca sexta.Archives of Insect Biochemistry and Physiology 69, 118–126.
Weevers, R.D., 1966. A lepidopteran saline: effects of inorganic cation concentra-tions on sensory, reflex and motor responses in a herbivorous insect. Journal ofExperimental Biology 44, 163–175.
Weirich, G.F., 1997. Ecdysone 20-hydroxylation in Manduca sexta (Lepidoptera:Sphingidae) midgut: development-related changes of mitochondrial and mi-crosomal ecdysone 20-monooxygenase activities in the fifth larval instar.European Journal of Entomology 94, 57–65.
Copyright © 2022 FDOKUMEN




















