
Hybrid Bacillus amyloliquefaciens X Bacillus licheniformisalpha-Amylases. Construction, Properties...
10
Eur. J. Biochem. 230,481-490 (1995) 0 FEBS 1995 Hybrid Bacillus amyloliquefaciens X Bacillus licheniformis a-amylases Construction, properties and sequence determinants Birgit CONRAD, Viet HOANG, Andreas POLLEY and Jiirgen HOFEMEISTER Institute of Plant Genetics and Crop Plant Research, Department of Molecular Genetics, Gatersleben, Germany (Received 23 September 1994/14 February 1995) - EJB 94 1455/2 A series of 33 single and mosaic hybrid a-amylases was constructed from the genes amyBA or amyLI, encoding the a-amylases from Bacillus amyloliquefaciens (AmyBA) and Bacillus lichenifomis (AmyLI). The hybrid proteins, consisting of the entire a-amylase sequence with a variable portion of AmyBA or AmyLI origin, were characterized in order to find enzymes with new properties (thermostability, temper- ature and pH optima, and substrate specificity), and to ldcalize the amino acid sequence regions responsi- ble for the changes. The thermostability of the AmyBAlAmyLI (AL-type) hybrid proteins correlated with the position and the length of the hybrid sequence. The hybrid enzymes fell into six groups retaining, in comparison to AmyBA, a certain value of the extra-thermostability of AmyLI or becoming more thermolabile than AmyBA. Four regions are proposed to contain thermostability determinants (TSDs). They map between amino acid residues 34-76,112-142, 174-179 and 263-276 of the respective hybrid enzymes, indicat- ing the dominance of the N-terminal half of AmyLI for these hybrid enzymes’ resistance against irrevers- ible inactivation. Two (TSD3 and TSD4) coincide with regions I and I1 that had already been suggested to stabilise AmyLI [Suzuki, Y., Ito, N., Yuuki, T., Yamagata, H. & Udake, S. (1989) J. Biol. Chern. 264, 18933 -18 9381. The temperature dependence of activity of the AL-type hybrid a-amylases was compared at pH 6.4 and pH 7.6 and the hybrid enzymes of one thermostability group were found to have similar temperature responses. A hybrid region between residues 34-76 is demonstrated to correlate with the a-amylases’ substrate specificity, i.e. either hydrolysis or accumulation of maltohexaose. This region was therefore named the G6G5 region. The exchange of internal sequences between residues 17-201 of AmyBA by the AmyLI counterpart in ALA-type mosaic hybrid a-amylases, with one exception (ALA99-429), unexpectedly destabilized the respective ALA-type hybrids. Two of these hybrid a-amylases (ALA17- 151 and ALA76-151) were less thermostable than AmyBA, while others (ALA112-151, ALA112-201) were enzymically inactive. These data support specific roles of the predicted A1-B domain portion between residues 17-201 of those Bacillus a-amylases probably for correct folding and enzymic activity. Keywords. a-amylase ; Bacillus ; hybrid enzymes ; protein mapping ; thermostability. A large spectrum of a-amylase genes from archaebacterial, eubacterial, fungal, animal and plant sources has been cloned and sequenced (Vihinen and Mantsala, 1989). They represent two independent gene families (Laderman et al., 1993). The largest family of a-amylases (EC 3.2.1.1) displays similarity with Taka-amylase A of AsperiZZus oryzae in their predicted triosephosphate isomerase @/a), barrel structure, as well as the pairs of amino acids within highly conserved sequence regions (Matsuura et al., 1984; Buisson et al., 1987a, b; MacGregor, 1987, 1988; MacGregor and Svensson, 1989; Jespersen et al., Correspondence to J. Hofemeister, Institut fur Pflanzengenetik und Fax: +49 039 482 280. Abbreviations. AmyBA, a-amylase of Bacillus arnyloliquefaciens; Amy’, phenotype of cellslcolonies exhibiting a-amylase activity or indi- cated by starch halo formation; Glc, and Glc,, maltopentaose and malto- hexaose; G6G5, sequence determinant correlating with the hybrid a- amylase specificity for maltohexaose; AmyLI, a-amylase of Bacillus li- clzeniformis; NpGlc,, p-nitrophenyl a-D-maltopentaoside ; NpGlc,, p-ni- trophenyl a-D-maltoheptaoside ; TSD, sequence determinant correlating with thermostability of the (hybrid) enzymes. Kulturpflanzenforschung, D-06466 Gatersleben, Germany Enzyme. a-Amylase (EC 3.2.1.1). 1991; Kim et al., 1992; Svensson and Sogaard, 1992; Sogaard et al., 1993). The a-amylases AmyBA and AmyLI (Talckinen et al., 1983; Yuuki et al., 1985; Gray et al., 1986) of Bacillus amyloliquefa- ciens and Bacillus licheniformis (Priest, 1991) are closely related and share a high degree of similarity even outside their con- served regions DVVXNH, GFRXDAXKH, AEYWQ, and FVXNHD (Janse et al., 1993), i.e. they exhibit only 91 amino acid substitutions out of 483 residues and two minor sequence gaps (Yuuki et al., 1985; Nakajima et al., 1986). Nevertheless, the enzymes differ with respect to their thermostabilities, tem- perature optima and substrate specificities (Fogarty, 1983 ; TO- mazic and Klibanov, 1988a,b). Various mutational approaches have been undertaken in order to determine amino acid residues responsible for the specificity and thermostability of Bacillus a- amylases (Smimova et al., 1987; Tomazic and Klibanov, 1988a,b; Suzuki et al., 1989; Holm et al., 1990; Vihinen et al., 1990; Svensson, 1991 ; Svensson and Sogaard, 1992; Nagas- hima et al., 1992; Takase et al., 1992). These have also been approached by the construction of hyperthermostable variants of AmyLI, thought to improve its biotechnological applications
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Hybrid Bacillus amyloliquefaciens X Bacillus licheniformisalpha-Amylases. Construction, Properties...

Eur. J. Biochem. 230,481-490 (1995) 0 FEBS 1995
Hybrid Bacillus amyloliquefaciens X Bacillus licheniformis a-amylases Construction, properties and sequence determinants
Birgit CONRAD, Viet HOANG, Andreas POLLEY and Jiirgen HOFEMEISTER
Institute of Plant Genetics and Crop Plant Research, Department of Molecular Genetics, Gatersleben, Germany
(Received 23 September 1994/14 February 1995) - EJB 94 1455/2
A series of 33 single and mosaic hybrid a-amylases was constructed from the genes amyBA or amyLI, encoding the a-amylases from Bacillus amyloliquefaciens (AmyBA) and Bacillus lichenifomis (AmyLI). The hybrid proteins, consisting of the entire a-amylase sequence with a variable portion of AmyBA or AmyLI origin, were characterized in order to find enzymes with new properties (thermostability, temper- ature and pH optima, and substrate specificity), and to ldcalize the amino acid sequence regions responsi- ble for the changes.
The thermostability of the AmyBAlAmyLI (AL-type) hybrid proteins correlated with the position and the length of the hybrid sequence. The hybrid enzymes fell into six groups retaining, in comparison to AmyBA, a certain value of the extra-thermostability of AmyLI or becoming more thermolabile than AmyBA. Four regions are proposed to contain thermostability determinants (TSDs). They map between amino acid residues 34-76,112-142, 174-179 and 263-276 of the respective hybrid enzymes, indicat- ing the dominance of the N-terminal half of AmyLI for these hybrid enzymes’ resistance against irrevers- ible inactivation. Two (TSD3 and TSD4) coincide with regions I and I1 that had already been suggested to stabilise AmyLI [Suzuki, Y., Ito, N., Yuuki, T., Yamagata, H. & Udake, S. (1989) J. Biol. Chern. 264, 18 933 -18 9381. The temperature dependence of activity of the AL-type hybrid a-amylases was compared at pH 6.4 and pH 7.6 and the hybrid enzymes of one thermostability group were found to have similar temperature responses.
A hybrid region between residues 34-76 is demonstrated to correlate with the a-amylases’ substrate specificity, i.e. either hydrolysis or accumulation of maltohexaose. This region was therefore named the G6G5 region. The exchange of internal sequences between residues 17-201 of AmyBA by the AmyLI counterpart in ALA-type mosaic hybrid a-amylases, with one exception (ALA99-429), unexpectedly destabilized the respective ALA-type hybrids. Two of these hybrid a-amylases (ALA17- 151 and ALA76-151) were less thermostable than AmyBA, while others (ALA112-151, ALA112-201) were enzymically inactive. These data support specific roles of the predicted A1-B domain portion between residues 17-201 of those Bacillus a-amylases probably for correct folding and enzymic activity.
Keywords. a-amylase ; Bacillus ; hybrid enzymes ; protein mapping ; thermostability.
A large spectrum of a-amylase genes from archaebacterial, eubacterial, fungal, animal and plant sources has been cloned and sequenced (Vihinen and Mantsala, 1989). They represent two independent gene families (Laderman et al., 1993). The largest family of a-amylases (EC 3.2.1.1) displays similarity with Taka-amylase A of AsperiZZus oryzae in their predicted triosephosphate isomerase @/a), barrel structure, as well as the pairs of amino acids within highly conserved sequence regions (Matsuura et al., 1984; Buisson et al., 1987a, b; MacGregor, 1987, 1988; MacGregor and Svensson, 1989; Jespersen et al.,
Correspondence to J. Hofemeister, Institut fur Pflanzengenetik und
Fax: +49 039 482 280. Abbreviations. AmyBA, a-amylase of Bacillus arnyloliquefaciens;
Amy’, phenotype of cellslcolonies exhibiting a-amylase activity or indi- cated by starch halo formation; Glc, and Glc,, maltopentaose and malto- hexaose; G6G5, sequence determinant correlating with the hybrid a- amylase specificity for maltohexaose; AmyLI, a-amylase of Bacillus li- clzeniformis; NpGlc,, p-nitrophenyl a-D-maltopentaoside ; NpGlc,, p-ni- trophenyl a-D-maltoheptaoside ; TSD, sequence determinant correlating with thermostability of the (hybrid) enzymes.
Kulturpflanzenforschung, D-06466 Gatersleben, Germany
Enzyme. a-Amylase (EC 3.2.1.1).
1991; Kim et al., 1992; Svensson and Sogaard, 1992; Sogaard et al., 1993).
The a-amylases AmyBA and AmyLI (Talckinen et al., 1983; Yuuki et al., 1985; Gray et al., 1986) of Bacillus amyloliquefa- ciens and Bacillus licheniformis (Priest, 1991) are closely related and share a high degree of similarity even outside their con- served regions DVVXNH, GFRXDAXKH, AEYWQ, and FVXNHD (Janse et al., 1993), i.e. they exhibit only 91 amino acid substitutions out of 483 residues and two minor sequence gaps (Yuuki et al., 1985; Nakajima et al., 1986). Nevertheless, the enzymes differ with respect to their thermostabilities, tem- perature optima and substrate specificities (Fogarty, 1983 ; TO- mazic and Klibanov, 1988 a,b). Various mutational approaches have been undertaken in order to determine amino acid residues responsible for the specificity and thermostability of Bacillus a- amylases (Smimova et al., 1987; Tomazic and Klibanov, 1988a,b; Suzuki et al., 1989; Holm et al., 1990; Vihinen et al., 1990; Svensson, 1991 ; Svensson and Sogaard, 1992; Nagas- hima et al., 1992; Takase et al., 1992). These have also been approached by the construction of hyperthermostable variants of AmyLI, thought to improve its biotechnological applications

482 Conrad et al. (Eur J. Biochem. 230)
Hae 111, 703
Hind 111, 1 ECOR v, 748 amyBA Mbo I, 904
Aha 111, 1105 Hind 111, 1484
AV8 11, 1945
ECOR I+ 2400
amyll
Pst I, 74 Aha 111, 544 HaeIII, 142 Ava II , 1378 EcoR I, 1781
Mbo I, 343
1 Psf I + EcoRl
Pst I + EcoRl pHPl3
/-y Ava II, 1305
lac Z .L
PL 7 6607 bp EccflI, 1707
1 Hind 111
Hind Ill r
Fig. 1. Construction of E. COW?. subtilis shuttle vector pL7 and pAL76. The PstI-EcoRI fragment with the amyLI gene (truncated at codon -4 of its signal peptide including the entire gene and its transcription terminator) was ligated into the multi-cloning site (MCS) of vector pHP13 (Haima et al., 1987) to give pL7. A HindIII fragment harbouring the entire 5’-portion of the amyBA gene (including the gene promoter, the open reading frame up to codon 297 of its mature amylase) was subsequently ligated into the HindIII-cut plasmid pL7 to give pAL7. In order to eliminate the restriction sites at the MCS in front of the tandem-gene cassette, the plasmid pAL7 was, after partial HindIII linearization, treated with exonuclease Bal31, ligated and recovered from transformants as derivative pAL76.
(Declerck et al., 1990; Joyet et al., 1992). The possibility to construct functional hybrids by in vivo recombination between the homologous a-amylases of Bacillus stearothermophihs, B. amyloliquefaciens or B. lichenifomis in order to create enzymes with new properties has also been demonstrated (Rey et al., 1986; Diderichsen et al., 1987). A similar hybridization ap- proach in combination with mutational studies led Suzuki et al. (1989) to discover sequence regions I and region I1 of AmyLI, which were proposed to enhance additively and independently the thermostability of AmyBA.
Here we report the construction and properties of a series of AL-type hybrid a-amylases which are composed of an AmyLI part that replaces the N-terminal AmyBA counterpart stepwise up to residue 276, as well as of mosaic hybrids with variable internal portions of either AmyLI or AmyBA, replacing different lengths of the respective amylase counterpart. Comparative in- spection of a selected 33 hybrids for activity, thermostability, temperature and pH optima, as well as the end products of starch hydrolysis, in relation to the position and length of the hybrid area led us to propose several sequence regions of the two en- zymes that influence those properties.
EXPERIMENTAL PROCEDURES Bacterial strains and culture media. The strains used were
Escherichia coli DHSa (Bethesda Research Laboratories, Gaith-
ersburg MD) and B. subtilis GSB26 aroI906 met85 sacA321 amyE str6 (a streptomycin-resistant derivative of strain QBll33 ; Steinmetz et al., 1976). The culture media were TBY, 2XTBY and NBY as described by Conrad et al. (1992). When required, media were supplemented with antibiotics as follows : ampicillin 100 pg/ml, chloramphenicol 10 pg/ml, erythromycin 50 pg/ml for E. coli; chloramphenicol 5 pg/ml and erythromycin 1 pg/ml for B. subtilis. The production of a-amylase by recombinant clones was identified on NBY agar plates containing 1% corn starch.
Transformation and DNA techniques. E. coli was trans- formed by the method of Hanahan (1983). Plasmid DNA trans- formation into B. subtilis competent cells was as described by Gryzcan et al. (1978) or into protoplasts according to the proto- col of Chang and Cohen (1979). Plasmid DNA was isolated from E. coEi and B. subtilis using standard methods (Sambrook et al., 1989) and purified using the Qiagen plasmid kit (Diagen GmbH Diisseldorf). Template DNA for sequencing was usually prepared from E. coli and processed according to the instructions for automated laser fluorescent DNA sequencer (Pharmacia Bio- systems GmbH) using an AutoRead T7 sequencing kit. Oligonu- cleotide primers for PCR or Fluoreprime-Amidite labelled prim- ers for sequencing were synthesized using a model 380B oligo- synthesizer (Applied Biosystems). PCR was performed using the bio-med Thermocycler 60 device (bio-med GmbH Theres, Ger- many) and standard protocols.

Conrad et al. (Eur: J. Biochem. 230) 483
Plasmids and hybrid gene construction. The plasmid vec- tors used were pUC18 (Yanisch-Perron et al., 1985), Bluescript pBS' (Stratagene), and the E. coZiB. subtilis shuttle vector pHP13 (Haima et al., 1987). The a-amylase gene amyLZ of B. lichenifomis strain ATCC 9789 (Ingle and Boyer, 1976; Morgan and Priest, 1981) on a 2.4-kb DNA insert was cloned using vec- tor pGB354 (Behnke et al., 1981) to construct the AmyLI ex- pression plasmid pSB3 (Steinborn et al., 1983).
The nucleotide sequence of this gene was found to be iden- tical with the sequence of homologous genes cloned from B. lichenifomis strains NCIB 8061 and ATCC 27 811 (Gray et al., 1986; Yuuki et al., 1985). The AmyBA expression plasmid pSB6 (Steinborn and Hofemeister, 1984) was the donor of the a-amylase gene amyBA of B. amyloliquefaciens strain ATCC 23 844 that was cloned using vector pGB354. The nucleotide se- quence of the AmyBA-encoding DNA was downstream from a 5'-terminal CZuI site identical with the published sequence of the a-amylase gene of B. amyloliquefuciens strain El8 (VTT 197; Takkinen et al., 1983). The two genes were used as donors and arranged on vector pHP13 to give the tandem-gene cassette of plasmid pAL7 as outlined in Fig. 1. Plasmid pAL76 was derived from pAL7 after limited Bul31 treatment of the linearized plas- mid DNA that was achieved by a partial HindIII digestion of pAL7 in order to eliminate restriction sites from one end of the tandem-gene cassette (Fig. 1). Due to the deletion of parts of each amylase gene, neither pAL7 nor pAL76 directed the syn- thesis of a-amylase in B. subtilis or in E. coli. These plasmid derivatives were used for the construction of hybrid amylase genes by homologous in vivo recombination (Weber and Weiss- mann, 1983), or in order to construct hybrids by the restriction fragment replacement technique using either existing or PCR- generated restriction sites (Higuchi, 1990), i.e. for HaeIII, EcoRV, MboI, AatII AhuIII, ClaI, AvaII as indicated in Fig. 2. The correctness of the sequence of the hybrid genes was subse- quently verified by automated nucleotide sequencing.
Amylase recovery and characterization. Amylase activity was estimated at 37 "C using an automated reading on microtiter plates. Samples of 10 pl each were mixed with 50 p1 amylase reagent from Sigma diagnostic kit (pH 7.0, procedure 577) into 96-well microplates with p-nitrophenyl a-D-maltoheptaoside (NpGlc,) as substrate. The microplate was incubated for either 5 or 10 min at 37°C. The reaction was stopped by the addition of 25 p1 1 M Na,CO, solution. The absorbance was measured at 405 nm with a Microplate Reader EL 312e (Bio-Tec-Instru- ments). An a-amylases solution containing 1 unit (U) catalyzes the hydrolysis of 1 pmol NpGlc,/min at 37°C.
Either wild-type or hybrid a-amylases were recovered from the supernatant fluid of B. subtilis GSB26 cultures harbouring the respective plasmid derivatives after 48 h of growth at 37°C in 2XTBY medium. For the heat treatment, cell-free (superna- tant) samples, containing about 0.8 - l .O U enzyme/ml,were in- cubated in a water bath, after the addition of 1 mM CaCI, but without added substrate, for an indicated period of time at given temperatures. The heat treatment was interrupted by placing ap- propriate samples on ice and the residual activity determined as described. Residual a-amylase activities were expressed as percentage activity of an untreated control sample. Each mea- surement was made at least three times.
The relative activities at different reaction temperatures at either pH 6.4 or 7.6 were determined on microtiter plates apply- ing 25 p1 of supernatant fractions with about 0.05 U enzyme in Sorensen phosphate buffer with 1 mM NpGlc, as substrate. The temperature was stepwise increased by 5 K. The desired temper- ature was reached after about 1 min of heating. The absorbance was subsequently measured in 15-s intervals 11 times using a Microplate Reader model 3550UV with a dual channel incubator
(BioRad). The absorbance increase at 405 nm was a measure of product formation.
The pH optimum of a-amylases was estimated in a total vol- ume of 0.1 ml containing 0.125 U enzyme in 0.1 mM buffer (piperazine dihydrochloride, glycylglycine and sodium hydrox- ide; Penin and Dempsey, 1974) at different pH values in steps of 0.2 in the range of 4.4-9.2. The substrate was soluble starch (0.5 mg/ml). The samples were loaded into microplate wells and incubated for 15 min at 37°C. After adding 1 vol. (0.1 ml) io- dine solution (0.5 g I,, 5 g KI in 1 M HCl), the residual starch concentration was assayed by a method modified from May and Elliot (1968) taking the absorbance at 590 nm.
HPLC analysis and paper chromatography. The products of the amylolytic degradation of oligomaltosides (Glc,, or Glc, ; Sigma) were analysed by paper chromatography on Whatman no. 1 paper (20x20 cm) and visualised as described by Robyt and French (1963). The a-amylase reaction products with solu- ble starch as substrate were analyzed with a Dionex HPLC sys- tem using the CarboPacTM PA1 column (P/N 35391). About 0.01 U enzyme in Sorensen phosphate buffer pH 7.0 containing 4 mg soluble starch in a total volume of 1 ml was incubated for about 3 h at 37°C in order to obtain the end products of the enzyme reaction. The starch concentration corresponds to 24.7 mM a-1 ,4-glycosidic bonds neglecting a-1,6-hranch points in the substrate molecules. After centrifugation, supernatant samples (100 pl) were filtered through a membrane (0.22 pm) and applied to the sample port. The end products (oligomalto- sides) were separated in a continuous gradient of sodium hy- droxide, sodium acetate and water using a pulsed electrochemi- cal detector (Dionex).
Polyacrylamide gel electrophoresis. Proteins of culture supernatants were separated by SDSPAGE using 4% stacking and 12% resolving gels of 2 mm thickness and stained with Coomassie blue dye as described by Laemmli (1970) using the rainbow protein markers (Amersham Life Science) as reference proteins.
RESULTS
Construction of hybrid amylase genes. The alignment of the nucleotide sequence of both amylase genes, umyBA and amyLI, coding for the mature enzyme proteins reveals about 71 % iden- tity and even a few restriction sites are conserved in both genes. We have thus taken advantage of predicted in vivo recombina- tion for generating single cross-overs (Weber and Weissmann, 1983), and made use of the existing or PCR-generated restriction sites (Higuchi, 1990) in both genes for restriction fragment re- placement in order to construct a series of hybrid amylase genes. The plasmid pAL76 (see Experimental Procedures, Fig. 1) was cut at the unique HindIII or PstI site and transformed into E. coZi DHSa(recA1) host cells. In parallel B. subtilis GSB26 com- petent cells were transformed with pAL7 DNA (without linear- ization). A series of transformants with Amy+ phenotype, i.e. starch halo formation, was screened for plasmid DNA, and the potential hybrid DNA was characterized by restriction mapping and nucleotide sequencing in order to test the template for cor- rectness and to localize the fusion region. The letters A and L in the hybrid names stand for the AmyBA and AmyLI portion, respectively, and the numbers for the amino acid residue(s) at the cross-over sites of the hybrid enzymes. The cross-overs were located within a stretch of 6-14 identical nucleotides, except for constructs AL167 and AL169, where this region consisted of 3 and 5 identical nucleotides in both genes, respectively. The outermostcross-over regions were found between bp228 (hybrid AL76) and bp828 (hybrid AL276), counting the nucleotides

484 Conrad et al. (Em J. Biochem. 230)
A
L
AL17
AL34
AL76
AL108
AL112
AL142
AL147
AL149
AL151
AL163
AL174
ALl79
AL185
ALl91
AL198
AL207
AL231
AL234
AL244
AL256
AL263
AL276
AL431
ALA17-151
ALA34-84
ALA76-151
ALA99-429
ALAl12-151
ALA112-201
LA1 9
LA43 1
LALl9-433
LAL19-153
1 A9
1 A7
l R 1
4 76
974
344
19 . 153
Stability group
V
I
I
I
I1
II
II
111
111
111
Ill
111
111
IV
nd
nd
nd
nd
IV
nd
nd
IV
IV
V
V
VI
V
vi I
nd
nd
V
I
V
111
G6G5 character
A
L
L
L
A
A
A
A
A
A
A
nd
A
A
nd
nd
nd
nd
A
nd
nd
A
A
A
A
L
L
A
A
nd
nd
A
L
A
A
Fig. 2. Compilation of a series of hybrid a-amylases of types AL, ALA, LA and LAL constructed in these studies. The letter A stands for the sequence derived from AmyBA = closed bars; the letter L stands for the sequence derived from AmyLI = dotted bars). The amino acid residue at the recombinatiodfusion sites is given for each hybrid. The vertical lines indicate the predicted limits of the domain regions A,, B, A, and C of Bacillus a-amylases as defined by MacGregor (1988). The restriction map of the DNA fragments encoding AmyBA (A) or AmyLI (L) used for construction of hybrids are shown in the upper part. The restriction sites newly introduced into the gene fragments by site-directed mutagenesis are labelled by stars (*I; nd in the column stands for not determined.

Conrad et al. (Eur: J. Biachem. 230) 485
S D D E l l Ill 1-1 V I m
Fig. 3. Thermostability of 24 hybrid a-amylases in relation to the parental enzymes AmyLI (L) and AmyBA (A). Thermostability is measured as the percentage of residual activity at 37°C after 90°C heat treatment for either 15 min (dark columns) or 30 min (dotted columns). The standard deviations from six determinations are included. The relative activites of the hybrid a-amylases before heat treatment were in the range 0.8-1.0 U/ ml. The response groups (I-VI) are indicated as described in the text. The inset shows the time course of thennoinactivation of one representative a-amylase selected from each response group at 90°C. The curves illustrate at which inactivation level after 15 min or 30 min of the heat treatment the values for the graph were taken. (v) L, AmyLI; (0) LA431, group I; (+) AL108, group 11; (X) AL1.51, group 111; (A) AL263, group IV; (0) LA19, group V; (m) ALA76-151, group VI; (0) A, AmyBA.
Table 1. Properties (substrate specificity, thermostability) of mosaic hybrid amylases related to the exchanged sequence regions. The TSD regions and response groups are defined according to Fig. 3. The sugar composition of the digest with soluble starch as substrate was determined by HPLC analysis as described in Experimental Procedures. Data are from a representative experiment showing the mean and standard error of triplicate determinations. The data are representative of two experiments. The percentage of glucose or maltooligosaccharides in the digest was calculated from the peak area relative to the total of the peak areas formed by the electrochemical potential plotted against the retention time; n = not tested.
Hybrid AmyLI sequence determinants sequence region TSDU TSD2 TSD3 TSD4 TS re- Percentage of sugars in the digest
G6G5 114-144 176-178 263-276 sponse 36-78 group G1 G2 G3 G4 G.5 G6 G7
LAL19- 153 - - + + I11 5 20 28 11 20 17 0 LAL19 -453 - - - - V 10 17 22 14 16 18 3 ALA17 - 15 1 + + - - VI 17 27 17 22 17 1 1 ALA76 - 15 1 - + - - VI 10 24 15 16 24 11 0 ALA34 - 84 + - - - V 12 23 16 23 20 1 0 ALAY9-429 - + + + I” 7 35 9 16 20 14 1 ALA112-201 - + + - -b n n n n n n n * ALA112-151 AmyBA - - - - V 6 21 24 14 30 19 0 ArnyLI + f + + I 13 18 23 1.5 17 0 0
- - - - - - - - _ c - - +
a The tbermoinactivation of this mosaic hybrid was subsequently tested at 93 “C and found to have about 40% residual activity after 30 min, like AmyLI.
No amylase activity in B. subtilis, but amylase activity in E. coli. No amylase activity in B. subtilis nor in E. coli.
from codon 1 of the gene encoding the mature ArnyBA amylase. The most N-terminal hybrids AL17, LA19, and AL34, as well as the hybrids AL151, AL431 and LA431, were subsequently constructed in vitro by the restriction fragment replacement
strategy, i.e. using either the Had11 site at bpS1 of amyBA (bpS7 of arnyLI), the EcoRV site at bpl02 of amyBA (or newly intro- duced at bp108 of amyLI), the Aha111 site at bp453 (of arnyLI) or the conserved AvaII sites (Fig. 2). The single cross-over hy-
486 Conrad et al. (Euz J. Biochem. 230)
brid genes were subsequently used to construct a spectrum of mosaic hybrids in combination with restriction sites shown in Fig. 2. They contain a distinct hybrid sequence replacing the counterpart of a partner gene that therefore encodes a sequence mosaic; these are LAL19-433 (using HaeIII and AvaII), LAL19-153 (using Hue111 and AhaIII), ALA112-201 (using AL112 and ClaI), ALA112-151 (using AL112 and AhaIII), ALA99-429 (using AatII and AvaII), ALA76-151 (using AL76 and AhaIII), ALA34-84 (using hybrid AL34 and MboI), ALA17-151 (using Hue111 and AhaIII). A scheme of the struc- ture of the 33 hybrid a-amylases that were further investigated is shown in Fig. 2.
Hybrid gene expression in E. coli and B. subtilis. With the exception of the mosaic hybrids ALA112-151 and ALA112- 201, all other hybrid genes constructed and illustrated in Fig. 2 showed starch halo formation in E. coli as well as in B. subtilis host cells. The mosaic hybrid ALA112-151, however, did not express a functional amylase, either in E. coli or in B. subtilis. This deficiency was either partly or entirely restored in hybrids ALA112-201 and ALA76-151. The hybrid ALA112-201 with 50 amino acids extending the replacement region towards the C-terminus showed amylase activities in E. coli but not in B. subtilis, while the hybrid ALA76-151 with 37 amino acids extending the replacement region towards the N-terminus ex- pressed an enzymically active (but thermolabile) hybrid a-amy- lase in both host organisms (Fig. 3, Table 1).
The hybrid a-amylases in the supernatant of a selected spectrum of B. subtilis cultures were compared by SDSPAGE and found to have about the same mobility and quantity as the parental enzymes (data not shown).
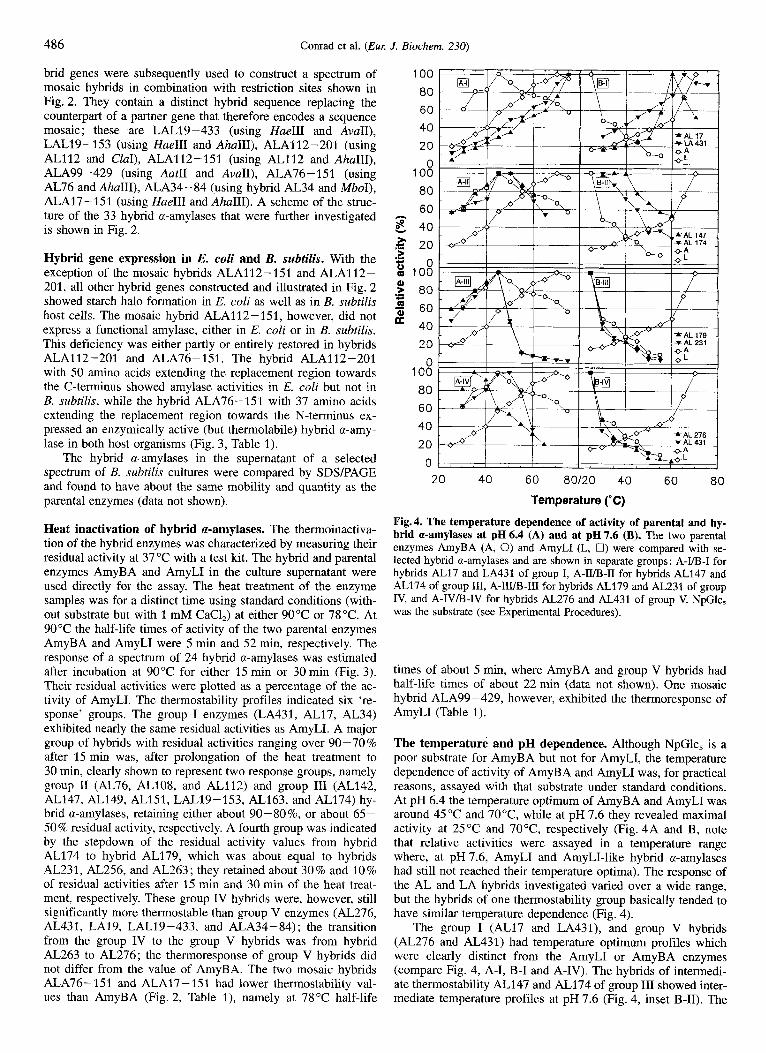
Heat inactivation of hybrid a-amylases. The thermoinactiva- tion of the hybrid enzymes was characterized by measuring their residual activity at 37°C with a test kit. The hybrid and parental enzymes AmyBA and AmyLI in the culture supernatant were used directly for the assay. The heat treatment of the enzyme samples was for a distinct time using standard conditions (with- out substrate but with 1 mM CaCl,) at either 90°C or 78°C. At 90°C the half-life times of activity of the two parental enzymes AmyBA and AmyLI were 5 rnin and 52 min, respectively. The response of a spectrum of 24 hybrid a-amylases was estimated after incubation at 90°C for either 15 rnin or 30 min (Fig. 3). Their residual activities were plotted as a percentage of the ac- tivity of AmyLI. The thermostability profiles indicated six 're- sponse' groups. The group I enzymes (LA431, AL17, AL34) exhibited nearly the same residual activities as AmyLI. A major group of hybrids with residual activities ranging over 90-70% after 15 min was, after prolongation of the heat treatment to 30 min, clearly shown to represent two response groups, namely group I1 (AL76, AL108, and AL112) and group I11 (AL142, AL147, AL149, AL151, LALl9-153, AL163, and AL174) hy- brid a-amylases, retaining either about 90-80%, or about 65- 50 % residual activity, respectively. A fourth group was indicated by the stepdown of the residual activity values from hybrid AL174 to hybrid AL179, which was about equal to hybrids AL231, AL256, and AL263 ; they retained about 30 % and 10 % of residual activities after 15 rnin and 30 min of the heat treat- ment, respectively. These group IV hybrids were, however, still significantly more thermostable than group V enzymes (AL276, AL431, LA19, LAL19-433, and ALA34-84); the transition from the group IV to the group V hybrids was from hybrid AL263 to AL276; the thermoresponse of group V hybrids did not differ from the value of AmyBA. The two mosaic hybrids ALA76-151 and ALA17-151 had lower thermostability val- ues than AmyBA (Fig. 2, Table I), namely at 78°C half-life
100
80 60
40 20
0 100
80 60
8 40
>
h
E 20
: 100 .- $! 80
.- - 0
CI = 60 ' 40 20
0 100
80 60
40 20
0
20 40 60 80120 40 60 80
Temperature ('C)
Fig. 4. The temperature dependence of activity of parental and hy- brid a-amylases at pH6.4 (A) and at pH7.6 (B). The two parental enzymes AmyBA (A, 0) and AmyLI (L, 0) were compared with se- lected hybrid a-amylases and are shown in separate groups: A-VB-I for hybrids AL17 and LA431 of group I, A-II/B-II for hybrids AL147 and AL174 of group 111, A-III/B-III for hybrids AL179 and AL231 of group IV, and A-IVB-IV for hybrids AL276 and AL431 of group V. NpGlc, was the substrate (see Experimental Procedures).
times of about 5 min, where AmyBA and group V hybrids had half-life times of about 22 min (data not shown). One mosaic hybrid ALA99-429, however, exhibited the thermoresponse of AmyLI (Table 1).
The temperature and pH dependence. Although NpGlc, is a poor substrate for AmyBA but not for AmyLI, the temperature dependence of activity of AmyBA and AmyLI was, for practical reasons, assayed with that substrate under standard conditions. At pH 6.4 the temperature optimum of AmyBA and AmyLI was around 45 "C and 70"C, while at pH 7.6 they revealed maximal activity at 25°C and 70"C, respectively (Fig. 4A and B, note that relative activities were assayed in a temperature range where, at pH 7.6, AmyLI and AmyLI-like hybrid a-amylases had still not reached their temperature optima). The response of the AL and LA hybrids investigated varied over a wide range, but the hybrids of one thermostability group basically tended to have similar temperature dependence (Fig. 4).
The group I (AL17 and LA431), and group V hybrids (AL276 and AL431) had temperature optimum profiles which were clearly distinct from the AmyLI or AmyBA enzymes (compare Fig. 4, A-I, B-I and A-IV). The hybrids of intermedi- ate thermostability AL147 and AL174 of group In showed inter- mediate temperature profiles at pH 7.6 (Fig. 4, inset B-11). The

Conrad et al. (Eul: J. Biochem. 230) 487
$AL17, LA19 L
AL34 ANLNGTLMQYFEWmPNDGQHLQNDSAY~~GITAVWIPPAYKGTSQADVGYGAYDLYDLGEFHQKGT 72
AL147d 4AL151 JAL163 L GRGSTYSDFKWHWYHFDGTDWDESRKLNRI A ... N..............A.......IS.. ........
--I------ TSD3
34 JAL244 L WGTWYANELQL KFSFLRDWVNHVREKTGKE ...... A .. I......S. .........Q A..QA..
L PLHYQFHAASTQGGGYDMRKLLNGTVVSKHPLKS QPGQSLESTVQTWFKPLAYAFILTRESGY 358 A ... FNLQ ... S........R..D.....E.A..E.A.. .............................. 358
>>- - - - - - - -- -----I---”----- >>>>>>>>>>>> -c--------
L P Q V F Y G D M Y G T K G D S Q R E I P A L K H K I E P I L K A R K Q Y A Y G A Q ~ ~ ~ ~ ~ ~ ~ D ~ V ~ ~ ~ ~ ~ ~ D S S V ~ S ~ ~ ~ ~ D 430 A ............. T.PK...S..DN.........E....P....~..P.V~.........A.K ......... 430
>>>>>>>>>>>> --------------- LAL431, LA431
L GPGGAKRMWGRQNAGETWHDITGNRSEPWINSEGWGEFKVGSVSI~QR 483
Fig. 5. Alignment of the (mature) a-amylases AmyLI (L) and AmyBA (A). Amino acids are shown in the single-letter code. Gaps are indicated by dashes. The extension of the sequence areas suggested to contain a distinct thermoresponse (TSD) or substrate specificity determinant (G6G5) is marked by boxes. The positions of the fusion site of the respective (single cross-over) hybrid a-amylases are indicated by an arrow L and the abbreviations for the hybrids are as used in Fig. 2. The predicted limits of the triosephosphate isomerase @/a),-barrel domains A1 and A2 (black letters) divided by domain B and the following domain C as calculated for AmyBA by MacGregor (1988) are positioned according to Declerck et al. (1990). The conserved regions found in a-amylases of various origin (Vihinen et al., 1990; Janse et a]., 1993) are shown in boxes. The amino acid positions Hisl33Tyr and Ala209Val as discussed by Joyet et al. (1992) are boxed and labelled by asterisks.
A .... S . . . . A . Z K . . . . . . Y . . . . . . , D T . R . G . D . , . . . . . . . . D . . . . . . . . K 483
response of hybrid AL276 was exceptional, as it clearly deviated at pH 6.4 from AmyBA in reaching highest activity at lower temperatures (see Fig. 4, A-IV).
We did not expect the hybrid a-amylases to differ much in their pH optima (at 37OC) from the two parental enzymes, since even the parental enzymes AmyBA and AmyLI have quite simi- lar and broad maxima between pH 6.4 and 7.6 (data not shown). The following hybrid a-amylases were, however, found to have exceptionally high activities at low pH (4.5): LA431, 15 %; ALA134-84, 60%; ALA17-151, 85%; LA19, 95% (relative to 100% activity at their pH optimum, data not shown).
Hydrolysis of starch and of oligosaccharides. The accumula- tion of Glc, among the hydrolysis products of soluble starch is one of the characteristics of the AmyBA enzyme, while AmyLI belongs in this respect to the a-amylases that hydrolyse Glc, (Fogarty, 1983; Fogarty et al., 1991). We made use of that character in order to compare the hybrid enzymes for either Glc, hydrolysis or accumulation. With maltohexaose as substrate and after paper chromatographic separation of the digest, the hybrids AL76, AL142, AL276 and LAL19-153 reacted like AmyBA.
They did not hydrolyse the Glc, substrate, while the hybrids AL17 and AL34 revealed a reaction pattern similar to that of AmyLI (data not shown). HPLC analysis of the products in the starch digest was subsequently made for a spectrum of single and mosaic hybrid a-amylases. The character Glc, accumulation/ Glc, formation of 19 hybrids changed only once, i.e. with the exchange of a sequence between amino acids 34-76 (data sum- marized in Fig. 2). This region overlaps with region TSDl (see above) and was therefore named the TSDUG6G5 region (Fig. 5). Analysis of the mosaic hybrid a-amylases confirmed the correlation between the substrate specificity and the origin of the TSDUG6G5 region (Table 1).
DISCUSSION Thirty-three single or mosaic hybrids of the two Bacillus
a-amylases were selected in an attempt to construct enzymes with changed properties and to identify as yet unknown se- quence regions that correlate with changed enzyme properties, e.g. their thermostability and substrate specificity. In a similar (hybridisation) approach, Suzuki et al. (1989) proposed two

488 Conrad et al. ( E m J. Biockem. 230)
regions of AmyLI to stabilise the enzyme against thermal inacti- vation.
The serial construction of the AL-type hybrids allowed us to map four TSD regions from AmyLI which correlated with the thermostability and one region (G6G5) that affects the substrate specificity of hybrid a-amylases. The residual activity was used to evaluate the efficacy of the TSD regions. Two of the determi- nants, our regions TSD3 and TSD4, overlap precisely with re- gions I and I1 that were already described by Suzuki et al. (1989). Their presence in the hybrid proteins of group 111 (TSD3 and TSD4) and group IV (TSD4) correlated after 15 min of the heat inactivation with residual activity values of about 80% and 30%, respectively (Fig. 3). A major group of hybrids with resid- ual activities ranging over 90-70% after 15 min was, after pro- longation of the heat treatment up to 30 min, clearly shown to fall into two separate response groups (Fig. 3). The residual ac- tivity values of the group I1 (AL76, AL108, ALll2) and group 111 hybrids (AL142, AL147, AL149, AL151, AL163, and AL174) ranged over 86-82% and 66-52%, respectively, and differed clearly by 20-30%. The loss of residual activity corre- lated with the loss of a sequence from either residues 34-76, or residues 112- 142, which we consequently proposed to repre- sent regions TSDl and TSD2. This assumption was confirmed by the group 111 character of the LAL-type (mosaic) hybrid LAL19-153. The loss of the regions TSDl and TSD2 from AmyLI correlated with a pronounced decrease (to about 66%) of the hybrid’s residual activity (Fig. 3). This correlation led Su- zuki et al. (1989) to conclude that regions I and I1 (TSD3 and TSD4 in our studies) affect the thermostability of the hybrid enzymes independently and additively.
We then faced, however, unexpected problems in confirming these correlations after construction of a spectrum of ALA-type (mosaic) hybrid a-amylases (ALA34-84, ALA17-152, ALA76-151, ALA99-429, ALA112-151, ALA112-201; see Fig. 2). They were expected to become, in a sequence mosaic, stabilised in comparison to AmyBA by the presence of AmyLI regions (TSD1, TSD2 and/or TSD3, respectively). With only one exception (ALA99-429), the stability values were, how- ever, in comparison to AmyBA, either not enhanced (ALA34- 84), or diminished (ALA17-151, ALA76-151) (Fig. 3). The exception (ALA99-429) contained only three, but the most effi- cient, TSD regions (2, 3 and 4) of AmyLI and had similar ther- mostability to AmyLI (data not shown). In contrast, two other mosaic a-amylases (ALA112-151, ALAl12-201) exhibited no activity in B. subtilis. These detrimental effects of AmyLl se- quences residues 17-201 in an AmyBA sequence mosaic might reflect specificities of these protein portions. In view of the pre- dicted triosephosphate isomerase barrel secondary structure of these Bacillus a-amylases (MacGregor, 1988 ; MacGregor and Svensson, 1989; Holm et al., 1990; Vihinen et al., 1990), these regions of the two Bacillus a-amylases cover parts of the pre- dicted domain A1 and nearly the entire domain B. The latter forms an extremely long loop between the predicted third (3- strand and third a-helix (MacGregor, 1987; Fig. 5). This loop is believed to stabilise the enzyme by binding a Caz+ ion (Mat- suura et al., 1984; Buisson et al., 1987; Boel et al., 1990). The relatively high frequency of amino acid substitutions, as well as mutational studies within this part of the AmyLI sequence, also suggested that this region affects the foldinghnfolding process and was therefore predicted to be a ‘weak part of the protein’ (Joyet et al., 1992). The properties of our ALA-type mosaic hy- brid a-amylases strengthen these conclusions.
From previous mutational studies with AmyLI, a Hisl334Tyr substitution was shown to further stabilise AmyLI (Declerck et al., 1990) and to give, in combination with a second amino acid substitution, Ala209hVal hyper-thermostable
AmyLI enzyme variants (Joyet et al., 1992). Both substitutions are originally present at the same positions in AmyBA (Fig. 5). In the AL-type hybrids of our series beginning with AL142, the replacement of N-terminal portions of AmyLI by the AmyBA counterpart did not, however, stabilise the respective hybrid a- amylases but resulted in hybrids of lower thermostability. This indicated that the Hisl334Tyr substitution in the context of re- spective AL-type and mosaic hybrids apparently becomes sup- pressed by the loss of the AmyLI TSD regions. We have not, however, tested the effect of an additional Ala209-Val substitu- tion in the respective hybrids.
We have now demonstrated an all-or-nothing change of the substrate specificity of hybrid a-amylases for maltohexaose to correlate with the exchange of a N-terminal region ranging over residues 34-74. Transfer of this region from AmyLI into AmyBA, or vice versa from AmyBA into AmyLI, generated hybrids ALA34- 84 and ALA99 -429, respectively. Their ther- mostability was nearly equal to either-AmyBA or AmyLI, but the content of the sugars (Glc, and Glc,) in the digest was clearly changed, i.e. to either that of AmyLI or AmyBA (Table 1). This region was therefore named the G6G5 region. The altered speci- ficity for maltohexaose correlated with the change of five amino acid residues, including the substitutions Ala56+Pro or His66+Gln in the predicted loop 2 of domain A1 . This loop was supposed to constitute sections of subsites A and/or B within the active site of AmyBA (MacGregor, 1987, 1988). Our finding in connection with the proposed function(s) of the predicted do- main A1 loop regions strengthens a distinct contribution of that protein area to the enzyme’s specificity for maltohexaose. In this concern it was also of interest that the length and folding of the eight loops of the predicted domain A had already been sug- gested to affect the substrate specificity of a-amylases (Matsuura et al., 1984; Buisson et al., 1987a,b).
The specific roles of the proposed sequence region(s) remain speculative until more is known about their contribution to the tertiary structure of these Bacillus a-amylases (Suzuki et al., 1990; Lee et al., 1991; SBgaard et al., 1993), but they could now be targeted for mutational and physicochemical studies.
The authors acknowledge the help of Susanne Konig in sequencing the hybrid genes, Ingo Wilke for the introduction to HPLC analysis and Thomas Walther for initial experiments. Manfred Klaas and Julian Phil- lips helped to improve the manuscript. We thank Heike Siedel and Silke Gorgulla for technical assistance. This studies were supported by the Forsckungskreis der Ernakrungsindustrie e. \.I (Bonn), the Arbeits- gemeinsckaft industrieller Forschungsvereinigungen, and the Ministry of Economics under project AIF-FV 101 D.
REFERENCES Behnke, D., Gilmore, M. & Ferretti, J. J. (1981) Plasmid pGB301, a
new multiple resistance streptococcal vehicle and its use in cloning of a gentamycinkanamycin resistance determinant, MoE. Gen. Genet. 182,414-421.
Boel, E., Brady, L., Brzozowski, A.M., Derewenda, Z., Dodson, G. G., Jemsen, V. J., Petersen, S. B., Swift, H., Thim, L. & Woldlike, H. F. (1990) $Xcium binding in a-amylases: An X-ray diffraction study at 2.1-A resolution of two enzymes from Aspergillus, Biochemistry 29, 6244-6249.
Buisson, G. I., Duke, E., Payan, F. & Haser, R. (1987a) Alpha-amylase tertiary structures and their interaction with polysaccharides, Food Hydrocolloids 516, 399 -406.
Buisson, G., Duke, E., Haser, R. & Payan, F. (1987b) Three dimensional structure of porcine pancreatic a-amylase at 2.9 A resolution: role of calcium in structure and activity, EMBO J. 13, 3909-3916.

Conrad et al. (Eur J. Biockem. 230) 489
Chang, S. & Cohen, S. N. (1979) High frequency transformation of Ba- cillus subtilis protoplasts by plasmid DNA, Mol. Gen. Genet. 168,
Conrad, B., Bashkirov, V. & Hofemeister, J. (1992) Imprecise excision of plasmid pE194 from the chromosomes of Bacillus subtilis pE194 insertion strains, J. Bacteriol. 174, 6997-7002.
Declerck, N., Joyet, P., Gaillardin, C. & Masson, J.-M. (1990) Use of amber supressors to investigate the thermostability of Bacillus li- cheniformis a-amylase, J. Biol. Chem. 265, 15481 -15 488.
Diderichsen, B., Outtrup, K. H., Schiilein, M. & Norman, B. E. (1987) Chimeric enzymes, European patent EP-87305781.
Fogarty, W. M. (1983) Microbial amylases, in Microbial enzymes and biotecknology (Fogarty, W. M., ed.) pp. 1-92, Applied Science Pub- lishers Ltd, London.
Fogarty, W. M., Bealin-Kelly, F., Kelly, C. T. & Doyle, E. M. (1991) A novel maltohexaose-forming a-amylase from Bacillus caldovelox: pattern and mechanisms of action, Appl. Micmbiol. Biotecknol. 36,
Gray, G. L., Mainzer, S. E., Rey, M. W., Lamsa, M. H., Kindle, K. L., Carmona, C. & Requadt, C. (1986) Structural genes encoding the thermophilic a-amylases of Bacillus stearothermophilus and Bacillus licheniformis, J. Bacteriol. 166, 635-643.
Gryzcan, T. J., Contente, S. & Dubnau, D. (1978) Characterization of Stapkylococcus aureus plasmids introduced by transformation into Bacillus subtilis, J. Bacteriol. 134, 318-329.
Haima, P., Bron, S. & Venema, G. (1987) The effect of restriction on shotgun cloning and plasmid stability in Bacillus subfilis Marburg, Mol. Gen. Genet. 209, 335-342.
Hanahan, D. (1983) Studies on transformation of Esckerickia coli with plasmids, J . Mol. Biol. 166, 557-580.
Higuchi, R. (1990) Recombinant PCR in PCR protocols: A guide to methods and applications (Innis, M. A., Gelfand, D. H., Sninsky, J. J. & White, T. J., eds) pp. 177-183, Academic Press, San Diego.
Holm, L., Koivula, A. K., Lehtovaara, P. M., Hemminh, A. & Knowles, J. K. C. (1990) Random mutagenesis used to probe the structure and function of Bacillus steamtkemopkilus a-amylase, Protein Eng. 3,
Ingle, M. B. & Boyer, E. W. (1976) Production of industrial enzymes, in Microbiology (Schlessinger, D., ed.) pp. 420-426, American So- ciety for Microbiology, Washington DC.
Janse, B. J. H., Stein, A. J. C. & Pretorius, I. S. (1993) Regional se- quence homologies in strach-degrading enzymes, Curr Genet. 24,
Jespersen, H. M., MacGregor E. A., Sierks, M. R. & Svensson, B. (1991) Comparison of the domain-level organization of starch hydrolases and related enzymes, Biochem. J. 280, 51-55.
Joyet, P., Declerck, N. & Gaillardin, C. (1992) Hyperthermostable vari- ants of a highly thermostable a-amylase, Bioflecknology 10, 1579- 1583.
h m , I.-C., Cha, J.-H., Kim, J.-R., Jang, S.-Y., Seo, B.-C., Cheong, T.- K., Lee, D. S., Choi, Y.-D. & Park, K.-H. (1992) Catalytic properties of the cloned amylase from Bacillus lickeniformis, J. Biol. Ckem.
Laemmli, U. K. (1970) Cleavage of structural proteins during the assem- bly of the head of bacteriophage T4, Nature 227, 680-685.
Laderman, K. A,, Asada, K., Uemori, T., Mukai, H., Taguchi, Y., Kato, I. & Anfinsen, C. B. (1993) a-Amylase from the hypertherrnophilic archaebacterium Pyrococcus furiosus, J. Biol. Chem. 268, 24 402- 24407.
Lee, S. Y., Kim, S., Sweet, R. M. & Suh, S. W. (1991) Crystallization and a preliminary X-ray crystallographic study of a-amylase from Bacillus lickeniformis, Arch. Biochem. Biophys. 291, 255 -257.
MacGregor, E. A. (1987) The relationship between structure and speci- ficity of a-amylases, Food Hydrocolloids 1, 407-41 1 .
MacGregor, E. A. (1988) a-Amylase structure and activity, J . Protein Chem. 7 , 399-415.
MacGregor, E. A. & Svensson, B. (1989) A super-secondary structure predicted to be common to several a-I ,4-~-glucan-cleaving enzymes, Biochem. J. 259, 145-152.
Matsuura, Y., Kusunoki, M., Harada, W. & Kakudo, M. (1984) Structure and possible catalytic residues of Taka-amylase A, J. Biockem. (Tokyo) 95, 697-702.
111 -115.
184-189.
181-191.
400-407.
267, 22 108-22 114.
May, B. K. & Elliott, W. H. (1968) Selective inhibition of extracellular enzyme synthesis by removal of cell wall from Bacillus subtilis, Bio- chim. Biopkys. Acta 166, 532-537.
Morgan, F. J. & Priest, F. G. (1981) Characterization of a thermostable a-amylase from Bacillus licheniformis NCIB 6346, J. Appl. Bacte- rial. 50, 107-114.
Nagashima, T., Tada, S., Kitamoto, K., Gomi, K., Kumagai, C. & Toda, S. (1992) Site-directed mutagenesis of catalytic active-site residues of Taka-amylase A, Biosci. Biotech. Biochem. 56, 207-210.
Nakajima, R., Imanaka, T. & Alba, S. (1986) Comparison of amino acid sequences of eleven different a-amylases, Appl. Microbiol. Bioteck- nol. 23, 355-360.
Pemn, D. D. & Dempsey, B. (1974) BufSersfor pH and metal ion con- trol, pp. 154, Chapman and Hall, London.
Priest, F. G. (1991) Isolation and identification of aerobic endospore- forming bacteria, in Bacillus, Biotechnology handbooks (Harwood, C. R., ed.) vol. 2, pp. 27-56, Plenum Press, New York and London.
Rey, M. W., Requadt, C., Mainzer, S. E., Lamsa, M. H., Ferrari, E., Lad, P. J. & Gray, L. G. (1986) Homologous a-amylases of Bacillus and generation of their hybrids in vivo, in Bacillus molecular genetic and biotechnology applications (Ganesan, A. T. & Hoch, J. T., eds) pp. 229-239, Academic Press, London.
Robyt, J. & French, D. (1963) Action pattern and specificity of an amy- lase from Bacillus subtilis, Archiv. Biockem. Biophys. 100, 451 - 467.
Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989) Molecular cloning: a laboratory manual, 2nd edn, Cold Spring Harbor Laboratory Press, Cold Spring Harbor NY.
Smimova, N. A,, Sorokin, A. V., Laptev, D. A,, Veiko, V. P. & Kozlov, Y. I. (1987) Two-codon mutagenesis of Bacillus amyloliquefaciens a-amylase gene, Mol. Biol. 22, 1257-1211.
SGgaard, M., Abe, J., Martin-Eauclaire, M. F. & Svensson, B. (1993) a- Amylases: structure and function, Carbohydrate Polymers 21, 137- 146.
Steinborn, G., Hofemeister, J., Bomss, B. & Behnke, D. (1983) Verfah- ren zur Herstellung von hitzestabiler Alpha-Amylase, GDR patent application, DD A5 281 414, 07.83.
Steinborn, G. & Hofemeister, J. (1984) Verfahren zur Herstellung von alpha-Amylase, GDR patent application, DD B5 233 852, 03.10.84.
Steinmetz, M., Kunst, F. & Dedonder, R. (1976) Mapping of mutations affecting synthesis of extracellular enzymes in Bacillus, Identity of sacU, amyB and pap mutations, Mol. Gen. Genet. 148, 281- 285.
Svensson, B. (1991) Structurelfunction relationships in starch-hydrolases and related enzymes, J. Jpn. SOC. Starch Sci. 38, 125-135.
Svensson, B. & Segaard, M. (1992) Protein engineering of amylases, Biockem. SOC. Trans. 20, 34-41.
Suzuki, Y., Ito, N., Yuuki, T., Yamagata, H. & Udaka, S. (1989) Amino acid residues stabilizing a Bacillus a-amylase against irreversible thermoinactivation, J. Biol. Chem. 264, 18933-18938.
Suzuki, A., Yamane, T., Ito, T., Nishio, T., Fujiwara, H. & Ashida, T. (1990) Crystallization and preliminary crystallographic study of bac- terial a-amylases, J. Biochem. (Tokyo) 108, 379-381.
Takase, K., Matsumoto, T., Mizuno, H. & Yamane, K. (1992) Site-di- rected mutagenesis of active site residues in Bacillus subrilis a-amy- lase, Biochim. Biupkys. Acta 1120, 281 -288.
Takkinen, K., Petterson, R. F., Kaikkinen, N., Palva, I., Soderlund, H. & K a ~ a i n e n , L. (1983) Amino acid sequence of a-amylase from Bacil- lus amyloliquefaciens deduced from the nucleotide sequence of the cloned gene, J. Biol. Ckem. 258, 1007-1013.
Tomazic, S. J. & Klibanov, A. M. (1988a) Mechanisms of irreversible thermal inactivation of Bacillus a-amylases, J. Biol. Chem. 263,
Tomazic, S. J. & Klibanov, A. M. (1988b) Why one Bacillus amylase is more resistant against irreversible thermoinactivation than an- other? J. Biol. Chem. 263, 3092-3096.
Vihinen, M. & Mantsala, P. (1989) Microbial amylolytic enzymes, Crit. Rev. Biockem. Mol. Biol. 24, 329-418.
Vihinen, M., Ollikka, P, Niskanen, J., Meyer, P., Suominen, I., Karp, M., Holm, L., Knowles, J. & Mantsala, P. (1990) Site-directed muta- genesis of a thermostable a-amylase from Bacillus stearothermophi-
3086-3091.

490 Conrad et al. (Eul: J. Biochem. 230)
lus: putative role of three conserved residues, J . Biochem. (Tokyo) 107,267-212.
Weber, H. & Weissmann, c . (1983) Fornation of genes coding for hy- brid proteins by recombination between related, cloned genes in E. coli, Nucleic Acid Res 11, 5661 -5669.
Yanisch-Perron, C. , Vieira, J. & Messing, J. (1985) Improved MI3 phage cloning vectors and host strains: nucleotide sequences of the M13mp18 and pUC19 vectors, Gene 33, 103-119.
Yuuki, T., Nomura, T., Tezuka, H., Tsuboi, A., Yamagata, H., Tsuka- goshi, N. & Udaka, S . (1985) Complete nucleotide sequence of a gene coding of heat- and pH-stable a-amylase of Bacillus licheni- f o m i s : comparison of the -no acid sequences, J. ~ i ~ ~ h ~ ~ , (Tokyo) 98,1147-1156.




















