
Tuberculosis in wild seals and characterisation of the seal bacillus
6
Tuberculosis in wild seals and characterisation of the seal bacillus DV COUSINS*, SN WILLIAMS* R REUTER'~ D FORSHAW? B CHADWICK§, D COUGHRA": P COLLINS and N GALES** SUMMARY: Tuberculosis was diagnosed in 3 otariid seals found dead on beaches at 3 locations on the south coast of Western Australian between May 1990 and March 1991. This confirms that tuberculosis Is present In the 2 native seals (Neophoca cinerea and Arctocepha/usforsterl) In Western Australian waters. Mycobacterium sp Isolatedfrom the lungs of 2 of the seals were studied to determinethe similarity of the strains to each other, to the strains isolated during 1986 from Australian sea lions and New Zealand fur seals kept in captivity at a marine park near Perth, Western Australia, and to a strain isolated In 1988 from a seal trainer who worked with the Infected captive seals for 3 years. After restriction endonuclease analysis (REA) with the endonucleases Bst Ell, Bcll and Pvu 11, one of the wild seal stralnsappeared to haveIdentical DNAfragmentpatterns to the strains from the captive seals and the seal trainer. The other wild seal isolate had Identical REA profiles using Bst Ell and Bcl I, but a minor difference was detected using PvuII. Differences in these isolates were more clearly seen in restriction fragment length polymorphisms after hybridisatlon with two DNA probes. The secretory proteln MPB70, present in Mbovis, was not detected In wild seal isolates using sodium dodecyl sulphate polyacrylamlde gel electrophoresis and Western blotting techniques. Analysis of protein and DNA fragment profiles indicated that seal tuberculosis Isolates form a unique cluster within the M tuberculosis complex. Aust Vet J 70: 92 - 97 Introduction Bacterial pneumonia is one of the most common diseases encountered in marine mammals, and is frequently associated with the stranding and death of seals and sea lions (Howard et al 1983). The bacteria most commonly isolated from stranded pinnipeds are similar to those reported in cetaceans and include Pseudomom. Staphylococcus, Streptococcus and Salmonella. Mycobacterium fortuiturn has been reported to cause cutaneous (Lewis 1987) and M srnegmatis generalised (Gutter et al 1987) mycobacteriosis in sea lions. There are reports of tuberculosis in seals in zoos (Blair 1913; Ehlers 1965). but details of bacterio- logical or histopathological findings were not described. In 1986, an organism identified biochemically as M bovis was considered to be the cause of tuberculosis in a colony of captive Australian sea lions (Neophoca cinerea) and New Zealand fur seals (Arctocephalus forsteri) at a marine park near Perth in Western Australia (Forshaw and Phelps 1991). Studies of this organism showed that although its pathogenicity and bio- chemical and drug sensitivity reactions were consistent with M bovis, it had antigenic and genetic properties that indicated it was a unique strain, and was classified as belonging to the M tuberculosis complex (Cousins eta1 1990). Inmid-1988, a seal trainer working at a zoological park on the east coast of Australia Australian Reference Laboratory for Bovine Tuberculosis, Depart- ment of Agriculture, South Perth, Western Australia6151 Veterinary Pathology Services. PO Box 96, Plympton, South Australia 5038 * Department of Agriculture, Albany, Western Australia 6330 Division of Veterinary Biology, Murdoch Veterinary School, Murdoch, Western Australia 6150 Department of Conservation and Land Management, Como, Western Australia 61 52 Department of Conservation and Land Management, Albany, Western Australia 6330 Atlantis Marine Park, Yanchep, Western Australia 6035 ' '* developed pulmonary tuberculosis caused by an organism iden- tified biochemically as M bovis. The seal trainer had worked with seals at the marine park in Western Australia from 1983to 1985. Restriction endonuclease analysis (REA) revealed that the strain from the seal trainer was identical to that found in the seals at the marineparknear Perth in 1986(Thompson el a1 1992), indicating that there was a common sourc,? of infection. Because of the unique characteristics of the strainisolatedfrom the captive seals and the history of tuberculosis in the captive colony, it was suggested that tuberculosis may have originated from at least 1 of the seals at the time of capture in 1981 (Cousins et al 1990). However, to our knowledge, tuberculosis caused by a member of the M tuberculosis complex had not been confumed bacteriol- ogically in wild seals. Although early estimations placed the numbers of Australian sea lions at about 5000, a more recent study indicates that there may be 9300 to 11 700 sea lions in Australian waters. Their distribution ranges from Kangaroo Island in South Australia to the Abrolhos Islands in Western Australia, with about 2700 to 3400 located in Western Australian waters (Gales N 1992 unpublished) (Figure 1). The New Zealand fur seal is found in New Zealand, its suban- tarctic islands, and along the south coast of Australia to Augusta in Western Australia. A recent survey estimates that there are 13 breeding colonies consisting of 27 200 New Zealand fur seals in South Australia and Western Australia, withmost of the popula- tion in South Australia (Shaughnessy and Gales 1990). It is estimated that there are at least 4000 New Zealand fur seals in Western Australia. The original stock of seals caught to establish the marine park in Perth were caught in 1981 at Daw Island, 1 of the breeding colonies in the Recherche Archipelago south of Esperance. We report 3 cases of tuberculosis in wild seals (1 New Zealand fur seal and 2 Australian sea lions) found dead on the southern coast of Western Australia.Polymerasechainreaction (PCR) was used to c o n f m the diagnoses, and antigenic (sodium dodecyl sulphate polyacrylamide gel electrophoresis and Western 92 Australian Veterinav Journal Vol. 70, No. 3, March 1993
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Tuberculosis in wild seals and characterisation of the seal bacillus

Tuberculosis in wild seals and characterisation of the seal bacillus DV COUSINS*, SN WILLIAMS* R REUTER'~ D FORSHAW? B CHADWICK§,
D COUGHRA": P COLLINS and N GALES**
SUMMARY: Tuberculosis was diagnosed in 3 otariid seals found dead on beaches at 3 locations on the south coast of Western Australian between May 1990 and March 1991. This confirms that tuberculosis Is present In the 2 native seals (Neophoca cinerea and Arctocepha/us forsterl) In Western Australian waters. Mycobacterium sp Isolated from the lungs of 2 of the seals were studied to determine the similarity of the strains to each other, to the strains isolated during 1986 from Australian sea lions and New Zealand fur seals kept in captivity at a marine park near Perth, Western Australia, and to a strain isolated In 1988 from a seal trainer who worked with the Infected captive seals for 3 years. After restriction endonuclease analysis (REA) with the endonucleases Bst Ell, Bc l l and Pvu 11, one of the wild seal stralnsappeared to have Identical DNAfragment patterns to the strains from the captive seals and the seal trainer. The other wild seal isolate had Identical REA profiles using Bst Ell and Bcl I, but a minor difference was detected using PvuII. Differences in these isolates were more clearly seen in restriction fragment length polymorphisms after hybridisatlon with two DNA probes. The secretory proteln MPB70, present in Mbovis, was not detected In wild seal isolates using sodium dodecyl sulphate polyacrylamlde gel electrophoresis and Western blotting techniques. Analysis of protein and DNA fragment profiles indicated that seal tuberculosis Isolates form a unique cluster within the M tuberculosis complex. Aust Vet J 70: 92 - 97
Introduction Bacterial pneumonia is one of the most common diseases
encountered in marine mammals, and is frequently associated with the stranding and death of seals and sea lions (Howard et al 1983). The bacteria most commonly isolated from stranded pinnipeds are similar to those reported in cetaceans and include Pseudomom. Staphylococcus, Streptococcus and Salmonella. Mycobacterium fortuiturn has been reported to cause cutaneous (Lewis 1987) and M srnegmatis generalised (Gutter et al 1987) mycobacteriosis in sea lions. There are reports of tuberculosis in seals in zoos (Blair 1913; Ehlers 1965). but details of bacterio- logical or histopathological findings were not described.
In 1986, an organism identified biochemically as M bovis was considered to be the cause of tuberculosis in a colony of captive Australian sea lions (Neophoca cinerea) and New Zealand fur seals (Arctocephalus forsteri) at a marine park near Perth in Western Australia (Forshaw and Phelps 1991). Studies of this organism showed that although its pathogenicity and bio- chemical and drug sensitivity reactions were consistent with M bovis, it had antigenic and genetic properties that indicated it was a unique strain, and was classified as belonging to the M tuberculosis complex (Cousins eta1 1990). Inmid-1988, a seal trainer working at a zoological park on the east coast of Australia
Australian Reference Laboratory for Bovine Tuberculosis, Depart- ment of Agriculture, South Perth, Western Australia6151 Veterinary Pathology Services. PO Box 96, Plympton, South Australia 5038 * Department of Agriculture, Albany, Western Australia 6330 Division of Veterinary Biology, Murdoch Veterinary School, Murdoch, Western Australia 6150 Department of Conservation and Land Management, Como, Western Australia 61 52 Department of Conservation and Land Management, Albany, Western Australia 6330 Atlantis Marine Park, Yanchep, Western Australia 6035
' '*
developed pulmonary tuberculosis caused by an organism iden- tified biochemically as M bovis. The seal trainer had worked with seals at the marine park in Western Australia from 1983 to 1985. Restriction endonuclease analysis (REA) revealed that the strain from the seal trainer was identical to that found in the seals at the marineparknear Perth in 1986 (Thompson el a1 1992), indicating that there was a common sourc,? of infection. Because of the unique characteristics of the strainisolated from the captive seals and the history of tuberculosis in the captive colony, it was suggested that tuberculosis may have originated from at least 1 of the seals at the time of capture in 1981 (Cousins et al 1990). However, to our knowledge, tuberculosis caused by a member of the M tuberculosis complex had not been confumed bacteriol- ogically in wild seals.
Although early estimations placed the numbers of Australian sea lions at about 5000, a more recent study indicates that there may be 9300 to 11 700 sea lions in Australian waters. Their distribution ranges from Kangaroo Island in South Australia to the Abrolhos Islands in Western Australia, with about 2700 to 3400 located in Western Australian waters (Gales N 1992 unpublished) (Figure 1).
The New Zealand fur seal is found in New Zealand, its suban- tarctic islands, and along the south coast of Australia to Augusta in Western Australia. A recent survey estimates that there are 13 breeding colonies consisting of 27 200 New Zealand fur seals in South Australia and Western Australia, withmost of the popula- tion in South Australia (Shaughnessy and Gales 1990). It is estimated that there are at least 4000 New Zealand fur seals in Western Australia. The original stock of seals caught to establish the marine park in Perth were caught in 198 1 at Daw Island, 1 of the breeding colonies in the Recherche Archipelago south of Esperance.
We report 3 cases of tuberculosis in wild seals (1 New Zealand fur seal and 2 Australian sea lions) found dead on the southern coast of Western Australia. Polymerase chainreaction (PCR) was used to confm the diagnoses, and antigenic (sodium dodecyl sulphate polyacrylamide gel electrophoresis and Western
92 Australian Veterinav Journal Vol. 70, No. 3, March 1993

Western Australia
ou Distribution of seals in Australian waters
A.p. doriferus A. forsteri KI
N. cinerea
igure 1. Map of southern Australia showing the distribution of seals in Australian waters, the locations where the tuberculous wild seals were )und in 1990 and 1991, and the site where marine park seals were captured in 1981.
blotting) and genetic studies (REA and hybridisation studies) were performed to assess the similarity of strains from the wild seals to the strain previously isolated from seals at the marine park near Perth and from the seal trainer who had worked with seals at the same marine park for 3 years.
Materials and Methods Animal History and Clinical Findings
Case I -On 9 May 1990, a mature male New Zealand fur seal was found dead at Yallingup Beach (Figure 1) on the south west coast of Western Australia. The seal had been sighted in the vicinity in the previous week. It was seen convulsing on the morning of 9 May 1990 and died later that day. The carcase was submitted to a local private veterinary practice for necropsy.
Case 2 -On 11 January 1991, a sub-adult male sea lion was noticed in the harbour at Albany on the south coast of Western Australia (Figure 1). He was in some distress, with laboured breathing and numerous skin lacerations. He was observed in the water in the evenings and lying on the beach during the day, apparently oblivious to his surroundings. The sea lion was found dead on the shore 3 days later and the carcase was submitted to the Albany Regional Laboratory of the Western Australian Department of Agriculture for necropsy.
Case 3 -A sub-adult male sea lion was found dead at Bremer Bay (Figure 1) on the south coast of Western Australia on the morning of 23 May 1991. A limited necropsy was conducted on the beach.
Pathological Examination Suspect granulomatous or other lesions as well as a range of
normal tissues were collected from these cases and submitted fresh or fixed in 10% buffered formalin for bacteriological or histopathological examination. In case 1, pleural fluid, lung and mediastinal lymphnode were submitted fresh to the local hospital laboratory for general culture because mycobacterial infection was not suspected. A wide range of tissues were submitted to the Murdoch Veterinary Hospital for histopathological examination. In case 2, tissues were collected for histopathological examina- tion and lung was submitted for general and mycobacterial culture. In case 3, fresh samples of lung, heart and kidney were sent to the Animal Health Laboratories, South Perth, for histo- pathological and bacteriological examination.
Bacteriology Mycobacterial culture and identification was perfomed using
standard mycobacterial methods (Vestal 1975). In addition, suspect colonies of mycobacteria were tested with a monoclonal antibody raised against M bovis (Agen Biomedical Ltd) using the immunoperoxidase test (Comer et a1 1988).
Polymerase Chain Reaction PCR was used to identify Mycobacterium sp isolated in culture
and in the direct detection of mycobacterial DNA extracted from sections of fixed lung tissue from each seal using a moditication of standard methods (Wilson 1988; Wright and Manos 1990). Sections from lung of a confiied case of bovine tuberculosis and a normal bovine lung were used as positive and negative controls. The PCR was performed using a multiplex PCR reported previously (Wilton and Cousins 1992).
SDSIPAGE and Western Blotting Both isolates obtained from the wild Australian sea lions were
compared with previous isolates from 2 of the captive seals (894 and 895) and the seal trainer (146-D) as well as reference strains of the M tuberculosis complex, namely M tuberculosis H37Rv, M bovis AN5, M bovis BCG, M africanum (TMC 3, 12 and 37) and M microti (NCTC 08710). Antigen was extracted using a freeze-thaw method (Duffield 1988) and analysed by SDSPAGE in duplicate. The separated protein bands from each strain were silver stained. The duplicates were transferred to a nylon membrane using Western blotting procedure and probed with a monoclonal antibody to M bovis as described previously (Cousins et a1 1990).
Restriction Endonuclease Analysis REA was performed on the 2 isolates from the wild sea lions,
the 2 from the captive seals, the 1 from the seal trainer and the 7 reference strains from the M tuberculosis complex group of organisms described above. Restriction digests were performed with each of the 3 endonucleases Bst 11, Bcl I and Pvu I1 (Toyobo Co., Ltd. Osaka, Japan) and analysed by gel electrophoresis.
Restriction Fragment Length Polymorphism DNA electrophoresis and Southern transfer - After
electrophoresis, DNA fragments from the REA digests above and
Australian Veterinary Journal Vol. 70, No. 3, March 1993 93

a digest using Alu I were transferred to positively charged nylon membrane under vacuum. Each of the membranes was hybridised with two probes.
Preparatwn and use ofpTBNl2 probe - An RNA probe was labelled with non radioactive digoxigenin-UTP by in-vitro transcription from the DNA clone pTBN12 (isolated from Mtuberculosis) (Ross et al 1992) with T7 RNA polymerase using an RNA labelling kit. Hybridisations were performed overnight at 42C essentially as reported by Ross et al(1991).
Preparatwn and use of IS61 10 oligonucleotide - A 31-base oligonucleotide incorporating a sequence matching the left hand inverted terminal repeat of IS61 10 (Thlerry et al1990) was end labelled with non radioactive digoxigenin-l1dUTP using ter- minal transferase and used as probe in hybridisations performed overnight at 69C.
Digoxigenin detection method - The presence of digoxigenin labelled probes were detected using the alkaline phosphatase conjugated antibody DNA detection kit with the chemilumines- cent substrate 3-(2’-Spiroadamantane)-4-methoxy4-(3”-phos- phoryloxy)-phenyl-l,2dioxetane (AMPPD) as recommended by the supplier (Boehringer, Mannheim, Germany). The images were recorded on X-ray film and compared by eye.
Results Pathological Findings
Case 1 - At necropsy of the New Zealand fur seal, the thoracic cavity contained about 5 to 8 L of cloudy fluid There was a large mass in the left lung about 40 to 50 mm in diameter. The right lung was collapsed, and the lung and the pleura contained numerous 5 mm lesions. Fibrinous, white caseous material covered the entire pleura. Associated lymph nodes were en- larged. There was a large scarred area on the spleen. The gastrointestinal tract was empty. Histopathological examination of the lungs and lymph nodes revealed a pyogranulomatous reaction with acid-fast bacteria detected by Ziehl-Neelsen stain. All lung and lymph node samples submitted (including pres- capular, bronchial and mediastinal lymph node) were affected. Only mild, multifocal inflammation was seen in the liver. No lesions were found in the kidneys and heart. These fmdings suggested a chronic, pyogranulomatous pleuropneumonia and lymphadenitis caused by a Mycobacterim sp. The lung appeared to be the primary focus of infection.
Case 2 -The sea lion showed evidence of severe dehydration. The stomach was empty. The trachea contained a large amount of thick, brown, mucopurulent fluid. There was pneumothorax with collapsed lungs. The apical and cardiac lobes of the left lung contained numerous small abscesses. The entire diaphragmatic lobe of the right lung was consolidated and grey-white in colour, with purulent material oozing from the cut surface. The bronchial lymphnodes were enlarged and moist. There were no other gross lesions. Microscopic examination revealed an extensive granulomatous pneumonia characterised by infiltration of lymphocytes, neutrophils, macrophages and epithelioid cells in the bronchi and alveolar lumens associated with foci of necrosis, minerdisation and interstitial fibrosis. Granulomas were also found in the spleen and bronchial lymph node. Many slender acid-fast bacilli typical of Mycobacterium sp were seen in macre phages and in the areas of necrosis in the lung, lymph node and spleen.
Case 3 - At necropsy there was considerable autolytic change present and lesions were evident only in the lung tissue. These consisted of extensive areas of pale consolidation in some lobes with smaller focal lesions in the periphery of othm. Histo- pathological lesions were similar in all areas of the lung ex- amined. They consisted of areas of coagulative and liquefactive necrosis surrounded by a mixed inflammatory cell population
Figure 2. Multiplex PCR on seal isolates after electrophoresis on a 2% agarose gel and staining with ethidium bromide. Lanes 1 and 8 molecularweight standards (lambda Hindlll and pUC 19 Hpa II), lane 2 negative control, lane 3 M intracellulare, lane 4 M avium , lane 5 M bovis, lane 6 isolate from case 2, lane 7 isolate from case 3.
consisting mainly of plump spindle and lymphoid cells. The reaction extended into the surrounding alveoli where many macrophages filled the air spaces. Many acid-fast bacilli were seen concentrated in the necrotic areas.
Bacteriology In the New Zealand fur seal (case l), a moderate number of
leucocytes and gram-negative bacilli were evident in the gram- stained impression smears from lung and mediastinal lymph node. On general culture, heavy mixed growth of gram-negative bacilli and heavy growth of Escherichia coli were found in the lung and lymph node. No growth was evident from the pleural fluid after 48 h. Mycobacterial culture of the lung from the 2 Australian sea lions (cases 2 and 3) revealed heavy growth of an acid-fast bacillus identified by biochemical and drug sensitivity tests as M bovis. Both of these isolates were negative when tested by the immunoperoxidase test.
Polymerase Chain Reaction Both of the Mycobacterium sp isolated from the lung of the 2
Australian sea lions and preparations from fixed tissue sections from each of the 3 seals and the positive control tissues produced DNA fragments of 1080 bp and 372 bp in size in the PCR for Mycobacterium sp and M tuberculosis complex, respectively (Figure 2). No amplifiedproduct was evident in the normal tissue negative controls.
SDSIPAGE and Western Blots SDSPAGE of mycobacterial protein patterns for the seal iso-
lates and other M tuberculosis complex strains were similar but differences weredetectedbetween species. Proteinprofiles of the isolates obtained from the 2 wild seals were more similar to each other and to profiles from captive seals and seal trainer isolates than to the profile from the reference strain of M bovis. In fact, the seal isolate profiles showed more similarity to the profiles of M tuberculosis and M bovis (BCG) than to the M bovis profile. After Western blotting, protein bands at 23 and 27 kDa were recognised by the monoclonal antibody to M bovis, only in the profile belonging to the reference strain of M bovis.
REAIRFLP The REA and RFLP DNA profiles obtained from seal isolates
were different from those obtained with the reference strains
94 Australian Veterinary Journal Vol. 70, No. 3, March 1993
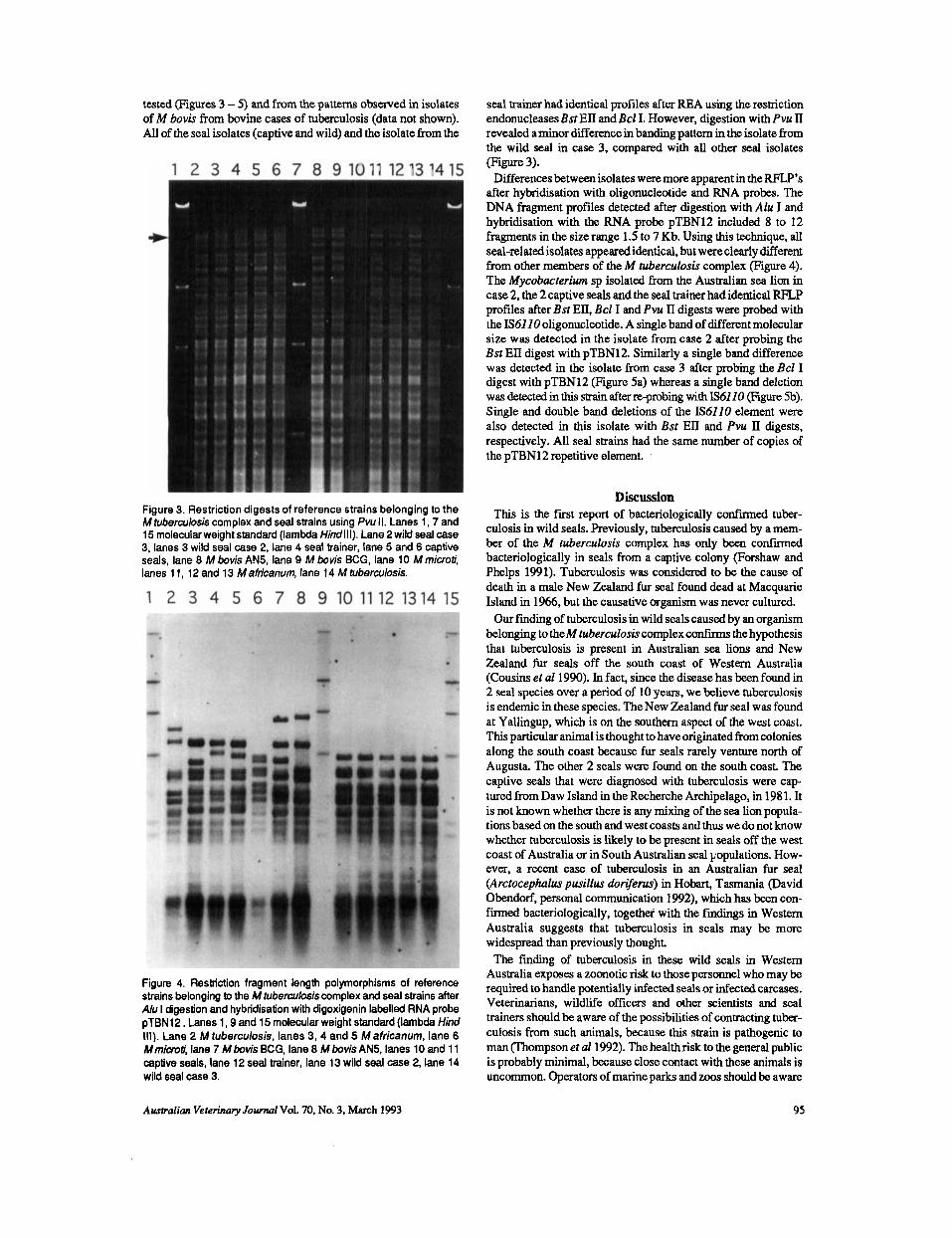
tested (Figures 3 - 5 ) and from the patterns observed in isolates of M bovis from bovine cases of tuberculosis (data not shown). All of the seal isolates (captive and wild) and the isolate from the
Figure 3. Restriction digests of reference strains belonging to the M tuberculosis complex and seal strains using Pvu II. Lanes 1, 7 and 15 molecular weight standard (lambda Hindlll). Lane 2 wild seal case 3, lanes 3 wild seal case 2, lane 4 seal trainer, lane 5 and 6 captive seals, lane 8 M bovis AN5, lane 9 M bovis BCG, lane 10 M microti, lanes 1 1, 12 and 13 M africanum, lane 14 M tuberculosis.
Figure 4. Restriction fragment length polymorphisms of reference strains belonging to the M hrberculosiscomplex and seal strains after A h I digestion and hybridisation with digoxigenin labelled RNA probe pTBN12. Lanes 1,9 and 15 molecular weight standard (lambda Hind Ill) . Lane 2 M tuberculosis, lanes 3, 4 and 5 M africanum, lane 6 Mmim?i, lane 7 M bovis BCG, lane 8 M bovis AN5, lanes 10 and 1 1 captive seals, lane 12 seal trainer, lane 13 wild seal case 2, lane 14 wild seal case 3.
seal trainer had identical profiles after REA using the restriction endonucleases Bst EII and Bcl I. However, digestion with Pvu II revealed aminor difference in banding pattern in the isolate from the wild seal in case 3, compared with all other seal isolates (Figure 3).
Differencesbetween isolates were more apparent in the W ’ s after hybridisation with oligonucleotide and RNA probes. The DNA fragment profiles detected after digestion with Alu I and hybridisation with the RNA probe pTBN12 included 8 to 12 fragments in the size range 1.5 to 7 Kb. Using this technique, all seal-related isolates appeared identical, but were clearly different from other members of the M tuberculosis complex (Figure 4). The Mycobacrerim sp isolated from the Australian sea lion in case 2, the 2 captive seals and the seal trainer had identical RFLe profiles after Bst ED, Bcl I and Pvu 11 digests were probed with the IS6110 oligonucleotide. A single band of different molecular size was detected in the isolate from case 2 after probing the Bst EII digest with pTBN12. Similarly a single band difference was detected in the isolate from case 3 after probing the Bcl I digest with pTBN12 (Figure 5a) whereas a single band deletion was d e t a t d in this strain after re-probing with IS61 10 (Figure 5b). Single and double band deletions of the IS6110 element were also detected in this isolate with Bst EII and Pvu 11 digests, respectively. All seal strains had the same number of copies of the pTBN12 repetitive element.
Discussion This is the first report of bacteriologically confiied tuber-
culosis in wild seals. Previously, tuberculosis caused by a mem- ber of the M tuberculosis complex has only been confiied bacteriologically in seals from a captive colony (Forshaw and Phelps 1991). Tuberculosis was considered to be the cause of death in a male New Zealand fur seal found dead at Macquarie Island in 1966, but the causative organism was never cultured. Our finding of tuberculosis in wild seals caused by an organism
belonging to theM tuberculosis complex confii the hypothesis that tuberculosis is present in Australian sea lions and New Zealand fur seals off the south coast of Western Australia (Cousins et a1 1990). In fact, since the disease has been found in 2 seal species over a period of 10 years, we believe tuberculosis is endemic in these species. The New Zealand fur seal was found at Yallingup, which is on the southern aspect of the west coast. This particular animal is thought to have originated from colonies along the south coast because fur seals rarely venture north of Augusta. The other 2 seals were found on the south coast. The captive seals that were diagnosed with tuberculosis were cap- tured from Daw Island in the Recherche Archipelago, in 1981. It is not known whether there is any mixing of the sea lion popula- tions based on the south and west coasts and thus we do not know whether tuberculosis is likely to be present in seals off the west coast of Australia or in South Australian seal populations. How- ever, a recent case of tuberculosis in an Australian fur seal (Arctocephlus pusillus dorgencr) in Hobart, Tasmania (David Obendorf, personal communication 1992), which has been con- firmed bacteriologically, together with the fiindmgs in Western Australia suggests that tuberculosis in seals may be more widespread than previously thought.
The finding of tuberculosis in these wild seals in Western Australia exposes a zoonotic risk to those personnel who may be required to handle potentially infected seals or infected carcases. Veterinarians, wildlife officers and other scientists and seal trainers should be aware of the possibilities of contracting tuber- culosis from such animals, because this strain is pathogenic to man (Thompson et al1992). The healthrisk to the general public is probably minimal, because close contact with these animals is uncommon. Operators of marine parks and zoos should be aware
Australian Veterinary Journal Vol. 70, No. 3, March 1993 95
of the risks of introducing tuberculosis into captive animal colonies from these sources.
The PCR identified these seals strains as belonging to the M tuberculosis complex. The PCR used does not discriminate M tuberculosis from M bovis, although one such method has been described (del Portilloef al 199 1). The use of a more discriminat- ing PCR may help in the more accurate characterisation of this organism. The PCR was also of benefit in c o n f i i g a histo- pathological diagnosis of tuberculosis caused by M tuberculosis complex in case 1, where no fresh tissue was submitted for culture. This technique shows considerable promise in the study of cases of tuberculosis where conventional mycobacterial cul- ture has not, or cannot, be performed. However, the sensitivity of this technique in detecting mycobacteria has still to be fully evaluated.
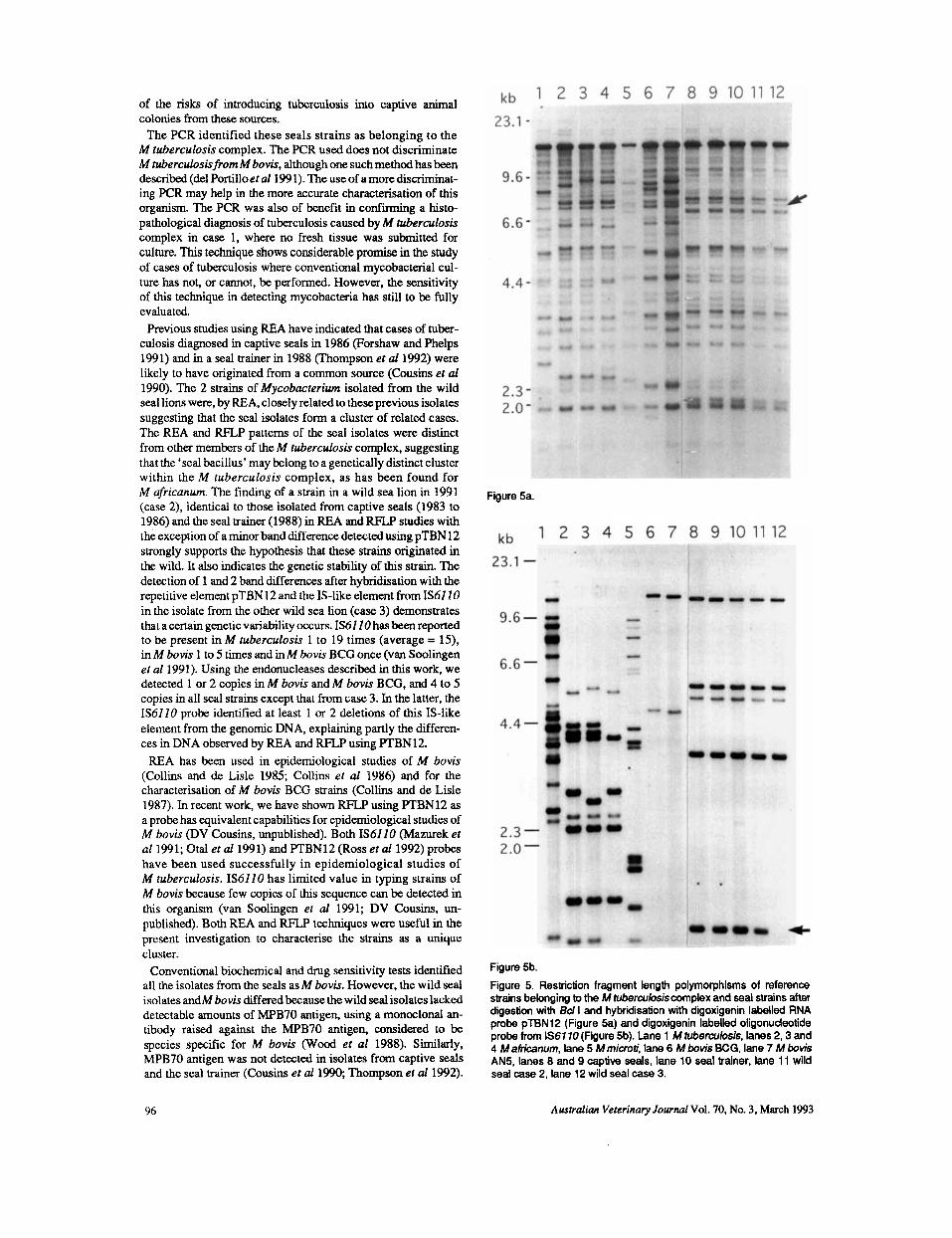
Previous studies using REA have indicated that cases of tuber- culosis diagnosed in captive seals in 1986 (Forshaw and Phelps 1991) and in a seal trainer in 1988 (Thompson et a11992) were likely to have originated from a common source (Cousins et uf 1990). The 2 strains of Mycobucterium isolated from the wild seal lions were, by REA, closely related to these previous isolates suggesting that the seal isolates form a cluster of related cases. The REA and RFXP patterns of the seal isolates were distinct from other members of the M tuberculosis complex, suggesting that the ‘seal bacillus’ may belong to a genetically distinct cluster within the M tuberculosis complex, as has been found for M dricanum. The finding of a strain in a wild sea lion in 1991 (case 2), identical to those isolated from captive seals (1983 to 1986) and the seal trainer (1988) in REA and RFLP studies with the exception of a minor band difference detected using pTBN12 strongly supports the hypothesis that these strains originated in the wild. It also indicates the genetic stability of this strain. The detection of 1 and 2 band differences after hybridisation with the repetitive element pTBN12 and the IS-like element from IS61 10 in the isolate from the other wild sea lion (case 3) demonstrates that a certain genetic variability occurs. IS61 10 has been reported to be present in M tuberculosis 1 to 19 times (average = 15). in M bovis 1 to 5 times and in M bovis BCG once (van Soolingen et a1 1991). Using the endonucleases described in this work, we detected 1 or 2 copies in M bovis and M bovis BCG, and 4 to 5 copies in all seal strains except that from case 3. In the latter, the IS6110 probe identified at least 1 or 2 deletions of this IS-like element from the genomic DNA, explaining partly the differen- ces in DNA observed by REA and RFLP using PTBN12.
REA has been used in epidemiological studies of M bovis (Collins and de Lisle 1985; Collins et al 1986) and for the characterisation of M bovis BCG strains (Collins and de Lisle 1987). In recent work, we have shown RFLP using PTBN12 as a probe has equivalent capabilities for epidemiological studies of M bovis (DV Cousins, unpublished). Both IS6110 (Mazurek el a1 1991; Otal et al 1991) and PTBN12 (Ross et ul1992) probes have been used successfully in epidemiological studies of M tuberculosis. IS6110 has limited value in typing strains of M bovis because few copies of this sequence can be detected in this organism (van Soolingen et al 1991; DV Cousins, un- published). Both REA and RFLP techniques were useful in the present investigation to characterise the strains as a unique cluster.
Conventional biochemical and drug sensitivity tests identified all the isolates from the seals as M bovis. However, the wild seal isolates andM bovis differedbecause the wild seal isolates lacked detectable amounts of MPB70 antigen, using a monoclonal an- tibody raised against the MPB70 antigen, considered to be species specific for M bovis (Wood et a1 1988). Similarly, MPB70 antigen was not detected in isolates from captive seals and the seal trainer (Cousins et al 1990; Thompson et al 1992).
Figure 5a.
Figure 5b. Figure 5. Restriction fragment length polymorphisms of reference strains belonging to the M tuberculosiscomplex and seal strains after digestion with Bd I and hybridisation with digoxigenin labelled RNA probe pTBN12 (Figure 5a) and digoxigenin labelled oligonudeotide probe from IS61 10(Figure 5b). lane 1 M tuberculosis, lanes 2,3 and 4 M airicanum, lane 5 M microti, lane 6 M bovis BCG, lane 7 M bovis AN5, lanes 8 and 9 captive seals, lane 10 seal trainer, lane 11 wild seal case 2, lane 12 wild seal case 3.
96 Awtralian Veterinary Journal Vol. 70, No. 3, March 1993

The immunoperoxidase test used routinely in our laboratory and others, has proved to be areliablemethod for the rapid identifica- tion of M bovis , because all isolates of M bovis have detectable levels of MPB7O antigen. These findings highlight the limitations associated with the phenotypic characterisation of unusual iso- lates of mycobacteria and show that molecular tests are essential for accurate taxonomic studies of mycobacteria.
The cases of tuberculosis reported here indicate that spread of' the disease was by inhalation, because the lung appeared to be the primary focus of infection. In addition, 5 of the 6 infected captive seals (Forshaw and Phelps 1991) and the seal trainer (Thompson et a1 1992) also had pulmonary infection. This is similar to the spread of tuberculosis in most other animals includ- ing man and cattle, where spread of the organism occurs most commonly by aerosols (Thoen el a1 1984). The histopathological changes were similar to those described in the captive colony of seals (Forshaw and Phelps 1991). All 3 cases of tuberculosis in the wild seals were found in older males. This may be because males forage in a wide area and occasionally haul out on main- land beaches. Females and younger seals are more likely to stay on off-shore islands. In some areas, sub-adult male sea lions tend to congregate in groups on islands after they become sexually mature until they are old enough to have their own family groups. Such habits may lead to the preferential spread of tuberculosis in animals of the same sex and age groups.
It is interesting to speculate on the original source of this infection in seals. As previously reported, it is unlikely that the seal trainer passed the infection to the seals in the captive colony (Thompson el a1 1992) or that the seals in the captive colony acquired the infection from infected cattle after capture (Cousins ef a1 1990). Whether it is possible for seals in the wild to become infected from infected cattle grazing coastal areas or islands off the south coast of Western Australia is a matter of conjecture. If this were so, the seals must have become infected before 1978, as the last case of bovine tuberculosis was recorded in that year. It is unlikely that the disease was passed from man to the seals in the days when sealing was a major industry, since any seal in close contact with sealers would, presumably, have been killed for its pelt. It is well known that adult male seals travel large distances foraging for food; their temtory may cover a 100 nautical mile radius (Gales ef al1992). Older seals, who are not active enough to catch live prey, have been known to scavenge for food. Though perhaps unlikely, there is a possibility that a seal may have eaten the corpse of a tuberculous human who was buried at sea in the days of transportation. This theory would partly explain the fact that the seal strain is more closely related to M tuberculosis than to M bovis. A further possibility, and perhaps a more likely explanation, is that tuberculosis may have been endemic in seals for much longer, perhaps hundreds or thousands of years, as long as M tuberculosis andM bovis have been present in human and cattle hosts, respectively. But if this were so, one might expect to find more genetic diversity in the strains isolated from seals, and more cases of tuberculosis in seals reported, from locations around the world.
It is hoped that further studies using DNA DNA homology, ribosomal RNA sequence analysis and gene expression studies may help to determine the genetic relationship between the seal strain and others of the M tuberculosis complex thus clarifying the taxonomic position of this strain. This should provide us with more insight into the possible origin of the strain.
Acknowledgments This work was supported by the Brucellosis and Tuberculosis
Eradication Campaign. We thank Tim Doran, CSIRO Division of Animal Health,
Parkville, Victoria, for supplying the oligonucleotide probe, and Dr Brian Dwyer and Dr Bruce Ross of Fairfield Hospital, Fairfield, Victoria, for providing us with the cloned DNA probe, pTBN12. We gratefully acknowledge the assistance of Dr John Brighton and Les C o p e in the collection of specimens. We thank Andrew Gregory, Barry Francis, Paul Ashley and Tai Lee for their excellent technical assistance and Beth Gow of the Mycobacteria Reference Laboratory for conventional identifka- tion of these organisms.
References Blair WR (1913) New York Zoological Society Sevenfeenth Annual Report,
Collins DM and de Lisle GW (1985) J Cfin Microbiol21:562 Collins DM and de Lisle GW (1987) J Gen Microbioll33: 143 1 Collins DM, de Lisle GW and Gabric DM (1986) J Hyg, C a d 9643 1 ComerLA,JohnM.BundesenPGandWoodPR(1988)VefMicrobiol18:191 Cousins DV. Francis BR, Gow BL, Collins DM. McGlashan CH ef a1 (1990)
del Portill0 P, Murillo LA and Patarryo ME (1991)JClinMicrobiol292163 Duffield BJ (1988) Immunological and Microbiological S fd ie s on Mycobacteriumbovb,PhDthesis, JamesCookUniversiry,Townsviue,p 100
Ehlen K (1965) Internafional Z w Yearbook 5149 Forshaw D and Phelps GR (1991) J Wildl DiF 27288 Gales NJ, Cheal AJ, Pobar GJ and Williamson P (1992) Awt Wildl Res 1 9
GutterAE. Wells SKandSprakerTR(1987)JZooAnLnMed18:118 Howard EB, Britt JOJ, Matsumoto GK. Itahara Rand Nagano CN (1983) In
Pathobiology of Marine Mammal Diseases, edited by EB Howard, CRC Press, Baco Raton. p 70
P 73
Res Vet Sci 48: 196
(in press)
Lewis JCM (1987)Aquat Mamm 13:105 Mazurek GH, Cave MD, Eisenach KD, Wallace RJJ, Bates JH ef a1 (1991) J
Otal I, Martin C, Vincent-Levy-Frebault V, ' I h i eny D and Gicquel B (1991)
Ross BC. Raios K, Jackson K and Dwyer B (1991) J Clin Microbiof 163:904 Ross BC, Raios K, Jackson K and Dwyer B (1992) J Clin Microbiol30:942 Shaughnessy P and Gales N (1990)AwtralianRanger Bulletin 5: 46 Thieny D, Cave MD. Eisenach KD. Crawford JT. Bates JH ef a1 (1990)
Nucleic Acids Res 18188 T h e n CO, Karlson AG and Himes EM (1984) In The Mycobacteria: A
Sourcebook Part B, edited by GP Kubica and LG Wayne, Marcel Dekker, New Yo&, p 1209
Thompson PI, Cousins DV, Gow BL, Collins DM, Williamson BW ef a1 (1992) Am Rev Resp Dis (in press)
van Soolingen D, Hermans PWM, de Haas PEW, Soll DR and van Emden JDA (1991) J Clin Microbiol29:2578
Vestal AL (1975) Procedures for the Isolation and Identifcafion of Mycobacteria, Centers for Disease Control, Atlanta, p 1 1
Wilson K (1988) In Current Protocols in Molecular Biology, edited by FM Ausebel,R Brent,RE Kingston,DD Mmre,JG Seidmanetal, John Wiley, New York, p 2.4.1
Clin Microbiol292030
J Clin Microbiol291252
Wilton S and Cousins D (1992) PCR: Methods and Applicaiwns 1269 Wood PR, Ripper J, Radford AJ, Bundesen PG, Rylatt DB el a1 (1988) J Gen
Microbiol1342599 Wright DK and Manos MM (1990) In PCR Profocols: A Guide to Methodr
and Applications, edited by MA Innis, DH Gelfand, JJ Sninsky and TJ White, Academic Press, Berkeley. p 153
{Accepted for publication 10 November 1992)
Australian Veterinary Journal Vol. 70, No. 3, March 1993 97




















