
Knox_Chromosomal_location.pdf - MSpace - University of ...
193
Chromosomal f,oõauion and. Irinkage Analysís of froose Smut Resiseancê in Wheat BY RONALD E, KNOX À Thesis Submitted to the Faculty of Graduate studies In Part'íal Fulfillment of the Requirements for the Degree of Doct.or of Philosophy Ðepartment of Plant Science University of Manitoba winnipeq, Mani Loba @ rebruary, 1994
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Knox_Chromosomal_location.pdf - MSpace - University of ...

Chromosomal f,oõauion and. Irinkage Analysís
of froose Smut Resiseancê in Wheat
BY
RONALD E, KNOX
À ThesisSubmitted to the Faculty of Graduate studiesIn Part'íal Fulfillment of the Requirements
for the Degree of
Doct.or of Philosophy
Ðepartment of Plant ScienceUniversity of Manitoba
winnipeq, Mani Loba
@ rebruary, 1994

l*l Nâional Librarv
Acquisilions andBibliographic Services Branch
395 W€llinglon SlreetOttawa, OntarioKlA ON4
Bibl¡othèque nat¡onaledu Canada
Direction des acquis¡t¡ons etdes services bibliographiques
395, rue WellingtonOnawa (Onlario)K1A ON4
You hte Vo¡te.èl&eoce
Ovt hle Na¡e ¡élüeñ.e
The author has granted anirrevocable non-exclusive licenceallowing the National Library ofCanada to reproduce, loan,distribute or sell copies ofhis/her thesis by any means andin any form or format, makingthis thesis available to interestedpersons.
The author retains ownership ofthe copyright in his/her thesis.Neither the thesis nor substantialextracts from it may be printed orotherwise reproduced withouthis/her permission.
L'auteur a accordé une licenceirrévocable et non exclusivepermettant à la Bibliothèquenationale du Canada dereproduire, prêter, distribuer ouvendre des copies de sa thèsede quelque manière et sousquelque forme que ce soit pourmettre des exemplaires de cettethèse à la disposition despersonnes intéressées.
L'auteur conserve la propriété dudroit d'auteur qui protège sathèse. Ni la thèse ni des extraitssubstantiels de celle.ci nedoivent être imprimés ouautrement reproduits sans sonautorisation.
rsBN 0-315-92243-5
C¿nadä

Nome -Dirt".t ose selecl lhe one sub¡ect which moslneorly describes lhe conlenl of your dissertolion. Enter lhe corresponding lourdigit code in lhe spoces provideä.(l¿rl t'9^!t".ql,:, ___.____ W¿U.M.ISubiecl Cotegories
ÍHE HUffiÀ¡{IflES AñIÐ SO€I,AT S€¡EN€ES(01'tMUNlGTl0NS AND IHE ARfS Psvcholoov ...............................0s25Archite<l!re..............................0729 Reodino::.................................0535Arr Hirorv................................0377 Relio oüs ..... .... .. . . . . os27Cinemo..1.................................0900 Sc;e-nc€s...................................021¿Donce......................................0378 Secondorv..........................0s33FineAns..................................0352 Soc;olScíences.........................053¿lnlormotonScience..................0723 Socioloov ol .............................03a0Journolkm................................0391 soeciol--:..................................0529LibrôryScience.........................0399 Tèoche¡T¡oinino........ 0s30Moss Communicot:ors ...............0708 Technoloav.....:........................0710MLsic .......................................0¿ 13 Têrs ondMea3urêmenh ........... 0288SoeethCommunicorion............0459 Vo<o1ono1................................07¿7TfieÞr .. .... .. .-.... .... . .... .. . .0¿ó5
lÄNGUAGI, lITIRATUR' ÀNDfDUCAïtOil UHGU¡SÌI(SYqrçrgr...1 . .. . . .. Y:l? ronorooe
ëeiêrol............... ....... . . oó7eAdultondContinu;no........... Oslóî;;i:,ii;"Ì- -" õ;i; Aacienr...............................028eÃil::ll::::, , ðiiã ljiHiilti: : 8331BilnguololdM'.rrfc,r!tu.o1.........0282 tir.rotur€8u!ine!s ...................................0ó88Comm¡nivCo¡æe.................027s Genero|.............................0¿01
ðJiirlliliàî'ñ,ì,;¿ìl;; öili !!.":'..-:l,r: P?11Eo,lv childhoôd........................05ì8ËË;{.;i;;,::* .ð!i; Medíevo,........................02eZ, Medern..............................029Bti¡\dîcê.................................... 02/ /ö;;j;;;;;J ë;;;;;l;;; ...... .. ò3jþ Akicon...............................031ó
iåi''::::::::::::l::Y ð¿Bó ame¡icon oseìiì;il;; ..................................... o;;5 Asior................................0305i.jii";;i ð!iõ coñodion lEnel¡hl..............03s2
' consdior lFr€'nchl ..............0355Home tconom¡ca ...................... 02 / Ai;¡;;i'iJ -: :-. ö¡ii !"|llil"t: : 3åliLônôùddê ond Lirerôturê 0279il1fiå%ìr:l': ::::l: ::: . . . ðiáó ro|n Americon 03r ?
Musìc.......................................0522 Middle Eosre¡n " "" 0315'phiË;;;i,;;ï .. ... . .õöiô Romonce...... .. 0313
i,iü¿:í.:i.:1... ...... .... ...... .. . . õ3á5 srovic ônd Eo5r EuroPeol 03r ¡
PHII.OSOPHY, RITIGION AIIDIHIOI,OGYPhi|osoohv................................0422
ëenero|................. ........0318BihlicôlShrdiês o3?lC|erov........................... 03ì9Hilr;ry of... .... .. . .. . .o32oPhilosôDhv of ......................0322
Theolosy.l...:............................0¿ó9
500Ât SqtN(tsA¡nericon Sbdier......... ........ ...0323Anlhropolosy
Archoeoloov...................... 032¿Culturol..11................ 0326Plìvsico| ..............................0327
Busiñeis Adñiñisrrôl;ôncenerol..............................03ì0Accounrino .........................0272Bonkins..:........... ..............0770Mônosemen1 ......................0¿5¿Mortêiinô 0138
conodion sddie' .....................0385Econor¡ics
uenero1...........-.......-.-.... . o5{JlAoricultùro1......................... 0503Cõmmerce-8usiness ........ .... 050sF¡nonce ..............................0508l-lirory................................ 0509Lobor'.............................. .0s1oTheorv................................051 I
ro|k|ore.1.......................... .......0:ssGêooroohy...-....-......................03óóGero-nt¡i]oqv .............................0351H;stoN -'
céne¡o|.... . .. . ...os7s
@

Disserlolion Abslrocls lnlernolional esf orgonisé en colégor¡es de sujels. Veuillez s,v.p, choìsir le suiet qui décrit le mieux volrelhèse et inscrivez le code numêrique opproprié dons l'espoce réservé ci-dessous.
|-|-l-n {JM.ISUJET
Cotégories por suiets
H¡'!AñA¡{¡IÊS ET SGIEN€ES SOGIAIEs
CODE DE SUJII
SE¡EN€85 ET INGÉNIER¡F
1ecrure.....................................0535Molhémorioues.................... 0280Musioue .1.. os22O¡¡eniorion êr conr!ho1ion....... .05ì9Philosoohie de l'éducotion ......... 0998Phvsio,ie 0523P¡óq¿mme! d érudes êr
enseionemenr .. .. 0727Psvcho!ãoie os255crences .................. ....... . .. . (J/ l4Sciences so<ioìes....................... 0534Sociolooie de l'èducorion........... 03¿0Technofôsie............... ......... .. 07ì0
I.ÂNGUT, TIflÉRAfURT TfUNGUT5Í0Utto¡gles. ,. _trenerolrlê! .................... .. .uôly
Ancieññes.... .... .................02891inouis|ioue...................... ..0290Mo?erne!.................. . 029t
LitlérolureGênêrolirés..................... ...04014nciennes........................... 029¿ComDorée ..........................0295Medíévo|e.... ......................02e2Moderne.............................0298Alricoine ............ .. .. . .......031óA¡éricoine ......................... 0591Anolôise....................... .. . 0593Asi¿lioue.-............. 0305conoúenne lAnoloisel ........ 03s2conodrenne lfronco,sel . ....0JJ5Ge¡monioue 03ì ìtorìno om'áricoine................ 03l 2Moyen.or;e.role......... .. 03r5Romõne 0313Slove êl €t.europænne .....0314
Çèolçie. . . 0372
Hvd;ol6oiå. ... o38sMi',¿¡olåoie or'ì ìOcèonosiophie physique . ... ....0415P.lèôhôrõ¡idL,ê O3¿5Poleðko|ooii........................ ..0¿2óP.lÉÒ¡r.ld:è or'l8Polèozooloäie...........................0985Polynolosie-........... .. .... ......... 0427
s(lt (ts Dt ta saNli It DtrrNvtR0 Ntft$NrÉconomie dome!rioue............. .. 038ó5c ences de I envroñnement 0/óaSciences de lo sontê
Générolirés .........................05óóA¿minislrorion des hìóitôux O7ó9Alimenrôlionelnukilitn . ..0570audiolø;e.... . o3oochimiôrÊ'érÒôie o99)Dênriterie..:...... .. ........ .....05ó7Déveloooementhrmoin 073AEnseioååmenr................ .. . 0350l.-,;"|""i. oga?1or5rrs.................................(J5/5Médecine du hovoilel
thérôó;e 035¿Mede¿ile er ¡hnu¡oie 05ó¿Obsréhioùe er ovñËcôlôôiê 0380oohrolmhlôore:i........ .:...... 038ìOhhoohonie-.. .... .. . .... OaóoPorhofooie O57lPhormocre .................... .....u5/2Pho¡mocolooie . .... .. . .... . .0a19Phv<iorhé¡oàie 0382RôiliôlMiÞ o57ÀSonré mËnrol-" . .... .... .........0347
5orn5 nlrrmreß ...............U5óvloxicologie . .... ..................0383
4ncienne...................-........ 0579Mediévo|e...... .... ...............0581Moderñe.......... .. ..........0582H¡roire des ¡oir'.... .... . .0328
Conodi€iñe........................033¿8rors.Unis ............... .......... 0337Eurooèenne 0335Movån-orienrole 0333Loti'no ornéricoiñe.... ........... 033óAsie, Aurrolie et Océonie....0332
Hisroire des sciences... 0585Loisìrs...................... . ........081¿Plonilicorion urboine ei
- régionole ........... .. . ....A999
çenero rles ..... ........ . uô l5Adrni¡irroiionÕuhlioùe 0617Droil et relôrio;!
internorionoles .............. ..0ól óSociolôo;e
Génirolites............... a626Aidê er bien.òrre sociol........ 0ó30
élobli";menisoénitenrioire3 O6t7
DémooroÞhie.......... .... ......0938Erudeidd l' indi"id, er
- de lo fomi |e........... .. .. ...0ó28ErL:des des relotions
inierethñ;ôùe3 erdes relorions rocioles 0ó31
Slruclure et develoooemenrso<iol.............11........ ozoo
Théorie er mérhodes ...........03¿¿TrovoileÌ relorions
industrie|jes......................0ó29Tronloorls ... . ..... 0709Tro'oìlsociol.................... 0¿52
Bioméd]co|e........... .. ..... .. 05alCholeur ei ther
modvnomioue.......... . .. .03¿8Condirìonne"i.*
{Embo |oqel .....................05¿9Gèñie oèroiooriol ...... . 0s38
Génie civil .:................... ...05¿3Génie élecìronioue et
elê(k;a!e . . .054aGénie inJurriel..... .. 05¡óGènie mèconioue................05a8çenenucleorre........... u5llMeconroùe ¡ôvôlê l)5Á/Mèro lurbie . .. .. o7Á3Science ðes rnorériou^ .... ....079aTechnioue du oèlrole . a765le.hnr.re mrnrÞrê (ì551Ìê.hni;1,ê( (ô.ir^i,êr pt
muni¿iooles.. .... .... . oss¿T€chnoloiie hvd¡oLJlioue.. . osa5
Mèconiouê ôåôliiuéê o:]r'¿,Céorechnoìosii . .......... O¿28
fTê¿h""!"l,i.l a7e5Recheiche ooé¡ãtiónnelle. . .. .. .079óTenieserris!us (Tech¡olosie) ....079¿
PSY(H0t0GtI(Jenerolrles |J621Pe¡sonnoliré............ .................0ó25Psvchobiolooie.... o3¿9Psicholøie-cl;nioue aó??Psicholdie d! c¿moo¡remenr o3B¿Pricholoãie du dev¿lÒóóéñênr 0ó20P!;choloãieexoérime;iirle.. 0ó23Psicholoðie in¿ur,ielle .. o62aPsicholoËie ohvsiolooioue........ 0989Psicholoãie lo.iole .: .:. . o¿51Psichom5r,ie .... .. ............ oó32
@
...0422
...........03r 0.........0454
...........0770
..........0272
..........0338
..........0578
o4730285
S(IINCTS PHYSIOUTS
Sciences Pu¡esChìmie
Genê¡ol'rés....... .................0¿85Biochimie . ............... .. ¿87Chi¡nie ooricole..... ....... . 07a9Chimie oñol¡io¡e 0Á86Chimie min¿'rolå........... .. . O¿8e(hrm e nucl-ÀoÍe ... . O/34Chim;e o¡oonioue ............... 0490Chimieohãrmoceurioue. 0491Phvsiouå... : oÄ9aPofvmCres ............. .... .. .0¿95Rodrolron.. .. . O/54
Molhémo1ioues..................... .. 0¿05
' c¿nè,olirés .........................oóosAcoulioLre.............. O98ó
orroohvlioue.-..... 0ó0óElecrroniqire èr élecrricir€ ...... Oó07rru des el Dlosmo......... . .. ..o/5yMéréo¡olobie . ....... . ...qéaqupnqu€,.,, -........................u/J2Porlicules {Phy!iquenucleonel.... ........0798Phvsioue orôñioùe 07 ÁgPhisiciue de l étår solide oól IPhísiciue moleculoire ........ .. oó09Pl,i¡iciuenuclæire.... 0óloRoijioi;on....... . .......025ó
Sloliliques ......................... .. . 04ó35ciences Appliqués Et
1nformorioJe........................ . .098¡lnqè¡ie¡ie
(Jenerolrles.. l)t.l/Asrico|e ................. .. .......0539Aulomobile . . 05r'O

CEROì{OSOHAL LOCATION AXD LINKAGE A¡TAIYSIS
OF TOOSE SUUT RESISTANC3 IN !¡EEAT
BY
RONAID E. KNOX
A Thesis submitt€d to the Faculty of G¡aduate Studies of the University of Manitobain partial fulfillment of the requirements of the degree of
DOCTOR OF PEILOSOPEY
@ 7994
Permission has been granted to the LIBRARY OF THE UNTyEITSITY OF MANTIOBAto lend o¡ sell copies of this thesis, to the NATONAL LIBRARY OF CANAÐA tomic¡ofilm this thesis and to lend or sell copies of the film, and LIBRARYMICROEILMS to publísh an abshact of this thesis.
The author ¡eserves other publication rights, and neithe¡ the thesis no¡ extensiveexhacts from it rray be printed or other-wise reproduced without the autho/s w¡ittenpermission"

ABSTR,ACT
The present knowledge of the genetics of toose smut resj-stance in wheat
is neither substant.ial nor coherent enough to a1low researchers Co
efficientLy develop molecular markers linked lo loose smut resistancegenes. Knowledge of the chromosome locat.ion of loose smut resislanceqenes can improve ¿he efficiency of analysÍs of genetic finkage tomapped molecular markers. The primary objec!íve of this study was todeterrnine the chromosorne location of a loose smut resistance gene inconmon wheaC, A second objective was Co evaluate seed proteins and
monoclonal antibodies to certain of those proteins as chromosome maïkers
for their role in cytogenetic analysis of loose smut resist.ance,
Linkage analysis between a loose smut' resistance gene and genetic
markers located within the same chromosone, and analysis of informationabout Lhe inheritance of loose smut resistancè were ancillaryobjectives, Segregating popula¿ions were generated by crossing loose
smut susceptible cytogenetic stocks wiCh sources of loose smu¿
resistance, and the progeny rr'ere evaluated for seed protein and loose
smut resistance traits. Seed protein and monoclonal antibody chromosome
markers assist.ed in chromosome detection, and were found to be a
reliable ¡ool in cytogenetic analysis. The chromosome-marker-assisted
cytogenetic analysl"s was used to determine Èhe chromosomal location of a
loose smut resistance gene in chromosome 6A. Evidence of an associationbetween the chromosome 6A loose smuL resistance gene and a restrictionfragment length polymorphism was found. Evaluation of cytogeneticstocks for loose smut resistance placed the 6À loose smut resistancegene in the short arm of chromosome 6A. Limited results on theinheritance of 1oo6e srnut resistance pointed to the occurrence ofrecessive and dominant genes, and cultivars possessing multiple genes
for resiseance. Loosê smut resistance genes can be located tochromosomes by using molecular markers to assist in the cytogenetic
av

analysis, and the knorvledge of the chromosome location of a loose smut
resistance gene all-ows the efficient establishment of linkage withnarkers known to be in the same chronosome.

ACKNO\ryLEDGEMENT
Research is noe done withouC the assiseance and input of rnany people. lwould like to take this opportunity to convey my gratit.ude to the people
who helped make this ¡hèsis project a success. First, f greatlyappreciate thè selfless support and guidance of my advisor, Dr. NeilHowes, who \,¡as al\À'ays available to discuss and help work out problems,
and has been a great inspiration and têacher. f also extend similarappreciation to Ðr. Jens Nielsen, whose door was always open fordiscussion and to answer my many questions related to the smut fungi.
Thank you to my advisory cornmitt.ee I Doctors Anita Brule-Babel, Lakhdar
Lamari, and .tim Reid for being very accorunodating and for providj-ng
constructive advice, f would like to give special thanks to Dr. percy
Thomas for his role as commiteee member throughout the thesis and foract.ing as advisor in Dr. Howes¡ absence during the examination process,
Thank you to the many other people who made my work much easier through
technical- assistance, contribueion of seed, use of facilities, and
discussion, a few of whom are: Carla Cohen, Wayne Mauthe, Reg Sims, Ed.
Czarnecki, Doctors Won Kim, Fred Townley-Smith, Eric Kerber, peter Dyck
and Taing Aung.
f also acknowledqe bhe A,griculture Canada Research station at SwiftCurrent, Saskatchewan for their support of my training, and inparticul-ar thanks to Dr. Ron De pauw and Dr. Àsh1ey O,Sullivan forbackinq me in this endeavour, Thanks to my family wanda, Âdam,
Veronica, and Riley, for enduring the long days and \,¡eeks at home in my
absence, and particular thanks Co my tvif e wanda for encouragement,
patience, and support.

FOR.\ryARD
ThÍs thesis starts with a general abstract, int.roduction and literaturereview, fotlowed by the presentation of four chapt.ers of experimentalreseaxch, each representing a particular research theme, The ¿hesis
concl-udes in a general discussion and conclusion followed by the list ofreferences cited throughouc the thesis, The formaÈ of each chapter isas followsl ehe abst.ract, introduceion, materials and meehods, results,and discussion on the theme of Ehe research within the chapCer. The
thesis is written in the style of the Canadi an .Jouxnaf of pLant Science.
Chapter 3 entitled ,The application of chromosome - speci f ic monoclonal
antibodies to wheat genetics, is published in Genome, Chapter 4
entitled 'A monoclonal antibody chromosome marker analysis used tolocate a loose smut resistance gene in wheat chromosome 6À, is submittedLo Theoretical and Applied Genetics, Chapter 5 itlusLrat.es linkageanalysis beeween a chromosome 6À loose smut resist.ance gene and othertraies in che same chromosome. chapter 6 brings toqether a collectionof data generated toward the objective of locat.ing a loose smut
resistance gene to a chromosome, Although providing no conclusiveresufts the data in Chapter 6 is of sufficient impact to affect thedirection of future research and therefore this information is reportedwithin the main body of the thesis. The list of references contains allreferences used throughout the thesis. Â.n appendix presents the 1ist.sof lines and results of their inoculation with a series of loose smut
races and r^¡hich were used as background information for the development
of the Chesis.

TABLE OF CONTE¡{TS
ABSTR.A.CT
ACKNOWLEDGEMENT
FORWÀRD
IJIST OF FIGURES
L]ST OF TABLES
CHAPTER 1r INTRODUCTION
CH,APTER 2: LITERATURE REVIEW
Wheat
cenetics
Àr¡aComy
Loose SmuC
Disease Cycle
Environmental Effects on Loose SmuL Expression
Inoculum and Inocul_at ions
Timing and Me¡hod
Host Range
cenetics of UstiTago tri ticiLoose Smut Races and the wheal Differential Set.
Anatomy of Resistance
cenetic SEudies of Res i s tance
cenetic Markers
Seed Proteins
Monoclonal_ Antibodies
DNA and Isoz]¡rne Markers
CHÂPTER 3: The .Application of Chromosome Specific Monoclonal
A¡tibodies to Wheat cenetics
ABSTRACT
PÀGE
ivvi
viixi
xiv1,
4
4
4
,7
8
I9
T2
13
1,4
t4
18
22
24
28
33
35
38
38

TNTRODUCTION
MÀTERI.N,LS ÀNÐ METHODS
Pl-ant Materiaf
Enzl¡me Linked fmmunosorbanl Assay (ELISA)
Sodium Dodecyl SuI fate - polyacryl amide ce1
Electrophoresis ( SDS - pÄcE)
Cytogenetic,Analysis
RESULTS
lsolation of Spontaneous 18 Nullisomic
Roblin N1B,/Prelude F, Analysis
RobIin NLB/Canthatchi//prelude/Diamant
Roblin N1B/TD6 / / prel,rde
D]SCUSSION
CHÂPTER 4: A Monoclonal AnLibody Chromosome Marker Anafysis Used
to Locate a Loose Smut Resistance cene in Wheat
Chromosome 6A
ABSTRACT
INTRODUCTION
MÄTERTALS .AND METHOÐS
RESULTS
DISCUSSION
CHÀPTER 5: The .â,ssociation Beeweèn a Wheat Chromosone 6A cene
Controlling Loose Smut ResisEance in Whea!, the Srg
and cliadin cene loci and an RFLP in the Same
Chromosone
ÀBSTRÀCT
INTRODUCTTON
MATERIÀLS ÂND METHODS
Plant Materiat
Enzyme Linked frnmunosorbant Àssay
Polyacrylamide cel Electrophoresis
38
40
40
47
41
42
42
43
46
48
s0
54
54
55
59
64
75
85
85
85
86
86
89
90

Loose Smut fnoculations
DNA Ã,nalysis
RESULTS
D]SCUSSTON
CHÀPTER 6: The Genetics of Loose Smut Resistance in Wheat
ABSTRÀCT
]NTRODUCTION
MÄTERTÀLS AND METHODS
RESULTS
D]SCUSSTON
GENERÀL D]SCUSSION
CONCLUSTON
],TST OF REFERENCES
APPENDTX A
90
90
93
103
106
106
107
109
110
134
r40
t4t158

FTGURE
Fiq. 3.1.
Fig, 3 .2.
¡ag. J,J.
Fiq. 3 ,4.
Fiq. 3 .5.
Fiq. 3.6,
Fig. 4.1,.
Fí9, 4,2.
LIST OF FIGURESPAGE
Relalive binding (to Prelude) of MÂb p24B to50% propan-2-ol extracts of half F2 kernels fromthe cross Roblin Nl-B/Pre1ude., .,...... 44
SDS-PÂcE separation of F, kernèfs of the Roblin/Prelude cross selected on the basis of M¡bbinding: slot l- and 18, Pretuder slot 2 and 19,Roblin; sloC 3-12, selection of seeds wíth a lowMÃb reading; sloL 13-l-7, selection of seeds witha hiqh MÀb reading. .......,... 45
Relative bindinq (to Canthatch) of MAb p24B to50? propan-2-o1 extracts of half kernels ofRoblin, 796-1, Canthaech and F1 Roblin N1B/Canthatch. ... ,.. , .. . . 47
Relative binding (to Neepawa) of MÀb p24B to 50%propan-2-o1 extracts of testcross F1 kernels fromthe cross Roblin N1B/Canthatch//prelude/DiamanE.. . ,.. . 48
Relative bindinq (to Prel-ude) of MAb p24B to 50%propan-2-01 extracts of testcross F1 kernelsfrom the cross Roblin N1-B/TD6 / /prelude. . . . . . ,. 49
SDS-PAGE separaLion of extracts from testcross F1kernels from Roblin NIB/,ID6/ /prelude cross! slot1, Neepawa and Prelude; slots 2-8, selectionswith hiqh MAb reacCion; slot's 9-16, selectionswith int.ermediaee M.Ab reaction; sl-ot 1?, TD6;slot 18, Prelude; slot 19, Neepawa. ......,.... 45
l"lAb 230/9 binding measured by absorbance of ELISASon seed extracts from the F, population: (À) Cadet6Aq(6A.) /cadet.; (B) cader 6AS(6A) /Kora; (c) cader6Àq (64),/Thaccher; (D) cadet 6As(6A) ,/TD18 (not.elpopulati.on and treatment sizes are not all equal) , ..,. 6jÐistributj-on of loose smue proport.ions of F3families classified by chrornosome based on ànt.ibodyreadings: (À) Cadet 6À9(64),/CadeC to race T19, (B)Cadet. 6Aq(6A.) /Thatcher to race T19r (C) Cadet6À9(6'A')/Koea to racè T19, (D) Cadet. 6Aq(6Â) /Kotato race T31; (E) Rescue 6À9(6A)/TD18 co race T19;(F) Rescue 6AS(6A') /TD18 to race T3L; (c) Rescue6As(6À) ,/TD18 to race T15i (u) nescue 6À9(6À) /TDl8to race T8 (note: population and treatment. sizesare not all equal) .. , , .1I
Crossing scheme used for the Cadet 6À95r6A5alien translocation line in a cross withChinese (Ch.) Spring Srg testcrossed with ¿healien chromosome substitution line Cadet 6Ä9(6À).,.,.. B?
Fig.5.1,

Fig, 5,2Ä. PÄcE analysis are as follows for the Lane andcullivarr 1, Chinese Spring Sr8; 2, Cadetr 3,Cadet 6Ä9(6.A') i 4, Cadet 6À95:6Ä,5; 5, ChineseSpring 6À5 t',; 6, Chinese Spring 6ÄL tt, , 7, preludesr13... .......95
Fiq, 5,28. PAGE analysis of five TC F3 seeds representingthree families (lanes 3-'l , 8-f2, 13-17) of thecadet 6À95 : 6Às/Chinese Spring sr8//Cadet. 6A9(6A)populatíon and controls Cadet (lanes 1 and 19)and Chinese Spring Sr8 (lanes 2 and 18) .,. . . ,. 95
Fiq, 5.3, Segregation of Cadet. 6A95 : 6À5 /Chines e Spring Sr8/ /cadeL 6Às(6À) TC F3 families for reaction to loosesmut. race T19..,. .,... 96
Fig. 5.4. DNÀ of homozygous recombinant lines for the 6A5chromosorne arm restricted wilh Hind III and probedwith probe 167 whexe lane and descriptíon of lineare as foll-ows: 1, Cadet 6Àgs:6Äs; 2, Cadet 6Âg(6Â) ;3, Chinese Spring Sr8, 4-8, loose smut resistantlines without band; 9-L5, loose smut susceptiblelines wÍth band. . . .. . fO2
Fiq. 6.1, Distribut.ion of prÕgeny of the Rescue 6AsrcA]) /H44population inoculatêd to race T15.... .,,...,..111Fig. 6.2. Loose smut xeaction of families of the population
Rescue 6À9 (6.A),/ctenlea inocutated with (A) T8, (B)TLo, (c) T15, (D) T19, (E) T31, (F) T39. . . . . ... .... . ..172
Fiq. 6,3, Loose smut reaction of families of the populationCadet 6A9(64) /KoCa inoculated with (A) TL5, andRescue 6.As(6À)/rD6 with (B) T8, (c) T10, (Ð) T15,(E) rjl ,......113
Fig. 6.4. Loose smut reaction of families of lhe populationcadet. 6A,q(6A) crossed with (A) Red Bobs and inoculatedwith T10, (B) TD3 and T10 and Rescue 6AS(6A) crossedand inoculated with (C) TD16 and T15, (D) TD16 andT10, (E) TDI-6 and T31.... ,....j.1-4
Fiq. 6,5. Loose smut reaction of families from populations ofRoblin NLB crossed onto a Kota and testcrossedwiÈh prelude and inoculated \r'ith races (A) T1O, (B)T15, (c) T19,... ......115
Fiq. 6,6. Loose smut reaction of families of the populationson which a loose smut resist.ant 1ine was crossedont.o Roblin N1B and test crossed with prelude,cultivar and race are as follows (À) TDI-B and T1O,(B) TD18 and T19, (C) Canthatch and T19, (D)Canthatch and T39..... . . . .116
Fiq, 6.7, Loose smut. reaction of families from the populationwith which Hy377 was crossed onto prelude MIB andtest crossed \,¡ith prelude, inoculated with (À) T2,(B) r10, (c) r19, (D) r31 (E) r39. ....77,7
xLl-

Fig. 6.8. Loose smut reaction of families from the populationsin which Rescue 6Ä,9(68) was crossed and ínoculatedwith (À) TD3 and r8, (B) TD3 and Tj-o, (c) Red Bobsand T8, (D) Red Bobs and T10.... ..,...L19
Fiq. 6.9, Loose smut. reaction to various families from thepopulations of (A) Roblin N1B/TD2 i/Pre1ude, (B) Rescue6Ãs(6À) /TD9, (c\ Rescue 6À9(68)/TD2, (D) Rescue6Ag(6D) /cIenlea. ......L19
x}J-.r.

LIST OF T,.{BLEST,AB],E PÂGE
Table 2.1.A. summary of races¡ virulence genes¿ anddifferentials used in genetic studies by Nielsen(L977, L982l and resistance genes assigned byMclntosh (1988) based on Lhe gene-for-geneprinciplè, along with race reaction as susceptible(S) or resistant (R) . ......... I7
Table 2,18. Phenotlt)es when combinations of resistance al_Ie1esat one locus and qfenes at ti¡o loci are cornpared withvirulence alleles at one locus and genes at two lociwhere (-) = susceptible or avirulent condition atother 1oci. ...,..,,... 17
Table 2.1C. Evidence for a gene dj-fference for resistance betweenTÐ2 and TD3, and between TD4 and TD5.... .,.... !7
Table 2,1D. Evidence that TD2 has an additional gene to TDI-O andl-ikewise TD3 has an additional gene to TD10..,........ I7
'Iable 2.2. A condensed differential set of ?riÈicum aesEivum(data collected by ,I. Nielsen) .., ... .. 19
Table 2.3, Races UstiTago triÈ.ici which differènt.iate theTriticum aestivum dif ferentials (from Nielsenl-987a), .......2!
Table 2.4, À sunìnary of genetic studies of resistance to loosesmut. of wheat.. ....... 25
Table 2.5. Chromosome and seed proteins associated with thatchromosome. (Âdapted from carcia-Olnedo et a1 .(1978, 1982) wirh addirions) ..... ..,.. 32
Table 3.1, Somatic chromosome number, electrophoretic bandingpattern, and reaction to M,Ab p24B of cuLtivars usedin crosses for cytogenet.ic analysis, .,.,,.,,,. 40
Table 3,2. Comparison of cyt.ological analysis of rooc tips toP24B MAb reaction of populat.ions Roblin NlB,/pïeludeand Roblin NtB/,rD6//prel-ude,... ....... 46
Table 3.3; Genetic assortment of M.Ab p24B reaction, occurrenceof PÀcE band and root t.ip count and chromosomemorphology of Roblin NlB/Canthatch/ / pretude/DiamantCestcross F2 progeny derived from Fl seed cfassedas intermediate ox high M.Ab binding, .. , .. . .. , . .49
Table 4,J.. Proportion of smutted to total plants (p) and totalplants (N) of wheat cultivars and lines inoculatedwith loose smut racês T19, T31, T15 and TB andmonoclonal ant.ibody 230/9 binding relative t.o Cade!(oD) and the standard deviat.ion (sd) usinq a samplesaze or sJ-x,.., ..,,... 65

Table 5.1. Sègregalion of Cadet 6À95:6A5 and Chinese SpringSxg for a 6A qliadin band and loose srnut xace T19resislance and a chi square to test deviation froma 1:1:1:1 segregation ratio,. ,......., 97
Table 5.2. Chi square values for goodness of fit of segregat.ionratios for qliadin against Sr8 rust resistance andloose smut resistance against Sr8 rust. resistance,.... 99
Table 5.3, Ä list of ÐNA probes located to the 6Â chromosomeand the source of bhe probe. . . ,... . , . 100
Table 5,4. Segregation of progeny from the Cadet 6À95¡6Ä5/chinese spring srg//cadet 6À9{6À) populábion fora 6A Hind III RFl,p band and (À) loose smut raceT19 resíst.ance, (B) stem rust resistance, (C)gliadin band... , .,... 101
Tab1e 6.1, Loose smut race T8, T15 and T19 reaction of CadetsubsbiLution lines into which individual Rescuèchromosomes have been substituted, .., i.22
Tabfe 6.2, Loose smut. race T6, T10, T15, T19, T31 and T39reaction of chinese spríng substitution linesinto which individual Thatcher chromosomes havebeen substituted.... .........1-23
Table .A.,1, Infection results of a group of Cadet cytogeneticstocks evaluafed with races T2, T6, TB, Tl_0, T15,T19, T31 and T39 of loose smut... .... 161
Table .4.2. Loose smut lace a2, T6, T8, T10, TL5, T19, T31and T39 reaction of various Rescue cytogenetic
Table .4,.3. l,oose smut incidencè in the hexaptoid wheatdifferentials inocutated Lo ,rZ, "f6, Tg, T1O, TL5,T19, T31, T39, . . .
Table À.,4. fnoculat.ion of miscellaneous cultivars andcytogenelic stocks with races T2, T6, Tg, T1O, Tj.5,T19, T31, T39.,,,
762
r.6 3
165

1
CT{APTER. 1
INTR.ODIJCTION
Loose smul is a disease of historical economic importance (Fischer and
Holton 1957) , and the disease continues to inflict. losses and add toproduction costs by forcing farmers Lo chernicatly treat sèed ofsusceptible cuttivars. Input costs to farmers can be reduced byproviding loose smut resistant varieties. À thorough understanding ofthe genetics of foose smut resistance provides more ef f .icient mêans ofmaintaining resistance during selection, and thus of reducing the costof breeding for resistance to lÕose smut. The development of geneticmarkexs linked to loose snut resistance genes provides the potential offureher rêducing this cos t.
Knowledge of the significance of Lhe host crop is important to putinto pexspective the impact of a disease such as loose smut. Wheat is a
widely adapt.ed crop, and is grown in most parts of the wortd at variousa1litudes and at. various times Ehroughout. the year. Because of its wide
adaptation, wheat covers more hectares of land, and total production isqreater, than any other crop in the world (Briggle and Curtis 1997). À
susceptible variety of wheat grown on the Canadian prairies, \dithout any
means of control of loose smut, will suffer an annual average yiel_d lossof eiqht percent (Niè1sen unpublished) , Such a potential loss on aworld-wide basis is considerable.
In Canada, genetic resistance to loose smut in whea! is the preferred.method of control, although through Che l_990s, several susceptibl-e wheat
varieties tr'ere registered (ÀDF 1993) . cenetic resistance provides a
favourable alternative or supplement to chemical control, and. effectiveresistance to a comprehensive collection of races from around the wortddoes exist. (Nielsen 1993). The greatest barrier to the use of foosesmut resisgance is the difficulty in evaluating breeding material for

2
this trait. Litt.le usefut informat.ion is available on the genetics ofloose smut resistance in wheat, part.ly because few studies have been
done, and partly because the information from the different studiescannot be interrelated due to inconsistene charac teri zat.ion of races.An understanding of the genetics of loose smut resist.ance will lead tomore efficient breeding, by allowing the charac teri za¿ion of genes interms of the nuÍìber required for effective resistance, the breadth ofresj-stance of each individual gene, and the expressivity and penetrance
of individual genes. Breeding efficiency wil_l also be gained by
improved ability to determine poputat.ion size, and bettexcharacteri zatíon of races. Determining the chromosome focat.ion assisCs
in identifying genes, and is useful in the development of markers tofoose smut reaistance genes. The development of markers closely IínkedLo loose smut resistance genes provides the potential of more efficientsefect.ion for resistance íf Èhe markers are more easily evaluated thanloose snut resis tance.
Many Cerms have been created to explain cytogenetic conditj-ons, a few
\{hich can be confusing are defined here. The phrase chromosome locacionwill be used to refer to the chromosome in which a gene resides, and
will be used in a different sensê from gene mapping. Gene mapping witlbe used t.o refer to the position of a gene relative to other genes or tothe centromere, Thê tern linkage will be used t.o indicaÈe the degree ofassociation of genes with each other. The traditional definition oftes¿cross is a cross of a homozygous recessive parent. onto an indivídualwhich is either homozygous dominant. ox heterozygous, Che segregation ofwhich is used to measure the condition of al1e1es at the loci inquestion, For the purpose of cybogenetic analysis the traditionaldefinicion is expanded to include the null altele, which results fromthe use of particular cytogenetic stocks, and which will be t.reated as a
recessive a1le1e (e.q. a nullisomic 18 stock will be considered to have
the null alleIe for any gene in that chromosome) .

3
Like breeding for loose smut resistance, genetíc analysis of loose
smut. resistance is difficult because of a requirement for a large familysize, imposed by the failure of the fungus to infect atl plants with a
susceptible genotype, This requirement for large families i.s compounded
for routine cytogenetic analyses used to locate genes to chromosomes,
because the most comnon procedures also require a large population size.The Cwo primary objectives at the beginning of ehis thesis project
were eo lay the groundwork for more efficient cytogenetic analysis usingmolecular chromosome markers in conjunction with segregating populaCions
derived from cylogenetic stocks, and to identify the chromosome locationof at least one loose smut resistance gene using the chromosome marker
t.echnique, If time permitted, a further objective was to establish a
linkage between a loose smut resistance gene located to a chromosome and
markers located to the same chromosome. The strategy used was to focusanalysis on a few chromosomes for which good chromosome markers were
available, and to croes severaf sources of loose smut. reeistance ontocytoqenet.ic stocks for the chosen chromosones. À series of races was
used to inoculat.e segregating progeny to improve the chances ofcharacterizing Índividual genes in lines which are multigenic for loo6e
smut resistance. Because chromosome markers were used, smal1
populaeions provided sufficient information to determine the associationof loose smut resistance with a chromosome. Even though smal1
populations were used, information on the nunbex of genes segregatingcould st.i11 be established because a series of races was used. toinoculate segregating progeny, A fourth object.ive of this project was
to analyze the data from all populations to provide preliminary evidence
of the nature and nunìlf,er of genes involved in loose smut resist.ance Ínthe tÍnes studied. Fina]ly, many genetic stocks were inoculated t.o
determine how they should be used in crossing, and loose smut resistantcytogenetic stocks were inoculated to try to obLain evidence of thechromosome l-ocation of the resisgance,

4
CH,dPTER 2
LITER.ATUR,E REVIEW
This litêrature review covers, in detail, aspects of the biology and
genetics of wheal and of the pathogen causing loose smut, Ustifagotritici (Pers.) Ros¿rup f.sp. tritici (Boerena et a1 . 1992) (referred tohenceforth as ustiTago tritici based on Nietsen (1985)). The
infor¡nation presented attempts lo provide insight into the factorsaffecting the study of genetics of wheat resistance Lo Ustifago tritici,and provides background for the development. of genetic markers in wheat,
markers for boCh chromosomes and resist.ance genes. Such basicinformation is essential if mapping genes for resistance Eo thispathogen is to be successful-. The disciplines from which information isdrawr include the anatomy and morphology, genetics, and pathology of theU. tri tici /wTreat. aegricorpus, and the cytogenetics, seed proteinchemistry, and molecutar biology of wheat. Review articles will be
reported where possible for the reader who desires more background,
Primary reports are normally referred to rvhen the information has a
potential impact on the course of research into the development ofmol-ecular markers Co loose smut resislance genes.
9lheat
cenetics
Corunon or bread wheat lTîiticum aestivun) is an a11opo1yp1oid
(hexaploid) cereal grain of relatively recent evolutionary origin, and
is a me¡nber of the tribe Triticeae of the family cramineae (Feldnan
1976; Kinber and Sears 1987). Such a complex genome confers bothadvantages and disadvantages on its study (Soxrelts 1992) , The
signifj-cance of three genomes is that many genes occur in moïe than one

5
dose (Schmidt 1973, Hart 1982) , províding the plant wit'h a high degree
of genetic plasticity, but making genetic analysis more difficult (e.q,
the three seed colour genes (Schmidt 1973)), The three genomes,
however, aI1ow aneuploid manipulations that cannot be accomplistred in a
diploid (À]lard L960) , wheat 1ínes with such cytogenetic aberrationsare known and used Ín localing qienes to chromosome arms, and in mapping
genes relative to the centromere. Because wheat is of relatively recentevolutionary origin, diversity at the DNÀ leve1 is expectèd to be
relatively low, in fact little polt¡morphism is detected. at this leve1(He et al , 1992; ,loshi and Nguyen 1993).
To accomplish the objective of tocating a loose smut resistance gene
to a chromosome, techniques prevíously used for locaÈing other traits tochromosomes can be adapted. These principles also apply to locatingtraits with poeential as genètic markers to chromosomes, À discussionof the principles behind different methods used to tocate genes tochromosomes, along with the advantages and disadvantages of each method,
is left t.o the introduction to Chapter 4. Evaluation of substiCutionlines or alien addition lines carrying pairs of homoeologous
chromosomes, monosomic analysis, telôsomic analysis, in situhybridization, and linkage to a lrait previously located t.o a chromosome
are all methods by which g:enes can be located to chronosomes,
Analysis of Lwo series of subseiEut.ion tines was used by Dhitaphichitand coworkers (1989) ¿o locate genes for loose smut resistance.Nicholson and coworkers (1993) used the method to determine thechromosomes involved in Septoria nodorum resistance. This meehod, alongwith analysis of ditelosomic, nullisomic-t.etrasomic, and alien additionlines, is the method of choice fox locating potential gènetic markeïs tochromosomes, Examples of polential markers localed using these meehods
include: Random Ãmplified pollmorphic DNÀ,s (RÀpDs) (Devos and cale1992); Restriction Fragment Length polymorphisms (RFLps) (Chao et a1 .
L988, 1989; Sharp et at, 1988a, 1989; Gitt et at. 1991; Harcourt and

6
Gale 199L; Heun et aI . 1991; Liu and Tsunewaki 1991, Ànderson et al .
1992; Devos el a1 . 1992b) ; isozymes (Hart et al . 1980; Diaz et al . 1996;
Chenicek and Hart 198?, Koebner et al . 19gg; Liu et al . l_990; petchey etal . l-990, Thiele and Seidel 1990; Liu and cale 1991, Friebe et aL. !992¡Sun and Dvorak l-992; Thiele and MeIz 1,992) i and seed proCeins (Orth and
Bushuk l-974, Âragoncillo et a1 . 19?5, Bietz et a1 . 19?5; payne eL al .
1980, 1981, 1982, t984; Brown and Flavetf 1981; Galili and Feldmân 1993;
lfackson et a1 , 1983; Fra-Mon et at. 1984, Lafiandra et al . L984, Burnouf
and Bietz 1985i Sanchez-Monge et at, 1986, Liu et aI . 1989, carcia eta1 . 1990; craybosch and Morris 1990; cupt.a and shepherd 1990, Masojc eC
a1 , 1993a),
Monosornic analysis is conunonly applied to locating genes forrèsistance to pests such as bhe rooL knot nematode (Kaloshian et al ,
199L) , leaf rust (Dyck and Kerbêr L97!, Lg'tj and 1991, Dyck et al .
1987), stern rust (McInCosh 7972 and 1978i Kerber and Dyck 1973i Sanghi
and Baker 1974; Sawhney ee al . 1981) , and yeltow rust (Kema and Lange
1992; Bariana and Mclntosh 1993, Jiazhi et. a1 . 1993). In addit.ion,monosomic analysis is used to identify the chronosome location of genes
of traits which have potential as genetic markers. For example, seed
proteins (Hueros et a1 . i.988; Sanz et al . L98B) as weLl as visualtraits such as kernel colour and glume shape (Bares and Kosner 1975) are
evaluated for chromosome focation by monosomic analysis,The identification of the chromosomal l_ocat.ion of eyespot resistance
provides an example hrhereby the association of a previously locatedtraj.t with that of a trait to be located provides evidence of thechromosomal focation of the trait to be Localed (Mena et al . 1992). The
evaluation of linkage of traits previously located to chromosomes withtraits such as RFLps which are being newly mapped allows theideneification of the chxomosome location of the newly mapped. traits(Liu and Tsunewaki 1991). This strategy also allows mapping of thet.raits previously located to chromosornes for whj.ch no lj-nkage data \,¡as

7
available by taking advantage of newly discovered associaCions ofpreviously located traits with common, for example, RFLP 1oci,
Ditelosomic or alien substiluted chromosome lines are used to locat.e
genes by segxegation analysís whereby a correlation is establishedbetween, for example, loose smut resistance and a particular chromosome
observed microscopically in the segregating progeny. This mêthod,
however, depends on much microscopy and is often restxicted to vexifyingg,ene locaCion done by othex means or for locating a grene to a chromosome
arm. This method is used to map genes for resistance to leaf rust(Rowland and Kerber 19?4, Kerber 1988), stem rust (McIntos:II L972, 1"978¡
\Iones et. al . 1991) and ye11ow rusL (Kema and Lange 1992; Bariana and
Mc]ntosh 1993).
Ànatomy
Àn understanding of the morphology and anatomy of Èhe wheat plant isimportant to understanding the disease cycle of loose smut and the
response of particular forms of resistance at the cel1u1ar level . Such
information is the basis of inoculaCion and assessment stxategies used.
for selecting for resistance to loose smut. .As a menìlcer of the
cxamineae, wheat displays the comnon grassy traies of Ehis family.Cult.ivated wheat is a self pollinated annual (Àllard 1960) . Afterpollination and fertilizacion of the ovary, the enìlcryo develops
sometimes as far as the beginning of Lhe fourth leaf (Briqqle 1967).
Upon germination, the embryo expands to expose the coleorhiza and thecoleoptile, and che second internode, the internode just above the pointof attachment of the coleoptile, elongates within the coleoptile. The
apical meristem conCinues to divide, layinq down further leaf initialsand ti1ler buds, After the plan! enters the reproductive phase, ehe
spike develops and is pushed upward with internode elongation. Anthesisnormally takes place after the spike has emerged from the boot (the ]astleaf sheach), with the lemma being forced away from lhe palea by the

I
lodicules, Cross pollination, or Che entry of loose smul spores intothe floxets, can occur at this lime (Agarwal and cupt.a 1989).
The fertilized ovary develops into the seed while the pericarp and
integuments form the seed coat of the fruiL, called a caryopsis (Esau
1977). The ernbryo devetops fu11y, prÍor !o the full development of the
endosperm (Briqqle 1967), From the standpoint of genetics of resistanceto loose smut, it is importanl to consider that. the integuments and
pericarp develop frorn maternal tissue, and reflect the genetics of the
female parent., Ànother consideration regarding genetic analysis j.s thatthe endosperm is 3n with 2 doses of each chromosome being maternal and
one paternal , The dosage effect of chromosomes, imparted on seed
proteins, can be Caken advantage of in certain tlæes of genetic studies(Uh1en and Ringlund 1987; Huèxos et al . 1988), and will be discussed.
lat.er as it relates to the use of monoclonal antibodies to seed Ðroteinsas chromosome markers.
L,oOSe Smut
Disease Cycte
The infection and invasion processes of flstiTago tritici in wheat and
UstiTago nuda in barley are si¡nilar, and have been reviewed thoroughlyby shinohara (1976), In fact, there has been much debate over Che years
as to whether or noÈ Ustj-laqo tritici and ustilago nuda shoutd be
classed as a single species (Nielsen 1972). Major studies on the
anatomy of infection are presented by Batts (1955a and b) , caskin (1959)
and, in barl-ey only, Gabor (1985) .
The loose smut fungus infecLs wheat at the time of flowering, when a
spore (or spores) of U, tritici ent.ers a floret and germinates tÕ form
haploid hl¡phae which fuse to form dikaryotic hyphae. The dikaryotichlæhae form an appressorium and infection peg on the ovary wa1l, whereby
the developing ovary is penetrated (Batts 1955b, Gabox 1985), The
fungus grows Èhrough the parenchl¡rna of ehe pericarp to the int.eguments

9
(testa) and nucellust the fungus permeates lhese lissues but, for the
most. part/ does not penetrate the endosperm. The fungus grorvs toward
the base of the ovary, where it. enters the embryo (baking about fivedays), before passing through ghe scutellum and establishing in the
apical meristem approximatêly 20 days after inoculation (Ba¿ts 1955a and
b; Gabor 1985). During maturaiion of the seed, the fungus gioes dormant
and is revitalized upon germination of the eeed.
During seedling dèvelopment the fungus grows with the meristem (Mantle
1961c), At. the time when the head is developing, the fungal mycelium
forms sporogenous hyphae, from which spores forrn by segmentation.
Nuclei fuse within the spores to form diploid nuclei. When the infectedspike emerges, nost of the tissue has been converted to a mass of dark
brown spores. The spores are disseminated by wind and rain, and are
parCicularfy suited to inpaction on narrow surfaces such as glumes
(Greqoxy 1952). Those spores which ent.er florets at anthesis start the
disease cycle over again,
EnvironmentaL Effects on Loose Smut Expression
Obtaining consj.stent and opCimum expression of disease is imporLant foraccuracy in genetic analysis. The leve1 of toose smut expression of a
particufar wheat line is influenced by genotype of the host., environmênt(Agarwal and cupta 1989, Dean 1969, Kavanagh 1"961, Tapke 1948) , and by
the genotl4)e of the pathogen, The penetrance of a trait is also oftenaffected by the genetic background in which a gene resides (ÀILard
l-960) . Minor genes imparting a 1ow 1evel of resistance (Mackenzie l_991)
may contribute to lhe penetrance of major genes. Sample size can be
adjusted to account for the leve1 of random variation for a particularexpected difference between resistant and susceptible classes, such thata real difference can be detected, Replication is required to determinethe extent of variation beyond random variation. A further discussionof the statistical approach to analyzing loose smut d.aÈa is present.ed. in

10
Chapter 4.
There is evidence that the envirorunenlal condiEions at the time ofinoculation can affect the amounC of infection. Whereas spike wetness
plays only a minor role, humidity has an influence on the 1eve1 ofinfec¿ion (Tapke 1931; Dean 1965, Loria et a1 . 1982). The importance ofhumidity in artificial inocuLation (presumably by water suspension
methods) was debated by Nietsen (1987a) . Tapke (1949) indicated,however, the significance of humidity on natural infect.ion, and thatspores on plants in drier air germinate more stowly, Tapke (194g) alsopointed out the importance of duration of f1o\,¡ering on the level ofinfection and that plants under water stress proceed through floweringmore quickly than those [rith ample water. In studies of barley, Malikand Batts (1960) came up with simifar results, showing that pollinaEionís delayed under moist cloudy conditions, and that florets remain open
longer than in dry warm weather. perhaps such a delay in ovarydevelopment is important in providing adequate t.ime for infec¿ion.Cextainly, prolonging floret opening provides more time for spores toenter,
water stress, it should be noted, is not only a function of humidity,¿emperature, and air speed, but also of the v¡ater - supplying capabililyof the soil, Care should be taken during the \.¡atering of plant.s growTl
under axtificj-al growth conditions and to be inoculated with loose smut,
so thal water stress does not become a variable associated with thelevel of infection, cregory fi-952) showed that srnut spores can be
carried in raindrops. Malik and Batts fi.960) did three tests of spore
transmission. In one treatment, water droplets carried spores and were
alLowed to impact on florets, ln a second treatmenl, dry spores \,rere
bfown onto spikes, followed by the application of water droplets;thirdly, dry spores were applied by brush or wind, without water. Loose
smut infect.ion was greater in the former two treatnents in which waierdroplets were involvêd. Inoculation techniques carried out under trigh

11
humidity and simulated light shoreers may be the bese for promoting
infection and thereby ensuring detection of susceptibility and
resist.ance.
The optimurn temperature for infection by the loose srnut patbogen isbetween 22 and 25oC (Tropova 1938r Agarwal et at. 1984). Temperatuxe
also affects the expression of loose smut symptoms, particularly incertain cornbinations of host and pathogen genotlt)es (Kavanagh 1959,
1961a and c; Ânon. 1963; Dean 1965 and 1969). The optimum temperature
for loose smut expression is around 23.C, with 1ower temperatures having
less of an effect on expression than higher temperatures. Tapke (1948)
discussed conflicting reports of the effect of the temperature to which
infected seedlings are exposed on the leve1 of loose smut at maturity.Kavanagh (1961-c) studied the effect of temperature on the level of smut
expression, and found that high temperature in some genotlt)es reduces
sporulation, The effect of temperature is most critical duringinternode elongation, with sporulation being reduced after longerexposure to hiqh temperature. Temperature is also associated withmycelial product.ion of U, nuda in culture (Kavanagh 1961c) . As the
temperaLure increased, thê growth in culÈure declined, ¡ean (1969)
provided an exarnple of low temperature reducing sporulation. Àlthough
mycelium is not eliminaEed from heads of some genotl¡pes when sporulationis absent, the heads are abnormal ,
Ànothex factor which affects the l-evel of loose smut incidence isdepLh of seeding, with shallow seeding being more favourable to loose
smut infection (agarwal and cupta 1989). The effect of seeding depLh
rnay, in turn, be relat.ed to seed size, because sma1l seeds are more
highly infect.ed than other seed sizes under natural infection condiEions(Taylor 1-927). Low soil fertility favours loose smut expression (Fromme
f920). No evidence of the effect of light quatity, or intensity on
loose smul infection could be found in the literature. In contrast tothe previously mentioned studj-es, where environment played a role in the

I2
level of loose smut expression, BatLs and Jeater (].959a) found. no
association between environment.al conditions during the growing periodof susceptible plants derived from inoculated seed, and the incidence ofloose smut.. The discrepancies between different reports an the effectof environment on the expression of loose srnut may be caused by thedifferential response of the genotlæes (Kavanagh 1961c) used by thedifferent workers.
Inoculum and Inoculations
Me¡hod and timing of inoculation are fuïther variabr-es in the evaluationof the Ieve1 of resistance of a part.icular genotlæe. Therefore, it j.s
important to understand the naLure of inocufum, and timing and. melhods
of inoculation, These were thoroughly reviewed by Nielsen (19g?a) .
Sexual recornbination ín UstiLago tz:itici occurs just. prior toinfection; therefore, variability in che leve1 of infection can occurbecause of the inoculum source used. Races of U, trit.ici that arehomozygous for virulence qenes are necessary for genetic studies on
resistance, À race homozygous for virulence loci may be obtained by a
series of passages of the isolate through a differential host. However,
the approach to homozygosity \,iill be delayed by linkage of chromatin bo
the matj-ng lype 1ocus, and because the probability of garnete fusion isrepresented by sampling without replacement (once two of the fourgameees fuse there is no choice but for the remaining two gametes tofuse) (Kirby 1984) ,
The inoculation procedure is dependenl on knowledge of the diseasecycIe. Knowing that spores pass from an infected plant Eo healthyflorets, where they germinat.e to infect the ovary and d.eveloping enbryoaround the tíme of anthesis, is critical_ to the success of artificialinoculation' fnoculation rnerhods must attempt to simulace this naturalprocess' Many different mèthods of inoculation have been t.ried over theyears (Nielsen 1987a) ; examples include the use of: inoculum susDended

13
in talc and placed in a bag over the wheat head (Loria et a1 . 1992),
spores brushed oneo the stigmas (Kilduff 1933) , dusting spores ontoflorets with the upper portion of the leruna and palea removed (Tinqey
and Tolman 1934), hj-gh pressure jet inoculation (À¡derson 1961) , partialvacuum with spores suspended in water (Heyne and Hansing 1955) , and thePoeblmann nethod (poèhlmann L945) , j-nvolving a hypodermic needle and
rubber bulb or syringe to inject a spore-water suspension into florets.The Poehlmann rûethod has a nunber of advantages for genetic sCudiès,
such as avoiding contamination, which is more difficult with the partialvacuum and high pressure jet inocufaLion methods. Furthernore, ehe
injection of spores with a syringe i6 sirnpLer and easier than the bag orbrush methods, Fina1ly, infection 1evels tend to be higher with the
Poehlmann method compared to the partial vacuum method (Malik and Batts1960).
TIMING ÀND METHoD
Timinq of inoculation is critical because a ralher narroh' infectionwindow is created by the requirement thaL spores must enter floretsaround the lime of flowering so that infection can occur, Several
studies were done to determine the opt.imum stage of inoculation.studies by Piekenbrock 092't) and Malik and. Batts (1960) favoured mid-anthesis or later for inocul-ation, whereas Tingey and Tolman (1934) and
cothwal (1972) suggested the optimum inoculaCion period to be just priorto anthesis. However, Che majority of studies point to mid-anthesis as
the optimum Lime for inoculation (Tapke 1929; Oort l_939, Ohms and Bever
1956; Bever and Ohms 1966, ,fonès and Dhitaphichit 1991) , The range intime axound mid anthesis for which optimum infeclion is maintainedvaries across the different studies, as wel_l varietal differences \,¡ere
noted in some studies (Tapke 1929; cothwal 1972). Nielsen (1987a)
considered mid-anthesis (growth stages 60-65 0n the zadocks scale) to be
a reasonable compromise as the optimum time for inoculation,

r4
Malik and Batts (1960) discovered that infection was greatest in seed
from mid-head, probably indicating that there is an optimum stage ofovary development for infection, and that flore¿s in a head of \,¡hea! are
at various st.ages of development at any one !ime, with the lower and
upper fforets on Ehe head usually being less mature than the niddle.The concentration of inoculum also influences the 1eve1 of infection(Oort l-939; Chatrath and BahI 1966; Jones and Dhiraphichit 1991) . One
gram of spores per litre of tvater is recornmended (Nielsen 1997a) . When
the Poehlmann method is used, the inoculum is injected into individualflorets on an entire spike, and at maturi¿y the seed is harvest.ed forpl.anting in rows in the greenhouse for evalualion.
Hos¿ Range
Ust.ìfago tritici, in addition Co being a pathogen of wheat, attacks a
nunber of relatives of wheat, including Aegijops spr.,, Agropyron spp,,.EJymu,s spp., Hordeum spp,, secal.e cereaJe afLd TriticaLe hexapioide, alldoEher genera of the tribe Triticeae (Nie1sen L9j3, lg7}a, b, 1985,
1987b). The host range of U. tritici has inplications on the use ofalien cytogenetic stocks in locatíng, within chromosomes, genes forresistance to loose smut, and on the potential of obt.aj-ning new aourcesof resistance for wheat from alien sources. It is important thatcycogenetic stocks be tested for l_oose smut react.ioni rel_ying on theinfection information of the progenitor stock can lead to wrong
conclusions if an alien chromosome is carrying a loose smut resistance
cenetics of. ItstíLago triticiMany races of ùstilago tritici exist and the interaction with q¡heat
tends to follow a gene-for-gene relationship (Holton 1953, Oort 1963;
Nielsen 1987a). Because of a gene-for-gene interaction, insÍqht intothe genetics of the pathogen also provides insight int.o lhe genetics of

15
resistance in the host. Furthernore, as Nielsen (198?a) pointed out,successful breeding for resistance to loose smut depends on a knowledge
of the biology of the pathogen, partÍcularly knowledge of the geneticsof virulence.
As mentioned in the discussion on inoculum, sexual reconbinalion isconsidered to cake place during spore germination, with karyogamy ordiploidization having taken place previously during the process of spore
maturacion (shinohara 19?6) , Meiosis resutts in haploid nuclei which
are distribuled within a prornycelium during spore germination, Haploidhypha format.ion, hl4)hal fusion, and dikaryotization were described indetail by popp (1955) .
The fact that. recombination occurs just prior to infection must be
consj.dered in the race concept of this pathogen (Christensen and
Rodenhiser l-940i Fischer and Holton 195?), The major implication isthat spores are not clonally propagated as are urediospores in therusts. In fact, when considering virulence loci, a sorus collected frorIL
the field represents a heterogeneous, heterozygous population if theinfectiÕn originated from the fusion of genetically different haploidhlæhae, This heterozygosity rr'ill be expressed as variation insubsequent infections of cultivars with pareicular resistance genes
(differentials), and this variation may be misconstrued as instabilityÍn the pathogen. Cherewick c-958) found variabitity remained inisol-ates even after 5-LO cycles of inbreeding,
Nielsen (1987a) considered Ehe event of spores of differing raceslanding in the same floret to be rare. An extrapolation of thisconsideration would mean racès wil] be hiqhly inbred.. One would expectthis situation only to apply when incidence of loosê smut Ís 1ow. Inone study where the incidence of two races of lJstiJ-ago nuda was high,over seven percent of infections were by hybrids (Thomas 1994). Thisrelatively híqh incidence of hybridization was found by Thomas (1928) tobe as much as 43% of the expected frequency, based on random mating of

.1"6
gametes produced by U, nuda in barley.Because of the great. deal of technical expertise and effort required
in crossing races and characterizing the progeny for virulence, reportson genetics of virulence of Ustijago are few. Nielsen (19gg) reviewed
the subject of the genetics of Ustilago ruda and U, tritici, and has
been the sole investigator in the geneÈic analysis of virulence in t/.
tritici (Nielsen L977, 79821 . In Nielsen,s work, four virulence genes
were identified, A sunmary of virulence for each race and the infectionof the differentiats is given in Tabte 2,1À (Nietsen 1"9j,1 , L982, 1987a).
Nielsen,s h'ork demonstraled lhat races possess multiple genes forvirufence and differentials possess multiple genes for resisbance.Nielsen (197?) first crossed races T1 and T2. He found that no
segregation for virulence in the pxogeny occurred on TD2 (where TD
represents U. trit¿ci differentiat (Nielsen 19gZ)) and TD3, and
concLuded the virulence was the sane in both, or that T2 has Lwo closelylinked virulence genes. Similarly, no segregation occurs on TD4 and
TD5, and here, too, the conclusion is that virulènce i.s the same inboth, or that T1 has two closely finked virulence genes. Segregationdoes occur between the TÐ2/TD3 group and the TD4/TD5 group, Such
segregation is expected from two races which qive Èhe patt.ern ofresistance and suscepCibility against. lwo sources of ïesistance thatthese tines do (Table 2.18 group tv) . Àny other comlcination of fewerthan two resistance genes produces a patlern of three susceptibte tlæes(Table 2.18 group I-I1) instead of two susceptible and two rèsistant1ines. Nielsen (1977) pointed out that there are races which have
additional but differing virulence than T1 and T2 because there areraces which give the pattern of group rv of Tabre 2,18 on differentialsTD2 and TD3 or TD4 and TD5. The comprement of the virulencè interactionis that. ?D2 and TD3, in addition to having a gene for resistance incorunon, must have a gene difference to produce the susceptible,/resistant conbination seen in Table 2.1c, Likewise, TD4 and TD5 must
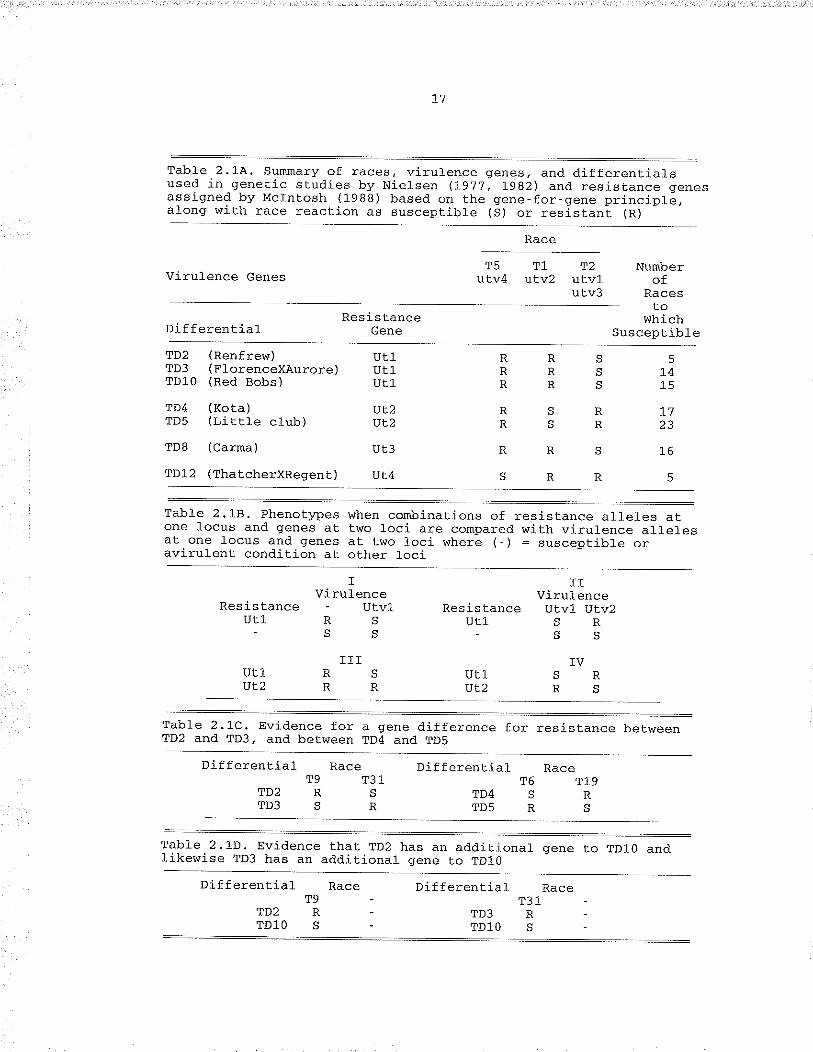
Table 2.1.A. Surunary of races, virul.ence genes, and different.ialsused in genet.ic studies by Nietsen lIg,tj, fgg2l and xesist.ance genesassigned by Mclntosh (1988) based on the gene-for-gene principlá,
Race
Virulence GenesT5 TL 12
utv4 uLv2 utvlutv3
ResistanceGene
Nurnberof
RacesCo
wnrchDi f ferent ia1 SuscepLible
s L4
R 17R 23
s 16
R5
RRRRRR
RSRS
RR
SR
TD2TD3TDJ.O
TD4TD5
TD8
(Renfrew)(FlorenceX.Aurore )(Red Bobs )
(Kota)(Little club)
(Carma)
utLut1U¿1
vL2ÍJL2
ut3
TD12 (ThatcherxRegent ) Ur.4
Table 2.18, Phenotlæes when conìbinations of xesistance a1leles aEone focus and genes at Lwo loci are compared with virufence allelesat.one locus and genes at two loci where (-) = susceptibfe oravÍrulent condition at othex loci
TVirulence
Resistance - UtvlUT1 RS
]Tvirulence
Resistance Utvl Utv2utl, s R
Table 2.1.C, Evj.dence for a gene difference for resiBtance betweenTD2 and TD3, and between TD4 and TDs
Differential Race DifferentialT9 T31
TDz R S TD4TD3 S R TD5
IVUT1 SRÍJL2 R S
rrrUT1 RSUL2RR
T6 T19SRRS
Table 2.1D. Evidencelikewise TD3 has an
fhat TD2 has an additional gene to TDLo andadditional gene to TDlO
Dif f erential,TD2
TDl O
Diffèrent.ial
TD3TDl O
T31R.
T9R.s-

18
have a gene difference. TÐ4 has an additional gene over TD5, and TD2
has an additional gene over TD3, because TD4 and ,ID2 af.e resistant ¿o
more races than TD5 and TD3 respectively (Table 2.1À) , based on person,s
(1959) theoretical analysis of resistance and virulence.Nielsen (1982) reported virulence on TD1O is lhe same as on TD2 and
TD3, indicating a gene in cornmon amo¡g the three. The pair TD2 and
TD10, as well as the pair TD10 and TD3, also differ in a resistance gene
(Table 2.1D) , and TÐ2 is likely to have one grene more than TD1O and TD3
because TÐ2 is resistant to more races than TD1O or TD3 (Table 2.1A),Virulence in races T5 and T2 on TDg and TD12 segregate from each otheras expected from the race paLtern shown in Table 2.1.å,. Virulence on TDg
also segregates from TD2, 3 and 10, indicat.ing a virulence gene
difference in race T2 conpared to race T5, and thus a differentresistance gene in TD8. The genes responsible for virulence on TD12 and
TD8 are different from the gene j_n T1, because neither T5 nor T2 isvirulent on TD4 or TD5. Therefore, based on the gene-for-gene concepb,
TD12 and TD4 differ in genes for resistance. TD12 Lost a gene duringimprovernent of this differential for agronomic traits by Nielsen and
Dyck (1988), demonstratinq that TD12 has more than one gene. Based on
Person's (1959) method of analysis, TDl.2 differs by tr\'o genes over TD4
(Tabte 2.1A) .
Iroose Smut Races and the Wheat DifferentiaL Set
Forty one races of loose smut have been identified by Nielsen (199?a) ,
thereby irnplyinq several genes for resistance. civen that 35 races Õn
hexaploid wheat exist, and assuming a gene-for-gene relationship is ineffect, at leasl six resistance genes are required for differentiation(2 to the powèr 6 = 64), Nielsen,s different.ials contain l_5 hexaploidwheat lines, although this number could be reduced to 10 with some
deletions and additions (Table 2.2). Some races have been
pooled in lable 2.2 because the incompatibility reaction is not

Table 2-2. A cond.ensed differential set of frj ticu.rn aestjyum (data collected by J. Nielsen)
Race (T- )
Differential 27 L6 1,L 34 35 37 28 5 1 7 6 23 4LL8 21, 1,7 38 831 20 9 2L2 1-5 LO 22 L3 19 39
Prelude/Diarant S S S S S S S S S S S S S S S S S S S S S S S S S S S S S
ChinookSSSSSSSSSSSSSSSSSSSSSSSSSS
îD5ASSSSSSSSSSSSSSSSSSSS
TD4ssssssssssssssTDSASSSSSSSSSSSSS
TD1O S S S S S S S S S S S S S
MoscovkâSSSSSSSSSS
SinLonSSSSSSSS
TÐ9 s s s s s s s s s
TDlsSSSSSS
TD2qc

20
considered. civen even 10 dj-fferentials, the nunìber of races j-s eitherseverely under - represented (2 to lhe power I0 = L024), or some of the
differentials possess more than one gene for resistance, and existingraces which could be further subdivided are not, or a conùcination of the
tv/o problems exists.In the same manner that dÍfferential wheat lines can identífy races,
races can be selected rvhich can dÍffêrentiate the wheat lines, fn the
same way Lhat differentials with more than one gene for rêsistance willunderestimate bhe number of virulence combinabions and thus races
(Person L959) , races not possessing full virulence minus one virulencegene will underestimate the number of resistance genes. The objectivein choosing races for genetic woxk on resistance is to use the races toident.ify the segregation of different genes for resistance. Ideally one
would like to have a set of races, wich each race conÈaining aIIvirulence genes except one, and each race deficien! in a different gene.
However, the next bèst choice is lhose races with as great a virulenceas possible, which will differentiate the differential cult.ivars. Eightdifferentiat races selected from Nielsen,s (1987a) cotlection are listedin Table 2.3. cenetic analysis to locate loose smut resistance genes tochromosomes will be made more difficult by Ehe segregation of multiplegenes, requiring the use of larger populations. Resistance t.o loose
srnut in a cult.ivar may be conferred by a single gene to which no race
with virulence is yet know¡, or resistance may be conferred by a
combination of resistance genes to which no corresponding combination ofvirulence genes is yet knov¡n in the pathogen. Because of the effortinvolved in locating genes to chromosomes, single gene lj-nes would be
ideal . Barring this ideal, the next best material- to use for initialstudies would consist of lines about which something of the inheritanceof resistance i6 known, or lines wibh a pedigree and to $'hich the known
races have been tested. civen that the latter is represented by the
differentials, these are the fiïSe choice for genetic studÍes.

Table 2.3. Races of ustilago tritici which differentiate the rriticum aestivl.¡ndifferentials (from Nielsen 19 g7a)
910 12 L4 15 16
Differential (TD- )
ssss
SSSSSSsssssssssssss
s
s
sssssss
Race
(r- )
26I
1015'L9
31

22
Many sources of resistance to all known races have been described by
Nielsen (1983), indicatinq that other resistance genes may still be
available for identificatÍon; however, cornbinations of existingresistance genes, to which the corresponding virulence combinations have
not yet been found, cannot be ruled out.
Anatomy of Resistance
From the developmental moxpbology and anatorny of the wheat plant and the
disease cycle of loose smut, five stages at which resistance can be
expressed have been identified (caskin and Schafer 1962r Loria et a1 .
1982) . Loria and coworkers (1982) talked about field resistance inwhich floret accessibility is considered the important feature. Any
restriction to the entry of loose smuL spores constitutes a fofln ofresistance by the escape of infection, and has been termed morphologicalresistance by Nielsen (personal corununication), and wil-I not be
considered further in this thesis. Mantle (1961b) , on the other hand,
talked about resistance at the tissue 1evel, through the use of lightmicroscopy, He descri.bed four tissues where mycelial proliferation isstopped. These tissuès are¡ ovary, scutellum, crown, and apíca1 growingpoint, Three of these locaÈions were examined by cabor and Thomas
(1987) in barley, namely: prevèntion of ovary penetration, enìlcryo
resj.sCance, and resistance in the seedling. Mantle (1961c) furtherobserved that the fungus in susceptiblè cultivars maintains itself verynear the apical growing point, dispelling the hlæothesis that Che
progress of the mycel_ium can fa11 behind the developmeng of the growingpoint and thaÈ the mycelium catches up to the growing point at some
later s tage .
caskin and Schafer (1.962) sunmarized the t]æes of histologicalreactions which can occur. Resistance to penelration is the second siteof defence by the host, after exclusion of spores from the fl_oret, butis the first site for physiological resistance. Ân import.ant

23
consideration j-n the study of genetics of loose smut resistance is thatthe outer layers of tissue of ehe ovary are maternal, and represent thegenot]¡I)e of the fernale. ln such an instance, a recessive gene for loosesmut resist.ance \\'ould appear doninant upon inoculation of the F1 seed.
Schaller (1949) believed a maternal tissue effecl occurs for the baxleycultivar Trebi, \,¡hich is resistant Eo U, nuda, Necrosis of invaded
cel1s occurs during the resistant reaction in Trebi (Shinohara 1976) .
Batt.s (1958b) , caskin and schafer (1962) and NíeIsen and Dyck (19g8)
observed a maternal tissue effect in wheat. It is inportant to considerthat even susceptible genotlæes become resistant t.o infecCion as theovary matures. The effect of ovary maeurity was discussed by Batts(1955b) , and is indicated in the section on Tining and Method ofInoculation in fhis review.
Resistance to ehe fungus that succeeds in penetrating the ovary can
occur at. two leveLs in relation to the ernbryo ¡ exclusion fïom the enbryo(caskin and Schafer 1962; popp 1959), and invasion of the enìlcryo butexclusion from thè plunular bud (Ohms and Bever L955, Batts and Jêater1958br Popp 1959) . These two forms of resist.ance were not discriminatedby Mantle (L961a) or popp (1951) . À fifth location of resj-stance occurswithin the seedling, in which case the mycelium does not' keep pace withthe apical meristem (caskin and schafer j.962). Even if the fungus
maintains itself within the meristern, resistance may be manifested bythe inhibition of growth and further development of the seêdling,referred to as the incompatibility reaction (oort 1944; Mantle l-961b).If the fungus becomes est.ablished in the growing point, it may be
excLuded from the inflorescence during the rapid meristematic groh'th ofthe seedlinq (Gaskin and Schafer 1962). Should the fungus succeed ininvading the tissue of the irunature inflorescence, it will be
transporCed upwards with the inflorescence as the stem elongates.An attempt to speed up the process of evaluating loose smut resistance
by evaluating embryos was demonst.rated by popp (1951 and 1959), Batts

24
and Jeater (1958b) , and caskin and Schafer (1962) . Inoculatingseedlings was demonstrated by Kavanagh (1959 and 1961b) and Jones and
Ðhitaphichit (1991) as a means of reducing lhe time from inocul_ation toinfection. Such methods, however, are unsatisfactory because only a few
forms of xesist.ance might meet the selec¿ion criLeria. In the case ofernbryo analysis, only resistance manifesLed in the maternal tissue up tothe scutellum, along the infection path, would be selected for; otherforms of rêsistance would be discarded, This method does not avoid thedifficult and time-consuming inoculation procedure, and¿ in fact, more
inoculations \4rou1d have to be done because certain forms of xesistancewould bê díscarded. More informat.ion is needed to conclude whether ornot. seedling inoculation would circumvent certain tl¡pes of resistance,
cenet,ic Stud.ies of Resistance
Sevêral studies of inheritance of resiseance to loose smut. have been
done in wbeat. Studies determining nunìbers of genes and al1eleinteraccion are summarized in Table 2.4. Many \À¡orkers agree that it isnot easy to determine the nurnber of genes involved in xesist.ance t.oloose smut, alChough some workers do indicale the nurnber of resistancegenes. In large part¡ the difficult.ies arise from not being able toclassify segregating progeny clearly as susceptible or as resistant.compared to detèrmining the nurnlcer of segregating genes, allelicrelationships are perhaps not so difficult to establish, based on theinoculation of F1 (crossed seed) or segregation in the F} of course,the particular race or races used in a study will influence the outcome,
because only those genes in wheat whiÕh are interacting with theavirufence genes of the pathogen will be observed. Metzger et al .
(1979) demonstrated the use of multiple races for evaluating Èhe geneticnature of conunon bunl resistance. Such a procedure can be easilyadapted to lhe evaluation of loose smut resistance by inoculatingdj-fferent heads of the same plant \eith different races, More

Table 2.4.
Cross
Grune Dame* XRun]çers Dickkopf
andRimpaus red schlanstedt
Grune Dame* XRoter Schlandstedter
Kota X Red Bobs*Kota X Garnet*
San Mart.in X 38 MA*
Hope* X FederationHope* x Dicklow No. 3Preston* X 01,-24
Pansar III* X -
Federation,/Khapl i * XMentana
vilmorin 2? X Jubile*Thatcher* X Redman
Dundee 48* X RaneeWhite Federation* X Ranee
Kawvale* X Clarkan
A sunmary of genetic studies of resistance to 1oose smut of wheat
Dominance Nunlcer of Genes ReÊerenceof Resistance Segregating
Recessive
Recessive
Recessive
IncompleteDominaûce
Dominant
Dominant
Dominant
Dominanl
DominantDominant
Dominant
(continued on next paqe)
Piekenbrock 1927
cxevel 1930
Kilduff L933
Rudorf and Rosenstiel 19 34
Tingey and Tolman L934
Larose and Vandervalle L937
Milan 1938
Milan 1939
Larose and Vandervalle 1937
caîpbe]l 1948
Pugsley 1953
Heyne and Hansing 1955

Table 2 .4. continued
Todd* X Knox orKawvale* X ClarkanRichelle* X J
Hope-Hussar* X KnoxPI19L533 * X KnoxKawvale* X Kr¡oxTremexino* X KnoxRieti* x Knox
NP824* X Rj-o Negro
NP775 X NP798 *
Cappelle* X Hybrid 46Picardie* X andKota* X Intercrosses
NP79O* X NP775
Bexencukskaya 98*Saratovskaya 36*Saratovskaya 29*
Kenya 340Y4A1 X IÀs52
Partlyrecessive
J
RecessiveI
I
I
¿
Dominant
Dominant
DominantIJ
Dominant
Ad.diLive
Dominant* Indicat.es resistant parent.- Indicatês information \^¡as not avaitable.
2
11,
22
1,
111
1
3
2
Richards 19 61
Gaskin and Schafer 19 62
Mathur and Kohli 19 63
Agrawal et al. J-9 63
Ribeiro 1963
Àgrawal and Jain L965
Shestakova and vj ushkov L974
Medeiros 1976

2'l
ofinformation about the segregation resislance genes can be obtained by
multiple race inoculations,Because there is no sEandard race classificalion for 1oose smut, it is
difficult to compaxe the conunonality of resistance genes in published.
studies. However, data for doninant and recessive modes of al1e1icinteraction show lhat recessive alleles occur frequently. Workers
report on both dominant (Tingey and îolman j-934r Milan j-938, 1939;
Pugsley 1953r Richards 1961, Mathur and Kohli 1963; Ribeiro 1963,
Àgrawal and,fain 1965; Nielsen and Dyck 1988) and recessive alleles(Piekenbrock 1927; Gaskin and schafer !962ì Aqrawa:- and ,Jain L965,
Nielsen and Dyck 1988), with some expressing complete resistance(irununicy) (crevel 1930, Rudorf and Rosenstiel 1934). oehers have
demonsirated incomplete dominance (tingey and Tolman 1934; Richards
1961) , white Campbell (1948) attempted to study i.nheritance ofresisbance of parents in which the susceptible parent had some level_ ofresistance, The study of moderately expressive genes allows for thestudy of gene interaction with xespect to a given race. Shestakova and
Vjushkov (1974) ïeported on additive gene action (althouqh the souïce
does not indicate v¡hether ox not such action was inter- or intragenic orboth) , while Kilduff (1933) reviewed reports describing genes havingcumulative effect.s, a condition also mentioned by Tingey and Tolman
(1934), Heyne and Hansing (1955) talked about minor genes. The
existence of minor genes and genes with cumulative effects indicatestbat conbinat.ions of genes can resulL in a greater level of resisÈanceto a given race lhan a single noderately expressive gene alone. Thisinformation, if true, would validate tbe development of genecic markèrs
to minor genes for the purpose of pyramiding the genes.
There are two reports of the inheritance of Che incompatibilityreaction, Ribeiro (1963) reported that incompatibility in Cappelle,Picardie, and Kota is inherited as a single dominant gene, and Medeiros(1976) indicated incompatibility in Kenya 340y4À1 is conferred by a

28
single dominant gene.
Dhitaphichit and cor,¡orkers (1989) studied subst.itution lines of wheat.
They demonsbrated that resistance to race T6 may be influenced by genes
in a nurìlcer of chromosones in the wheat cultivars Hope and Thatcher, butthat the most effec¿ive resistance in Hope resides in chromosome ?À, and
j-n Thatcher in chromosome 78, The Hope and Thatcher genes are ofinterest because of tbeir potential use as sources of resistance inbreeding, and ¿heir potential durability, Hope resistance is considered
to derive from Vernal emmer, whereas Thatcher resist.ance derives frorn
Iumillo durum (Nielsen 1969).
Ribeiro (L963) recommended that genes for loose srnut resistance inwheat be designaeed ,Ut, and Mclntosh (1988) used these slmìbols t.o
characterize pulat'ive genes, based on the resisÈance-virulence
interact.ion worked out by Nietsen (1,977 ar:rd f9g2) .
Genetic Markers
The advantages of knowledge of the genetics of resistance to loose smut
to breeding for resistance have been pointed out in the int.roduction tothis disserEation. The chromosomal Location of a gene is one aspect ofgenetic knowledge which can assist in the determination of genetic
markers located in tbe samè chromosome. Improving the efficiency ofcytological analysis by using biochemical chromosome markers can speed
up the rate aE which genes such as those controlling l-oÕse smut
resistance can be located in chro¡nosomes (Ànderson et al . 1992). Thissection provides a discussion of genetic markers because of their roleas chromosome markers in cytogenetic analysÍs, and because of Cheirpot.ent.ial as markers to genes for loose smut resist.ance. À more
detailed discussion of the use of chromosome markers for locating genes
to chromosomes is left to Chapter 3.
The phenotlæè of any gene previously located to a chromosome can be
used as a genetic marker for the chrornosome in which it resides.

29
Ho\qever, ehe most useful genetic markers are lhose which are easy toobserve, other than a few simple morphological traits, the easiestmarkers to observe are sêed proteins vísualized by electrophoresis.This is becausè seed proteins can be sampled by taking a portÍon of theendosperm, leaving the embryo end of the seed inlact and viable (Bietz
et aI . 1975; du Cros and Hare l_985) . In addition, the seeds do not have
to be grown until the t.est results are known, saving the effort and
space involved in plantinq the seed and growing the plants. The
analysis of seed proteins with antibodies is especially useful .
Artibodies exist which discriminate between proteins (Skerxi¡t and
Underwood 1986; Dawood et al . 1989; Donovan et al . 1999, Skerritt and
Robson l-990; Zawistowski et a1 . 1992; Brett et al . 1993; Graybosch eC
a1 . 1993; Masojc eL a1 . 1993b) , The presence or absence of a proteinindicates the presence or absence of a gene, and perhaps even alle1lcforms of the protein coded by a gene at a particular locus, Monoclonal
aneibodies have the added advantage Õver electrophoresis ofdiscrÍminating among l-ines having different gene dosage. lsozl¡mes,
disease reaction, and DNA poly orphism can also be used as markers.
Seed Proteins
Because of the ímpact of seed proteins on quality, a great deal_ ofresearch has been done on !hem. In facb, several reviews are avail-ableon the subject of seed proCeins (carcia-Olmedo et al . 1978 and 1992;
Payne and Rhodes 1982r Kreis et a1 . 1985r Shewry and Miflin 1995; pal¡ne
J.987; de Man 1.990, MacRitchie et a1 . 1.990r TaEham et. al . 1990).
Seed proteins are classified based on differential solubilities of thedifferent protein components, and specific polypeptides are identifiedby electrophoretic procedures which separate thê polypeptides based. on
size and chargê, It is useful Eo study the chemistry of these proteinsin a little more depth if they are to be used effectj-vely as chromosome
markers. De Man (1990) described the Osborne classif.ication of \,rhea!

30
seed proteins, whêreby proteins are classified and named based on
solubility. The Osborne sysLem is as followst Atbumins are water
soluble prot.einsi clobulins are neutral salt solubte proteins; profamins
are aqueous-ethanol ex¿¡actable; clutelins are soluble in diLute acid oralka1i, Shewry and ¡tiflin (1985) considered an addibional subclass ofproteins the CM or chloroform/methanol soluble proteins, which, as the
name implies, are soluble in chloroform - rnethanol . These proteìns are a
subclass within the protamins. As welf, Shewry and Miflin (1985)
grouped the glutenins as prolamins because the glut.enins are soluble inaqueous alcohol, tùith the distinction between the glutenins and obherprolamins being that a reducing agent is required fox solubility of theglutenins. Solubility of gliadins (a class of prolamins) is notdependent on the pxesence of a reducing agent. The glutenins containcysteine which can participate in intermolecular disulphide bonds
(shewry et aI . 1992). cliadins, which possess sutphur containing amino
acids (sulphur rich gliadins), form intramolecular disulphide bonds,
?he glutenins and gliadins combine to form glutèn.
îhe important consideration for using seed proteins as molecularmarkers is ho\,¿ they can be separated and visualized. Normally, theglutenins are separated by SDS-pÀcE (Sodium Dodecyl suÌfatePolyacrylamide ce1 El ectrophores i s ) , which also allows an estimation ofmolecular \,¡eight of proteins (Bietz et al . 1975; Ng and Bushuk 1997).
The remaining groups of proteins are normally sêparat.ed using acid-pÀGE
in a one step system (carcia-Olmedo et al . 1992). However, because ofthe large nunìlcer of proteins present in seeds, various modificationshave been applied both to extraction procedures and separatingprocedures, including two-dimensional methods, These will not be
discussed further because this thesis emphasizes the rapid screening oflines based upon markers that can be identifièd by one dimensional_
elec trophores i s , Resolution during the separation of qliadins isoptimized when the gefs are run at between 7 and 1O.C (Lookhart et al .

31
1985) . Firm gels are obtained by lotvering the tevêls of caCalyst (Khan
et al . 1985), and when acrylamide concentration is in the range of 11 to16% (craybosch and Morris 1990).
The albumins and globulins usually coincide with many of the seed
proteins whích have metabolic and strucLural activity (Shewry and Mlflin1985; Gupta et a1 . 1991) ; whereas the gliadins and glutenÍns are
associated with the starch macrix, and, as such, provide a role innitrogen, carbon, and sulphur storage. Because of the diversity ofalbumins and globulins, two-dimensional separation often has been the
only way to characterize the individual components (Aragoncillo e! al .
1975; Fra-Mon et al, 1984, Sanchez-Monge et al , 1986; Forsyeh and
Koebner L992), Therefore, more work needs Lo be done to improve
resolution in a one-dimensional systen.
Locating genes coding seed proteins to chromosomes is an impoxtanL
area of research. Reviews on this subject were presented by Garcia-Olmedo and coauthors (1978 and 1982). Gliadin loci are located in che
group l- and lhe group 6 chromosomes (Brow¡ and F1avel1 1981; Ga1i1i and
Feldman 1983, Lafiandra et al . 1984; payne et al . 1984, craybosch and
Morris 1990). Trans - tegulation may occur, considexing Howes et a1 .
(1990) found suppression of band 45 in prelude and desuppression of band
40 in Tet.ra Preludei however, this could also be explained by post-
translational processing. Brov¡n and Flave1l (1981) found thatchromosome 2,4, affects the synthesis of gliadins genes in chromosome 6D.
The hiqh molecular weight gfut.enins are located in the group 1
chromosomes (Orth and Bushuk 19?4; Bietz et a1 . 1975; payne et al . 19g0,
t98f, 1982), although Bietz et at. (1975) indicated such a locus alsooccurs in 4D, Loh¡ molecular weight glutenins are coded by group 1
chromosomes (.fackson et al , 1983; Burnouf and Bietz 1995; cupi:a and
Shepherd 1990) ; whereas globulins and albumins are locaeed in a varietyof chromosomes (Aragoncillo et al . l_975, Bro\a'n and Flavell 19gl_, Fra-Mon
et a1 , j.984; Sanchez-Monge et a1 . 1986; Liu et a1 , 1989i carcia et a1 ,

Table 2.5. ChromosomeGarcia-Olmedo et al.
Chromosome
1À1B1D
'A
2D
3A3B
4À4B4D
and seed proteins associated with that chromosome. (Adapt.ed fron(1,978, 1982, with additions)
cliadin (s), clurenins (L), purothionins (L)cliadin (S), clutenins (L), purothionins (L), Globutincliadin (s) , clutenins (L) , purorhionins (t,)
Gliadin, Alpha - amylase inhibitorAlpha-ajnylase inhibitorcliadin, Àlpha- amylase inÏ¡ibitorAlbumln (S)Albumin (S), Alpha-amylase inhibiror (S), GlobutinÀlbumin (S), Alpha-amylase inhibitor (S), clobulinÀlbumin, clobulinCM, Àlbumin - HMw {L)CM, Albumin - IIMW (L), clobufinA.lbumin - HMW (L) , PurothioninAlbumin (L), PuroLhionin, clobulinAlbumin (L), Purothionin
Gliad.in (S) , AlbuminGliadin (S), GlobulinGliadin (S), Alpha-anylase inhibitor (s), clobulinLMW gliadins (S), Albunin (r,)CM, À.1bumin (L), clobulinCM, LMW gliadins (L), Albumin (L), Globutin
Protein ( chrornosome arm location)
5À5B5Ð
6A6B6D
7L7B7D
HMW = high molecular weight, LMw = low molecular weight, cM = chtoroform-methanor,long arm, S = short arm.

33
1990; Forsyth and Koebner 1992; Mâsojc ef al . 1993a) . Each chromosome
is know¡ to be associated with at. leasl one lype of seed protein (Table
2.5). Therefore, it is possible to have a seed protein chromosome
marker for each chromosome in wheat..
Àn impori:ant requirement of a genelic marker is that it. demonst.rate
al1e1ic variation. Àlthough alletes of the HMW glutenins have been
sorted out (Larerence and Shepherd l-981; payne eh a1 . j-981, payne and
Lawrence 1983t cupea and shepherd 1988), much work remains to sort outthe alleles of other tlæes of seed proteins.
MonoclonaL Àntibodies
Monoclonal antibodies (MÀbs) are antibodies derj-ved from a clonal lineof b - lt¡mphocytes thaE have inunortality as a result of being fused witban immortal myeloma to produce a hybridoma (Milstein 1990; calfre and
MilEtein 1981), .A,ntibodies produced by such cells aïe often hiqhlyspecific. I'IÀbs are created by purifying an ant.igen, such as a seed
storage protein, which is injected into a mouse to stimulate an inmune
response. The b-celIs are extracted from the spleen of the mouse, and
cultured and fused with myefoma ce1ls. The resulting clones continue todivide, and rnost continue to produce antibodies. These cells are then
diluted out, and colonies derived frorn single cells are screened forspecificity using, for example, an enzl¡me linked inìrnunosorbant assay(El,IsÂ) . There are many general references such as Klein (1990) whichprovide the fundamentals of imnunology. Monoclonal antibodies inparticular are discussed by Milstein (1990) and calfre and Milstein(1981) . Inmunology as it applies eo certain aspects of cereal researchis reviewed by Vaag and Munck (1997) , and skeïritt and coauthors (1990) .
However, the experimental work reported in this dissertation is mainlydirected at the exploitation of existing MAbs through their use as
genetic markers, and further discussion will be focussed. on monoclonalantibodies to seed proteins as chronosome and qenetic markers.

34
Because advantagès exist fxom using seed prot.eins as genetic markers?
and because of thè convenience and rapid testing through the use ofmonoclonal ant.ibodies (skerrítt et al . 1990) , applying monoclonal
antibodies to seed proteins as markers takes advantage of the benefitsof both (Konarev 1981) . To date, tdÀbs as chromosome markers are limitedto the detection of alien rye and Agropyron chromosome segments in wheat
(Howes et. aI . 1989, 1993; Brett et at. 1990).
Antibodies have been produced to a nunber of seed proteins rangingfrom gliadins to alpha-amylase inhibitor proteins (Skerritt and
Underwood 1986, Dawood et a1 . 1989; Donovan et al , 19g9; Skerritt and
Robson 1990; zawistowski et a1 . l-992, BretL eC a1 . 1993, craybosch etaI . 1993; Masojc et al . 1993b). The bínding specificity of antibodiesvaries, but for antibodies to be used as chromosome markers thefollowing qualities are usefulI The antibody binds tÕ most if not. allstrains of wheat possessing the chromosome; The antibody has a highaf f j-nity and specificity for Lhe protein, with 1iEtle cross reaction toproteins coded by other chromosomes; The level of binding is hiqh,allowing quantification. One of the advantages of such an antibody overelectrophoresis is ehat the protein content can be quantified (fannet.Ca
et al . 1993), v¡hich allows Che discrimination of g,ene dosage (Skerritte! aI , 1987, Masojc et al . 1993b). such discrÍmination al1ows thedetection of heterozygosity, or t.he dêtection of a single dosê of a
chromosome. ln the case of a complex elêclrophoretic gel pattern, orwhere a nulf allele exists at a locus, or where two cultivars in a crossshare the sarne a1lelic form of a prolein used as a marker,
electrophoresis may not a11ow discrimination between a heterozygote and.
a homozygote,
skerritt and Robson (1990) pointed out bhat antibodies made toglutenins often cross-react with glutenins coded by 1oci in otherchromosornes. MÀbs raised agaj.nst total gl_iadin exCracts range inspecificities to gliadins, with somè even binding to gÌutenins (Mills et

35
al . 1990; Skerritt et a1 . 1991) . The t}æe of immunoassay (e.q. ELISÀ)
affects the 1evel of cross-reactivity (skerritt and Hj_ll 1990).
Furthermore, the tlæe of extractant. and solid phase affects crossreactivÍty.
The poeential exists for antibodies developed Lo other cereals to be
used in wheat. Singh and coworkers (1989) found that an antibody to an
oat globulin also binds to triticin of wheat, Skeïrit.t and Lew (1990)
found antibodies to wheat qliadins bind to proteins from rye, oats, and
barley. In fact, 230/9, a\ antibody discussed in Chapter 4, is arnong
this group. Àlthouqh cross-reaction of MAbs to proteins coded bydifferent genes is undesirable for use as a chromosome marker, beingable to draw on antibodies developed to other cèxea1 grain seed. protej.nsexpands bhe potential- batCery of antibodies availabl-e for a particularLask, if conditions can be manipulated to obtain the desiredspecificiLy,
DNA and Isozl¡me Markers
Isozymes as genetic markers offex great diversity, which is an advangage
for their use as genetic markers (Koebner et a1 , 198g, McIntosh 19gg;
Liu et aI . 1990; Petchey et at. 1990; Thiele and Seidef 1990; Liu and
Gale 1991r Sacco et a).. !992t Sun and Dvorak 1992, Thiele and Melz
1992) . However, as pointed out by Sharp and coworkers (1999), isozl¡me
analysis requires a diversity of methods be used to evatuate each
marker. Because of this difficulty with isoz!¡mes, DNÀ markers, withtheir uniformity of analysis and potentiafly greac diversjty, have
greater pot.ential as genet.ic markers.
A nurìber of Elæes of DNÀ marker strat.egies on cereal crops, includingsequence tagged sites (Tragoonrung et al . 1992), random amplifj-edpolymorphic DNA (RÀpD) markers (Devos and cale !g92ì He et al . 1992;
Dweikat. et al . 1993), variable nunber Candem repeats (VNTRs ) (Talbertand CLack 199L; szurmak and Dobrzanska 1993), and restriction fragment

36
length poll¡morphism (RFLP) markers (Chao et al . 1988, sharp et af.1988a), have been evaluated. The fevel of polymorphism for DNA markersis lovr in wheat. However, some researchers have found that. denaturinggradienl gel electrophoresis improves the detection of pol]morphism inRÀPD markers in wheat (He et al . 1992, Dweikat et al . 1993). Because
RÀPD markers are a more recent development, the pool of useful markersis 1ow, and map development has not be as g.reat as v/ith RFLP markers.In fact, a nunìlcer of maps in wheat of various tlæes has been produced
using RFLP markers (Chao et a1 , 1998, 1989; Sharp et a1 . 19gga, 1999;
Gil.l et aI . 1991r Heun et. a1 . 1991; Liu and Tsunewaki 1991, .A,nderson etaI. 1,992t Devos and cate 1993; Devos eC al . 1993). RFLps were chosen as
markers as part of the research for this thesis because of the knowledge
of their chromosome location, and, in some cases, their map location has
already been determined. Mapped RFLps allow their selective use inLinkage analysis t.o loose smut resistance genes which have been locatedto particular chromosones.
Generally, RFLps make good chronosome markers because there is oftenpolymorphism among genomes when no poll¡morphism occurs within a gienome
(Sharp et al . 1988a, 1989r Talbert et al . 1991, Ànderson et al , 1992).In fact, cale and coworkers (1989) listed a nurnber of applications forRFLPg in cytogenetic analysis. Many of the polymoïphisms detected areconsidered to be caused by deletions or insertions of DNA. in thenoncoding region associated with the gene which the RFLP pïobe detects,as opposed to point mutat.ions which affect resLriction sites (Sharp etal , 1988b; Sabelli et a1 . 1992), particular restriction enzl¡mes arecapable of producj-ng poll¡morphism, whereas others will not (Chao et al ,
L989; Graner et al . 1990) . Àmong the most informative enzl¡mes in wheatare Eco RV, Ssp I, Xba I, Hind III, and Dra I. Some probes detece onlysingle-copy sequences, whereas others detect multiple- copy sequences(Chao et al . 1989). Such mulbipl-e-copy sequences may be due to homologyof sequences of homoeologous chromosomes, or due to multiple copies of a

3',?
gene within a chromosomê (Chao et aI 19Bg; Liu and ?sunewakj- j-991_,
Anderson et al . 1992),

3B
CIIAPTER. 3
The Application of Chrornosome-specific
Monoclonal Antibodies to Wheat Genetics
ABSTRACT: À monoctonal antj-body (p248) to a \dhear gliadin proLein
coded by a gene in the short arm of chromosome 1B was used as a
chromosome marker. Sonatic chromosome nunìber, as predicted by the levelof bindinq of the anEibody to extracts from seeds of single cross F2 and
testcross F1 populations, was confirmed with SDS-pÀcE qelelectrophoresis and root-tip analysis. Thè implications of the use ofmonoclonal antibodies as lools for cytogeneeic analysis are d.iscussed.
TNTRODUCTION
Cytogenetics has played an imporbant rol_e in wheat breedinS, both intransferring new sources of stress and disease resistance from related.wild species, and in identifying the chromosome locaEion of specificgrenes once transferred (ÀIlard 1960; Knott 1997b; Mclntosh 1987).Locating a gene eo a chrornosone requires the analysis of progeny fornullisomic, monosomic, and disomic chromosome composition, as well as
evaluating the expression of the st.ress or disease rêsistance phenotjæe.However, cytological analysis is time-consuming (GaIe et aI . 19g9), thusthe method is limited to the study of relatively smal1 populations,Biochemical markers are available to aid in identifying the presence ofspecific chromosomes. These includè DNA probes (Chao et a1 . 19g9; Sharpet al , 1989), isozymes (Masojc and cate 1990), potyacrylamide ge1

39
electrophoretic kernel protein bandj_nq patt.erns (Bietz eb al . l-975), and
monoclonal antibodies (MÀbs) (Horves et. al . 1989 and 1993). The use ofbiochemicaL markers to track chromosomes promises to increase the
efficiency of cytogenetic anafysis (ca1e e¿ a1 . 1989). MAbs to seed
storage prot.ej-ns are very appealing as markers because they are easy touse and allow selecti-on to be done on sèeds rather lhan at some laLerstage in plant devetopment., fn \,¡heat, t,labs specific to endosperm
gliadin proteins coded by the 1BS chromosome arm have been used torapidly screen ['heat lines for ¿he presence of the 1B],/1RS wheat-ryetranslocation (Howes et. al . L989),
Monosomic analysís has been used extensively in wheat to locatedomínant genes, such as disease resistance, Lo specifj.c chromosomes
(McIntosh 1987). This method requires the anatysis of sufficient.numbers of progeny to distinguish aberrant genetic ratios, such as those
deviating from a 3:1 in the case of a single (dominant.) gene segregation(Dyck et aI. !987¡ Kaloshian et al . 1991) . ceneeic studies ofresistance to some diseases, such as loose smut, requires progeny
Eesting using F3 or testcross F, families to provide sufficient data toaccorunodaee problems such as disease escapes or lo\,¡ penetrance of thetrait (Thomas and Metcalfe 1984). Àlthough the amount of cytologicalanalysis is not extensive, population sizes required to characterize thesegregation of the disease resistance traiC are unwieldy, crossing a
resistant parent to a nullisomic or monosomic sêries, and classifyingselfed or testcross progeny as nullisomic, monosomic, and euploid tl4)es,requj-re rel-atively few of these plants in a progeny test to establish a
correlation between the presence or absence of the trait. of interest and
presènce or absence of a particular chromosome. Nevertheless, thelocating of genes to chromosomes has been restricted to those that aresimply inherited with high penetrance and high expressivity, because offarge population sizes otherwise required for either cytogeneticanalysis or segregation analysis, Rapid identification of chromosomes

40
wiCh markers will improve the efficiency of cytogenetic anafysis oftrait.s which are less easily studied by conventional means.
fn thj.s study, the usefulness of a MÀb Eo identify nuLlisornic,monosomic, and disomic lines for the 18 chxomosome was evaluated. The
MÀb procedure was verified \,¿i th polyacrylamide gel electrophoresis and
convencional cytogenetic cechniques.
MÀTERIAIJS ÀÀTD METHODS
PLant, Mat,eriaL
Five lines listed in Tabte 3,L were used in crosses to produce
segregating populations for examination with the MÀb method. ofchromosome analysis, The crossês were as follows: Roblin N1B/pre1ude,
Roblin N1B/Canthatchi/ / prelude/Diamant, Robtin N1B/TD6//ereluð.e. The twotestcross popuÌations atlowed independent verificalion of the MÄb,s
ability to quantify protein as a reflection of 18 chromosome nurnlcer byproviding differences in seed protein electrophoretic banding pattern.Às demonstrated in Table 3.1, prerude and. TD6 differ in their chronosome
18 alleles for hiqh molecutar weight (HMw) gtutenins. As wel1, prelude
is missing a lower molecular weight band present in TD6.
Table 3.1, Somatic chromosome nunìlcer,pattern, and reaction to MAb p24B offor cytogenetic analysis
electrophoretic bandingcultivars used in crosses
Cul tivar SomaticchromosorneNurnber(2n=)
HMW Binding byGlu¡enin l.ÍÀb p24B to1Bl, bands 1BS qliadin
Roblin Nulli 1B (801-L)PreludePrelude,/Diamant F6CanthatchTD6 (Pr69282)
4042424242
null7+87+9'l+97+9
++++
- = no binding, + = binding occurs,1BS = chromosome 18 short arm, LBLnul1i = nutlisomic,
HMW = hish molecufar weight,= chronosome 18 long arm,

41
SimÍIarly, the line Prelude/Diamant is missing the lower molecular
weight band presen¿ in Canthatch. The Roblin N1B (nullisomic forchromosome LB) used in these crosses was identified frorn a population offive thousand half kernels of breeder seed of Robtín (Campbêfl and
Czarnecki l-987) exa¡nined with IrÀb p248, In all crosses Roblin N1B was
used as the female, and in the three-way crosses the F1 plant was used
as the female. Ànothe¡ line identified from lhe Roblin breeder seed
population designated 796-l- was used as a Low MÀb binding conerolinstead of the nullisomic 18 because of scarcity of seed of this lat.ter1ine. Parents, F1 plants, and testcross F1 plants were girown in a
grov¡th cabinet at 18 and 15"c on a 16 and 8 h day/night cycle.
Enzlme lrinked. Im.munosorbang Àssay (ELISÀ)
The procedure for ELISA determinations was the same as that describedpreviously (Howes et al . 1989), with the foltowing modifications. The
propan-2-o1 extracts t{ere diluted 1:15 in T0% ethanol, and triplicate100-uL aliquots were bÕund t h at 25oC and rinsed in 20 mM TRIS - 0.15 M
NaCl - Tween B0 (TBS-Tween) , Blocking and MÀb p24B binding was as
described previously. Bound MÀb was detected with a 1 in 2000 dilutionof affinity purified goat anti-nouse alkaline phosphatase (Bio-Rad
Laboratories), and developed with substraCe solution (0.5 mg/ml p-
nitrophenyl phosphaEe, 1- M diethanolamÍne pH 9.6, 0.5 mM Mgclr) 1 to 16
h uneil the cont.rol samples gave readings of one to two absorbance unitsread at. 405 n¡n. Neepawa or prelude was used as the control within and.
across ELISÀ plates.
sodiun Dod.ecyl Su1fat,e.polyacrylanide ce1 ELectrophoresis
Sodium dodecyl sul fat e - polyacrylamide gel electrophoresis (SDS-pÀGE) was
pexformed using the method of payne and Corfield (19?9) , but using an
1L% acrylamide - 0.05% bisacrylamíde separating ge1 for the RoblinNlB/Prelude population, and 17% acrylamide - 0.08% bisacrylamide for the

42
Roblin N1B/canthatch/ / prelude/Ðiamant and Robfin NI.B/TD6 / / prelude
populations. Endosperm pieces, following extraction with propan-2-01,
were extracted with 150 uL 16% dimethyl formamide - 5% 2 - mercaptoethanol- 0.062 M TRIS pH 6.8 - 2% SDS (Smith et al . 1986) at 6ooc for t h.Extracted samples (30-40 uL) were applied to gel slots. Neepawa was
used as a reference for the identification of HMw glutenin bands (Ng and
Bushuk 1987).
After el ectrophores i s, the gels were fixed in 12% TCA for 1-3 h,stained in 0.04% Coomassie Brilliane Blue R250 in 5O% methanol - 10%
acetic acid - H2o overnight, rinsed 4 h in 50% me¿hanol - l_O% aceticacid - H2o, L5 min in Hro, stained in pAcE 90 stain (Blakesley and Boezitg'l'l\ 1,6 h, and cleared in 2O% ammonium sulfate solution, photography
was performed with cool white fluorescent. back lighting, using an orangef i1t.er.
Cytogenetic Analysis
Root-tip analysis was done on a sample of plants from the RoblinN1B,/Prelude F,, Roblin N1.B/TD6 / / prelude testcross Fl, and Robl-in
N1B,/Canthatch / / pr elude/Dlamant testcross F, populabions, Embryo ends ofseed were placed on rnoist sand for five days. Root tips were collectedín ice water, held aL o-L"C for 22 h, tberr fixed in a 3¡1 solution of95% ethanol to glacial acetic acid, and stained wíth Feulgen solutionfollowing hydrolysis in 1M HCt for I min at 6OoC, The root tips were
squashed in 1.5% acetocarmíne solution, and the morphology and nunìber ofmitot.ic chromosomes v¡ere recoïded.
RESULTS
IsoLatÍon of Spontaneous 1B NullisomícOut of the 5,000 Roblin breeder seed kernels analyzed by ELISÀ, L2
kernels were identified as having Iow or zero bindinq to MÀb p248.
Eighe out of the 12 kernels germinabed, bu! two failed to develop a

43
shoot, and of Che six seeds that grew, four were normal (2n=42), withprogeny kernels producing a normal binding reaction to MÀb P248, One
pl-ant, designated 796-1, had 42 chromosomes, bue none of the progeny
kernêls reacted to M-Ab P248, SDS-PAGE showed that 796-L lacked 1BS
gliadins, but had 1BL HMW subunits 7+8. Root tip analysis of this tineproduced no visual evidence of a deleLion of chromatin, however a
mutation or small intrastilual deletion in the qliadin locus could
explain lhe resulls. The other plant was monosomic (2n=41) with three
satelliLe chromosomes, and setfed progeny kernels segregatêd for MAb
binding. Of the low-reacting progeny kernels, most were monosomic
(2n=41-) but one was nullisomic (2n=40) , and SDS-P,AGE sho\4red L¡lat the 1BS
gliadi.ns and 1BL HMw glutenins were absent in the nullisomic. This
plant, designated 801-1, had a slower growth rate and a lower seed set(five kernels,/spike) than Roblin (27 kernels/spike), The 801-1 line was
used as the female parent in Che Roblin N1B/Canthatch cross, whileselfed seeds confirmed to be nutlisomic 1B (chromosone 18 ischaracterized by its large satellite, very similar to chromosone 68, yelthe 1B satellitè is smaller and its short arm is shorter than that of68) were used for the Roblin N1B/TD6 and Roblj-n NlBi/prelude crosses.
Robtin N1B/Frelude F2 À¡raLysis
F2 seed of the cross Roblin Nl-B/Prelude was examined bo determine the
ability of the ant.ibody ¿o distinguish aneuploid línes within a
segregating poputation, One hundred and sixty F2 half kernels were
examined by ELISÀ using MAb P248, resulting in a range of absorbance
vafues which are summarized in Fig. 3.1 (in Fiq. 3.1 and subsequent
histograms, dividinq marks on the horizonEal axis represent boundaries
of the class intervals, and the values between the interval boundaries
represenb. bhe class midpoints) . À farge peak was apparent over lherange of intermediate rel-aCive absorbance readings, while srnaller peaks
were observed in each tail, Ten kerneLs tchich had 1ow antibody bÌndinq

18
oT,-o loc)
l1)14
Nt!
o
b 10-oCRl-z
6
0.0 5 0.21
Fiq. 3,1. Relative bindinqext.racts of F, half kernels
o,37 0,53 0.61 0.85Absorbonce (+O S nm)
(to Prelude) of M.Ab P24B to 50% propan-2-otfrom Ehe cross Rob1in NLB/prelude,
reactions determined by low relalive absorbance values, and five kèxne1s
l'hich had intermediate antibody binding reactions delermined by
int.ermediate relative absorbance values, were selected for SDS-pÄGE.
À11 five of the intermedia be - reac Cing kernels had HMW subunibs 7+g
characteristic of the prelude chromosome LB (Fiq. 3.2). Six of the LO
1ow-reacting kernels did not have HMW subunits 7+9, indicating Che
absence of chromosome LB, while the others (except one) had fainterbands for HMW subunits 7+8, relative Co the euploid control-, suggestingthey were monosomic 18. The ineensity of bands is in part rel_ated. tothe seed size used for extraction. qhe one extract expected to have av¡eek band 7+8 (Fig, 3.2 lane 10), but did not, may have been caused by
the extraction of a larger píece of seed,

Fig. 3.2. SDS-PÀGE separation of F, kernels of the Roblin/prelude crossselected on the basis of MÀb binding: slot l and 18, prelude; slot 2 and19, Roblj-n, slot 3-l-2, selection of seeds with a l-ow MÀb reading(monosomic and nullisomic) , slot 13-L?, selection of seeds with àintermedíate IIÀb reading (a11 monosomic).
Fiq-.3,6, SDS-PAGE separation of extracts from testcross Fl kernels fromRoblin N1B/TD - 6,/ /prelude cross¡ slot 1, Neepawa and prelud-e; slots 2-8,selections with high MÀb reaction (disornic) ; slocs 9-16, selections wichintermediat.e MÄb reaction (monosomic) r slot l-?, TD-6; slot 18, prelude;slot 19, Neepawa,
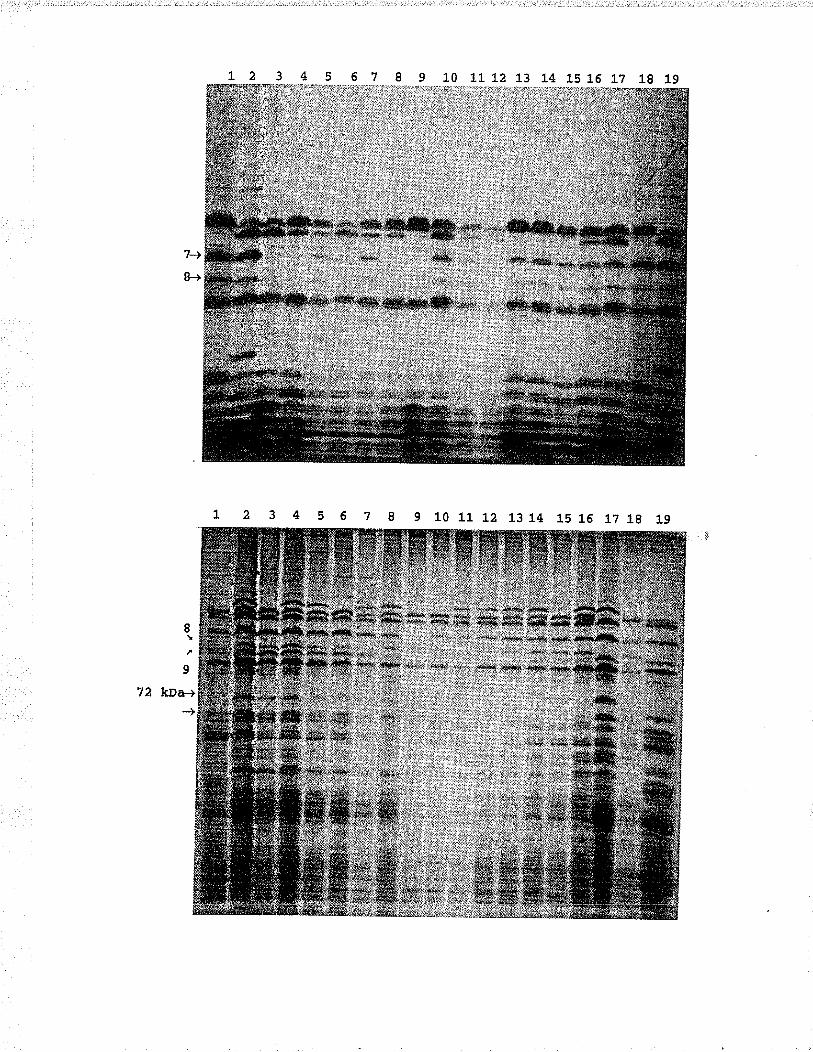
L0 tt t2 13 14 15 16 17 18 19
2 3 4 5 6 ? I 910 11 L2 L3L4 15 16 17 18 19

46
The I'lÀb and PAGE analyses were followed up wit'h cytologicalTable 3.2. Cornparison of cytologicaL analysis of roo! tips toP248 MAb reaction of populations Robl-in N1B,/pretude and RoblinN1B,/TD6 / /PreIude
Populat.ion Somatic cell MAb P24B reactionnumber (2n= ) Low fntermediate Hj-qh
2v4500
404t42
Roblin N1B/Prelude
Roblin N1B/TD6 / /prelude 4041-
YNumber of plants in each c1ass.zfndicates no samples were taken.
examination of root tips. The results are shown in Table 3,2. The fiveseeds which had an intermediate ìrÀb bindinq reaction and PAGE bands 7+g
had 41 chromosomes. Four of the 10 l-oh' samples could not be obtainedbecause of lack of germinabion, of the xernaining six, Lwo were 2n=40.
These two seeds corresponded to both low ¡,tAb binding xeaction and the
absence of PÂGE bands 7+8. The other four seeds were 2n=41, Three ofthe seeds corresponded to fow MAb binding and pAcE bands 7+9, while one
aeed corresponded to tow MÀb binding and thê absence of bands ?+8, The
absence of 7+t ç¿n be explained by lhe possible loss of the gene codingfor these bands. However, other èxplanations are possible, such as tbeoccurrence of a univalent shif¿, or technical error such as poor
extraction, sample t.ransfer, Coo sma11 a seed piece, or other error.
Roblin N1B/Canthatch//pretude/oianant
The testcross Roblin N1B/Cantha bch/ /prelude/Diamant. was studied. because
the endosperm of the progeny of such a cross is genetically more uniformthan for an F2 population, To demonstrate the chromosome dosage
discriminaeion of M.Ab p24B the extracts of Fl, half-kernels of the crossRoblin NlB/Canthatch were compared wi.th extracts of Roblin, 796-1 (a 1ow
bindinq control) and Canthat'ch in an ELISÀ. Binding of the MÀb p24B to
-00-90-06

F1 seeds was les s
than 33% of the value
of Canthatch or
Roblin (Fiq. 3.3) .
The Robl inN1B/Canthatch F1
plants were
testcrossed wi th
Prelude,/Diamant as a
poflen parent, and
relative binding ofMÃb P24B to each
testcross F1 kernel
is shown in Fig. 3,4,
Seeds could be
separat.ed into two groups based on antibody bindinq, those with an
íntermediate bì.ndinq (11-60% of Neepawa) and those with hiqh bindinq(80-L15% of Neepawa) . The ernbryo ends of seeds used in the above
analysis \,rere planted, and one spike on each plant was bagged, To
confirn the results of the ELISA done on F1 seed, 10 testcross F2
kernels from the bagged heads were analyzed by ELISA, and SDS-PAGE and
root-tip counls wexe done on the seeds with the lowest binding extxacts(Table 3.3).
without exception, all families with uniformly hiqh l.,fÂb bindingreactions derived from parents having a high À,1¡,b bindinq reaction.Likewise, afl famílies segregating for MAb binding reaction derived fromparents wj-th an intermediate r,fÃb binding reaction. The prelude/Diamant
and Roblin N1B lines lacked a protein band when compared with Canthatch,SDS-PÃ,cE indicated, again \!'ilhout exception, that families which did notcontain the band derived from parenes with Íntermediate M.Ab bindinqreactions. The intermediate MÄb reaction is expected to coïrespond to
1
EÇ
n 0.8
o" o.o
!.)oc-3 o.¿
o
{ o,
796-1 Fl Roblin Conlholch
Fig. 3,3, Relative binding (to Canthatch) of M,\bP24B Lo 50% propan-2-o1 extracts of half kernelsof .Roblin, 796-f, Canthatch and F1 RoblinN1B/Canthatch.

J! 12
9 ro
t!1 I
¡--
Efzo
the henizygous
condition, with the
single LB chromosome
being derived from
Prelude,/DiamanL ,
Families which
contained the band
derived from parents
with high M.Ab bindinq
reaction, which were
expected to be
disomic, with the
Canbhabch chromosome
producing the band.
A sampfe of tes tcros s
Absorbonce (405 nm)
Fiq. 3.4. Relative binding (to Neepawa) of MÀbP24B Lo 50% propan-2-o1 exi:ract.s of tesLcross Flkernels from the cross RoblinN1B/Canthatch,/ /Prelude/Diamant,
F2 seeds v¡as analyzed for somatic chromosome nurìber. AII low-binding
reaction seeds contained three sat.elli.te chromosomes, and 88% of these
seeds conlained 41 chromosomes. All high-reacling seeds contained foursatellite chromosomes, and all contained 42 chromosomes.
RobLin N1B/TD6//Prelude
Alle1ic differences in seed storage proteins occurred between parents
TD6 and Prelude (Table 3.1) , The gene locus for the ?+8 bands is in a
different arm than ehab of the gene coding a product recognized by MÀb
P24B (Dawood et a1 . 1989, payne 1987), SDS-pÀcE of this population was
considered to be a very good confirmation of thê Ì,jÀb, s performance inpredicting the segregation and dosage of the 18 chromosome.
The endosperm portion of F1 seeds of lhe Roblin N!B/ID6 / /prelude cross
were assayed by an ELISA (riq. 3,5). As with Roblin Nl-B
/Canthatch/ / Prelude
/Diamant, a bimodal distribution was produced, The sanples with the

Table 3.3. cenet.ic assortment of M,Ab p24B reacCion, occurxence ofPAGE band and root tip count and chromosome morphology of RoblinNlB/canthatch/ /ÞTelvde/Diamant testcross F2 progeny dérÍved fromF1 seed classed as internediace or high MAb binding
Tesrr..ir ôss ,'^ Testcross F1 MAb reaction
Testcross F2Charact.eristic Intermediate High
Genetic assortment toMAb
PÀGE band occurrence
Root - tip count
Nunber of satel l i techromosomes
segregating (16)z Nonsegregating (11)
Absent (13 )
40 or 41 (9)
3 (s)
Present (11)
42 (',t )
4 17)
zNunlcer of seeds or plants lested are in parentheses.
highest and fowest
absorbance readings
were run using SDS -
PÀGE (Fig. 3 ,6, page
45) . Â close
relalionship occurred
between the magnitude
of EITIS.A values and
pxesence or absence
of HMW subunit 9
characteristic of
TD6, cel slots treo
lo eight contained
samples wi th high ì,fÂb
!c
Ø6
t-oL2¡Efzo
Absorbonce (40S nm)
Fiq, 3.5, Relative bindinq (to prelude) of MÀbP24B to 50!È propan-2.o1 ext.racts of testcross F1kernels from lhe cross Roblin N1B/TD6 / /prelude , -
binding reaction and all contained band g (from prelude) and. band 9
(from TD6) . In slot eight, band 9 was presenc but weak. Two otherbands (arrorqs Fig, 3.6, page 45) absent in prelude appeared only in thehigh ¡,ÍÄb samples. The one band rneasured aL 72 kÐa (Ng and Bushuk L9g7)

50
has previously been shown to be associated with the 18 short chromosome
arm (Howes et al . 1990) . samples in slots 9 to 16 had an interrnediateMÀb binding reaction and contained only band 8,
Chromosome counts were carried out on root-tip cefls from Lestcross Fl
seed of Roblin N1B/TD6 / / pretude. À strong relationship between
chromosome counts and MAb bindinq react.ion was observed. (Table 3.2).Those plants with hiqh Mab binding reactions possessed 42 chrornosomes
with four satellite chromosomes, while those wieh low binding reactionspossessed only 41 chromosomes with three satellite chromosomes.
DISCUSSION
The MAb test for \a¡heat chromosome 1B was applied to two tl4)es of geneticanafysis: (1) a single-cross monosomic-18 F1 derived F, populat.ion frÕm
which disomic' monosomic and nullisomic kernels courd be ident.ified, and(2) a testcrosses from which monosomic or disomic kernels for chromosome
LB could be identified, Throughout this study, high molecular weightglutenins coded by chronosome 1BL (clu 81) and an unidentified proteinseparated by SDS-PAGE were used as an independent check on the MÀb test.Monosomic and disomic prants derived from the selfed monosomic could notbe discinguished from each other with SDS-PAGE because of the same
banding pattern. The results demonstrate that gel electrophoresis can
be used t.o further enrich the population after MÀb analysis, so thatcytogenetic analysis need only be applied to a very few seeds.
Cytological analysis is still required to confirm the identity ofprescreened 1ine6, because regulatory mutancs or smal1 deletions can
also result in a low binding reaction to the MAb, v¡ithout the loss ofthe chromosome,
The M.Ab screen enables very large numbers of seeds tÕ be non-
destructively examined (Howes et a1 . 1999) so that even spontaneous 1B
monosomic planCs could be detected. In wheat, on avêrage, a selfedmonosomic plant segregaees to produce about 73t monosomic, 24% euploid,

51
and 3% nullisonic offspring (McIntosh 198?). Â trimodal distributionwas observed using the MAb p24B as a chronosome marker (Fiq. 3.1) . The
peaks were not well separaled, but because the seed was from a
het.erogeneous and heterozygous population of F, seed, a range ofexpression of genes could be expected. Furthermore, the endosperm
tissue is triploid, and t4Àb binding values of monosomic lines will be oftwo tlt)es (3n=62 with 2 doses of 18 and 3n=61 with 1 dose of LB,
although 3n=61 t]æes will predominate) , depending on whether chromosome
1B is passed through tbe male or the female gamete. Afthough the MAb
may not. reliably reflect the dosage of a particular chromosome, it can
be used to enrich the nuÍìlcer of aneuploids h,ithin a populat.ion of seed.
Monosomic kernels were generated from testcxosses ueing monosomic 1B
plants as the female parenE, and euploid 1ines as the pollen parent.îhe Roblin NIB/TD6 / /pre:-ude testcross was chosen with cultivarsdiffering in c1u 81 a1le1es, subunit 7+g or I+9, so that monosomic and
disomic kernè1s could afso be identified by the presence of one
(monosomic) or two (disomic) c1u 81 alteles, The RoblinNLB,/Canthatch//Prelude/Ðiamant test.cross population fortuitouslysegregated for the presence or absence of a different band. The strongrelationship between MÀb reaction and banding pattern for bobh
populations confirmed the cytogenetic interpretation made based. on theMÀb binding reaction.
The triploid nature of the endosperm tissue was exploiLed through thetestcross method. The monosomic Fl female either contribubes 2 doses ofthe 18 chromosome or does not contribute any 18 chrÕmosome to thetriploid endosperm t.issue. The pollen parent cÕntributes only one-thirdof the chromosome number in endosperm tissue, which is reflected in theamount of protein, that is, endosperm lissue of all testcross kernels iseither 3n=61 with chromosome 18 present in a single dose or 3n=63 withl-B present in 3 doses. Às a result of the differential chromosome
dosage effect, che quant.itative nature of che M.Ab test could be applied,

52
Every kernel could be identified as being monosomic 18 (and thus derivedfrom the fertilizalion with the euploid pollen parent.) or dísomic 1a,
which was reflected in the bimodal distriburions (Fiqs. 3.4 and 3,5) ofthe test.cxoss Fl seed of the two differenL populations tested. As
expected, Che t.est.cross produced a preponderance of intermediate - MÀb -
reacting kernels compared Eo the nurnlcer of high-MAb-reacting kernels.Testing a number of seeds of a lest.cross F, family by MÄb analysis or bygel electrophoresis can be used as a check on the MÃb result of the one
progenitor lestcross F1 seed used to classify that seed as monosomic ordisomic, The testcross method can be expected t.o produce more
consistent' and clear results because of consistency in chromosome number
in the endosperm tissue, but also because of homogeneity among al_l
testcross individuals,In these seudies, disease-resistant genotypes were crossed to a plant
nullisomic for chromosorne 18. The nethod can be exlended by crossingthe euploid, for example a disease-resistant genotlæe (ma1e pareng) , toa suscept.ible monosomic plant (female parent) , and selecting by tlÀb
analysis only monosomic kernels which must contain the 1B chromosome
from the resi-stant genotlæe. The monosomic plant can then be
testcrossed, and MÀb analysis used to classify monosomics and disomics.Because many monosomic series are available, this would often be the
method of choice, Às few as 10 monoso¡nic plants are needed to be t.ested
to determine if a disease resi.stance gene is located in chromosome LB
(P>0,00L) , .A disadvantage is thal recessive genes cannot be as readilystudied. The method applied to hybrid progeny of a selfed double
monosomic wheal alien substitution line has the advantages of both ofthe above meehods (Howes et a1 . 1993),
The results demonstrate that. ì,{Abs can bê used as chromosome markers inaneuploid analysis, particularly in a testcross, thereby improving theefficiency of cytogenetic analysis. Àt present, this method cannot be
applied to al-I chromosomes since only MÀbs specific to 18, 6A and. 6D

53
chromosomes have been identified (Howes et at. 1989 and 1993; skerritt1991; skerritt et a1 . 1991) . Endosperm proteins are coded by group 1
and 6 chromosomes (Wrigley and Shepherd 19?3; payne et al . 1991), and an
alpha amylase inhibit.or has been associated with a group 2 chronosome
(Masojc and Gale 1990). Àlbumins and globulins have been assocíatedwiEh groups 3, 4, 5,6 and ? chromosomes (Fra-Mon et a1 . 1994; cupta etal . 1991) , and alpha arnylase j-nhibitor genes in the group 3 and group 6
chromosones (Sanchez-Monge et al . 1986) . Thus, iC should be possible toisolate specific MÀbs for alL whea¿ chromosomes.

54
CTI.A.PTER. 4
A Monoclonal Antibody Chrornosome Marker
Analysis Used to Locate a Loose Smut R.esistance
Gene in Wheat Chromosorne 6A
ABSTRACT: Many genes have been located in specific wheal
chromosomes, yet little is known about the location of genes forresistance Eo tlstiTago triLici, which causes loose smut, Crosses \,¿ere
made between the loose smut susceptible alien substitution lines Cadeg
6¡\S(6À) and Rescue 6ÀS(6A.) (lines in which wheat. chromosome 6A issubstitueed by Agïopyron chromosome 6) , and four cultivars resistant torace TL9: Cadeb, Kota, Thatcher and TD18. The segregating progeny were
tes¿ed for reaction to loose smut race T19, and for the leve] of bindingwith a monocLonal antibody specific to a chromosome - 6À - coded seed
protein. The antibody, which binds only in the presence of the 6À
chromosome, \rras used as a chromosome marker. Àn associat.ion was
established between resistance Lo U. tritici race TL9 and the presence
of chromosome 6À for each of the cult.ivars tested. The resultsindicated that resistance to race T19 resides in chromosome 6A for allfour cultivars. Race T19 resistance in Cadet appea¡s t.o be located inthe shor¿ arm of chromosome 6Ã,, based on the evaluation of the cadet 6À
long ditetosomic stock, which was susceptible, and the Cadet 6À-shortr 6
Agropyron-short alien translocat.ion stock, which \r'as resistant..Chromosome markers can be used in conjunction with cytogenetic stocks tolocace genes to chromosornes.

55
INTRODUCTION
Control measures for loose smut are difficult Co develop and maintainbecause they must take into account lhe intimate relationship between
this obligate fungal pathogen and its host, but control is jusci.fiedbecause of 1ow, but consislent yearly yietd losses (Schafer 19g?, Wiese
3,98'1). cenet.ic resistance, which obviates the environmental hazardsposed by seed treatment, has beên used effect.ively to conCrol loose smut(Schafer 1987). Knowledge of ¿he chromosome location of loose smut
resistance genes can be used to improve the efficiency of breeding forresistance to this disease.
Breeding and genetic analysis of loose smut resistance are difficultbecause florets of the wheat spike must be individuatly inoculated, and.
progeny evaluation requires extensive growth cabinet and gireenhouse
space (Lfonea and Dhitaphichit 1991-). Not onty is the evaluation ofloose smut reaction done on a family basis (caskin and schafer 1962) ,
nearly two generations of the host must be grown i one for inoculationand the second for sl¡mptom expression. with advances in the development
of molecular markers, the breeding for resistance Co loose smut can
potentially be more efficíent through the use of markers ctosely lj.nkedto loose smut resistance genes.
Às naps of molecular markers in the Triticeae become commonplace (Chao
et aI . 1989i Sharp et at. 1989; cil1 et a1 . i.991; Heun et at. 1991; Liuand Tsunev¡akÍ l-991, .A,nderson et al . 1992; Devos et a1 . 1992 and 1993;
Devos and cale 1993) , the use of such markers will be limiEed by thenunber of genes mapped to chromosones L'ithin the particular populationsused to develop the rnaps, cenes for traits not. polynorphic withinexisting mapping populations will not be so ea6i1y mapped, thusrequiring the study of parents and populat.ions for which molecularmarkers and maps have not been developed, but which possess the desiredtraits of interest such as loose smut resistance.

56
Ànalysis of two chromosome substiEution series was done by
Dhitaphichit el a1 , (1989) to locate two foose smut resistance genes towheat chromosomes 7À and ?8. Results of analysis of varietalsubstitution 1ines, however, should not be considered concl-usive
evidence of the locacion of a gene (Mclntosh 198?; Koebner et. al . 1988t
Yen and Baenziger 1992) because such backcross lines are prone toincomplete genotype rest.oration. Furthernore, heterogeneity or residualheterozygosity in either or both the recurrent and donor parents used .in
the developrnent of substicueion lines can result in misteading
conclusions. In addition, chromosomal localization is resLricted tothose genes wiChin the available cyÈogenetic stocks. Strategiesinvolving segregation analysis, using confirmed cytogenetic stocks, notonly provide stronger evidence of the chromosomal loca¿ion of a gene,
but also a11ow analysis of traits not carried within existingcytogenetic stocks.
Monosomic analysis ínvolving segregating populations is commonly uEed
for locating genes in chrÕmosomes (sears 1969). Thè problem with thismethod is that large F2 populatj-ons are required to distinguishsegregation ratios. As an example, a line possessing a dominant loose
srnut xesist.ance gene can be crossed onto a monosomic series. Fivepercent nullisomic progeny will be assumed in the case of the criticaLcross. A minimum population size of 5? is required to distinguish the
ratio involving ttte crilical chromosone of 19:1 disomic and monosomi-c
(resistant) to nullisomic (susceptible) , from the ratio of 3:1 resistantto susceptibte for the noncritical chromosorne populations (99 percentprobabilicy) (Hanson l-959), Because of the wide variation for foose
smut reaction within a genotlt)e, analysis is complicated by overlap inlhe reaction of ¡esistant and susceptible families such thatdelimitation of the cl-asses is difficutt (Ribeiro 1963) , and arbitrarydelimitation should be avoided (Knotr 1907a) .
Chapter 3 provides a discussion on a method Õf chromosome tracking

57
using seed proteins as chromosome specific molecular markers, so thattraits can be locaCed to chromosomes using relatively small populati_ons(i,e, fewer than the 57 families required for monosomic analysis) . The
crosses involved the use of nullisomic cytogenetic stocks to avoidcrossing over for a part.ìcular chromosome. The advaneage of rapidanalysis using monoclonal antibodies (MÄb) to seed proteins is utilized.(HoweE et aI . 1989) when MAbs are used as chromosome rnarkers . MÂ.bs arealso used as chromosome narkers in populations invotving aliensubstitution lines (Howes et al . 1993). The present study made use ofalien substitution lines to avoid crossing over while using a M.Ab as a
chromosome marker.
fn principfe, to use alien substitution Iines in conjunction withchromosome markers, crosses would be made as in monosomic analysis, butinstead of crossing onto a monosomic series, crosses would be made
between a source of loose smu! resistance and a series of susceptiblealien chromosome subst.itution 1ines, èach representing a dÍfferentchromosome. For each cross, progeny would be classified using (from a
series of chromosome markers such as antibodies specific to particularchromosomes) the marker which is pollaTìorphic between the resistantparent and the corresponding alien chromosome substitution Iine in thatcross. Complete association of the resistance trait with a wheat
chromosome, and susceptibility with the alien chromosome in one of thecrosses, would indicate the chromosome location of the gene, ,All oehercrosses would show random segregation for resisEance and susceptibilitywithin each chromosome c1ass.
Rather than use a fu1l series of 21 afien chromosome substitutionlines crossed onto a sj-ngle source of resistance, the approach used herewas to evaluate those chromosomes for which good seed protein markersare avaj-lable, and to cross cyloqienetic stocks that represent thechromosomes with markers onto different sources of 1oÕse smug
resistance, The group 1 and group 6 wheat chromosomes coding for lhe

58
seed storage proteins offered the great.est potential of providing
poll¡rnorphic chromosome markers (Payne 1987). The sources of resistance
chosen for evaluation included: lines from Ehe U. tritici race
different.ial set (Niel-sen 1987a) , other sources of resistance of
historical significance in spring wheat brèeding, and the progeniEors of
cytogeneCic stocks found to be resistant during Lhe course of thisstudy.
This study was undertaken primarily to provide evidence of the
chromosome locat.j-on of a loose smut resistance gene or genes, but also
to validate the use of MÀbs as molecular chromosome markers inconjunction with alien substituÈion lines, Àftex a firsC round of
crossing of cyeogenet.ic sCocks with sources of resistance had been
initiated, the concurrent evaluation of cytogenetic stocks for loose
smut reaction indicated that the cultivar CadeC was resistant to loose
smut race T19, while |L¡Le Agropyron alien substiLution line Cadet 6À9(6,A.)
(Whelan 1988) was susceptible to this race, This informaEion indicated
the location of a resisEance gene in chromosome 6A.. Attention was
focused on lhe study of the segregation analysis of this 6A/T19
resistance, and chromosome l-ocation of this resistance on chromosome 6A
in four culLivars is the main subject of this report.Inilial testing of populations from ehe first round of crosses
indicated a Tl-9 resistance gene to be in the 6A chromosome for Kota, a
progenitor of the differeniial TD4 (Hanna 1937, Nielsen 1987a) . A
second round of crossing, including Cadet and Thatcher crossed onto
Cadet 6A9(64), was initiat.ed after the inoculation of Cadet and cadel
6AS(64) indicated resistance io be in chromosome 6A,. Thatcher is a
parent of Cadet (Zeven and Zeven-Hissink 1-976) and progenitor to the
differential TD12.A, (Nielsen 1987a) . The CadeE 6As(5À)/Cadet cross was
made because it could allo\.¡ aasessment of the chromosone marker
technique for locatíng loose smut resistance genes to chromosomes, based
on the evidence for the location of the T19/6À resistance gene already

59
available from the inoculation of Cadet and Cadet 6Aq(6À) . Evaluationof the Cadet 6AS(6À) /Cadet and Cadet 6Àq(6A) ,/Thatcher poputations
indicated a T19 resist.ance gêne in chromosone 6A for both populations.Evaluation of a fourth populatíon having TD1B, a loose smut
differenlial, and Rescue 6Àq(6A) (whetan 19gB) as parents indicat.ed yet.
another line wi¿h a TL9 resistance qene in chromosome 6Ä.
Usinq the chromosome 6A seed-protein specific Ml;r. 230/9 (Skerritb eta], 1991; Ho\ees et al , 1993) as a chromosome marker and the aliensubstitution 1ínes Cadet 6ÀS(6A) and Rescue 6Àq(6A) to avoid crossingover of the 6À chromosome¡ T19 resistance in Cadet, Kota, Thatcher and
TD18 was found to be associated with wheat chromosome 6À. The gene
location was accomplished using a minimum of resources, and confirms thevalue of molecul-ar markers as chromosorne markers (in particular, MAbs toseed proteins), in co¡nbinatj-on with alien substitution lines, as a
mebhod of locating genes to chromosomes,
Knowledge of the chromosomal location of loose smut resistance genes
within the differentiar see witl make easiex che eventual development ofsingle gene differential lines used to assess races of U, tritici.
MATERTÀIJS .AIVD METHODS
cadet 6A9(64) and Rescue 6Às(6A) (alien chromosome substitution lines inwhich the 6A chromosome is substituted with the homoeologous Agropyron
eTongatum chromosome 6A9) were two of a group of subsbituti_on lines used
as females in crosses with several sources of loose smut. resistance,Dr, E.Ð.P. Whelan of thê Àqriculture Canada Research station, Lethbridgeis gratefufly acknowledged for the provision of the group 6 cytogeneticslocks. the loose smut reaction of the cytogenetic stocks was unknown
during a first round of crossing. Because at least half a year isrequired to evaluate a genotlæe for resistance, resistance evaluation ofstocks was done concurrently with crossing. Kota and TDlg were among
the sources of resistance in this round of crossing, fn a second round

60
of crossing, the resistanL culCivars Cadet and Thatcher were included.
TD18 was crossed onto Rescue 6ÀS(6Ä.) , whereas the other three resistanL
sources were crossed onto Cadet 6ÀS(64) . Fl plants were grown from each
cross and the heads were bagged.
Eight races of loose smut were initially chosen as inoculum, wieh an
attempt to select races r,¡hictr maximize virulence and at the same tíme
allow for di f ferênt.iation of resistance within the loose smut
differentials described by Nj-eIsen (1987a). Only races T8, T15, T19,
and T31 were pertinent and therefore used on the parental conbinations
in thie s budy,
The lrÀb 230/9 was provided by Dr. J,H. Skerritt at CSIRO in Sydney
Australia, to rr'horn we are grateful . F2 seed was analyzed for thepresence or absence of the wheat 6.4 chromosome, based on the bindinq ofMl¡ 230/9. Antibody bindinq was based on an ELISÀ (Enzl¡me Linked
Immunosorbant Àssay) described by Howes and coworkers (1989), A portionof the brush end of each seed was removed and placed in a mu1tiwellp1ate, and extracted in L50 uL of 50% propan-2-o1 overnight at 3?ôC.
Seed pieces were squashed with a blunt-ended rÕd and allo\^red to incubateanother 30 min. Three repLicaÈes, of a 1 in 10 dílution of each extractÍn 70% ethanol, were bound to a plate aC room temperature for 30 min and
blocked with 1% milk for 1.5 min, The tdÀb 230/9 \tas diluted 1 in 10,000
and applied to plates for 90 min, followed by ehe addition of goat-
antimouse antibody conjugated with alkaline phosphatase diluted 1 in2000 applied for 2 h. Ethanolamine buffer fi..0 M, pH 9.5) containing 1
mg/ml p-nilrophenyl phosphate as substrate for the phosphatase \{ras
added, and the colour reaction was allowed to develop. Absorbance was
measured using a Tit.re Tek muttichannel spec trophotome ter at wavelength
405 nm, Incubations of milk and antj.body steps were done ac 37oc.
Washes between each step were done using 20 nM Tris pH 7.4 - 0,15 M NaCl
- 0.05% Tv¡een 80 (TBS-Tween), Àntibodies wexe prepared in O,05% milkdiluted i-n TBS - Tween.

61,
Up to 80 F2 seeds for each population were analyzed with the MAb
230/9, aTong with Thatcher and Cadet as high antibody binding controls.Ten of the highest binding seeds, expected lo be disomic for the wheaL
6À chromosome, and 10 of the lowest binding seeds, expected to be absent
for the 6À bue disomic for the Agropyron 6 chromosome were selected(Howes et a1 . 1993), À.bsorbancè readlngs were converted to relaLivereadings, using the high controls as a basis, The population size of l-O
hiqh-bindinq and L0 low-binding seed was determined based on
distinguishing between the two most likely causes of the expected
outcome of all 1ow-binding seed (disomic Agropyron 6À9 and no antigenfor antibody) selections being susceptible and all high-binding seed
(disomic wheat 6À and possessing ant.igen to antibody) selections beingresistant. These two causes are chance segregation or a realassociat.ion between foose smut resistance and the hrheat chromosome,
Considering the segregaeion possibiliby, chance segregat.ion would have
to work in favout of the one phenotlæe (e.g, resistant) for the one
chromosome class (e.g. wbeat) , and at the sane time hrorking in favour ofthe other phenotjt)è (susceptible) for the other chromosome class(Agropyron) to provide the same outcorne as a real association between
the chromosome and resistance. Because races with complex virul_ence
were chosen, few multiple-gene segregations for resistance to the same
race were assumed. fn an F2 population, 3:1 (or 1:3) is the expected
segregatj-on ratio for a síngle gene, If resistance was dominant and
there was no associalion between resistance and the chromosome in a
particular cross, then by chance the suscept.ible genotlæe was expêcted
to occur 25 percent of the time within lines classified as disomic forthe alien chromosome. To minimize the chance to one percent of decidingin favour of a real association of resistance with a chromosome, when infact chance segregation occurrèd, a population of 4 indj-viduals must. be
stìrdied (p=.25{=0,004) within the alien chromosome class, If theresistance was recegsive, the wheat chromosome class is the most

62
important to consider. When the dominance relationship is unknown, as
was the case in this sludy, both classes must be consj.dered for a totalpopulation of eight. The population of 20 chosen in this study was
considered ample.
The embryo end of the seeds having the desired antibody reactions were
pregerminated and planted in seerile soil to minimize microbialinfestation of ¿he seed and prevent foreígn seed contamination. The
Cadet 6Aq(64)/Kota population was inocutated with race T19 as a
priority, and, i.f fert.ile heads remained inoculation was performed withrace T31 to determinê the association between T31 and TL9 resist.ance.
Likewise, the Rescue 6Àq(6Â) /TD18 population was inoculat.ed \.¡i th T19
first, and T15, T31 and T8 were inoculated onto laeer spikes. The Cadel
6Àq(6'A),/cadet and Cadet 6Aq(6À.) /Thatcher populations were inoculatedwith only race TL9.
Plants to be inoculated were grown in a growth cabinet at 19"c, with16 h light and I h dark. plants were inoculated using a l-O mL
hlt)odermic syringe with a 22 gauge needle to inject a water suspension
of spores (approximately 1 mg mt 1) ineo florets at mid-anthesis.Following inoculation, the head was bagged Lo minimize cross pollinationand to improve infection conditions (Nj-elsen 199?a) .
À minimum sample size was calculaled using Loose smut values of theparental lines (Table 4,1). The famity size needed co distinguishbetween p1=.74 (the probabili.ty of occurrence of factor one, here equal
to the proportion of smutted plants in Cadet 6A9(6À) ) and p2=,26 (theprobability of occurTence of factor thro, here equal to the proportion ofsmuLted plants in Kota) at a 99 percent probabiliey is 16 (Hanson 1959)
ior the populat.ion Cadet 6À9(6À) and Kota, which required the greabest
family size of any of the paxental combinations studied. where
possible, at least two heads (often 3 to 4 heads) on each F? plant. were
inocul-ated. Because of the statistical unreliability of data on
families of fewer than j.6, due to the inftuence of sma1l sample effect.s,

63
such data was not used for the analysis and interpretation of the datapresented here. However, it should be noted that the data of familiesof fewer than 16 would not have distorted the inLerpretation of thedata,
Inoculated heads were harvested and threshed separately a¿ maturity.Seed from one head from each family for each race was planted in thegireenhouse, followed by planting of a second head when the space became
available, and so on until all heads had been planted and evaluated,The nurnlcers of l-oose smut infeceed plants and healthy plants were
counted. Proportions of smutted plants were calculated, with the range
in sample (family) sizes occurring for each population (mean sample sizein parentheses) as follows: Cadet 6Àg(6Â) /Xoea - T19: 1g Lo I44 (75), -
T31: 16 to 49 (36); Cader 6Àq(6A) /Tharcher - T19: 34 to 106 (66) ; Cadet
6As(6A)/Cader - Tl_9: 48 ro 142 (102); Rescue 6Às(6À) /,rDL} _ T8: 18 ro ?1
(45) , - T15: 19 to 10? (61), - T19: j"g Lo !!2 (60) , - T31: 2! Lo II2(41).
To confirm the M.Ab 230/9 binding of the single F2 seeds, 10 F3 seeds
of populations Cadet 6Aq(6À) ,/Kota, Cadet 6ÀS (6À),/That.cher, and Cadet
6A9(6À)/Cadet. were tested to the antibody in an ELIS.A.
The alien translocation lj-ne Cadet 6Àgsr6A5 (whelan and Lukow L990),in which the long arm of chromosome 6 of Cadet is replaced by the shortarm of chromosome 6 from Agîopyron, and the ditelosomic Cadet 6ÂL t'r ,
missing the short arms of the homologous 6A chromosomes, were
inoculated. These inoculations were done with race T19 after reactionto race T19 of Èhe Cadet 6Aq(6.A) and Cadet had been determined, The
ditelosonic and translocation lines were inoculated with race T19 t.oprovide inforrnatj-on on the arm locatj-on of the resistance gene.
statistical analysis of much of the data throughout ehis study was
superfluous because of the nagnitude of differences between lreatments,nonetheless, statistical analyses were done and will be reported, À t-tese was performed on the absorbance readings of the parents (Table

64
4,1), Thatcher, Cadet, Kota and Cadet 6AS(6À) , and TD18 and Rescue
6As(64) loose smut reactions were compared using Fisher,s exact 2.tail-test. Pairwise, 2 X 2 lesls of independence using Fisher,s exacl 2-tailtest were done using Ehe loose smut reaction data from Cadet, Cadet
6AS(64), Cadet 6AqS:6À5, and Cadet 6Altrr (a C-test could not be used
because values of the resistant cultivars were not normallydistributed) . Loose emut dat.a from the segregating popul.ations were
arcsine transformed to ob¿ain normality before applying a t-t.est.All tests were run using the computer procedures of the StatisticalAnalysis System fnsCitute (1989).
RESUIJTS
This section presents data from a selection of lines and populations
Eaken from a laxger group of lines and populations used initially in the
study. The results represenC those lines and populations analyzed afCer
the focus was Curned to the 6À chromosome, based on initial results from
the inoculat.ion of the Cadet 6Â9(6À) parent and its progenitor Cadet.
only results of populaLions that indicated T19 resistance to be inchromosome 6A are presênted; however, results of the reaction of thesepopulations to other races are also presented to provide a cont.rast
between results that demonslrated resistance to be associated with the
\a'hea! 6A chromosome, and results that demonstrated resist.ance not to be
associated with the wheat 6À chronosomê.
The l4Äb 23019 bindinq !o seed extracts of Ehe two atien substitutionlines Rescue 6AS(6À) and Cadet 6Àq(6À) was signifj-cantly (p>0,005 tvhen
the nul1 hlæothesis assumes no difference in mean absorbance readings)
lower than Chat of Ehe euploid cultivars, providing the pol-I,morphism
necessary to use this antibody as a chrornosome rnarker (Tab1e 4.1_) .
Absorbance readings from the alièn subst.ilution lines and the Cadet 6ÂL
trr were significantly lo\rer than the euploid lines or Cadet 6A95!6Às (p>
0.00s) .

Table 4.1- Proportion of smutted to toLal plants (p) and total plants (N) of wheat cultivarsand lines inoculated with loose smuL races T19, T31, T15 and T8-and monoclonal antibody 230,/9binding (oD) relaLive to cadet and the standard deviation (sd) r¡si¡n a sampte size of iixRace
CadetCadet 6Àq(64)Cadet 6ÀL t"Cadet 6À95:6A5Kota
ThatcherTD18Rescue 6å9r (6À)
zlnoculaLion not. done -
T19
0.11 3090.74 2040.55 2990.22 930.26 23
0 . 1"6 r070.00 380.39 s5
T3L
0.960.93
_z
0 - 00
0.730.00o -r7
28
s6
18
TL5
0.00 1180.13 to1
0.16 37
0.00 360.00 3'10.64 55
0.000.00
0-55
0.000.000.33
i.0 462-
49
593648
OD
1.00 0.2950.05 0.0280.09 0.034L -26 0.5r.70.86 0.323
1.05 0.361i..08 0.0540.10 0.014
sd

66
The loose smut reactions of the wheat lines used as parents in thisstudy are also present.ed in Table 4.1-. Of the four loose smu! races
shorvn, the most not.er,¡orthy results were fo¡ the cultj-vars Cadeg and
Cadet 6Aq(6À) because of their significant (p>0.001) difference inresponse to race Tl-9, These results indicated that resistance to race
T19 resj-des in the wheat 6A chromosome. To get an indicat.ion of the
chromosome arm location and a further affirmation of the chromosomal
location, two additional cyeogenetic sCocks were evaluated: the Cadet
6ÀL t''r and the Cadee 6A95!6A5 translocation. Resulgs of the inoculationwith T19 of the Cadet 6.A,95:6A5 and the Cadet 6À,L t,, are also shown inTable 4.1, There was no reason to inoculate these two lines with theother three races. The proportion of smutted plant.s in Cadet 6A95:6AS
was significantly lower than in Cadet 6À9(6.A) or Cadet 6ÀIr t , and
Likewise for Cadet compared wiCh Cadet 6ÀS(6A) or Cadet 6ÄL ttr,demonstrating that loose smut reaction r4as not independent of chromosome
condition (p>0.001 for each of the tests) .
Whereas the loose smut' reaction to each of the races was reasonably
clear-cut for most lines in Table 4.1, the reactions for Rescue 6Aq(6À)
were less c1ear, For example, the proporlion of infected plants to raceT3l- was 0.17 for Rescue 6A9(6À) whereas the proportion in Rescue was
0.82 1¡¡=22¡, Thaecher and Kota differed siqníficantly for react.ion toloose smut race T19 from Cadet 6A9(6Â) , and TD18 from Rescue 6Aq(6A)
(p>0.001) , The results of the inoculations of the parents dictatedwhich races could bê used on the segregating populations.
l'1Ab 230/9 binding to F2 seed extracts was expected to ïepresent thewhea! chrornosome dosage. The results of ELfSÀs for the four populations
cadet 6Àq(6Ã)/cade¡, cader 6Ä9(6A)/Kora, cadet 6Äs(6À) /Tharcher and
Rescue 6Àq(6À)/TD18 are shown in Fig. 4.1A to 4.1-D respectively (in thisfigure the values of the horizÕntal axès represenb refative absôrbance
readj-ngs; in this and subsequent histograms, tick marks of horizontal

ÌvlAb binding (obsorbonce 405 nm)
t5
I,1U)ro 13c.) .-ü''Ø11
NtoLr- I9_Eo7t-6o.E1fJz2
1
0
Fig, 4.1, llþb 230/9 binding measured by absorbance of ELISÀS on seed91t{qc!q from F2 poputarionsr (A) cader 6As(6À) /cadet¡ (B) cadet6Às (6.A.) ,/Kota;
MAb binding (obsorbonce 405 nm)
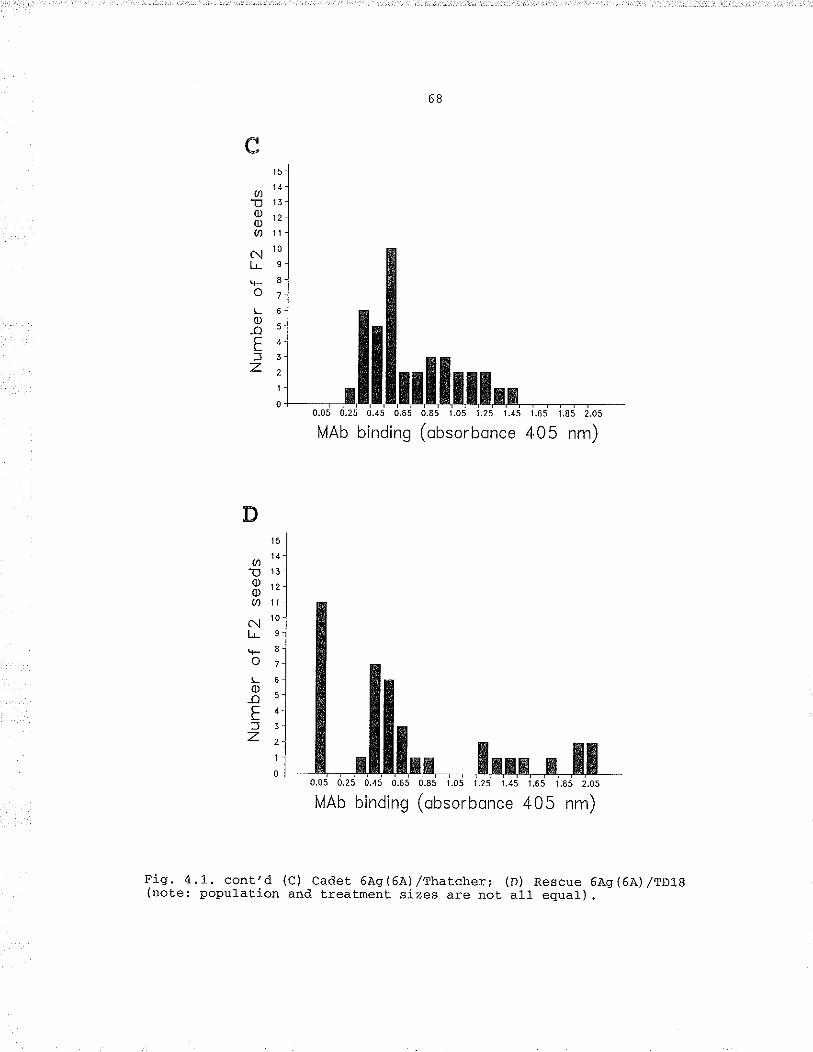
0.0s 0.25 0,.15 0.65 0.6s 1.05 1,25 L15 1.65 1.85 2.0s
MAb binding (obsorbonce 405 nm)
Fiq. 4,1. cont'd (c) cadet 6À9(6Â) /rhatcher; (Ð) Rescue 6As(6A)/TD1g(note: population and treatmenL sizes are not all equat).
15
,n 'o!1Jc) ."c) ''Ølt
c.i 10
Lr- Iri- 6
o7!-6c)
¡lE1f5z2
I
0
MAb binding (cbsorbonce 405 nm)

69
axes demarcate class intervals, and the values represent the midpoint ofthe int.erval). The seeds represented by the far left-hand tail of the
distribution were expected to lack wheat chromosomes, reflected in a 1ow
1eve1 of binding of the ant.ibody. Instead of possessing the wheat 6A
chromosome, these enbryos were expected to be mostly disomic for the 6-
Agropyron chromosome (endosperm Erisomic 6À9) , \{hereas enìlcryos of seed
represented by the far righl-hand tait were expected Lo be disomic forche 6A-v¿heac chromosome (endosperm trisomic 6À and hiqh binding of the
antibody) . Äs seen in Fiq, 4.18, analyEis of the Cadet 6À9(6A) /Kotapopulation pxoduced a trimodal distribution, as did the Rescue
6Às (6À) /TD18 (Fig, 4, LD) , whereas bhe cadet 6Às (6A) /cadet (Fig. 4.l-À)
pxoduced few low-reacting seed, and Cadet 6Aq(6A) ,/Thatcher produced ¡one(Fiq. 4.lc) . The seeds with extremely high or extrenely 1ow MÃb bindinqselected from the tails of each distribution were carried on for loose
smut analysis and further tLAb analysis. The intermediate - binding seeds
were expected to be double monosomics, and, with the exception of theThatcher population, were dropped fron further study. A l_0-seed sample
from each F, plant (F¡ seed) was used to confirm the reaction of thesingle F, seeds. The nunber of double monosomics accidentally selectedwas L in 20 for the Cadèt 6Àq(6Ä) /Kota poputation, and 5 in 20 for theCadet 6Aq(64)/Cadet population, where the 20 seeds consisted of both lowand high tlæes, However, none of the cadet 6AS (6A) /Thatcher poputat.ion
were disomic for 6À9. These results were consistênt with thedistribuÈions shown in Fig. 4,1, which showed Chat the Cadet
6A9 (64),/Cade! and the Cadet 6Â9(6À) /Thatcher produced few very 1ow-
binding seeds. F3 seed of TDI-8 \,¡as nol evaluat.ed.
The results of segregation of F2 progeny for loose smut in l_inespreviously classified for chromosome 6A or 6A9 are shown for Cad.et
6À9(64) /Cadeb. in Fiq. 4,2A, Cadet 6Aq(6A) /Thatcher in Fiq. 4,28, Cadet
6ÀS(64)/Kota in Fig, 4.2C and D, and Rescue 6À9(6Â)/TD1g in Fig. 4.2E to4.2H. Segregation response to different races are shown for cultivars

70
Kola and TD18 in Fiq. 4.2C Lo 4.2D and 4.2E Lo 4.2H respectively. The
reactions of the Cadet 6Aq(6,A) /KoLa F2 seqregating lines to U, triticiracè T19 (based on the evaluation of F3 families) are show¡ in Fiq. 4.2C
for the disornic 6A and the disomic 6À9 chromosome classes (based on MAb
230/9 binding) . Those progeny with the wheaC 6A chromosome had a
significanlly lower proportion of smut ehan those with the 6À9
chromosome, with no overlap bet\4¡een the two chromosome classes (the
probability of 't, being near infinitely smatl when the nu1l hl,Þothesis
assumes no effece of chromosome) . The Cadet 6ÀS(6À) /Kota poputatÍon
sample size used was 18 divided inlo two groups of 9 based on tl4)e
ofchromosome. The probability of not obCaining the alCernatj.vephenotlæe in one chromosome class, based on chance segregatj-on, was lessthan 0,0001. Similar resulls for reaction to race Tl_9 were obtained
with the Rescue 6À9(64) /TD18 (Fiq. 4.2E) and the Cader 6A9(6À) /Cadet(Fig. 4,2À) populations; howêver, the size of the alien chromosome classwas only five for the Cadet 6Ãq(6A) /Cadet poputat.ion. The low MAb
binding progeny (reprêsenting chxomosome 6À9) shown in Fig, 4.28 for theCadet 6Aq ( 6À) ,/lhatcher population represented double monosomic plantsinoculaeed wi¿h race T19. plott.ed values for race T19 infection of aI1four populations produced similar distributions lîíq. 4.2), The mean
infected propottion for F, families derived from F2 plants, selected as
having the subs¿ituted 6À9 chromosome fox each population, tvas ofsj-milar infect.ion level- to Ehe susceptible parent in the cross, whilethe mean of lines selected as having the wheat. 6A chromosome for each
population was similar to the mean of the resistant parenc in the cross.fn contrast. to the results from inoculation with T19, when the same
plants from the Þopulation Cadet 6Àq(6À) /Kota were inoculated wj_th raceT31, to which Kota was resistant but Cadet 6Âq(6A) was suscêptible, no
association (t-test probabiliey is 0.65) between chromosome 6Ã and
resistance could be ascèrtained (Fj.q, 4.2D) . A further illust.ration of

A10
9
8
7
6
5
4
5
2
1
00.05 0.25 0.45
Proporlion of0.6 5 0.85
smuifed plonts
10
I8
7
6
5
4
3
2'1
00.05 0.25 0.45 0.65 0,85
Proportion of smutled plonfs
Fiq. 4.2. Distribution of loose smut proport.ions of F3 familiesclassified by chromosone based on antiboãy readings: (e) C.a"t6À9(6À)/cadet to race T19; (B) cadet 6Às(6Ã) /That¿her t'o race T19,
U)0)
.E
o
oLo)
_o
Efz
B
v)a)
=Eo
oL()
-oEfz

c10
9
I7
6
5
4
3
2
1
0 0,05 0.25 0.45 0.65 0.85Proporlion of smutted plonts
10ô
I7
6
5
4
3
I
1
00.0 5 0 .25 0.4s 0.6 5 0.8 s
Proportion of smutTed plonts
Fí9. 4.2 cont'd (C) Cadet 6À9(6A)/Ko¿a to race T19; (D) Cadet6Às (64),/Kota to race T3l-t
l1)
.q
Eo
oLo
_o
Ef,z
Ito
=Er1
(+-
oLc)¡Efz
D
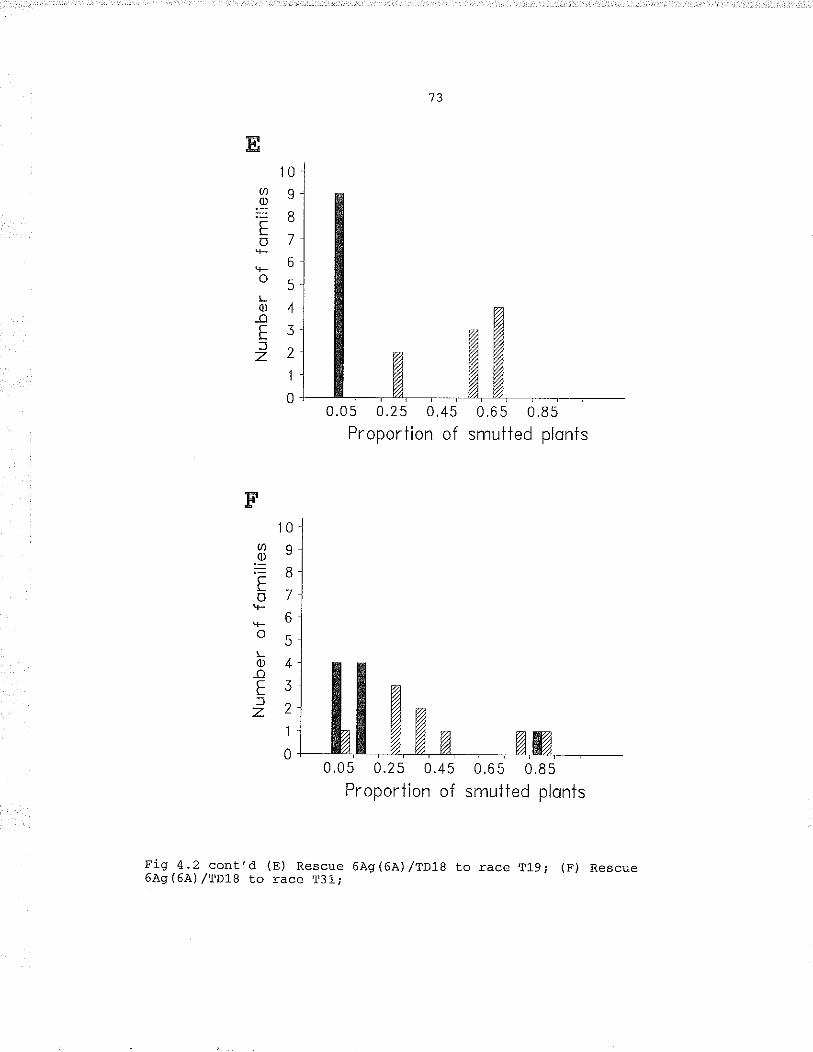
E10
I8
7
6
4
3
2
1
00.05 0.25 0.45 0.65 0.85
Proportion of smutied plonts
l09
8
7
6
5
4
3
2
1
00.05 0.25 0.45 0.65 0.85
Proporlion of smuTted plonls
Fiq 4.2 cont,d (E) Rescue 6A9(6À) /TÐ19 to race T19; (F) Rescue6As(64) /TD18 to race T31;
tJ')
c)
=Eo
oLAJ¡Efz
u)o
.E
o
oL0)
l_J
Efz
F
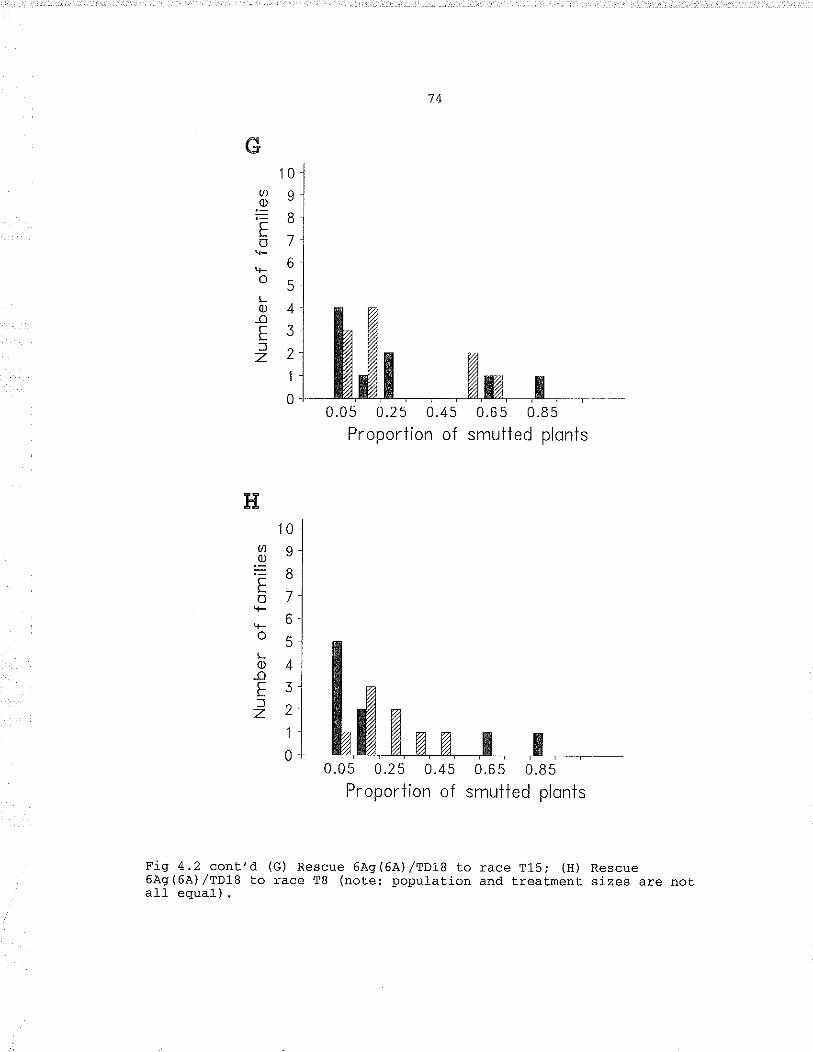
G
a
ÉEo
oLo¡Efz
U)
ÊEo
oLc)-oEfz
10
9
I7
6
5
4
J
2
1
0
10o
I7
6
5
4
3
2
1
0
0.0 5 0.25 0.4s 0.6 5 0.8 5
ProporTion of smulled plonts
H
0.05 0.25 0.45 0.65 0.85
ProporTion of smuTted plonts
Fiq 4,2 cont,d (c) Rescue 6Â9(6A)/TD18 to race TL5i (H) Rescue6AS(6À)/TD18 Co race T8 (not.e: population and treatment sizes are notall equal),

'75
the distribution of progeny when no association occurred is demonstrated
in Fig. 4,2c by the progeny of the Rescue 6À9(6A) /TD1B population when
tested Co race T15 (t-test probability of 0.80) . The lack ofassociation of the 6A chromosome with resistance to races T31 and Tg was
not so clear cut in Che Rescue 6Â9(6A) /TD18 population (Fiq, 4.2F and
H).
one further result noted from this study was the fower ferbility and
viability of lines containing alien chromosomes, compared to euploidplants. This was demonstraCed by the nurnber of F3 seed and plants ineach chromosome group, and the fact that most families tbat were
excluded from analysis because of being too small were classed as havingthe 6A9 chromosome, Às an example, esseneially all seed of the Rescue
6ÀS(64) /TD18 populabion was inoculated. From the 2992 inocutated seeds
planted from plants that were classed as having the wheat. 6À chromosome,
2304 plants, representing Che first. 83 heads inoculaled, survived to be
rated. of che 2617 inoculat.ed seeds from plants classed as having lhe6A9 chromoeome, only 1457 plant.s, representing seed from the first gg
heads, survived to be rated.
DTSCUSSION
The location of a T19 resistance gene appeared to be in chrornosome 6A
of Cadet, based on evidence from segregatíon analysis and cytogeneticstock evaluation, À M,Ab chromosome marker enabled the use of smallpopulations for testing the chromosomal location of ehe loose smut
resistance genes, T19 resist.ance in three other cultivars, which have
rel-ationships to loose smut differentials, or to Cadet, was found to be
associated with the 6A chromosome with the aid of the ¡4A,b marker. The
chromosomal location of genes wiehin the loose smut differential set can
be used to help identify the genes for resistance in the differentialset. îhe T19 resistance gene in Cadet appeared to be in the 6À shortchromosome arm, based on cytogenetic stock evaluation,

76
The prime objective of this study was eo focate a gene or genes forloose smut resist.ance to a chromosome in wheat through analysis of a
segregating population, with the aid of a chromosome marker and an alienchromosome substitution stock. The low levef of loose smut race T19
infection of cadet, compared to the high level of infecti-on of Cadet
6Àq(6À) , indicated thal the location of resis¡ance to race Tl-9 was ínchromosome 6Â, This information al]owed greater effort to be focused on
¿he study of segregating populaCions possessing resistance to race T19,
in which one of the parents possessed an alien chrornosorne substitubionin which the wheat 6À chromosome was replaced. The Cadet 6Â9(6A)/Cadet
and Cadet 6Aq(6À.) /Thatcher popula¡ions were generaCed, and the Cadet
6A95:6À5 and Cadet 6AL t" were inoculated based on the preliminaryinformation lhat a loose smut resistance gene resided. in chromosome 6À,
Ànalysis of the Cadet 6Aq(6Ä)/Cadet poputation not only allowed
confirmalion of the chromosomal location of a loose smut race T19
resistance gene in the Cadet 6A, chromosome, it also affirmed the valueof the MÀb chromosome narker,/alien subscitut.ion fine method for locatinggenes to chromosornes, One segregate from the Cadet 6A9(6A) /Cadetpopulagion, classed as having the 6À chromosome, was more susceptiblethan iÈs other 6Ä siblings (Fig. 4,2e) . This rnay be simply rand.om
variation or a second gene for partial resistance to race T19 may be
segregating. The important point to not.e is bhat if the resistance gene
were not in 64, the probabilify that all the lines classed as not havingthe 6A chromosome afso segregabed as susceptible is remote,
Tbe results aLso demonstrated thae TL9 resislance in Thatcher, TD1g,
and Kota was associated with the 6À chromosome, Nielsen (19g7a)
reported tbat ED4, a selection fror0 Kota, produced infection levels ofless than 10% when inoculat.ed wilh race T19, The Kota used in thisstudy was inoculated to confirm Nielsen,s report (Tab1e 4,1); however,
Che Kota used in this study did not appeax to be as resistant to T19 as
the TD4 described by Nielsen, The segregation for the T19 resistance in

77
the Kota used in this study confj-rmed that the Kota possesses
resistance. The difference in reaction between the Kota here and TD4
used by Nj.el-sen (1987a) may be due to random variation, or Nielsen,s TD4
may have a differenl gene or modifier gene if the original Kota were
heterogeneous, fn addition to the resulls indicating that Kota had a
gene for resistance in chromosome 6À, the results also indicated thaLKota had a second gene for resistance to T31, which was not inchromosome 6À. The same progeny that vrere inoculatêd with TL9 were
inoculaeed wj-fh T3L, yet progeny within each chromosome class were
mainly resistant.Although no dj-somíc 6A9 progeny were identified in the Cadet
6ÀS(6A') /That.cher populalion, doubfe monosomics were evaluated forresistance to race T19 along with the lines disomic for the wheaC 6À,
chromosorne. The susceptibility of Ehe Cadet 6ÀS (6A),/îhatcher doublemonosomics to race T19 indicates the gene is hemizygous ineffective orrecessi-ve. Because all of the lines classed as disomic for Che wheat 6À
were resistan!, and a1l of the lines double monosomic for b.he wheat 6Â
anð. Agropyron 6Ä9 chromosomes were susceptible, the evidence is strongthat the Tl-9 resistance in Thatcher resides in the wheat 6A chromosome,
Unlike Cadee 6ÀS(6À) , \a'hi. ch was highty susceptible, Rescue 6AS(6À) was
only moderateLy suscepeible t.o race TL9. The moderate susceptibitity ofthe Rescue 6ÀS(6A) may have been due to mi s - inoculation. The resutCssho\,ring that. the suscepLible segregates of bhe Rescue 6As(6A) /TD18population vJere more susceptibte than Che Rescue 6A9(6À) tends eo
support ehe mis-inoculaLion hlæothesis. Às for the reaction to raceT3L, Rescue 6AS(6A) appeared moderat.ely resistant, yet, Rescue
inocutated to T31 was highly susceptible. Highly susceptible progeny
did segregat.e from the cross of Rescue 6À9(6Ä) with TD1g, as shown byinoculations with race T31 (Fig, 4.2F), indicatinq that the data for T31
on Rescue 6AS(64) may be an underestimate of the true susceptibility ofthis cultivar to race T3L, In general, workers evaluating the genetics

't8
of loose smut resistance over bhe years have been frustrated by thevariable reaction of this disease (Ribeiro 1963),
The T31 and T8 results were not clear-cut for thê Rescue 6Â,9(6À) /?D1gpopulatj.on. À two-gene system could explain the result.s of loose smut
incidence values in xesponse to inoculation with T31 and Tg. À majorgene segregating for resistance, but not in chromosome 6À, would explainthe approximate 3:1 segregation (aceual nunbers .lt2) of resistant tosusceptible in the 6A9 chromosome class (Fig 4.2F), But, in general,the 6Ä lines were more resistant than the 6,A,9 lines, A second gene in6A could explain the additional xesistance. The difference in loosesmut reaction between the 6A classified lines and the 6A9 ctassifiedlines was not' sCat.istically siqnificani: for the Rescue 6ÀS(6À) /TD18population inoculated to T31 or TB (the probabilities being O.10 and
0.65 respectively) , but these values were confounded by the susceptiblesegregation class. civen the hl¡pothesis that two genes are functioningin a cumulative fashion, four classes resultr R1R2, RLs2, S1R2, S1S2
where R is resistant and S susceptible and L is locus one and. 2 is Locus
2. If a second gene residing on 6A is removed (i.e. 6Àq lines) twogroups of resistance can be formed, those segregating for two genes (6À
lines R1R2, R1s2, S1R2) and those segregating for one gene (6À9 linesR1) . The nu11 h14)othesis is that chromosome 6À does not possess a loosesmut resistance gene influencing the reaction to race T31 or Tg,
susceptibl-e segregates are excruded from this analysis to d.eeermine theeffect of R2. When the t-test was recalculated with t.he susceptiblesegregates excluded, a sÍgnificant difference between the 6A and 6À9
groups was detected at. the 5% probability 1evel, indicating thatchromosome 6À indeed had an effec! on loose smut resistance to races Tg
and T31 in the TD18 popul_ation. Modifier genes, and genes withcumulative and additive effect.s, have previously been implicated inloose smut resistance (Kilduff 1933; Heyne and Hansing 1955, shestakova
and vjushkov 1974). Further study is needed to confirm this potengial

'79
cumulative effect of resistance genes on loose smut.
The segregates susceptible to race Tg fro¡n the populat.ion Rescue
6A9(64) ,/TD18 were also the most. susceptible to race T15, indicating thatthe same gene conferred resistance to these two races, The ratio of 14
resistant. to 5 susceptible suggests that genetic control for resistanceto race T15 could be from one dominant gene, A progeny susceptible toT15 and Tg was resistant to T31, and a different progeny susceptible toT31 was resistant to both T15 and T8, indicating yet another gene inTÐ18. That TD18 could have 3 genes for resistance to loose smut issupporled by lhe few races virulent on it as a differential (Nielsen
19I7a, Person 1959).
The low loose smut reaceion of Cadet 6A95r6À5 and the high reacEion ofCadet 6ÀL t'r indicaeed the presence of the T19 xesistance gene to be inthe short arm of chromosome 6À,
WiCh regard to Che method used t.o locate the T19 resistance qenes tochromosome 64, the Mtic. 230/9 is part.icularly useful as a 6A chromosorne
marker. This antibody binds to gliadins produced at the 6A locus(Skexrit.t et al . 1991; Howes et al . 1993), but did not bind to seed
extracts which had the 6À chromosome substituted by the 6 Agropyron
chromosome (Table 4,1). The absorbance readings of the aliensubstitution lines were consistent L'ith those reÞorted by Howes êt al .
(1993). The 1or.¡ antibody -binding of Che CadeC 6AL t.Í is consistent withthe fact that. the gliadin locus is located in the 6À5 chromosome arm
(Garcia-olmedo et. at, 1982), Bècause this antibody bound. to seed
extracts possessing lhe wheat 6A chromosome, but did not bind toextracts of sèed in rvhich the wbeat 6À chromosomes were replaced by 6À9
chromosomes of Agropyron, the poll¡morphism needed to classifysegregating progeny was satisf ied.
ln principle, the MAb chromosome marker/alien chronosome procedure ismore efficient in terms of the population size required to make a
decision on chrornosome location of a gene than is monosomic analysis (a

80
population of 57 for monosomic analysis compared to g for chromosome
marker analysis). Thè procedure appears to bè robust, accommodating thesometimes \.¿ide variation seen in 1oose smut observalions. The
chromosome marker/a1ien substieution line method seems to be able toacconmodate problems of variation r\'ithin a lrait being analyzed, because
of precise classificatÍon of the chronosome condition. Because sma1lpopulatj-ons are used, re-evaluation of outtiers is feasible forclassification of the seed by the antibody, and, if necessary, highlydeviant smut reactions can be verified by inoculaLing a sample of F3
plants. The method provides cleax results when Che genotype beingstudied possesses a single highly expressive g,ene, The meEhod couldalso be used in more compfex genetic situations, where, for the crossinvolvj-ng ehe critical chromosome, ¿he segregation ratio for n genes inthe wheat. chromosome class is compared to a ratio of n-1 genes in theal.j-en chromosome subs¡itution class, where n is the nunùcer of genes,
obviously, in systems involving more than one giene, population sizesmust be larger when races cannol be selected to selectively remove theeffects of all but one of the resistance g.enes.
The minimum population of lines with and without a particular wheat
chromosome required to deLermine lhe chromoEomal residence of a singlegene may be sma1l, but a population size somewhat larger is useful .
This is because misclassification of F2 seed.s may occur. Àlthough threêchromosome classes are expected, four endosperm classes are expected inthe F2 progeny of a double monosomic plant, A,s was pointed out inChapt.er 3, the level of endosperm protein encoded by a particular alle1ewill vary depending on whether or not the al1e1e is contributed by thefenale or the male (i.e. whether or not the endosperm is triploid),Therefore, the class of doubte monosomics actually represented two
cfasses (one class of endospèrm possesslng Ewo 6-Agropyron chromosomes
and one 6A-wheat, and the second class possessing two 6A-tvhea¿
chromosomes and one 6-Agropyron). The other two classes are reDresented

81
by progeny possessing disomic alien chromosomes and by progeny
possessing disomic wheat chromosomes, Figure 4.1 demonstrates thatpeaks tend to merge. Amonq the most obvious factors which can affectthe leve1 of antibody binding and cause chromosome dosage classes tomerge are (in addition to gene dosage (Chapter 3)) the environmentat
condit.ions for which the developing seed is exposed (Shewry and Miflin1985) , the size of seed piece used for extractíon¡ and the precisionwith rr'hich samples axe dilutèd and handled. Other reasons for having agreater than theoretical minimum population size inctude, that not a1fseeds may survive, and plants may pos€ess problems such as poor vigouror f erti li ty.
With loose smut, there is a second population factor to be considered,namely the family size of inoculated plants. When the leve] of diseaseexpression between the resistant and suscept.ible parent dectines, thenthe family size (i.e, the number of inoculated seed from a single plant)must be increased. In this study, there is evidence for a reduct.ion infertility and/or viability when a J.ine possesses an alien chromosome inplacê of a wheat chromosome. This factor must be accounted. for when
determining the nunber of heads t.o be inoculated.The success of the chromosome rnarker meehod depends on being able to
evaluate a large initial F, population of seed for their chromosome
constituCion. TheoreticaLly the alien chromosome should segxegate withits homoeologous wheat chromosome in a 1:2:L ratio. In this study theproportion of disomics was considered when large populations of F, seed
were initially evaluated to classify the seed based on chromosome ¡nake
up (FiS. 4.1) , A rnajor advantage of monoclonal ant.ibodies to seed
proteins is Cheir ease of use in an ELISÂ, in which many seeds can be
evaluated with minimal effort (Howes et al , 1999), In Lhe experiment.s
here, even though initíal populations were large (between 60 and g0
seeds) they may not have been large enough, as dernonstraled by the Cadet.
6Ä9(64) /That.cher and cadet 6As(6À)/cade! populations (Fig, 4,1À and

82
4.1C) . No explanation can be offered for the cadet 6À9(6A) /Tha¡cherpopulation not generatinq 6A.9 disomic progeny, except that perhaps poor
maintenance of the alien chromosÕrne in thè disonic form in certaincrosses may be a poeential drawback of the use of atien substitution1ines. The fact that segregabion for the disomíc alien chromosome 6A9
was under represented was indicated by thè antibody, and, in hindsight,larger populations of F2 seed could have been evaluat.ed to improve theopportunity of selecting disomic 6.A9 1ines. Not just seeds with thelowest xeadings should be selected, but' those with the lowesL readingswhich are comparable to the low check should be selected. obviously,si"mplicily and precision of classifying the chronosome marker traitcontribute to the efficiency of the overalL analysj-s.
The conmon chromosome location of resistance to race T19 in Che
cultivars Kota, Cadet, That.cher and TDI_8 is evidence thaL these
cultivars may possess the same or a related gene, Knowledge of thechromosomal location of genes withj-n the differential set would providesome indication of the relationship of genes possessed by those
differenÈia1s, and would assist in the developrnent of single gene
differential 1ines, The crosses in this study provide a preliminarystudy of the relaeionship of resistance to race Tl.9 among the fourcul-livars, all of which are related to or are differentials. Kota and
Thatcher are progenitors to differentials TD4 and TD12 respectively, and
TD18 is a differential (Nielsen 1997a) . The Thalcher cross was made
based on the observation that TD12 was resist.ant to race T19, and when
the differential was crossed to make it day-length insensitive as
described by Nielsen and Dyck (j.ggB), T19 resistance teas lost in theimproved different.ial TD12A. Nielsen and Dyck (1988) showed thatThatcher,s value as a differential was not diminished by the loss of T19
resistance during agronomic improvement of this differential . This can
be explained by the fact that the gene lost was d.uplicated in anotherdifferential, for which the present sludy provides evid.ence (both TD4 or

83
TD18 nay have the same Tl-9 resistance gene as Thatcher) , Thaecher and
Kota are of interesL not only because of their use in race
differentials, but also because their differentiat derivatives TD4 and
TD12 were used in the analysis of virulence of races of loose smut
(Nielsen 197'1 and t982\ , cenes for resistance to loose srnut in TD4 and
TD12 have been named by Mclntosh (1988), based on virulence inCeractionsof Loose smut races (Nielsen 1,9jj and !9821 .
Nielsen's (L977, f982) genetic analyses of virulence showed Kota has a
gene (Ut.2) different from TD12 (UtA), Because TD12A becamê susceptibleto only race T19 during agronomic improvement as a differential, the T19
resisCance must be conditioned by a second resistance gene in TD12.
Race T5 is virulent. on TDI_2 but avirulent. on TD4, and was one of theraces used by Nielsên (1982) as a parent in the cross for virulencesègregation analysis. For this difference in reaction between TD4 and
TD12 to occur, T5 nust have virulence on the T19 resistance gene inTD!2. The avirulence of T5 on TD4 indicates Ut2 is the effective geae,
and the TL9 resistance gene (assuming the T19 resistance in Kota and
TD12 is the same) is a thiïd gene between the two differentiaLs,Ribeiro (1963) first suggested Ut as a sl¡nbol for genes resistant toloose smut of wheat. He labelled Kota resistance to race C2 as Utc2.¿,
meaning the fourth gene discovered to be resistant to race C2. Mclntosh(1988) designated 4 genes for resistance based on the gene-for-gene
concept of hosC pathogen interactions and Nietsen,s (19?7 and 19g2)
identification of virulence genes. Mcfnt.osh designated FlorencexAurore,Renfrew, and Red Bobs as having Ut1, Kota and iJittle Club as having Ut2,
Carma Ut3, and ThatcherxRegent Ut4. Therefore, I propose the secondqene in TD12 be designated Uts. Further study is required to determinewhether or not this is the same gene as that found in Kota, Cadet, and
TD18 ,
The results of this study demonstrate the value of using a seed-
protein-specific monoclonal antibody as a chromosome marker in

84
conjunction with an alien subsCitution line in the cyt.ogenetic analysisof wheat. Gains in efficiency are realized in two ways: The correlativemarker method is robust and requires a smaller populalion size than
monosomic analysis; By using M.Abs, more rapid analysís of a populationis achieved through an EL]SA and sampting seed êndosperm, in contrase toconventional cytogenet.ic analysis of root tips of seedtings or of po1len
mother cel1s with microscopjc observation. The use of al-iensubst.itution lines, and Che production and analysis of F2 populationsfor disomic individuals, a11ows for evaluation of recessive genes wieh
the sane ease as dominant genes.
The M.Ab 230/9 i,s a good indicacor of chromosome 6A consticution based
on the relationship achieved with loose smut xeaction in the segregatingcadet 6A9(6Ã.)/cadet population compared with the evidence from
inoculation of the cytogeneeic stocks indicat.ed a 6À short location forT19 loose smut resistance in Cadet. This study is indicative of thepotential value of other chromosome marker/arien chromosome substitutionline combinaeions for locating g.enes to chromosomes. These resuLtsprovide the first denonstrat.ion, ehrough genetic segregation analysis,of the chromosomal location of a ]oose smut resistance gene, and thefirst tentative locatj-on of such a gene in a chromosome arm. The
monoclonal - antibody -based cytogenetic technique applied here can be used
to assist ehe chromosomal location of a wide variety of difficult-to-evaluate wheat t.raits, The identification of the chromosomal locationof genes should be seen as a way of facilitating future efforts of gene
taqging with biochemical- markers such as RFLps (rest.riction fragmentlength poll¡morphisms) or RÀpD (random amplified polymoïphic DNÀ),

85
CTTAPTER 5
The AssociatÍon Befween a Wheat Chromosome
6A Gene Controlling Loose Smut Resistance in
Wheat, the Sr8 and Gliadin Gene loci, and an
RFLP in the Same Chromosome
ABSTRÀCT: Linkage was tested anong a group of chromosome 6À, traits.
Homozygous recorìbinant progeny for the 6À short. chromosome arm of lhepopuLation Cadeb. 6AqS ! 6A5,/Chinese Sprinq Sr8//Cadèt 6À9(6À) (where Ag
represents Agropyroo. chromosome and S, short arm) were tested t.o stem
rust race C10 and loose smut race T19, and evaluated for gliadin bandingpattern using polyacrylamide ge1 electrophoresis. DNA samples takenfrom the progeny were cut using restriction enzymes, and the cut sarnples
were probed to detect RFLps. The loose smut resistance gene was nèitherlinked bo the gliadin, nor to the stem rust resistance genes, However,
the 6Â loose smue resist.ance gene was associaeed \arith an RFtp.
Geneb.ic markers eiqhtlyare potentiatly easier
for resistance to loose
INTRODUCTION
linked Lo genes conferring loose smut resistanceto evaluate than reaction to the disease, À gene
smut race T19 has been located to chromosome 6.A

86
(Chapter 4 of this ¡hesis) , and is likely in the short arm of thischromosome, Âlso located in the short arm of the 6À chromosome are a
gliadin gene (shepherd 1968; carcia-olrnedo et a1 . 1978, 1982i payne
1987; Mclntosh 1988) and the stem rust resistance gene SrB (Sears et al ,
1.957r Mclntos}j, 1972t Sawhney eE al . 1981) . .As well, RFLps have been
located to thj-s chromosome (Sharp et al , 1988a, l_989; Heun et al . 1991;
Liu and Tsunewaki 1991; Ànderson et al . 1992).
In this study, a population was esCablished which demonstrated
poll¡morphism for a11e1es at the gliadin locus and the loose smut
resistance 1ocus, Progeny were also evaluated for the Srg st.em rusbresistance gene, and a sample of RFLP probes was evaluaeed forpoll¡moxphism between the parents and among the progeny. The purpose ofthis study was to det.ermine the l-inkage relationship between a 1oose
smut resistance gene located in chromosome 6À with the gliadin locus
also in the same chromosome, lhus allowing assessment of the value ofthe gliadin locus as a marker for the Loose smut. resistance gene.
Linkage analysis was also done between the loose smut resistance,gliadin and the Sr8 stem rust resistance gene 1oci, and a group ofrestriction fragment. length poll¡morphism (RFLP) probes located in thischromosome .
MATERIAIJS A}IÐ METHODS
PLant Material
The translocaÈion line Cadet 6Ä93:6ÀS (Dr. E,D,p. Whelan, Agriculturecanada Research st.ation, Lethbri-dge, Àlberta provided this and otherCadet cytogenetic stocks) , resistant Lo U. tritici race TL9, was used as
the female parent in a cross tvith Chinese Spring Srg (from the 1ate Dr.E.R. Sears, Dèp. of Àgronomy, University of Missouri, Colunìlcia, MO)
(Fig, 5,1). The latter line was deveÌoped by backcrossing the srg stem
rust resistance gene ineo the cultivar Chinese Spring (Sears et al .
1957). rhe Cadet 6À9S!6AS is an alien translocation tine in irhich the

87
Cadet 6À95:6À5 X Ch. Sprinq Sr8 6A
6.qgs 6Ä5 6ÀL 6Às
iifrìfÈåïiþr EilI
-r
t/11T19 / cl i<-'-+sr8
Resis È. ////////
J,F, X cadet 6À9 (6À)
6Àss:6Äs ä,ì$,{{f*Ìi,.- li,fltdÆl{ilÍ{lå1#"trå.Ìl.ilJÍ *"6À F,- ijüiiiillili';iii#'jjf;if"d,^"I
6À short arns undergo I
crossiDg over I
I.!TC Fr progeny
6Ä95 arns pair double monoso¡nic:between 6À95:6À5 no pairing bêtween
and 6À9 the 6À and 6À9
ör-tll"lif'ht'r o¡ rxr Èit$lllf.Hån#..ffi¡dl!.ififliÌ[':ilüãiÍ,i11*rüï-iliil.ir1r:ïi+tii:"in*i tïtl; r rìr.riq
segregatj.on MÀb Tc F2 Proqen'ratio binding- ÈË,iÎt$t'*'È-
-xI
t hiqh i*li:"Î- l*#ji,f+þ,r
-xIsifìr'#.lïþr nxr ëÌifflüffiTi.ffiåLi$jïtr#z lnÈelmediate ;;i;,_"".i#iqiUil id¡!;iitii i4iii i.i i l+ll
Èir[,1filf¡.T¡iíìirntiÏÌiiiÍ]i ri fl,Ìfrfit"tt$t$¡l. iijgÌlüÌ1 low rfrfrfÍl]l: !+"1rlri'"iis4.ll u tt*t
l¡ii"li"lilüÞ,iiiililii$iiiîi ÈiüiiittlËJ$ är¡$Tl#flfl-'
I+S. 5 ,1. Crossing scheme used for the Cadet 6A95:6A5 alien translocationline in a cross with chinese (ch.) Sprinq Srg úestcrossed \eith the atienchromosome substilution line Cadet 6ÀS(6Ál .

88
long arm of chronosome 6 of Cadet is replaced by the short arm ofchronosome 6 from Agropyron (v.¡helan and Luko\,¡ l_990) . Two plants ofChinese Spring Sr8 (designated CS Sr 8À and CS Sr 88) were used as
pollen sources. when selfed progeny of each were tested with slem rusbrace C10, only the CS Sr8À plant was found to be resistan¿. A testcrosswas made using Cadet 6AS(6À) as the testcross female parent, This linelacks the wheat 6å, chromosome and the Tl_9 resistance gene (Chapter 4
this thesis) , The h/heat 6À chromosome from the chinese Spring Srg and
the 6À5 chromosome arm from cadet 6Ã,gsr6Às will have no pairing parlnerwhen the F1 of Chinese Spring Srg and Cadet 6AqS:6ÀS is crossed withCadet 6A9(6À), fn such a cross, the only pairing event allowingcrossing over, occurs in the 6À short arm of chromosome 6A duringmei-osis of the initial crass.
Eighty testcross (TC) F1 seeds were planted and grown !o maturity. Inan attenpt to minimize growth cabinet usage, five TC seeds per pot wereplanted, with tillers trinuned in an att.empt. to produce one good head perplant. Â11 eighty seeds devel_oped plant.s, only two of which produced
less than 10 seeds. Up to 10 seeds {TC Fr) from each plant were
evaluated with the monoclonal antibody (MÀb) 230/9 (provided by Dr. !T.H.
Skerritt CSÌRO, Sydney Australia) , which binds to the protein coded bythe gliadin locus in chromosome 6A (Howes et al. 1993). The level ofantibody binding indicates gene and therefore chromosome dosage (Chapter
4 of ghis thesis), The one seed with the highest bindinq from eachgroup of 10 seeds h'as planted, in an att.enpt to recover planEs disomicfor either the 6À chromosome or the 6A9516A5 chromosome. The resultingplants v¡ere grown in the gro\,¡th cabinet in individual pots, and two tothree heads h'ere j.noculated wiCh a water suspension of teliospores of f/.
tritici race Tl-9 spores (1 ng mt'l) . The loose smut inoculated heads(TC F3 seed) were harvested individually and plant.ed in lhe greenhouse.
Seed was collected from 63 plants, being those remaining after some
families produced no high an antibody bindinq seed in the F, and lack

89
of emergence or sterility in others, Five uninoculated TC F3 seeds fromeach TC F2 plant were used for electrophoretic and antibody analysis,The embryo ends of thesè seeds were grow¡ out and plant material was
harvested for DNA extraclion. Two sets of five TC F3 seeds of plantsderived fxom the CS Sr BA rust resistan! parent were also grown and.
inoculated wíth st.em rust race c10, and, as well_ Cs Sr gÀ and cadet wereplanted as coneroLs.
Rust inoculations were done l¡ith race C1O, which is aviru].ent on theSrB gene. Rust spores were rubbed on damp leaves with lhe fingers, orby mistíng of a spore suspension from a spray bottle. Inoculated plantsr.¡ere placed in a dew chanlcer overnight. plants were removed from thedew chamber in the morning, and covered with clear plastic caps tomaintain moisture. The plants were placed under bright light for theremainder of the day at which time the ptastic caps were removed,
Growth cabinet8 and greenhouses were operated at 15 to l-g"C, ¡{ithsupplemental liqht provided in the greenhouse. Day 1ength was 16 hlightandShdark,
Enzyme Línked fmnunosorbant Àssay
MÀb analysis \a'as donè using an enzyme linked immunosorbant assay(ELISÀ). For detailed procedures, see chapt.er 4 of this thesis. Seeds
were cut. j.n hatf so that the eÍìbryo end coutd be grown for otheranalyses, while the endosperm end was extracted in 50% propan-2-o1 . The
extract was bound co microtitxe plates, and ELISAS were performed usingMÃb 230/9, which is specific for protein coded by wheat chromosome 6A,
As ment.ioned, 10 Tc F, seed from each plant were analyzed in an attemptto obtain disomic seed for eiLher ¿he 6À9516À5 chromosome or for lhe 6À
chromosome. Five TC F3 seeds werè evafuated to confirm the disomiccondition.

90
Polyacrylamide ce1 ELect,rophoresÍs
Acid PAGE gels were run on seed èxtracts of Cadet., Cadet 6À93:6A5, Cadet
6As(6À) , chinese spring sr8, and Chinese sprinq 6AL ditetosomic (t,,),Chinese Spring 6A5 tÍ, prelude Sr13 (Che Chinese Spring cytogeneticstocks were originally from the IaLe Dr. E.R. Sears, and maintained and
provided by Dr, E.R. Kerbèr, and prelude Sr13 by Dr, D. Harder both atÀgriculture Canada Research Station, Winnipeg, Manitoba) to dêterminewhich gliadin band(s) corresponded to the 6À short arm 1ocus. The
extracts fron TC F3 seed used for the ELISÀ were also used inpolyacrylamide gel eleccrophoresis (pÀcE) at 1ow pH. six percentacrylamide with 1,25% bis-acrylanide was prepared at pH 3.1 usingalumínum lactate buffer. cels were run for f 3/4 h at 540 V at 7oC withsodium lact.ate bottom buffer pH 3.1 and aluminum lactate top buffer pH
3.1. Thê antibody bindinq was correlatêd to pÀcE band variation todetermine wheeher or not the antibody 230/9 could detect atlelicvariation,
Iroose Smut InocuLationg
ïnoculum was prepared and inocufaLions were done using a syringe and
needle as described in Chapter 4 of this thesis. Àt teast two hèads ofeach TC F, plant were inocul-ated, and selected F3 plants were inoculatedto confirrn results of the F, inoculat.ion.
DNA Ànalysis
Plants derived from the embryo end of lhe seeds used for acid pÀcE
analysis were germinated and allowed to groh, to the four Lo five leafstage. The leaf material from the up to five seedlings of the same
family v¡as harvesCed into perforated polyethylene bags as bulks.Bulking of leaf material from the up to 5 planEs was based lheirhomozygosity for the 6A5 chromosome arm. Irunediately after each samplewas harvesled the leaf material was cut into 2 to 3 cm pieces and

9T
submerged in liquid nitrogen. The plant nìaterial was transferred fromliquid nitrogen to a freeze dryer, and lyophilized at 10 um Hg vacuum
pressure at room têmperature fox Cwo days. FollowÍng drying, aamples
were stored on sílica ge] at -20'C until the DNÄ was extracted.The following steps were used for DNÀ extraction. Up to 250 mg of
plant material was ground in a 1l_5 run mortar with a pêst.le, \,¡ith 3 g ofacid-washed sand and 25 mL of liquid Nr. The sample was transferred toa 50 mL Teflon cent.rifuge tube, To each Lube was added 20 mL of lysisbuffer consisting of 1,Lt CTÀB (hexadecyl trimethylammonium bronide) , 55
nM Tris pH 8, 55 mM EDTA, ( ethyt enediaminetetraace Cic acid) pH g and l-.54
M NaCl aE pH 8, followed inunediately by the addition of 50 uL proteinase
K (1- mS/mL). The samplè was vortexed a\ð. 2.2 mL 20% SDS was added,
followed by gentle mixing. The samples were allowed to digest tor 2 hat 65oc wich intermittent gentle mixing. Tubes were fil1ed with 24:1chloroform: i soamyl alcohol (about 15 mL) . Samples were rocked and
allowed to mix for L/2 h. Samptes were centrifuged for 20 min in a
table-top centrifuge at abouC 2,500 rpm at room t.emperature, The upperliquid phase was transferred to a cl_ean Teflon tube, A volume ofpropan-2-ol equal to 0,6 times the volume of the aqueous extract (about
9 mIJ) was added to precipitat.e the DNA. The DNÀ was spooled onto a
glass rod and washed with 70% ethanol . The ethanol was alfowed to drainfrom Che DNÀ, which was then di-ssolved in 2 mL of steril-e water in a 30
mL Teflon tube overnight. at 4.C. Each sample was trea¿ed with 7.2 uL of10 mg,/ml beat-t.reated ribonuclease (RNÀase) in TE (10 nìM Tris, L mM
EDTÂ) for 2 h at 37"C. To reduce the amount of carbohydrate extractedwith the DNÀ (the carbohydrate int.erfered with dissolving the samples inwater), a second extraction with l_% CTAB, 1 M NaCl and 1X TE hras done
for each sar|lple. Six millilitres of phenol ¡ chloroform: i soamyl alcohol(25|24:L) was added to each sample with mixing. samples were
centrifuged for 15 min, and the upper aqueous sorution was transferredto a clean tube, Chloroformr isoamyl atcohol (4.5 mL, 24:1) was ad.ded.

92
and mixed, and again centrifuged for 15 min. The upper aqueous solutionwas saved in a clean lube, precipitated with 95? ethanol and 1,/1_0 volume
3 M sodium acetate (pH 5.2), and stored at -20"c. The ÐNÀ was rinsed.twice in 70% êthanol and dissolved in sterile water, The concencrationof the DNÀ in each sample was determined and adjusted to 1.5 ug/ul afterquantification spectrophotome trical ly at 260 nm. Restriction of the DNA
of each of the parents was done with restriction enzymes Eco RV, Hínd.
fII, Ðra I, Xba I, and Bst IIE (cibco BRL), and probed with a series ofchromosome group 6 pxobes (provided by Ðr. Ä. craner, InsÈ.ítue furResi s tenzgene tik, crunbach, cermany; Dr. M.D. ca1e, TPSR, Canbridge, UK,
and Dr. Won Kirn, Àgriculture Canada Research Station, Winnipeg, Canada
as part of the North Ànerican Barley cenome Mapping group) bo determinethose which produced poll,'norphism between CS SrB and Cadet 6À95:6À5 and
to see which bands were associated with the 6À chromosome. Eco RV and
Hind III restrictions resulted in polyrnorphism, and these enzl¡mes were
used on DNÀ of the progeny, Fifteen micrograms of DNÀ Ì^,as cut with 40
units of the restriction enzl¡me in the appropriate reaction buffer, The
samples \,¡ere incubated overnight, loaded on a 0.9% agarose gel (20 x 25
cm and 5 nm thick) , and run for L10O v-h. The ge1 leas blotted onto a
zeta Probe cT (Bio-Rad) nylon membrane for 14 h overnight, using an
alkal-íne blottinq syscem, The membrane was rinsed in 2X SSc (sodj-um
chloride-sodium cit.rate made as a 2OX seock with 173 q NaCl 8g.2 g
sodium citrate adjusted to pH ?.0 with NaOH and made to 1 L withdeionized (Dr) wat.er) (Sanìlcrook et a1 . 1989) and baked at 80.C for 40
min.
Probing involved first blocking the menìbrane with 3 mL of 5 mg/ml ofautocfaved salmon sperm DNÀ boiled 10 min (to denature the DNA) in a
prehybrj.di zation solution consisting of 19 mL of water, 6 mL of 5X HSB
(30,3 q PIPES Ínto 300 mL Dl wat.er pH 6.8 and 600 mL 5 M NaCl and 40 mL
0.5 M EDTA Nar2Hro made to 1 L with DI water) , and 3 mL of modifiedDenhart,s III (2 g 1% BSA (bovine serum albunin) , 2 q Fico1t_4l}, 2 g

93
PVP-360 (polyvinylpyrol idone ) , 10 q sDs, 5 g Naapro?-HrO and 100 mf, ÐI
wat.er, pH 8,0) at 65'C for 6 h in a HyBaid oven, The probes were
label-Ied by the randon primer labelling method using 32p adenosine (BRL
Randorn P¡imers DNÀ Labelling system), and precipitated with the additionof L0 uL of autoclaved salmon sperm DNA, 1/10 volume of 3 M sodium
aceiate, pH 5.2, and 2,5 volume of 95% et.hanol . The solution was placed
at -20oC tor 2 h, centrifuged for 25 min ín a microcentri fuge, \,,/ashed
once with cold 70% ebhanol, dissofved in TE pH 7.6, and. placed in a
boilinq water bath for L0 min. The probe was suspended in 30 mL freshprehybridi zation solution to produce the hybridization solution. The
prehybridi zation solution was poured off the blots and the hybridizationsolution was added. probes were allowed to bind for 16 h at 65oC,
Blots were removed from the hybridization tubes, transferred to trays,and washed two times for 15 min in 2X SSC wiLh 1t SDS, and tLro times for10 min in 0.35X SSC with 1% SÐS at 65oC. Excess wash solution was
blotted from the menbranes, and bloLs were wrapped in Saran plaslicwrap. The blots were exposed to Kodak XARs XO mat X-ïay film incassettes containing intensifying screens for up to five days at -?0" C.
RESUIJTS
segregates of the cross Cadet 6À95 | 6A5 /Chines e Spring Srg//cadet 6A9(6A)
were classified for loose smut resistance, 6À gliadin proteins, stem
rust resistance, and an RFLP. The TC F2 seed within a famí1y \{ere
expected to consist of dísomics for the 6.A95:6ÀS translocation,hetexozygotes for the 6Ä95:6ÂS translocation, and 6A9, and disomics for6À9 in a 1:2:1 ratio (Fig. 5.L) . Alternatively, a family could possess
disornics for che 64, double monosomics fox the 6À and 6A9, and disomicsfor the 6À9 in a 112¡1 rahio. The seeds with the highest antibodybindinq were expected to be either those disomic for the 6AgS:6Ã,s, or

94
those disomic for the 6À chromosornes. Eiqht of the families did. not.
contain seeds producinq high bindinq, therefore seed from those familiesr,¡as not. planted, The remaining 72 families possessed seed ranging from
low binding to high bindinq. Because double monosomics were not
available as checks to provide a mean and standaxd deviation to which tocompaxe the segregating seeds, and because the binding values of the
double monosomic distributíon probably overlapped wj-th disomic 6A
values, it was impossible to predict which intermediate values were from
double monosomÍcs, allowing some double monosomics to be selected. The
lowest. binding seed kept had a corrected absorbance (relative t.o thehigh control) of 0.2. Nine Tc F2 plants either did not survive or didnot produce seed, leaving 63 families. One plant was inocufat.ed withloose smut but no additional seed was available for PAGE and the ELISA,
dropping the total number of useful families to 62.
The PÀcE protein band designaCed Cdt in Fiq. 5.2A was absent in Cadet
6AS(6À) and Chinese Spring 6AL ttt indicating this band is coded by the6.4. qliadin locus on 6ÀS, Chinese Spring Sr8 had a different band in thesame region of the gel as 6À band in CadeE. Five TC F3 seeds
representing each family were evaluated for their gliadin bandingpatLern, using Àcid PÀcE, and the same èxtracts were also used for theELISÀ. Fiq, 5.28 provides an example of lhe segregation for the 6À band
in lhree families. Note that. Chinese Spxing Srg.A has an additional fastrunning band at the bottom of lane 1 (Fiq, 5.28) compared to Chinese
spring srSB (lane Lg) , addinq Èo the evidence of heterogeneity in thisseock, There \,Jere no discrepancies between Acid pÀcE bands and the1evel of l.mb binding for thÕse families which uniformly showed eitherthe Cadet 6A gtiadin allele or the Chinese Spring Srg gliadin aftele.That. is to say, thêre were no families in which a low absorbance
corresponded Co a 6À band, Ho\,¡ever, this \r'as not the case for familiesj.dent.if ied as homogeneous not expressing a 6À band; four families showed

Fig. 5,2A.. PÀcE analysís is as follo\as for the lane and cultivarrchinese Spring Sr8;2, Cadet; 3, Cadet 6Aq(6À) ;4, Cadet 6Äqs:6À5;Chinese Spring 6.A5 t'tr 6, Chinese Sprinq 6ÂL tnr ?, prel_ude Sr13.
r,
FiS. 5,28, PAGE analysis of five TC F3 seeds represenling three famil-ies(1anes 3-7, 8-L2, f3-171 of Che cadet 6Âgs : 6À5 /¿hines e SirLnq SrB//cadeL6As(6À) population and controls chinese Sprinq Sr8À (Ianè 1)
-ana òLineseSpring SrSB (lane 18) and Cadet (lanes 2 ãna fg) .
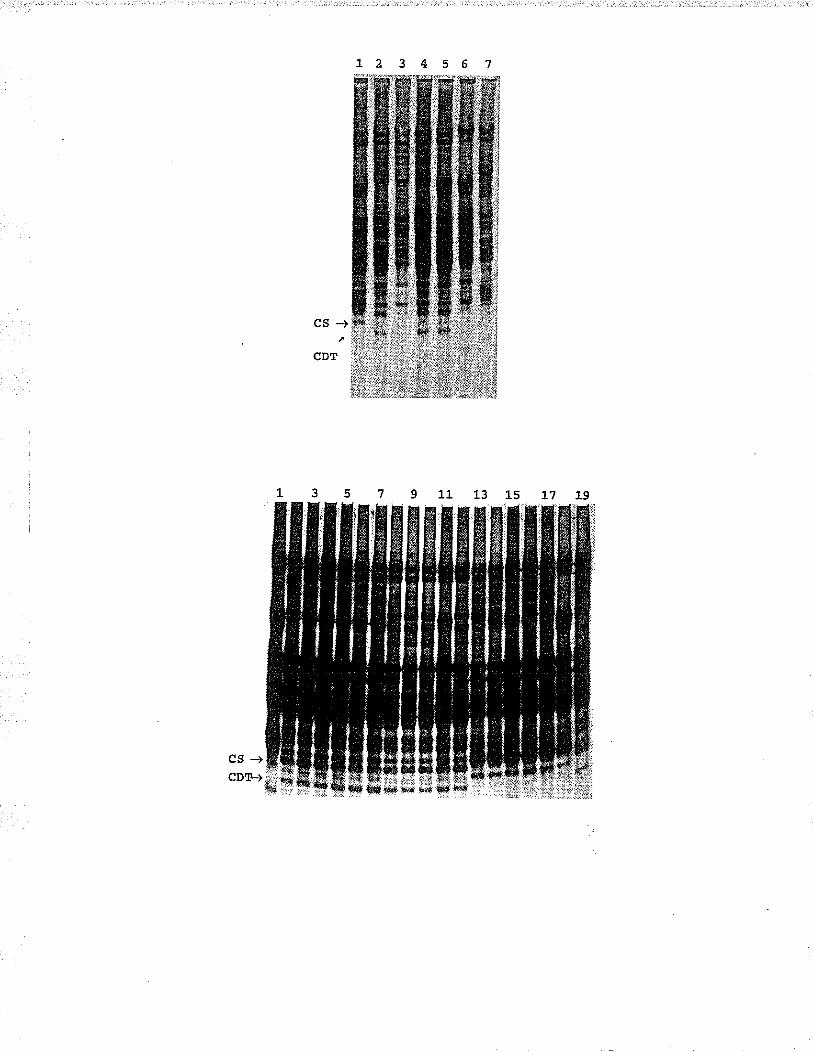
Cs -)
CDT
7 911 13 15 L7 19

96
dj-screpancies between Àcid pÀcE result's and antibody bindinq (i.e.with 1ow antibody binding possessed the pÂcE band).
6
4
2
0 25 45 65 85Percent loose smut
Fiq,..5.3. Segregation of Cadet 6À95 | 6ÀS/Chinese Springsrg/ /cade| 6Âq(6À) TC F3 famities for reaction to toose smuirace T19.
Thirty-nine families were uniforrn for Acid PAGE banding (and EIJISA)
across the five seeds tested, Thirty-seven of the thirey-ninehomogeneous families had sister seed inoculat.ed to race Tl.9 of toose
snut. The segregation of families for loose smue reaction is shown inFig, 5.3 (tick narks on the horizÕntal axis demarcate class intervalsand g¡re values represeng bhe midpoint for each interval). There
appeared Eo be ts¡o populations¡ those having greaeer than 25% loose
U)
ÊEo
o
0)_tl
Efz
12
10
8

9',7
smut, and those having less than 25% loose smut. To determine whether
or not loose smut resistance to race TL9 was linked to the qliadin1ocus, the population was divided into families having the Cadet 6À
qliadin atlele and those having the chinese spring Srg qliadin a11e1e.
Each of the qliadin classes were further divided as being resistant orsuscepeible to T19 (Table 5.1) . A Chí square test showed only randorn
assoxtment of alleles, indicat.ing that these genes were not linked,
Table 5.1. Segxegation of Cadet 6À95:6A5 and ChinesêSpring Sr8 for a 6Â gliadin band and loose smut xace T19rèsistance and a chi square to test deviation from a1:1:1:1 segregation ratio
Res IRace
Resis
Tot
Ratio1:11t 1,
Trai tcliadinLoose smutconbined
chi square00.1081.054
Prob, =3.843.847 -82
0.0s
NSNS
df11,
3
Several setbacks occurred in trying to obt.ain stem rust reactions ofthe population and checks. Seedlings from the two parental 1Ínes ofChinese Spring sr8 showed variation for reaction to race C36, One linegave a tlæe - tv¡o reaction, whereas the other gave a llæe-four reaction,See l(nott (1987a) for a description of reacLion tlæes, Based on thisinformation, only those families derived from the resistant Chinese
Sprinq Sr8 parenl were tested for sEem rusl reaction. ÄIthough race C36
is avirulent on Sx8, it was found that Cadet. and Cadet 6À95:6A5 were
also resistant (qiving a tlæe two react.ion) to this race. Race C1O was
select,ed because it. has wide virulence, yet is avirulent on Srg (Green
istance toe T19
cl-iadin À]lele
ToLaCadet 6À95 | 6A5 Chinese Spring
tant L0 7 L7
usceptible I L2 20
18 19 37

98
1981) . Cl-o was found to be virulent on Cadel (reaction t]¡pe 3 to 4) ,
vJhile avirulent on Chinese Spring' Sr8 {tlt)e-two xeaction) , À shortage
of Cadet 6À95r6.A5 seed hindered the evaluation of rust react.ion on lhislj-ne. In a first round of inoculat.ions of the progeny, up to fiveplants per family \,¡exe inoculaeed wieh race C10. Àlthough few clearlyresistant lines could be detected (i.e. characterized by the Llæe - tr^ro
reaction) , some lines showed a greater level of resistance than others.The families were cfassified as susceptible, questionable, and
resistant, A second inoculation of the families, this time at thejointinq stage, produced símj.lar results, Ägain, there were those h'hich
were clearly susceptíb1e and those which were difficult to classify. An
aCLempt t'o evaluate additional races on the parents for further study ofChe progeny failed because of a powdery mildew infestation,
One optíon was tÕ group lhe stem rust susceptible famílies togetherand group those with questionabfe sten rust reactions together with theresistant class. When lhe results were compared with the values forloose smut reaction and with gliadin banding pattern of ¿he same
families, linkaqe was indicated by the low nurnber of reconlcinant
families (Tables 5.2A, and 5.2C) . The parental tl4)es axe circled inthese tables, and are the predominant classes. The low nunìber ofreconbinant progeny was true for both the stem rust resistance and theloose smut. resistance conbination (Table 5.2D) , and for the rustresistance and gliadin combination (Tabte 5.28) when the families withquestionable rust ratings were classified as rust susceptible. The
population size of L8 is somewhat small for the Chi square tèst, where a
minimurn of five observations for the smallest class for a totat of 20 isrecommended, with the caution of the population size in mind, the Chi
square test was run on each of the groupings, and is shown in Table 5.2.When tesged to a 1:1:1:1 expected segregation ralio, the Chi squareprobabilities were not signiflcant ag a 5% leve1 for each grouping of
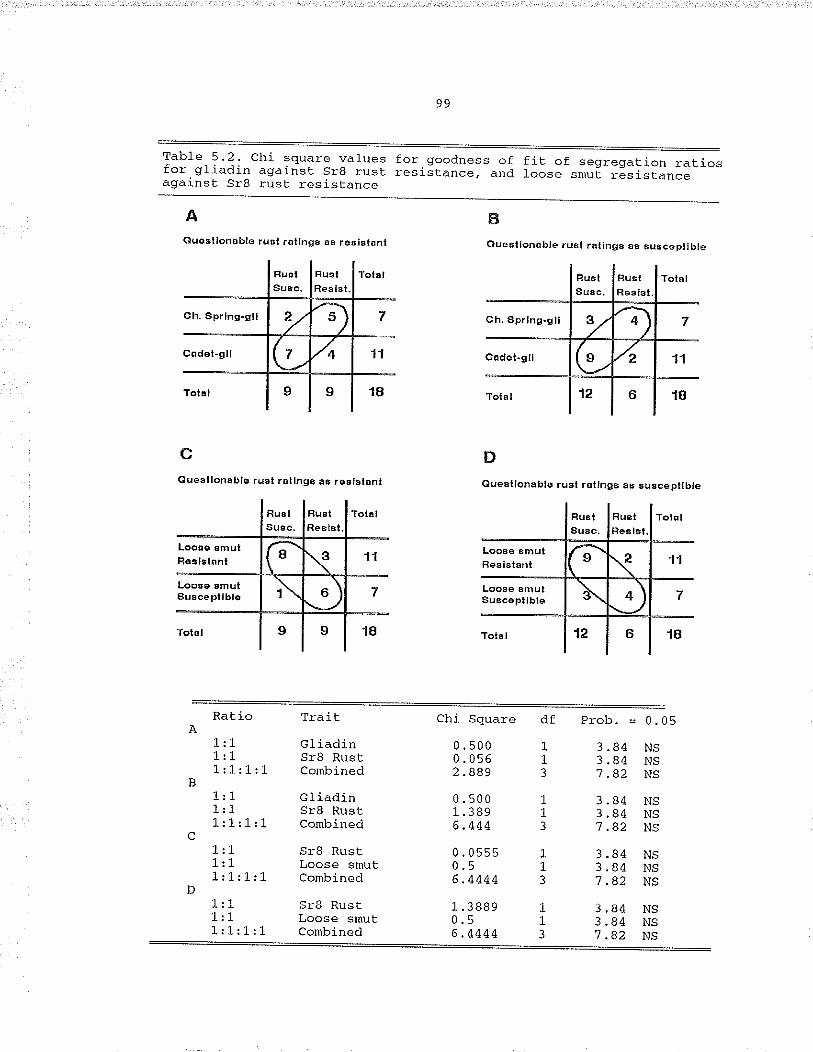
99
Table-5.2. chi square values for goodness of fie of segregation ratiosfor,gliadin againse sr8 rust resiãtance, ana iãå.e smut resistancêagainst Srg rust res i s Eance
AõuBBl¡onable ¡usl ¡'åtlngs ôs reslslanl
cOuesllontrble rust rallhgs as reslslant
B
Quesllonable rusl ¡'alings as 5uÊcepllble
D
Gueatlonable rusl râllngs aB susceÞl¡bfe
Rat io
L¡1
1:L:1:L
1:L1:11.1.1,'1
'I .1
1¡11r1:L¡l-
Trai t
Gl iadinSrg RustCornbined
GI iadinSrB RustCombined
Srg RusgLoose smutcombined
Sr8 RuslLoose srnutcombined
Chi Square
0.5000.0562.889
0.5001.3896 .444
0.05550,56 .4444
1.38890.56 .4444
Prob.
3 .843.84'7 .82
3 .843.847 .82
3 .843.847 .82
3.843 .84
= 0.05
NSNSNS
NSNSNS
NSNSNS
NSNSNS
df
113
1,
1:LL¡1:1:1

100
values, Because the probability of obtaininq the Chi square value
obtained for three of Che combinations was reasonably low (between 5 and
10%), and considering the small sample size used, linkage of lhese
traj-ts should not be ruled oue without further testing. Tèn DNÀ probes
\¡¡ere evaluat.ed against Chinese Spring Sr8 and Cadet 6A95:6.A.5 and Cadet
6ÂS(6À) DNÀ (Table 5,3) in search for poltaTìorphisrn that could be used as
Table 5.3. Ä list of DNA probes located to lhe Triticeae 6chromosomes and the source of the probe
Probe Armidentity probed
Source
MWG514 LMWG620 S
CMt¡lG6 5 2 ScMwG653 scMwG669 LMWG820 S
2238458
1,6'7 s754 L
Dr. A. GranerFederal Biotogical Research Centrefor Agriculture and ForestryInstieube for Resistance ceneticsGrunbach, Germany
Ðr, w. Kin (NÀBGM)Agriculture CanadaResearch Stationwinnipeg, canada
Dr, M.D. caleCa¡¡b¡idge LaboratoryNorwich, U. K-
additional markèrs for loose snut resistance, and as a chromosome marker
for locating the loose smut resistance gene relaCive t.o the cent.romere.
Up to five restriction enzl,nes \eere evafuated in an attempt to improve
the chance of obtaining a useful poll¡morphism. All- probes bound tosouthexn blots producing two or more bands. One probe was found to be
associated with Lhe 64, chromosome because a band produced by cadet
6ÂqS:6AS l\¡as not produced by Cadet 6AS(6À) , but no polyrnorphism between
Cadet 6ÀqS:64,5 and Chinese Spring Srg was found, In most cases no
pollanorphism and no association with the 6A chromosome occurred. when
polymorphism did occur it. was inconsj_stent across enzymes, i,e,, some
enz]¡mes produced poll¡morphism and some did not., One probe (l_67)
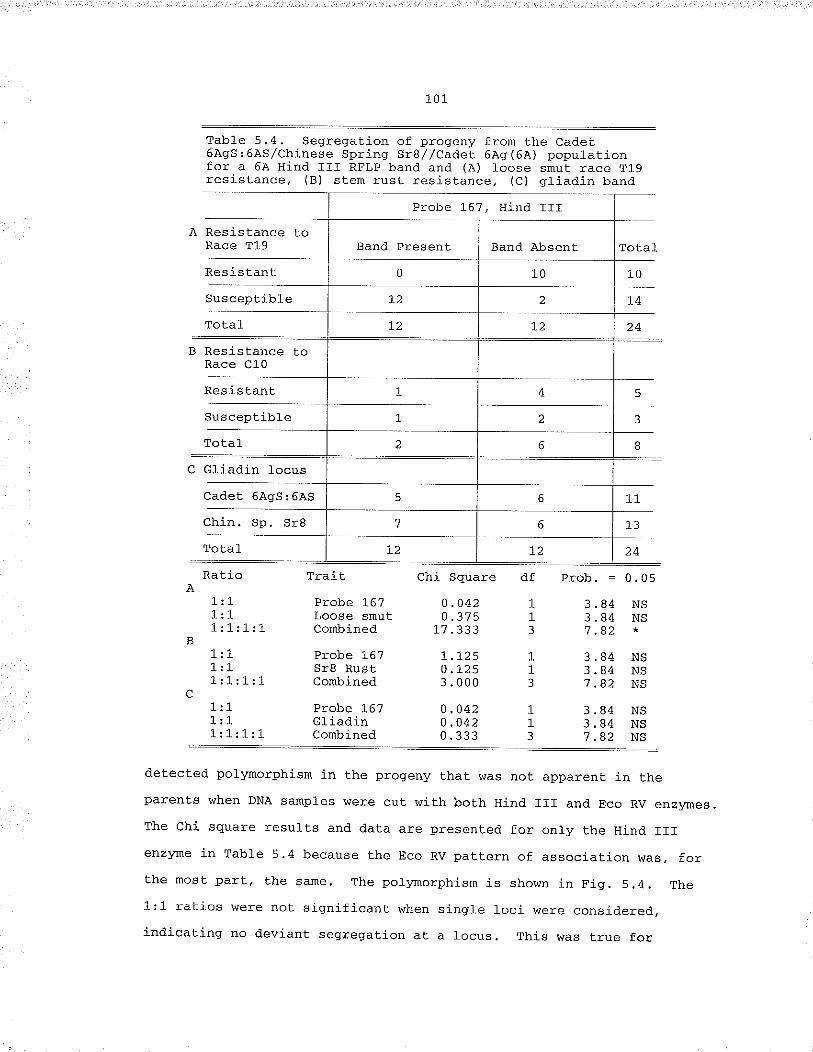
101
Table 5.4, Segregation of progeny from the Cadet6Aqs : 6As /chinese spring sr8//cadet. 6Às(6Ä) popularionfor a 6À Hind III RFLP band and (Ä) loose smut race Tl_9reEistance, (B) stem rust resistance, (C) gtiadin band
B Resistance toRace C10
A Resistance loRace T19
Res i s tant
Susceptible
Total
Resisgant
Susceptible
c GJ-aadfn locus
Cadet 6A95:6AS
chin. sp. sr8
11
Total
RatioA
1.11:1:l-:L
B1_:1
1:L:L:1c
l-:1
1:l-:L:1
Probe 167toose smutCombined
Probe 167Sr8 RustCoÍìlcined
Probe 167Gl iadinCombined
ChÍ Square
0.0420.375
17.333
1, .1250 .I253.000
0.0420.0420.333
Prob. =
3.843.847,82
3.843.847 .82
3.843.841.42
df 0.05
NSNS
NSNSNS
NSNSNS
detect.ed polyrnorphism in the progeny thal was not apparene in theparents when DNÀ samples rvere cut with both Hind III and Eco RV enzl¡mes.
The Chi square rêsults and data are presented for onfy the Hind IIIenzl¡me in Table 5.4 because the Eco RV pattern of association was, forthe most part, the same, The polt¡morphism is show¡ .in Fiq. 5,4. The
1:1 ratios were not significant when single loci were considered,indicat.ing no deviant segxegation at a locus. This was true for
Probe 167, Hind III
Band Prês ent Band Àbsen!

Fig. 5,4, DNA of homozygous recombinant lines for the 6AS chromosome armcut with Hind rrr and probed wich probe 167 where lane and a."".ipiiårr-ãiline are as followsi 1-7, loose smut suscepeible lines ,iti, e.ã fu-;;;ã;8-12, loose smut resistant lines without 6.6 kb band; f:, Cirinese SprinóSrSi 14, Cadet 6A9(6À) r 15, Cadet 6Àgs¡6À5,

t! !2 13 t4 15
<-6 . 6kb

103
segregation involving the RFLP (Hind III and Eco Rv generated) with thest.em rust, qliadin, and loose smut 1oci. À significant Chi square was
produced for the combined segregation of the loose smut locus and lheRFLP generated by the Hind III enzl¡me, (as was the case \eith the loosesmut locus and two polymorphisms generaled by Eco RV) .
DISCUSSION
The cross between a norrnal wheat and an alien translocation line inwhich lhe long arm is replaced will a11ow pairing only of the wheat
short arms. Traits in the short arm only are free to recornleine,
allowing linkage analysis, Because a test.cross was done with the aliensubst.itucion line, homozygous recombinant lines were produced for the6À5 chromosome arm. The process of genèxating the homozygous
recombinant lines is similar to the process of developing reco¡ïbinantsubstitution lines described by ,Jones ee al , (1991). Homozygous
reconìbinant fines a110w the lines to be propagated indefinitely withoutdi-sturbinq, due to crossing over, the linkage rel_ationship of genes inthe short arm of 6A.
The most reliable resul-ts from lhis study were those for loose smut
reaceion to race T19, and gliadin evaluation for which the popuLationsize v¡as the largest. These data indicated Chat the TL9 resistance ofchromosome 6Â was not linked to the gliadin locus in the same
chromosome. The rust data were the least reliable because of Ehe smallsample size, and because of the poor difference in reactj-on betwèen
resistant and suscept.ible tlt)ês. There was a preponderance of parentaltræes for both the rust resistance and gliadin combination and the rusLresistance and loose smut resistance combination. Detection of a \,reak
linkaqe between the rust resístance locus and boLh the gliadin and theloose smut resistance loci may be possible. With a larger sample size,precision of the linkage estirnat.e could be improved. Better conditionsfor expression of the srg gene, or the use of a different race which may

104
better detect. the Sr8 gene, could also improve the retiability of thislinkage analysis.
Àlthough a nurnber of studies have described linkage to seed proteinloci (McIntosh (1988)i .lones et aI . (1990); Czarnecki and Luko\,¡ (Lgg2)\,only a few studies have shown linkage between RFLps and disease or pestresistance loci in wheat (Chao et al . 1989; Mena et aL. !992¡ Conner etaI . 1993). Only a limited nunìber of markers are available at this timefor such studies in tùheat, However, in the present study, an
associat.ion was found between an RFLP and tbe foose smuh resistancegene, although furthex testing is required to determine if this is a
chance occurrence or if it. is truly linked, and to answer the questionwhy the poll¡norphisn did not show up in the parents. À linkage valuecannot be assigned because the parents do not show the poll¡moïphism.
However, if the most prevalent tl4)es are assumed to be the parentaltlq)es, the recornlcination frequency is eight percent, A reconbinationfrequency of eight percent would make thj_s RFLP a useful marker. onê
possibility for the lack of po]_ymorphism in the parents is that only one
plant of each parent was used for DNA extraction and a geneticallyaberrant plan! may have been chosen, In which case in future studiesmore parental plants should be used as a DNA source,
Because the long arm of Cadet is replaced by the 6Ã,9 short arm, Che
Cadet 6.Ags : 6As/chinese Sprinq Sr8//cadet 6Aq(6À) population will providethe opportunity in the future to determine the recornbination frequencyof wheat short arrn traits with Ehe centromere, ?herefore, any maïkeridentified for the long arm of Chinese Spring SrB can be used as a
cenlromere marker, because lhe long arrn wilJ- remain unchanged throughoutmeiosis. Because appropriate poltmorphism for the group 6 longchromosomè arms could not be found from Lhe probes used in this study,possible linkage of the loose smut resistance to the centroneïe courdnot be tested. In addition lo RFLP markers, a nuÍìber of isozymes may
act. as centromere markers in this populaeion, or as markers for the

L05
loose smut resistance gene. Isoz]¡mes \Àrhich have beèn associated withthe 6Ä chromosome (Hart et at. 1980; Mclntosh 1988) include esterase(,faaska 1980), aminopeptidase (Haxt and tangston 19?7), aconitase(Chenicek and Hart i-987), qlutamic oxaloacetic transaminasê (Harf 1975),
alpha arnylase (Nishikavra and Nobuhara 19?1; cale et. al . 1993), and
dipeptidase, (coldenberq 1986).
In conclusion, the qliadin locus is not a suit.abl_e marker for race T19
loose smut resÍst.ance located in the 6A chromosome, but an RFLP ispromising,

CHAPTER. 6
The Genetics of Loose Smut Resistance in Wheat
ABSTRACT: rnformaeion on the number and location of loose smut
resistance genes xesiding in various wheat. cytogenet.ic stocks, linesused as loose smut differen¿iaIs, and cultivars cofiìnonly used inbreeding programs can assist in their further genetic analysis and use
in resistance breeding. À nuÍìlcer of sources of resistance, and
cytogenetic stocks were inoculat.ed with up to eight races of U. tritici(T2, T6, T8, T10, T15, T19, T31, and T39). À survey of a number ofsmal1 segregating populations from crosses designed to locate loose smut
resistance to chromosomes allo\yed preliminary information on the nunbeï
of genes segregating, because of inocufation of each progeny with a
series of races. Some lines demonstrated mult.i-gene segregation forloose smut resistance, Both donínant and recessive genes wexe found. À
gene for resistance to race T15 in the cultivar H44 was located inchromosome 6À by segregation analysis, and evidence \qas found that a
gene resistant to race T10 was located in chromosome 28 in the cultivarChinese Spring. The location of the T10 resiseance \Â,as found by
inoculation of Chinese Spring-Thatcher substitubion 1ines. preliminary
informaeion about the number of genes can be obtained by the inoculationof each meÍìlcer of srnall populations with a nunber of races. fnformatj-onobtained about Ehe location of loose smut resistance genes from thej.noculation of cytogenegic seocks can help to focus future geneticresearch,

1,07
INTRODUCTION
Many sources of loose smut resistance in wheat have been identified,several of which display complete resistance to existing races (Anderson
1961t Nielsen 1983), Mulliple genes seem to contribute to resistance inat least some of the U. Èrjtjcj race differential lines (Chapter 4 ofthis thesis; Tingey and Tolman 1934). Fe\ù cytogenetic stocks have been
evaluated for resistance to loose smut (Mclntosh i.9gg) , with the
exception of the Chinese Spring-Hope and Chinese Spring-Thatcher inter-variet.al substitution lines (Dhitaphichit et a1 . 1989), Inoculation ofsuch cytogenetic stocks can immediately yield useful information such as
the influence of Hope and Thatcher chromosomes 7À and 7B respectively on
dramatically reducing the incidence of infection by race T6
(Dhitaphichif et al , 1989). Not. onl-y witl breedinq for resistance toloose smut benefit fron the direct knowledge of nurnber and conbinalionsof genes involved in resistance, but benefits will be seen through thebetter characteri zation of races and the easier devetopment of genetic
narkers (Allard 1960, Fleischmann and Baker 1971, Morris 198?). The
assessment of cytogenetic stocks for loose smut resistance will a1lovJ
for eheir systematic use, as well as occasionally providing informationon chromosome location should the stocks bxj-ng together the rightconÙ3inati-on of genetic factors, such as a susceptible chromosome
subscituted for a resistant. chromosome (ChapCer 4 this thesis,Dhitaphichit et a1 . 1989), The determination of the genetic makeup ofthe lines used to dífferentiaee races will contribute to the development
of single-gene differentiaf lines, and ¿o a set of lines which can be
used as a reference set to which all other genes can be compared
systematically (Äl]ard 1960, Fleischmann and eaker 19?1). Characterizedgenes in resistant lines used in breeding can be compared to previouslyidentified qrenes, so that new genes or alfeles Õf exj.sting genes can be
identified (Dyck and Kerber 1995) . Knowledge of the chromosomal

108
location of a g,ene helps in the gene identification process (Bariana and
McTntosh L993). Knowledge of the nurnber of genes and dominance
relationship of alleles helps in locating genes to chronosones, and
knowledge of the chromosomal location of genes helps in the development
of markers to those genes (Morris 1987),
The purpose of this chapt.er is to reporl preliminary informalion on
Che nuÍùcer and nature of genes found ín a nuÍìlcer of diffêrengial and
resistant st.ocks. Informalion on bhe reaction of cytogenetíc stocks toraces of loose smut is also provided, so that genes can be more
efficiently located to chromosomes. preliminary Cesting of cytogeneticstocks and lines of po¿ential value in breeding or genetic studies was
done to a11ow an understanding of segregation when these lines were
int.ercrossed, and to confirm findínge reported by others such as
Anderson (1961) and Nielsen (1983 and 1987a) .
The differential lines of Nielsen (1987a) and three highly resistantCanadian hexaploid wheats (clenlea, H44 and Hy3?7) were used as parenis
in crosses in the present study. ?entat.ive evidence for a resistancegene in chromosorne 18 from Hy3?7 L'as obtained from lhe backcross
development of this cultivar (Howes, personal communication) . Hy37? isa backcross line possessing a pxotein band that occurred in the loose
smut resistant donor parent, but not the recurrent loose smut
susceptible parent. The protein was know¡ to be coded by chromosome LB.
The recurrent parent a11ele was expected to be transferred to Hy3?7
after fivè backcrosses, but was not, In the present study a cross was
made with HY377 onto the prelude rnonosomic 18 (M1B) cytogenetic stock t.o
try bo confirm the location of a LB loose smut resistance gene, The
cultivaxs Glenlea and H44 weïe evaluated because they possess complete
resistance to all races in the Nielsen collection (Nielsen 1983) and areused as a source of resistance in breeding programs.

109
MATERIAIJS A}ID METHODS
F2 Populalions were established using Rescue 6À9(6À) , Rescue 6ÀS(68) ,
Rescue 6A9(6D) , and cadet 6AS(6À) as the female parents in selectedcrossing combinaeions with the cultivars c1enlea, H44, Kota, Red Bobs,
TÐ2, TD3, TD6, TD9, and TD16. Testcross populations \,¡ere estabtishedusing Roblin nullisomic LB (N1B) as the female in crosses with Kota,
Canthatch, TD2, and TÐ18, wifh the resulting single cross F1 pLants
crossed rvith the susceptible prelude. prelude MlB was used as thefemale in a cross with Hy37? and the subsequent monosomic Fr was crossed
with Prelude. Segregat.ion of lhe 18 chromosome was evaluated by the
same methods as reported in chapter 3 using monoclonal anlibody p24B as
a chromosome marker, and segregation of the 6À chromosome was evaluatedusing the sane methods reported in Chapter 4 using monoclonal antibod.y
230/9 as a chrornosome marker. polyacrylarnide ge1 electrophoresis was
used Lo track protein bands as chromosone markèrs for the populations inwhich the 6B and 6D chromosomes were being studied. Electrophoreticmethods and staining of gels are described in Chapter 4. progeny ofthese populations were inoculated with one or more of races ,12, î6, Ig,T10, T15, T19, T31, and T39, The procedures usêd for inoculation were
the same as those described in Chapter 4.
Inter-varietal subsi:ieution lines were gro\!,n as described for lines in
.Chapter 4, The Chinese Spring-Thatcher fines were those developed by ,f,Kuspira (cenetics Department, University of ,A]berta) , and maintained and
provided by Dr, P.t. Dyck (Àgriculture Canada Research station,winnipeg, Manitoba) . The cadet-Rescue lines were those developed byR.I Larson, and maintained and provided by Dr, E,Ð.p, Whelan
(Agriculture Canada Research Station, Lethbrj-dge, Àlberta) . The Chinese
Spring - That.chèr inter-varietal substitution lines were evaluated withraces T6, T10, T15, T19, T31, and T39 !o identify chromosomes affectingresistance t.o t.hesê races. The inter-varietal substitution lines ofCadet.-Rescue were inoculated with races TB/ T15, and T19 to identify

110
chromosomes affecting resistance to these races.
RESUIJTS
The results of j-nocufations of progèny of segregating populations willbe provided firsE, followed by the results of inoculat.ions of ¿he
substitution 1ines. The proportion of loose smut in the parental lines,from the inoculation with up to eight. races, are listed in .A,ppendix 1
(resul-ts of inocufat.ions of other lj-nes not discussed in this chapterare aLso found in Àppendix l-) , The j-nterpretation of Che segregation ofthe progeny of populations derived from these cyt.ogenetic s¿ocks and
Lines is based on the loose smut reactions of these lines.Figure 6.1 (the values for proportion of smutled plants on the
horizonlal axis represent Ehe mid-point of the int.erval for which the
endpoints are marked by ticks, for this and subsequent figures)demonstrates that those progeny of the Rescue 6AS(6A),/H44 populationmissing the 6À chromosome were, in general, more susceptible to loose
smut. race Tl-5 than those progeny possessing the 6À chromosome. The mos!
susceptible plants of this population were characterized as weak and
much less fertile than plants in the resisÈant group. One planc which
derived from a seed with a low anL ibody - bindinq had a 1ow loose smue
infection, and was a vigorous, fert.ilê plant. like Lhe plants derived.
from high antibody - binding seed.
The results obtained from the remaining populations, in which
chromosomes LB, 6À, 6B and 6D segregated, did not show the chromosomal
location of any further genes. These poputations did provide some
indicalion of the presence of a11el_es dominant for loose smut
resistance, and, for lhose populations inoculated with more than one
race, an indication of the nunìlcer of genes was also obtained. The
presence of dominant genes was the most. likely explanation for lheresults of segregation of populations involving: clenlea tÕ aL1 raÕes
tesged (Fig. 6.2, 6.9Dr; Kota and CadeÈ 6A9(6À) to T15 (Fig, 6,3A) ; TD6
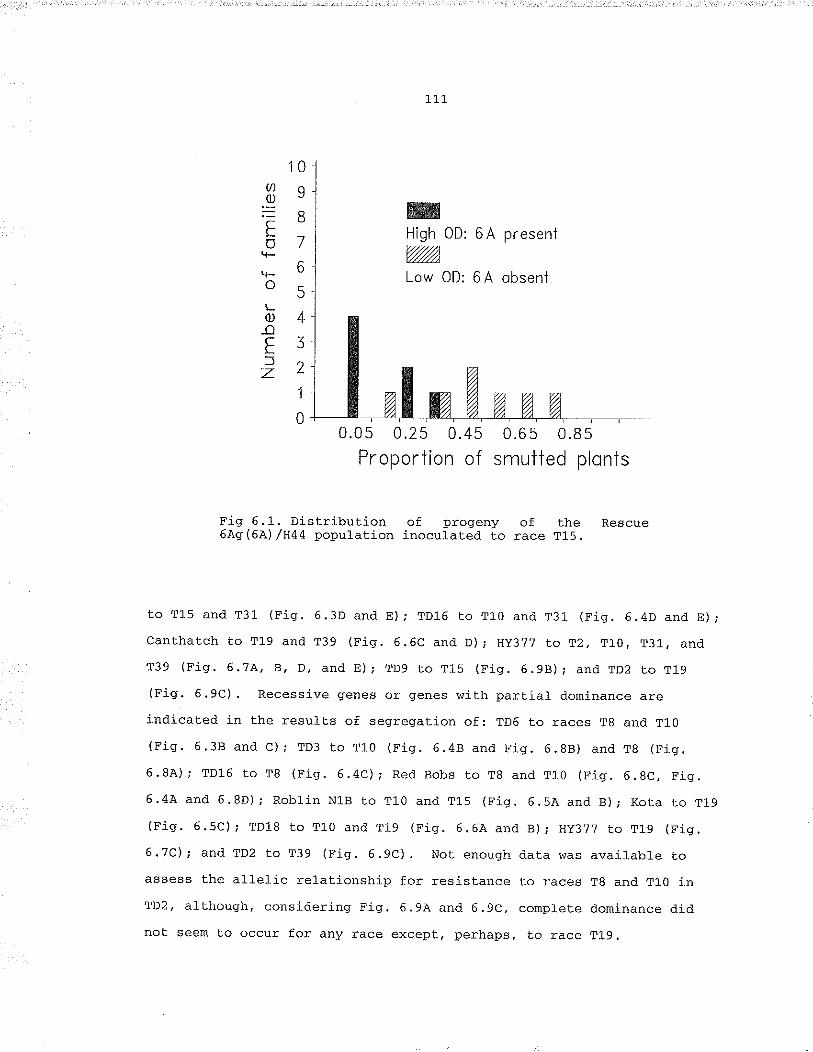
(f)
.9;=Eo
t{-
q-oL0)
-oEfz
10
IIl6
5
4
3
2
1
0
IHigh 0D: 6A preseni
ØLow 0D: 6A obseni
0.05 0.25 0.45
Proportion of0.65 0.85
smutted plonts
Fiq 6.1, Ðistribution of progeny6AS (6À) /H44 population inoculated to
Rescue
Lo T15 and T31 (Fig, 6.3D and E); TDL6 to T1O and T31 (Fiq. 6.4D and E) i
Canthatch to T19 and T39 (Fiq. 6.6C and D) r Hy3?7 Lo T2, T1O, T31, and
T39 (Fig, 6.'lA, B, D, and E) , TD9 to T15 (Fig. 6,98) ; and TD2 to T19
(Fig. 6.9C) , Recessive genes or genes with partial dominance areindicated in the results of segregatíon of: TD6 to races T8 and T1O
(Fiq. 6,38 and C) ; TD3 to T10 (Fiq. 6.48 and Fig. 6.BB) and Tg (Fiq.
6.84); TD16 to Tg (Fig. 6.4C) ; Red Bobs ro Tg and T1O (Fig. 6.8c, Fig.6.4À and 6.8Ð) ; Roblin N1B to T1O and Tt5 (Fiq. 6.5À and B) ; KoEa t.o Tl-9
(Fig. 6.5C) ; TD18 Eo T1O and T19 (Fig. 6.6À and B) i Hy3?7 ro T19 (Fiq.
6,7C); and 9D2 to T39 (Fiq. 6.9C) . Not enough data was available t'o
assess lhe alle1ic relationship for resistance to races Tg and T10 inTÐ2, although, considering Fig. 6,9A and 6.9C, complete dominance didnot. seem !o occur for any race except, perhaps, to race T19,
of therace T15.

Àio
Eo67q_6o.
J
o4¡l^Z¿
1
0
EIJJ 1O
øao-5çc-al.,. 6
os04¡E3f,"zt
I
0
Drot¡0
t-0/\-6os04-0E3l"z¿
j
0
t- ioOQo-E¿tol,,- 6
0.o4l)EJf^
I
0
trr l0(¿ôEqc"L-ol
6
0È04
_o
Erf,^
1
0
0.05 0.25 0.15 0.65 0,85
Proporfion of smulled plonls
0.05 0.25 0.45 0.65 0.85
Proporlion of smutled plonts
0.05 0.25 0.45 0.65 0,85
Proporlion of smulled plonls
Fiq. 6,2, Loose smut react.ion of6Â9 (6À) /clen1ea inoculated withT31 , (F) T39 .
1L2
0.05 0.25 0.45 0,65 0.85
Proportion of smutled plonls
0,05 0,2s 0.15 0.65 0.85
Proportion of smuiled plonts
0.05 0.25 0.45 0.65 0.85
Proportion of smulled plonls
families of the populat.ion Rescue(Ã) 18, (B) r10, (c) r15, (D) T19,
v)0)
'¡ao
o
0)
-oEfz
(E)

B10ooo"E¡L-ol
6
o.J
o1¡f"z¿
1
0
IHigh 0D: 6A present
ruLow 0D: 6A cbsenl
0.05 0,25 0.45 0.65 0.85
Proporlion of smulted plonts
0.05 0,25 0.45 0.65 0.85
Proporlion of smutled plonts
Fig. 6.3. Loose smut reaction of6À9 (6Ä) /Kota inoculated with (À)T8, (C) 110, (D) rLs, (E) 131.
113
0,05 0.25 0.45 0.65 0.85
Proporfion of smulled plonts
0,05 0.25 0.15 0,65 0,85
Proporiion of smulled plonts
0,05 0.25 0.45 0,65 0,8s
Proportion of smulted plonts
families of thê population CadetT15, and Rescue 6À9(6À,) /TD6 rvith
ciooô0l"Ect_ol,.- 6
0.
0)4¡E3l"z¿
I
0
E10Øõ;8t_ol.,- 6
o.J
04¡E3l"zt
I
0
DroEp67..6o!
0d¡E3f"zt
f
0

L1-4
Aio
=¡L-o/q_6o.
J
04¡EJf"
I
0
u 10
øo0"
L-o/e-6o.o4-oE3l"zz
I
0
Broø0EqcEI
Â
o-¡!04-oE3l^z¿
1
0
EtooooJEcL0/,'- 6
o. ¡o.r 4-0E3f,"zL
I
0
Dio
=rc-ol6
o"J
Lo4¡E3f"zt
f
0
IHigh 0D: 6A present
%Low 0D: 6A obsent
0.05 0.25 0,45 0.65 0.85
Proporlion of smutled plonls
0.05 0.25 0.45 0.65 0.85
Proportion of smulled plonls
0,05 0.25 0.45 0.65 0.85
Proporlion of smulted plonts
0,05 0.25 0.45 0.65 0.85
Proportion of smulled plonls
0.05 0.25 0.15 0.6s 0.85
Proportion of smulled plonts
Fiq. 6,4. Loose smut reaction of families of Che population Cadet6Às(64) crossed wirh (A) Red Bobs and inoculaLed;ilh T1O, (B) TD3 andT10 and Rescue 6A9(6A) crossed and inoculated with (C) TD16 and Tg, (D)TÐ16 and T10, (E) TDI-6 and T31.

r'1 B disomic
m1B monosomic
a
Écc0
0
La.)
_0
Efz
crooooJ"Eat-o7\-6o.
J
oi4¡E3l"zt
1
0
l0
I8
1
6
5
4
3
2
1
0
EI¡J 10
ø00"Ecc"L-ol
6
JLa4
_0
tJf"zL
1
00.05 0.25 0.45 0.65 0.85
Proportion of smulled plonls
0.05 0,25 0.45 0,65 0.85
Proporlion of smutled plonts
0.05 0,25 0.45 0,65 0.85
Proporlion of smufled plontsFig. 6.5. toose smut reaction of families from populations of Roblin N1Bcrossed onto a Kota and testcrossed rr'i t'h prelude and inoculated withraces (À) T10, (B) T15, (C) Tt9.

A10aõ0"
c_ol6
0.c
0)4¡t.rf4z1
1
0
a
ÉEo
oLOJ¡Efz
c
T1B disomic
%1B monosomic
f00
I7
6
5
4
3
2
f
0
0,05 0.25 0.45 0.65 0.85
Proporfion of smutted plonts
0.05 0.25 0.45 0,65 0,85
Proporlion of smulted plonts
0.05 0.25 0.45 0,65 0.85
Proporlion of smulled plonls
0.05 0.25 0.45 0,6s 0.85
Proporlion of smulled plonls
ttÐ 10
OQ()-Eat_Ol
6
ô.J
!04_0
tJf"z¿
1
0
Dio00()"Ecc"L.El.,. 6
o.:)
Io4¡I ..)
f"z¿1
0
Fiq. 6.6. toose smut reaction of farnilies of the populations on which aloose smul resistant line was crossed onto Roblin t¡1g and test crossedwith prelude, cultivar and race are as follows (a) t¡18 à"a tio,-f¡l --TD18 and T19, (C) Canthatch and T19, (D) Canehat.ch and T39.

1,17
A10q¡ a0J'E¡t_ol\.6os04¡EJl"z¿
I
0
Cjo
E¡t-olr-6o5o4-0E55"z¿
1
0
Bio
;sc_al,,_ 6
o.J
o4!Ê3f,"z¿
I
0
Eroo00l"Enc-olt-60¡
o4-0
E3tr"z¿1
0
Ð10øoE¡c"ol
6
0.Lo4-0E3f"z¿
1
0
T1 B disomic
m1B monosomic
0.05 0.25 0,45 0.65 0.85
Proporlion of smutted plonts
0.05 0.25 0.15 0.65 0.85
Proporlion of smulled plonls
0,05 0,25 0.45 0.65 0,85
Proporiion of smutled plonls
0,05 0,25 0.45 0.65 0.85
Proporlion of smulted plonls
0,05 0,25 0.45 0.65 0.85
Proporlion of smulted plonls
Fiq. 6.7, Loose smut reaction of families from the population wj-th \,¡hichHY377 was crossed onto prelude MLB and test crossed with prelude,inoculated wir.h (À) T2, (B) T10, (c) T19, (D) T3L (E) 139.

Broo00"
=8c-õl6
ô.J
La4_0
E3l4zt
1
0
Arooqú)-5rc-ol
6
o.:l
04¡l-
f
0
D10oo0'Iqc-ol.,6o-
J
o4-oE3l"z¿
1
0
t- t0o00)"EaFo7*6o-¡Lo4¡E3f"z¿
1
0
rI6B pr esenl
ru6B obsenl
0.05 0.25 0.45 0,6s 0.85
Proporlion of smulled plonts
0.05 0,25 0.45 0.65 0,85
Proporfion of smutled plonls
0.05 0.25 0.45 0.65 0.85
Proportion of smulted plonts
0,05 0.25 0.45 0.65 0.85
Proportion of smulled plonts
Fiq. 6.8, Loose smut reaction of families from Ehe populations in whichRescue 6A9(68) was crossed and inoculated with (Ã) f¡3 and T8, (B) TD3and TLo, (C) Red Bobs and T8, (D) Red Bobs and T10.

Race
nT8
mT10
NT15ffiT19
ffiT3I
Brooq0-Iqc_Ol
6
o.Lo4
_0
E3l"z¿
1
0
D10oo0l-Eçc"o7
6
o"J
04¡E3f"z¿
1
0
c10oo0'c-L-ol
A
o-J
o4¡c1l"z¿
I
0
AtoOQ0J-Eqr_Ol
6
o"J
LoJ4
_0
EJl4zL
1
00.0s 0.25 0.4s 0.65 0.85
Proporlion of smulled plonts
0.05 0.25 0.45 0,65 0,85
Proporlion of smulled plonts
0.0s 0,25 0.45 0,6s 0.85
Proportion of smutted plonts
0,05 0.25 0.45 0.65 0.85
Proporlion of smutled plonfs
Fiq. 6,9. Loose smut reaction to various families from the populationsof (À) Roblin NLB/TD2 / pretude, (B) Rescue 6As(6.¡\) /epg, (C) neicue6AS(68) /TÐ2, (D) Rescue 6Ãs (6D) /ctentea.

120
More than one gene seqregating was determined by the differentialsegregation of the same pxogeny to different races, Some of thepopulations tested to rûore than one race indicated more lhan one
segxegating gene. The inoculation of the Rescue 6Àq(6A) /c1en1eapopulation with three different races (T15, T19, and T39) resulted in a
different susceptible segregate Eo each race (Fig. 6,2C, D and F),There was a preponderance of resistant bl4)es to aI1 ïaces, and no
susceptible segregales occurred upon inoculation with races Tg, T10, and
T31 (Fig. 6,2A, B and E) .
The two l-ines of lhe Rescue 6Àq(6À) /TÐ6 population which were most
susceptible to race T31 were also the most susceptiblè to race Tl-5,
indicat.ing that' the gene or genes in TD6 resistant to race T3L were the
same genes conferring resistance bo T15 (Fig. 6.3D and E) , Comparing
Fig. 6.38 and C to Fiq. 6.3D and E shows that more susceptiblesegregates occurred when the progeny were inoculated with races Tg orT10 than when inoculated with T15 or T31, Most families susceptible torace T8 were also suscepLible to race TLo, Only one family differed inthis respect.
fn the Cadet 6Àq(6À) ,/Kota poputation (Fiq, 6.3A), both parents were
resislant to race T15. One susceptible segregate was produced,
indicating that Cadet 6ÀS(6À) and Kot.a each possess different singledominant gienes for resistance t.o race T15. When the same progeny werej.noculated with races T15 and T19, thê progeny most susceptible to T15
h¡ere not the same as the progeny most susceptible to T19 (Chapter 4) .
No highly susceptible segregates to race T10 were observed arnong the17 families from the poputation Cadet 6À9(6A)/Red Bobs (Fiq. 6,4A) .
Like the progeny of the Cadet 6Ag(6A) /Red Bobs poputabion, the resÍstantsegregates from the population Cadet 6Àq(6Â) /TD3 were onLy moderatel-y so
(Fiq.6.48) . The progeny of the Rescue 6Aq(6À) /TDL6 population most
susceptible to TB (Fiq. 6.4C) were the same progeny whj.ch were most
suscept.ibfe to T10 (Fig. 6,4D) from the cross of . The progeny

121
susceptibte to races T8 and T10 $¡ere resistant to T31 because no
segregates susceptible to race T31 werè identified (Fig 6.4 E) .
In the testcross populat.ion of Roblin N1B/Canthatch/ /prelude resistantprogeny predominated when inoculated with T39 (Fig, 6,6D) , bu¿ not. when
inoculat.ed with T19 (¡'ig. 6.6c). Like canthaLch, Hy3?7 $,as evaluated ina eesbcross, and most' progeny clustered toward the resistant end of the
spectrum when inoculated with T2 (Fig. 6.7À) and T39 (FiS. 6.78), and tothe susceptible end when inoculated with T19 (Fiq, 6,7C), The two
progeny most susceptible to race T31 (Fiq. 6,7D) were not among theprogeny most susceptible to other races. In a t.est.cross involving Kota,
a susceptible phenotlt)e to race T19 predominated in most of the progeny
(Fiq. 6.5C). Kota and Roblin are resistant to T15, whereas only Roblinis resistant to TLo. The progeny of the cross Roblin NlB/Kota// prelude
were susceptible to race T10 (Fig, 6.5A) and T15 (Fig. 6,58) , as
menlioned earlier, indicating segregation of a recessive gene or genes.
Susceptible and moderately susceptible segregates were common among
the progeny of TD3 and Red Bobs \,¿hen inocutated with either TB or T1O
(Fiq, 6.8). The segregates most susceptible to race TB are the same
progeny susceptible to T10 when TD3 was the parent (Fiq. 6.9À and B) ,
and similarly for Red Bobs (6.8C and D) .
Results of inoculation of Cadec-Rescue and Chinese Spring-Thatcher
inter-variet.al substitution lines are highlighted in the remainder ofthis sect.ion. Thatcher was found to be susceptible to U. Êritjcj races
T10, T31, T39, wiEh Ínconplete resistance to race î19, and certainChinese Spring sources were susceptible to T6 and T15 (Table A.4) .
Cadèt is resistant to races T8, TL5, and T19 (Tab1e À.1), and Rescue issusceptible to races TL5 and T19, with incomplet.e resistance to race Tg
(Table À.2). Table 6.1 1i-sts the loose smut reachion of the series ofcadet varietal substitution 1ines, in which each Rescue chromosome was
substituted separately into CadeE, replacing its Cadet homologous
chromosome counterpart. The data were incomplete for race Tg, but the
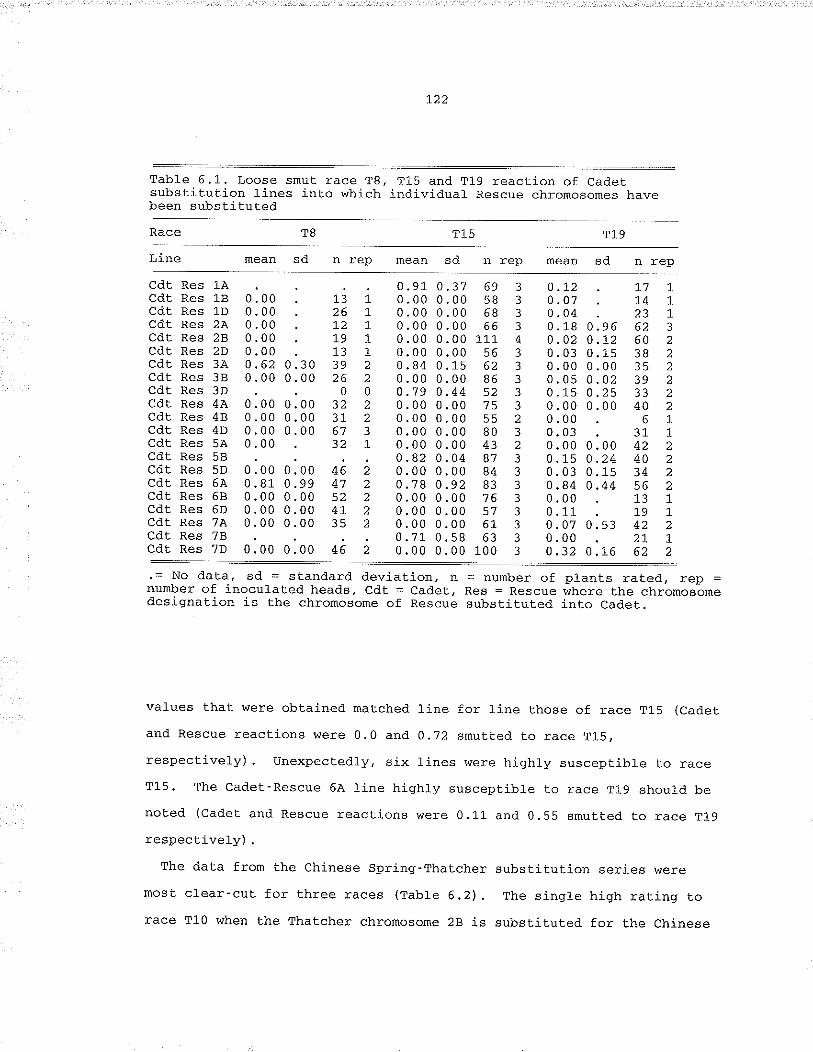
Table 6.1. Loose smut race T8, T15 and T19 rèaction of CadeLsubstitution lines into which individual Rescue chromosomes havebeen substituted
T15 T19
Line mean sd mean sdn rep n rep mean sd n rep
Cdt Res 1ÀCdt Res 18Cdt Res 1DCdt Ree 2ACd¿ Res 2BCdt Res 2DCdt Res 3ACdt Res 38CdC Res 3DCdt. Res 4ACdt Res 48Cdt Res 4DCdt Res 5ÃCdt Res 5BCdt Res 5DCdt Res 6Â,CdC Res 68Cdt Res 6DCdt Res 7ACdt Res 7BCdt Res 7D
0.000.000 ,000.000.000,62 0.300,00 0.00
0.00 0.000.00 0.000.00 0.000.00
o. oo o. oo 460.81 0,99 470.00 0.00 520.00 0.00 4L0,00 0.00 35
0.00 0.00 46
0,91 0.37 690.00 0.00 580.00 0.00 680.00 0.00 660.00 0.00 1r,10.00 0.00 560.84 0.15 620.00 0.00 860.79 0 .44 520,00 0.00 '150.00 0.00 550.00 0.00 800.00 0.00 430.82 0,04 870.00 0.00 84o.'78 0,92 830.00 0.00 760,00 0.00 570,00 0.00 6t0.71 0.58 630.00 0.00 100
0.!2 770.0? 140.04 230.18 0.96 620.02 0.12 600.03 0.15 380.00 0.00 350.05 0,02 390.1s 0.25 330.00 0.00 400.00 60.03 3l-0.00 0.00 420.15 0.24 400.03 0.15 340.84 0 .44 s60,00 130.11 L90.07 0.53 420.00 2t0.32 0,16 62
1å
L913
260
3167
L
L
1,
322222
2202231
à
.= No data, sd = standard deviation, n = nu¡nber of plants xated, rep =nurnber of inocufaeed heads, Cdt = Cadet, Res = Rescue where the chromosõmedesignation is the chromosorne of Rescue substituted into cadeÈ.
values that were obtained matched li_ne for line bhose of race T15 (Cadet
and Rescue reactions were 0.0 a d O.j2 smutCed to race T15,
respectively) . UnexpecÈ.edly, six l-ines were highly susceptible to raceT15. The Cadet-Rescue 6Â line highly susceptible to race TL9 should be
not.ed (CadeL and Rescue reactions were 0.11 and 0.55 smutted t.o race T19
respectivel-y).
The data from the chinese spring-Thatcher substitution series were
nos! clear-cut for three races (Table 6,2), The single high rating torace Tl-0 when the Thatcher chromosome 2B is substituted for the Chinese

1,23
Table 6.2. Loose smut race T6, T10, 11,5, T1,9, T31 and T39reaction of Chinese Spring substiCution lines j.nto \.¡hichindívidual Thatcher chromosomes have been substitutedRace T10
Line mean mean
T15
CS Tc 1ÀCs ?c 18CS Tc 1DCS Tc 2ACS Tc 2BCS Tc 2ÐCS Tc 3A
CS Tc 3DCS Tc 4ÀCS Tc 4BCS Tc 4DCS Tc 5ACS Tc 5BCS Tc 5DCS Tc 6.4CS Tc 68CS Tc 6DCS Tc 7ÀCS Tc 78CS Tc ?D
0.00 250.68 310.07 140.08 !20 .59 340.30 300 .67 240.00 r_s
o.23 300.54 260.69 290,20 600.88 25
0.33 330 .42 450.00 320.30 200.00 l-60. L7 29
0.00 s0.00 Lr0.00 60.00 200.75 200.00 L90.00 330,00 180.00 330,00 2L0.00 190.00 330.00 290.00 90.00 20,00 60.00 60.00 t40.00 110.00 70.00 26
0.67 300,66 320.62 130.76 330.88 250 .62 s00,60 25u. tz 250.73, 4L0.87 230,'76 290.7r 340.49 4L0.70 300 .67 210.86 420.74 190 .42 L90.37 l_90.00 180.'14 27
T31T19 T39
Line mean
CS Tc 1ACS Tc 18CS Tc 1DCS Tc 2ÀCS Tc 28CS Tc 2DCS Tc 34.CS Tc 38CS Tc 3DCS Tc 4ÀCS Tc 4BCS Tc 4DCS Tc 5ACS Tc 58CS Tc 5DCS Tc 64,CS Tc 68CS Tc 6DCS Tc 7ÀCS Tc 78CS Tc 7D
0.000.000.001,00
0.050,000.000.000.000.000.00
0.000.000.000.000.000.00
0.000.000.000.000.000.000,000.000.000.000.000.000.000.000.000.000.000.000.000.390.00
414
92
1',?
520262318
181,71316
9),
236
9394037t9
33
0.00 90.00 160.00 150.00 220.37 270.00 280.00 t40.00 I0.00 740.00 100.00 200.00 r_00.00 300.43 2r0.00 10.00 30.23 130.00 I0.00 270.49 720.00 20
.= No data, n = nunìlcer of plants CS = Chinêse Spring, Tc =Thatcher, where the chromosome designation is the chromosomeof Thatcber substituted into Chínese Spring.

L24
spring 2B, indicated that the chinese spring resistance to race T1O
resides in chromosome 28 (Chinese Spring and Thalcher reactìons Eo race
Tl-0 are 0.0 and 0.96 respectively) . Likewise, the only susceptiblèreacCíon to race T31 in the Chinesê Spring-Thatcher series was for thechromosome 7B line, which indicat.ed thaL resistance to this race residesin chromosome 7B (Chinese Spring and Thatcher rèactions to race T31 were
0.0 and 0.73 respectively) , The chinese Spring - Tha t.cher 7B line was theonly line fully resistane to race T15 (Chinese Spring and Thatcherreactions to race T15 were 0.61 and 0.0 respeccively). Limitations on
lhe number of heads available fox inoculation meant not all inoculationscould be accomplished wiÈh race T19. Four lines showed no incidencewhen inoculated reith T6, with one of these lines being Chinese Spring-Thatcher 78. Inoculation of the Chinese Spring-Thatcher substitutionlines with T39 pxoduced the unusual result of four suscept.ible. The
level of loose smut infection to T39 was moderale in each of the fôurlines (chinese spring and îhatcher reactions to race T39 axe 0,0 and.
0 .64 respect ively) ,
DTSCUSSION
The data der0onstrate the occurrence of gènes with different. levels ofdominance. Multiple qenes for resist.ance are indicated by inoculationresults from both the segregat.ing populations and bhe inter-varietalsubst.ítulion lines, Evidence for the number of genes acting in a
particular genotlt)e comes from the different reaction of the same
progeny to different races, Ribèiro (]-963) used two races to help sortout the nunber of resistance genes to loose smut, while Ward and Metzger(1970) , and Met.zger et al , (1979) used muttiple races to indicate thenumber of resj-stance genes segregating against conmon bunt. The
proporeion of resistant to susceptible progeny also gives an idea of thenunìber of genes involved (i{eyne and Hansing 1955) . À populationinoculated to different races can d.emonstrate different leve1s of

1,25
dominance to each of the races. This may indicate different. resj-stance
genes or modifying fact.ors (Samborski 1963). Cornbining these forms ofevidence from multiple populations, whlch agree in terms of the nunìber
of genes estimat.ed, strengthens the conclusions about the nunìlcer ofgenes in the resist.ant parent,
When most of the F, progeny of a segregatinq populabion are resistant,doninance is indicated (Heyne and Hansing 1955) , which was the case r{ithnost of the populations studied, Conversely, when most of the progeny
are suscept.ible and a parent is known to be resistant, then the gene orgenes are recessive (Gaskin and Schafer L962). lf the progeny clusterin the rnidrange region, then the resistance is partially dominant(Lamari and Bexnier 1991-) . Examples of boeh of these 1atter situationswere al-so seen. Considering Table 2.4 of Chapter 2, the appearance ofdominant, partially dominan!, and recessive a11eles is not surprising,
Àn argument can be made to support the hl¡pothesis that clenleapossesses at least three genes for resistance, The progeny from thepopulation Rescue 6À9(6Â.) ,/Glenlea susceptible to races T15, T19 and T39
differed, indicating Ehat each progeny had a resistance gene; however,
Èhe proporlion of resist.ant Co susceptible progeny either approached orv¡as a 15¡1 ratio, depending on the race, indicating two genes forresistance to any one race, for a total of three genes. Metzger and
coworkers (1979) used the same strategy to help determine the aunber ofcomnon bunt resist.ance genes involved in the line CI7O90, The lack ofprogeny susceptibLe to races T8, T10, and T31 further supports thehlæothesis of more than one gene for resistance in clenlea (Fiq, 6,2).That Glenlea would have three genes for resistance is not surprising,considering its background. One of the parents of clenlea is derivedfrorn lines known to possess high Ièvels of loose smut resistance,including Pernlcina, Thatcher, Redman, Regent, Marquis and H44 (Anderson
1961, Nielsên L983, Zeven and zeven-Hissink 19?6). Tingey and Tolman
(1"934) reported lhree genes for 1oose smu! resistance in Hope, a sib of

L26
H44 (zeven-Hissink 1976). The other parent of clenlea (CB1OO) is alsoderived from lines possessing loose smub resistance (Nielsen personal
coÍununication) . An important. irnplication of the finding that virulenceexists to the genes making up the rèsistance in clenlea, should it be
borne out with more detailed study, is that virulence exj-sts to an
important source of resistance used in breeding of Canadian prairiewheat. À11 that is required to overcome the clenlea resistance is a
recombj.nation of the existing virulence genes, ÀnoLher consideration isthat. when clenlea is used as a source of resistance, a large breedingpopulation will be required to recoveï all three genes, One segregate
moderately susceptible to race T39 was produced out of the five members
of the Rescue 6À9(6D)/clenlea population, indicat.ing as litt1e as one
gene, or providing evidence that a gene resides in chromosome 6D, The
population size of Rescue 6À9(6Ã,) was not big enough to determine
whelher or not any of lhe genes are in the 6A chromosome of clenlea.The population size would have to be big enough (abouL 40 progeny ineach chromosome class (Hanson 1959)) to disLinguish beCween a 3:1 ratioin the class missing the 6Â chromosorne, and a 1511 ratio in the class inwhich the 6A chromosome is present.
The simplest explanation of the segxegation data from the TD6-derivedpopulation is that two genes are segregating (Fiq, 6.38 to E) .
Resistance to races T8 and T1O is common because the susceptiblesegregates were, for the rnost part, the same for Lhe two races, and thegene appears to be partly dominant. Resistance to races TL5 and T31 isconmon becausê the susceptÍbfe segregate was the same for the two races,and the gene appears to be dominant. progeny segregating for a
partially dominant gene to one group of races, and for a domjnant. gene
to a second group of races, is an indícation of two genes. That a gene
resides in 6A in TD6 cannot be rulêd out, TD6 is the line pI692g2 and
was used by Níe1sen (198?a) in place of the late-maturing cultivar van
Hoek from oort.,s dífferential set (Nielsen 19g7a) ,

1,27
Both Kota and Cadet 6Àq(64) are resistant to T15. The segregation ofresis¿ance from Kota when crossed onto Cadet 6Âq(6À) would indicat.e thatthese two cultivars differ in two dominant genes for resistance t'o race
T15 (Fig, 6,3À) , The testcross of Robtin NlB/Kota with prelude does notcontradict. dominance to race T15 in Kota (Fiq, 6,58) . Segregation toT31 was also dominant (Fiq. 4,2D) , The testcross of Roblin
NlB/Kota/ / Prelude indicates that resistance in Kota to race T19 isrecessive. The difference in reaction of the progeny of the Cadet
6Aq(6A') /Kota to race T19 and T31 indicated two gene6 in l{ota (Chapter
4) , The one dominant and one recessive response to two different races
is a further indication of two genes for resiseance in Kota. Ribeiro(1963) reported that Kota possesses a single dominant gene to the racedesignated as C2, although in a study by Kilduff (1933) Kota was thesusceptible parent.. Kota was added to Oort,s differeneial set by
Chexewick (Nielsen 1987a).
Only one parlly dominant gene seemed Eo be segregating from Red Bobs
(Fig. 6.8c and D) . Kilduff (1933) used Red Bobs as a loose smut
resistant source in a study of the genetics of loose smut resistance,Àlthouqh he did not feel confident enough to make a conclusion about theinheritance of resist.ance in Red Bobs, his data indicate a singledominant gene. Red Bobs, like Kota, was added to Oort,s differentialset by Cherewick (Nielsen 1987a). TD3 displayed incomplete dominance toraces Tg and T10, and only one gene can be said t.o be segregating (Fig,
6,8.A' and B). TD3 is a cultivar derived from Flourence x Àurore, and isone of Oort,s original differential lines (Nielsen 1997a) , The
segregation pattern of TD16 is best explained by two genes, because ofthe lack of a susceptible Clæe from Ehe 10 progeny inoculated with raceT31, yet susceptible progeny segregated when inoculated with TB and T1O
(Fiq. 6,4C to E) . One gene would seem to confer resistance to both raceT8 and T10, TD16 is ehe ]ine Marroqui 5gg, and was added to thedifferential set by Nielsen (1997a) .

I28
All chromosome LB analyses were set up as tesCcrosses. No chromosome
association with resisEance was seeni however, there are some
in¡eresting points to note. The testcrÕss can give an indication of the
dominance of a11eles. Testcross F1 plants are all eíther heterozygous
resistant or honozygous susceptible, if resistance is dominant (nibeiro1963). Roblin N1B was obtained from a monosomic 18 planC from Roblin
breeder seed (Chapter 3). Roblin N1B has the same pLant tlæe and seed
protein profile as Roblin except for the nissing 18 protej-n bands in bhe
nullisomic when seed proteins were run using pÀcE, Roblin is resist.antto race T10 (Tab]e 'A,4), The resistance in Roblin N1B to race T1O iseilher recessive, or Roblin, the progenilor of Rob1in Nl_B, has lost a
gene fox resistance (Kota ìs susceptible to T10) (Fig, 6.5A).Indications are that T19 resistance in TDI-B recessive, like that in
Kota. TD18 is the line C.T. 439, and was added to the differential set.
by Nielsen (1987a) , Recessive resistance to T19 in TDlB and Kota would
support. a hlæothesis that. TD18 and Kot.a possess lhe same gene. As
indicated in Chapter 4, TD18 may have two oEher dominant genes forresistance. This is not surprising, considering that TDlg has bothThatcher and Regent in its background (zeven and zeven-Hissink 19?6) .
Two dominant genes were indicated to race T39 in canthatch (Fig, 6.6Ð)
because of a greater nunùcer of resistant to susceptible progeny in thelestcross, where a 3:1- segregation indicates two genes (Ribeiro 1963).
Neither the nunber of genes involved, nor the gene action for resistanceto race TL9 is indicared (Fiq, 6,6C). À study by Campbell (1948)
indicated at least one gene for loose smut resist.ance in Thatcber, and
from his data the gene appeared to be dorninane. Canthatch is a
backcross derivative of Thatcher (Zeven and Zeven-Hissink l-976).
Two genes can explain the segregation in crosses involving Hy377 (Fjg.
6.71 . Resistance to races T2, T31,, and T39 shows up as dominant,
whereas resistance to T1O and T19 seems to bè only partly dominant,
although dominance reversal cannot be ruled out. It. is reasonabLe to

r29
assume Chat lwo genes confer resistance to races T2 and T39, because offer,¡ susceptible segregates t.o these tv¿o races. Furthermore, T31-
susceptible progeny are resistant when inoculated with othèr races.
There appears to be a single dominan¿ gene segregating to racê T15 inTD9 (Fig. 6,98) . TD9 is the cultivar Kearney, and was added to the
differential set by Nielsen (1987a) . Resistance in TD2 may bè parttydominant (Fiq. 6.9À and C) . TD2 ís Renfrew, and is one of Oort,soriginal differential lines (Nielsen 1987a) .
The Cadet-Rescue inter-variètal substitution lines were originallydeveloped to evaluate milling and baking quality (Kosmolak et al . j.ggo).
Rescue and Cadet also differ in their loose smut reaction to variousraces (Tables Ä.1 and A,.2). Inoculation of the Cadet-Rescue
substitution lines with different U. tritici races produced some
unexpected resul-ts. TwÕ of the subst.itution lines tesbed t.o race Tg
\Àrere suscèptible (four lines were not inoculated) , and six lines were
susceptibl-e to race T15. possible explanat.ions for multiple linesusceptibiliCy include that either Rescue or Cadet. were het.erogeneous
for resistance to races Tg and T15, or that a univalent shift may have
occurxed during the development of certain lines (cale et a1 . 1989). Às
welI, mixing may have occurred during the handling of the seed stocks.Translocat.ions or mutat.ions would also be difficult to deEect, althoughit is diffj-cult to explain six susceptible lines when only one was
expect.ed, There is no evidence to indicate Chat the resistance to raceT15 is different from the resistance to race T8. Even though the datais questionabte, the results still suggest a focus for future research.ft. is tikely that one of the six susceptible lines truly represent.s theCadet chromosome possessing the resistance gene to T15, which has been
knocked out by substitution rvith one of the Rescue chromosomes. À more
detailed study of six chromosomes in cytogenetic stocks is a substantialxeduction in effort from analysis with 21 cytogenetic stocks. The
inoculations of the Cadet-Rescue series with race T19 provided clear

13 0
evidence that ¿he T19 resistancè is in chromosome 6À, which supports thefindings discussed in Chapter 4. The levels of infection for the otherlines are within the expecLed range of infection for cadet,
Some of the same problems of high infection on more than one
substitution 1j.ne occurred, as vrell, t¡ith inoculations of Chinese
Sprinq-Thatcher substitution lines (Table 6.3). The Chlnese Spring-That.cher lines were developed to assess many traits, but noC loose smut
resist.ance (Kuspira and Unrau 195?). Chinese Spring is rèsiseant toTl-0, T19, T3L, and T39, whereas Thatcher is susceptible to chese races(Table ,A,.4). Conversely, That.cher is resistant to races T6 and T15, yetchinese spring i6 susceptible. Therefore, inocul_ation of these
substieution lines offered an oppor¡unity to locate resisCance qenes inChinese Spring when knocked out by a susceptible Thatcher chromosome,
and to locate resiscance genes in Thaecher when a particular Thatcherchrornosome makes Chinese Spring resistant to race T6 or T15,
It is not unreasonable to believe that the gene conditioningresiseance to race T10 in chrornosome 28 of Chinese Spring al-so
conditions resistance to race T19, because of the similar reaction ofthe lines to these two races, Results for T6 and T39 are lesss traightforward, and may be due to faceors previously descrj-bed. as a
possible explanation for six susceptible tlnes in the Cadet.-Rescue
substitution line series, Considering that race T6 virulence is notbroad across the differentials, indicating this race has few viïulencegrenes (Nielsen 1987a) , it would not be surprising tha¿ Thatcher has more
than one gene for resistance to race T6, although four genes in fourchromosomes seems a very high number. Dhitaphichit et a1 , (1989) alsoinoculated the Chinese Spring-Thatcher substitution lines to race T6,
and they found infecEion in al1 lines but the Chinese Spring-Thatchêr78. They concluded that the major factor controlling loose smut
resistance in Thatcher is in chromosome 78. The face that four l-inesare susceptible to race T39 is most difficult to explain. It is

131
interesting bhat one suscept.ibl-è line to race T39 is Chinese Spring-
Thatcher 28 (chromosome 28 of Chinese Spring seems lo confer resistanceto xace T10) , and a second is Chinese Spring-Thatcher 7B (chromosome 7B
of Chinese Spring seems ¿o confer resistance lo race T31) . It is alsointeresting that only partial susceptibility occurs. perhaps there is a
series of chromosomes, each contributing some resistance lo race T39,
such that only partial resistance occurs when genes are removed, butwhen ¿ogether, complete resistance occurs. Tingey and Tolman (1934)
proposed Chat such a mechanism may occur in the cultívar Hope,
T15 resistance ín That'cher was demonstrated to be in chromosome 78,
whereas resístance to T3L in Chinese Spring resides in chromosome 78.
lf they are at the same locus, obviously they must. be different alle1es.However, it. is possible that rnore Ehan one loose smut resistance locus
occurs in chromosorne 78, which would favour breeding for resistance,because mult.ip]e loci can be recornbined so resist.ance genes aretogether, whèreas multiple atleles cannot. Resistance to T6 may be
conferred by the gene thaÈ confers resistance to T15 in Thatcher.ln sunmary, this repoxt provides evidence either through segregat.ion
rati.o or differences Ín race infection of different progeny, that morè
than one loose smut resistance g.ene occurs in lines in the loose smut
race differential set or their progenitors (Kot.a, Thatcher, TD18, TD6,
TD10, and TD16) and in clenlea and Hy3?7. Examples of dor0inance,
partial dominance, and recessive a1leLe interaction exists. Because ofthe sma11 sample size used in the present study, further work needs tobeen done t.o be conclusive as to the nature of the genetic makeup of thedifferentials and of key genetic sources of resistance used in breedingprograms, ff Glenl-ea does carry three genes for resistance, and onlythe combination of genes prevenEs disease, it follows Lhae if thissource of resis¿ance became widespread j.t may not be long-lasting. The
dependency of breeding for resistance on a narrow gene base empbasizes
the need for molecular markers to genes for resistance to allow

r32
pyramidinq of genes, even inbo a background such as clenlea for which no
knovm race possesses all the required corresponding virulence.The evidence Lhat a second resistance gene to race T15 may reside in
chromosome 6A in H44 should be foltowed up using a larger population. Tt
can be shown that the puCative H44 gene cannoC be the same gene as thatcontrolling resia¿ance to race TL9 in Kota, because Kota is susceptibleto race T15. However, the H44 resistance may be allelic to T19
resistance, which could be determined by an intercross of H44 and a T19
resistant source.
Evidence that the T19 resistance in Kota and TD18 is recessive cones
from the testcross progeny of populations Roblin NIB/Kota/ /prelude and
Roblin N1B/TD18 /prelude , That the T19 resistance is recessive, and inchromosome 6À strengthens the h1¡pothesis that the T19 resigtance in Kota
and TD18 is due to the same gene, Seronger evidence Ehat one gene isshared among these three cultj.vars would come from crossing each
conìlcination and determining whether or not segreqiation had occurred.
fnoculat.ion of the Cadet-Rescue chromosome substitution series provided
further evidence thae a gene conferring resistance to race T19 residesin chromosome 6À.
The information on race Tl-5 from the Cadet-Rescue substitution linesindicales that further study should be focused on chromosomes 1A, 3,A,
3D, 58, 6À, and 7B. Cadet is a derivative of That.cher (Zeven and Zeven-
Hissink 1976) , and Chinese Spring was made resist.ant to race T6 \,rhen theChinese spring chromosome was replaced by the Thatcher ?B chromosome, as
determined in a study by Dhitaphichit et al. (1989). These results were
confirmed in the presenE study, and inoculation of these substitutionlines with T15, to which Chinese Spring is susceptible, showed that when
the Thatcher 78 chromosome replaces the Chinese Sprinq 78, resistanceoccurs. À11 of these pieces of information strongly support thecontention that a loosê smut resistance gene in Thatcher is located inchromosome 78. The inocul-at.ion of the Chinese Spring-Thatchêr

133
substitution lines further indicates a resistance giene to race TLO inChinese Spring residing in chromosone 28. This information combined
with the results of Chapter 4, that. a T19 resistance gene resides incbromosome 6A of Thatcher, would indicate Thatcher has three genes forloose smut resistance.
Future work should fol1Õw up q'ith segregat.ion analysis of the
tentative gene locations determined by inoculations of inter-varieealsubstit.ution 1ines, Identifyinq the chromosomal location and
intercrossing of the different.ial lines is required Lo help identify thegenes and to produce single-gene lines for loose smut resistance. Such
single gene lines wj-]f aid in better discrimination of xaces and the
identification of new loose smut resistance genes.

134
GENERAT-, DISCUSSIOI\
A monoclonal antibody to a seed sLorage protein coded by the 1B
chromosome was used as a chromosome marker. The MAb chromosome marker
was used to track the 18 chromosome in a hybrid populalion in whjch one
of the parents was nullisomic for the 18 chromosome. Through the use ofauch a cytogenetic sCock, lhe 18 chromosome could segregate withoutrecombination of the genes within the chromosome, Although a MAb to a
seed Etorage protein has several advantages as a chromosome marker,
there are drawbacks to the use of nullisomic lines, the biggest being1ow fertility. In an effort to avoid the fertility probtems ofnullisomic lines, the usefulness of other cytogenetic stocks was
investigated. Àlien chromosome substitution lines tend to conpensat.e
for the loss of a homologous pair of chromosomes (Sears 1969). A MAb toa chromosome 6A seed proeein was used as a chromosome marker in a
population in which one parent was an afien substitution 1ine. Not onlydid the study confirm the value of a ¡lÄb as a marker, but alsodemonstrated that alien substitution li-nes can be easier to work with.By using ¡.t¡bs in conjunction with the appropriate cytogenet.ic stocks,chromosome analysis with a microscope is reduced to investigating theoccasional unusual xesult based on MAb analysis, and the population sizecan be reduced compared to monosomic analysis. Use of the MÀb
chromosome marker,/alien chromosome substitution line allowed thechromosomal- location of a toose smut resistance gene to be in chromosome
6À, With the knowledge of the chromosomal location of the l_oose smut
resistance gene, the focus of atteneion was turned to linkage analysisand mapping of the loose smut resistance gene with other chromosome 6A
traits. An association of the loose smut resistance gene with an RFLP
was found, although no association could be found between the foose smut
resistance gene and che gliadin locus. The use of several races to

135
inoculat.e each population allowed discrimination of more than one
segregating gene even \,,¡hen the population size was smal1.
Sharp and co\,¡orkers (1989) point out the convenience of using RFLps as
chromosome markers because they provide a uniform protocol of analysisused to mark all traÍts, compared to isozl¡mes, for example, which
require many different assays, An ELISÂ ís a uniform prococol for theanalysis of monoclonal antibodies, and such a test. is easier to perform
than either RFIJP or isozyme analysis. When M.Abs are used to assay seed
Þroleins, lhey provide a genetic rnarker method with a tevel ofefficiency unlike any o¿her narker proeocol . However, because thepotential number of seed protein gene loci is 1o\4' rel_ative t'o the number
of RFLP loci, seed proteins and MÄbs lo those proteins should be
considered mainly as chromosome markers and not as general genetic
markers. Ànother disadvantage to MÀbs is the 1arge amount of effortrequired to obtain a suitable ant.ibody (Klein 1990), sèed proteinsobserved through electrophoresis are more simply evaluated than RFtps,
and sufficient seed protein loci have been identified co mark each
chromosome in \-\¡heat. (Tab1e 2.5) . RFtp analysis requires a relativelylarge sample of DNÀ, too much to bê obtained from the endosperm end. of a
seed. Therefore, RFLP analysis requires ¿he germination of seeds and
lheir development into a tillering plant to provide sufficient DNÀ,
whereas seed protein analysis can be done on a porCion of the endosperm
of a resting seed. By using seed proteins as markers, the seed does nothave to be germinated and girown unLil after the results of the assay arekno\,ù¡ .
Random amplified poll¡morphic DNÀ (RÀpD) marker anafysis is a promising
alternative to seed proteins as chrornosome markeïs. R¿,pD analysisrequires less DNA than RFLP analysis (Williams and St, Clair 1993),
however, a question to be answered is whegher or not enough DNÀ can be
extracted frorn an ungerminated seed to make ehis method as efficient as
seed protein analysis. .Another drawback to the uêe of both RFLps and

13 6
R-APDS in wheat ís the lack of pollanorphism (He èt aI. L992¡ Dweikat eta1 . 1993; Joshi and Nquyen 1993), which nay be overcome through the use
of denaEuring gradient 9e1 ètectrophoresis (He et a!. 1,992; Ðweikat etal . 1993), Being able to detect chromosome dosage based on the amount
of bindinq of the MÃb is another advantage of the M.Ab. Classificationof progeny by the number of chromosornes, in addition lo identity of the
chromosome, improves the efficiency of the analysis. R-APD markers t.end
not to be codominant (Devos and cale 1992). The inabí1íty todisCinguish heterozygotes, hemizygotes or double monosomics diminishesthe value of RÀPD narkers as chromosome narkers.
Â.n import.ant condilion for tt¡e success of any chromosome marker
technique iE that the chromosome be inherited \\¡ithoue recombinaeion
taking place so that a correlation between a partj.cular trait and itschromosome can be esCablished, Although crosses were performed between
a loose - smut - suscept.ibl e nullisomic and a loose-smut-resist.ant source, a
testcross was required to recover ehe aneuptoid gametes. Monosomic
lines were suggest.ed as a substieute for nultisomic lines (Chapter 3) ,
however, one more evaluat.ion step is required to confirm that the
monosomic line is indeed monosomic and a testcross is st.i1l required torecover the aneuploid gametes, À1íen substitution lines were used withgood success, Âdvantages of the alien substitution lines h'ere lhe good
fertility of the lines, and, as a result, the lack of a requirement of a
testcross, An alien substitution line lacks a homologous pair ofchromosomes, but. this deficiency is compensated for by the addition of a
pair of homoeologous chromosomes (Sears j_969), When the loose-smut-susceptible 6 Agropyron alien substitueion line was crossed to a loose-smut.-resistanE line, a double monoso¡nic was generated in which a single6A chromosome from the resistant. parent. was presenl with a single 6A9
chromosome from the susceptible alien substitution 1inè. Because of thelack of pairing between the alien chrornosome and the wheat chromosome,
no recombination can occur wilhin lhe wheal chromosome.

1,37
Loose smut resistance is located in lhe 7A and 78 chxomosomes, based
on the inoculation of inter-varietal substitution lines (Ðhi¿aphichie eta1 , 1989), the drawbacks Èo the use of which were previously described(Chapt.er 4). This study successfully used the ¡,fÀb chromosome marker/
alien substitution line method to locate a loose smut resistance gene tÕ
chromoso¡ne 6À in four cultivars. Inoculation of inter-varietalsubstitutíon tines and alien chromosome subsCitution lines confirmed thegene l-ocation in the culÈivar Cadet. Inoculation of a ditelosomic and
an alien translocation line provided evjdence that the loose smut
resist.ance gene in thè culeivax Cadet is in the short arm of chromosome
64. Inoculat.ion of the chinese spring-Thatcher substitution linesconfirmed the resulls of Dhitaphichit and coworkers (1999) that a gene
resides in chromosome 78, Evidence of a gene for resistance in Chinese
Spring in chromosome 2B was also found.
A question tha! needs to be addressed is why locate genes tochromosomes at all? The location of genes to chromosomes is part of theprocess used to identify and characterize a qene (Bariana and Mclntosh1993). Locating a gene to a chromosome rnakes mapping lhe gene easier,and allows the more efficient determination of linkage wiLh markers
already mapped co the same chromosome. A putative RFLP marker h¡as found
by knowing ¿he chromosomal location of the gene and focusing linkageanalysis on RFLPS associated \,¡i th that chromosome. Knowledge of the 6A
chromosomal location of the loose smut resistance gene provided theopportunity to do linkage analysj-s with the qliadin locus although no
linkage was det.ected, RÃpD markers can be evaluated quickly compared toRFLP markers (Devos and cale 1992) , for exampfe, makinq it feasible toconcentrate on the development of a marker to a singl-e gene. Ä, map ofRÀPD markers woufd, however, a11ow the more efficient development ofmarkers to specific genes if the chromosonal location of bhe gene iskno\,m, Linkage analysis could be performed be¿ween the gene and allR.A,PD markers known to be in the same chromosome, rather than to all R¡,pD

L38
markers .
When considering the use of genet.ic markers? it. is important toconsider Che dra\,¡backs as well as thê advantages so that the most usefuL
markers can be foundr 1, The marker must be more efficient Co use than
the evaluat.ion of the trait it replaces (i.e,, cheaper, fast.er, less
labour int.ensive) ; 2, The marker must be sufficienEly closely linked(up to 15 cent.imorgans would be useful) Co the t.rait of interesC toprovide a hiqh probability that under select.ion for the marker the traitwill be pul1ed along, ídeally with Ehe maxker being pleiotropic with the
trait of interesti 3, The marker cannot confer any dêtrimental effecton individuals which carry the marker, and, ideally, the marker itselfshould be a trait with positive effecesi 4. The marker must be easilyidentified; 5. The marker must be in a cis configuration with the traitso that the known allele can be transferred wiEhout concern lhatdetrimental allefes exist for the same marker tocusi and 6. Àl1elicforms of the marker must exist, with the rare a]lele of the marker beingmos! useful .
The locaCion of a loose smut rèsistance gene to a chromosome, using a
monocfonal antibody as a chromosome marker, in conjunction with an alienchromosome substitution line, indicates that not only can loose smut
resistance genes be located co chrornosomes with this method, but thatother genes could be located to chromosornes in a similar manner. Ifmonoclonal antibodies were developed to seed proteins coded by each
chromosone, and a complete set of alien substieution lines weïe broughttogether, any gene could be located t'o any chromosome using this method,
These seed proeein 1oci, in conjunct.ion with cytogenetic stocks, couldbe used to determine the chromosomal location of genes cÕntrollingtraits of interest, and thus make the search for genetic markêrs easier.
The work here is just lhe beginning of what can be done not only inloose smut genetics, but in the development of molecular chromosome
markers. Future research can take a number of directions, including:

13 9
the study of the lnhêriLance of sources of loose smut resisÈance; the
breakdow¡ of sources of loose smut resistance into single-gene
components to simplify marker development and to provide an improved
race diffexential set, the development of MÀbs ¿o the seed proteinscoded by addj.¿ional chromosomes in wheat.; the determination of the
chromosomal location of additional loose smut. resislance genes; and
genet.ic analysis to identify closely linked markexs which coutd be
applied in breeding programs. More specific questions arising from thisresearch include: is the T19 resistance in chrornosome 6A in Kota,
Thaecher, Cadet and TD18 allelic; and does Che race T15 resistance gene
in H44 reside in chromosome 6À, and is it allelic co the T19 resistancegene? Segregat.ion analysis should be done to confirm the location of a
Chinese Spring loose-smut-resistance gene in chromosone 28, and,
similarly, to follow up on the work done by Dhitaptrichit and coworkers(1989) for the location of resisCance genes in chromosomes 7A and ?8.
Âs new markers located to chromosome 6À become available, linkageanalysis should be done with the 6Ä loose smut resistance gene in an
attempt to develop a more closely linked marker in the event that the
Cadet gene, or a nore effective allele of the Cadet gene should become
importanC in breeding. The T19 loose smut resistance gene should be
isolated into a susceptible genetic background to det.ermine how
effective the gene is against the spectrum of races, and to determineits penetrance in the absence of other genes for resistance.

CONCLUSIOI[
À gene for resistance to looee smut race T19 was shov¡n to be inchromosome 64, after the classification of segregating populations forchromosome make-up and for loose smub reaceion. Chromosomal locationanalysis is very efficient when monoclonat antibodj-es are used as
chrornosome markers in conjunction with alien chromosome substitut'ion1ines. The population size used to locate a loose smut resistance giene
to a chromosome was 20 or fewer. The location of a gene to a chromosome
facilitates the analysis of linkage with other traits and markers
located !o the same chromosome. Analysis of the race T19 loose smut
resistance indicat.ed that ghe gliadin locus in chromosome 6Â vJas notlinked; however, an RFLP located to the 6À chromosome might be
associated with the resistance. It should be possible to develop
molecular markers linked to loose smut resistance genes, for use inbreeding for resistance Eo toose smut..

L4L
LIST OF REFERENCESADF 1993. Varieties of grain crops for Saskatchewan l_993. SaskatchewanAgriculture Development Fund, Saskatchewan Agriculture and Food. 17 pp.
ÀgarwaL, V,K, and cupea, M.C, 1989. Loose smuC of wheat - À threat toseed production. Seed Res. 17: 55-68.
Àgarwal, V.K., Verma, li,S., Àgarwat, M, and cupta, R.K. L984. Studies onl-oose smul of wheat, II. Factors influèncing seed infection and seedtransmission. Seed Res. L2: 72 -'17 .
Àgrawal, R,K, and,Jain, K.B.L. 1965. Inheritance of resist.ance of N.p.790 to loose smut (Ustjlago tritici). Indian ,f , cenet. plant Breed. 25:376-380.
Agrawal, R.K., Rao, M.V. and ,Joshi, A,B, 1963. fnheritance of toose-smulxesistance in an inter-varietal- cros6 of Triticum aestivum L. Indian ,f,Genet. Planl Breed, 23: 135-140,
.ã.lLard, R.W. 1960. prínciples of plant Breeding. ,lohn Wiley and Sons,Tnc., USA. 485 pp.
Anderson, .t.À., Ogihara, y., Sorrells, M.E. and Tanksley, S.D. 1992.Development of a chromosomal arm map for wheat based on RFLP markers,Theor, Àppl. cene¿. 83: 1035-1043.
Ànderson, R.G, 1961 , The occurrence of loose smut resistance ln 42,28and 14 chromosome wheats. Can. .f. plant. Sci. 41: 828-835.
A¡on, 1963. Research in Che school of Âgriculture. Rep, Sch. Àgric,Nottingham, f962, pp. ]-9-34.
ÀragonciLLo. C., Rodriguez.Iroperèna. M.A., Carbonero, p, and carcia.Olmedo, F. 1975, Chromosomal conCrol of non-gliadin proteins from 70%ethanol extracC of wheat endosperm, Theor. Àppl . cenet, 45r 322-326.
Bares, I. and Kosner, ,J. 1975. Location of genes controlling somecharacters in the spring wheat variety Z1atka by monosomic ãnalysis,Euphytj-ca 24t 55'1 -563.
Bariana. H.S. and Mclntosh, R.A. 1993. Cytogenetic studies in wheat. XV.Location of rust resisiance genes in vpMl and their genetic linkage withother disease resistance genes in chromosome 2À. Genóme 36t 4j6-4g2.Bages, C.C.v. 1955a. Infection of wheaE by ]oose smut, UsÈi.Zagro t.ritici(Pers,) Rostr, Natu¡e 1?5r 467-468.
Batt,Ê, C.C.v. 1955b. Observacions on the infection of wheat by loosegmut (UsÊìJa9o trìt.ici (Pers.) Rostr.). T¡ans. Br. Mycot. soc. 3B¡ 465-
Batts, C.C.V. and üeater, A. 1958a, The development of loose srnut(UstiTaqo tritici) in suscepÈible varj-eties of wheat., and someobservations on field infection. Trans. Br. Mycol . Soc. 4!: LL5-L25.
Batts, C.C.V. and 'Jeater, A. 1958b. The reaction of wheat varieties toLoose smut as determined by embryo, sèedling, and adult. plant tests.^Ann. Appl. Biol .46:23-39,

1,42
Bever, W.M. and Oh¡ns, R.E, 1966. Effect of time of inocutation of winterk'heat with Ustitago tritici on t.he percen¡age of eÍìlcryos infected. and onthe abundance of hlæhae. phytopathology 46: 15?-158.
Biêtz, 'J.À., Shepherd, K.W. and Wall, J,S, 19?S. single-kernel analysisof glutenin¡ Use in wheat genet.ics and breeding. cexeal Chem. 52: SL:-
BlakesLey, R.W. and BoezÍ, ,J.À. I97j. A new staining technique forproteins in polyacrylamide gels using coomassie BïiilianE Eiue c250.Ànal . Biochem. 82 ¡ 580-582.
Boerema, G.H., Piet,ers. R. and ]tamers, M.E.C. 1992. Check-1ist forscientific names of common parasitic fungi. Suppl_ement. Series 2b(additions and corrections) : Fungi on fi¿Id cróps: cereals and grasses,Nelh, ,f , Pl . Path. 98 Suppl. !: L-32.
Brett, G.M., Mit1s, E.N.C.. parnar, S.. Tatham, À.S,, Shelrry, p.R. andMorgan, M,R.A. 1990, Monoclonal antibodies that recognizè the repeatmotif of the S poor prolamins, .t. CereaÌ Sci. 12: 245-255,
Brett, c.M,, Mills, E.N,C., Tatham, À,S,, Fido. R,J., ghewry, p.R, andMorgan, M.R.À. 1993. Irnmunochemical identification of LMw sùbunits ofglutenin associated with bread-making quality of wheat flours. Theor.A,pp1 , cenet.86t 442 - 448 .
BriggL€, IJ,W. 196?. Morphology of the wheat ptant. pages 89-116 jn K.S.QuisenberLy, ed. wheat and wheat Improvemencl amerj.cai society ofÀgronomy, Inc., Madison, Wisconsin, USÀ.
BriggLe, L.W. and Curtis, B.C. 1982. wheat Worldwide. American Societyof Agronomy, Inc., Madison, Wisconsin, USÀ. 1-32 pp.
Brown, J.W.S. and FLavel1, R,B, 1981. Fract.ionation of wheat gliadin andglutenin subunits by two-dimensional êleccrophoresis and lhe iole ofgroup 6 and group 2 chromosomes in gliadin synthesis. Theor. Appl .cenet. 59 | 349-359.
Burnouf, T, and Bietz, \T.À. j.9g5. Chromosomal control of gluteninsubunite in aneuploid tines of wheat: analysis by reverseá-phase high-performance liquid chromatography. Theor. Àppl . cenet. ?O: ÈfO-efg.-CampbelL, A.B. 1948, The inherieance of loose smut reaction in a crossbeLween two sprÍng wheaLs¡ Redman and That.cher, Master,s Thesis,University of Manitoba, Winnipeg.
CampbelL, B. and Czarnecki, E. 199?. Roblin hard red spring wheat. Can.¡f , Pl-ant . Sci . 67 : 803 - 804 .
Chao, S., Shar¡r, P.,t. and cal.e. M,D. 1998. À linkage map of q,heathonoeologous group 7 chromosomes using RFLP markerÀ. pages 493-49g jnT,E. MilLer and R.M.D. Koebner, eds, Þroceedinss of lhe SevêlthInternational Wheat cenetics Sl¡mposium, Institúte of plant ScienceResearch, Canbridge,
Chao' S., Sharp, P,,J., worland, A.,J., warham, E.,t., Koebner, R.M,D, analGa1e, M,D. 1989. RFLP based genetic maps of wheat homoeol_ogolr" gro,rp ichromosomes. Theor. Àppl , cenet, Zgr 495-504.
Chatrath, M.S, and BahL, N. 1966, Effect of concentration of inoculum oninfectj-on in loose smut of v¡heat. Indian phycopáEhology !9t 3!2-3!3,

143
Chênicek, K.,t. and Hart, G.E. 198?. Identification and chromosomallocations of aconitase gene loci in Triticeae species. Theor. Àpp1 .Genet.74:26I-268,Cherewick, W.,t. 1958. Cereal smut races and their variability. Can. ,f.Plant. Sci. 38 ¡ 481-489.
Chrisbensen, J,'J, and Rodenhiser, H.A, 1940. lhysiologic specializationand genet.ics of the smu¿ fungi, Bot, Review 6: Zgg-¿Zi.
Conner, R,Ir., Whetan, E.D,p., IJaroche, À. and Thomas, J.B, i.993.Reaction of alien chromosome substitution and addition lines of hard redspring wheat to corunon root rot and btack point. cenome 36: 173-180.
Czarneckí, E.M. and Lukow, O,M, L992. Linkaqe of sten xust resist.ancegene sr33 and Lhe gliadin (cli-D1) locus on chromosome l_DS in wheat.Genome 35: 565-568.
Dawood, M.R., Hones, N.K, and Bushuk, W. 1989. preparation of monoclonalani:ibodies
. agaj-ns t specific gliadín proteins and pieliminaryinvestigaeion of bheir ability to diicriminate cereal cultivars. .t.Cereal Sci. 10: l-05 - 112.
Dean, w.M, 1965. some effects ofof wheat. [Rep. Sch, Ã.gric. Univ.510 .
Dean, W.M, 1969. The effect of t.emperature on loose smut of wheat(Ust.iTago nuda) . Ànn. Appl, Biot. 64: 75-83.
de Man, ,t.M. L990. principtes of Food Chemistxy. van Nostrand. Reinhotd.,New York. 469 pp,
Devos. K.M. and caLe, M.D. 1992. Thè use of random amplified poll,rnorphicDNÀ markers in wheat. Theor. Appl , cenet. 94: 56j-570.Devos, K,M. and caLê, M.D. 1993, Extended genetic maps of thehomoeologous group 3 chromosomes of wheat, rye and bãr1ey. Theor, ,AppI .Genet.85¡ 649 - 652.
Devos, K.M., Àtkinson, M,D., Chinoy, C,N., Iríu. C.,t. and caLè, M,D.1992. RFLP-based genetic map of the homoeologous group 3 chromosomes ofwheat and rye. Theor. .ã,ppl_. cenet, 83: 931-939.
Devos, K,M., Millan, T. and cale, M.D. 1993. Comparative RFLP maps ofthe_ homoeologous group-2 chromosomes of wheat, rye and barley, Tireor.Àpp] . cenet.85t 784-792.
Dhitaphichit, P., ,tones, p. and Keane. E.M. 1999. Nuclear andcytoplasmic gene control of resistance to l_oose smut (Ustjlago tritici(Pexs.) Roscr.) in wheat (rriticun aestivum L.), Theor. Appl: cenet.. 7gl897-903.
Diaz, F., ¡'ernandez, ,J.4. and Jouve, N, 19g6, Structure and chromosomallocation of malat.e dehydrogenase (zone Z) isozlrmes in common an¿ aurumwheats. Euphytica 35: 509-513.
Donovan, G,R., Skerri!9, 'J,H. and Castle, S.¡,. 1999, Monoclonalantibodies used to charact.erize CDNA clones expressing specific wheatendosperm proieins, !7. Cereal Sci. 9¡ 9.1 -I]1-.
temperature and humidity on loose smutNote, 1964:50-55.1 Rev. Àpp1 . Myc. 44t

1,44
du Cros, D.L. and Hare, R.A. 1985. Inheritance of gliadin prot.einsassociated q'ith qualÍty in dururn wheat. Cïop Sci. 25¡ 6j4-67.t.Dweikat, I., Mackenzie, S., i,ew, M. and Otun, H, 1993. pedigreeassessment using RÃPD-DGGE in cereal crop species. Theor. Àipt. cenet.85:497-505.
Dyck, P.i,. and Kerber, E.R, L9?1. Chromosome location of three gienes forleaf rust resistance in common wheat. Can. ,J. cenet. Cytol . 13: 4g0_483.
Dyck, P,L. and Kerber. E.R. 197?. fnheritance of leaf rust resistance inwheat cultivars Rafaela and EAp 261-27 and chromosome location of geneLrL7. Can. ,-T. cenet.. Cytol , 19¡ 355-358.
Dyck, P,L. and Kerber, E.R, 1995. Rêsistance of the race-spêcific type.Pages 469-500 in À.P. Roelfs and w.R. Bushnel] eds. The Ceieaf nustê-,Vo1 . I1. Âcademic press, Orlando, Floxida.Dyck, P.L., Kerber, E.R. and l¡ukow, O.M, 1992. Chronosome location andl-inkage of a new gene (Lr 33) for reaction Lo puccinia recondita inconunon wheat, cenome 29: 463-466,
E91u, K. 1977. .â,naeorny of seed plants. ,John Wiley and Sons, New york.550 pp.
Feldman, M. 1975, Triticum spp. (cramineae-TriEicinae), pages 1,20-!28 inN.W. Simmonds, èd, Evolution of Crop plants, Long,rnan Inc., New york.
Fischer, c.W. and Holton, c,S, 1952. Biology and Control of the smutFungi. The Ronald press Co,, New york, 622-þp,
FLeischnann, c. and Baker, R.,t. L921. oat crown rust racedi f f erentiation: repracement of the standard differential varietiès witha new set of single resj-st.ance gene lines derived from Àvena steriTis.Can. ,f, Bot. 49: 1,433-f437.
Forsyth, S.À. and Koebnêr. R.M.D. 1992. Wheat endosperm hiqh molecularr{eight albumins and beta amyrasesr genetic and erecirophorátic evidenceof thej.r identity. ,1. Cerèal Sci. 15: !3.1 -L4L,
Fra-Mon, P. Salcedo, c. and carcia- Olnedo, F, L994, chromosomalassìgnment of genes conCrolling salt-solubie proteins (aLbumins andglobulins) in wheat and relat.ed species. rheoi. appl . cenet. 69: 167-
Friebe, 8,, ZelLer, F.!t., Mukaí, y.¡ !'orster, 8.p., Bartos, p. and.Mclntosh, R.À. 1992. Charac terj zaeion of rust-resistant w:nea:-- Aqropyroninternedìut, derivatives by c-banding, in situ hybridization u"d-i;ã;V.;analysis, Theor, Appl . cenet, g3 ¡ 775-792.
FroÍune, F.D. 1920. The development. of loose smub of wheat as modified bysoil fertility. phytopathology 10¡ 53.
Gabor, B. 1985, The histology of loose smut resistance in barley.Master's Thesis, University of Manitoba, winnipeg.
Gabor, B.R, and Thomas, p.I,. ].98?. Ung atlefe for loose smut resistanceassociated with necrosis in embryos of infected barley. phytopatholoqy77: 533-538.

t4s
Gale, M,D,, IJa!¡, C.N., Chojeckí, A.!t. and Kempton, R.À. 1993. Genet.iccontrol of alpha-amylase production in wheat. Theor. Àpp1 . Genel. 64r309-316.
GaIe, M.D,, Sharp, P,\T., Chao, S, and Law, C.N. 1989. Àpplicatj-ons ofgenetic rnarkers in cytogenetj.c manipulation of the wheal genome. Genome3]-:137-142,
Galfre, c. and MiLseeÍn, C. 1981, preparation of monoclonal antibodies:Strategies and procedures. Methods Enzl¡rnol . 73t 3-52,
Galiti, c, and FeLdman, M. 1983, cenetic controt of endosperm proteinsín wheat. 1, The use of high resolution one-dimensional gè1el ect.rophores i s for the allocation of qenes codÍng for endosperm proeeinsubunits in the comnon wheat cultivar "Chinese Spring', , fneoi. appt,ceneC . 64 : 97 - 10f .
Garcia, M,F., Marana, C., Mena, M., carcÍa, O.F. and Carbonero, p. 1990.cloning of CDNA and chromosomal locat.ion of genes encoding the threetl¡pes of subunits of the wheat tetrameric inhibitor of insect alphaamylase. Plant Mol . Biol , 14: 945-853.
carcia-Olnedo, F,, Carbonero. p.. Àragoncj-LLo, C, and Salcedo, e, fg7g.chromosomal control of wheat endosperm proteinsr A criticat review,Paqes.555-566 in IÂEA, ed. Seed protein fmprovement. by NuclearTechniques, International Atomic Energy egèncy, Viennã, Austria.Garcia-Olmedo, F,, Carbonero, p. and Jones, B.Ir. 1992. Chromosomallocations of genes lhat control wheae endosperm proteins. pages L-4? inY, Pomeranz, ed. Advances in Cereal Science Technology, AmericanAssociat.ion of cereal ChemisLs, Inc, st. pau1, Minnesõta, UsA.
Gaskin, T,À. 1958, Developmeneal and qenetic basis of resistance inwheat to loose smut. Diss. Àbst. 18.
Gaskin, T.A. and gchafer, J.F. L962. some histological and geneticrelation8hj-ps of resistance of \aheat to loose smut. phyeopaÈholoqy 52:602 - 607 .
Gíl1, K.S., Lubbers, 8.L., cill, 8.S., RauÞp, W.,J. and Cox, T.S. 1991. Agenetic linkage map of ?riCjcun Eauschii (DD) and its relationship tothe D genorne of bread wheat (ÀÀBBDD) . cenome 34t 362-374.
GoLd.enberg. E.M. 1986. Chromosornal location of peptidase, pEpT-1, genesin friÈicum aestivum var. Chinese Spring, cenet. Res, 48t 1,9-2O.
GothwaL, B,D, L972. proper stage for the inoculat.ion of wheat plantswibh the loose smut pathogen, fndian ,f . Mycol , plant. pathol . 2: 89.
Graner, A.' Siedler, ¡I., ,Jahoor, 4.. Herrmann, R.c. and wenzel, G. 1990,Àssessment of the degree and tl4)e of restrictj-on fragment. Iengthpolj¡moxphism in barl-ey (Hordeum vuLgare). Theor. Appl . cenet. 80: 826-832 .
Graybosch, R.A, and Morris, R, X990. Ân improved SDS pÄcE method for the;îily"t"
of wheat endosperm storage proreins, ,J. Cereal Sci, 11 : 201-
Graybosch, R,, geo, Y.W. and Peterson, C.,t. 1993. Detection of \,¡heat - ryechromosomal LransfocaÈions using an antisecalin monocfonal antibody,Cereal Chem. 70: 458-463.

746
Green, c.,t. 1981. Identificacion of physiologic races of pucciniagraminis f ,sp, Èrjtjci in Canada. Can. ,f. plant paChol , 3: 33-39.
Gtegory, P.H. 1952. Fungu6 spores. Txans. Br, Mycol . Soc.35: 1-18.
Grevel, F.K. 1930. Un¿ersuchungen úber das Vorhandensein biologischerRassen des Flugbrandes des Weizens (UstiTaqo tritici), phytopa¿hot, Z.2t 209 -234.
Gupta. R.B. and Shepherd, K,W. 1988. Inheritance of novel high motecutarweight gfutenin subunits in the Tunisian bread wheat ,BT 22gg,, cenome30t 442 - 445.
Gupta, R.B. and. Shepherd., K.W. 1990, Two step one dimensional SDS pAcEanalysis of LMV{ subunit.s of glute1in. l_. Variation and genetic controlof the subunits in hexaploid wheat.s. Theor. Appl . cenet. gO: 65-74,
Gupta, R.8., Shepherd, K,W. and MacRitchie, F, 1991. cenet.ic control andbiochemical properties of some high molecutar wêight albumins in breadwheat. ,t. Cereal Sci. ]-3: 22t-235.
Hanna, W,F. 1937, physiologic forms of loose smut' of wheat. Can, J, Res.Ser. C, 15: 141- 153.
Hanson, W.D, 1959, Minimum family sizes for the planning of geneticexperiment.s. Àgronony ,Journal 5L: 711-715,
Harcourt, R.IJ. and caLe, M.D. 199L. A chromosome - speci f ic DNÀ sequencewhich reveals a high leve1 of RFLP in wheat. Theor. Appl . cenet. g1:397 - 400 .
Hart, G.E, 1975. Glutamate oxaloacetate eransaminase isozl¡mes ofTriticumt Evidence for multiple systems of tripticate structural genes.Pages 637-657 i¡ C.L. Market, ed. fsozymes, Àcademic press,
Hart, G.E, 1987. cenetic and biochemical studies of enzlmes, pages 199-21,4 in E.G. Heyne, ed. wheat and wheat Improvement, Arneiican soðiety ofAgronony, lnc,, Madison, Wísconsin, USA.
Hart, G.E. and. Irangston, p.J. !977. Chromosomal locatj_on and evotutionof isozlrme structural genes in hexaploid \,¡heat. Heredi Ly 39t 263-277.
HarC, G.E., Islan, A.K,M.R. and Shepherd, K,w, 1990. Use of isozl¡mes aschromosome markers in the isolation and characteri z ation of wheal.-barleychromosome addition 1ines. cenet. Res. 36: 31-:- -325.
He, S., Ohm, H. and. MacKenzie, S. !992. Detection of DNA sequencepoll¡morphisms among wheat varieties. Theor, Appl . Cenet, 94i 5:3-57g.Heun, M., Kennedy, .A'.8., .Anderson, J.À., Lapitan, N.L.V., SorrellË, M.E.and TanksLey, g.D. 1991, construction of a restriction fragment. fångfh
'
pollmorphism map for barley (Hordeum vuJgare). cenome 34t Ã3j -44'1 . -
Hel¡ne, E.G. and Hansing, E.D, 1955. Inherit.ance of resistance to loosesmut of wheaC in the croases Kawvale x Clarkan. phytopathology 45: g-10.
Holton, C.S, 1953, physiologic specialization and gênetics of the smutfungi. II. Bot. Review !9: !8j -208.
¡Iowes,.N,K., Í,ukow, o.M., Dawood, M.R. and Bushuk, W, 1999. Rapiddet.ect.ion of che 1BL/1RS chromosome transfocation in hexaploid wheats bymonoclonal antibodies, ,J. Cereal Sci. 10: 1-i.4.

t47
llor,res, N.R., Kovacs, M.I. and Rerber, E.R, 1990. D genone suppression ofgliadin electrophoretic band 45 in hexaptoid wheat ánd the eiiect uponquality. cenome 33: 395 - 399.
Howes. N.K., Whei-an, E.D.p., IJukow, O.M. and Skerritt. ,t.H. 1993.rdentification of alien chromosome subsritutions that confer resistanceto colonization by the whea t. curl mite using r0onoclonal antibodiesspecific to the short arm of chromosome 6Ä óf common wheat. cenome (Inpress) .
Hueros, G., Sanz, ,t.C. and ltouve, N. 1988. Chromosomal location by Flmonosomic analysis of endosperm pxoteins in bread wheat. 1. Onedimensj-ona1 electrophoresis of glutenins and gliadins. Theor. AppL.cenet. 76: 787-'187,
Iannetta, P.P,M., ,tames, E.K., McHard.y, p,D,, Sprene, ,f .I. and Minchin,F,R. 1993, Àn ELISA procedure for quantification of relaLive amounts oiintercellular glycoprolein in legurne nodules, Ànn. Bot. 71¡ 95-90,,taaska, V. 1980, Electrophorecic survey of seedling esterases in wheatsin relatíon to their phylogeny. Theor. Appl , cenet. 56: 2.13-284.
,tackson. E..A,., l¡o1t, Ir.M. and palme, p.f. 1983. Characteri zat.ion of highmolecular weight gliadin and low molecular weight glutenin subunits ofwheat endosperm by two dimensionaÌ electrophorèsis and the chromosomallocalisation of their controlling genes. Theor. Àppl , cenet. 66t 29-37.
'tiazhi, P., Rongj in, s,, zhigang, y. and ciuyinq, z. ],ggg. Monosomicanalyses of ye11ow rust and leaf rust resistancè genes in winter wheatcultivar 'Fengkang,. plant Breeding 11Or 153-156.
Jones, P. and Dhieaphichit, p. 1991. cornparison of response Co floret.and seedling inoculation in wheat Ítstifago t.rit.ici and barley ÍlstiJ.agonuda conìlcinations. plant pathol . 40,t 268:2.17.
üones, S,S., Dvorak, ,J. and eualset, C.O. 1990, Linkage relations of cliDI, Rg2, and Lr21 on thê short arm of chronosome 1D in wheat. Genome 33:937 -940 .
qon-eq, q:s., Dvorak, ¡I., K¡ott, D,R. and pualsêt, c.o. 1991, use ofdouble ditelosomic and normal chromosome îD reconùcinant substitucionlines to map Sr33 on chromosome arm 1DS in wheat. cenome 34¡ 505-50g.
üoshi, C.P. and Nguyen, H,T, 1993, Âpplication of the random amplifiedpolymoxphic DNÀ technique for the detection of poll¡morphism amoirg wildand cultivated tetraploid wheats. cenome 36: 60r-609.
Kaloshian, I., Waines, ,J.c., Roberts, p,À. and Thomason, Lif. 1991,chromosomal locat.ion of root - knot - nematode resistance gàne in the ogenome of wheat. ,J. Hered. 82r 254-256.
Kavanagh, T. X959. À technique for seedling inoculation wiehchlamydospore s of UstiTago nuda and U, t.riticì, The effect oftemperature on the development of loose smut in adult plants of \,¡heatand baxley. Phytopathotogy (Älcst) 49t 542-543,
Kavanagh, T. 1961a, Infect.ion and developmental seudies on ustiJago nudaand u, È-ritici. Diss. abst. 2l¡ 2849-2BSO,
Kavanagh, T. L961b. fnocutating barley seedlings with UstiTago nuda andwheat seedlings with U. tritici, phytópathology St: tlS-tll ,-

148
Kavanagh_, T. 196Lc. Ternperature in relation to loose smut in barley andr,¡heat. Phytopathology 51: 189 - 193.
Kena, G.H.'J. and lJange, tñ. ]-992, Resistance in spet t h'hea t. to yellowrust. II. Monosomic analysis of the Iranian accession 415. Euphytica 63:2r9 -224.
Kerber, E,R. 1988. Telocentric napping in wheat of gene L\32 forresistance to leaf rust. Cxop Sci. 28: 179-ú9.Kerber, E.R. and Dyck, p.IJ. 1973. Inheritance of stem rust resistancetransferred from diploid wheat (?ritjcum monococcum) to tetraploid andhexaploid wheat and chromosome location of the gene involved. Can. ,t.Genet. Cytol . !5t 397-409.
Khan, K., Hamada, À.S. and Patek,electrophoresis for wheat varietygel properties. Cereal Chem. 62¡
KiLduff, T. 1933, Inheritance ofcertain othex characters in KotaRes, 8: 147 -!72.
,t, 1985. Polyacrylamide gelidentificationr Effect of variables on
310 - 313 .
bunt and loose smut reaction and ofx Red Bobs and Garnet. crosses. Can. ,t.
Kirnber, c. and Sears, E.R. 198?, Evolution in the genus Tîiticum and theorigin of cultiva¿ed wheat. pages !54-L64 jjt E,c. Heyne, ed. Wheat andWheat Improvement, Àmerican Society of Âgronomy, Inc., Madison.
Kirby, G.C. 1984. greeding systêms and heterozygosity in populations oftetrad forming fungi, HerediÈy S2t 3S-4!.
K1ein, 'I. 1990. Inmunology. BlackwelI Scientific pubtications, Inc,,Boston. 508 pp.
Knott, D,R. 1987a. The Wheat. Rusts - Breeding for Resistance. Springer-Verlag, cermany, 20j. pp.
Knoet, D.R. 1987b. Transferring alien genes to wheat, pages 462-47I inE,c. Heyne, ed. Wheat and wheat frnprovement, Àmerican So¿iety ofAgironomy, Inc., Madison,
Koebner, R.M,D., Miller, T,8., Snape, J,W. and IJaw, C.N. 19gg. wheatendopeptj-dase; genet.ic control, polymorphism, intrachromosomal genelocation, and alien variation. Genome 30: fg6-Lg2.
Konarev, A.v. 1981, The genome specific grain proteins and thephylogenetic inCerrelaEion beEweèn rritiêun L,-, ETytriqia oesi., al¡zmusL. a\d Agropyron caertn, Theor, Àppt. cenet., 59t fj.,l -Ir!.Kosmoj-ak, F,c., Larson, R,f . and McKenzie. H. L9gO. Millinq and bakingquality of Rescue-cadet reciprocal- substitution tines. can] J. piã"È 'Sci, 60: 1333 - 1341.
Kreis' M.,_ Fglqg, B.G., slrewry, p.R., wi11iaÌnson, M,, p¡¡r¡e1I, iL, rJewís,H,H.A. and MifLin, B.ù. 1995. Isolation and characteri zãtion' oi ;.;;åi-'seed protein genes, Adv. Agric, Biotechnol , LO6-107.
Kuspira, ,t, and Unrau, J. L95?. cenetic analysis of certain charact.ersin.common wheat using whole chromosome subst-itution lines. Can. ,¡. piãntSci..37:300-326.
Lafj-andra, D,, Kasarda, D.D. and Morïis, R. 1994. Chromosomal assignnentof genes coding for the wheat. gliadin protein component.s of the

1,49
cultivars 'Cheyenne, and ,Chinese Spring, by two dimensional (two pH)el-ectrophoresis . Theor. Äpp1 . cenet. 68: 531-539.
Lamari, L. and Bernier, C.C. 1991. cenetics of tan necrosis andextensive chlorosis in tan spot of wheat caused by pyrenophora tritici-repentis. Phylopathology 81r 1092 - 1095,
Irarose, E, and Vand.ervalle, R, 1937, Some re6ults obtained in theartificial infection of wheat by Ustilago nuda triLici schaff. leuelquesresultants d,infection artificielÌe d'Ustijago nuda tritici schaff. Jur1e Fromenl. 8u11. Inst, agron. cem.lcloux, VI:81-B?l . Rev. ,Appl_. Myc. 16!'7 32 -'133 .
IJawrence, c.L. and Shepherd, K.W. 1991. Inherj-tance of glutenin proteinsubunj-Cs of wheat.. Theor, .A,ppt. cenet. 6Or 333-337.
Iriu, C.,J. and cate, M,D. 1991. The chromosomal locat.ion of genesencoding NÀDH dehydrogenase isozymes in hexaploid wheat and relatedspecies. Genome 34r 44-5L.
l,iu. C.,t., Chao, S. and cat-ê, M.D. 1999. Wsp-1, a set of genescontrolling water-soluble proteins in wheat and related species, Genet.Res. 54 | 173 - 181.
Liu, C.,J., Chao, S. and cale, M.D. 1990. The genetical control oftissue-specific peroxj.dases, per-1, ner-2, eei-3, per-4 and per-5 inwheat. Theor, Àppl-. cenet. 79: 305-313.
IJíu, Y.c. and Tsunewaki, K. 1991. Restriction fragmene lengthpoll¡morphism (RFLP) analysis in wheat. Tf. Linkage Maps of the RFLPsit.es in cornmon wheat . (Iap. ,J. cenet . 66 | 6L:. - 633 ,
Lookhart, G.Ir., Cooper, D.B. and Jones, B.L. 1995. EffecC of temperatureand alternate lacCate buffer syst.ems on resotution of wheat gtiaãinproteins by polyacrylamide gel et ectxophorea i s . Cereal C¡rem. 62t lg-22,IJoria, R., Wiese, M. and ¡Jones, A.Ir. 1992. Effects of free moisture,head developmenc and ernbryo accessibility on infection of whêat byUsEilago tritici. phytopathology't2t f27O-L2j2,
Mackenzie, D,R. 1991. progress in plant disease resistance research. FAOPlant Prot. 8u11, 39: tLi -i.54,
MacRitchie, F,, Cros, D.L.du and Wrig1ey, C.W. 1990. Flour pottæeptid.esrelated to wheat quality. pages 79-145 jn y, pomeranz, ed. Àdvãàcès inCereal Science and Technology, Àmerican Assoc, of Cereal Chemists, Inc.,SE, Pau1, Minnesota.
Malik, M,M.S. and Batts, C.C,V. 1960. The deLermination of the reactionof barley varieties to loose smut. Ànn. Appl . Biol , 49: 39-50,Mantle, P.c. 1961a, The effect of vernalizatj-on on ernbryos of fieldresietant wheat varieties infected by loose smut (us¿il;go tritic.i, .À¡n. .Appl . Biol. 49: 692-703.
Mant,Le, P.G. 1961b. Further observatj-ons on an abnormal reaction ofwheat. to loose smut, Trans. Br. Mycol , Soc, 44¡ 529-545.
MantLe, P.c. 1961c. persistence of loose smut ìnfection wiehin thegrowing point region of susceptible h'heat varieties. Tïans, Br, Mycol .Soc. 44; 546 - 548 .

1s0
Masojc, P, and Gale, M.Ð, 1990. The factor nodifying alpha-amylasei"9?yT" pattern from rye endosperm is an endogenóus
- alpËa - amyiaseinhibitor. Hereditas 113 ¡ 151-155.
Masojc, P., zawistowski, ,t,, Howes, N.K., Àung, T. and. cale, M,D, 1993a.Poll¡morphism and chromosomal location of endogenous alpha-amylaseinhibit.or genes in conmon wheat. Theor, Àppt, Genet. 95: 1043-L048.
Masojc, P,, zawisto!¡ski, ,J., Zawistowska, U. and Howes, N.K. 1993b, .Acombined monoclonal and polyclonal antibody sandwich EiISÀ, forquanCifícation of the endogenous alpha-amylase inhibitor in barley andwheat . !T , Cereal Sci . L7 : 1,1,5 - 1,24 .
Mabhur, H.C. and Kohli, S.p. 1963. Inheritance of resistance to loose_snul in Triticum aestivum. Indian J. cenet. 23: 256-ZSg.
McIntoÊh, R.À, L972. Cytogenetical sLudies in wheat VI . Chromosomelocation and linkage studies involving Sr13 and Srg for reaction toPuccinia graminis f. sp. ¿ritjcj. Àust. ,f. Biot. Sci, 25¡ 765-.173.
McIneosh, R.À, 1978. CytogeneticaL sCudies in wheat. X, Monosomicanalysis and linkage studies i.nvolving genes for resj-stance to puccj¡tjagraminis f. sp, trjticj in cuttivar Kotå. Heredity 4Lt 7I-82.Mclntosh, R.À, 1987. cene location and gene mapping in hexaploid wheat.Pages 269-28'1 in E.c, Helme, ed. wheat and Wheal Improvemenl, ÀmericanSociety of Âgronomy, Inc., Madison.
McIntosh, R.À. 1988. Catalogue of gene sl¡rnbol s for wheaC. pages 1225-!323 in T,E. Mi]ler and R.M.D, Koebner, éds. Seventh Internationat WheatGenetics Symposium, IpSR, Cambxidge,
Medeiros, M.C. 1976. occurrence of physiologic races of loose smut of\a¡heat, usËj-taqo tritici (pers.)Rosti., in Biazil, sources of resistanceand inheritance of xesistance to race T2 in the int.ervarietal crossKenya 340 Y.4.Ã.1 X IAS 52 . Master,s Thesis, UniversiLy of Manitoba,Winnipeg .
Mena, M., Doussinault, G., Lopez, 8.I., Àguaded, S., Garcia, O.F. andDelibes, A, 1992. Eyespot resisEance gene pch 1in iI 93 wheaC fines.-Evidence of tinkage to markers of chrómosome group 7 and resotution fïomthe endopeptidase locus Ep D1b, Theor. Ã.ppt. Cenet. 83t !044-I04j.Metzger, R..I., SchaLler, C,W. and Rohde, C.R. 1929, Inheritance ofresistance to corrunon bunt. in wheat, C,f . ?090. crop Sci. 19: 309-312.Milan, À. 1938. Summer experiments on early wheat varieties withUstiTaqo tritici (pers).Tens. Iprove estj-ve sul1, ,.ustiJago tîitici,(Pers.) .fens, convarieta di Grano precoci. Riv. pat. veq. 2.1 ¡2g7-296(1937)1, Rev, .App1 , Myc, 17r 166.
Milan, À. L939, The susceptibility Lo UstiLago tritici (pers,) Jens. ofsome normal hybrids of wheat lsensibilita per la Ustj-Zago tïitici(Pers.) Jens, di alcumi ibridi di frumeneol Riv, pat, veg. 2927!-g4.(1939). Rev, Appl . Myc. 29¡ 71"-84.
Mi1ls-, E.N.C., Burgess, S.R., Tatham, A.S., Sheyrry, p.R., Chan, H.W.S,and MorgÀn, M,R.À. 1990, Characteri zation of a panel of monoclonal antigliadin antibodies. ,.f. Cereal Sci. L1: g9-101.
MiLstein, C. 1980. Monoclonal antibodies. Sci, .Aner. 243: 66-i4.

151
Morris, R. 1987. Origins and analyses of genes and qenones in wheat andits relatives, Pages f52-153 in E,c. ueynè ed. Whea¿ and wheatfmprovement, American Society of Àgronomy, Inc., Madison,
Ng, P.K.w. and Bushuk, W, 1987, cluteni-n of Marquis wheat as a referencefor esLimating rnolecular weights of glutenin subunits by sodium dodecyfsulfate polyacrylamide gel electrophoresis. Cereal chem, 64t 324-32,1 ,
Nichotson, P., Rezanoor, H.N. and Wor1and, À,,J. 1993. chromosomatlocation of resistance to Septoria nodorum in a synthetic hexaploid.wheat determined by the study of chromosomaf subsÈitution tineã in'Chinese Spring, wheat. plant' Breeding L!0¡ I'17-!84.NieLsen, ,I. 1969, Â race of Ustijago tritici virulent on Thatcher wheatand its derivatives. plant Dis. Rep. 53: 393-395.
NÍelsen, J. 1972. fsolation and culture of monokâryoLic haplonts ofUstilago t.titici, observacions on their physioloqyl and thè taxonomicrelationship betneen U. tritici and t_r. nuda, Can. J, Bot, 5O: 1,j75- j,,tg:-,
Nie1sen, ¡J, 1973, Reaction of triticaLe and spring rye to toose smut of\4'heat. Can. ,J. p1ant. Sci. 53 t 749-,153.
NieLgen. J. !977, Inheritance of virulence of loose smut of wheat,rJstì7ago tîitici, on the differential cultivars Renfrew, Florence xAurore, Kota and Little Club. Can. ,f. Bot.. 55: 260-263.
NieLsen. J. 19?8a. HosC range of the smut species Us|iLago nuda andUstiTago tritici in the ¿ribe Triticeae, Can. ¡. not, 56¡ 901-915.t_{,i?f::en, ,J. 1978b. Tr4o grasses as conmon hosts for al1 species ofUstiTago parasitic on wheat, barley and oats. Can. ,1. Bot. 56t 271-2-2716 ,
NieLsen, J. 1982. Inheritance of virulence of tJstitago tritjci on thedifferential cultivars carma, Red Bobs, and a derivaÉive of ttre cróssThatcher X Regent Spring wheat. Can. .1. Bot. 60r 11.91-1193.
Ni?Lqeq, ,t. 1983. Spring wheats i¡nrnune or highly resisrant to nsËj-Zag¡otrit.ici, plant Dis. 6?: 860 - 863
Nielsen, ,1. 1985. UstiTaqo spp. pathogenic on Äeg,jJops. I!.. UstiLagotritici. can, .I. Bot. 63 t 765-i71,
Nietsen, .T, 1987a. Races of ustiLago tritici and bechniques for theirstudy, Can, ,f , Plant pathol .9:9L-105,Nielsen, !T. 198?b, Reaction of. Hordeum species to the smut fungiUstiTago nuda and û. tritici. Can. J, eol. 65: 2024-2027,
Nielsen, ü, 1988, UstiTago spp,, smuts. .Adv. p1ant, pathot, 6: 483-490,Nielsen, !t. and Dyck, p.L. 1998. Three improved differential hosts toidentify races of ustiJ-ago Eritici, catt, 3. plant pathol . !0t 327-331.Nishikawa, K. and Nobuhara, M, 19?1, cenetic studies of alpha-amytaseisoz¡rmes in v¡heat.. l. Location of genes and variation in tètra- åndhexaploid \{heat. Jap. ,f . cenet. 46; 345-353,
Ol:lns, R.E. and Bever, W.M. l9SS. T1æes of seedling reaction of Kawvaleand wabash winter wheae to chree physiorogic raceÁ of IJsëi|ago tritici.Phytopathology 45r 513 - 516

152
Ohms, R.E. and Bever, W.M. 1956. Effect of time of inoculation of wintert1¡heat wi¿h UstiJ-ago tritici on the percentage of ernbryos infected andthe abundance of h],T)hae. phytopathology 46: 157-158.
Oort. A.J,P, 1939. Inocutation experiments with loose smuts of wheat andbarley (UsÈj-Zago trit.ici and U. ¡uda) . phytopathology 29t .1L7-j28.
Oort, À.¡I.P. 1944. Studies on loose smut. II. Hl4)ersens i ¿ivi ty of wheatto loose smut (usÈj-zaqo tritici) [onderzoekingen over stuifbrãnd rr.Overgevoel igheid van Tarwe voor etuifbrand (UstiLago trit.ic.i). Tijdskr.o. Plant.enz. 50: ?3-106, 1944.1 Rev. Àpp1 . Myc. 35: 160-161,
Oort, A.,t.P. 1963. À gene-for-gene relationship in the ?rj tj cum- us tiLagosyseem, and somè remarks on host-pathogen combinations in general . Neth.,t. P1 . Path.69:104-l-09.
OrCh, R.A, and Bushuk, W. 19?4. Studies of glutenin. vI . Chromosomallocation of genes coding for subunits of glútenin of cornmon wheat.Cereal Chem. 51: 118 - 126,
Palme, P.L L987. cenetics of wheat storage proteins and the effect ofallelic variacion on bread-making quality. À;n. Rev. plant physiol , 3g:141.-153,
Pa]me. P.L and Corfield. R.c. 19?9. Subunit composition of wheatglutenin proteins, isolated by ge1 filCration in a dissociating med.ium.Planta 145:83-88.
Palme, P.L and lJawrence, c.J. 1993, catalogue of alle1es for thecomplex gene Ioci, clu A1, clu 81, and clu Dl which code for highmolecular weight subunits of glutenin in hexaploid wheat. CereaÍ Res.Conm. 11r 29 -35.
Pa]me, P.I. and Rhodes, À,p. 1982. Cereal storage proteins: structureand role in agriculture and food technology eenéCið aspects, processing.Encycl , Plant Physiol . l-4A.¡ 346-369.
Palme, P,f., Iraw, C.N, and Mud.d, E.E. 1980, Controt by homoeol_ogousgroup 1 chromosomes of the high moteculax weight subunÍts of glútenin, amajor protein of wheat endospérm. Theor. Appll Genet, 59: 1-L3-L20.
Palme, P.f., Ho1t, L.M. and, taw, C.N, 1991, Structural and genetical:!gdig" on the hiqh motecular weight subunirs of wheat qluténin. 1,A,1Ielic variation in subunits amongst varieties of wheai (friticumaestivum), Thêor. .App1 . cenet', 60¡ 229-236.
Patmê, P.I., golt, L,M., WorLand, À,,t, and IJa$r, C.N, 1992. Structuratand gene¡ical studies on the hiqh molecular weight subunits of wheatglulenin, Part 3. Telocentric mapping of the subunit genes on the 1ón91lT"_9I the honoeoloqous group 1 chromosomes. Theor. Appl . Genet. 63:129 - 1,38 .
Palme, P,I., ,Jackson, 8.A., Holt, L.M, and Law, C,N, L984. ceneticlinkage between endosperm storage protein gene; on each of the shortarms of chromosomes l-A and 18 in wheat. Thèor. Appl . ceneÈ. 5Z: 235-243,
Person, C. 1959. Gene-for-gene relationships in host-parasite systems.Can.,.f. Bot, 37: 1101- 1130.
Petchey, 8.M.. Koebner, R.M.D, and. caLe, M.D. 1990. ceneticcharacteri zalion of a further homoeoallelic series of grain est.erase1oci, Est-6, jn wheat'. Theor, AppI . cenet, 79! 294-296,

153
Píekenbrock, P, t927, Invest.igations on ¿he behaviour of. Ustijaqotritici t.owards variet.ies and hybrids. funtersuchungen über dasverhalt.en des UstiJ.ago tTitici an Sorten und Krenzungen. Kuhn Arch.15:411-456 (f927].1, Rev. Appl . Myc. 7: 435.
Poehlmann, J.M. 1945. A simple method for inoculating baxley wieh toosesmut. Phytopathology 35t 640-644,
Popp, W. L95L. Infection in seeds and seedlings of wheaL and barley inrelation Co development of loose snut. phyCopatholoqy 4!; 26!-275.
Popp, ry. 1955. À comparative sludy of spore germination of UstijagoEritici and U. nuda. phytopathology 45: 585-590.
Popp, W. L959, A new approach co the enìlcryo test for predicting loosesmue of wheat in adult plants. phytopathology 49t 75-I7.PugsLey, À.T. 1953. THe resistance of White Federation 45 and Ðundee 4gwheat to Ustilago tritici (loose smut of wheat). J, Àustr. Inst. .A,gric.Sci . 19 t 238-240.
Ribeiro, M.A.M.do vaLl.e 1963. The inheritancè of the resistance of wheatto two races of UstiTago ¡uda. Trans. Br, Mycol . Soc. 46: !79-!92.Richards, H.R. 1961. fnheriÈance and expression of loose smut resistancein wheaC. Diss. .Abst. 2Lt 7696.
RowLand, G.c. and Kerber, E.R. 19?4, Telocentric mapping in hexaploidwheat of gre¡l.es for leaf ru6t resistance and other chãraóters derivedfTom AegiTops squarcosa. Can. ,J. cenet. Cytol . L6: I37-L44,Rudorf, W. and. Rogenstiel, K.V. 1934. Studies on the resj_stance of wheatvarieties to Loose smut, t/sËj-¿açro tritici, and on its inheritance incrosses [Untersuchungen übex die widertands fähiqkei t bei weizensortengegen Wei z enf lugbrand, Ustilago tritici, und über ihre Vererbung inKrenzungen. Zeitschr. f. zuchtung. !9t324-332.1 Rev. .A,ppl , Myc. 13:621,SabellÍ, P.A.¡ lJafiandra, D. and ShevJry, p.R. 1992. ReÊtriction fragmentanaLysis of ,nu11, forms at fhe cli 1 loci of bread and durum wheaEõ,Theor. Appl . Genet. 83¡ 428-434.
Sacco, F,, Tranquilli, c., corgoschidse, L. and suarez, E. Lgg2.Àminopept.idase 81 ¡ a centromere marker for chromosome 68 of wheat..cenome 35 | 26L-263,
Sambrook, ù., Fritsch, E.F. and Maniatas, T. 1989, Molecu1ar cloningr ÀLaboratory Manual , Vol . 1 to 3, 2nd ed, CoId Spring Harbor press, cól_dSpring Harbor, New york. 1626 pp.
Sa¡nborski, D..T. 1953. A mutation ín puccinia recondita Rob, ex. Desm, f.sp, trj¿jcj to virulence on Transfer, Chinese Spring X AegijopsunbefTulata Zhuk. Can. ,J. Boe. 4!t 4i5-4i9.Sanchez -Monge, R., Barber, D., Mendez, E,, carcia-Olmedo, F. andSaLcedo, c, 1986, cenes encoding alpha amylase inhibitors are 1ocabed inthe short arms of chromosomes 38, 3b and 6¡ of wheat (Triticum aestjvumL.). Theor. Àppl . cenet. '12¡ L08-1!3,
Sanghi, A.K. and Baker, E.p. 19?4, Monosomic anafysis of genesconditioning stern rust resist.ance in two conmon wñea¡ cultivars, Can. ,J.cenet. Cyt.ol . !6¡ 28f-284,

1,54
Sanz, J.C,, Hueros, c,, Go zalez, ¡1.M. and ,Jouve, N. 1998. Chromosomallocation by F1 monosomic analysis of endoclperm proteins in bread wheaJ.2. Two dj-mensional fract.ionaeion of qliadins. fheor. Àpp1 . cenet. 761933-940,
Sa\,¡hney, R.N., Sínqh, D. and ,foshi, B,C. 1991. Monosomic anal_ysis ofg-enes for resistance against stern rust puccjnia graminis race-s in breadwheat. Theor. Appl . cenet. 59: 313-316.
Schafer, ,t.F. L987. Rusts, smuts and powdery Mildew. paqes 542-584 j¿E,G. Heyne, ed. Wheat and Wheat Improvement, American Sóciety ofÄgronomy, Inc. , Madison.
Schaller, C.W, 1949. Inherit.ance of resj-slance to loose smut, usti_Zaqonuda, in barley. phytopathology 39t 959-979,
Schmidt, ,t.W. 1973. Breeding and genet.ics. pages g-30 jn c.E. Ing1ett.,ed. Wheat: producCion and Utilization, The Ã,Vi publishinq Co., wéstpo;t,Connecticut,
Sears, E.R. 1969, wheat CytogeneCics, Ann. Rev. cenet. 3: 451-468.
Sears, E.R., Loegering, W.e. and Rodenhiser, H.A. 1957. Identificationof chromosornes carrying genes for scem rust resisgance in four varj-etiesof conunon wheat., Agronomy Journal 49¡ 208-21-2.
Sharp, P.,t., Desai, 9., Chao, S. and caLe, M.D. 19gga, Isolation,charac leri za t.ion and applications of a set of 14 RFLP probes ideíbifyinqeach homoeologous chromosome arm in the Triticeae, paqès 639-646 in ô,f .Milfer and R.M.D. Koebner, eds, proceedinge of the Seventh fnternationalWheat cenet'ics Syrnposium, Institute of p1ãnt Science Research,Canìlcridge.
Sharp, P,J,, DesaÍ, S. and ca1e, M.D. 1948b. Isoenzyme variation andRFLPS at. the bet.a-amylase loci in whèaL, Theor, Àppl . cenet.. Z6: 691_699.
Sharp, P.!t,, Chao, S., De6ai, S. and cale, M.D. 1999. Thê isolation,characteri- zation and application Ín the Triticeae of a set of wheat RFLPprobes identifying each homoeologous chromosome arrn. Theor. Àpp1 . cenee,78:. 342-348,
Shepherd, K.w. 1968, chromosomal cont.rol of endosperm proteins in v¡heatand rye, Pages 86-96 jn K.W, Finlay and K.W, Shepherd,-eds. eroceedlngsof the Third International wheat cenetics Sl¡mposium, Àust, Âcad. Sci,;Canberra .
ShesÈakova. À.P. and Vjushkov, A.À. 19?4. Inheritance of spring wheatresist.ance to loose smut ttsÊj.Zaçro tîitici (pers.) ,fens. tc;netíka t0;17_241 . Biol . Abstr, 6O: 4?54.
Shewry, P.R. and. Míflin, B.¡J. 1995. Seed storage proteins ofeconomically important. cereals. pages 1-93 j¡ Í. pomeranz, ed, Âdvancesin Cexea1 Science and Technology, Ámerican .A,ssãciation of CerealChemists, St Paul, Minn.
Shewty, P,R., HaLford, N.c. and Tathan, A.S. 1992. High molecular weightsubunit.s of wheat glutenin. J. Cereal Sci. 15: 105-120.

155
Shinohara, M, 1976. Behaviour of UstiLago nuda (,tens.) Rostrup and U.tyitici (Pers.) Rostrup in their host tissues. Rev. p1ant. prót. Res. 9:r24 - 1,42 .
Singh,.N.K,, Shepherd, K.w,. IJangridge, p,, cruen, L.C., Skerrit.t, ,7.H.and Wricley, C.W. 1988, rdenlificatiòn of fegumin like proteins in\,/heat. PlanÈ Mo1 , Biot. 11 : 633 - 639 .
Skerritt, ,t.H, 1991. Ä simple antibody based test for dough strength. I.Development of method and choice of antibodies. cereal ch¿m, 69: ¿6?-4'14 .
Skeffiet, ü,1¡, and Hill, A,S, 1990. Homologies between grain storageprot.eins of different cereal species. 2. Eifects of assáy format aádgrain extractant on antibody cross reactivity. .1. Cereal Sci, 11 ¡ 123-L4! .
Skerritt, ,J,H, and Ler.r, p.y. 1990. Homologies between grain storageproteins of different cereal species, l_. Monoclonal ancibody ,"."Éio.,'i,¡i th total protein extracts, ,.f. Cereat Sci. 11: !03-12!.Skerrítt, ,t.H, and Robson, IJ.G. 1990, Wheat low molecular weighCglucenin subunits structural relacj_onship to other glucen proÉeinsanalyzed using specific antibodies. Cereal Chèm. 67; 250-257,
Skerritt, ,J.H. and Underwood, p,À, 19g6. Specificity characteristics ofmonoclonal ancibodies to wheat grain storage proteins. Biochim. Biophys,Act.a 874: 245 - 254 .
Skerritt,, iI.H,, Martinuzzí, O, and l{riqLey, C.I¡I. 1997. Monoclonalantibodies in agricultural testing¡ quántitation of specific \,rheat
TåöiUt"" affectêd by sulphur deficiency. can. J, planr. Sci. 67t L2!-
Skerrit.t, ,t.H., HiLL, À,S. and McÀdan, D,p, l-990. fnmunology invadeschemistry: Testing cereals with antibodies, Chemistry in aúËtrafia(Sept.): 297 -300,
Skerritt, ,J.H., MarEinuzzí, O, and Metakovsky, E,V. 1991. Chromosomalcontrol of wheat gliadin protein epitopesr analysis with specificmonoclonal an¡ibodies. Theor. Äppt, cenet. 82r ¿4-53,
Smith, D.8.. tister, p.R., and Hanson, p.R. 1996. Discrimj-nation ofbarley varieties by electrophoresis of end.osperrn prot.ein extractabieinto a mixture of sodium dodecyl sulphate 2 -mercaptoethanol_ , anddime thyl formamide. ,-t, Cereal Sci, 4; !07-LL6.Sorrells, M.E. 1992, Development and application of RFLps in poll¡p1oids.Crop Sci, 32: L086 - 1091,
StatistícaL Ànalysis System fnstitute, 1989, SAS procedures cuide,Version 6. SAS lnstitute, lnc., Cary, N.C, ?05 pp.
S!t, l¡. and Dvorak, J. ]-992. Chromosornal location of adenylate kinase, 6phosphogluconate dehydrogenase, and glutamate pyruvate trãnsaminasestructural loci in wheae¡ barley, and t ophopyrun eLongatum, cenome 35t147 - 154 .
Szurmak, B. and Dobrzanska, M, 1993. .A Large ÐNÀ repeat of thelignersion pattern conunon to rr,heat and rye genomes. plant Mot, Biol, 21:9t9 -927.

156
TaLbere, IJ.E. and Clack, T.H. 1991. Identification of D-genomechromosomes oÊ Chinese Spring on Southern Blots using genome-specificrepe¿itive DNÀ. .T. Hered, 82: 509-51,2.
Talbett, Ir,E,, Magyar, c.M., Iravin, M., Blake, T.K. and MoyLan, S.Ir,1991, Molecular evidence for the origin of the s-derived. gènomàs otpollæloid Triticum species, .Amer, !T. Bot. 7g: 340' 34g.
Tapke, v.F. 1929. Influence of variecal resistance, sap acidiEy, andcertain environmental factors on the occurrence of loose s¡nut in wheat..f. Àgric. Res. 39: 3l-3 - 339.
Tapke, V,F. 1931. Influence of humidity on ftoral infection of wheat andbarley by loose smut. !T. Agric. Res. 43: 503-516.
Tapke, V,F. 1948. Environment. and the cereal smuts. Bot, Review 14r 359-4r2 .
Tathan, 4.S,, Shev,ry. P.R. and Belton, p.S. 1990. Structural sEudies ofcereal prolamins, including wheat gl-uten. Adv, Cexeal . Sci. Technof. 10:L-7B.
TayLor, J,W, f927, Effect of continuous selection of sma1l and larqewheat seed on yie1d, bushel weight, variet.al purity and loose smutinfection.,f. Àmer. Soc. Ä,gron. 20: 856-86?.
Thiele, v. and MeLz, c. L992. chromosomaf location of genes controllinglactate dehydrogenase in rye, wheat, and barley, Cenome 35: 32-34.
Thiele, v, and Seidel, À, 1990. chromosomat location of a calalase genein wheat using rye-wheat-additions. plant Breed. Z. pf.Lar.zer.zucht 105:'18-79.
Thomas, P.L. 1978. potential for natural hybridizat.ion between strainsof Ustifago nuda, Can. !T. Bot, 561. 721,-.126,
Thomas, P.L. 1984. Recombination of virulence genes followinghybridization bêEween isolates of Ustilago nudå infecting bailey undernatural conditions, Can. J. plant pathol . 6: 101-104.
Thomas, P.f,. and. MetcaLfe, D.R, L994. Loose smut resislance in twointroductions of barley from Ethiopia. Can, ,f. p1ant. Sci, 64t 255-260.Tingey, D.C, and Tolman, B. 1934. Inheritance of resistancê to loosesmut in certain wheat crosses. ,f, .Agric, Res, 49: 631-655.
Tragoonrung, S,, Kanazin, V., Hayes. p.M. and B1ake, T.K. 1992.Sequence- tagged- site- f acilit.ated pCR f or barley genome mapping, Theor.Ã.pp1 . cenet, 84: 1002 - 1008.
Tropova, À.T. 1938. Surnmary of the scientific research work of CheInstitute of plant protection for the year 1936. part I. pests anddiseases of cereals and sheller belts. lpub] . Off. pan-Sov. V. I. LeninAcad, agric. Sci. Leningrad 1937.1 Rev. Appl . Myc. L?; 434.
UhLen, A.K. and, Ringlund, K. 199?. Gene dosage effects on storageproteins in wheat (Tîiticum aestjvum). .I. ceieal sci. 6r 21,9-22á.
Vaag, P. and Munck, IJ. 1987. Immunochemical methods in cereal researchand gechnology. Cereat Chern. 64¡ 59-72.

L57
Waud, 'f,,L. and Metzger, R.,t. 1970. Inheritance of a new factor (Btg) forresistance to cofiunon bunt in wheat. Crop Sci, j-0¡ 'lO3-704.
Whê14n, E.D.P, 1988, Transmission of a chromosome fron d.ecaploidAgropyron elongatum that confers resislance to the wheat curl mite inconmon wheat., Genome 30: 293-298,
WheLan, E.D.P. and Lukow, O.M. 1990. The genetics and qliadin proteincharacteristics of a wheat-alien Cranstocation that confers reÈistanceto colonization by the wheat curl miee. cenome 33r 400-404.
Wiese, M.V. 1987. Compendiurn of wheat Diseases, AmericanPhytopathological Society, St. pau1, Minn. 106 pp.
WÍl-1ians, C.E. and Sb. Claj-r, D.A. 1993. phenetic relaCionships and1evels of variability detected by restriction fragment lengthpo1]¡moxphism and random amplified pol_t¡morphic DNA. analysis of cultivatedand wild accesgions of Lycopersicon esculentum. cenome 36: 619-630,
WrigLey, C,w, and. Shepherd, K.W. 19?3, Electrofocusing of grain proteinsfrorn wheat genotlæes, Ânn. N,y. Acad, Sci. 2Ogt LS4-f62, -
Yen, Y. and Baenziger, p.S, 1992, A bêcter way to construct recombinantchromosome lines and their controls, cenome 35¡ g27-g30,
Zawistowski, ,t., .Ansell, M., Zar,Jistowska, U. and. Howes, N.K, !gg2. Amonocfonal ant.ibody to endogenous alpha amylase inhibitor of barley. ,f.Cereal Sci, L5: 1-4.
Zeven, À.C. and Zeven-Hissink, N.C, 1975, cenealogies of 14000 wheatvarieties. centre-NcC, Wageningen, The Netherland;. 119 pp.

158
APPET{DIX A
TNTRODUCTION
This appendix lists a series of lines inocutated with up to eight racesof U. trit.ici. The data is presented mainly for referei.ce pur;oses,Ã.lthough, some unusual findings are poj-nted out in the discuss-ionsection to follow. These results weie not included. in a separatechapter in the body of che thesis because the daea as it stãnds was notconsidered strong enough for any conclusions to be made.
MÀTER]ÀLS ÃND METHODS
Lines listed in the tables of this appendix were grown and inoculated asdescribed for lines in Chapcer 4, The chinese sp;ing -.AegrjIopsunbelTuLat.a (CS-U) ,and - Hordeum chiLense (CS-Hch) tines were- provid.ed byDr, T.E, Miller (Cambridge Laboratory, Cotney Lane, Norwich, üx,), tn"ChÍnege Sprirlq-.Agropyron internedium (CS-Ai) lines \¡¡ere provided by Dr.R.A. Mclntosh (University of sydney, plant Breeding Institute, cobÈicty,NSW, AustTalia), the Chinese Spring-Hope (CS-Hope) substieution lineswere those of Dr, .I. Kuspira, maintained and provided by Ðr. p.L, Dyck(Àgricutcure canada Research stat'ion, winnipeõ, ¡¿anitobã), the Cadel andRescue linès were provided by Dr. E.D.p. Wfrè1ãn (¡qrjculture CanadaResearch Station, Lethbridge, Alberta) and the loose srnut differèntiallines (TD-) were provided by Dr. .1. Nietsen (Agrículture canada neseaichStation, Winnipeg, Manitoba) .
RESUTTS ÀND DISCUSSION
Table À,1 provides a list of reactions of all the Cadet cytogeneticstocks evaluated during this sCudy. The most significant reÃu1ts werepointed out in Chapter 4. However, a few things are worth noting. Theprogeny of the double monosomic 6À/6Ä9 population inoculated to ;ace T19were híghly susceptible providing evidencè that the T19 resistance isrecessive. Substitution of the wheat chromosomes 4A, 4D and 5D with thealien chromosornes 44,g, 4Ag and 5Ag respectively did not affect tt. i..ruiof xesistance of Cadet to any of the races test.ed. Substj.tution of the6À chromosome with the 6À9 in addition to affecting resistance to i19also seemed to affect the leve1 of expression t.o race TL5. Whereas, allother Cadet. lines were conpletefy resistant to T15 Cadet 6Ag(6À) shòwedL3% loose smut. This is an indication that loose srnut resiÁianâe maywork jointly to produce cumulative effects in some cases.
Table A,.2 summarizes data of races inoculated onto various Rescuecytogenetic stocks, There were no interesting reactions that made anysense, Three reactions that did not make sense were the lack ofinfection in Rescue 6Àg(6A) Co racè T2t Rescue 6A9(6D) to race T2 andRescue 6A9(6D) to race T31. In general Rescue and the Rescue linesproduced quite variable infection responses both among and \a,ithin race-line conìlcinalions (see standard devialions in Table À.2 compared to, forexample, the standard deviati.ons for the U. tritici race diifereneíálsin Table À.3). Incon€istent inoculation may be responsible for thesàextreme deviations. Ànother explanation is that treierogenelty oå-uisamong the Agropyron chromosomes, however, lhese usually-derivè from onestock. Alternatively heterogeneity in Rescue may be aiotherexplanation.
Inoculations of the differentials were done bo verify race reactionsand the results for the most part reftected the expectèd react.ion (Table

159
Ã'.3) (Nielsen 1987). Discrepanciès which should be noted wereresislance Eo T15 by Red Bobs (TDI-O), moderate susceptibility of Kota(TD4) to race T19, suscept.ibility of TD3 to T31. The reactiòns of RedBobs (TD10) and Kola (TD4) can bè explained based on heterogeneity ofthe sample. The genetic research was initiated on Red Bobs and Xo¿abefore consideration had been given to the use of the differentials asthe genetic studies. Fox these two lines Ehe inbred differential wasnot used for race evaluation, but' insLead the original variety had beenused. The problem of heterogeneity was noticed too late Ín t.ñe studyfor re-evaluations of lines to be performed. However, the consisten¿yof reactions of the oeher differenLials to Che races used in this stuáycornpared to the reactions expected based on Nielsen (1997) wouldindicate the races axe correclly classified, There is no explanat.ionfor the resistance in TD3 to race T31.
À number of other lines were inocufated to evaluate leve1s ofresístance for a variety of reasons. sorne were inoculated forevaluation as suscepts, some as sources of xesistance. Some wereinoculated as cyt.ogenet.ic stocks to see if chromosome variation affectedthe 1evel of resistance. À series of lines were evaluat.ed to determinewhether or not Canehatch resistance was due to foose smut resistancebeing carried with stem rust resistance during the development ofCanthat.ch or to see if the recurrent parent Thaccher was heterogeneousfor resistance.
Canthatch received from Dr. Eric Kerber (Ägriculture Canada,Winnipeq) , Canthatch RL2935, Eqypt Na1O1/Mqi"i/L2, FKN pre1ude16 sr7,Hope 48 saR, Kenya Farmer, Lee6/KF, prel 4 Sr7b, Red Thatcher, fci/KF,That.cher and Thatcher RLL945 were inoculated with race T39. The commonefement of several of thesè lines is the possession of the Sr? stem rusLresistance gene aC vari-ous levels of backèrossing. The Sr? gene wasintroduced inho Canthatch or derivat.ives of That¿her, the báckcrosslines were evaluated to determine if xesistance ho racè T39 wasassociated with Sr7 and ttte different. sources of Thatcher werej-noculated t.o determine if heteroqeneity existed within Thatcher, The1ow react.ion of Canthatch relative to Thatcher to race T39 (Tab1e À.4)waa what. spurred the study, The Eric Kerber Canthatch is a single plangselection which subsequently was furthex inbred several generatíonsl alook at Canthat'ch RL2936 sho\À'ed a higher reacEion to T39 with a highstandard deviation. The resul¿s were based on up to five inoculatãdheads from up to five different plants. These rèsults suggêstheterogeneity, Comparison of three Thatcher lines Red fhálcher,That.cher and Thatcher RLL945 also indicaeed heterogeneity when thevariation in reaction to T39 was compared (Table À:4). Êvidence thatloose smut resiseance was not carrieã with sr? resistance was seen whenKenya Farmer was evafuated showing 49% incidence of susceptible plantsto race T39. Kenya Farmer was the source of Sr7 resistan¿e in -Canthatch.
- comparing . C! inese Spring 1U(1À) , 1U(18) , 1U(1D) to Chinese spring
showed no difference in resistance to seven of the eight races-.However, reaction to T6 may be Bomewhat differenl whièh again may be dueto heterogeneity of Che lines ox Che 1U chromosome may carry someresiscance to loose smut. .Analysis of ¿he group ? cyiogeneiic stocks ofchinese spring is.complicated by the tac¿ thae -ene clj-nese spring usãato produce the alien cytogenetic stocks was not provided, rñereiorethree Chinese Spring sources were used as controis. Chinese Spring (EK)received fxom Dr. Eric Kerber, Chinese spring (pI sc) received. froñ tneDeparlment of plant Science, Univexsity of Manitoba and Chinese Spring(NH) provided by Ðr, Neil Howes \,¡ere uèed as controls. These stoóksdiffered sonewhat in there resistance t.o T6 bub were most diffexent inxeaction to T15, whereas, Chinese Spring plsc and NH qrere susceptibleto T15 Chinese Spring EK was resistant. Comparison of the group 7cytoqenetic stocks indicat.es that there may Ëe resist.ance cárriãA inthese chromosomes in chinese spring. Chinèse Sprinq 7Äi (7D) was

160
susceptible to T39 and T19. However, none of the other chromosome sevensubsCitution lines were susceptible indicating thaÈ the aljenchromosomes 7U and THch carry resistance or an er¡or occurred in thedevelopment of the Chinese Spring 7Ai (7D) line. a similar response ofChinese Spxinq 7Ai (78) to T6 and ?Ai (?B) to T2 and T6 occurred ashappened with chinese spring 7Ài (?D) \r¡i th lhe same explanation,However, it is odd that the 7À and 7B substitution tines weresusceptible \a'hile the 7D substitution line was resistant whensubstituted with 7Ài, indicating Ehat the Chinese Spring used here mayhave been susceptible to T6 and that the 7D substitution line was mis-inoculat.ed. Considering resistance to race TL5 rnosC of the gxoup ?stocks were susceptible, indicating that Che original Chinese Spxing wasalso susceptible. The 7Àí group of chromosomes may be an exception,where only Chinese Spring 7Äi (7Â) was susceptible, This and Chesusceptible reaction of Chinese Spring ?Ai (?A) to T2 was the strongestevidence that a gene for lÕose smut rèsistance in Chinese Spring residesin chromosome 7À, The fact that Dhitaphichit and coworkers- (1999)reported resÍstance in Hope in chromosome 7À and which was confirmedhere supports Che fact that Chinese Spring could and Hope does haveresistance in 7À,, À cross between chinese Spring and Hópe would have tobe made Co determine if Lhey have the same gene, alleles of the samegene or a differene gene. It is interestinq thae Chinese Spring Srg wassusceptible to several xaces to which even Chinese Spring plsc ;asresistant. Because the Srg resistance is in chromosóme 6a it ispossible that. incorporation of the Srg resistance also resulted. in theíncorporation of a loose smut susceptible alleIe in 6À or that yet'another genotl4)e of Chinese Spring was used to develop the Srg 1ine,
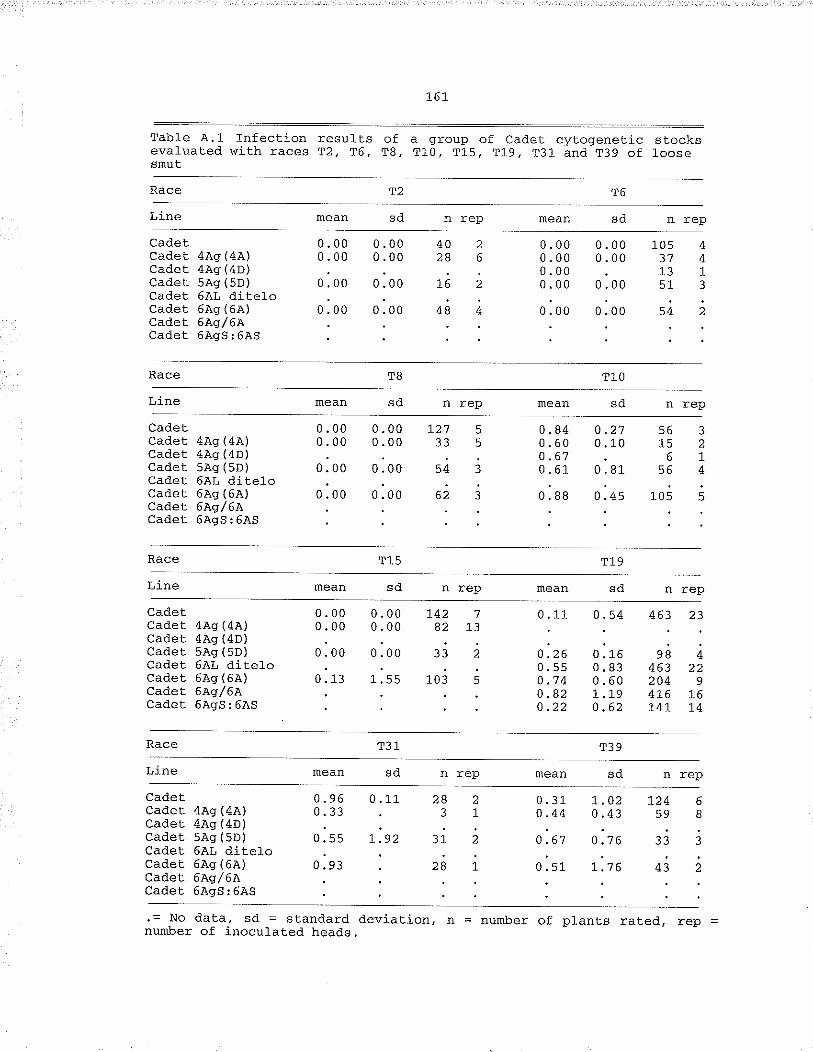
161
Table A.l- fnfection results of a group of Cadet cytogenetic stockseval-uated reith races T2, î6, T8, T10, T15, T19, T31 and T39 of 1oosesmut
Race
Line n rep n rep
CadetCadet 4Aq (4¡{)Cadet 4Aq (4D)cadet 5Aq (5D)Cadet 6ÄL diteloCadet 6As (64)Cadet 6À916Ä'Cadet 6À95 | 6À5
0.00 0.000.00 0.00
0.00 0.00
0.00 0.00
0,00 0.000.00 0,000.000.00 0.00
0.00 0.00
::
40228616 2
4ö ;
r.0 5371351
54
:
Tt0
Line n rep mean n rep
Cadetcader 4AÛ (4A)Cadet 4Àq (4D)cadet 5Àg (5D )Cadet 6ÀL di telocadet 6À9 (6Ã)Cadet 6Agil6ÀCadet 6A95:6A5
0.00 0.000,00 0.00
0.00 0.00
o.oo o.oo
0. 84 0.2'10.60 0,100.670.61 0.81
0.88 0.4s
s6 3-15 Z
6ts6 4
1-05 s
33 5
543623
T15 T19
Line n rep nean n rep
CadetCadet 4Àg (4À)cadee 4Aq (4D)Cadet 5Aq (5D)Cadet 6ÀL diteloCadet 6À9 (64)CadeL6À9164Cadet 6A95 | 6A5
0.00 0.000.00 0.00
0,00 0.00
0, L3 1,55
0.11 0 .54
0.26 0.16 98 40.5s 0.83 463 220,74 0.60 204 90 . 82 L .1,9 476 !60.22 0 ,62 141 1-4
1,42 782 13
103 5
Race T31 T3 9
Line n rep n rep
CadetCadet 449 (4À)Cadet 4Ag (4D)cadet 5Ãq (5D )
Cadet 6ÀL di teloCadet 6A9 (64)Cadet 6A9164Cadet 6AqS:6A5
0.96 0,11
0,55 r.92
0,3L 1.020.44 0,43
0.6'1 0.76
o,sr r'.76
r24 659 I33 3
4åi
28231
31 2
28i
.= No dat.a, sd = standard deviation, n = nunùoer of plants rated, rep =nunber of inoculated heads.
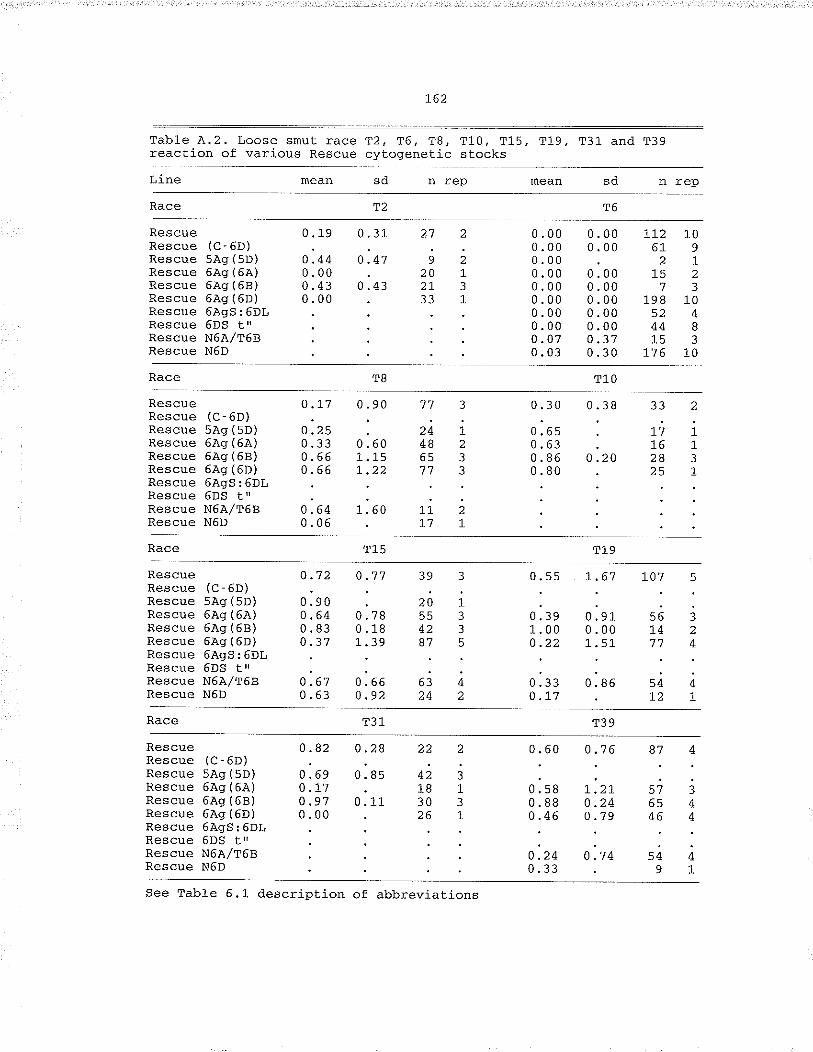
1,62
Table ,A..2. Loose smut race T2, T6, T8, T10, T15, T19, T31 and T39reaction of varj-ous Rescue cytogenetic stocks
mean sd n rep mean sd n rep
RescueRescue (C - 6D)Rescue 5Ag ( 5D)Rescue 6A9 ( 6A)Rescue 6À9 ( 68)Rescue 6Â,9 (6D)Rescue 6À95:6DLRescue 6ÐS t"Rescue N6A/T6BRescue N6D
0.19 0.31
o .44 0 .470.000,43 0,430.00
0.00 0.000.00 0.000.000.00 0.000.00 0.000.00 0.000.00 0.000,00 0.000.07 0.370.03 0.30
2',l tL2 106L9
15 273
198 1052444815 3
L76 10
9220L21, 333 r
T10T8
RescueRescue (C- 6D)Rescue 5Àg (5D)Rescue 6A9 ( 6A)Rescue 6A9 (68)Rescue 6A9 ( 6D)Rescue 6À95:6DLRescue 6DS t¡lRescue N6A/T6BRescue N6D
0,r7 0.90
0.33 0,600.66 l-.1s0.66 t.22
0,30 0.38 33
0.65 L70.63 160,86 0.20 280.80 . 25
77
244865'77
0"64 r.eo ri0.06 17
Race T15 T19
RescueReEcue (C - 6D)Rescue 5Ag ( 5Ð)Rescue 6A9 ( 6A)Rescue 6Â9 ( 68)Rèscue 6Â9 (6D)Rescue 6Ã,95:6DLRescue 6DS t'rRescue N6À,/T6BRescue N6D
0.72 0.7'1 39
0.9 0 200.64 0.78 550.83 0.18 420 ,37 1.39 87
0.55 r .67
0.39 0.911.00 0.000 .22 1.51
3
i33
L07
0,6'1 0.660.63 0,92
0.33 0.860.L7
56 31,4 277 4
544L2 1
aãl¿242
Race ?31
RescueRescue (C - 6D)Rescue 5Àg ( 5D)Rescue 6Aq (6.4)Rescue 6A9 (68)Rescue 6A9 (6D)Rescue 6A95:6DLRescue 6DS t,'Rescue N6A/T6BRescue N6D
0.82 0.28 22
0,69 0.85 420.17 180.97 0.11 300.00 26
0.60 0.'16
0.58 I.210.88 0.240.46 0.'19
0.24 0.740.33
87
siä6s 4n1
:54491
see Table 6.1- description of abbreviacions
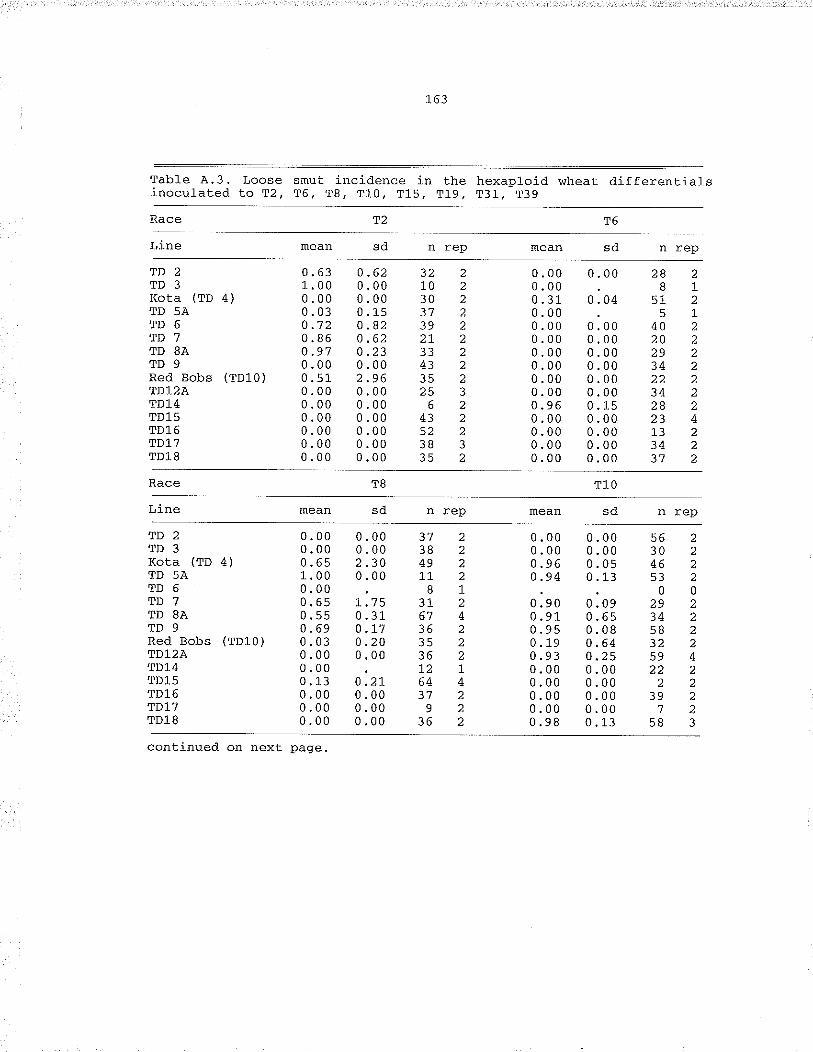
163
Table À.3. Loose smut. incidence in the hexaploid wheaL differentialsf"" ,Race T2
Line mean n rep mean n rep
TD2TD3Kora (TD 4)TD 5ÀTD6TD7TD 8ATD9Red Bobs (TD10 )TD12ATD].4TD15TD16\D!7TD18
0.63 0 .621.00 0.000.00 0.000.03 0,150.72 0.820,86 0 .620 .9'7 0 .230.00 0.000.51 2.960.00 0.000,00 0,000.00 0.000.00 0.000.00 0.000.00 0.00
0,00 0,000.000.31 0.040.000.00 0.000.00 0,000.00 0.000.00 0.000.00 0.000.00 0,000.96 0.150.00 0.000.00 0.000.00 0.000,00 0.00
103037
2r334335
643
3835
222222222
288
515
40202934223428
133437
T10
Line mean sd n rep
TD2TD3Kota (TD 4)TD 5ATD6TD7TÐ 8ÀTD9Red Bobs (TDl- 0 )TD12ATD14TD].5TD16TD17TD18
0.00 0.000.00 0.000.65 2,301.00 0,000.000.65 L ,'150,55 0.310.69 0.L'70.03 0.200.00 0.000.000,13 0.210.00 0.000.00 0.000.00 0.00
0.00 0.000.00 0.000.96 0,050.94 0,13
0.90 0.090.91 0.650,95 0.080.19 0,640,93 0.250.00 0.000.00 0.000.00 0.000.00 0.000.98 0,13
56 230 246200
29234 258 2322594tt at)
39 272
58 3
37384911
831673635361,2
379
36
4222r4222
continued on next page.

Table À.3 continued
T15 T19
Line sd n rep sd n rep
TD2TD3Kora (TD 4)TD 5ÀTD6TD7TD 8ÀTD9Red Bobs (TDL 0 )TD12ÀTÐ14TD15TD16TD17TD18
0.!2 0.2L0.88 0.380.1-6 0.040.880.00 0.000.95 0.010.00 0,000,00 0.000.03 0.150.00 0.000,16 0,370.11 0 ,320.95 0.310.03 0.160.00 0.00
0.00 0.000.88 0.310.43 t.200.94 0.430.78 0.100 .26 r .420.25 0 .470 .19 0 ,'t 90.12 0.?so .71, L.370.06 0.120.00 0.000,93 0.471.000.00 0.00
39 224242232232235 228291 4t'| 2
150 736 32]-440 220 1
81343'13438393'7353834
2839
37
422122222223222
Race T3 t-
Line mean n rep sd n rep
TD2TD3Kota (TD 4)TD 5ATD6TD7TD 8.ê,
TD9Red Bobs (TD10 )
TD12Ã,TD14TD 1.5TD].6TD17TD]-8
0,90 0.72l-.00 0,000.00 0.000.00 0.000.00o.92 0.000.00 0.000.00 0.000.97 0.261.00 0.000,00 0.000.00 0.000.00 0.000.00 0.000.00 0.00
0,00 0.00 39 20.94 0.03 3s 20.91 0.03 44 20-70 0.77 37 20.56 1.89 34 20.61 1,55 28 20.81 0.04 36 20.33 0.47 40 20.2'1 0.87 48 30,95 L.92 22 20.09 0.56 32 20.00 0.00 31 30.81 0.'1'1 36 21.00 0.00 33 20.52 1,2'7 29 2
2I
222323332
see Table l for definition of abbreviations

165
Table Â.4. Inoculatj-on of niscellaneous cultivars and cytogeneticstocks wibh races T2, T6, T8, T10, T15, T19, T31, T39
Race T2
Line mean n rep mêân
783802r.A8BerilloBiggar BSR
Bihar
cs 1u ( l-A)cs 1u (18)cs 1u (1D)
cs 7Ài (7À)cs 7Ài (78)cs 7Ai (7D)cs THch (7A)CS THch (78)
cs THch (7D)cs 7u (7À)cs 7u (78)cs 7u (7D)CS Hope 5A
CS Hope 68CS Hope 7Acs N1D/T1BCS NHCS PlSc
CS Sr8CanthatchCanehatch N7ÐCanthatch RL2936Diamant
Eql¡ptNa101/Mq12l12FKN Pre16 Sr7GfenleaH44HY368
0.00 0.000.00 0.00
0.00
1.000.00 0.000.000.00 0.000.00
0.42 0.640.000,000.00
.
0.000,000.00 0.000,00 0,000.00
0.780.72 0.460,000.00 0.000,00
0.00 26
.t7
. 1"2
75739
5
3220.000,000.000.000.0028
542
!4
6631
,1
11
535618
0.88 L70,'ìr 310.00 220,03 39
5224434469
0.00 0.00 780.00 0.00 18
0.00 720.00 33
0.00 0.00 39
0.82 0.61 38
0.000.000 .020,020.38 !.23
0.68 0.s80.05 0 .2t0.250.35
0,91 0,540.00 0.000.00
1.00 0.00 52
42
iI
2
6244
I!7
4627
"l
0.000.000.8s
0.00
0.45
35 248315 3
5;i25 140 2
0.00 0.000.00 0.000.00 0.00
continued on next page
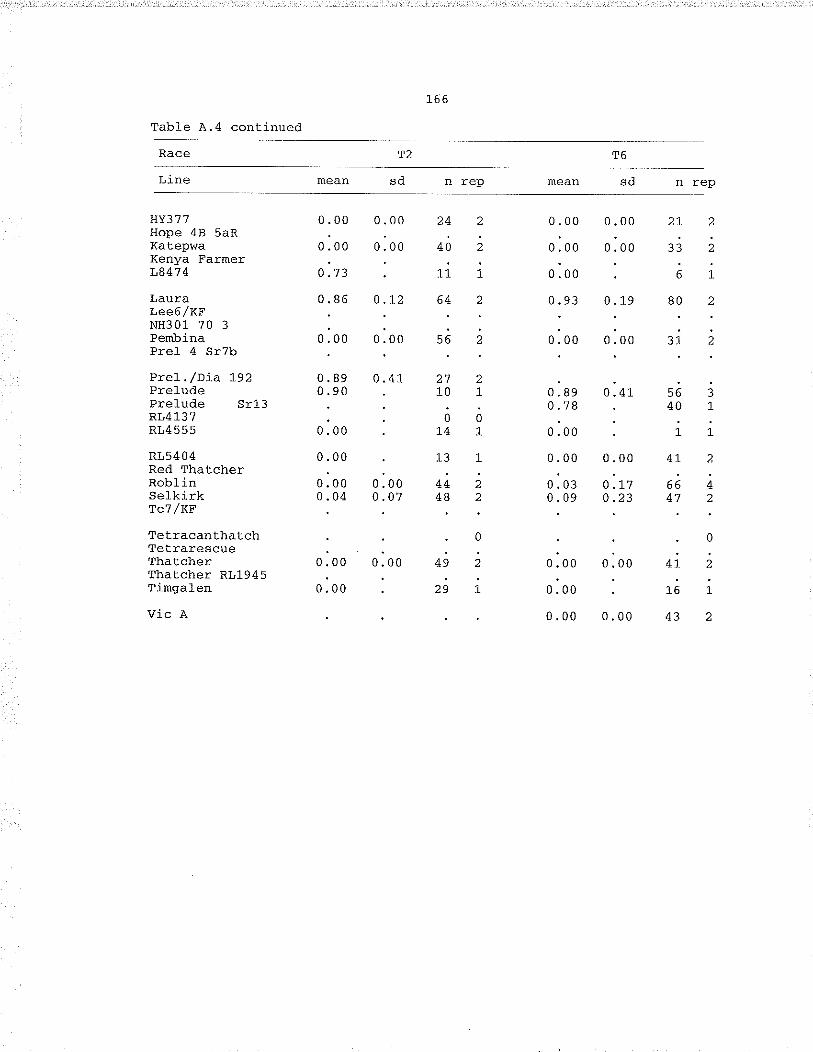
1.6 6
Table À.4 continued
T6
l,ine n rep mean sd n ïep
HY377Hope 4B 5aRKatepwaKenya Farmerr,847 4
LauraLee6 /RFNH301 70 3PenìlcinaPrel 4 SrTb
Prel. /Dia !92PreludePrelude Sr13Rr,413 7RL4555
Rr,5404Red ThatcherRobl inSelkirkTc7 lKF
TetracanthatchTetrarescueThatcherThatcher RLI-945Timgalen
Vic À
0.00 0.00
0.00 0,00
o.73
0,86 0.I2
0.00 0.00
0,00 0.00 21 2
0.00 0.00 33 2
0.00 6 l-
0.93 0.19 80 2
0.00 0.00 31 2
24
40
rL
b4
:56
2
à
i2
0,89 0.410.90
:
0.00
0.00
0.00 0.000 .04 0.07
0.89 0.410.78
0.00
0,00 0,00
0.03 0.r70 ,09 0.23
s¿ 340 1
11
4].2664472
27t-0
01,4
l-3
444A
21
ó1
L
à2
0.00 0.00
0.00
::0.00 0,00
0.00
0.00 0.00
49
29
0
;i
0
à
i2
4r
1,6
43

Table Ä.4 continued
Race T10
Line mean n rep
783802LA8BerilloBiggar BSR
Biharcscs 1u (1À)cs lu (18)cs 1u (1D)
cs 7Ai (7À)cs 7Ài (78)cs 7Ài (7D )
Cs THch (7.A)cs THch ( 7B)
cs THch (7D)cs 7u (7A)cs 7u (78)cs 7u (7D)CS Hope 5À
CS Hope 68CS Hope 7Acs NLD/T1BCS NHCS Pl Sc
CS SrgCanChat.chCanthatch N7DCanthatch RL2936Diamant
Es:yr,LNal1r /Mqr2 /72FIC.I Prel6 Sr7clènleaH44HY368
Hv37'.lHope 48 5aRKatepwaKenya FarmerL847 4
0.00 0.000.001.00
0.08
1.000.00 0.000.000.00 0.000,00
0.000.070.000.00 0.00
0,000.000,000.000,00 0.00
0.00 0.000.00 0,00
0.00
1.000.00 0.000.000.00 0,000.00
38
L7,o1,'1
283
0.00 190.00 230.00 190.00 50.00 9
0.000.000.000.000.00 0.00
31I
t7
50
32
4032
L640
875
230
9t:
tt3
L6,7
15
0.00 0.00 250,00 0.00 24
0.00 1
0,00 0.00 430.00 1-'l
0.99 0.L2 68
0.000.00
0.000.10 0.08
0.92 0.54
20132r
651
71,t2:I
748
0.000,00
41
i1
I2
38 2
42
i
21,
3
0.00
0.00 0.000.00 0.000,67 1.81
0.35 1,.87
0.00 0.00
o'-g2
0.00 0.00 440.00 230.96 0 .22 49
0.00 0.00 r7
0.59 0.61 39
o.oe . ré
¿àà44295 3
't4 3
472
13 1,
continued on nêxt page

Table A..4 continued
T8 T10
Line n rep n repmean sd
LauraLee6,/KFNH301 70 3PembinaPrel 4 SrTb
Prel . /Dia 192PreludePrelude SrL 3RL4137RL4555
RL5 4 04Red ThatchexRoblinSelkirkTc7 /Kp
TetracanthaLchTetrarescueThatcherThatcher RL19 4 5Timgalen
Víc A
0.92 0.19 0,63 1.24 63 2
0.00 0.00
1.00 0.000.98 0.19
0.940,L7
0.00 0,00
0.00 0.00o.1,2 0.48
0.00 0.00
0.91 0.220.88 0,5L0.'74
0.87
0 .47
0.06 0.000.15 1.10
81
1824
23
4343
34
i1
2
à2
442674277
237
15 1,
36240 2
0.00 0,00 59
0.79 38
0.96 0.05 53
0 ,46 l-3
continued on next page

169
Table À.4 continuèd
r15 T19
Line mean sd n rep
7838021,A8BerilloBiggar BSR
Bihar
cs lu (1Â)cs 1u (18)cs Lu (1D)
cs 7Ai (7À)cs 7Ai (78)cs 7Äi (7D)Cs THch (74)cs THch (78)
cs THch (7D)cs 7u ( 7A)cs 7u (78)cs 7u (7D)CS Hope 5À
CS Hope 68CS Hope 7Àcs N1D/TLBCS NHCS Pl sc
CanthatchCanthatch N7DCanthatch RL2936Diamant
E91ærNa1O1/MqL2l12FKN Pre16 Sr7cLenleaH44HY368
0 .00 0,000.000.68
0.00
0,610.010.000.000.00
0.87 0,420.551.000.000.00
0.900.00 0.000,000,00 0.000.00
0,140.000,330.00 0.00
0.1130
1032511
'7
1,4
2440
289844
L'7
31 219 L20 118 123 1
7L 250 211 l-
542021,!423
46420 125 1
427
0.38 0.650.060.000.62 0.400.00
0,52 0.'1r0.160.560.29 0.580.66 0.61
0.72 0 .620,000.000.'120.66
0.930.00 0.000.00 0.00
0.92 0. s1
116332L65
1,
8625346553
0.00 . 620.00 230.00 . 240.00 300.00 0.00 s6
0.00 0.000.000.000.000.00
9649
29
294823
3',? 0.95 0.30 39 2
0.000.030.79
0.00o.21,0.89
0.92 0.480.00 0.000.00
122
;
tI3
4',1 2J.L Z
89 3
o.oo o.oo ¿i0.00 180.91 0 .22 s3
continued on next page

Table A.,4 continued
T15 T19
Lane sd n rep mêan n rep
Hv3't 7Hope 48 5aRKatepwaKenya FarmerL84'7 4
LauraLee6,/KFNH30L 70 3Penìlc i naPrel- 4 SrTb
Prel . /Dia 192PreludePrelude Sr13RL4137RLA555
RL54 0 4Red ThatcherRO-b-LanselkirkIc7 /KF
TetracanthatchTetxarescueThatcherThatcher RL19 4 5Timgalen
Vic Ä
0.03 0.1-2
0.00
o'.82 ,
0.81 0.53
0.00 0.00
0.00 0.00
r. oo :
0.65 0.86
2
;i2
L8
zo
79
29
ti88
18 182436 1
35 2
77 440 2
9940
ta
8748
111,07 6
97
55 3
2
1
i2
0.00 0,00 ¿i 2 o. oo o. oo f,at 2
0.87 0.380.78 L.94
0.82
0.00 0.00
0.01 0.100.33 2.04
1.000.90 0.430.81
0.00
0.00 0.00
o.77 1 .020,00 0.00
52
1
2
42
0,000.00 0.00
0.s4
o. oo :
0.16 0,56
0.33
0.00 0.00
t36
28
012
i
continued on next page

Table À.4 continued
Race T31 T39
f,ine sd
7838021A8BerilloBigqar BSR
Bihar
cs 1u (1A)cs 1u (18)cs i_u fi-D)
cs 7Ai (?Â)cs 7Ài (78)cs 7Äi (?Ð)cs THch (7À)cs THch (78)
Cs THch (7D)cs 7u (7.A)cs 7u (78)cs 7u ( 7D)CS Hope 5A
CS Hope 68CS Hope 7Acs N1D/T1BCS NHCS Pl Sc
CanthaechCanthat.ch N7DCanehatch RIr29 3 6Ðiamant
EslætNa101/Mq12 / 12FKN Pre16 Sr7clenl eaH44HY36B
continued on next. page
o.4'7 0,¿5 380.00 14
0.00 56
0 .62 0 .620.200.77
0.00
1.000.000.000 .000.00
0.04
0.240.00
0.00 150.00 r-60.00 250.00 160.00 0.00 35
0.00 0,00 3'7
51448
I1,27
212153
503368
0.870.00 0.000.000.000.00
0.000.000.000.00 0.00
0,000.000,000.000.00 0.00
0.00 0.00
0.000,00
0.49 0.39
':0.91 0.25
0.00 0.000.02 0.130.88 0.29
. 1,70.00 34,8
0.00 16-a
313332'1a
I
1,623
9
193'72935
23,2013
L'7
úr'l
2
1L
11,
2
1
;iL
1L
1"
2
11
21"6
152
l-08123
l_0r2
49200
10 1147
43
55
452322
0.000.00
0.75 0.250.08 0.900.000.24 L. r_80.98 0.1s
0.58 0.740.48 0,830.000.00 0.000,85 0,33
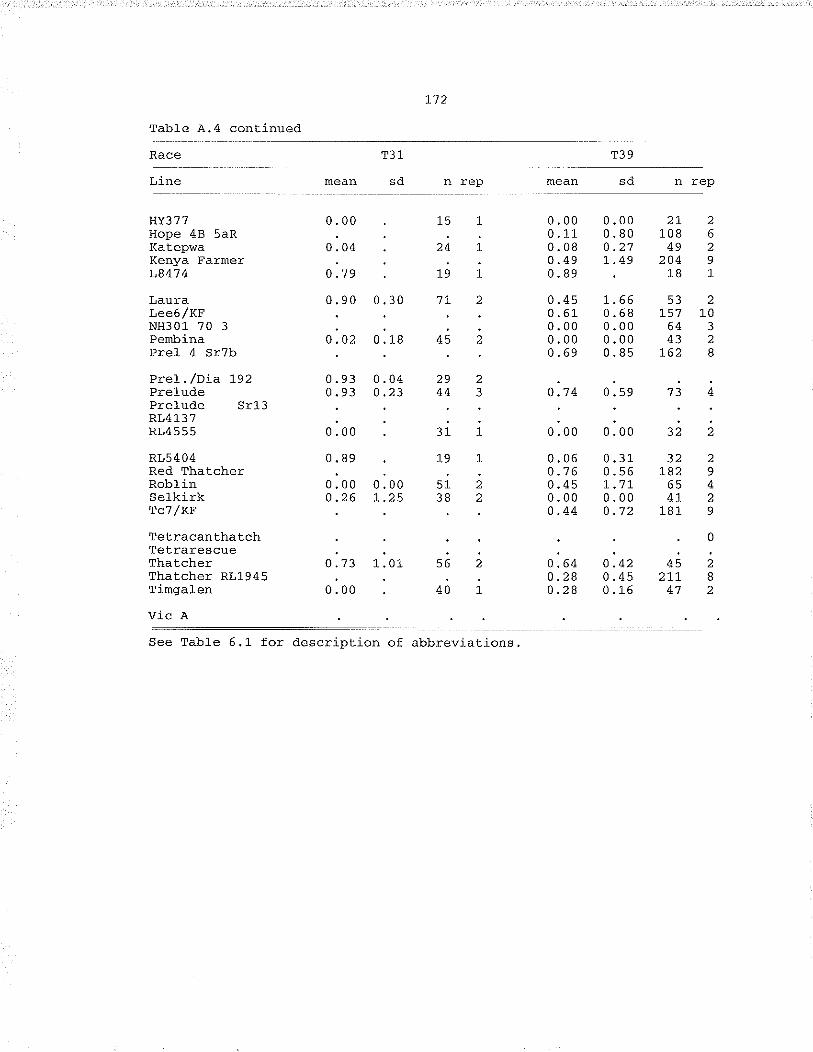
Table 'A..4 continued
T31 T39
!ane n rep n rep
Hv3'1',?Hope 4B 5aRKatepwaKenya FarmexL847 4
LauxaLee6 /KFNH301 70 3Pernlc í naPrel 4 SrTb
f,rel . /Dia !92PreludePrelude sr13RL413 7Rr,4 555
RL5 4 04Red ThatcherRoblinselkirkT.c'l /KF
TetracanthatchTetrarescueThatcherThatcher RL19 4 5Timgalen
vic À
0,00 1s
0.0¿ 24
0 .'19 19
0.90 0.30 ,:
0 .02 0.18 45
0.93 0,04 290.93 0.23 44
0.00 0.00 2! 20,11 0.80 108 60,08 0.27 49 20 ,49 7.49 204 90,89 18 1
0.45 1, .66 53 20.61 0.68 Ls7 100.00 0.00 64 30.00 0.00 43 20.69 0.85 J,62 8
0.'14 0.s9 73 4
1
ii2
0.00
0.89
0.00 0.000 ..26 t ,,25
0 .'13 1.01
0.00
31
19
o.oo o.oo ú à
0.06 0.31 32 20.76 0.56 L82 90.45 1,.77 65 40.00 0.00 4L 20 .44 0.72 181 9
.00.64 0 .42 45 20.28 0.45 2r1- I0.28 0. L6 4'7 2
5-L ¿38 2
56 2
40 r
See Table 6.t for descripLion of abbreviat.ions,

J U¡Jt\llñt¡ Iù riÐE i,-'J.Lji P,ørj.a
Hesearah Êta lonRæ8rch ErEnch, Ágnflnurê ÇånadâP,O. Bo|t03ûÈìYlt flrrÌff|, EElt(åtchéwansH 3r2
FACSIMILE rHANSfri¡TTÁ¡" HOÌIQ€
TelÈptþn€: (3{¡t)7FñÆa1FAx: (3û6F/:¡-Ð123
ÞAlg: FebEuåsy 3, 1994Ho- of F¡agce (lnch¡dhg lrrnsrnÌttål slteel) å
Tor (Nemo A Addrsså)
lytnNRCC{Í¡çñeOtÊ,Ârra
FAr( No,: (613 ) 982-?656
Fmm¡ (ltamo à Áddr€sB)
Ron KnorcAgæiculÈure CanráaAeEsa¡ch staÈionBor( f030SwlfE Curreor. Såð k.
FAx No.: (3o€)î7$9I29
Su¡¡ecî: Appreval for Ehesl6 ÞuþIlcaeion
liffiage;Thia nessage íe ín raeponse co yqur rdquest over lha tålêÞhone gbls ñornitrg !håÈI s€nd yêu Ehê forfû thal I am !Êquired b¡y thÉ alíona! LLb¡*:y of qantda Èo iillin. NoE only iE lhe¡e é lofin fËoñ Ehê Nagionèl Librèry of cånàdè þut å1so â lormt!órl Èhc Univerrfty oË ¡{àníÈobê which I mutE llll in. I havc rcac cepicB of Èheflu,ed ifl forms qa rdcll at bLânk för¡¡ç foË Þoth lnärlÈutioho,
6y way of þtckground Èhc ¡runue c:i¡rt, publiehed Ln Cenomê 1â enc,leied '.ìÞÞ1lcâÈ iêtlof ehromceene-specl I tc ¡ncnêê1êqal anribodj..s in -h-idt- geneeics' , 1992. oÈne¡no,35r831-837 by R.E. ¡tnô.:(, N.fi. llèwes ànd 1. Aunç, Thir nraauscripÈ Ëcpreâcnge oneof lix cheptrr¡ ln na Èhesis êntiElcd chlorr'otÇÊâI Loc{eion añd :inkqde Àna]'¡slsof lqese sfiuq R.oisÈôncs ù1 tdhrè-t.. For ghls Forej.on of Èhê Ehâ$it plôaaa-i.ndic¿tq approvaliT reprcar¡go¡¡ ànd dlelE:Þurlên *! indlcai€d on Èhr Nationå¡llÞrêry cÉ côa¿då ê,rd UlllycrslËy oÉ HaaitoÞè lorrnr,
I appecclate youÌ ã,sóirtanec ln this lnâgtê¡. thank you {n advansa.
Sfhcerely,npí ßnox
"Permission is granted for use of the materj.al describedabove, provided acknowledgement is given to the source. "
Mrs.Re s e archNa t ional
yth HoImes, DirectorJournaLsResearch council
February 14, . L994




















