
Flexible fate commitment of E2-2high common DC progenitors ...
Upload
independentCategory
view
4download
0
Original Article
Isolation and angiogenesis by endothelial progenitors in the fetal liver
Running title: Characterization of fetal liver endothelial progenitors
Stephanie Cherqui1, Sunil M. Kurian1, Olivier Schussler1, Johannes A. Hewel2, John R.
Yates III2 and Daniel R. Salomon1
1 Department of Molecular and Experimental Medicine 2 Department of Cell Biology The Scripps Research Institute, 10550 North Torrey Pines Road, La Jolla, California 92037, USA. *Corresponding author: Daniel R. Salomon, M.D. Department of Molecular and Experimental Medicine
The Scripps Research Institute, 10550 North Torrey Pines Road, La Jolla, California 92037, USA. Phone: 858 784 9381 Fax: 858 784 2121 [email protected]
Received February 18, 2005; accepted for publication June 21, 2005. ©AlphaMed Press 1066-5099 doi: 10.1634/stemcells.2005-0070 TSRI Manuscript #: 16902-MEM Word Count: 5240 Financial Statement: This work was funded by the National Institutes of Health, R21 DK62598, the Juvenile Diabetes Research Foundation, 3-2003-738 and the Molly Baber Reseach Fund. No support for this work was derived from any commercial source and the authors have no direct financial interest in any aspect of the manuscript.
1
Stem Cells Express, published online August 11, 2005; doi:10.1634/stemcells.2005-0070
Copyright © 2005 AlphaMed Press

Abstract
Endothelial progenitor cells (EPC) have significant therapeutic potential. However, the
low quantity of such cells available from bone marrow and their limited capacity to
proliferate in culture make their use difficult. Here, we present the first definitive
demonstration of the presence of true EPC in murine fetal liver capable of forming blood
vessels in vivo connected to the host’s vasculature after transplantation. This population
is particularly interesting because it can be obtained at high yield and has a high
angiogenic capacity as compared to bone marrow-derived EPC. The EPC capacity is
contained within the CD31+Sca1+ cell subset. We demonstrate that these cells are
dependent for survival and proliferation on a feeder cell monolayer derived from the fetal
liver. In addition, we describe a novel and easy method for the isolation and ex vivo
proliferation of these EPC. Finally, we used gene expression profiling and tandem mass
spectrometry proteomics to examine the fetal liver endothelial progenitors and the feeder
cells to identify possible proangiogenic growth factor and endothelial differentiation-
associated genes.
Word Count = 169
2

Introduction
Endothelial progenitor cells (EPC) have the capacity to proliferate and
differentiate into mature endothelial cells. An unexpected and exciting development was
the discovery that endothelial progenitors, resident in peripheral blood [1-3] and bone
marrow [4-7], can be recruited from the circulation and participate in angiogenesis at
sites of tissue injury and/or ischemia [8, 9]. Vascular progenitors can also be isolated
from skeletal muscle and can differentiate into endothelial cells during ischemic injury-
induced neovascularization [10]. Similarly, embryonic lung mesenchyme contains
endothelial precursors [11].
Many have suggested the therapeutic potential of progenitor-driven angiogenesis
[9]. However, the differentiation of EPC into functional blood vessels in vivo is complex
and incompletely understood. For example, what is the biological significance and
regulation of angiogenesis mediated by EPC at sites of tissue injury in contrast to
angiogenesis mediated by local endothelial cells? Moreover, the quantity of EPC within
the circulation, even mobilized from the bone marrow, is low and only a small percentage
of CD34+ hematopoietic stem cells have endothelial progenitor capacity [12, 13].
Therefore, understanding growth factor and cell signal pathways that direct EPC
proliferation, survival and differentiation is an important strategy to enrich for
angiogenesis-competent cells suitable for clinical applications.
Fetal liver is a source of stem cells that can give rise to hepatocytes and biliary
epithelial cells [14, 15]. It is also well established that fetal liver contains Sca1+
hematopoietic stem cells capable of differentiation to myeloid and lymphoid lineages
3

[16-18]. A subset of fetal liver stem cells has been shown to express endothelial
progenitor cell makers in vitro [2, 9, 12] suggesting that they might be EPC. However,
this commonly accepted fact has never been proved by demonstration of angiogenesis in
vivo mediated directly by participation of these putative EPC. Our interests in the
potential of manipulating tissue compartment-specific progenitors as a means of
enhancing revascularization of cell transplants during tissue engineering led us to develop
a new method to purify stem cells from murine fetal liver. We characterized a
CD31+Sca1+ population of cells that contains the EPC. While it is certainly possible that
some hematopoietic stem cell activity is also contained within this population, the present
work is focused on their endothelial progenitor and angiogenic potential. Thus, we
established that the CD31+Sca1+ cells have a high efficiency of angiogenesis in vivo. We
then used biology, genomics and proteomics to better characterize these EPC and
possible growth factors and receptors required for survival, proliferation and maturation.
4

Methods
Animals. Balb/c and C57BL/6 mice were obtained from The Scripps Research Institute’s
animal facility. NOD/SCID mice were obtained from our own colony. Transgenic Tie2-
GFP mice were purchased from Jackson Laboratory (strain FVB/N-
TgN(Tie2GFP)287Sato). Protocols were approved by the Institutional Animal Care and
Use Committee. This program is AAALAC accredited and conforms to all USDA and
OPRR guidelines.
Liver Endothelial Progenitors (LEP) isolation. Livers obtained from 5 to 7 fetal
embryos (15-21 days post-implantation) were dissected, minced and washed. After
spinning 30 seconds at 99 xg, the tissue was digested with 3 mg/ml collagenase P (Sigma,
St Louis, Missouri) in 4 ml of HBSS with 1M CaCl2 and hand-agitated at 37 oC for 3.15
minutes. Digestion was stopped by adding 20 ml of HBSS with BSA 0.35% and cooling
in ice 10 minutes. After discarding 20 ml of supernatant and adding 5 ml of HBSS+BSA,
the pieces were triturated 6 times using a 14G needle attached to a 10 ml syringe. The
mixture was then pelleted by centrifugation (99 xg for 30 seconds), 5 ml of buffer was
replaced and this entire procedure was repeated twice. The digested tissue pellet was then
resuspended in 30 ml of complete RPMI-1640 medium (Cambrex, East Rutherford, New
Jersey), 10% fetal calf serum, 4 mM glutamine, 1 mM sodium pyruvate, 100U
penicillin/streptomycin (Invitrogen, Carlsbad, California), split in a 6-well plate with 5 ml
per well and cultured at 37 oC and 7% CO2. The media was changed every two days. We
performed all our studies with cells harvested at 8 days.
5

Bone marrow stem cell isolation. Bone marrow cells were extracted by flushing from the
tibias and femurs of C57Bl/6 mice at 6 to 10 weeks of age.
Sorting of Sca1+ stem cells. Sorting of both fetal liver and adult bone marrow cells was
performed using anti-Sca1 antibody conjugated to mini-magnetic beads (Miltenyi Biotec,
Inc., Auburn, California) according to the manufacturer’s instructions. Sca1+ cells are
eluted with a purity of better than 90% by flow cytometry and >99% viability by vital dye
exclusion.
Proliferation and survival assay. 105 cells per well were seeded in triplicate in 96-well
plates and cultured for 3 days in medium or supplemented with 10%, 20% and 40% of 48
h conditioned supernatants harvested from a feeder cell monolayer. The cells were
harvested with trypsin-EDTA and counted. Apoptosis and cell viability were measured
using annexin V and propidium iodide according to manufacturer’s protocols (Molecular
Probes, Eugene, Oregon). Proliferation was measured with [3H]thymidine in round
bottom 96-well plates added at 48 h [1 µCi ml-1] and 16 h later the cells were harvested
onto glass fiber filters and counted by liquid scintillation (MicroBeta, Wallac).
Matrigel Vascular-like Tube-Forming Assay. Matrigel (BD Biosciences, San Jose,
California) was added to the wells of a 24-well plate in a volume of 300 µl and allowed
to solidify at 37oC for 30 min. After the Matrigel solidified, LEP (1x106 cells) were
added in 1 ml of media: EBM-2 supplemented with FCS 2%, hydrocortisone, hFGFb,
6

VEGF, IGF1, hEGF, ascorbic acid and heparin (EGM-2 Bulletkit, Cambrex). The cells
were incubated at 37oC, 7% CO2 for 7 days and then photographed.
Endothelial Progenitor Colony-Forming Unit (EP-CFU) Assay. 5 x 106 LEP/well were
resuspended in 2 ml of Endocult Liquid Medium (StemCell Technologies, Vancouver,
British Colombia), plated on fibronectin-coated 6-well culture dishes (BD Biosciences)
and incubated for 2 days at 370C, 7% CO2. The non-adherent cells were then collected
and plated at 5 x 105 cells/well on fibronectin-coated 24-well culture dishes in 1ml of
Endocult Liquid Medium. After three days the CFU were counted and photographed.
Transplantation of Matrigel Templates. 106 cells were mixed in 500 µl of iced Matrigel
Basement Membrane Matrix to prevent gelification and injected subcutaneously into the
flank of mice using a 23G needle.
Estimation of Blood Vessels. Mouse tissues or Matrigel templates explanted 2 weeks
post transplant, were minced and digested in collagenase P (1.6 mg/ml; Sigma) and
DNaseI (10 U/ml; Roche Biochemicals, Indianapolis, Indiana) for 2 hours at 37 oC and
resuspended by pipetting every 30 minutes. After filtering through a 70 µm filter (BD
Biosciences), cells were collected and stained with blood vessel-specific antibody:
PhycoErythrin (PE) anti-CD31 antibody, a PE-conjugated rat IgG2a was used as the
isotype control (Caltag, Burlingame, California). A PE-labeled anti-αIIbβ3 antibody
(EMFRET Analytics, Wurzbug, Germany) and the corresponding PE-Rat IgG2b isotype
control was used to determine the quantity of platelets in the CD31+ subset. PE-
7

streptavidin was used to reveal binding of biotinylated Griffonia simplicifolia lectin I
isolectin B4 (Vector, Burlingame, California). The lectin (25 µg) was injected in the tail
vein of NOD/SCID mice 20 minutes before harvesting the transplanted Matrigel
templates.
Antibody phenotyping of LEP. Anti-mouse FcR CD16/CD32 (BD Biosciences) at 1 µg
per 106 cells incubated 10 minutes on ice was used as a blocking step. The following
directly conjugated anti-mouse antibodies were used at 1 µg per 106 cells and incubated 1
hour on ice: Fluoroscein IsoThioCyanate (FITC)-anti-Sca1 (D7 Ly-6A/E, eBioscience,
San Diego, California), FITC-anti-CD45R/B220 (BD Biosciences) and the isotype
control, FITC-Rat IgG2a. Similarly, we used PE-anti-CD31 (Caltag) with PE-Rat IgG2a
as isotype control and FITC-anti-F4/80 (Caltag) with FITC-Rat IgG2b as isotype control.
Histology. Explanted Matrigel templates were fixed in 4% Paraformaldehyde 4 hours at
room temperature and incubated overnight in 10% sucrose at 4 °C. Specimens were
frozen in embedding medium (O.C.T, Tissue-Tek, Redding, California) at –80 oC and 8
µm thick frozen sections made. After blocking with 1% BSA, 10% donkey serum in PBS
for 1 hour at room temperature, sections were stained with rabbit anti-human von
Willebrand Factor (vWF) (Dako, Carpinteria, California) at 1:200 dilution for 1 h at room
temperature, followed by donkey anti-rabbit IgG conjugated with Cy5 (Jackson
ImmunoResearch, West Grove, Pennsylvania) at 1:200 dilution for 1 h at room
temperature. Cy5-streptavidin (1:100 dilution) identified biotinylated Griffonia
8

simplicifolia isolectin. Sections were visualized using a BIO-RAD MRC1024 laser
scanning confocal microscope (Hercules, California).
RT-PCR. Total RNA was prepared in 1 ml of Trizol (Invitrogen), purified using RNeasy
columns (Qiagen, Valencia, California) and quality confirmed on an Agilent 2100
BioAnalyzer. Five µg of total RNA, treated with DNase (DNA-free; Ambion, Austin,
Texas), was reverse transcribed using the SuperScript First-Strand Synthesis System with
oligo dT primers (Invitrogen). First strand DNA was treated with DNase-free RNase
(Invitrogen). PCR was performed as follows: 94 °C, 30 seconds; 55-60 °C, 1 minute; 72
°C, 1 minute for 40 cycles. Primer sequences for the following genes are published:
Angiopoietin-1 (Ang-1), Angiopoietin-2 (Ang-2), Tie-1, Tie-2, Flk-1 and Flt-1 [5], Flt-4
[19], Cd34 [20], Endoglin [21], c-kit [22], Aa4 [23], Vcam-1 [24]. We also designed
primers for: Hhex, 5’-ATCTCAGAGGATTCCGACCAGG-3’ forward, 5’-
ATTCCCCAATGTTGCCCCCAC-3’ reverse (513 bp); Cd133, 5’-
GGAAAAGTTGCTCTGCGAACC-3’ forward, 5’- TGCTTGTTTGCTGGAGGGTC-3’
reverse (608 bp); and Tal1, 5’-GCCCAAAGATTTCCCCAATG-3’ forward, 5’-
AAACCCAGTGCCCCAAACAC -3’ reverse (543 bp); VE-Cadherin 5’ -
CAGCCAGCATCTTGAACCTG – 3’ forward, 5’ - GAGATTCACGAGCAGTTGGT -
3’ reverse (506 bp) and vWF 5’ - TGTTTTGTGGCGTGTATGTGAGG - 3’ forward, 5’
- GTGTTCTGGGTTTTCTGGAGTTTG - 3’ reverse (584 bp).
DNA Microarrays. Affymetrix GeneChip (Santa Clara, California) protocols were used
for all hybridizations. Samples were hybridized to MOE430A GeneChip arrays. Data was
9

analyzed using GeneChip Operating Software (GCOS) Version 1.0 from Affymetrix,
which computes signal intensity and P values for each probe set (Wilcoxon Rank Sum
test) and generates Present/Absent calls. We used RMA Express for signal normalization
[25] and BRB ArrayTools for class comparisons (http://linus.nci.nih.gov/BRB-
ArrayTools.html) and Cluster/TreeView for creation of heat map displays [26].
Microarray data for all the GeneChips are available at the Gene Expression Omnibus
(GEO) website (www.ncbi.nlm.nih.gov/geo) under the series ID GSE1727.
Protein extraction. Proteins were extracted with isopropyl alcohol from phenol-ethanol
supernatants of Trizol extracts after RNA was removed. Samples were allowed to
precipitate for 10 min (25 °C) and sedimented at 12,000 xg (10 min, 4°C). The protein
pellet was washed three times in two volumes 0.3 M guanidine hydrochloride in 95%
ethanol for 20 minutes at 25 °C and then centrifuged at 7,500 xg (5 min, 4 °C). The pellet
was then vortexed in 2 ml of 80% ethanol and centrifuged at 7,500 xg (5 min, 4 °C). The
protein pellet was dried at room temperature for 10 min and stored at –20 °C.
Multidimensional protein identification technology (MudPIT). We used
multidimensional protein identification technology (MudPIT) for this analysis as
described previously [27]. Protein samples were analysed using two different techniques
for cleavage. One replicate from each sample was enzymatically cleaved using
Endoproteinase Lys-C (Roche Biochemicals) followed by digestion with sequencing
grade modified Trypsin (Promega, Madison, Wisconsin). The other replicate was
10

chemically cleaved with Cyanogen Bromide (CnBr) in addition to enzymatic cleavage
with Trypsin and Endoproteinase Lys-C.
Analysis of tandem mass spectra. MS/MS spectra were analyzed using the SEQUEST
software analysis protocol as described [28]. A filter, called 2to3 [29], determined the
charge state (+2 or +3) of multiple peptide spectra and poor-quality spectra were deleted.
Each MS/MS spectrum after analysis and filtering was searched against the SwissProt
database (Release 42.0) and EntrezProtein (July 24) using SEQUEST [28]. DTASelect
was used to filter peptide identifications. Filter criteria were set to Xcorr values >2.2 for
1+ spectra, >2.5 for 2+ spectra and >3.5 for 3+ spectra with DeltCn of 0.1. For the
proteins hits of mRNA microarray data, moderate stringency was applied with Xcorrs of
>0.8 and Delta Cn of 0.01 followed by manual validation of each peptide spectrum based
on two main criteria: 1) more than 3 of the most intense fragment ions must show a
match and, 2) the b and y ion series must show continuity for at least 3 fragment ions
above background noise.
Statistical analysis. Data are expressed as mean ± standard error (SE) of at least 3
independent experiments. ANOVA was used to detect differences in cells survival and
proliferation. A P value of <0.05 was considered statistically significant.
11

Results
Phenotypic characterization of the isolated LEP. We developed a novel way to easily
isolate a population of cells containing endothelial progenitor cells from mouse fetal
livers in large numbers as detailed in the Methods. Between six and twelve days after
isolation, weakly adherent and highly light-refractile round cells grow out in clusters on a
firmly adherent cell monolayer (Figure 1A). We call these round cells LEP (Liver
Endothelial Progenitors). We can obtain these cells in large quantity, 5.5±0.5 x106 cells
per 5 to 7 murine fetal embryos harvested 15-18 days post-implantation. Flow cytometric
analysis of LEP after 8 days in the cultures demonstrated populations of cells expressing
the progenitor-associated markers, CD31 (46.3%±1.6) and Sca1 (Ly6a, 26.0%±7.0). On
the other hand, LEP were essentially hematopoietic lineage negative. Flow cytometry
analysis demonstrated that the cells are not macrophages (F4/80, 1.8%±0.6 vs. isotype
control, 1.2%+0.2). Staining for CD45R/B220 marking mature T, B and NK cells as well
as lymphocyte and macrophage progenitors in fetal liver was 3.8%±0.3 vs. the isotype
control of 1.2%+0.2. LEP are only found in fetal, not in adult, liver preparations.
Our working hypothesis was that the monolayer of cells that underlies the LEP
clusters represents a feeder cell population. To test this hypothesis, supernatant from the
feeder cells was added at various concentrations: 0, 10, 20 and 40% by volume to
harvested LEP. After 72 hours we collected the cells for counting and stained with
annexin V and propidium iodide to determine the quantity of dead cells by flow
cytometry. The number of LEP increased significantly between 0 and 10% conditioned
media (P=0.04) and it was maximal at 20% (Figure 1B upper panel). Similarly, we
12

observed a significant decrease in the quantity of dead cells between 0 and 10% of
conditioned media (P=0.004) after which this measure reached a plateau (Figure 1B
lower panel). The same data was obtained with a proliferation assay using 3H-Thymidine
(4,606±285 cpm for 0% vs. 16,557±525 cpm for 10%; P=0.04). These data prove that
factors produced by the underlying feeder cell monolayer are essential for the survival
and proliferation of LEP. One possibility is that the conditioned media might support
LEP survival and proliferation but not the maintenance of their endothelial progenitor
function. To test this hypothesis we cultured LEP in 20% conditioned media for 2 weeks
after which we demonstrated they could still form blood vessels in vivo after
transplantation into immunodeficient mice (data not shown) in the Matrigel template
assay described below.
LEP can form blood vessels in vitro. A well-established assay for the angiogenic
potential of endothelial progenitors is the development of vascular-like tubes in a
Matrigel in vitro culture [30]. LEP isolated from the feeder cell monolayers after 8 days
were plated in Matrigel and cultured for 5 days in an endothelial cell growth media.
Figure 2 is a photomicrograph of a representative culture showing the characteristic
vascular-like tubes developing from the LEP.
LEP can form blood vessels in vivo after transplantation. We used an angiogenesis
assay based on collagenase digestion and flow cytometry to measure the potential of LEP
to form new vessels in vivo post transplant. The underlying premise is that endothelial
cells released by collagenase digestion of a tissue and stained with endothelial cell-
13

specific antibodies can be counted by flow cytometry and expressed as the percent
positive cells in the unlabeled background. Thus, the number of endothelial cells in a
given tissue is a function of the number of vessels. To validate this assay, we harvested
kidney, lung, heart and brain from four transgenic Tie2-GFP mice, where only
endothelial cells express GFP [31]. Cells harvested from the collagenase-digested tissues
were then analyzed for expression of CD31, as an independent marker for mature
endothelial cells, and GFP to detect the Tie2+ cells. These results confirmed that all GFP+
cells were also CD31+ and represent mature endothelial cells (data not shown).
Next, we tested the use of Matrigel templates as a platform technology to verify
the endothelial capacity of LEP isolated from transgenic Tie2-GFP mice. LEP cells were
seeded in Matrigel (106 LEP per 500 µl) and injected subcutaneously into NOD/SCID
mice. After two weeks the Matrigel templates were explanted and analyzed by histology
and the collagenase-based flow cytometry assay described above. By flow cytometry,
GFP+ cells (i.e. Tie2+) are observed in templates containing LEP (Figure 3A) and these
cells are also CD31+ representing the mature endothelial cells derived from the LEP.
CD31+ endothelial cells comprise 35.5±1.6% (corresponding to ~1.3x106 cells) of all the
cells in the explanted Matrigel templates. In turn, the GFP+CD31+ cells derived from the
LEP represent 9.3±0.4% (corresponding to ~3.4x105 cells) of the total cells in the
template or 26% of the endothelial cells. Note that the presence of host-derived
endothelial cells (GFP-CD31+) cells in both the LEP and control templates is consistent
with the fact that Matrigel is a proangiogenic material.
One possible concern for use of anti-CD31 staining as a parameter for estimating
tissue angiogenesis is that this molecule is also expressed on platelets. To exclude the
14

possibility that marginated platelets and/or postmortem thrombi are contributing to the
staining we stained with an antibody reacting with murine integrin αIIbβ3 (Leo.D2),
specifically expressed by platelets [32, 33]. The results were that staining with this
antibody represents 2 to 3% of the total CD31 staining observed, excluding platelets as a
source of error.
That LEP form blood vessels in vivo in the Matrigel post transplant was
confirmed by frozen sections of explanted templates stained with the blood vessel-
specific anti-vWF antibody (Figure 3B-D). The key point is a co-localization between
vWF staining and the LEP-derived GFP+ cells lining the vascular lumens, which have
differentiated into a mature endothelial cell phenotype (Figure 3D).
To test whether these new vessels were connected to the host vasculature, we
injected the biotinylated endothelium-specific isolectin B4 in the tail vein of NOD/SCID
mice two weeks after the transplantation of LEP-containing Matrigel templates. The
explanted templates were stained with PE-streptavidin and we showed by flow cytometry
that GFP+ cells (Tie2+) are also isolectin B4+ (data not shown). We obtained the same
result by confocal microscopy by staining with streptavidin-Cy5 to identify the
biotinylated lectin bound to endothelial cells. Co-localization between the GFP+ cells and
the isolectin B4+ cells is clearly seen in Figure 2G. In conclusion, the LEP are endothelial
progenitor cells, mature into endothelium and incorporate into blood vessels that are
connected with the host vasculature after transplantation.
Finally, for the sake of a functional comparison, the same experimental design
was performed with bone marrow-derived Sca1+ cells from adult Tie2-GFP mice. Flow
cytometric analysis of the explanted Matrigel templates demonstrated that only
15

1.2±0.03% of the total cells were GFP+/CD31+ endothelial cells. These results
demonstrate the high angiogenic efficiency of the fetal liver-derived LEP as compared to
the adult bone marrow-derived Sca1+ cells to form blood vessels in vivo.
LEP Sca1+ cells contain the endothelial progenitor cells. Fetal livers were harvested,
collagenase digested and the resulting cell mixtures were placed in culture. After 8 days
the LEP were collected from the underlying feeder layer and magnetically sorted using
anti-Sca1 antibody. The presence of EPC was tested in LEP, LEP Sca1+ and LEP Sca1-
populations using the Endothelial Progenitor Colony-Forming Unit (EP-CFU) assay. We
counted the number of colonies characterized by a central cluster of rounded cells
surrounded by radiating, thin, flat cells (Figure 4): LEP 23.5±0.5, LEP Sca1+ 56.5±5.5
and LEP Sca1- 3.5±0.5. The quantity of CFU is significantly enriched in the Sca1+ subset
(P=0.01) and lower in Sca1- (P=0.001).
Sca1+ and Sca1- LEP from Tie2-GFP mice were then transplanted (106
cells/Matrigel template). Two weeks post transplant, we injected biotinylated isolectin B4
by tail vein and then explanted the templates, collagenase digested and examined the cells
by flow cytometry. The results were that GFP+/Tie2+ endothelial cells represent
0.9±0.8% of the total cells analyzed in the empty template control, 1.6±0.9% for Sca1-
and 8±2.4% for the Sca1+. Two color analysis for PE-labeled isolectin staining revealed
0.6±0.5% for the empty template control, 1.5±0.4% for Sca1- and 6.1±2.1% for the
Sca1+. Therefore, all these data prove that the LEP Sca1+ cell subset contains the
endothelial progenitors.
16

Gene expression profiling of LEP Sca1+ and Sca1- cells. In order to investigate potential
factors that drive EPC differentiation, we compared the genes expressed by these two
populations. We first performed RT-PCR on LEP Sca1+ and Sca1- cells testing 17 genes
associated in the literature with hematopoietic and endothelial progenitors (Figure 5). The
genes uniquely expressed by LEP Sca1+ are the VEGF receptors, Kdr, Flt1 and Flt4, and
also Cd34, Vcam-1, Cd133 and VE-cadherin. In contrast, vWF, Endoglin, AA4, Hhex,
Tal1 are expressed by both Sca1+ and Sca1- populations. All the primers were verified
using the feeder cell monolayer, a negative control of NIH 3T3 fibroblasts and a positive
control of a mouse endothelial cell line, MS1 [34] (data not shown).
Next, we performed gene expression profiling comparing LEP Sca1+ and LEP
Sca1- using Affymetrix MOE430A GeneChips (n=2 Sca1+ and n=3 Sca1-). The signal
intensities were normalized using RMA Express and a class comparison for differentially
expressed genes was performed with BRB ArrayTools with the P value filter set at <
0.005. This analysis demonstrated 445 differentially expressed genes: 228 genes were up-
regulated in the Sca1+ subset and 217 genes were up-regulated in the Sca1- subset. We
then filtered the entire set of differentially expressed genes for functions related to
angiogenesis and differentiation on the basis of the published literature as identified in
OMIM, PubMed and MGI as well as functional classifications of known genes listed at
NetAffyx and GO database. There are 54 proangiogenic genes expressed by Sca1+ cells
and only 3 by Sca1- (all different than those expressed by Sca1+). Figure 6 depicts a heat
map and cluster analysis of the proangiogenic and anti-angiogenic genes differentially
expressed by both LEP subsets. A full set of all differentially expressed genes is available
as Supplemental Data (Table S1).
17

Candidate growth factor pathways. LEP depend for survival and proliferation on the
underlying feeder cells or their supernatants. We propose that this dependence creates a
model system to identify growth factor and differentiation signal pathways for EPC that
may also be relevant to angiogenesis. Therefore, we performed gene expression profiling
using Affymetrix MOE430A GeneChips to compare the LEP to the feeder cell
monolayer. To focus on potential proangiogenic growth factors secreted by the feeder
cell monolayer, we selected candidates whose genes were expressed only by the feeder
cells and where the genes for their receptors were expressed by the LEP. This analysis
identified 9 growth factor/receptor pathways potentially involved in the survival and
proliferation of LEP: Bone Morphogenic Protein-4 (BMP-4), Epidermal Growth Factor
(EGF), Leukemia Inhibitory Factor (LIF), Fibroblast Growth Factor-1 and –2 (FGF1,
FGF2), Insulin-like Growth Factor-2 (IGF2), Platelet-derived Growth Factor Alpha
(PDGFα), Transforming Growth Factor Beta (TGFβ), and Vascular Endothelial Growth
Factor (VEGF). We then used the literature and web-based tools as already described as
well as Biocarta (www.biocarta.com) to create gene lists for each growth factor-
stimulated receptor signaling pathway potentially activated in the LEP. The results of this
analysis are provided as Supplementary Data (Table S2) and demonstrate that 92 out of
124 total genes in this candidate pathway table (74%) are found “Present” by gene
expression profiling in the LEP.
We also tested the expression of the candidate genes described above using
tandem mass spectrometry proteomics with the Multidimensional Protein Identification
Tool (MudPIT) [35]. Thus, we matched the protein candidates identified in LEP (1,817
18

proteins) and feeder cells (1,602 proteins) to the 9 growth factor/receptor pathways
identified using gene expression profiling (Table S2). In total, 75 candidate proteins out
of the 124 (60%) in the original set of gene expression-defined pathways were confirmed
by MudPIT proteomics.
19

Discussion
Stem cells have the potential of being tools for tissue engineering and
regenerative medicine. A major limitation at the present time is that endothelial
progenitor frequency in bone marrow and mobilized peripheral blood cells is extremely
low and the efficiency of angiogenesis is poor. Thus, a major challenge is developing
strategies to engineer the differentiation and proliferative expansion of endothelial
progenitors (EPC) in culture. In the present paper we describe a model of endothelial
progenitor cells that is particularly suitable for experiments designed to investigate
molecular mechanisms of EPC survival, proliferation and differentiation.
Specifically, we demonstrate that a population of stem cells we have called LEP
(Liver Endothelial Progenitors) are found in the murine fetal liver and can be readily
isolated. We prove that CD31+Sca1+ LEP are endothelial progenitors that form new
vessels at high efficiency (~26% of the total endothelial cells in a transplanted Matrigel
template) and are connected to the blood stream of the host after transplantation. These
data represent the first definitive demonstration that true EPC are indeed present in fetal
liver. Another cell population derived from the fetal liver, called LEP feeders, provide a
set of growth factors that drive LEP proliferation and survival in culture, while
maintaining their angiogenic potential.
Though EPC have been identified in bone marrow, cord and peripheral blood,
they represent only a small fraction of the cells in these compartments, typically less than
2% [12, 13, 36]. In contrast, we demonstrate that collagenase digestion of the murine
fetal liver is a relatively easy method that yields a large number of readily collected LEP
(typically 5.5±0.5 x106 cells per pregnant female) and the Sca1+ subset comprising the
20

EPC activity represents 26% of these cells. We transplanted Sca1+ cells obtained from
both fetal livers and adult bone marrow to assess their relative angiogenic efficiency. This
comparison demonstrated that 9.3% of the mature endothelial cells were derived from the
Sca1+ LEP as compared to only 1.2% from the Sca1+ adult bone marrow cells, supporting
our conclusion that the LEP are highly efficient in angiogenesis.
Based on our demonstrations that the Sca1+ LEP are functional EPC in vitro and
in vivo, we next evaluated the differences between Sca1+ and Sca1- LEP in a number of
known proangiogenic molecules by gene expression profiling using RT-PCR and high
density DNA microarrays. The key point is that defining the endothelial progenitor
potential of a new cell population must start with an array of well-established
angiogenesis assays rather than inferring such activity a priori by measuring a panel of
putative cell surface markers for EPC, many of which are contradictory, mark
overlapping cell populations of various lineages, may also stain mature endothelial cells
and none of which are presently proven to be endothelial progenitor-specific as single
markers.
We tested the endothelial progenitor lineage of the LEP Sca1+ by RT-PCR,
demonstrating that only the Sca1+ express the VEGF receptor genes, Kdr, Flt1 and Flt4,
as well as Cd133, VE-cadherin, Cd34 and Vcam-1. That VEGF receptors are expressed
by the Sca1+ cells is important because VEGF is a key regulator of angiogenesis [37] and
Kdr, Flt1 and Flt4 are involved in vascular development [36, 38, 39]. VE-cadherin and
CD133 are cell surface markers generally agreed upon to be expressed by endothelial
progenitors.
21

We extended our analysis of differential gene expression by using high density
DNA microarrays. A large number of genes (228) are differentially up-regulated in the
Sca1+ cells and 56 are linked functionally to angiogenesis including only 2 that are anti-
angiogenic (Figure 6). Among the 54 proangiogenic genes up-regulated, we note the well
known endothelial progenitor-associated genes, Kdr and VE-cadherin (cdh5), consistent
with our RT-PCR data. Indeed, Sca1+Kdr+VE-cadherin+ cells have been shown to define
a bone marrow-derived mouse endothelial progenitor [9]. A number of the up-regulated,
angiogenesis-associated genes in the Sca1+ LEP represent candidates for further studies.
Pleiotrophin and Midkine represent a family of mitogenic and angiogenic growth and
differentiation factors expressed in cells in early differentiation [40, 41]. The secreted
Frizzled Related sequence Protein-1 (sFRP-1) is strongly expressed during early phases
of the vascularization process in embryonic vasculature and modulates vascular cell
proliferation [42]. It is interesting that the Frizzled homolog 2 (Fzd2) is also up-regulated
in Sca1+ LEP but its precise physiologic function has not been determined yet and so this
was not counted in our list of angiogenesis-associated gene candidates [43]. Tweak
receptor (Tnfrsf12a) induces angiogenesis in vivo [44]. Laminin alpha4 (lama4) and
Nidogen (Nid1) are both up-regulated in Sca1+ LEP and form a stable complex that plays
an important modulatory role in angiogenesis [45]. On the other hand, only 6 genes are
up-regulated and angiogenesis-associated in the Sca1- LEP including 3 that are anti-
angiogenic (Figure 6).
In conclusion, the genomic profiling of LEP Sca1+ versus Sca1- cells
distinguished these two populations by differential gene expression. Specifically, LEP
Sca1+ cells are characterized by up-regulation of primarily angiogenesis-associated and
22

endothelial lineage-specific genes, while the LEP Sca1- cells reveal a majority of
hematopoietic lineage-specific genes. This separation by gene expression profiling
supports the biological results we obtained showing that the Sca1+ comprise the
endothelial progenitor activity of the LEP.
Taking advantage of the evidence that LEP need the feeder cells to survive and
proliferate, we investigated the key question: what are the growth factors required to
maintain EPC in culture? Commonly, monoclonal antibodies or supplementation of
cultures with recombinant growth factors are used to identify expression of specific
growth factor receptors. In the present study, we used a combined strategy of gene
transcript, protein expression profiling and bioinformatics to identify candidate pathways.
Therefore, we organized the gene expression profiling by selecting pathways in which the
feeder monolayer cells expressed the growth factor and the LEP expressed the receptor.
This integrated analysis with bioinformatics allowed us to identify 9 different growth
factor pathways. All these growth factors are well known and can be involved in cell
proliferation and survival. The LEP feeder cells may also have a role in EPC
differentiation. Indeed, among the 9 growth factors, 5 have at least one of their receptors
only expressed on the Sca1+: Acvr1, Egfr, Fgf1, Tgfbr2, and Vegfr-1, 2, 3. Beside VEGF
receptors that are well known for enhancing angiogenesis, the function of the others in
endothelial differentiation is less well known and they represent promising candidates for
further studies of endothelial differentiation.
In conclusion, we have proven that Sca1+ endothelial progenitors are found in the
fetal liver and that these form blood vessels integrated with the host’s vasculature when
transplanted in Matrigel templates. We have developed an isolation strategy that is easy
23

and yields a significant number of progenitors for study that proliferate in culture and
also have a high efficiency of angiogenesis in vivo. Thus, we suggest that this murine
system has promise as a means for advancing studies of endothelial progenitor
differentiation, angiogenesis and transplantation. Finally, we have used gene expression
profiling and tandem mass spectrometry proteomics as a first approach in identifying
candidate genes potentially involved in EPC growth and survival and endothelial
differentiation and angiogenesis.
24

Acknowledgements
We acknowledge the kind gift of the anti-phosphotyrosine-specific antibodies from Dr.
Martin Schwartz (University of Virginia). This work was funded by the National
Institutes of Health, R21 DK62598 (DRS), the Juvenile Diabetes Research Foundation, 3-
2003-738 (SC) and the Molly Baber Research Fund.
25

References
1. Asahara T, Murohara T, Sullivan A, et al. Isolation of putative progenitor endothelial cells for angiogenesis. Science 1997;275(5302):964-967.
2. Shi Q, Rafii S, Wu MH, et al. Evidence for circulating bone marrow-derived endothelial cells. Blood 1998;92(2):362-367.
3. Schatteman GC, Hanlon HD, Jiao C, Dodds SG, Christy BA. Blood-derived angioblasts accelerate blood-flow restoration in diabetic mice. Journal of Clinical Investigation 2000;106(4):571-578.
4. Crisa L, Cirulli V, Smith KA, Ellisman MH, Torbett BE, Salomon DR. Human cord blood progenitors sustain thymic T-cell development and a novel form of angiogenesis. Blood 1999;94(11):3928-3940.
5. Jackson KA, Majka SM, Wang H, et al. Regeneration of ischemic cardiac muscle and vascular endothelium by adult stem cells.[see comment]. Journal of Clinical Investigation 2001;107(11):1395-1402.
6. Orlic D, Kajstura J, Chimenti S, et al. Bone marrow cells regenerate infarcted myocardium.[see comment]. Nature 2001;410(6829):701-705.
7. Rafii S, Meeus S, Dias S, et al. Contribution of marrow-derived progenitors to vascular and cardiac regeneration. Seminars in Cell & Developmental Biology 2002;13(1):61-67.
8. Rafii S. Circulating endothelial precursors: mystery, reality, and promise.[comment]. Journal of Clinical Investigation 2000;105(1):17-19.
9. Rafii S, Lyden D. Therapeutic stem and progenitor cell transplantation for organ vascularization and regeneration. Nature Medicine 2003;9(6):702-712.
10. Majka SM, Jackson KA, Kienstra KA, Majesky MW, Goodell MA, Hirschi KK. Distinct progenitor populations in skeletal muscle are bone marrow derived and exhibit different cell fates during vascular regeneration.[see comment]. Journal of Clinical Investigation 2003;111(1):71-79.
11. Akeson AL, Wetzel B, Thompson FY, et al. Embryonic vasculogenesis by endothelial precursor cells derived from lung mesenchyme. Developmental Dynamics 2000;217(1):11-23.
12. Peichev M, Naiyer AJ, Pereira D, et al. Expression of VEGFR-2 and AC133 by circulating human CD34(+) cells identifies a population of functional endothelial precursors. Blood 2000;95(3):952-958.
13. Shaw JP, Basch R, Shamamian P. Hematopoietic stem cells and endothelial cell precursors express Tie-2, CD31 and CD45. Blood cells, Molecules and Disease 2004;32:168-175.
14. Strain AJ, Crosby HA, Nijjar S, Kelly DA, Hubscher SG. Human liver-derived stem cells. Seminars in Liver Disease 2003;23(4):373-384.
15. Zhang Y, Bai XF, Huang CX. Hepatic stem cells: existence and origin. World Journal of Gastroenterology 2003;9(2):201-204.
16. Cumano A, Paige CJ, Iscove NN, Brady G. Bipotential precursors of B cells and macrophages in murine fetal liver. Nature 1992;356(6370):612-615.
17. Rebel VI, Miller CL, Eaves CJ, Lansdorp PM. The repopulation potential of fetal liver hematopoietic stem cells in mice exceeds that of their liver adult bone marrow counterparts. Blood 1996;87(8):3500-3507.
26

18. Ema H, Douagi I, Cumano A, Kourilsky P. Development of T cell precursor activity in the murine fetal liver. European Journal of Immunology 1998;28(5):1563-1569.
19. Deckers MM, Karperien M, van der Bent C, Yamashita T, Papapoulos SE, Lowik CW. Expression of vascular endothelial growth factors and their receptors during osteoblast differentiation. Endocrinology 2000;141(5):1667-1674.
20. Aube M, Lafrance M, Brodeur I, Delisle MC, Carreau M. Fanconi anemia genes are highly expressed in primitive CD34+ hematopoietic cells. BMC Blood Disord. Jun 2003;16(3):1.
21. St-Jacques S, Cymerman U, Pece N, Letarte M. Molecular characterization and in situ localization of murine endoglin reveal that it is a transforming growth factor-beta binding protein of endothelial and stromal cells. Endocrinology 1994;134(6):2645-2657.
22. Schrans-Stassen BH, van de Kant HJ, de Rooij DG, van Pelt AM. Differential expression of c-kit in mouse undifferentiated and differentiating type A spermatogonia. Endocrinology 1999;140(12):5894-5900.
23. Petrenko O, Beavis A, Klaine M, Kittappa R, Godin I, Lemischka IR. The molecular characterization of the fetal stem cell marker AA4. Immunity 1999;10(6):691-700.
24. Schawalder A, Oertli B, Beck-Schimmer B, Wuthrich RP. Regulation of hyaluronan-stimulated VCAM-1 expression in murine renal tubular epithelial cells. Nephrology Dialysis Transplantation 1999;14(9):2130-2136.
25. Bolstad BM, Irizarry RA, Astrand M, Speed TP. A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinformatics 2003;19(2):185-193.
26. Eisen MB, Spellman PT, Brown PO, Botstein D. Cluster analysis and display of genome-wide expression patterns. Proc Natl Acad Sci U S A Dec 8 1998;95(25):14863-14868.
27. McDonald WH, Ohi R, Miyamato DT, Mitchison TJ, Yates JRI. Comparison of three directly coupled HPLC MS/MS strategies for identification of proteins from complex mixtures: single-dimension LC-MS/MS, 2-phase MudPIT, and 3-phase MudPIT. International Journal of Mass Spectrometry 2002;219(1):245-251.
28. Eng JK, McCormack AL, Yates JRI. An approach to correlate tandem mass spectral data of peptides with amino acid sequences in a protein database. Journal of the American Society of Mass Spectrometry 1994;5(11):976-989.
29. Sadygov RG, Eng J, Durr E, et al. Code developments to improve the efficiency of automated MS/MS spectra interpretation. Journal of Proteome Research 2002;1(3):211-215.
30. Kubota Y, Kleinman HK, Martin GR, Lawley TJ. Role of laminin and basement membrane in the morphological differentiation of human endothelial cells into capillary-like structures. Journal of Cell Biology 1988;107(4):1589-1598.
31. Motoike T, Loughna S, Perens E, et al. Universal GFP reporter for the study of vascular development. Genesis: the Journal of Genetics & Development 2000;28(2):75-81.
32. Phillips DR, Charo IF, Scarborough RM. GPIIb-IIIa: the responsive integrin. Cell 1991;65(3):359-362.
27

33. Shattil SJ. Signaling through platelet integrin alpha IIb beta 3: inside-out, outside-in, and sideways. Thrombosis & Haemostasis 1999;82(2):318-325.
34. Arbiser JL, Moses MA, Fernandez CA, et al. Oncogenic H-ras stimulates tumor angiogenesis by two distinct pathways. Proceedings of the National Academy of Sciences of the United States of America 1997;94(3):861-866.
35. Durr E, Yu J, Krasinska KM, et al. Direct proteomic mapping of the lung microvascular endothelial cell surface in vivo and in cell culture. Nature Biotechnology 2004;22(8):985-992.
36. Salven P, Mustjoki S, Alitalo R, Alitalo K, Rafii S. VEGFR-3 and CD133 identify a population of CD34+ lymphatic/vascular endothelial precursor cells. Blood 2003;101(1):168-172.
37. Ferrara N. Role of vascular endothelial growth factor in regulation of physiological angiogenesis. American Journal of Physiology - Cell Physiology 2001;280(6):C1358-1366.
38. Yamashita J, Itoh H, Hirashima M, et al. Flk1-positive cells derived from embryonic stem cells serve as vascular progenitors.[see comment]. Nature 2000;408(6808):92-96.
39. Marchetti S, Gimond C, Iljin K, et al. Endothelial cells genetically selected from differentiating mouse embryonic stem cells incorporate at sites of neovascularization in vivo. Journal of Cell Science 2002;115(Pt 10):2075-2085.
40. Zhang N, Deuel TF. Pleiotrophin and midkine, a family of mitogenic and angiogenic heparin-binding growth and differentiation factors. Current Opinion in Hematology 1999;6(1):44-50.
41. Deuel TF, Zhang N, Yeh HJ, Silos-Santiago I, Wang ZY. Pleiotrophin: a cytokine with diverse functions and a novel signaling pathway. Archives of Biochemistry & Biophysics 2002;397(2):162-171.
42. Dufourcq P, Couffinhal T, Ezan J, et al. FrzA, a secreted frizzled related protein, induced angiogenic response.[see comment]. Circulation 2002;106(24):3097-3103.
43. Zhao Z, Lee CC, Baldini A, Caskey CT. A human homologue of the Drosophila polarity gene frizzled has been identified and mapped to 17q21.1. Genomics 1995;27(2):370-373.
44. Wiley SR, Cassiano L, Lofton T, et al. A novel TNF receptor family member binds TWEAK and is implicated in angiogenesis. Immunity 2001;15(5):837-846.
45. Nicosia RF, Bonanno E, Smith M, Yurchenco P. Modulation of angiogenesis in vitro by laminin-entactin complex. Developmental Biology 1994;164(1):197-206.
28
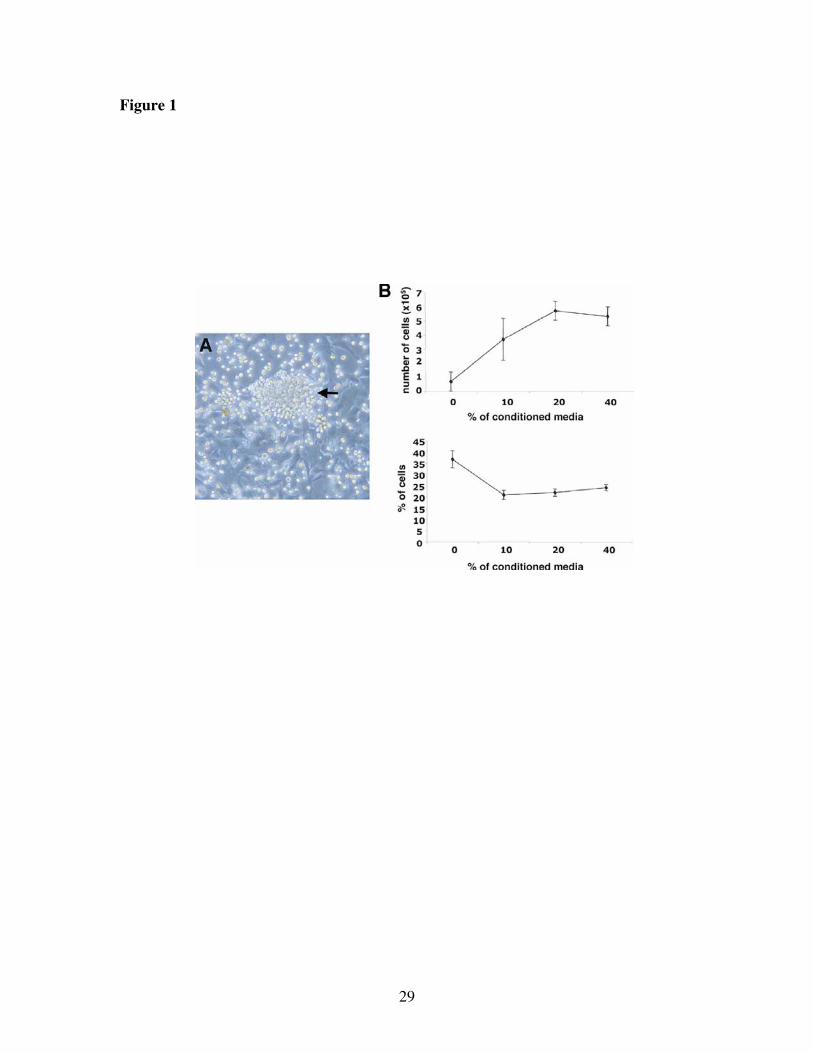
Figure 1
29

Figure 2
30

Figure 3
31

Figure 4
32

Figure 5
33

Figure 6
34

Figure Legends
Figure 1. Growth of Liver Endothelial Progenitor (LEP) cells on feeder cell
monolayers. (A) Light microscopy picture of LEP growing out of the collagenase
digestions of fetal mouse liver. The LEP are weakly or non-adherent and highly refractile
round cells that grow out in clusters (arrow) on a firmly adherent cell monolayer. (B) To
determine the effect of feeder cell-derived growth factors on LEP proliferation and
survival we harvested the LEP from the feeder cell monolayer and cultured them in 0, 10,
20 and 40% concentrations of conditioned feeder cell supernatants for 72 hours. The cells
were then counted (upper panel) or stained with annexin V and propidium iodide (lower
panel) to determine the quantity of dead cells.
Figure 2. Matrigel tube-formation assay. Light microscopy picture of LEP forming
tube-like structures (arrow) in Matrigel after 7 days in vitro. Magnification 200x.
Figure 3. Demonstration that LEP differentiate into endothelial cells in vivo. (A)
Analysis of LEP isolated from Tie2-GFP mice collected from Matrigel templates 2 weeks
post transplant. The two left panels marked ratIgG2a represent the isotype controls for the
anti-CD31 staining shown in the two right panels. GFP+ endothelial cells (R1) are present
in the Matrigel with LEP (lower) but not in the Matrigel control (upper). The two
GFP/CD31 plots show that all the GFP+ (Tie2+) cells in the Matrigel+LEP are also
CD31+ (R2). Based on R2, the number of endothelial cells generated from LEP in the
Matrigel is 8.8%. (B-G) Demonstration that LEP differentiate into endothelial cells in
35

vivo by confocal microscopy. LEP were transplanted in Matrigel templates and harvested
2 weeks post transplantation. GFP+ marking of the LEP is seen in the green channel (B)
and anti-vWF staining of blood vessels is shown in blue (C). The merged image shows
co-localization of GFP+ LEP cells with anti-vWF+ blood vessels (D). To demonstrate that
LEP-formed vessels are connected to the host vasculature we harvested the Matrigel
templates after injection of the biotinylated, endothelium-specific isolectin B4. GFP+ LEP
are seen in the green channel (E) and isolectin staining is shown in blue (F). The merged
image (G) shows the co-localisation of GFP+ LEP and isolectin staining, proving that
LEP-derived vessels are connected to the blood stream of the host. The scale bars
represent 21 microns.
Figure 4. EPC colony-forming assay comparing LEP, LEP Sca1+ and LEP Sca1-.
Phase contrast micrographs of Endothelial Progenitor Cell Colonies characterized by a
central cluster of round cells surrounded by radiating flat cells. Many colonies were
observed in the wells seeded with LEP Sca1+ (B; 56.5±5.5), less in the unseparated LEP
(A; 23.5±0.5) and almost none in LEP Sca1- (C; 3.5±0.5). Magnification 200x.
Figure 5. Comparisons of LEP Sca1+ and LEP Sca1-. RT-PCR analysis of
hematopoietic and vascular-specific gene expression.
Figure 6. Heat Map for differential gene expression of 62 angiogenesis-associated
genes comparing LEP Sca1+ and LEP Sca1-. We identified 445 differentially
expressed genes: 228 genes were up-regulated in the Sca1+ subset and 217 genes were
36

up-regulated in the Sca1- subset. We filtered this set by all genes associated with
angiogenesis in the literature and public databases. These 62 genes were then clustered
and relative expression levels are displayed in the heat map for the LEP Sca1+ vs. Sca1-
subsets as a color gradation from high (magenta) to low (aqua). The names of the anti-
angiogenic genes are highlighted in red to emphasize that 3 of the 6 differentially
expressed angiogenesis-associated genes in the Sca1- subset (see bottom of figure) are
anti-angiogenic.
37
Copyright © 2022 FDOKUMEN




















