
Effects of Cd injection on osmoregulation and stress indicators in freshwater 3 Nile tilapia 4 5
Upload
independentCategory
view
3download
0
ORIGINAL ARTICLE
Intraspecific variation in expression of candidategenes for osmoregulation, heme biosynthesisand stress resistance suggests local adaptationin European flounder (Platichthys flesus)
PF Larsen1,2, EE Nielsen1, TD Williams3 and V Loeschcke2
1Department of Inland Fisheries, Danish Institute for Fisheries Research, Technical University of Denmark, Silkeborg, Denmark;2Department of Biological Sciences, University of Aarhus, Aarhus, Denmark and 3School of Biosciences, The University of Birmingham,Birmingham, UK
Despite the recent discovery of significant genetic structuring ina large number of marine organisms, the evolutionarysignificance of these often minute genetic differences are stillpoorly understood. To elucidate the adaptive relevance of lowgenetic differentiation among marine fish populations, westudied expression differences of osmoregulatory and stressgenes in genetically weakly differentiated populations of theEuropean flounder (Platichthys flesus), distributed across anatural salinity gradient. Flounders were maintained in a long-term reciprocal transplantation experiment mimicking naturalsalinities in the North Sea and the Baltic Sea. Applying real-time quantitative PCR and microarray analysis we studiedexpression of four candidate genes (hsp70, angiotensinogen,Na/K-ATPase-a and 5-aminolevulinic acid synthase (ALAS)) ingill, kidney and liver tissues. Genes involved in osmoregulativeprocesses (Na/K-ATPases-a and angiotensinogen) showed
highly plastic but similar expression in the two populationsdependent on environmental salinity. However, we observeda unique sixfold up-regulation of hsp70 in kidney tissue offlounder from the North Sea following long-term acclimationto Baltic salinities. Similarly, significant differences betweenNorth Sea and Baltic flounders in expression of ALAS inresponse to different salinities were found in gill and livertissue. These findings strongly suggest that gene expressionin flounders is shaped by adaptation to local environmentalconditions. This identification of adaptive differences in highgene flow marine organisms adds a new dimension to ourcurrent understanding of evolutionary processes in the seaand is of paramount importance for identification, protectionand sustainable management of marine biodiversity.Heredity (2008) 101, 247–259; doi:10.1038/hdy.2008.54;published online 18 June 2008
Keywords: gene expression; quantitative PCR; local adaptation; genetic population structure; microarray
Introduction
Many species of marine fishes have large continuousdistributions in the oceans and are characterized byhaving high dispersal abilities both as juveniles andadults. In particular the free flowing pelagic phase ofeggs and larvae allows transport of these relativelypassive life stages over large distances by wind and/oroceanic currents. The implication of such a high geneflow scenario caused by the specific life history char-acteristics of ‘classical’ marine fishes is that the geneticdifferentiation is expected to be very limited comparedwith most other organisms (DeWoody and Avise, 2000).Low levels of genetic structuring in marine fishescompared with freshwater and anadromous fisheshas generally been confirmed by studies of geneticpopulation structure using different types of selectively
neutral genetic markers (reviewed by Ward et al.,1994; Waples, 1998). However, recent studies employinghighly variable neutral genetic markers, such as micro-satellites, have revealed small albeit statistically signifi-cant genetic differentiation among apparent localpopulations on a much smaller geographical scale thanpreviously assumed (Knutsen et al., 2003; Nielsen et al.,2004; Jorgensen et al., 2005). These relatively weaksignals of population structure have been suggested torepresent semi-independent evolutionary units capableof adapting to the local environment (Conover et al.,2006).
Marine fishes inhabit a diverse array of physical andbiological environments, favoring adaptive divergenceamong populations (Waples, 1998) and ultimately spe-ciation (Doebeli and Dieckmann, 2003). In general highlevels of neutral population differentiation for marinefishes has been shown to coincide with environmentalclines (Nielsen et al., 2003; Bekkevold et al., 2005;Hemmer-Hansen et al., 2007b) providing indirect evi-dence of local adaptations. Recently, genetic markersassociated with functional genes such as Pan I in cod(Pogson and Fevolden, 2003; Case et al., 2005) and heat-shock cognate gene (hsc70) in European flounders
Received 5 October 2007; revised 27 March 2008; accepted 26 April2008; published online 18 June 2008
Correspondence: Dr PF Larsen, Department of Inland Fisheries, DanishInstitute for Fisheries Research, Technical University of Denmark,Vejls�vej 39, Silkeborg, Silkeborg 8600, Denmark.E-mail: [email protected]
Heredity (2008) 101, 247–259& 2008 Macmillan Publishers Limited All rights reserved 0018-067X/08 $30.00
www.nature.com/hdy

(Hemmer-Hansen et al., 2007a) has demonstrated thatlevels of genetic differentiation associated with suchgenes are much higher than for neutral markers. Finally,regulatory variation in gene expression among estuarineand marine fishes has been shown to be pronouncedacross environmental gradients (Oleksiak et al., 2002;Picard and Schulte, 2004; Whitehead and Crawford,2006).
Salinity is expected to be one of the most importantenvironmental drivers for evolution in the oceans andsalinity gradients have been shown to coincide withsuspected adaptive population divergence in marinefishes (Nielsen et al., 2003, 2004). A very well-studiedsalinity gradient stretches from the high-saline North Seainto the brackish Baltic Sea (Johannesson and Andre,2006). Along a relatively short transect (400–500 km) thesalinity changes rapidly from marine conditions in theNorth Sea (33–35 p.p.t.) through the Kattegat, the DanishBelts and Øresund to brackish and almost freshwaterconditions in the northeastern part of the Baltic Sea(Johannesson and Andre, 2006). Recent populationgenetic studies have revealed relatively high levels ofgenetic structuring between marine fish populationssampled in the North Sea and the Baltic Sea, respectively,with FST values in cod (Gadus morhua), herring (Clupeaharengus) and turbot (Scophthalmus maximus) rangingfrom 2 to 4.5% (Nielsen et al., 2003, 2004; Bekkevold et al.,2005). Intraspecific differences in fitness-related repro-ductive traits were found for various populations ofmarine fish across the North Sea/Baltic Sea salinitygradient, for example, dab (Limanda limanda), plaice(Platichthys platessa), flounder (P. flesus) and Atlantic codin different saline environments (Nissling and Westin,1997; Nissling et al., 2002). Spermatozoa mobility, fertiliza-tion rates and salinity at which eggs became neutrallybuoyant were measured for different local populations ofmarine fish maintained at different salinities (Nissling andWestin, 1997; Nissling et al., 2002), mimicking naturalsalinities in the Baltic Sea and in the transition area.Following transfer of individuals from marine to brackishconditions these reproductive parameters remained es-sentially the same and their results strongly suggestedspecific adaptations to local salinities for these importantreproductive traits. These so-called ‘local adaptations’ aredefined as when individuals from one populationdemonstrate improved fitness in their native environmentcompared to other populations in the same environment(Kawecki and Ebert, 2004).
Recent population genetic studies conducted onEuropean flounder sampled across the North Sea/BalticSea transect have provided estimates of extremely lowgenetic differentiation (Larsen et al., 2007; Hemmer-Hansen et al., 2007b). Using variation in nine micro-satellite markers they found that flounders from the twoareas were almost genetically identical (FST ranging from0.005 to 0.006), thus indicating high levels of gene flowthereby reducing the likelihood of finding local adapta-tions within populations. However, despite the apparentlack of genetic differentiation Larsen et al. (2007) recentlyfound strong indications of local adaptations in overallpatterns of gene expression between flounder popula-tions in the North Sea and the Baltic Sea. Assessing geneexpression patterns in 3341 genes using microarrayanalysis they detected that 5% (158) of genes expressedin liver tissue were significantly differently expressed in
North Sea and Baltic Sea flounders subjected to the samesalinities under controlled conditions.
It is widely accepted that variation in gene expressionhas large functional consequences and that it is animportant component for environmental adaptationamong natural populations (Oleksiak et al., 2002; Babuand Aravind, 2006; Derome and Bernatchez, 2006). Littleeffort has, however, been diverted toward investigatinghow variation in gene expression could be related toosmoregulatory capacities or temperature toleranceunder salinity and temperature stress in natural fishpopulations. However, a recent study has demonstratedan apparent strong link between expression of heat-shock protein genes and fitness traits in fishes (Fangueet al., 2006). This study showed a tight coupling ofthermal tolerance with expression of hsp70 in gill tissueof killifish (Fundulus heteroclitus), originating from localpopulations inhabiting different thermal habitats (Fan-gue et al., 2006). Therefore, studies of the pattern andvariation in ‘candidate gene’ expression (for example,genes involved in temperature and salinity tolerance)among natural populations shows high potential forrevealing important local adaptations to different envir-onmental conditions in fishes.
In this study we wanted to test the hypothesis ofintraspecific variation in gene expression and adaptationto local marine conditions in European flounder sampledfrom the North Sea and the Baltic Sea. To do that westudied tissue-specific gene expression of four candidategenes for local adaptation and compare gene expressionprofiles from gill and kidney tissues with microarraydata for expression in liver tissue. Two candidate genesfor salinity adaptation, 5-aminolevulinic acid synthase(ALAS) and angiotensinogen, involved in heme biosynth-esis and blood pressure maintenance in fishes, respec-tively, were identified by Larsen et al. (2007) as the mostpromising candidate genes in a microarray study of geneexpression variation in liver tissue. ALAS was observedto be differently expressed between flounder populationsfrom the North Sea and the Baltic Sea, whereasangiotensinogen was identified as being highly plastic inexpression dependent on salinity acclimation (Larsenet al., 2007). Moreover two previously described andhighly likely candidate genes for local adaptation wereincluded in this study, hsp70 and Na/K-ATPase-a. Heat-shock proteins, and in particular hsp70, are known to beinduced by multiple stressors, including osmotic stress(Sorensen et al., 2003) and Na/K-ATPases are known tobe critical for maintaining ion balance and homeostasisof teleost fishes in relation to salinity acclimation (Evanset al., 2005).
We hypothesized that flounders originating from thehigh-saline North Sea and the brackish Baltic Sea arebetter adapted to their native saline environment than tononnative conditions. Accordingly, we expected to finddifferences in patterns of gene expression in floundersoriginating from the two populations. First we measuredhsp70 gene expression, which has been shown tobe highly correlated with fitness in other organisms(Sorensen et al., 2003; Fangue et al., 2006). Next wemeasured expression of genes involved in osmoregula-tive processes (Na/K-ATPase-a and angiotensinogen),where we expected high plasticity under different salineconditions, potentially explaining the high euryhalinityobserved for European flounder (Jensen et al., 2002).
Salinity-dependent gene expression in European flounderPF Larsen et al
248
Heredity

Finally we quantified the expression of ALAS to evaluateits expression in tissues directly involved in osmoregula-tion compared with the previous observations by Larsenet al. (2007) where analyses of liver tissue displayedpopulation differences. Gene expression responses fol-lowing long-term acclimation (50 days) as well as acute(1 day) were quantified to evaluate both initial and long-term regulation of stress and osmoregulative genes indifferent tissues. The results are discussed in relation topopulation structure and potential for local adaptationsin a classical high gene flow marine fish species.
Materials and methods
Tank experimentsImmature European flounders were collected usingdragnets from the North Sea at Skagen and in the BalticSea at the Island of Bornholm, Denmark (Figure 1) inAugust and September 2005. By the termination of theexperiment sampled flounders varied in length from 10.2to 14.5 cm and no significant difference in length ofsampled flounders was found between locations ortreatment-groups (one-way analysis of variance (ANO-VA): F3,18¼ 2.142, P¼ 0.1305). North Sea and Baltic Seaflounders were transported to the aquarium facilities atthe University of Aarhus, Denmark and held at10±0.5 1C in 1000-liter tanks containing recirculatedwater mimicking natural salinities of the North Sea(33±0.5 p.p.t.) and Baltic Sea (9±0.5 p.p.t.). All flounderswere maintained at a 16 h light/8 h dark cycle andallowed to acclimate to laboratory conditions for at least2 months at their natural salinity prior to starting theexperiments. Following acclimation, flounder from theNorth Sea and flounder from the Baltic Sea populationswere randomly assigned to a control group and a
reciprocal transplantation group with 5–6 individualsin each group (one tank for each salinity). In thetransplantation experiments tank water salinity wasgradually reduced from 33 to 9 p.p.t., or elevated from9 to 33 p.p.t. to end up at reciprocal salinities, over a 2 hperiod, to minimize acute stress responses.
An experimental design of reciprocal transplantationwas used to investigate differences in gene expressionbetween the two populations of flounders sampled.Acclimation was set to 50 days to assure full acclimation(Deane and Woo, 2004). By applying the reciprocaltransplantation experimental design the physical envir-onment can be standardized and allows for studyinggene expression variation depending on genetic originassociated with local adaptations (Mitchell-Olds andSchmitt, 2006). Finally, to evaluate whether flounderswere in fact stressed in response to salinity changesfollowing reciprocal transfer, we conducted a short-termexperiment, where flounders were sampled following 1day at nonnatural reciprocal salinities to study whetherflounders were in fact physiologically stressed followingtransplantation.
Flounders were initially fed with rainbow trout(Oncorhynchus myskiss) and mussels (Mytilus edulis) cutinto pieces, but later food was supplemented withcommercial trout pellets (BioMar, Denmark), up to 5days before they were killed. No acute stress effects, suchas behavioral changes or death, were evident within theexperimental period. All experiments were carried outaccording to Danish legislation concerning the use ofexperimental animals.
Tissue collection and RNA extractionFollowing termination of the tank experiment approxi-mately 150 mg of gill, kidney and liver tissue was
Figure 1 Map showing the flounder sampling locations Skagen and Bornholm.
Salinity-dependent gene expression in European flounderPF Larsen et al
249
Heredity
dissected and immediately placed in RNase-free Eppen-dorf tubes containing excess RNAlater following themanufacturer’s instructions (Qiagen, Hilden, Germany).RNAlater-preserved samples were stored at –20 1C untilbeing processed for RNA extraction.
Total RNA was extracted using the RNeasyminikit (Qiagen). Tissue (20 mg) was removed from theRNAlater preservation solution and immediatelyimmersed in 600ml of RLT buffer containingb-mercapthoethanol. The tissue was then homogenizedusing a Tissue-Tearor rotor homogenizer (BioSpecProducts, Bartlesville, OK, USA). Extractions wereperformed as recommended by the manufacturer exceptfor an additional DNAse treatment step for 15 min at25 1C to remove any remaining genomic DNA. Finally,total RNA was eluted from the Qiagen spincolumns using two times 30ml of RNAse-free waterand stored at –20 1C. Concentration of extractedRNA was determined at 260 nm in a standard Hellmacuvette (path length 10 mm) using the GeneQuantII (RNA/DNA Calculator; Pharmacia Biotech,Uppsala, Sweden). Total RNA quality was routinelyanalyzed using 2% agarose gel electrophoresisand applying capillary electrophoresis with the Agilent2100 bioanalyzer (Agilent Technologies, Palo Alto, CA,USA).
Reverse transcription of RNA and real-time quantitative
PCR
Reverse transcription of total RNA into cDNA wasperformed using the SuperScriptII RNase H-ReverseTranscriptase kit (Invitrogen, Carlsbad, CA, USA) in areaction volume of 20 ml containing 1� reaction buffer,5 mM MgCl2, 1 mM deoxyribonucleotide triphosphatemixture, 0.3 ml of anchored oligo-(dT)20 primer(2.5 mg ml�1), 1ml of SuperScriptII reverse transcriptaseand 3 mg of total RNA. Following reverse transcription,samples were diluted by 10-fold and stored at –20 1Cuntil analysis.
For the real-time quantitative PCR (qRT-PCR) analysisgene-specific primer pairs were designed using thePrimer-3 software (Rozen and Skaletsky, 2000) based onsequences available in GenBank. Primer sets weredeveloped for the following P. flesus genes: 5-aminole-vulinic acid synthase (ALAS) (GenBank accession no.AJ302076); ALAS-F 50-aggaacagcggtgctaagaa-30, ALAS-R,50-tctcaaaggccacgatcttc-30 product size 109 bp; angiotensi-nogen precursor (GenBank accession no. DV567747);angio-F 50-cccaggaggacaagctactg-30, angio-R 50-gctgctggagttcagctctt-30 product size 89 bp, hsp70 (GenBankaccession no. AF187726); hsp70-F 50-acaagagagctgtccgtcgt-30, hsp70-R 50-ctccctcatacagggagtcg-30 product size
NS 9 NS 33 BS 9 BS 33 NS 9 NS 33 BS 9 BS 33
NS 9 NS 33 BS 9 BS 33
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
Rel
ativ
e ge
ne e
xpre
ssio
n *
*
0
0.05
0.1
0.15
0.2
0.25
Rel
ativ
e ge
ne e
xpre
ssio
n
*
0
0.2
0.4
0.6
0.8
1
1.2
1.4
Rel
ativ
e ge
ne e
xpre
ssio
n
Na/K ATPase α Kidney Na/K ATPase α Gill
Na/K-ATPase α Liver
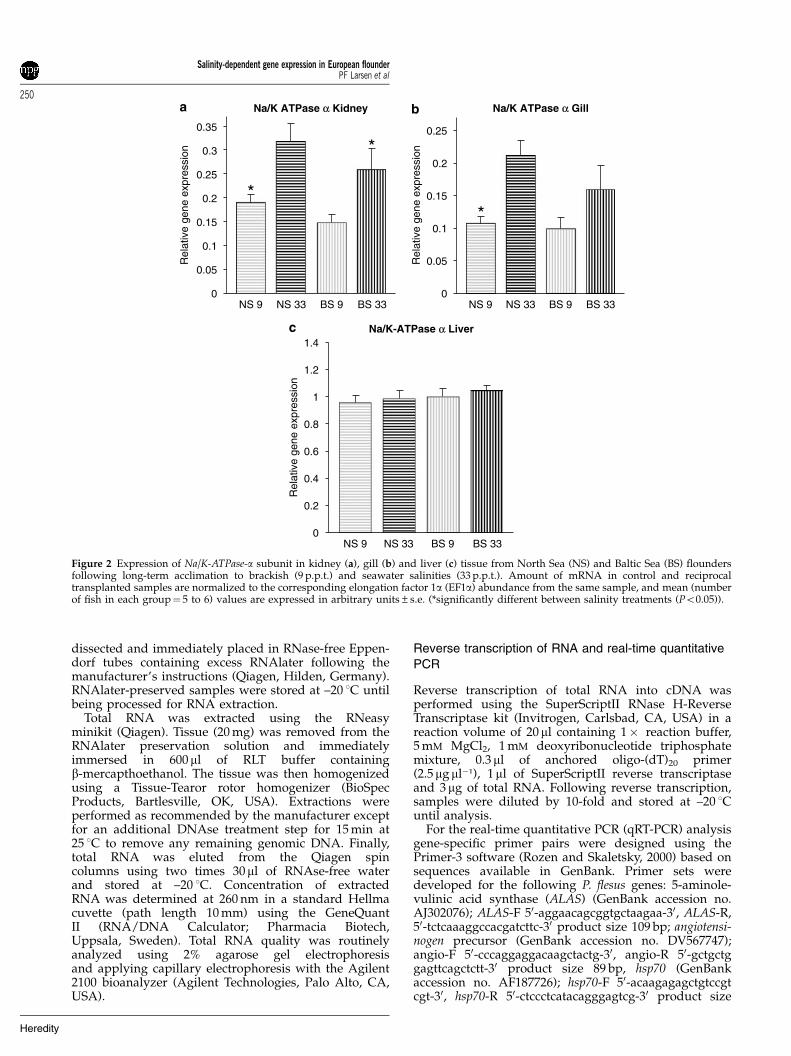
Figure 2 Expression of Na/K-ATPase-a subunit in kidney (a), gill (b) and liver (c) tissue from North Sea (NS) and Baltic Sea (BS) floundersfollowing long-term acclimation to brackish (9 p.p.t.) and seawater salinities (33 p.p.t.). Amount of mRNA in control and reciprocaltransplanted samples are normalized to the corresponding elongation factor 1a (EF1a) abundance from the same sample, and mean (numberof fish in each group¼ 5 to 6) values are expressed in arbitrary units±s.e. (*significantly different between salinity treatments (Po0.05)).
Salinity-dependent gene expression in European flounderPF Larsen et al
250
Heredity
103 bp, Na/K-ATPase-a subunit (GenBank accessionno. DV566003); NaATPa-F 50-aatcgcatcctcatctttgg-30,NaATPa-R 50-tggcttgaggggatacattc-30 product size105 bp and the housekeeping gene elongation factor 1a(EF1a) (GenBank accession no. EC378684); EF1a-F50-cccaggaggacaagctactg-30, EF1a-R 50-gctgctggagttcagctctt-30 product size 89 bp.
PCR products from all primer pairs were checkedon a 2% agarose gel to verify primer specificity andPCR product length and to ensure that they onlyproduced a single amplicon when flounder cDNAserved as template. Furthermore, dilution series wereperformed to verify amplification efficiency of primerpairs (Stahlberg et al., 2004).
qRT-PCR was performed on the Ligthcycler 1.2 (RocheDiagnostics, Mannheim, Germany) using SYBR Greenchemistry (LigthCycler FastStart DNA Master SYBRGreen I kit; Roche) and the Lightcycler relative quanti-fication software 3.5 (Roche Diagnostics). Using thissoftware we employed the fully automated method forCT determination called the ‘2nd Derivative Maximum’calculation. The CT value is the cycle where the secondderivative is at its maximum and ideally this shouldalways be in the heart of the log-linear portion of thereaction. Moreover this method is more objective and
reproducible than human-mediated crossing pointsettings (Lightcycler relative quantification softwaremanual). All qRT-PCR reactions were performedas follows: 10 min at 95 1C, followed by 40 cycles of95 1C for 15 s, 60 1C for 10 s and 72 1C for 10 s.Melting curve analysis was performed following eachreaction to confirm the presence of only a singleproduct in the reaction. Negative control reactionswere performed for all samples using RNA thathad not been reverse transcribed to control for thepossible presence of genomic DNA contamination.Genomic DNA contamination was present in mostsamples, but never constituted more than 1:3000 ofstarting cDNA copy numbers (data not shown). There-fore genomic DNA contamination reflects only a minorfraction of the final PCR product and only a minimumsource of error for the data analysis. In the presentpaper gene expression results for kidney and gill tissuesare produced from qRT-PCR analysis, whereasgene expression results for liver tissue are based onnormalized microarray data from the same fishes(microarray raw data are available from ArrayExpress,accession no. E-MAXD-22). Information on normaliza-tion and analysis of microarray results are available inLarsen et al. (2007).
Hsp70 Gill
0
0.0005
0.001
0.0015
0.002
Rel
ativ
e ge
ne e
xpre
ssio
n
Hsp70 Kidney
NS 9 NS 33 BS 9 BS 33 NS 9 NS 33 BS 9 BS 33
NS 9 NS 33 BS 9 BS 33
0
0.0005
0.001
0.0015
0.002
0.0025
Rel
ativ
e ge
ne e
xpre
ssio
n
* #
Hsp70 Liver
0
0.2
0.4
0.6
0.8
1
1.2
1.4
Rel
ativ
e ge
ne e
xpre
ssio
n
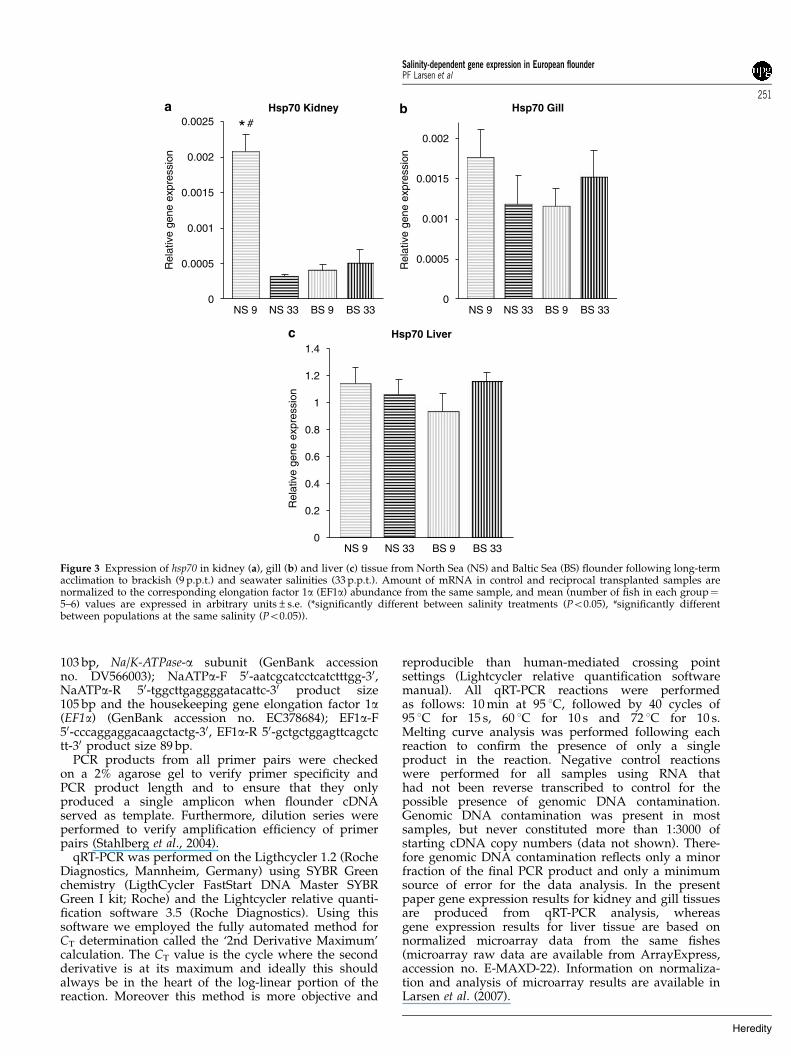
Figure 3 Expression of hsp70 in kidney (a), gill (b) and liver (c) tissue from North Sea (NS) and Baltic Sea (BS) flounder following long-termacclimation to brackish (9 p.p.t.) and seawater salinities (33 p.p.t.). Amount of mRNA in control and reciprocal transplanted samples arenormalized to the corresponding elongation factor 1a (EF1a) abundance from the same sample, and mean (number of fish in each group¼5–6) values are expressed in arbitrary units±s.e. (*significantly different between salinity treatments (Po0.05), #significantly differentbetween populations at the same salinity (Po0.05)).
Salinity-dependent gene expression in European flounderPF Larsen et al
251
Heredity
Statistical analysisRelative gene expression values were calculatedusing the ‘Comparative method’ according to Livakand Schmittgen, (2001). Data are presented as meanexpression levels ±s.e. Gene expression levelswere compared using one-way ANOVA (fixed effectsmodel), and multiple pairwise comparisons wereconducted using Tukeys test between salinities(within populations) or between populations(maintained at the same salinity). Finally, two-wayANOVA was conducted to test for an interaction effectof salinity treatment and population origin. Significancelevel was set at a¼ 0.05 in all analyses and analyseswere conducted using the PAST statistics software(Hammer et al., 2001).
Results
Long-term experimentNo mortality was observed throughout the period of theexperiment. In the long-term experiment the Na/K-ATPase-a subunit gene was observed to be expressed1.5- to 2-fold higher at high salinities than at lowsalinities in kidney and gill tissues from both popula-tions (Figures 2a and b). For North Sea flounder this
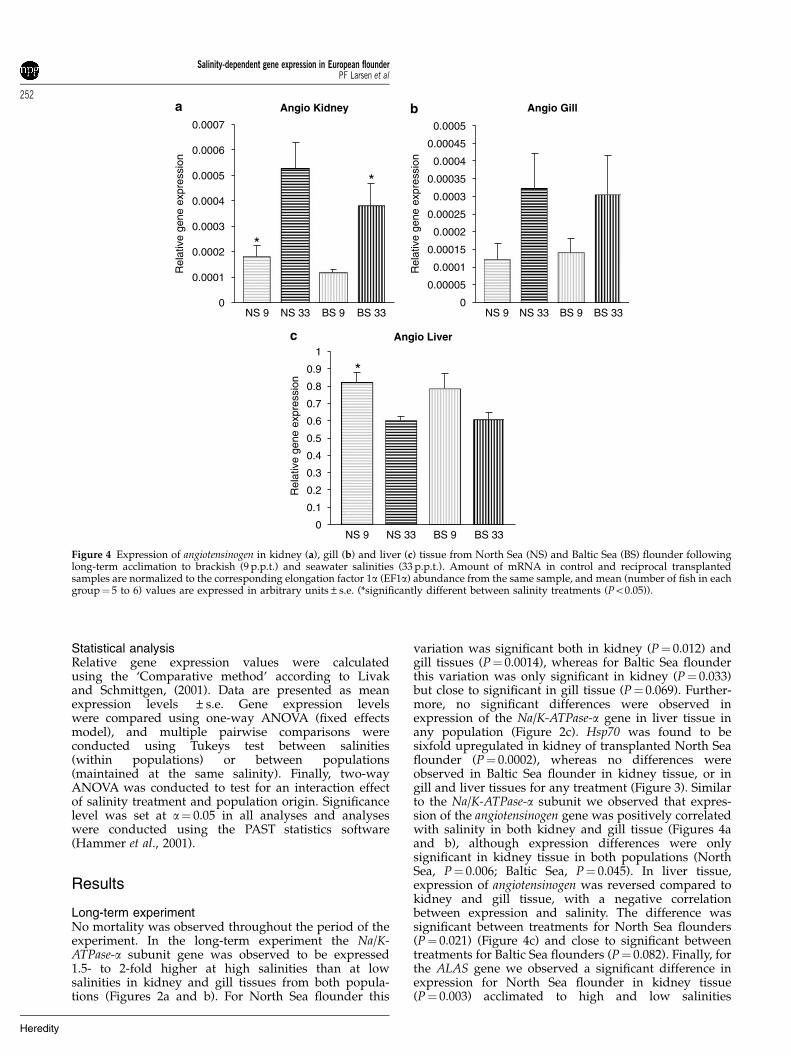
variation was significant both in kidney (P¼ 0.012) andgill tissues (P¼ 0.0014), whereas for Baltic Sea flounderthis variation was only significant in kidney (P¼ 0.033)but close to significant in gill tissue (P¼ 0.069). Further-more, no significant differences were observed inexpression of the Na/K-ATPase-a gene in liver tissue inany population (Figure 2c). Hsp70 was found to besixfold upregulated in kidney of transplanted North Seaflounder (P¼ 0.0002), whereas no differences wereobserved in Baltic Sea flounder in kidney tissue, or ingill and liver tissues for any treatment (Figure 3). Similarto the Na/K-ATPase-a subunit we observed that expres-sion of the angiotensinogen gene was positively correlatedwith salinity in both kidney and gill tissue (Figures 4aand b), although expression differences were onlysignificant in kidney tissue in both populations (NorthSea, P¼ 0.006; Baltic Sea, P¼ 0.045). In liver tissue,expression of angiotensinogen was reversed compared tokidney and gill tissue, with a negative correlationbetween expression and salinity. The difference wassignificant between treatments for North Sea flounders(P¼ 0.021) (Figure 4c) and close to significant betweentreatments for Baltic Sea flounders (P¼ 0.082). Finally, forthe ALAS gene we observed a significant difference inexpression for North Sea flounder in kidney tissue(P¼ 0.003) acclimated to high and low salinities
Angio Kidney
NS 9 NS 33 BS 9 BS 33 NS 9 NS 33 BS 9 BS 33
NS 9 NS 33 BS 9 BS 33
0
0.0001
0.0002
0.0003
0.0004
0.0005
0.0006
0.0007
Rel
ativ
e ge
ne e
xpre
ssio
n*
*
Angio Gill
0
0.00005
0.0001
0.00015
0.0002
0.00025
0.0003
0.00035
0.0004
0.00045
0.0005
Rel
ativ
e ge
ne e
xpre
ssio
n
Angio Liver
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
Rel
ativ
e ge
ne e
xpre
ssio
n
*
Figure 4 Expression of angiotensinogen in kidney (a), gill (b) and liver (c) tissue from North Sea (NS) and Baltic Sea (BS) flounder followinglong-term acclimation to brackish (9 p.p.t.) and seawater salinities (33 p.p.t.). Amount of mRNA in control and reciprocal transplantedsamples are normalized to the corresponding elongation factor 1a (EF1a) abundance from the same sample, and mean (number of fish in eachgroup¼ 5 to 6) values are expressed in arbitrary units±s.e. (*significantly different between salinity treatments (Po0.05)).
Salinity-dependent gene expression in European flounderPF Larsen et al
252
Heredity
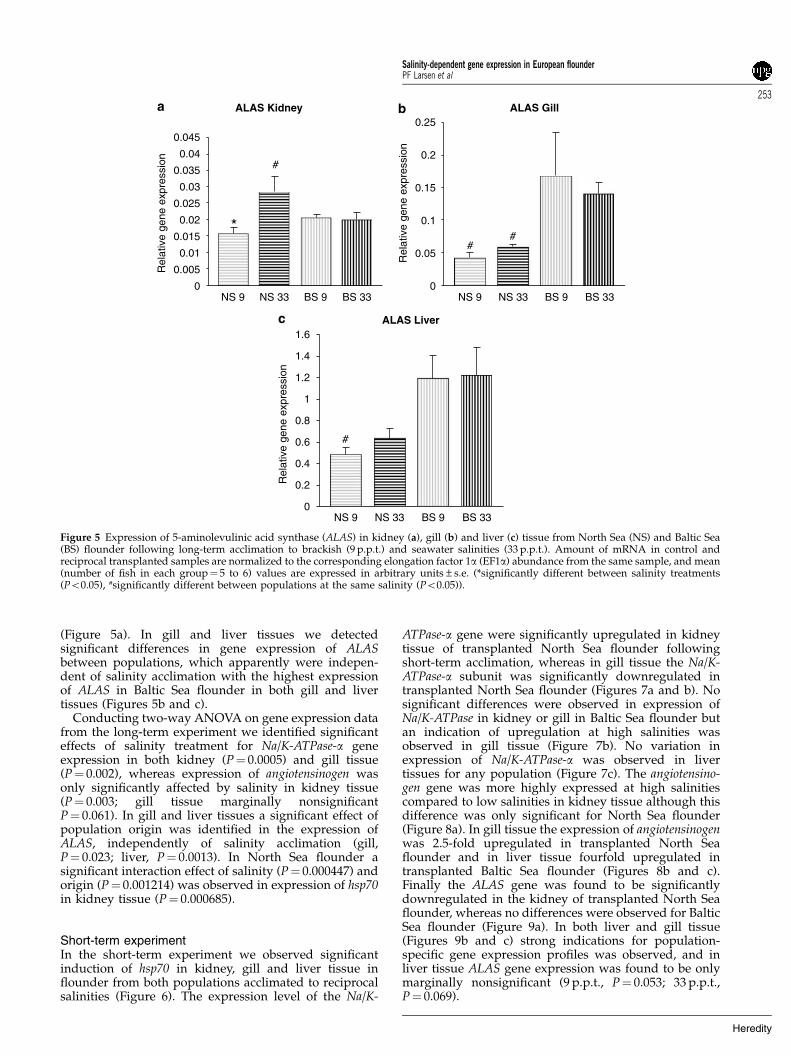
(Figure 5a). In gill and liver tissues we detectedsignificant differences in gene expression of ALASbetween populations, which apparently were indepen-dent of salinity acclimation with the highest expressionof ALAS in Baltic Sea flounder in both gill and livertissues (Figures 5b and c).
Conducting two-way ANOVA on gene expression datafrom the long-term experiment we identified significanteffects of salinity treatment for Na/K-ATPase-a geneexpression in both kidney (P¼ 0.0005) and gill tissue(P¼ 0.002), whereas expression of angiotensinogen wasonly significantly affected by salinity in kidney tissue(P¼ 0.003; gill tissue marginally nonsignificantP¼ 0.061). In gill and liver tissues a significant effect ofpopulation origin was identified in the expression ofALAS, independently of salinity acclimation (gill,P¼ 0.023; liver, P¼ 0.0013). In North Sea flounder asignificant interaction effect of salinity (P¼ 0.000447) andorigin (P¼ 0.001214) was observed in expression of hsp70in kidney tissue (P¼ 0.000685).
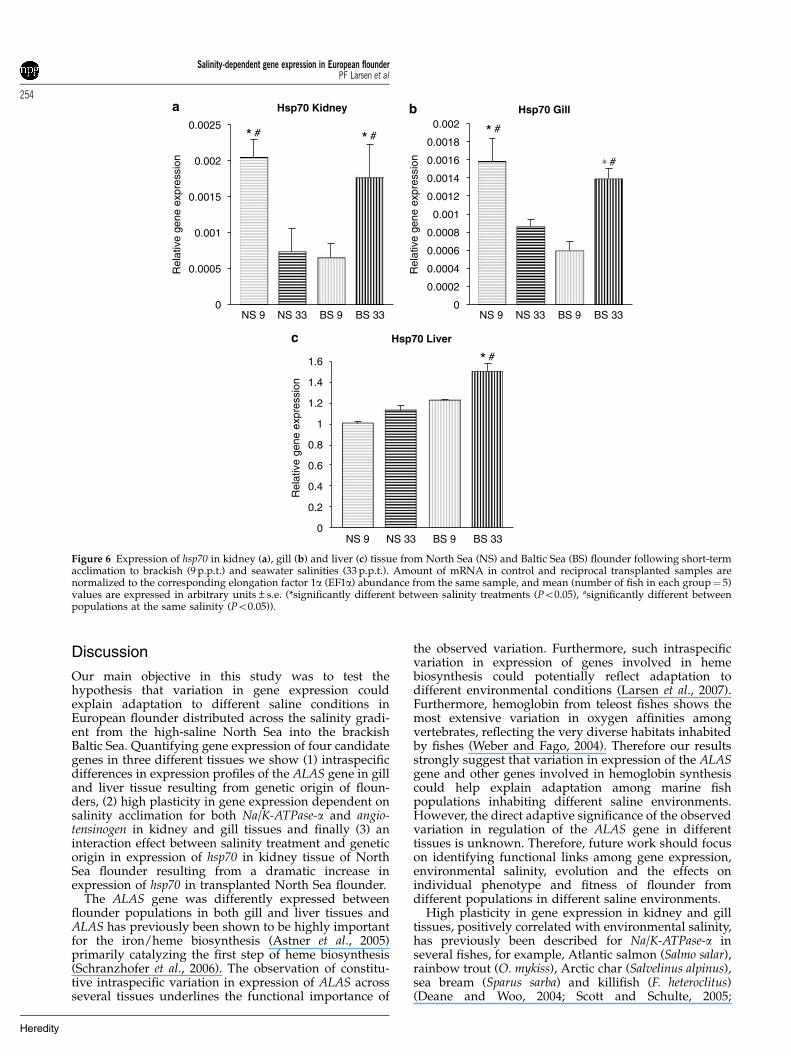
Short-term experimentIn the short-term experiment we observed significantinduction of hsp70 in kidney, gill and liver tissue inflounder from both populations acclimated to reciprocalsalinities (Figure 6). The expression level of the Na/K-
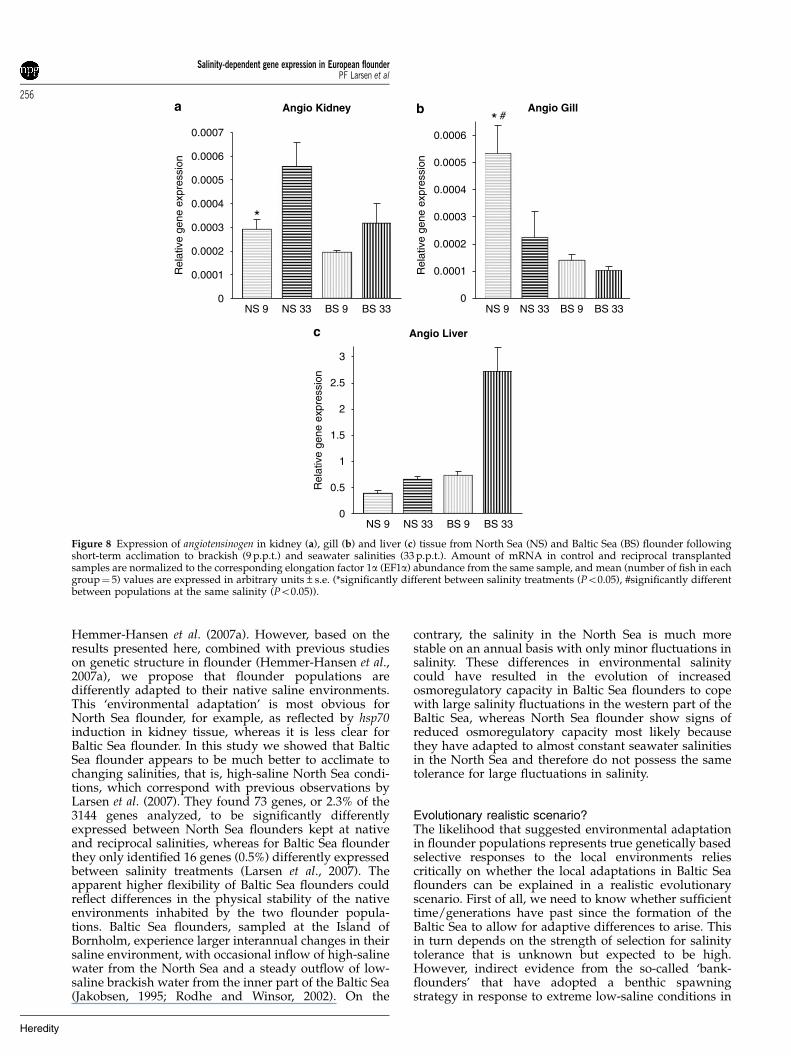
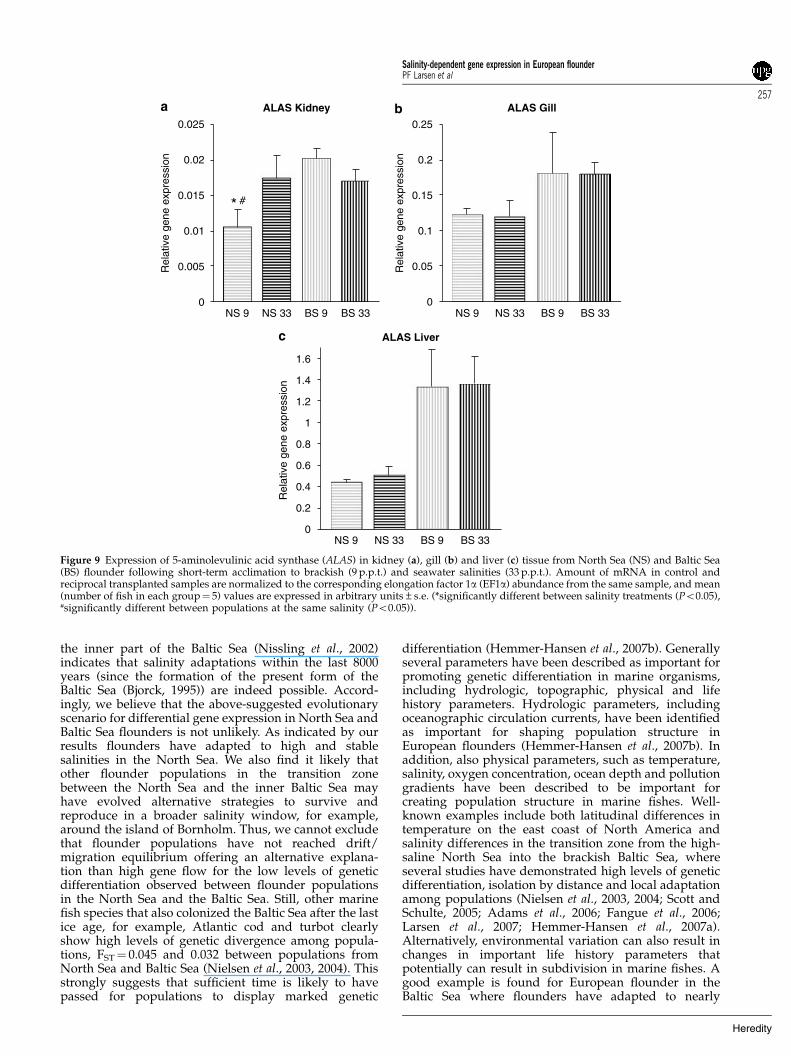
ATPase-a gene were significantly upregulated in kidneytissue of transplanted North Sea flounder followingshort-term acclimation, whereas in gill tissue the Na/K-ATPase-a subunit was significantly downregulated intransplanted North Sea flounder (Figures 7a and b). Nosignificant differences were observed in expression ofNa/K-ATPase in kidney or gill in Baltic Sea flounder butan indication of upregulation at high salinities wasobserved in gill tissue (Figure 7b). No variation inexpression of Na/K-ATPase-a was observed in livertissues for any population (Figure 7c). The angiotensino-gen gene was more highly expressed at high salinitiescompared to low salinities in kidney tissue although thisdifference was only significant for North Sea flounder(Figure 8a). In gill tissue the expression of angiotensinogenwas 2.5-fold upregulated in transplanted North Seaflounder and in liver tissue fourfold upregulated intransplanted Baltic Sea flounder (Figures 8b and c).Finally the ALAS gene was found to be significantlydownregulated in the kidney of transplanted North Seaflounder, whereas no differences were observed for BalticSea flounder (Figure 9a). In both liver and gill tissue(Figures 9b and c) strong indications for population-specific gene expression profiles was observed, and inliver tissue ALAS gene expression was found to be onlymarginally nonsignificant (9 p.p.t., P¼ 0.053; 33 p.p.t.,P¼ 0.069).
ALAS Kidney
NS 9 NS 33 BS 9 BS 33 NS 9 NS 33 BS 9 BS 33
NS 9 NS 33 BS 9 BS 33
0
0.005
0.01
0.015
0.02
0.025
0.03
0.035
0.04
0.045
Rel
ativ
e ge
ne e
xpre
ssio
n
*
#
ALAS Gill
0
0.05
0.1
0.15
0.2
0.25
Rel
ativ
e ge
ne e
xpre
ssio
n
##
ALAS Liver
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
Rel
ativ
e ge
ne e
xpre
ssio
n
#
Figure 5 Expression of 5-aminolevulinic acid synthase (ALAS) in kidney (a), gill (b) and liver (c) tissue from North Sea (NS) and Baltic Sea(BS) flounder following long-term acclimation to brackish (9 p.p.t.) and seawater salinities (33 p.p.t.). Amount of mRNA in control andreciprocal transplanted samples are normalized to the corresponding elongation factor 1a (EF1a) abundance from the same sample, and mean(number of fish in each group¼ 5 to 6) values are expressed in arbitrary units±s.e. (*significantly different between salinity treatments(Po0.05), #significantly different between populations at the same salinity (Po0.05)).
Salinity-dependent gene expression in European flounderPF Larsen et al
253
Heredity
Discussion
Our main objective in this study was to test thehypothesis that variation in gene expression couldexplain adaptation to different saline conditions inEuropean flounder distributed across the salinity gradi-ent from the high-saline North Sea into the brackishBaltic Sea. Quantifying gene expression of four candidategenes in three different tissues we show (1) intraspecificdifferences in expression profiles of the ALAS gene in gilland liver tissue resulting from genetic origin of floun-ders, (2) high plasticity in gene expression dependent onsalinity acclimation for both Na/K-ATPase-a and angio-tensinogen in kidney and gill tissues and finally (3) aninteraction effect between salinity treatment and geneticorigin in expression of hsp70 in kidney tissue of NorthSea flounder resulting from a dramatic increase inexpression of hsp70 in transplanted North Sea flounder.
The ALAS gene was differently expressed betweenflounder populations in both gill and liver tissues andALAS has previously been shown to be highly importantfor the iron/heme biosynthesis (Astner et al., 2005)primarily catalyzing the first step of heme biosynthesis(Schranzhofer et al., 2006). The observation of constitu-tive intraspecific variation in expression of ALAS acrossseveral tissues underlines the functional importance of
the observed variation. Furthermore, such intraspecificvariation in expression of genes involved in hemebiosynthesis could potentially reflect adaptation todifferent environmental conditions (Larsen et al., 2007).Furthermore, hemoglobin from teleost fishes shows themost extensive variation in oxygen affinities amongvertebrates, reflecting the very diverse habitats inhabitedby fishes (Weber and Fago, 2004). Therefore our resultsstrongly suggest that variation in expression of the ALASgene and other genes involved in hemoglobin synthesiscould help explain adaptation among marine fishpopulations inhabiting different saline environments.However, the direct adaptive significance of the observedvariation in regulation of the ALAS gene in differenttissues is unknown. Therefore, future work should focuson identifying functional links among gene expression,environmental salinity, evolution and the effects onindividual phenotype and fitness of flounder fromdifferent populations in different saline environments.
High plasticity in gene expression in kidney and gilltissues, positively correlated with environmental salinity,has previously been described for Na/K-ATPase-a inseveral fishes, for example, Atlantic salmon (Salmo salar),rainbow trout (O. mykiss), Arctic char (Salvelinus alpinus),sea bream (Sparus sarba) and killifish (F. heteroclitus)(Deane and Woo, 2004; Scott and Schulte, 2005;
Hsp70 Kidney
Rel
ativ
e ge
ne e
xpre
ssio
n
Rel
ativ
e ge
ne e
xpre
ssio
n
Rel
ativ
e ge
ne e
xpre
ssio
n
Hsp70 Gill
∗ #
* #* #* #
* #
Hsp70 Liver
0.0025
0.002
0.0015
0.001
0.0005
0NS 9 NS 33 BS 9 BS 33 NS 9 NS 33 BS 9 BS 33
NS 9 NS 33 BS 9 BS 33
1.6
1.4
1.2
1
0.8
0.6
0.4
0.2
0
0.002
0.0018
0.0016
0.0014
0.0012
0.001
0.0008
0.0006
0.0004
0.0002
0
Figure 6 Expression of hsp70 in kidney (a), gill (b) and liver (c) tissue from North Sea (NS) and Baltic Sea (BS) flounder following short-termacclimation to brackish (9 p.p.t.) and seawater salinities (33 p.p.t.). Amount of mRNA in control and reciprocal transplanted samples arenormalized to the corresponding elongation factor 1a (EF1a) abundance from the same sample, and mean (number of fish in each group¼ 5)values are expressed in arbitrary units±s.e. (*significantly different between salinity treatments (Po0.05), #significantly different betweenpopulations at the same salinity (Po0.05)).
Salinity-dependent gene expression in European flounderPF Larsen et al
254
Heredity
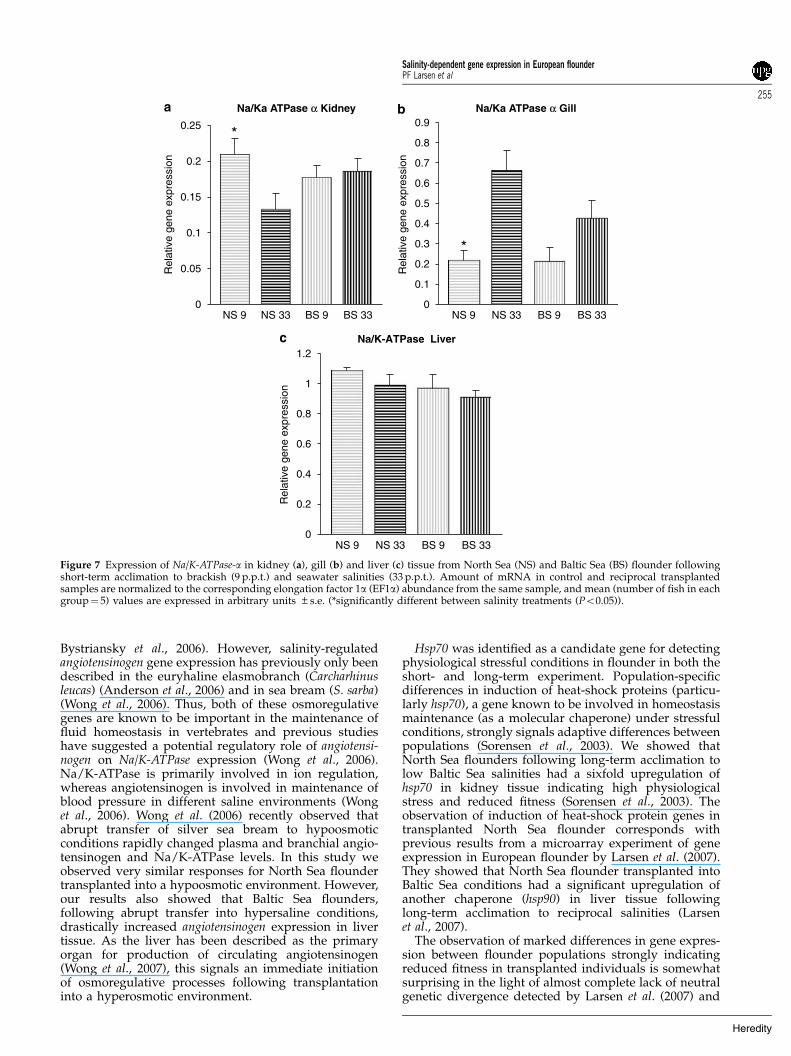
Bystriansky et al., 2006). However, salinity-regulatedangiotensinogen gene expression has previously only beendescribed in the euryhaline elasmobranch (Carcharhinusleucas) (Anderson et al., 2006) and in sea bream (S. sarba)(Wong et al., 2006). Thus, both of these osmoregulativegenes are known to be important in the maintenance offluid homeostasis in vertebrates and previous studieshave suggested a potential regulatory role of angiotensi-nogen on Na/K-ATPase expression (Wong et al., 2006).Na/K-ATPase is primarily involved in ion regulation,whereas angiotensinogen is involved in maintenance ofblood pressure in different saline environments (Wonget al., 2006). Wong et al. (2006) recently observed thatabrupt transfer of silver sea bream to hypoosmoticconditions rapidly changed plasma and branchial angio-tensinogen and Na/K-ATPase levels. In this study weobserved very similar responses for North Sea floundertransplanted into a hypoosmotic environment. However,our results also showed that Baltic Sea flounders,following abrupt transfer into hypersaline conditions,drastically increased angiotensinogen expression in livertissue. As the liver has been described as the primaryorgan for production of circulating angiotensinogen(Wong et al., 2007), this signals an immediate initiationof osmoregulative processes following transplantationinto a hyperosmotic environment.
Hsp70 was identified as a candidate gene for detectingphysiological stressful conditions in flounder in both theshort- and long-term experiment. Population-specificdifferences in induction of heat-shock proteins (particu-larly hsp70), a gene known to be involved in homeostasismaintenance (as a molecular chaperone) under stressfulconditions, strongly signals adaptive differences betweenpopulations (Sorensen et al., 2003). We showed thatNorth Sea flounders following long-term acclimation tolow Baltic Sea salinities had a sixfold upregulation ofhsp70 in kidney tissue indicating high physiologicalstress and reduced fitness (Sorensen et al., 2003). Theobservation of induction of heat-shock protein genes intransplanted North Sea flounder corresponds withprevious results from a microarray experiment of geneexpression in European flounder by Larsen et al. (2007).They showed that North Sea flounder transplanted intoBaltic Sea conditions had a significant upregulation ofanother chaperone (hsp90) in liver tissue followinglong-term acclimation to reciprocal salinities (Larsenet al., 2007).
The observation of marked differences in gene expres-sion between flounder populations strongly indicatingreduced fitness in transplanted individuals is somewhatsurprising in the light of almost complete lack of neutralgenetic divergence detected by Larsen et al. (2007) and
NS 9 NS 33 BS 9 BS 330
0.05
0.1
0.15
0.2
0.25
Rel
ativ
e ge
ne e
xpre
ssio
n
*
NS 9 NS 33 BS 9 BS 33
NS 9 NS 33 BS 9 BS 33
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Rel
ativ
e ge
ne e
xpre
ssio
n
*
Na/K-ATPase Liver
0
0.2
0.4
0.6
0.8
1
1.2
Rel
ativ
e ge
ne e
xpre
ssio
n
Na/Ka ATPase α Kidney Na/Ka ATPase α Gill
Figure 7 Expression of Na/K-ATPase-a in kidney (a), gill (b) and liver (c) tissue from North Sea (NS) and Baltic Sea (BS) flounder followingshort-term acclimation to brackish (9 p.p.t.) and seawater salinities (33 p.p.t.). Amount of mRNA in control and reciprocal transplantedsamples are normalized to the corresponding elongation factor 1a (EF1a) abundance from the same sample, and mean (number of fish in eachgroup¼ 5) values are expressed in arbitrary units ±s.e. (*significantly different between salinity treatments (Po0.05)).
Salinity-dependent gene expression in European flounderPF Larsen et al
255
Heredity
Hemmer-Hansen et al. (2007a). However, based on theresults presented here, combined with previous studieson genetic structure in flounder (Hemmer-Hansen et al.,2007a), we propose that flounder populations aredifferently adapted to their native saline environments.This ‘environmental adaptation’ is most obvious forNorth Sea flounder, for example, as reflected by hsp70induction in kidney tissue, whereas it is less clear forBaltic Sea flounder. In this study we showed that BalticSea flounder appears to be much better to acclimate tochanging salinities, that is, high-saline North Sea condi-tions, which correspond with previous observations byLarsen et al. (2007). They found 73 genes, or 2.3% of the3144 genes analyzed, to be significantly differentlyexpressed between North Sea flounders kept at nativeand reciprocal salinities, whereas for Baltic Sea flounderthey only identified 16 genes (0.5%) differently expressedbetween salinity treatments (Larsen et al., 2007). Theapparent higher flexibility of Baltic Sea flounders couldreflect differences in the physical stability of the nativeenvironments inhabited by the two flounder popula-tions. Baltic Sea flounders, sampled at the Island ofBornholm, experience larger interannual changes in theirsaline environment, with occasional inflow of high-salinewater from the North Sea and a steady outflow of low-saline brackish water from the inner part of the Baltic Sea(Jakobsen, 1995; Rodhe and Winsor, 2002). On the
contrary, the salinity in the North Sea is much morestable on an annual basis with only minor fluctuations insalinity. These differences in environmental salinitycould have resulted in the evolution of increasedosmoregulatory capacity in Baltic Sea flounders to copewith large salinity fluctuations in the western part of theBaltic Sea, whereas North Sea flounder show signs ofreduced osmoregulatory capacity most likely becausethey have adapted to almost constant seawater salinitiesin the North Sea and therefore do not possess the sametolerance for large fluctuations in salinity.
Evolutionary realistic scenario?The likelihood that suggested environmental adaptationin flounder populations represents true genetically basedselective responses to the local environments reliescritically on whether the local adaptations in Baltic Seaflounders can be explained in a realistic evolutionaryscenario. First of all, we need to know whether sufficienttime/generations have past since the formation of theBaltic Sea to allow for adaptive differences to arise. Thisin turn depends on the strength of selection for salinitytolerance that is unknown but expected to be high.However, indirect evidence from the so-called ‘bank-flounders’ that have adopted a benthic spawningstrategy in response to extreme low-saline conditions in
Angio Kidney
NS 9 NS 33 BS 9 BS 330
0.0001
0.0002
0.0003
0.0004
0.0005
0.0006
0.0007
Rel
ativ
e ge
ne e
xpre
ssio
n
*
Angio Gill
NS 9 NS 33 BS 9 BS 33
NS 9 NS 33 BS 9 BS 33
0
0.0001
0.0002
0.0003
0.0004
0.0005
0.0006
Rel
ativ
e ge
ne e
xpre
ssio
n
* #
Angio Liver
0
0.5
1
1.5
2
2.5
3
Rel
ativ
e ge
ne e
xpre
ssio
n
Figure 8 Expression of angiotensinogen in kidney (a), gill (b) and liver (c) tissue from North Sea (NS) and Baltic Sea (BS) flounder followingshort-term acclimation to brackish (9 p.p.t.) and seawater salinities (33 p.p.t.). Amount of mRNA in control and reciprocal transplantedsamples are normalized to the corresponding elongation factor 1a (EF1a) abundance from the same sample, and mean (number of fish in eachgroup¼ 5) values are expressed in arbitrary units±s.e. (*significantly different between salinity treatments (Po0.05), #significantly differentbetween populations at the same salinity (Po0.05)).
Salinity-dependent gene expression in European flounderPF Larsen et al
256
Heredity
the inner part of the Baltic Sea (Nissling et al., 2002)indicates that salinity adaptations within the last 8000years (since the formation of the present form of theBaltic Sea (Bjorck, 1995)) are indeed possible. Accord-ingly, we believe that the above-suggested evolutionaryscenario for differential gene expression in North Sea andBaltic Sea flounders is not unlikely. As indicated by ourresults flounders have adapted to high and stablesalinities in the North Sea. We also find it likely thatother flounder populations in the transition zonebetween the North Sea and the inner Baltic Sea mayhave evolved alternative strategies to survive andreproduce in a broader salinity window, for example,around the island of Bornholm. Thus, we cannot excludethat flounder populations have not reached drift/migration equilibrium offering an alternative explana-tion than high gene flow for the low levels of geneticdifferentiation observed between flounder populationsin the North Sea and the Baltic Sea. Still, other marinefish species that also colonized the Baltic Sea after the lastice age, for example, Atlantic cod and turbot clearlyshow high levels of genetic divergence among popula-tions, FST¼ 0.045 and 0.032 between populations fromNorth Sea and Baltic Sea (Nielsen et al., 2003, 2004). Thisstrongly suggests that sufficient time is likely to havepassed for populations to display marked genetic
differentiation (Hemmer-Hansen et al., 2007b). Generallyseveral parameters have been described as important forpromoting genetic differentiation in marine organisms,including hydrologic, topographic, physical and lifehistory parameters. Hydrologic parameters, includingoceanographic circulation currents, have been identifiedas important for shaping population structure inEuropean flounders (Hemmer-Hansen et al., 2007b). Inaddition, also physical parameters, such as temperature,salinity, oxygen concentration, ocean depth and pollutiongradients have been described to be important forcreating population structure in marine fishes. Well-known examples include both latitudinal differences intemperature on the east coast of North America andsalinity differences in the transition zone from the high-saline North Sea into the brackish Baltic Sea, whereseveral studies have demonstrated high levels of geneticdifferentiation, isolation by distance and local adaptationamong populations (Nielsen et al., 2003, 2004; Scott andSchulte, 2005; Adams et al., 2006; Fangue et al., 2006;Larsen et al., 2007; Hemmer-Hansen et al., 2007a).Alternatively, environmental variation can also result inchanges in important life history parameters thatpotentially can result in subdivision in marine fishes. Agood example is found for European flounder in theBaltic Sea where flounders have adapted to nearly
ALAS Kidney
NS 9 NS 33 BS 9 BS 33 NS 9 NS 33 BS 9 BS 33
NS 9 NS 33 BS 9 BS 33
0
0.005
0.01
0.015
0.02
0.025
Rel
ativ
e ge
ne e
xpre
ssio
n
* #
ALAS Gill
0
0.05
0.1
0.15
0.2
0.25
Rel
ativ
e ge
ne e
xpre
ssio
n
ALAS Liver
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
Rel
ativ
e ge
ne e
xpre
ssio
n
Figure 9 Expression of 5-aminolevulinic acid synthase (ALAS) in kidney (a), gill (b) and liver (c) tissue from North Sea (NS) and Baltic Sea(BS) flounder following short-term acclimation to brackish (9 p.p.t.) and seawater salinities (33 p.p.t.). Amount of mRNA in control andreciprocal transplanted samples are normalized to the corresponding elongation factor 1a (EF1a) abundance from the same sample, and mean(number of fish in each group¼ 5) values are expressed in arbitrary units±s.e. (*significantly different between salinity treatments (Po0.05),#significantly different between populations at the same salinity (Po0.05)).
Salinity-dependent gene expression in European flounderPF Larsen et al
257
Heredity

freshwater condition in the most northern part of theBaltic Sea. In this area flounders have been reported tohave changed the spawning strategy from pelagic tobenthic (Nissling et al., 2002). Under marine conditionsflounders are known to spawn pelagic eggs, thus, theso-called ‘bank’ flounders tend to spawn benthic eggs onshallow water most likely due to low salinity resulting inthe eggs sinking to unfavorable oxygen conditions on theocean bottom (Nissling et al., 2002). Accordingly, a majorgenetic barrier has recently been demonstrated betweenthe two life history forms in the Baltic Sea (Hemmer-Hansen et al., 2007a).
For this study flounders were caught as juveniles, withno differences in size, and they were treated identicallyin the laboratory. Furthermore, high heritability (Schadtet al., 2003; Brem and Kruglyak, 2005) and adaptivedivergence (Whitehead and Crawford, 2006; Larsen et al.,2007) is generally observed for gene expression profiles.Therefore, we find it unlikely that nongenetic factors,such as maternal, early developmental or epigeneticeffects arising from interplay between genetic andenvironmental components should affect gene regulationwhen individuals are maintained in a long-term recipro-cal transplantation setup.
The observation of stress responses in floundertransplanted to nonnative saline habitats in this studyand previously described by Larsen et al. (2007) hasimportant bearings in relation to a future scenario ofglobal change. Increased rainfall is expected to lower thesalinity in the Baltic Sea in the future imposing evenfurther salinity stress on the Baltic population. Therefore,this study underlines the importance of understandingpopulation structure in the sea, not only focusing onneutral markers presenting information about drift andmigration, but also focusing on genes and expression ofgenes involved in local adaptation and stress responses.Further knowledge on the presence of local adaptationamong marine fish populations will increase ourpossibility to properly manage marine fishes, andimprove our ability to predict the distribution andabundance of marine species in the light of the changingevolutionary forces imposed by global change.
Acknowledgements
We thank Palle Holm Hansen for help with fishmaintenance, Karen-Lise Dons Mensberg and DorteMeldrup for assistance in the laboratory and HenrikBaktoft for graphical assistance. This project wassupported by grants to Peter Foged Larsen from theSLIP research school under the Danish Network forFisheries and Aquaculture Research financed by theDanish Ministry for Food, Agriculture and Fisheries andthe Danish Agricultural and Veterinary Research Coun-cil, the Danish Institute for Fisheries Research and theElisabeth and Knud Petersens Foundation.
References
Adams SM, Lindmeier JB, Duvernell DD (2006). Microsatelliteanalysis of the phylogeography, Pleistocene history andsecondary contact hypotheses for the killifish, Fundulusheteroclitus. Mol Ecol 15: 1109–1123.
Anderson WG, Pillans RD, Hyodo S, Tsukada T, Good JP, TakeiY et al. (2006). The effects of freshwater to seawater transfer
on circulating levels of angiotensin II, C-type natriureticpeptide and arginine vasotocin in the euryhaline elasmo-branch, Carcharhinus leucas. Gen Comp Endocrinol 147: 39–46.
Astner I, Schulze JO, van den Heuvel J, Jahn D, Schubert WD,Heinz DW (2005). Crystal structure of 5-aminolevulinatesynthase, the first enzyme of heme biosynthesis, and its linkto XLSA in humans. EMBO J 24: 3166–3177.
Babu MM, Aravind L (2006). Adaptive evolution by optimizingexpression levels in different environments. Trends Microbiol14: 11–14.
Bekkevold D, Andre C, Dahlgren TG, Clausen LA, TorstensenE, Mosegaard H et al. (2005). Environmental correlates ofpopulation differentiation in Atlantic herring. Evolution 59:2656–2668.
Bjorck S (1995). A review of the history of the Baltic Sea,13.0–8.0 Ka Bp. Quat Int 27: 19–40.
Brem RB, Kruglyak L (2005). The landscape of genetic complex-ity across 5,700 gene expression traits in yeast. Proc Natl AcadSci USA 102: 1572–1577.
Bystriansky JS, Richards JG, Schulte PM, Ballantyne JS (2006).Reciprocal expression of gill Na+/K+�ATPase alpha-subunitisoforms alpha 1a and alpha 1b during seawater acclimationof three salmonid fishes that vary in their salinity tolerance.J Exp Biol 209: 1848–1858.
Case RAJ, Hutchinson WF, Hauser L, Van Oosterhout C,Carvalho GR (2005). Macro- and micro-geographic variationin pantophysin (PanI) allele frequencies in NE Atlantic codGadus morhua. Mar Ecol Prog Ser 301: 267–278.
Conover DO, Clarke LM, Munch SB, Wagner GN (2006).Spatial and temporal scales of adaptive divergence in marinefishes and the implications for conservation. J Fish Biol 69:21–47.
Deane EE, Woo NYS (2004). Differential gene expressionassociated with euryhalinity in sea bream (Sparus sarba).Am J Physiol Regul Integr Comp Physiol 287: R1054–R1063.
Derome N, Bernatchez L (2006). The transcriptomics ofecological convergence between 2 limnetic coregonine fishes(Salmonidae). Mol Biol Evol 23: 2370–2378.
DeWoody JA, Avise JC (2000). Microsatellite variation inmarine, freshwater and anadromous fishes compared withother animals. J Fish Biol 56: 461–473.
Doebeli M, Dieckmann U (2003). Speciation along environ-mental gradients. Nature 421: 259–264.
Evans DH, Piermarini PM, Choe KP (2005). The multifunctionalfish gill: dominant site of gas exchange, osmoregulation,acid-base regulation, and excretion of nitrogenous waste.Physiol Rev 85: 97–177.
Fangue NA, Hofmeister M, Schulte PM (2006). Intraspecificvariation in thermal tolerance and heat shock protein geneexpression in common killifish, Fundulus heteroclitus. J ExpBiol 209: 2859–2872.
Hammer Ø, Harper DAT, Ryan PD (2001). PAST: paleontolo-gical statistics software package for education and dataanalysis. Palaeontologia Electronica 4: 9.
Hemmer-Hansen J, Nielsen EE, Frydenberg J, Loeschcke V(2007a). Adaptive divergence in a high gene flow environ-ment: Hsc70 variation in the European flounder (Platicthysflesus L.). Heredity 99: 592–600.
Hemmer-Hansen J, Nielsen EE, Gr�nkjær P, Loeschcke V(2007b). Evolutionary factors shaping the genetic populationstructure of marine fishes; lessons from the Europeanflounder (Platichthys flesus L.). Mol Ecol 16: 3104–3118.
Jakobsen F (1995). The major inflow to the Baltic Sea duringJanuary 1993. J Mar Syst 6: 227–240.
Jensen FB, Lecklin T, Busk M, Bury NR, Wilson RW, Wood CMet al. (2002). Physiological impact of salinity increase atorganism and red blood cell levels in the European flounder(Platichthys flesus). J Exp Mar Biol Ecol 274: 159–174.
Johannesson K, Andre C (2006). Life on the margin: geneticisolation and diversity loss in a peripheral marine ecosystem,the Baltic Sea. Mol Ecol 15: 2013–2029.
Salinity-dependent gene expression in European flounderPF Larsen et al
258
Heredity

Jorgensen HBH, Hansen MM, Bekkevold D, Ruzzante DE,Loeschcke V (2005). Marine landscapes and populationgenetic structure of herring (Clupea harengus L.) in the BalticSea. Mol Ecol 14: 3219–3234.
Kawecki TJ, Ebert D (2004). Conceptual issues in localadaptation. Ecol Lett 7: 1225–1241.
Knutsen H, Jorde PE, Andre C, Stenseth NC (2003). Fine-scaledgeographical population structuring in a highly mobilemarine species: the Atlantic cod. Mol Ecol 12: 385–394.
Larsen PF, Nielsen EE, Williams TD, Hemmer-Hansen J,Chipman JK, Kruh�ffer M et al. (2007). Adaptive differencesin gene expression in European flounder (Platichthys flesus).Mol Ecol 16: 4674–4683.
Livak KJ, Schmittgen TD (2001). Analysis of relative geneexpression data using real-time quantitative PCR and the2-[Delta][Delta]CT method. Methods 25: 402–408.
Mitchell-Olds T, Schmitt J (2006). Genetic mechanisms andevolutionary significance of natural variation in Arabidopsis.Nature 441: 947–952.
Nielsen EE, Hansen MM, Ruzzante DE, Meldrup D, Gronkjaer P(2003). Evidence of a hybrid-zone in Atlantic cod (Gadusmorhua) in the Baltic and the Danish Belt Sea revealed byindividual admixture analysis. Mol Ecol 12: 1497–1508.
Nielsen EE, Nielsen PH, Meldrup D, Hansen MM (2004).Genetic population structure of turbot (Scophthalmus maximusL.) supports the presence of multiple hybrid zones formarine fishes in the transition zone between the Baltic Seaand the North Sea. Mol Ecol 13: 585–595.
Nissling A, Westin L (1997). Salinity requirements for successfulspawning of Baltic and Belt Sea cod and the potential for codstock interactions in the Baltic Sea. Mar Ecol Prog Ser 152:261–271.
Nissling A, Westin L, Hjerne O (2002). Reproductive success inrelation to salinity for three flatfish species, dab (Limandalimanda), plaice (Pleuronectes platessa), and flounder (Pleur-onectes flesus), in the brackish water Baltic Sea. ICES J Mar Sci59: 93–108.
Oleksiak MF, Churchill GA, Crawford DL (2002). Variation ingene expression within and among natural populations. NatGenet 32: 261–266.
Picard DJ, Schulte PM (2004). Variation in gene expression inresponse to stress in two populations of Fundulus heteroclitus.Comp Biochem Physiol A Mol Integr Physiol 137: 205–216.
Pogson GH, Fevolden SE (2003). Natural selection and thegenetic differentiation of coastal and Arctic populations of
the Atlantic cod in northern Norway: a test involvingnucleotide sequence variation at the pantophysin (PanI)locus. Mol Ecol 12: 63–74.
Rodhe J, Winsor P (2002). On the influence of the freshwatersupply on the Baltic Sea mean salinity. Tellus Ser A DynMeteorol Oceanogr 54: 175–186.
Rozen S, Skaletsky HJ (2000). Primer3 on the WWW for generalusers and for biologist programmers. In: Krawetz S,Misener S (eds). Bioinformatics Methods and Protocols: Methodsin Biology. Humana Press, Totowa, NJ, pp 365–386.
Schadt EE, Monks SA, Drake TA, Lusis AJ, Che N, Colinayo Vet al. (2003). Genetics of gene expression surveyed in maize,mouse and man. Nature 422: 297–302.
Schranzhofer M, Schifrer M, Cabrera JA, Kopp S, Chiba P, BeugH et al. (2006). Remodeling the regulation of iron metabolismduring erythroid differentiation to ensure efficient hemebiosynthesis. Blood 107: 4159–4167.
Scott GR, Schulte PM (2005). Intraspecific variation in geneexpression after seawater transfer in gills of the euryhalinekillifish Fundulus heteroclitus. Comp Biochem Physiol A MolIntegr Physiol 141: 176–182.
Sorensen JG, Kristensen TN, Loeschcke V (2003). The evolu-tionary and ecological role of heat shock proteins. Ecol Lett 6:1025–1037.
Stahlberg A, Hakansson J, Xian XJ, Semb H, Kubista M (2004).Properties of the reverse transcription reaction in mRNAquantification. Clin Chem 50: 509–515.
Waples RS (1998). Separating the wheat from the chaff: patternsof genetic differentiation in high gene flow species. J Heredity89: 438–450.
Ward RD, Woodwark M, Skibinski DOF (1994). A comparisonof genetic diversity levels in marine, fresh-water, andanadromous fishes. J Fish Biol 44: 213–232.
Weber RE, Fago A (2004). Functional adaptation and itsmolecular basis in vertebrate hemoglobins, neuroglobinsand cytoglobins. Respir Physiol Neurobiol 144: 141–159.
Whitehead A, Crawford DL (2006). Neutral and adaptivevariation in gene expression. Proc Natl Acad Sci USA 103:5425–5430.
Wong MKS, Ge W, Woo NYS (2007). Positive feedback ofhepatic angiotensinogen expression in silver sea bream(Sparus sarba). Mol Cell Endocrinol 263: 103–111.
Wong MKS, Takei Y, Woo NYS (2006). Differential status of therenin-angiotensin system of silver sea bream (Sparus sarba) indifferent salinities. Gen Comp Endocrinol 149: 81–89.
Salinity-dependent gene expression in European flounderPF Larsen et al
259
Heredity
Copyright © 2022 FDOKUMEN




















