
Inhibition of IGF-I–induced Erk 1 and 2 activation and mitogenesis in mesangial cells by bradykinin

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Molecular cloning of IGF-1 and IGF-1 receptor and their expression pattern in theChilean flounder (Paralichthys adspersus)
Sebastian Escobar a,1, Eduardo N. Fuentes a, Erika Poblete a, Juan A. Valdés a, Diego Safian a, Ariel E. Reyes b,d,Marco Álvarez c, Alfredo Molina a,d,⁎a Universidad Andres Bello, Facultad de Ciencias Biológicas, Laboratorio de Biotecnología Molecular, Av. República 217, Santiago, Chileb Universidad Andres Bello, Facultad de Ciencias Biológicas, Laboratorio de Biología del Desarrollo, Av. República 217, Santiago, Chilec Universidad Andres Bello, Facultad de Ciencias Biológicas, Laboratorio de Biología Celular y Molecular, Av. República 217, Santiago, Chiled Millennium Institute for Fundamental and Applied Biology, Santiago, Chile
a b s t r a c ta r t i c l e i n f o
Article history:Received 1 July 2010Received in revised form 22 March 2011Accepted 23 March 2011Available online 5 April 2011
Keywords:IGF-1IGF-1RNotochordSomitesChilean flounder
Insulin-like growth factor-1 and insulin-like growth factor-1 receptor (IGF-1 and IGF-1R) play main roles invertebrate growth and development. In fish, besides contributing to somatic growth, both molecules exhibitpleiotropic functions. We isolated complete cDNAs sequences encoding for both IGF-1 and IGF-1R in theChilean flounder by using RT-PCR and rapid amplification of cDNAs ends (RACE) techniques. We analyzedgene expression in pre-metamorphic larvae and different organs of juvenile fish through whole mount in situhybridization and RT-PCR, respectively. In addition, we studied the presence of calcified skeletal structures inpre-metamorphic larvae through the fluorescent chromophore calcein. The IGF-1 cDNA sequence displays anopen reading frame of 558 nucleotides, encoding a 185 amino acid preproIGF-1. Moreover, IGF-1R contains anopen reading frame spanning 4239 nucleotides, rendering a 702 amino acid subunit alpha and a 676 aminoacid subunit beta. The deduced mature IGF-1 and IGF-1R exhibited high sequence identities with theircorresponding orthologs in fishes, especially those domains involved in biological activity. RT-PCR showedexpression of IGF-1 and IGF-1R transcripts in all studied tissues, consistent with their pleiotropic functions.Furthermore, we observed IGF-1 expression in notochord and IGF-1R expression in notochord, somites andhead in larvae of 8 and 9 days post fertilization. Complementarily, we detected in larvae of 8 days postfertilization the presence of calcified skeletal structures in notochord and head. Interestingly, bothmRNAs andcalcified structures were found in territories such as notochord, an embryonic midline structure essential forthe pattern of surrounding tissues as nervous system and mesoderm. Our results suggest that IGF-1 and itsreceptor play an important role in the development of the nervous system, muscle and bone-relatedstructures during larval stages.
© 2011 Elsevier Inc. All rights reserved.
1. Introduction
The insulin-like growth factor signaling system plays an importantrole in promoting growth and development in vertebrates (Moriyamaet al., 2000). This pathway involves the coordinated function of atleast two ligands, two cell surface receptors and six high affinitybinding proteins (Duan et al., 2010). The biological effects of the IGFsystem are mediated mainly by the interaction of IGF-1 ligand withIGF-1 receptor (IGF-1R) modulated through IGF binding proteins(IGFBPs) (Duan et al., 2010).
IGF-1 is synthesized in the liver which is the principal source ofendocrine IGF-1. The primary stimulus for synthesis of liver IGF-1 andits release into the circulation is growth hormone (GH). In additionIGF-1 is expressed not only in liver but also in several other organswhere it most likely acts in an autocrine/paracrine manner. IGF-1 isproduced as a pre-pro-hormone, which undergoes at least twoprocessing events: cleavage of the signal peptide and the C-terminalpeptide (Etherton, 2004; LeRoith et al., 2001). The mature IGF-1 is asingle chain polypeptide composed of 70 amino acids, which containsdomains A and B separated by a C domain, and a D carboxy domain(Humbel, 1990). The IGF-1 receptor is synthesized as a single chainpre-pro-receptor, with a 30 residue signal peptide that is co-translationally cleaved and a 1337 amino acid pro-receptor that isprocessed at a tetrabasic cleavage site to generate alpha and betasubunits (LeRoith et al., 1995). Themature IGF-1R is comprised by twoalpha subunits and two beta subunits linked by disulphide bonds,forming α2β2 heterotetramers. The alpha subunits contain an
Comparative Biochemistry and Physiology, Part B 159 (2011) 140–147
⁎ Corresponding author at: Universidad Andres Bello, Facultad de Ciencias Biológicas,Laboratorio de Biotecnología Molecular, Av. República 217, Santiago, Chile. Tel.: +562661 8319; fax: +56 2661 8415.
E-mail address: [email protected] (A. Molina).1 Present address: Consejo Superior de Investigaciones Científicas (CSIC), Instituto de
Acuicultura de Torre de la Sal (IATS), Ribera de Cabanes 12595 Castellón, Spain.
1096-4959/$ – see front matter © 2011 Elsevier Inc. All rights reserved.doi:10.1016/j.cbpb.2011.03.003
Contents lists available at ScienceDirect
Comparative Biochemistry and Physiology, Part B
j ourna l homepage: www.e lsev ie r.com/ locate /cbpb

Author's personal copy
extracellular ligand-binding domain and the beta subunits arecomposed of a single transmembrane domain and a highly conservedintracellular TK domain (LeRoith et al., 1995).
At a cellular level the IGF-1 acts in an autocrine/paracrine mannerto control physiological processes such as protein synthesis, cellproliferation, differentiation, and apoptosis (Jones and Clemmons,1995). In vertebrates, the insulin-like growth factor system has beenshown to be unique among growth factors, playing an important rolein the early patterning and muscle development. Over the pastdecade, IGF-1 cDNAs' partial and complete sequences have beenisolated from several fishes, including Teleost (Wood et al., 2005),Elasmobranchs (Duguay et al., 1995) and Agnathans (Nagamatsu etal., 1991) revealing an evolutionary conserved sequence (Wood et al.,2005). In zebrafish four distinct genes encoding IGF peptides (IGF-1a,IGF-1b, IGF-2a, and IGF-2b) have been cloned and identified revealingan important role in the regulation of midline and notochorddevelopment during embryogenesis (Zou et al., 2009).
Additionally full length and partial cDNA sequences encodingIGF-1R have been reported in several teleost such us turbot (Elies et al.,1999), zebrafish (Ayaso et al., 2002; Maures et al., 2002), Japaneseflounder (Nakao et al., 2002), and seabream(Perrot et al., 1999) amongothers. To date the duplication of two functional IGF-1R genes (IGF-1Raand IGF-1Rb) in Japanese flounder (Nakao et al., 2002) and zebrafish(Maures et al., 2002) has been reported, with a differential expressionpattern suggesting different roles in the regulation of growth anddevelopment of vertebrates.
Chilean flounder (Paralichthys adspersus) is a marine fish widelydistributed throughout the Chilean coast, which is raising a higheconomic value. However, slow growth rate has been recognized as amajor problem inmarinefish, increasing thefinal production cost. Thus,the viability to farm these species requires new knowledge in order todevelop new strategies to improve fish growth (Delgado et al., 2008). Inthis regard, considering the relevant function of the IGF-1 signalingpathways in promoting growth and skeletal muscle development infish, we describe here as a first approximation, the isolation andcharacterization of the full length IGF-1R and IGF-1 cDNAs, whichinclude the codifying regions and the 5′- and 3′-untranslated regions ofboth transcripts. Additionally, we studied the mRNA expression in pre-metamorphic larvae and in different tissues of juvenile fish and thepresence of calcified skeletal structures in larvae.
2. Materials and methods
2.1. Fish
Chilean flounder (P. adspersus) were collected from the Centro deInvestigaciones Marinas de Quintay (CIMARQ) (V Region, Valparaíso,Chile). The fish were maintained under natural temperature andphotoperiod conditions corresponding to geographic localization ofCIMARQ (33°13′S 71°38′W) and were fed twice daily with turbotpellet (Biomar, Chile). Juvenile fish (average mass 200±10 g) weresacrificed through an overdose of anesthetic (3-aminobenzoic acidethyl ester) (300 mg/L). The heart, kidney, gills, intestine, gonads,spleen, liver, stomach, white muscle and red muscle tissues werecollected, directly frozen in liquid nitrogen and stored at −80 °C.
Larvae were obtained after in vitro fertilization of eggs by malebroodstock sperm. Embryos were maintained under intensive-cultureconditions in conic larval culture tanks at 19 °C±2 °C.
Larvae at pre-metamorphic stages were collected, fixed in 4%paraformaldehyde in PBS for 2 h at 4 °C, dehydrated in methanol andstored at −20 °C.
2.2. RT-PCR and cDNA cloning
Total RNAwas isolated from liver using Trizol reagent following themanufacturer protocols (Invitrogen, Carlsbad, CA, USA). A total of 5 μg
of RNApreviously treatedwithDNase I (1 U/μL)was used for RNAfirst-strand cDNA using M-MLV reverse transcriptase (Invitrogen). PCRprimers for cloning IGF-1 receptor and IGF-1 cDNA (Table A1,supplemental material) were designed from a consensus analysis ofconserved coding regions of known IGF-1 receptor and IGF-1 ligandsequences from Japanese flounder, turbot and zebrafish. The PCRreaction, containing the cDNA template, 10 μL of 10X PCR buffer,200 μM of each dNTP, 1 μg of the forward and reverse primers, and2.5 U of Pfu Taq DNA polymerase (Promega, Madison, WI, USA), wascarried out in a final volume of 50 μL. The IGF-1R and IGF-1 PCRproducts were cloned into the pGEM-T easy vector (Promega) and theclones were completely sequenced and assembled in only onesequence. The IGF-1R and IGF-1 full-length 5′-terminal region,including the transcription start site, was completed using the FirstChoice RLM-RACE kit (Ambion, Austin, TX, USA) according to themanufacturer's instructions. Briefly, a RT-PCR with an adapter primerand the IGF-1R gene specific primers (5IGF-1ROP 5′-GACAGACAG-CATCAGACCCCAAAACA-3′, 5IGF-1RIP 5′-TGCCAGTCACAGGATACTTG-3 ′) or the IGF-1 gene specific primers (5IGF-1OP 5 ′-AAAAGCCTCTCTCTCCACACAC-3′, 5IGF-1IP 5′-TCTCTCCACACACA-CAAACTGCAG-3′) using a CIP/TAP mRNA as a template in a nestedreaction. The IGF-1R 3′-region was obtained using the gene-specificprimers (3IGF-1ROP 5′-ACCCAGGTCCTACCCCCACTCAAA-3′, 3IGF-1RIP5′-TTCTCCCTTCGGGGAAAT GAGTTT-3′). The IGF-1 3′-region wasobtained using the gene-specific primers (3IGF-1OP 5′-ACCTGGA-GATGTACTGTGCAC-3′, 3IGF-1IP 5′-CAAGACTAGCAAGGCAGCTC-3′).
2.3. Sequence analysis
Amino acid sequences translated from the cDNA sequence werecompared with sequences in the GenBank public database, by usingthe NCBI-BLAST application (http://www.ncbi.nlm.nih.gov/blast).Multiple amino acid sequences' alignment for the IGF-1 receptorand IGF-1 were performed using clustalW (Thompson et al., 1994).
2.4. Tissue expression and distribution of IGF-1 receptor and IGF-1 bysemi quantitative RT-PCR analysis
Total RNA was extracted from juvenile Chilean flounder(P. adspersus) using the RNeasy Mini Kit (Qiagen, Austin, TX, USA)from several tissues (white muscle, red muscle, liver, spleen, kidney,intestine, stomach, heart, gill and gonad) following manufacturerrecommendations. RNA was quantified using NanoDrop technologywith the Epoch Multi-Volume Spectrophotometer System (BioTek,Winooski, VT, USA). Assessment of RNA quality was performed byelectrophoresis on a 1.2% formaldehyde agarose gel containingethidium bromide. Only RNAs with an A260/280 ratio between 1.9and 2.1 were used for cDNA synthesis. 800 ng of RNA was reversetranscribed into cDNA for 30 min at 42 °C using a Quantitect reversetranscription kit (Qiagen) as per manufacturer's recommendations.
PCR was performed using 7.5 μL of Go taq green master mix(Promega), 6 μL of cDNA (50-fold dilution), and 500 nM of each primer.Gene-specificprimerswereused to amplify a fragmentof 400 bp for IGF-1(F: CTTCAAGAGTGCGATGTGCTGT; R: GGCTGCTGTGCTGTCCTACG),645 bp for IGF-1R (F: GACCTTGCTCTCAAGTGCAC; R: CTAACGAGACAAC-GAGTCACA) and 679 bp for β-actin (F: GTCCCTGTATGCCTCTGGTCG; R:CAGGAGTACGATGAGTCTGGA), which was used as constitutive geneexpression control. Thermal cycling conditions for IGF-1, IGF-1R andβ-actin were the following: initial activation of 10 min at 95 °C, followedby 35 (IGF-1) or 33 (IGF-1R), or 23 (β-actin) cycles of 30 s at 95 °C; 30 s at60 (IGF-1) or 52 °C (IGF-1R) or 57 °C (β-actin); 30 s (IGF-1) or 45 s (IGF-1R and β-actin) at 72 °C, and a final extension of 10 min at 72 °C. Semiquantitative expression analyses were carried out through densitometricanalyses of the bands using the Image J program (National Institutes ofHealth (NIH), USA), performing a ratio of the gene of interest (IGF-1 or
141S. Escobar et al. / Comparative Biochemistry and Physiology, Part B 159 (2011) 140–147

Author's personal copy
IGF-1R) with the gene of constitutive expression (β-actin). Results arepresented as arbitrary units (A.U) and expressed as means±SE (n=3).
Statistical analyses were carried out with the software STATISTICA 7(Tulsa, OK, USA) using the general linear model (GLM) with repeatedmeasurements ANOVA followed by Tukey's analyses as post-test. Aprobability level of Pb0.05 was used to indicate significance.
2.5. Whole-mount in situ hybridization
A1604 bp fragment corresponding to the IGF-1Rcoding sequencewasamplified by PCR using gene-specific primers (5′-GCCTTTCCAGAACAT-CACAGAG-3′, 5′-TTGAACTCCTTCATGACGGAGG-3′) and the same cDNAdescribed before as a template. This fragments were cloned into pGEM-TEasy Vector System (Promega) originating the pL3F4R1604, which waslinearized with NdeI or SpHI restriction enzymes to synthesized sense(control) and antisense riboprobes DIG-UTP-labeled (Roche Diagnostics,Mannheim, Germany) using SP6 and T7 RNA polymerases (Promega)respectively. A 454 bp fragment corresponding to the IGF-1 codingsequence was amplified by PCR using gene-specific primers (5′-CCTCTCCACTACTGCTGTGTGTC-3′, 5′-ATGTCTGTGTGGCGTTGTGCAC-3′)and the same cDNA described before as a template. This fragmentswere cloned into pGEM-T Easy Vector System (Promega) originating thepLeigfI454,whichwas linearizedwithNdeI orNcoI restriction enzymes tosynthesized sense (control) and antisense riboprobes DIG-UTP-labeled(Roche Diagnostics, Mannheim, Germany) using SP6 and T7 RNApolymerases (Promega) respectively. The riboprobes were purifiedusing mini Quick Spin Columns (Roche Diagnostics) to eliminateunincorporated labeled nucleotides. Whole mount in situ hybridizationwas performed according to Fuentes et al. (2008). Briefly, after bleachingtreatment, embryos and larvaewere pre-hybridized overnight at 60 °C inhybridization buffer and then incubated overnight at 65 °C in hybridiza-tion buffer including 50 ng of sense or antisense IGF-1R and IGF-1riboprobes. After hybridization, embryos and larvae were washed in asolutionwith decreasing formamide concentration in 2XSSC, followedbytwowash-stepswith SSC 0.2X for 30 min at 65 °C. Larvaewere incubatedfor 4 h in a blocking buffer at room temperature. For immunodetection,samples were incubated overnight at 4 °C with anti-digoxigenin-APantibody (Roche Diagnostics). After washes with PBT to eliminate non-bounded antibodies and three additional washes with AP-buffer, stainswere performed with NBT/BCIP (75 mg/mL and 50 mg/mL, respectively)(Promega) for 6 h at 37 °C. The experiment was performed four timesusing n=15 individuals from each developmental stages. After in situhybridization, samples were observed in a Leica MZ12.5 stereomicro-scope and photographed with a Leica DF300 camera.
2.6. Larvae calcein staining
Calcein immersion was carried out as described by Du et al. (2001).Briefly, Chilean flounder larvae were netted and immersed in calcein(Sigma-Aldrich Chemical, St. Louis, MO, USA) solution (0.2%) in Petridishes for 10 min. After the immersion, larvae were rinsed a number oftimes in fresh water, and then allowed to set for 10 min to allow theexcess, unbound calcein to diffuse. The larvae were mounted on glassslides with methyl-cellulose (3%). Observation was carried in LeicaMZ12.5 stereomicroscopewithafluorescencefilter set andphotographedwith a Leica DF300 camera. Composite images of the larvae wereproduced with Image J program (National Institutes of Health (NIH),USA).
3. Results
3.1. Cloning and characterization of the IGF-1R and IGF-1 cDNA
The complete Chilean flounder IGF-1R and IGF-1 cDNA sequences(cfIGF-1R and cfIGF-1) were obtained using RT-PCR coupled to RACEmethod (GenBank number FJ438475 and EU017533, respectively).
The length of the complete IGF-1R cDNA sequence was 5033 bp,which includes a 622 bp 5′-untranslated region (UTR), an openreading frame (ORF) of 4239 bp and a 172 bp 3′-UTR. The ORFencodes a putative protein of 1412 amino acid residues (Fig. A1,supplemental material). The sequence analysis reveals that theChilean flounder cDNA IGF-1 receptor is organized into severalmajor domains including a signal peptide sequence of 30 aminoacids, an extracellular alpha subunit of 702 amino acids and anintracellular beta subunit of 676 amino acids (Fig. A1, supplementalmaterial).
The length of the complete IGF-1 cDNA sequence is 980 bp, whichincludes a 152 bp 5′-untranslated region (UTR), an open reading frame(ORF) of 558 bp and a 270 bp 3′-UTR, encoding a putative protein of185 amino acid residues (Fig. A2, supplemental material). Thecomparative IGF-1 sequence analysis with other species reveals thatthe ligand deduced protein is subdivided into six structural domains,including a signal peptide sequence of 44 amino acids, the B domain of29 amino acids, the C domain of 10 amino acids, the A domain of 21amino acids, the D domain of 8 amino acids and the E domain of 73amino acids (Fig. A2, supplemental material).
Amino acid sequence alignment of cfIGF-1R (FJ438475.1) withdifferent vertebrate orthologs, including mammalian (human,X04434.1 andrat, AF056187.1), birds (chicken,AJ223164.1), amphibians(Xenopus, AF055980.1) and fish (zebrafish, AF400275.1 and Japaneseflounder, AB065098.1) was performed (Fig. A1, supplemental material).The cfIGF-1R sequence was found to have 61%, 61%, 63%, and 62% ofidentity with IGF-1R of human, rat, chicken and Xenopus, respectively.Higher degrees of identity were foundwith other fish IGF-1R sequences,including a 74% with zebrafish and as much as 97% with another fishbelonging to the Paralichthys genus, the Japanese flounder (Fig. 1A).
A potential proteolytic cleavage sequence R-X-R-R was conservedin all the species compared. The cysteine-rich domain, into the alphasubunit of the cfIGF-1R, contains 24 cysteine residues, which werealso observed in all vertebrate IGF-1Rs (Fig. A1, supplementalmaterial). In the beta subunit, an IRS-1 and IRS-2 binding site (NPEYand GVLY), a potential ATP binding site (G-X-G-X-X-G-21-X-K), anautophosphorylation motif (YETDYY) and seven tyrosine residues inthe tyrosine kinase domain, were highly conserved in all the studiedspecies (Fig. A1, supplemental material). A lesser conserved regionwas found in the carboxyl-terminal, where large insertions wereobserved in the teleosts IGF-1R compared with those from highervertebrates (Fig. A1, supplemental material).
Amino acid sequence alignment of the cfIGF-1 (EU017533.1) withthose from several other species, including human (M27544.1), rat(NM_001082479.1), chicken(M32791.1),Xenopus (M29857.1), zebrafish(BC114262.1) and Japanese flounder (AJ010602.1) was performed(Fig. A2, supplemental material). The cfIGF-1 sequence was found to be60%, 65%, 69%, 66%, 73%, and 97% homologous to the IGF-1 of human, rat,chicken, Xenopus, zebrafish and Japanese flounder, respectively (Fig. 1B).
The comparison between the known IGF-1 sequences, revealshigher conserved regions at both B and A domainswhile the C, D and Edomains differ significantly (Fig. 1B). There were six conservedcysteine residues in themature peptide, two in B domain and four in Adomain, observed in all vertebrates. Additionally amino acid residuesinvolved in IGF binding with IGFBP and IGF-1 receptor in the A, B andC domains are highly conserved.
3.2. Tissue distribution of IGF-1R and IGF-1 mRNA
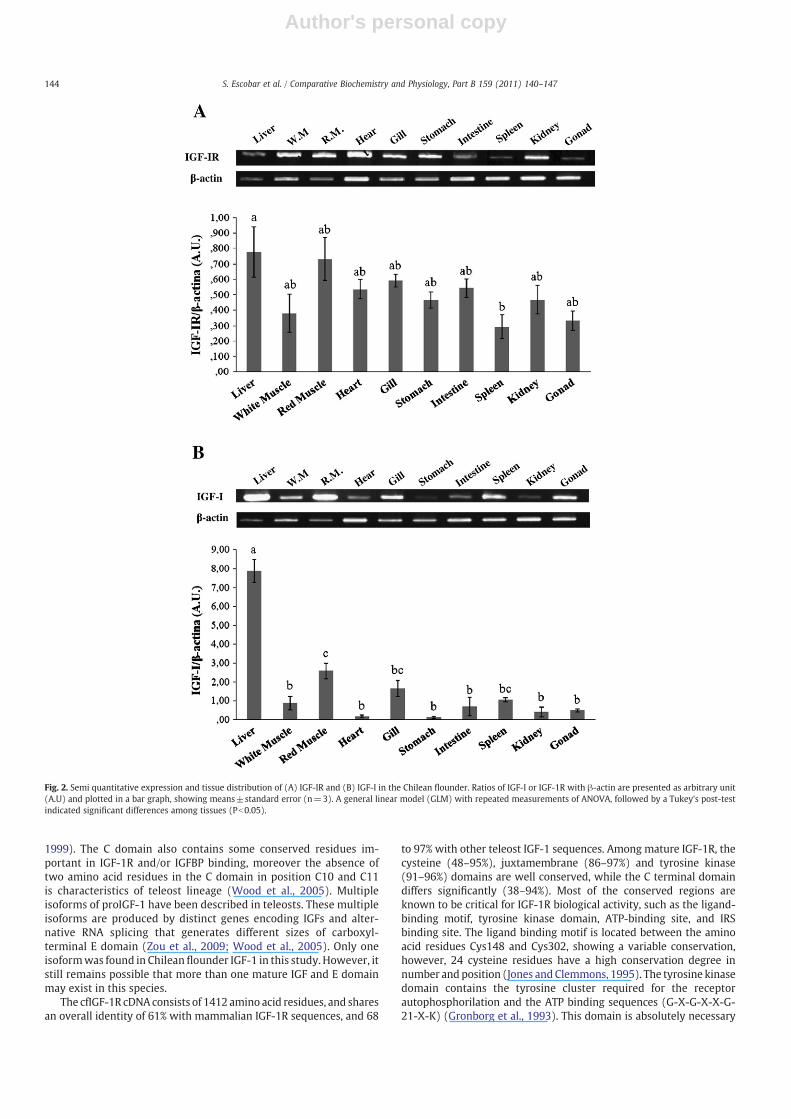
IGF-1R and IGF-1 mRNA semi-quantitative RT-PCR experimentswere performed to study the expression of mRNA in different tissuesof juvenile fish using β-actin as a constitutive expression control(Fig. 2). The IGF-1R and IGF-1 transcripts were detected in allinvestigated tissues. High levels of IGF-1R mRNA expression wereobserved in the liver, white muscle, red muscle, heart, gill, stomach,intestine, kidney and gonad (Fig. 2A). The IGF-1R mRNA expression
142 S. Escobar et al. / Comparative Biochemistry and Physiology, Part B 159 (2011) 140–147

Author's personal copy
detected in spleen was at low levels (Fig. 2A). Moreover, high IGF-1mRNA expression was observed in liver (Fig. 2B). Lower IGF-1 mRNAexpression levels were detected in white muscle, red muscle, heart,gill, stomach, intestine, spleen, kidney and gonad (Fig. 2B).
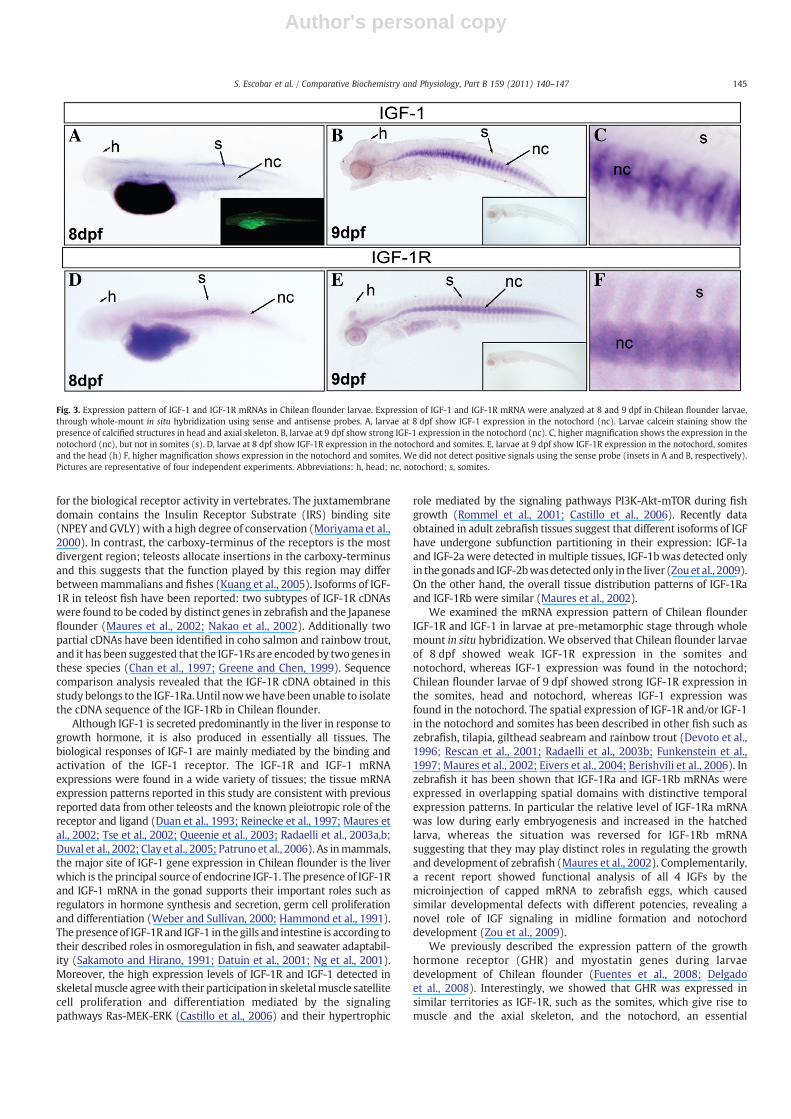
In addition, we studied the expression pattern of IGF-1R and IGF-1mRNA in embryos using whole mount in situ hybridization in Chileanflounder larvae from 8.0 and 9.0 days post-fertilization (dpf) (Fig. 3).The cfIGF-1R mRNA was detected at 8 and 9 dpf, larvae exhibited IGF-1R mRNA expression in the somites, notochord and cartilaginoustissues in the head. Additionally, cfIGF-1 mRNA was detected only inthe notochord. Sense probe was included as a negative control in all insitu hybridization experiments. No signal was detected, showing thatRNA hybridization was specific. In order to analyze skeletal calcifica-tion in Chilean flounder embryos, larvae of 8.0 dpf were immersed incalcein solution and observed under a fluorescence microscope.Fluorescent signal could be detected in larvae of 8 dpf in head regionand notochord (axial skeleton) (Fig. 3A).
4. Discussion
In this work we reported the complete cDNA sequence of the IGF-1and IGF-1 receptor from the flat fish Chilean flounder (P. adspersus),and we analyzed gene expression in pre-metamorphic larvae and
different organs of juvenile fish through whole mount in situhybridization and RT-PCR, respectively. These results complementprevious reported data of myostatin and the growth hormonereceptor (GHR) (Fuentes et al., 2008; Delgado et al., 2008), allimportant genes for growth and development in vertebrates (Daytonand White, 2008).
The deduced protein sequence of cfIGF-1 shares an overall identityof 60 to 65% with mammalian IGF-1 sequences, and 73 to 97% withother teleost IGF-1 sequences. As other vertebrates, cfIGF-1 cDNAsequence is predicted to encode a preprohormone, with a length of 185amino acid residues, which contain a signal peptide and B, C, A, D, and Edomains. The removing of putative signal peptide and E domain yieldsthe mature cfIGF-1 peptide of 68 amino acid residues. The cfIGF-1sequence showed six conserved cysteine residues CysB6, CysB18,CysA6, CysA7, CysA11, and CysA20 which are also located at the samepositions as mammal IGF-1 which is responsible for maintenance oftertiary structure (Hober et al., 1992). The analysis of themature cfIGF-1reveals that the B (83–100%) and A (86–95%) domains of the peptidesarewell conserved,while the C (100–42%), D (25–88%) and E (23–73%)domains differ significantly. The importance of this high sequenceidentity of B and A domains in different species can be attributed to thefunctional roles of these regions, which are involved in the binding ofIGF-1 with its receptor and IGFBP (Zhang et al., 1994; Magee et al.,
A
cenietsydomain
juxtamembranedomain
Tyrosine_kinasedomain
C_terminaldomain
Whole protein
Ch_flounder/j_flounder 95 97 96 94 97Ch_flounder/turbot 63 95 91 38 67Ch_flounder/carp 72 95 92 63 76Ch_flounder/zebrafishIGFR1a
69 92 92 66 74
Ch_flounder/zebrafishIGFR1b
58 92 92 50 68
Ch_flounder/xenopus 50 93 90 48 62Ch_flounder/chicken 52 90 91 47 63Ch_flounder/rat 48 86 91 42 61Ch_flounder/human 49 88 91 41 61
B
Bdomain
Cdomain
Adomain
Ddomain
Edomain
Whole protein
Ch_flounder/j_flounder 97 100 95 88 73 97 Ch_flounder/turbot 100 100 91 88 49 96 Ch_flounder/carp 97 58 95 25 27 74 Ch_flounder/zebrafish 83 67 95 38 27 73 Ch_flounder/xenopus 97 67 86 50 26 66 Ch_flounder/chicken 93 42 91 38 26 69 Ch_flounder/rat 93 42 86 38 25 65 Ch_flounder/human 86 42 86 38 23 60
α βCystein domain
5’UTR 3’UTR
Juxtamembrane TK C-terminal
B domain C domain A domain D domain E domain
5’UTR 3’UTR
Fig. 1. Comparison of the Chilean flounder IGF-1R and IGF-1 deduced domains with Japanese flounder, turbot, carp, zebrafish, Xenopus, chicken, rat and human IGF-1R and IGF-1.A) GenBank accession numbers: human (Homo sapiens) GenBank: X04434.1; rat (Rattus norvegicus) AF056187.1; chicken (Gallus gallus) AJ223164.1; African clawed frog (Xenopus laevis)AF055980.1; zebrafish (Danio rerio) AF400275.1 and BC163723.1; Carp (Cyprinus carpio) AY144591.1; turbot (Psetta maxima) AJ224993.1; Japanese flounder (Paralichthys olivaceus)AB065098.1, andChileanflounder (Paralichthys adspersus) FJ438475.1. The top linedrawing represents the cDNAstructureofhuman IGF-1R. The comparisonbetweenChileanflounderandother species was performed using 170, 43, 253 and 155 amino acids corresponding to the cysteine domain, juxtamembrane domain, tyrosine kinase domain and c-terminal domain,respectively. B)GenBank accessionnumbers: human (Homo sapiens)M27544.1; rat (Rattusnorvegicus)GenBank:NM_001082479.1; chicken (Gallus gallus)M32791.1;African clawed frog(Xenopus laevis) M29857.1; zebrafish (Danio rerio) BC114262.1; Carp (Cyprinus carpio) BAA11878.1; turbot (Psetta maxima) ACL14947.1; Japanese flounder (Paralichthys olivaceus)AJ010602.1; Chilean flounder (Paralichthys adspersus) EU017533.1. The top line drawing represents the cDNA structure of human IGF-1. The comparison between Chilean flounder andother species was performed using 29, 10, 21, 8 and 73 amino acids corresponding to B, C, A, D and E domains, respectively.
143S. Escobar et al. / Comparative Biochemistry and Physiology, Part B 159 (2011) 140–147
Author's personal copy
1999). The C domain also contains some conserved residues im-portant in IGF-1R and/or IGFBP binding, moreover the absence oftwo amino acid residues in the C domain in position C10 and C11is characteristics of teleost lineage (Wood et al., 2005). Multipleisoforms of proIGF-1 have been described in teleosts. These multipleisoforms are produced by distinct genes encoding IGFs and alter-native RNA splicing that generates different sizes of carboxyl-terminal E domain (Zou et al., 2009; Wood et al., 2005). Only oneisoformwas found in Chilean flounder IGF-1 in this study. However, itstill remains possible that more than one mature IGF and E domainmay exist in this species.
The cfIGF-1R cDNA consists of 1412 amino acid residues, and sharesan overall identity of 61% with mammalian IGF-1R sequences, and 68
to 97% with other teleost IGF-1 sequences. Among mature IGF-1R, thecysteine (48–95%), juxtamembrane (86–97%) and tyrosine kinase(91–96%) domains are well conserved, while the C terminal domaindiffers significantly (38–94%). Most of the conserved regions areknown to be critical for IGF-1R biological activity, such as the ligand-binding motif, tyrosine kinase domain, ATP-binding site, and IRSbinding site. The ligand binding motif is located between the aminoacid residues Cys148 and Cys302, showing a variable conservation,however, 24 cysteine residues have a high conservation degree innumber and position (Jones and Clemmons, 1995). The tyrosine kinasedomain contains the tyrosine cluster required for the receptorautophosphorilation and the ATP binding sequences (G-X-G-X-X-G-21-X-K) (Gronborg et al., 1993). This domain is absolutely necessary
Fig. 2. Semi quantitative expression and tissue distribution of (A) IGF-IR and (B) IGF-I in the Chilean flounder. Ratios of IGF-I or IGF-1R with β-actin are presented as arbitrary unit(A.U) and plotted in a bar graph, showing means±standard error (n=3). A general linear model (GLM) with repeated measurements of ANOVA, followed by a Tukey's post-testindicated significant differences among tissues (Pb0.05).
144 S. Escobar et al. / Comparative Biochemistry and Physiology, Part B 159 (2011) 140–147
Author's personal copy
for the biological receptor activity in vertebrates. The juxtamembranedomain contains the Insulin Receptor Substrate (IRS) binding site(NPEY and GVLY)with a high degree of conservation (Moriyama et al.,2000). In contrast, the carboxy-terminus of the receptors is the mostdivergent region; teleosts allocate insertions in the carboxy-terminusand this suggests that the function played by this region may differbetweenmammalians and fishes (Kuang et al., 2005). Isoforms of IGF-1R in teleost fish have been reported: two subtypes of IGF-1R cDNAswere found to be coded by distinct genes in zebrafish and the Japaneseflounder (Maures et al., 2002; Nakao et al., 2002). Additionally twopartial cDNAs have been identified in coho salmon and rainbow trout,and it has been suggested that the IGF-1Rs are encoded by two genes inthese species (Chan et al., 1997; Greene and Chen, 1999). Sequencecomparison analysis revealed that the IGF-1R cDNA obtained in thisstudy belongs to the IGF-1Ra. Until nowwehave been unable to isolatethe cDNA sequence of the IGF-1Rb in Chilean flounder.
Although IGF-1 is secreted predominantly in the liver in response togrowth hormone, it is also produced in essentially all tissues. Thebiological responses of IGF-1 are mainly mediated by the binding andactivation of the IGF-1 receptor. The IGF-1R and IGF-1 mRNAexpressions were found in a wide variety of tissues; the tissue mRNAexpression patterns reported in this study are consistent with previousreported data from other teleosts and the known pleiotropic role of thereceptor and ligand (Duan et al., 1993; Reinecke et al., 1997; Maures etal., 2002; Tse et al., 2002; Queenie et al., 2003; Radaelli et al., 2003a,b;Duval et al., 2002; Clay et al., 2005; Patruno et al., 2006). As inmammals,the major site of IGF-1 gene expression in Chilean flounder is the liverwhich is the principal source of endocrine IGF-1. The presence of IGF-1Rand IGF-1 mRNA in the gonad supports their important roles such asregulators in hormone synthesis and secretion, germ cell proliferationand differentiation (Weber and Sullivan, 2000; Hammond et al., 1991).Thepresenceof IGF-1Rand IGF-1 in thegills and intestine is according totheir described roles in osmoregulation in fish, and seawater adaptabil-ity (Sakamoto and Hirano, 1991; Datuin et al., 2001; Ng et al., 2001).Moreover, the high expression levels of IGF-1R and IGF-1 detected inskeletalmuscle agreewith their participation in skeletalmuscle satellitecell proliferation and differentiation mediated by the signalingpathways Ras-MEK-ERK (Castillo et al., 2006) and their hypertrophic
role mediated by the signaling pathways PI3K-Akt-mTOR during fishgrowth (Rommel et al., 2001; Castillo et al., 2006). Recently dataobtained in adult zebrafish tissues suggest that different isoforms of IGFhave undergone subfunction partitioning in their expression: IGF-1aand IGF-2a were detected in multiple tissues, IGF-1b was detected onlyin the gonadsand IGF-2bwasdetectedonly in the liver (Zouet al., 2009).On the other hand, the overall tissue distribution patterns of IGF-1Raand IGF-1Rb were similar (Maures et al., 2002).
We examined the mRNA expression pattern of Chilean flounderIGF-1R and IGF-1 in larvae at pre-metamorphic stage through wholemount in situ hybridization. We observed that Chilean flounder larvaeof 8 dpf showed weak IGF-1R expression in the somites andnotochord, whereas IGF-1 expression was found in the notochord;Chilean flounder larvae of 9 dpf showed strong IGF-1R expression inthe somites, head and notochord, whereas IGF-1 expression wasfound in the notochord. The spatial expression of IGF-1R and/or IGF-1in the notochord and somites has been described in other fish such aszebrafish, tilapia, gilthead seabream and rainbow trout (Devoto et al.,1996; Rescan et al., 2001; Radaelli et al., 2003b; Funkenstein et al.,1997; Maures et al., 2002; Eivers et al., 2004; Berishvili et al., 2006). Inzebrafish it has been shown that IGF-1Ra and IGF-1Rb mRNAs wereexpressed in overlapping spatial domains with distinctive temporalexpression patterns. In particular the relative level of IGF-1Ra mRNAwas low during early embryogenesis and increased in the hatchedlarva, whereas the situation was reversed for IGF-1Rb mRNAsuggesting that they may play distinct roles in regulating the growthand development of zebrafish (Maures et al., 2002). Complementarily,a recent report showed functional analysis of all 4 IGFs by themicroinjection of capped mRNA to zebrafish eggs, which causedsimilar developmental defects with different potencies, revealing anovel role of IGF signaling in midline formation and notochorddevelopment (Zou et al., 2009).
We previously described the expression pattern of the growthhormone receptor (GHR) and myostatin genes during larvaedevelopment of Chilean flounder (Fuentes et al., 2008; Delgadoet al., 2008). Interestingly, we showed that GHR was expressed insimilar territories as IGF-1R, such as the somites, which give rise tomuscle and the axial skeleton, and the notochord, an essential
Fig. 3. Expression pattern of IGF-1 and IGF-1R mRNAs in Chilean flounder larvae. Expression of IGF-1 and IGF-1R mRNA were analyzed at 8 and 9 dpf in Chilean flounder larvae,through whole-mount in situ hybridization using sense and antisense probes. A, larvae at 8 dpf show IGF-1 expression in the notochord (nc). Larvae calcein staining show thepresence of calcified structures in head and axial skeleton. B, larvae at 9 dpf show strong IGF-1 expression in the notochord (nc). C, higher magnification shows the expression in thenotochord (nc), but not in somites (s). D, larvae at 8 dpf show IGF-1R expression in the notochord and somites. E, larvae at 9 dpf show IGF-1R expression in the notochord, somitesand the head (h) F, higher magnification shows expression in the notochord and somites. We did not detect positive signals using the sense probe (insets in A and B, respectively).Pictures are representative of four independent experiments. Abbreviations: h, head; nc, notochord; s, somites.
145S. Escobar et al. / Comparative Biochemistry and Physiology, Part B 159 (2011) 140–147

Author's personal copy
structure for the proper formation of the nervous system andmesoderm, the last of which gives rise to the somites later in embryodevelopment (Richardson et al., 1998; Stemple et al., 1996).Furthermore, ligands just as myostatin and IGF-1 mRNAs, weremostly found in the notochord. Taken together, these observationssuggest there could be synchronization between positive and negativegrowth signals originated in the notochord that plays a crucial role inthe control of somite development (Delgado et al., 2008). In acomplementary way, we detected the presence of calcified structuresin notochord and head in 8.0 dpf Chilean flounder larvae by the use ofthe fluorescent chromophore calcein; this methodology has been usedto follow the development of the skeletal structures in zebrafishembryos detecting fluorescent labeling in head (Meckel's cartilage,the palatoquadrate, and the ethmoid plate) and axial skeleton (Du etal., 2001). Our results suggest that IGF-1 and its receptor play animportant role in the development of the nervous system, muscle andbone-related structures during larval stages.
In summary, the complete cDNA sequence of the IGF-1R and IGF-1was cloned from the Chilean flounder fish. The protein sequenceincludes all the structural domains and motifs responsible for theinteraction between ligand-receptor and IGF-1 mediated signaltransduction. Indeed, our results contribute to the knowledge of theIGF-1 system in the larvae and juvenile stages, both of which arecrucial periods for developing successful farming of the Chileanflounder.
Supplementarymaterials related to this article can be found onlineat doi:10.1016/j.cbpb.2011.03.003.
Acknowledgements
This work was supported by Grant Nos. 1090416 from FONDECYTand 15-03/28-04/13-06I fromUNAB Research Fund to A.M., FONDECYTNo. 1095128 to A.E.R., and the Millennium Institute for FundamentalandBasic Biology (MIFAB).We thank JuanManuel Estrada for technicalassistance and animal care in the Centro de Investigación Marina deQuintay (CIMARQ) and Ashley VanCott from The University of Nevada,Reno (USA) for improving and correcting the Englishof themanuscript.
References
Ayaso, E., Nolan, C.M., Byrnes, L., 2002. Zebrafish insulin-like growth factor-I receptor:molecular cloning and developmental expression. Mol. Cell. Endocrinol. 191,137–148.
Berishvili, G., Shved, N., Eppler, E., Clota, F., Baroiller, J.F., Reinecke,M., 2006. Organ-specificexpression of IGF-I during early development of bony fish as revealed in the tilapia,Oreochromis niloticus, by in situ hybridization and immunohistochemistry: indicationfor the particular importance of local IGF-I. Cell Tissue Res. 325, 287–301.
Castillo, J., Johnsen-Ammendrup, I., Codina, M., Navarro, I., Gutierrez, J., 2006. IGF-I andinsulin receptor signal transduction in trout muscle cells. Am. J. Physiol. Regul.Integr. Comp. Physiol. 290, R1683–R1690.
Chan, S.J., Plisetskaya, E.M., Urbinati, E., Jin, Y., Steiner, D.F., 1997. Expression of multipleinsulin and insulin-like growth factor receptor genes in salmon gill cartilage. Proc.Natl Acad. Sci. U.S.A. 94, 12446–12451.
Clay, L.A., Wang, S.Y., Wolters, W.R., Peterson, B.C., Waldbieser, G.C., 2005. Molecularcharacterization of the insulin-like growth factor-I (IGF-I) gene in channel catfish(Ictalurus punctatus). Biochim. Biophys. Acta 1731, 139–148.
Datuin, J.P., Ng, K.P., Hayes, T.B., Bern, H.A., 2001. Effects of glucocorticoids on cartilagegrowth and response to IGF-I in the tilapia (O. mossambicus). Gen. Comp.Endocrinol. 121, 289–294.
Dayton, W.R., White, M.E., 2008. Cellular and molecular regulation of muscle growthand development in meat animals. J. Anim. Sci. 86, E217–E225.
Delgado, I., Fuentes, E., Escobar, S., Navarro, C., Corbeaux, T., Reyes, A.E., Vera, M.I.,Alvarez, M., Molina, A., 2008. Temporal and spatial expression pattern of themyostatin gene during larval and juvenile stages of the Chilean flounder(Paralichthys adspersus). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 151,197–202.
Devoto, S.H., Melancon, E., Eisen, J.S., Westerfield, M., 1996. Identification of separateslow and fast muscle precursor cell in vivo, prior to somite formation. Development122, 3371–3380.
Du, S.J., Frenkel, V., Kindschi, G., Zohar, Y., 2001. Visualizing normal and defective bonedevelopment in zebrafish embryos using the fluorescent chromophore calcein.Dev. Biol. 238, 239–246.
Duan, C., Duguay, S., Plisetskaya, E., 1993. Insulin-like growth factor I (IGF-I) mRNAexpression in coho salmon, Oncorhynchus kisutch: tissue distribution and effects ofgrowth hormone/prolactin family peptides. Fish Physiol. Biochem. 11, 371–379.
Duan, C., Ren, H., Gao, S., 2010. Insulin-like growth factors (IGFs), IGF receptors, andIGF-binding proteins: roles in skeletal muscle growth and differentiation. Gen.Comp. Endocrinol. 167, 344–351.
Duguay, S.J., Chan, S.J., Mommsen, T.P., Steiner, D.F., 1995. Divergence of insulin-likegrowth factors I and II in the elasmobranch, Squalus acanthias. FEBS Lett. 371,69–72.
Duval, H., Rousseau, K., Elies, G., Le Bail, P., Dufour, S., Boeuf, G., Boujard, D., 2002.Cloning, characterization, and comparative activity of turbot. IGF-I and IGF-II. Gen.Comp. Endocrinol. 126, 269–278.
Eivers, E., McCarthy, K., Glynn, C., Nolan, C.M., Byrnes, L., 2004. Insulin-like growthfactor (IGF) signalling is required for early dorso-anterior development of thezebrafish embryo. Int. J. Dev. Biol. 48, 1131–1140.
Elies, G., Duval, H., Bonnec, G., Wolff, J., Boeuf, G., Boujard, D., 1999. Insulin and insulin-likegrowth factor-1 receptors in an evoluted fish, the turbot: cDNA cloning and mRNAexpression. Mol. Cell. Endocrinol. 158, 173–185.
Etherton, T.D., 2004. Somatotropic function: the somatomedin hypothesis revisited.J. Anim. Sci. 82, E239–E244.
Fuentes, E., Poblete, E., Reyes, A.E., Vera, M.I., Alvarez, M., Molina, A., 2008. Dynamicexpression pattern of the growth hormone receptor during early development ofthe Chilean flounder. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 150, 93–102.
Funkenstein, B., Almuly, R., Chan, S.J., 1997. Localization of IGF-I and IGF-I receptormRNA in Sparus aurata larvae. Gen. Comp. Endocrinol. 107, 291–303.
Greene, M.W., Chen, T.T., 1999. Characterization of teleost insulin receptor familymembers. II. Developmental expression of insulin-like growth factor type I receptormessenger RNAs in rainbow trout. Gen. Comp. Endocrinol. 115, 270–281.
Gronborg, M., Wulff, B.S., Rasmussen, J.S., Kjeldsen, T., Gammeltoft, S., 1993. Structure–function relationship of the insulin-like growth factor-I receptor tyrosine kinase.J. Biol. Chem. 258, 23435–23440.
Hammond, J.M., Mondschein, S.E., Samaras, S.E., Canning, S.F., 1991. The ovarianinsulin-like growth factors, a local amplification mechanism for steroidogenesisand hormone action. J. Steroid Biochem. Mol. Biol. 40, 411–416.
Hober, S., Forsberg, G., Palm, G., Hartmanis, M., Nilsson, B., 1992. Disulfide exchangefolding of insulin-like growth factor-I. Biochemistry 31, 1749–1756.
Humbel, R.E., 1990. Insulin-like growth factors I and II. Eur. J. Biochem. 190, 445–462.Jones, J.I., Clemmons, D.R., 1995. Insulin-like growth factors and their binding proteins:
biological actions. Endocr. Revs. 16, 3–34.Kuang, Y.M., Li, W.S., Lin, H.R., 2005. Molecular cloning andmRNA Profile of Insulin-like
Growth Factor Type 1 receptor in orange-spotted Grouper, Epinephelus coioides.Acta Biochim. Biophys. Sin. 37, 327–334.
LeRoith, D., Werner, H., Beitner-Johnson, D., Roberts, C.T., 1995. Molecular and cellularaspects of the insulin-like growth factor I receptor. Endocr. Revs. 16, 143–163.
LeRoith, D., Scavo, L., Butler, A., 2001. What is the role of circulating IGF-1? TrendsEndocrinol. Metab. 12, 48–52.
Magee, B.A., Shooter, G.K., Wallace, J.C., Francis, G.L., 1999. Insulin-like growth factor Iand its binding proteins: a study of the binding interface using B-domainanalogues. Biochemistry 38, 15863–15870.
Maures, T., Chan, S.J., Xu, B., Sun, H., Ding, J., Duan, C., 2002. Structural, biochemical, andexpression analysis of two distinct insulin-like growth factor I receptors and theirligands in zebrafish. Endocrinology 143, 1858–1871.
Moriyama, S., Ayson, F.G., Kawauchi, H., 2000. Growth regulation by insulin-like growthfactor-I in fish. Biosci. Biotechnol. Biochem. 64, 1553–1562.
Nagamatsu, S., Chan, S.J., Falkmer, S., Steiner, D.F., 1991. Evolution of the insulin genesuperfamily. Sequence of a preproinsulin-like growth factor cDNA from the Atlantichagfish. J. Biol. Chem. 266, 2397–2402.
Nakao, N., Tanaka, M., Higashimoto, Y., Nakashima, K., 2002. Molecular cloning,identification and characterization of four distinct receptor subtypes for insulin andIGF-I in Japanese flounder, Paralichthys olivaceus. J. Endocrinol. 173, 365–375.
Ng, K.P., Datuin, J.P., Bern, H.A., 2001. Effects of estrogens in vitro and in vivo oncartilage growth in the tilapia (O. mossambicus). Gen. Comp. Endocrinol. 121,295–304.
Patruno, M., Maccatrozzo, L., Funkenstein, B., Radaelli, G., 2006. Cloning and expressionof insulin-like growth factors I and II in the shi drum (Umbrina cirrosa). Comp.Biochem. Physiol. B. 144, 137–151.
Perrot, V., Moiseeva, E.B., Gozes, Y., Chan, S.J., Ingleton, P., Funkenstein, B., 1999.Ontogeny of the insulin-like growth factor system (IGF-I, IGF-II, and IGF-1R) ingilthead seabream (Sparus aurata): expression and cellular localization. Gen. Comp.Endocrinol. 116, 445–460.
Queenie, P., Vonga, K., Chana, M., Leungb, K., Cheng, C., 2003. Common carp insulin-likegrowth factor-I gene: complete nucleotide sequence and functional characterizationof the 5V-flanking region. Gene 322, 145–156.
Radaelli, G., Patruno, M., Maccatrozzo, L., Funkenstein, B., 2003a. Expression and cellularlocalization of insulin-like growth factor-II protein and mRNA in Sparus aurataduring development. J. Endocrinol. 178, 285–299.
Radaelli, G., Domeneghini, C., Arrighi, S., Bosi, G., Patruno, M., Funkenstein, B., 2003b.Localization of IGF-I, IGF-I receptor, and IGFBP-2 in developing Umbrina cirrosa(Pisces: Osteichthyes). Gen. Comp. Endocrinol. 130, 232–244.
Reinecke, M., Schmid, A., Ermatinger, R., Loffing-Cueni, D., 1997. Insulin-like growthfactor I in the teleost Oreochromis mossambicus, the tilapia: gene sequence, tissueexpression, and cellular localization. Endocrinology 138, 3613–3619.
Rescan, P.Y., Collet, B., Ralliere, C., Cauty, C., Delalande, J.M., Goldspink, G., Fauconneau,B., 2001. Red andwhite muscle development in the trout (O. mykiss) as shown by insitu hybridization of fast and slow myosin heavy chain transcripts. J. Exp. Biol. 204,2097–2101.
146 S. Escobar et al. / Comparative Biochemistry and Physiology, Part B 159 (2011) 140–147

Author's personal copy
Richardson, M.K., Allen, S.P., Wright, G.M., Raynaud, A., Hanken, J., 1998. Somite numberand vertebrate evolution. Development 125, 151–160.
Rommel, C., Bodine, S.C., Clarke, B.A., Rossman, R., Nunez, L., Stitt, T.N., Yancopoulos, G.D.,Glass, D.J., 2001.Mediation of IGF-1-induced skeletalmyotubehypertrophy by PI(3)K/Akt/mTOR and PI(3)K/Akt/GSK3 pathways. Nat. Cell Biol. 3, 1009–1013.
Sakamoto, T., Hirano, T., 1991. Growth hormone receptors in the liver andosmoregulatory organs of rainbow trout: characterization and dynamics duringadaptation to seawater. J. Endocrinol. 130, 425–433.
Stemple,D., Solnica-Krezel, L., Zwartkruis, F., Neuhauss, C.F., Schier, A.F.,Malicki, J., Stainier,D.Y.R., Abdelilah, S., Rangini, Z., Mountcastle-Shah, E., Driever, W., 1996. Mutationsaffecting development of the notochord in zebrafish. Development 123, 117–128.
Thompson, J.D., Higgins, D.G., Gibson, T.J., 1994. CLUSTAL W: improving the sensitivity ofprogressive 416 multiple sequence alignment through sequenceweighting, position-specific gap penalties andweight417matrix choice.Nucleic Acids Res. 22, 4673–4680.
Tse, M.C.L., Vong, Q.P., Cheng, C.H.K., Chan, K.M., 2002. PCR-cloning and gene expressionstudies in common carp (Cyprinus carpio) insulin-like growth factor-II. Biochim.Biophys. Acta 1575, 63–74.
Weber, G.M., Sullivan, C.V., 2000. Effects of insulin-like growth factor-I on in vitro finaloocyte maturation and ovarian steroidogenesis in striped bass, Morone saxatilis.Biol. Reprod. 63, 1049–1057.
Wood, A.W., Duan, C., Bern, H.A., 2005. Insulin-like growth factor signaling in fish. Int.Rev. Cytol. 243, 215–285.
Zhang, W., Gustafson, T.A., Rutter, W.J., Johnson, J.D., 1994. Positively charged sidechains in the insulin-like growth factor-1 C and D-regions determine receptorbinding specificity. J. Biol. Chem. 269, 10609–10613.
Zou, S., Kamei, H., Modi, Z., Duan, C., 2009. Zebrafish IGF genes: gene duplication,conservation and divergence, and novel roles in midline and notochorddevelopment. PLoS ONE 4, e7026.
147S. Escobar et al. / Comparative Biochemistry and Physiology, Part B 159 (2011) 140–147
Copyright © 2022 FDOKUMEN




















