
Scorpions et piqures de scorpions en Jordanie (Scorpions and scorpion stings in Jordan)

Ds
Pa
b
a
ARRAA
KSASPBC
1
Sqrc2srfGst
h0
Acta Tropica 134 (2014) 43–51
Contents lists available at ScienceDirect
Acta Tropica
jo ur nal home p age: www.elsev ier .com/ locate /ac ta t ropica
eep intraspecific divergences in the medically relevant fat-tailedcorpions (Androctonus, Scorpiones)
. Coelhoa, P. Sousaa,b, D.J. Harrisa,b, A. van der Meijdena,∗
CIBIO, InBio, Centro de Investigac ão em Biodiversidade e Recursos Genéticos, Universidade do Porto, Campus Agrário de Vairão, Vairão 4485-661, PortugalDepartamento de Biologia, Faculdade de Ciências, Rua do Campo Alegre, Porto FC4 4169-007, Portugal
r t i c l e i n f o
rticle history:eceived 9 October 2013eceived in revised form 28 January 2014ccepted 1 February 2014vailable online 11 February 2014
eywords:corpionismndroctonuscorpionshylogenyiogeographyryptic diversity
a b s t r a c t
The genus Androctonus, commonly known as fat-tailed scorpions, contains 22 species distributed fromTogo and Mauritania in the west, North Africa, through the Middle East and to as far east as India. With13 species, a substantial amount of this genus’ diversity occurs in North Africa, which is a major hotspotof scorpion sting incidents. Androctonus are among the most medically relevant animals in North Africa.Since venom composition within species is known to vary regionally, the improvement of therapeuticmanagement depends on a correct assessment of the existing regional specific and sub-specific variation.In this study, we assessed the phylogeographical patterns in six species of Androctonus scorpions fromNorth Africa using mitochondrial DNA markers. We sequenced COX1, 12S, 16S and ND1 genes from110 individuals. Despite lacking basal resolution in the tree, we found taxonomical and geographicallycoherent clades. We discovered deep intraspecific variation in the widespread Androctonus amoreuxiand Androctonus australis, which consisted of several well-supported clades. Genetic distances betweensome of these clades are as high as those found between species. North African A. australis have a deepsplit in Tunisia around the Chott el-Djerid salt-lake. A novel split between A. amoreuxi scorpions was
found in Morocco. We also found deep divergences in Androctonus mauritanicus, corresponding to areasattributed to invalidated subspecies. In addition we uncovered a clade of specimens from coastal southMorocco, which could not be ascribed to any know species using morphological characters. Based onthese findings we recommend a reassessment of venom potency and anti-venom efficacy between thesedeep intraspecific divergent clades.© 2014 Elsevier B.V. All rights reserved.
. Introduction
Worldwide, 1.2 million people are stung by scorpions every year.corpionism, defined as the severe to lethal incident as a conse-uence of a scorpion sting (Lourenc o and Cuellar, 1995) may beesponsible for 3250 global annual mortalities which are mostlyoncentrated in a few high-risk areas (Chippaux and Goyffon,008). North Africa in particular is considered a high-risk area forcorpionism (Chippaux and Goyffon, 2008), with the genera Leiu-us Ehrenberg, 1828 and Androctonus Ehrenberg, 1828 being the
oremost cause of serious envenomation in this area (Goyffon anduette, 2005; Graham, 2011; Habermehl, 1994). Five Androctonuspecies are considered as dangerous to man, particularly Androc-onus mauritanicus (Pocock, 1902) and thewidespread Androctonus∗ Corresponding author. Tel.: +351 916712100.E-mail address: [email protected] (A. van der Meijden).
ttp://dx.doi.org/10.1016/j.actatropica.2014.02.002001-706X/© 2014 Elsevier B.V. All rights reserved.
australis (Linnaeus, 1758), which are the most dangerous Androc-tonus in the Maghreb region (Morocco, Algeria, Tunisia) (Goyffonand Guette, 2005). A. australis is known for envenomating humansand possessing a high toxicity (LD50 = 0.32 mg/kg in mice; Wattand Simard, 1984). For this reason, A. australis was one of the firstspecies of scorpions to have its venom purified for neurotoxin char-acterization (Miranda et al., 1966). As in snakes (Daltry et al., 1996;Prasad et al., 1999), scorpion venom is known to have consider-able intraspecific regional variation in composition (Devaux et al.,2004; El Ayeb and Rochat, 1985; Newton et al., 2007; Smertenkoet al., 2001), and thus a different response to antivenom treatment(Omran and McVean, 2000). Furthermore, other species such asAndroctonus amoreuxi (Audouin, 1826) may also cause more casesof scorpionism than currently thought (Goyffon et al., 2012). Itis therefore important to study the phylogeographical patterns of
Androctonus over a great part of their distribution as it may havedirect applications in therapeutic management.Androctonus is present in deserts and semi-arid regions fromTogo to Morocco in the Atlantic coast of Africa (Lourenc o and

4 Tropic
QwA1ALto
ecwtaweatvAdSMagfot(
btgpsvtil
lD
Fd
4 P. Coelho et al. / Acta
i, 2007; Lourenc o, 2008) to the Maghreb countries and Egypthere they are also present in relatively elevated areas like thetlas Mountains and the Sinai Peninsula mountain range (Vachon,952), the Middle East (Levy and Amitai, 1980), reaching acrossfghanistan (Vachon, 1958) to India (Tikader and Bastawade, 1983).ittle is known about the biogeography of Androctonus, althoughhe radiation of buthids has been associated with the aridificationf the Palearctic Region (Fet et al., 2003, 1998).
In this work, we assess five species of the Maghrebian countriesxcept for Libya, plus Egypt and the Sinai Peninsula. A. mauritani-us occurs in Morocco. This country shares a further three speciesith Algeria, Tunisia and/or Egypt (Androctonus liouvillei, A. aus-
ralis and A. amoreuxi), while Androctonus bicolor occurs in Algeriand Tunisia. A. amoreuxi is known to occur mostly in sandy desertshile A. australis and A. mauritanicus can be found in anthropogenic
nvironments (Stockmann and Ythier, 2010). The country in ourrea of study with the most species of Androctonus, Morocco, is alsohe most orographically diverse. The mountain chains that subdi-ide Morocco, North to south the Rif Mountains, Middle Atlas, Hightlas and Anti-Atlas Mountains, are known to be important in theiversification of the scorpion genus Buthus (Husemann et al., 2012;ousa et al., 2012; Pedroso et al., 2013). Similarly, the Tell Atlasountains and Aurès Mountains of Algeria and Tunisia are associ-
ted with Buthus (Pedroso et al., 2013). Although Androctonus areenerally not as orophilic as Buthus, these mountain chains mayorm a barrier for dispersal of the lowland species. However, at leastne Maghreb Androctonus species is known to be associated withhe Hoggar mountains in Southern Algeria; Androctonus hoggarensisPallary, 1929)
The scorpion genus Androctonus was first described by Ehren-erg in 1828. Vachon (1952) stabilized the genus’ taxonomy,ransforming it into a morphological and geographical coherentroup with seven species known in North Africa. Lourenc o (2005)roduced an important taxonomical revision of the genus: the sub-pecies of A. australis and A. mauritanicus were no longer consideredalid, Androctonus crassicauda gonneti Vachon, 1948 was raised tohe status of species, Androctonus aeneas C. L. Koch, 1839 was placedn the synonymy of A. bicolor Ehrenberg, 1828 and A. liouvillei (Pal-
ary, 1924) was raised to the species level.Recently, molecular tools have been used to assess the phy-ogeny of Androctonus in Tunisia. Ben Ali et al. (2000), using nuclearNA ITS regions (ITS-rDNA), found paraphyletic clades in three
ig. 1. Map representing the sampling locations across North Africa of Androctonus scorpioata, and are not shown. Symbols correspond to the phylogenetic clades (see Fig. 2).
a 134 (2014) 43–51
well-accepted taxa (A. bicolor, A. australis and A. amoreuxi). BenOthmen et al. (2004), using allozymic differentiation, found littlesupport for the monophyly of A. australis and A. amoreuxi individ-ually. However three well-supported monophyletic lineages using16S-rDNA, each corresponding to a species were recovered in a sub-sequent study, thus demonstrating the usefulness of this gene asa barcoding marker (Ben Othmen et al., 2009). Furthermore, theyfound a phylogeographic pattern of A. australis in Tunisia, whereeach of its two lineages are distributed to the north or south ofthe Chott el-Djerid salt lake, situated in central Tunisia. Howevertheir molecular dating makes it unlikely that the salt lake forma-tion has generated a vicariant evolution of Tunisian A. australis. Inrecent years, molecular studies using the mitochondrial COX1 genealready uncovered considerable cryptic diversity in other scorpiongenera in the Maghreb region (Gantenbein and Largiadèr, 2003;Sousa et al., 2012, 2011, 2010). However, a molecular phylogeneticstudy of the medically relevant scorpions of the genus Androctonusacross the Maghreb region has thus far not been performed.
In this study, we assess the patterns of diversity estimatedfrom COX1, 16S and 12S sequence data across the Maghreb regionand Egypt. A separate reduced dataset was also made from ND1sequences. We here provide the first molecular phylogeny forAndroctonus scorpions in Morocco, Algeria and Egypt.
2. Materials and methods
2.1. Taxon sampling
We collected samples in the field in three countries: Morocco,Tunisia and Egypt (Fig. 1). Algerian samples were donated by Dr.Said Larbes. Additional specimens were purchased through thepet-trade for which no locality data were available other than thecountry of origin. Sampling sites are illustrated in Fig. 1 and fur-ther details are provided in Table 1. Scorpions were captured withlong forceps and preserved in 96% ethanol. To minimize the impacton scorpion populations, non-lethal sampling methods were usedwhen possible, and consisted of removing the distal part of oneof the second leg pairs. The scorpion was subsequently placed
in the sand, facilitating the clotting process and thus minimiz-ing haemolymph loss. Scorpions can recover partially amputatedappendages after a few molts and it is common for scorpions to beactive and functional even when missing appendages (pers. obs.).ns. Inset shows Egyptian samples. Pet trade acquired samples were without locality
P. Coelho et al. / Acta Tropica 134 (2014) 43–51 45
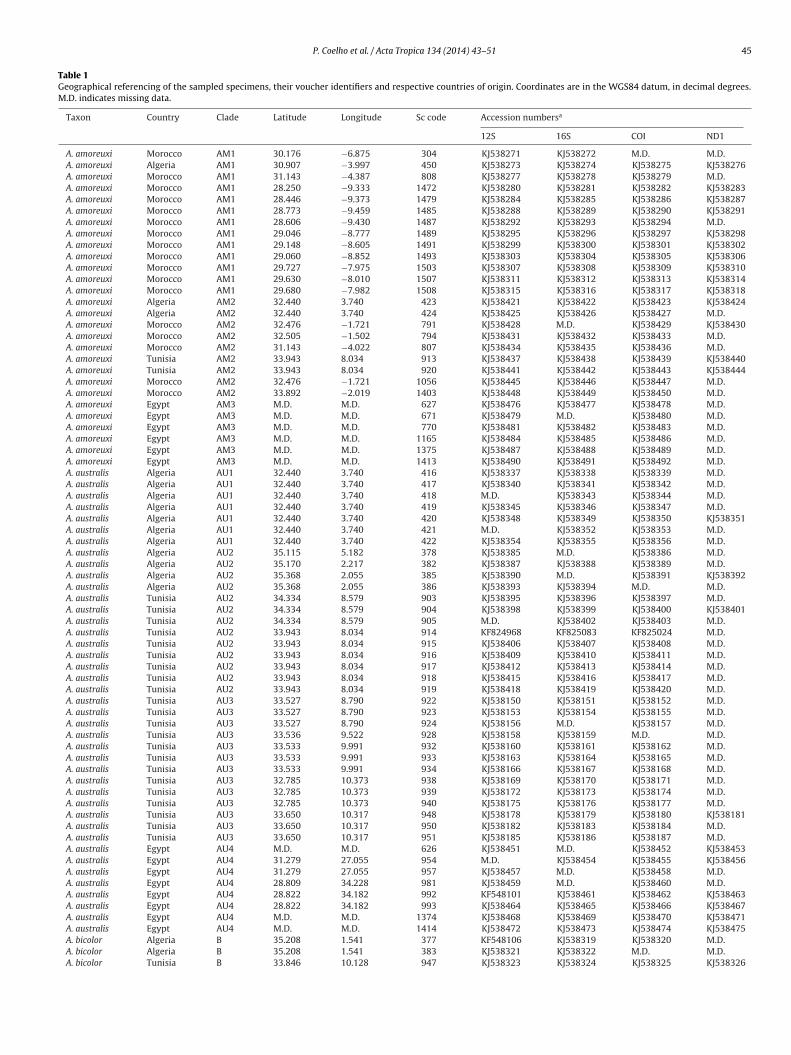
Table 1Geographical referencing of the sampled specimens, their voucher identifiers and respective countries of origin. Coordinates are in the WGS84 datum, in decimal degrees.M.D. indicates missing data.
Taxon Country Clade Latitude Longitude Sc code Accession numbersa
12S 16S COI ND1
A. amoreuxi Morocco AM1 30.176 −6.875 304 KJ538271 KJ538272 M.D. M.D.A. amoreuxi Algeria AM1 30.907 −3.997 450 KJ538273 KJ538274 KJ538275 KJ538276A. amoreuxi Morocco AM1 31.143 −4.387 808 KJ538277 KJ538278 KJ538279 M.D.A. amoreuxi Morocco AM1 28.250 −9.333 1472 KJ538280 KJ538281 KJ538282 KJ538283A. amoreuxi Morocco AM1 28.446 −9.373 1479 KJ538284 KJ538285 KJ538286 KJ538287A. amoreuxi Morocco AM1 28.773 −9.459 1485 KJ538288 KJ538289 KJ538290 KJ538291A. amoreuxi Morocco AM1 28.606 −9.430 1487 KJ538292 KJ538293 KJ538294 M.D.A. amoreuxi Morocco AM1 29.046 −8.777 1489 KJ538295 KJ538296 KJ538297 KJ538298A. amoreuxi Morocco AM1 29.148 −8.605 1491 KJ538299 KJ538300 KJ538301 KJ538302A. amoreuxi Morocco AM1 29.060 −8.852 1493 KJ538303 KJ538304 KJ538305 KJ538306A. amoreuxi Morocco AM1 29.727 −7.975 1503 KJ538307 KJ538308 KJ538309 KJ538310A. amoreuxi Morocco AM1 29.630 −8.010 1507 KJ538311 KJ538312 KJ538313 KJ538314A. amoreuxi Morocco AM1 29.680 −7.982 1508 KJ538315 KJ538316 KJ538317 KJ538318A. amoreuxi Algeria AM2 32.440 3.740 423 KJ538421 KJ538422 KJ538423 KJ538424A. amoreuxi Algeria AM2 32.440 3.740 424 KJ538425 KJ538426 KJ538427 M.D.A. amoreuxi Morocco AM2 32.476 −1.721 791 KJ538428 M.D. KJ538429 KJ538430A. amoreuxi Morocco AM2 32.505 −1.502 794 KJ538431 KJ538432 KJ538433 M.D.A. amoreuxi Morocco AM2 31.143 −4.022 807 KJ538434 KJ538435 KJ538436 M.D.A. amoreuxi Tunisia AM2 33.943 8.034 913 KJ538437 KJ538438 KJ538439 KJ538440A. amoreuxi Tunisia AM2 33.943 8.034 920 KJ538441 KJ538442 KJ538443 KJ538444A. amoreuxi Morocco AM2 32.476 −1.721 1056 KJ538445 KJ538446 KJ538447 M.D.A. amoreuxi Morocco AM2 33.892 −2.019 1403 KJ538448 KJ538449 KJ538450 M.D.A. amoreuxi Egypt AM3 M.D. M.D. 627 KJ538476 KJ538477 KJ538478 M.D.A. amoreuxi Egypt AM3 M.D. M.D. 671 KJ538479 M.D. KJ538480 M.D.A. amoreuxi Egypt AM3 M.D. M.D. 770 KJ538481 KJ538482 KJ538483 M.D.A. amoreuxi Egypt AM3 M.D. M.D. 1165 KJ538484 KJ538485 KJ538486 M.D.A. amoreuxi Egypt AM3 M.D. M.D. 1375 KJ538487 KJ538488 KJ538489 M.D.A. amoreuxi Egypt AM3 M.D. M.D. 1413 KJ538490 KJ538491 KJ538492 M.D.A. australis Algeria AU1 32.440 3.740 416 KJ538337 KJ538338 KJ538339 M.D.A. australis Algeria AU1 32.440 3.740 417 KJ538340 KJ538341 KJ538342 M.D.A. australis Algeria AU1 32.440 3.740 418 M.D. KJ538343 KJ538344 M.D.A. australis Algeria AU1 32.440 3.740 419 KJ538345 KJ538346 KJ538347 M.D.A. australis Algeria AU1 32.440 3.740 420 KJ538348 KJ538349 KJ538350 KJ538351A. australis Algeria AU1 32.440 3.740 421 M.D. KJ538352 KJ538353 M.D.A. australis Algeria AU1 32.440 3.740 422 KJ538354 KJ538355 KJ538356 M.D.A. australis Algeria AU2 35.115 5.182 378 KJ538385 M.D. KJ538386 M.D.A. australis Algeria AU2 35.170 2.217 382 KJ538387 KJ538388 KJ538389 M.D.A. australis Algeria AU2 35.368 2.055 385 KJ538390 M.D. KJ538391 KJ538392A. australis Algeria AU2 35.368 2.055 386 KJ538393 KJ538394 M.D. M.D.A. australis Tunisia AU2 34.334 8.579 903 KJ538395 KJ538396 KJ538397 M.D.A. australis Tunisia AU2 34.334 8.579 904 KJ538398 KJ538399 KJ538400 KJ538401A. australis Tunisia AU2 34.334 8.579 905 M.D. KJ538402 KJ538403 M.D.A. australis Tunisia AU2 33.943 8.034 914 KF824968 KF825083 KF825024 M.D.A. australis Tunisia AU2 33.943 8.034 915 KJ538406 KJ538407 KJ538408 M.D.A. australis Tunisia AU2 33.943 8.034 916 KJ538409 KJ538410 KJ538411 M.D.A. australis Tunisia AU2 33.943 8.034 917 KJ538412 KJ538413 KJ538414 M.D.A. australis Tunisia AU2 33.943 8.034 918 KJ538415 KJ538416 KJ538417 M.D.A. australis Tunisia AU2 33.943 8.034 919 KJ538418 KJ538419 KJ538420 M.D.A. australis Tunisia AU3 33.527 8.790 922 KJ538150 KJ538151 KJ538152 M.D.A. australis Tunisia AU3 33.527 8.790 923 KJ538153 KJ538154 KJ538155 M.D.A. australis Tunisia AU3 33.527 8.790 924 KJ538156 M.D. KJ538157 M.D.A. australis Tunisia AU3 33.536 9.522 928 KJ538158 KJ538159 M.D. M.D.A. australis Tunisia AU3 33.533 9.991 932 KJ538160 KJ538161 KJ538162 M.D.A. australis Tunisia AU3 33.533 9.991 933 KJ538163 KJ538164 KJ538165 M.D.A. australis Tunisia AU3 33.533 9.991 934 KJ538166 KJ538167 KJ538168 M.D.A. australis Tunisia AU3 32.785 10.373 938 KJ538169 KJ538170 KJ538171 M.D.A. australis Tunisia AU3 32.785 10.373 939 KJ538172 KJ538173 KJ538174 M.D.A. australis Tunisia AU3 32.785 10.373 940 KJ538175 KJ538176 KJ538177 M.D.A. australis Tunisia AU3 33.650 10.317 948 KJ538178 KJ538179 KJ538180 KJ538181A. australis Tunisia AU3 33.650 10.317 950 KJ538182 KJ538183 KJ538184 M.D.A. australis Tunisia AU3 33.650 10.317 951 KJ538185 KJ538186 KJ538187 M.D.A. australis Egypt AU4 M.D. M.D. 626 KJ538451 M.D. KJ538452 KJ538453A. australis Egypt AU4 31.279 27.055 954 M.D. KJ538454 KJ538455 KJ538456A. australis Egypt AU4 31.279 27.055 957 KJ538457 M.D. KJ538458 M.D.A. australis Egypt AU4 28.809 34.228 981 KJ538459 M.D. KJ538460 M.D.A. australis Egypt AU4 28.822 34.182 992 KF548101 KJ538461 KJ538462 KJ538463A. australis Egypt AU4 28.822 34.182 993 KJ538464 KJ538465 KJ538466 KJ538467A. australis Egypt AU4 M.D. M.D. 1374 KJ538468 KJ538469 KJ538470 KJ538471A. australis Egypt AU4 M.D. M.D. 1414 KJ538472 KJ538473 KJ538474 KJ538475A. bicolor Algeria B 35.208 1.541 377 KF548106 KJ538319 KJ538320 M.D.A. bicolor Algeria B 35.208 1.541 383 KJ538321 KJ538322 M.D. M.D.A. bicolor Tunisia B 33.846 10.128 947 KJ538323 KJ538324 KJ538325 KJ538326

46 P. Coelho et al. / Acta Tropica 134 (2014) 43–51
Table 1 (Continued )
Taxon Country Clade Latitude Longitude Sc code Accession numbersa
12S 16S COI ND1
A. bicolor Tunisia B 33.650 10.317 949 KJ538327 KJ538328 KJ538329 KJ538330A. bicolor Egypt B M.D. M.D. 1093 KJ538331 KJ538332 KJ538333 KJ538334A. bicolor Egypt B M.D. M.D. 1371 KJ538335 KJ538336 M.D. M.D.A. cf. gonneti Morocco G 28.479 −11.212 1453 KJ538372 KJ538373 KJ538374 M.D.A. cf. Gonneti Morocco G 28.479 −11.212 1454 KJ538375 KJ538376 KJ538377 KJ538378A. cf. Gonneti Morocco G 28.479 −11.212 1455 KJ538379 KJ538380 KJ538381 KJ538382A. cf. Gonneti Morocco G 28.479 −11.212 1456 KJ538383 KJ538384 M.D. M.D.A. liouvillei Morocco L 30.668 −6.380 210 KJ538188 KJ538189 M.D. M.D.A. liouvillei Morocco L 34.286 −3.169 780 KJ538190 M.D. KJ538191 KJ538192A. liouvillei Morocco L 34.286 −3.169 781 KJ538193 KJ538194 KJ538195 M.D.A. liouvillei Morocco L 33.892 −2.019 786 KJ538196 KJ538197 KJ538198 M.D.A. liouvillei Morocco L 33.213 −2.019 787 KJ538199 KJ538200 KJ538201 M.D.A. liouvillei Morocco L 32.087 −1.241 793 KJ538202 KJ538203 KJ538204 KJ538205A. liouvillei Morocco L 32.571 −2.015 796 KJ538206 KJ538207 KJ538208 M.D.A. liouvillei Morocco L 32.571 −2.015 797 KJ538209 KJ538210 KJ538211 KJ538212A. liouvillei Morocco L 33.289 −3.780 816 KJ538213 KJ538214 KJ538215 M.D.A. liouvillei Morocco L 33.508 −3.528 817 KJ538216 KJ538217 M.D. M.D.A. liouvillei Morocco L 33.892 −2.019 830 KJ538218 KJ538219 KJ538220 M.D.A. liouvillei Morocco L 32.087 −1.241 839 KJ538221 KJ538222 KJ538223 M.D.A. liouvillei Morocco L 33.213 −2.019 1055 KJ538224 KJ538225 KJ538226 M.D.A. liouvillei Morocco L 33.892 −2.019 1207 KJ538227 KJ538228 KJ538229 M.D.A. liouvillei Morocco L 33.892 −2.019 1394 KJ538230 KJ538231 KJ538232 M.D.A. mauritanicus Morocco M1 32.225 −8.166 15 KJ538253 KJ538254 KJ538255 KJ538256A. mauritanicus Morocco M1 32.526 −7.863 287 KJ538257 KF825084 KJ538258 KJ538259A. mauritanicus Morocco M1 32.661 −7.793 290 KJ538260 KJ538261 KJ538262 M.D.A. mauritanicus Morocco M1 32.661 −7.793 291 KJ538263 KJ538264 KJ538265 KJ538266A. mauritanicus Morocco M1 32.661 −7.793 292 KJ538267 KJ538268 M.D. M.D.A. mauritanicus Morocco M1 M.D. M.D. 625 KJ538269 M.D. KJ538270 M.D.A. mauritanicus Morocco M2 30.183 −9.580 1467 KJ538233 KJ538234 KJ538235 KJ538236A. mauritanicus Morocco M2 30.183 −9.580 1468 KJ538237 KJ538238 KJ538239 KJ538240A. mauritanicus Morocco M2 30.183 −9.580 1469 KJ538241 KJ538242 KJ538243 M.D.A. mauritanicus Morocco M2 30.159 −8.481 1549 KJ538244 KJ538245 M.D. KJ538246A. mauritanicus Morocco M2 30.059 −9.084 1558 KJ538247 KJ538248 KJ538249 KJ538250A. mauritanicus Morocco M2 30.098 −8.938 1589 KJ538251 KJ538252 M.D. M.D.A. mauritanicus Algeria M3 29.068 −10.248 438 KJ538357 KJ538358 KJ538359 M.D.A. mauritanicus Morocco M3 29.087 −9.898 1443 KJ538360 KJ538361 KJ538362 KJ538363
VaeV
etkOu
2
folua
RoTuRScP
A. mauritanicus Morocco M3 29.087 −9.898
A. mauritanicus Morocco M3 29.087 −9.898
oucher specimens were collected at almost all sites. All specimensre deposited in the collection of CIBIO, Centro de Investigac ãom Biodiversidade e Recursos Genéticos, Universidade do Porto,airão, Vila do Conde, Portugal.
Some key species that could not be sampled in the field,.g. Egyptian A. bicolor, were obtained through reputable animalraders. Morphological identification was done in the lab usingeys by Vachon (1952) and Lourenc o (2005). A single specimen ofpisthacanthus asper (Peters, 1861) (Scorpiones: Liochelidae) wassed as outgroup (details provided in Table 1).
.2. DNA extraction and PCR conditions
Fresh or preserved leg muscle tissue (or metasoma muscleor smaller specimens) was used for DNA extraction. Dissectionccurred, preferentially, from the third leg in order to minimize theoss of important taxonomical characters. Total DNA was extractedsing proteinase K digestion (10 mg/ml concentration) followed by
standard salt extraction protocol (Bruford et al., 1992).Four mitochondrial fragments were amplified: 16S ribosomal
NA (16S rRNA), 12S ribosomal RNA (12S rRNA), cytochrome Cxidase subunit 1 (COX1) and NADH dehydrogenase 1 (ND1) (seeable 2). Polymerase chain reactions were performed in a final vol-me of 25 �L and using 1.0 �L each of 10 pmol primer, 12.5 �L
EDTaq ReadyMix PCR Reaction Mix with MgCl2 (Sigma–Aldrich,t. Louis, USA), 1.0 �L of the purified DNA and 9.5 �L of water. Diffi-ult PCR reactions were amplified with a touchdown PCR approach.urified PCR templates were unidirectionally sequenced using1444 KJ538364 KJ538365 KJ538366 KJ5383671445 KJ538368 KJ538369 KJ538370 KJ538371
dye-labeled dideoxy terminator cycle sequencing on an ABI 3730XLat Macrogen Inc. The sequencing primers were the same as thoseused in the PCRs.
2.3. Data analysis
Chromatographs were checked manually for sequencing errorsusing FinchTV 1.4.0 (Geospiza, Inc.; Seattle, USA). Sequences wereedited using MEGA 5 (Tamura et al., 2011) and aligned using defaultsettings of MUSCLE (Edgar, 2004) for non-coding sequences andClustalW (Larkin et al., 2007) for COX1 and ND1.
Phylogeny reconstruction was performed using Maximum Like-lihood (ML) and Bayesian Inference (BI) methods. A homogeneitypartition test executed in Paup* 4.0 rejected homogeneity of thedifferent genes. We therefore carried out both a partitioned anda combined analysis. The best fitting models of sequence evo-lution were determined by the AIC criterion in JModeltest 2.1.2(Darriba et al., 2012) both for the separate genes and the com-bined dataset. ML tree searches were performed using PhyML,version 3.0 (Guindon et al., 2010). Bootstrap branch support valueswere calculated with 1000 replicates. The BI analysis was con-ducted with MrBayes 3.2.2, with 5,000,000 generations, samplingtrees every 10th generation (and calculating a consensus tree afteromitting the first 12,500 trees). Mean genetic distances between
and within clades were calculated with MEGA5 based on COX1sequences. Variance was estimated with 4000 bootstrap replicates,with uncorrected p-distances (Table 3). GenBank accession num-bers are given in Table 1.
P. Coelho et al. / Acta Tropica 134 (2014) 43–51 47
Table 2List of primers and PCR conditions used for molecular analyses. PCR conditions start with temperature (◦C) of each step followed by the time in seconds in brackets.
Gene Primer name Sequence (5′ → 3′) Source PCR conditions
16S rRNA18-mer (forward) CGATTTGAACTCAGATCA Simon et al. (1994) 94(180), [94(30), 50(45), 72(60)]
× 35, 72(300), 12(∞)20-mer (reverse) GTGCAAAGGTAGCATAAT Gantenbein et al. (2000)
12S rRNA12S-F AvdM AGAG-TGACGGGCAATATGTG
Van der Meijden et al. (2012)94(180), [94(30), 52(45), 72(60)]× 35, 72(300), 12(∞)12S-R AvdM CAGCGGCTGCGGTTATAC
COI
LCO1490 (forward) GGTCAACAAATCATCATAAAGATATTGGFolmer et al. (1994)
94(180), [94(30), 48(45), 72(60)]× 35, 72(300), 12(∞)HCO219 (reverse) TAAACTTCAGGGTGACCAAAAAATCA
COI avdm F WTYCTACIAATCAYAARGATATTGGVan der Meijden et al. (2012)
94(180), [94(30), 49(45), 72(60)]× 35, 72(300), 12(∞)COI avdm R TAMACYTCIGGGTGWCCAAAAAAYCA
Oow
3
3
a1otpig
3
Wtosbssc
TG4
ND1ND1-LR-N-12945 (forward) CGACCTCGATGTTGAATTAAND1-N1-J-12261 (reverse) TCGTAAGAAATTATTTGAGC
In addition we downloaded all 16S rDNA sequences in Benthmen et al. (2009) from GenBank and we combined these withur 16S dataset. Phylogeny reconstruction of this extended datasetas performed with ML (tree not shown).
. Results
.1. Sequence data
We sequenced 110 Androctonus specimens and one outgroup (O.sper) (Table 1). The combined dataset consists of an alignment of457 basepairs (bp) (606 bp of COX1, 374 bp of 16S rRNA and 477 bpf 12S rRNA). The dataset included 777 variable sites (53.3% of theotal nucleotide positions), 680 bp were constant (46.7%) and 511ositions were parsimony informative (35.1%). Due to difficulties
n amplifying ND1 only 41 sequences were available, and thus thisene was not included in the combined analysis.
.2. Overall phylogeny
Four of the six species were recovered as monophyletic (Fig. 2).e identified 13 clades that correspond to biogeographical and/or
axonomical units. Most of the deeper branches present low valuesf bootstrap support (ML) and posterior probability (BI). Con-equently, the relationships between species could not always
e resolved. ML and BI trees using a dataset that included ND1equences (not shown) did not differ in the topology of well-upported clades. Trees derived from the individual genes did notontradict the groupings recovered from the concatenated dataset.able 3enetic distances between and within groups. Numbers below the diagonal are mean p-d000 bootstrap replicates, are shown above the diagonal. Mean p-distances within clades
Distance between clades
AM1 0.010 0.011 0.011 0.011 0.011 0.011 0.011 0.010 0.AM2 0.075 0.011 0.012 0.011 0.012 0.012 0.010 0.011 0.AM3 0.092 0.089 0.012 0.011 0.011 0.011 0.012 0.012 0.AU1 0.090 0.102 0.090 0.007 0.009 0.011 0.012 0.011 0.AU2 0.087 0.102 0.093 0.039 0.010 0.010 0.012 0.011 0.AU3 0.090 0.098 0.097 0.061 0.069 0.009 0.012 0.011 0.AU4 0.094 0.093 0.089 0.080 0.080 0.056 0.013 0.011 0.B1 0.088 0.086 0.096 0.093 0.090 0.097 0.108 0.008 0.B2 0.077 0.082 0.100 0.093 0.087 0.087 0.096 0.042 0.B3 0.094 0.100 0.099 0.106 0.099 0.098 0.107 0.079 0.074
L 0.087 0.081 0.096 0.099 0.099 0.108 0.108 0.087 0.085 0.M1 0.084 0.098 0.095 0.087 0.087 0.081 0.096 0.085 0.080 0.M2 0.110 0.103 0.115 0.109 0.108 0.099 0.116 0.096 0.099 0.M3 0.103 0.104 0.097 0.107 0.102 0.100 0.107 0.109 0.104 0.G 0.101 0.087 0.111 0.100 0.104 0.096 0.102 0.109 0.109 0.
AM1 AM2 AM3 AU1 AU2 AU3 AU4 B1 B2 B
Hedin (1997)94(180), [94(30), 47(45), 72(60)]× 35, 72(300), 12(∞)
3.3. Species-level clades
A. amoreuxi, is divided into three clades: clades AM1 andAM2 contain specimens from Morocco, clade AM2 also containsspecimens from Tunisia and Algeria and clade AM3 consists of indi-viduals from Egypt. Clades AM1 and AM2 are placed as sister groupswith a high support (BI = 1.00; ML = 87) and the AM3 clade has nowell-supported affinities but may be sister taxon to A. australis. Thewestern part of the range, from Morocco to Tunisia, is divided intotwo clades; a northeastern (AM2) and a southwestern clade (AM1);the Southern Moroccan amoreuxi (AM1) clade extends South, fromthe regions of Assa and Zag in the South, to the southern area ofthe Anti-Atlas Mountains and North toward Taouz, in the Erfoudarea, close to the Morocco–Algerian border (Fig. 1). The A. amoreuxiclade AM2 groups specimens from three countries: in Morocco itis distributed in the Erfoud area and also to the areas surroundingBouarfa and Ain-Beni-Mathar (High Plateau in Northeast Morocco);In Algeria it is represented in one location, Ghardaia (between theGreat Ergs); in Tunisia it is represented in one location, close toTozeur (between Chott el-Gharsa and Chott el-Djerid salt lakes).The third clade, AM3 groups specimens that were acquired via thepet-trade from Egypt, none of which are georeferenced. Pairwisegenetic distances between these clades are in a range between7.5% and 9.2% (Table 3). Downloaded 16S sequences of A. amoreuxiare split between clades AM1 and AM2 with a sequence from BenOthmen et al. (2009) (andr05) placed in the AM2 clade whereas thesequence of Fet et al. (2003) is placed within our AM1 clade (tree
not shown).Clade L (the liouvillei clade) unites all Moroccan A. liouvilleiscorpions. A total of 15 specimens were collected, ranging fromthe High-Atlas Mountains, to the Northeastern range of the
istances calculated between clades whereas the corresponding variances, based on are shown to the right.
Within clades
011 0.010 0.010 0.011 0.011 0.012 0.016 0.002 A1011 0.010 0.011 0.011 0.011 0.011 0.013 0.003 A2011 0.011 0.011 0.012 0.011 0.013 0.003 0.001 A3012 0.012 0.011 0.012 0.012 0.012 0.004 0.002 AU1012 0.011 0.011 0.011 0.011 0.012 0.012 0.003 AU2011 0.012 0.010 0.011 0.011 0.012 0.009 0.002 AU3012 0.012 0.011 0.012 0.012 0.012 0.004 0.001 AU4011 0.011 0.012 0.011 0.012 0.012 n/c n/c B1010 0.011 0.011 0.011 0.011 0.013 0.010 0.004 B2
0.011 0.010 0.011 0.011 0.012 n/c n/c B3083 0.011 0.011 0.011 0.011 0.008 0.002 L090 0.096 0.011 0.011 0.012 0.012 0.003 M1094 0.099 0.105 0.010 0.011 0.023 0.004 M2095 0.098 0.093 0.090 0.011 0.014 0.003 M3106 0.085 0.111 0.104 0.088 0.004 0.002 G
3 L M1 M2 M3 G p-Distance St. err.

48 P. Coelho et al. / Acta Tropica 134 (2014) 43–51
Fig. 2. Bayesian estimate of phylogenetic relationships of Androctonus (outgroup not shown). Posterior probabilities values and bootstrap support values are shown aboveand below nodes respectively. These samples are marked with the same colors and shapes in all figures.

Tropic
MAbp
sStmpsssbd
ihfM
Tac(biatsstfCsrfvtpfAet
Noitcdsc
4
hADtlbrg
P. Coelho et al. / Acta
iddle-Atlas and to the High Plateau along the Algerian borderland.verage genetic distance within the clade is of 0.8%. It appears toe sister taxon to A. bicolor (ML 85% bootstrap support, BI posteriorrobability 0.96).
A. mauritanicus, is presented in three clades; M2 and M3 areister clades (BI = 0.60; ML = 80) whereas M1 was unrelated to these.pecimens from the M1 clade are located in the northern range ofhe Marrakech-Tensif-El Haouz administrative region. The speci-
ens from the M2 clade occur in a horizontal stretch from theeaks of the Anti-Atlas along the Souss valley with the westernmostample being 6 km from the seacoast. The M3 clade is comprised ofpecimens from two sampling locations, both from the southwestlope of the Anti-Atlas Mountains (see Fig. 1). Genetic distancesetween clades range from 9.0% to 10.5% while the highest geneticistance within these clades, in clade M2, is 2.3% (Table 3).
A group of four dark-colored Androctonus, which morpholog-cally resemble Androctonus gonneti most closely, are designatedere as A. cf. gonneti (see Section 4). These specimens (clade G) were
ound in one sampling point in the southern limit of the Anti-Atlasountains near the mouth of the Oued Draa.A. australis forms a monophyletic unit, divided into four clades.
he AU3 (east Tunisian australis) and AU4 (Egyptian australis) cladesre sister groups in the Bayesian tree (BI = 0.80), which is not thease in the ML analysis. Clades AU1 (Algerian australis) and AU2northern Algeria and west Tunisian australis) are sister groups inoth trees (BI = 0.95; ML = 99). Starting from the East, the AU4 clade
s exclusive to Egypt. Some of the specimens present in this cladere not geographically referenced, as they were obtained throughhe pet-trade. The AU3 clade (East Tunisian australis) is distributedolely in Tunisia: specimens range Southeast of the Chott el-Djeridalt lake, north to the Chot el-Fejaj salt lake and as far south ashe Tataouine desert region (Fig. 1). The AU2 clade is distributedrom Algeria in the east, and westward to the Chott el-Djerid andhot el-Gharsa salt lakes in Tunisia. The AU1 clade is comprised ofpecimens from a single location between the Great Ergs of Alge-ia (Fig. 1). Pairwise genetic distances between these clades rangerom 3.9% to 8.0%. The AU1 and AU4 clades present the lowestalues of within group genetic distance (0.4%). Among A. australis,he highest within group distance was found in clade AU2 (mean-distance = 1.2%). Our samples from the Sinai Peninsula confirmormer reports on this species’ occurrence in the Sinai (Levy andmitai, 1980). The extended 16S dataset shows that Ben Othment al. (2009)’s “northern and southern lineage” sequences are placedhroughout the AU2 and AU3 clades respectively (tree not shown).
The B clade (bicolor clade) unites all A. bicolor specimens.otably, this clade has longer branches within it than any of thether clades in this study. For this reason we divided this cladento three subclades (B1, B2 and B3) and quantified the genetic dis-ances between them. A. bicolor is present in the Tell Atlas in Algeria,entral Tunisia (Fig. 1) and Egypt (no GPS point available). Geneticistances between these subclades range from 4.2% to 7.9%. Theseubclades were too small for mean p-distance within clades to bealculated.
. Discussion
Genetic methods, using one or very few mitochondrial genes,ave proven very successful in uncovering cryptic diversity in Northfrican scorpions (Froufe et al., 2008; Sousa et al., 2012, 2011).espite using three genes with a high number of informative sites,
he dataset did not provide sufficient resolution above the species
evel. The relationships among these species therefore could note resolved. Species-level clades, however, were well-resolved andeceived high support, and we resolved several novel and biogeo-raphically coherent clades within the species.a 134 (2014) 43–51 49
For the past 20 M.a. the paleoclimatic history of North Africahas been characterized by drastic and oscillating events. TheMediterranean basin experienced extreme changes from the clos-ing of the Mediterranean Sea’s eastern end (15–19 M.a.) to itsisolation from the Atlantic Ocean leading to the Messinian salin-ity crisis (5.96–5.33 M.a.) producing significant sea-level changes.Afterwards, the violent re-flooding of the Mediterranean basinsubmerged large areas of North Africa and created islands in cur-rent Morocco and Algeria (Krijgsman et al., 1999; Steininger andRogl, 1984). Furthermore, during the last 10 M.a. glacial cycles pro-duced climatic oscillations between wetter and drier conditions inNorth Africa. Consequently, the Saharan desert experienced con-tractions and expansions depending on the formation of heavilyvegetated landscapes or hydrographic systems (Douady et al., 2003;Schuster et al., 2006). Fragmentation during these climatic oscilla-tions of more xeric habitats (Drake et al., 2011; Le Houérou, 1997),such as those favored by Androctonus species, may have also con-tributed to the isolation and subsequent diversification in thesescorpions.
The Atlas Mountains, formed in the mid-late Miocene, are amajor biogeographical barrier, and are thought to be the cause ofdiversification in several taxa (e.g. Fonseca et al., 2009; Gonc alveset al., 2012; Habel et al., 2012), and seem to have also played a role inMoroccan Androctonus. The distribution of the clades of A. mauritan-icus suggests that the mountain ranges could act as barriers to geneflow. A. mauritanicus is found in median and low altitudes (e.g. cladeM2 and M3). The High-Atlas is situated between M1 and M2 cladesand the Anti-Atlas between M2 and M3 clades. It seems from thispattern that orography might be a probable cause for differentiationin this species, as is the case in Buthus scorpions (Habel et al., 2012).Alternatively, the rivers associated with these mountains may havealso formed north–south barriers to dispersion, thus allowing dif-ferentiation within A. mauritanicus. Vachon (1952) described twosubspecies in Morocco: A. m. mauritanicus and A. m. bourdoni. Thedistribution of clade M1 largely coincides with the distribution ofthe former subspecies, while the distribution of clade M2 coincideswith the distribution of the latter. Although Lourenc o (2005) didnot find evidence to support these two subspecies, the genetic dis-tances between them are at a level of those found between fullspecies of Androctonus (Table 3). In addition, clades M2 and M3are also separated by a similar species-level genetic distance. Anexploratory morphological analysis of the specimens of clade M3found a very granulated prosoma and tergites, which does not fit thedescription of A. mauritanicus. These morphological characters alsodistinguish them from the specimens in clade M2, showing thesescorpions are distinct both genetically and morphologically. How-ever, a more detailed morphological assessment is clearly neededprior to a taxonomic revision. A. mauritanicus is, together with A.australis, among the most dangerous scorpions in the Maghreb notonly due to their venom toxicity but also because these speciesoccurrence often overlaps regions with relatively high human pop-ulation densities (Chippaux and Goyffon, 2008). Between 1996 and2006 in Morocco, 53% of deaths that resulted from sting casescome from incidents with A. mauritanicus (Touloun et al., 2012).The genetically highly divergent subpopulations of A. mauritanicusmay, similar to A. australis (Devaux et al., 2004) and other scorpionspecies (El Ayeb and Rochat, 1985; Newton et al., 2007; Smertenkoet al., 2001), display differences in venom composition. This shouldbe taken into consideration in both the development of antiseraand the treatment of scorpion stings.
A. liouvillei, although distributed in at least as large an area asA. mauritanicus, is not as deeply split into separate clades. This
may be due to the less orographically complex area it inhabits. Itis mainly found on the high plateau east of the Middle Atlas, andeastward into Algeria. Although the A. liouvillei clade is sister tothe A. bicolor clade, genetic distances between them are near the
5 Tropic
sa
A(AcramitAdttgotwbp2Atseoitnabbwtetemtct
iEsiAcelnrt(APbp
RAcws
References
Ben Ali, Z., Boursot, P., Said, K., Lagnel, J., Chatti, N., Navajas, M., 2000. Compari-
0 P. Coelho et al. / Acta
pecies-level, corroborating Lourenc o’s (2005) morphology-basedssessment that A. liouvillei is a distinct species from A. bicolor.
A. australis is one of the most widespread scorpions in Northfrica and one of the leading causes of scorpionism in North Africa
Chippaux and Goyffon, 2008). There is a deep split between cladesU1 and AU2 on one side, and clades AU3 and AU4 on the other. Thelades in Tunisia (clades AU2 and AU3) are therefore more closelyelated to clades distributed in countries as far east as Egypt (AU4)nd as far west as Algeria (AU1) than to each other. Substantial poly-orphism for three toxins in the venom of A. australis was reported
n Tunisia (Devaux et al., 2004). Differences were found betweenhe venoms of specimens from localities corresponding to cladesU2 or AU3. This shows that it is highly likely that phylogeneticallyivergent groups may have different venom compositions in A. aus-ralis. The genetic distance also reflects a higher distance betweenhem than between the pairs AU1–AU2 and AU3–AU4. This sug-ests either a strong biogeographic barrier between these clades,r that their current proximity is an area of potential secondary con-act. There is a large salt lake, Chott el-Djerid, between the regionshere clades AU2 and AU3 are in closest proximity. Salt lakes as
ig as Chott el-Djerid might act as physical barriers for scorpion dis-ersion (Ben Othmen et al., 2009) and other taxa (Kornilios et al.,010). Although Ben Othmen et al. (2009) dealt only with Tunisianndroctonus, had fewer samples, and used only 16S rRNA data,hey also found a deep divergence between the north-western andouth-eastern Tunisian A. australis. Their “northern” and “south-rn lineages” are genetically and geographically congruent withur AU2 and AU3 clades respectively. They explored the possibil-ty of the lake’s formation causing the vicariant event. Howeverhe relatively recent dates for the lake’s formation (90–150 ka) doot coincide with their divergence time estimation (3.43–9.90 Ma),nd they suggest that the recently formed salt lake acts only as aarrier to secondary contact, rather than having caused vicarianceetween these two populations. Their divergence time estimateas not based on calibration points, but rather on a constant muta-
ion rate, and should therefore be regarded with caution. Pedrosot al. (2013) have found a similar high divergence between popula-ions of Buthus scorpions to the North-West and South-East of Chottl-Djerid. Due to a lack of dependable internal calibration points forolecular dating, we have refrained from calculating divergence
ime estimates in our study, and placing the divergences betweenlades and species in a paleogeographic or paleoclimatic contextherefore remains tentative.
Surprising was the find of two distinct clades of A. amoreuxin Morocco. The distribution of clades AM1 and AM2 meet in therfoud area. Although there are no obvious geographical formationseparating them, the genetic distance between clades is 7.5%, whichs similar to the distance recovered between the A. australis clades.s currently recognized the species appears to be paraphyletic, withlade AM3 sister to A. australis although with very weak support lev-ls. Although some A. amoreuxi specimens were separated by theargest geographic distance of any species in this study, we couldot find morphological differences between them in taxonomicallyelevant characters (Lourenc o, 2005; Vachon, 1952). The fact thathe genetic distance between clades AM2 and AM3 is relatively high8.9%) corroborates the division of A. amoreuxi into two subspecies:. a. amoreuxi in the Maghreb and A. a. levyi Fet, 1997 in the Sinaieninsula. However, to what taxonomic level these groups shoulde ascribed requires further morphological study of the interveningopulations.
Near Tan Tan Plage, a coastal town near the mouth of the Draaiver, we found four specimens morphologically different from
. mauritanicus and Androctonus sergenti Vachon, 1948 (also, darkolored species). Morphologically, they seem to be closer relatedith A. gonneti. The 4 adult male specimens we captured have theame pectinal teeth count (33) as reported by Vachon (1952) for A.
a 134 (2014) 43–51
gonnetti. A. liouvillei and A. sergenti are reported to have a smallerpectinal teeth count (Lourenc o, 2005; Vachon, 1952). As no otherspecies are known for that region, we designated the G clade speci-mens as A. cf gonneti, a species known from this region. Our estimateof relationships based on mtDNA clearly indicates that these aredistinct from A. liouvillei (Fig. 2). Further sampling will be neces-sary to study the relationships between this population and otherspecies from Morocco, but it appears to be A. gonneti, a variant ofthis species, or an undescribed new species.
Summarizing, we were able to identify cryptic variation in A.australis, A. mauritanicus and A. amoreuxi. Some cases were sim-pler, in which we could corroborate the validity of a distinctgroup, such as A. liouvillei. Further work should aim at sequenc-ing nuclear genes in order to resolve deeper relationships withinthe genus. It would also be very interesting to obtain samples ofother species like Androctonus aleksandrplotkini, Androctonus hog-garensis, Androctonus maroccanus Lourenc o et al., 2009, A. sergentiand A. gonneti. Those, together with a more extensive sampling ofAlgeria, Libya and Mauritania could provide a prime perspectiveon the patterns and processes shaping Androctonus evolution andcurrent distribution in North Africa.
5. Conclusions
We here show that A. australis, A. mauritanicus and A. amoreuxihave deeply divergent subclades. Such variation can be reflected inthe venom these animals produce, as seen in other scorpion species(Borges et al., 2010; Newton et al., 2007; Omran and McVean, 2000;Smertenko et al., 2001). We therefore suggest that the venom of A.australis, A. mauritanicus and A. amoreuxi should be studied withthese deep divergences in mind. Antivenom developed for scor-pions in one region should be tested for efficacy against venomsfrom regions (Fatani et al., 2010) where scorpions correspond todifferent clades as identified here.
Furthermore despite lack of basal resolution, our results haveclear bearing on the taxonomic status of some of the (sub)speciesincluded in our study. In addition, we identified a population of A.gonneti-like specimens near Tan-Tan (clade G) which merits furtherstudy as a potential new species.
Acknowledgements
We are indebted to Dr. Said Larbes of the Département de Biologie,Faculté des Sciences Biologiques et Agronomiques, Université M. Mam-meri, Tizi-Ouzou, Algeria for supplying important samples fromAlgeria. We thank Abdullah M. Nagy of the St. Catherine wildlifepreserve, Sérgio Henriques, Diana Pedroso and our colleagues at theCIBIO institute for assistance in the field. Thanks to Arendo Flipsefor supplying some of the Egyptian scorpions. This work was fundedby Fundac ão para a Ciência e Tecnologia, BIABDE/74349/2006(to D.J.H.), SFRH/BD/74934/2010 (to P.S.), SFRH/BPD/48042/2008(A.v.d.M.) and a FCT I&D project (PTDC/BIA-EVF/2687/2012) toA.v.d.M. DJH is supported by “Genomics and evolutionary biology”co-financed by North Portugal Regional Operational Programme2007/2013 (ON.2 – O Novo Norte), under the NSRF, through theEuropean Regional Development Fund (ERDF).
son of ribosomal ITS regions among Androctonus spp. scorpions (Scorpionida:Buthidae) from Tunisia. J. Med. Entomol. 37, 787–790.
Ben Othmen, A., Chatti, N., Ben Ali-Haouas, Z., Ouldbrahim, I., Said, K., 2004.Allozymic differentiation of Tunisian populations of Androctonus species andButhus occitanus (Scorpiones: Buthidae). Biol. J. Linn. Soc. 81, 255–265.

Tropic
B
B
B
C
D
D
D
D
D
E
E
F
F
F
F
F
F
G
G
G
G
G
G
G
H
H
H
H
P. Coelho et al. / Acta
en Othmen, A., Said, K., Mahamdallie, S.S., Testa, J.M., Haouas, Z., Chatti, N., Ready,P.D., 2009. Phylogeography of Androctonus species (Scorpiones: Buthidae) inTunisia: diagnostic characters for linking species to scorpionism. Acta Trop. 112,77–85.
orges, A., Bermingham, E., Herrera, N., Alfonzo, M.J., Sanjur, O.I., 2010. Molecularsystematics of the neotropical scorpion genus Tityus (Buthidae): the histori-cal biogeography and venom antigenic diversity of toxic Venezuelan species.Toxicon 55, 436–454.
ruford, M.W., Hanotte, O., Brookfield, J.F.Y., Burke, T., 1992. Single-locus and mul-tilocus DNA fingerprinting. In: Hoelzel, A.R. (Ed.), Molecular Genetic Analysis ofPopulations: A Practical Approach. IRL Press, Oxford, pp. 225–270.
hippaux, J.-P., Goyffon, M., 2008. Epidemiology of scorpionism: a global appraisal.Acta Trop. 107, 71–79.
altry, J., Wuester, W., Thorpe, R., 1996. Diet and snake venom evolution. Nature379, 537–540.
arriba, D., Taboada, G.L., Doallo, R., Posada, D., 2012. jModelTest 2: more models,new heuristics and parallel computing. Nat. Methods 9, 772.
evaux, C., Jouirou, B., Naceur Krifi, M., Clot-Faybesse, O., El Ayeb, M., Rochat, H.,2004. Quantitative variability in the biodistribution and in toxinokinetic studiesof the three main alpha toxins from the Androctonus australis hector scorpionvenom. Toxicon 43, 661–669.
ouady, C.J., Catzeflis, F., Raman, J., Springer, M.S., Stanhope, M.J., 2003. The Sahara asa vicariant agent, and the role of Miocene climatic events, in the diversificationof the mammalian order Macroscelidea (elephant shrews). Proc. Natl. Acad. Sci.U.S.A. 100, 8325–8330.
rake, N.A., Blench, R.M., Armitage, S.J., Bristow, C.S., White, K.H., 2011. Ancientwatercourses and biogeography of the Sahara explain the peopling of the desert.Proc. Natl. Acad. Sci. U.S.A. 108, 458–462.
dgar, R.C., 2004. MUSCLE: multiple sequence alignment with high accuracy andhigh throughput. Nucleic Acids Res. 32, 1792–1797.
l Ayeb, M., Rochat, H., 1985. Polymorphism and quantitative variations of toxins inthe venom of the scorpion Androctonus australis Hector. Toxicon 23, 755–760.
atani, A.J., Ahmed, A.A.E., Abdel-Halim, R.M., Abdoon, N.A., Darweesh, A.Q., 2010.Comparative study between the protective effects of Saudi and Egyptianantivenoms, alone or in combination with ion channel modulators, against dele-terious actions of Leiurus quinquestriatus scorpion venom. Toxicon 55, 773–786.
et, V., Gantenbein, B., Gromov, A.A.V., Lowe, G., Lourenc o, W.R., 2003. The firstmolecular phylogeny of Buthidae (Scorpiones). Euscorpius 4, 1–10.
et, V., Polis, G.A., Sissom, W.D., 1998. Life in sandy deserts: the scorpion model. J.Arid Environ. 39, 609–622.
olmer, O., Black, M., Hoeh, W., Lutz, R., Vrijenhoek, R., 1994. DNA primersfor amplification of mitochondrial cytochrome c oxidase subunit I fromdiverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 3 (5), 294–299,http://dx.doi.org/10.1371/journal.pone.0013102.
onseca, M.M., Brito, J.C., Paulo, O.S., Carretero, M.A., Harris, D.J., 2009. System-atic and phylogeographical assessment of the Acanthodactylus erythrurus group(Reptilia: Lacertidae) based on phylogenetic analyses of mitochondrial andnuclear DNA. Mol. Phylogenet. Evol. 51, 131–142.
roufe, E., Sousa, P., Alves, P.C., Harris, D.J., 2008. Genetic diversity within Scorpiomaurus (Scorpiones: Scorpionidae) from morocco: preliminary evidence basedon CO1 mitochondrial DNA sequences. Biologia (Bratisl.) 63, 1157–1160.
antenbein, B., Kropf, C., Largiadèr, C.R., Scholl, A., 2000. Molecular and morpholog-ical evidence for the presence of a new Buthid taxon (Scorpiones: Buthidae)on the Island of Cyprus. Rev. Suisse Zool. 107 (1), 213–232, Retrieved fromhttp://www.cabdirect.org/abstracts/20000507188.html
antenbein, B., Largiadèr, C.R., 2003. The phylogeographic importance of the Straitof Gibraltar as a gene flow barrier in terrestrial arthropods: a case study withthe scorpion Buthus occitanus as model organism. Mol. Phylogenet. Evol. 28,119–130.
onc alves, D.V., Brito, J.C., Crochet, P.-A., Geniez, P., Padial, J.M., Harris, D.J., 2012.Phylogeny of North African Agama lizards (Reptilia: Agamidae) and the roleof the Sahara desert in vertebrate speciation. Mol. Phylogenet. Evol. 64 (3),582–591, http://dx.doi.org/10.1016/j.ympev.2012.05.007.
oyffon, M., Dabo, A., Coulibaly, S., Togo, G., Chippaux, J., 2012. Dangerous scorpionfauna of Mali. J. Venom. Anim. Toxins Incl. Trop. Dis. 18, 361–368.
oyffon, M., Guette, C., 2005. Scorpions dangereux du Niger. Bull. Soc. Pathol. Exot.98, 293–295.
raham, M.R., 2011. Scorpions of the World Scorpions of the World. Roland Stock-mann, Eric Ythier 2010. N.A.P. Editions, France, ISBN 978-2-913688-11-7, pp.567, J. Arachnol. 39, 166–167.
uindon, S., Dufayard, J.-F., Lefort, V., Anisimova, M., Hordijk, W., Gascuel, O., 2010.New algorithms and methods to estimate maximum-likelihood phylogenies:assessing the performance of PhyML 3.0. Syst. Biol. 59, 307–321.
abel, J.C., Husemann, M., Schmitt, T., Zachos, F.E., Honnen, A.-C., Petersen, B., Par-makelis, A., Stathi, I., 2012. Microallopatry caused strong diversification in Buthusscorpions (Scorpiones: Buthidae) in the Atlas Mountains (NW Africa). PLoS ONE7, e29403.
abermehl, G., 1994. Gift-Tiere und ihre Waffen: Eine Einführung für Biologen,Chemiker und Mediziner. Ein Leitfaden für Touristen, 5th ed. Springer Verlag,Heidelberg.
edin, M.C., 1997. Molecular phylogenetics at the population/species interface in
cave spiders of the southern Appalachians (Araneae:Nesticidae:Nesticus). Mol.Biol. Evol. 14 (3), 309–324.usemann, M., Schmitt, T., Stathi, I., Habel, J.C., 2012. Evolution and radiation in thescorpion Buthus elmoutaouakili Lourenco and Qi 2006 (Scorpiones: Buthidae) atthe foothills of the Atlas Mountains (North Africa). J. Hered. 103, 221–229.
a 134 (2014) 43–51 51
Kornilios, P., Kyriazi, P., Poulakakis, N., Kumlutas , Y., Ilgaz, C., Mylonas, M., Lym-berakis, P., 2010. Phylogeography of the ocellated skink Chalcides ocellatus(Squamata Scincidae), with the use of mtDNA sequences: a hitch-hiker’s guideto the Mediterranean. Mol. Phylogenet. Evol. 54, 445–456.
Krijgsman, W., Hilgen, F.J., Raffi, I., Sierro, F.J., Wilson, D.S., 1999. Chronology, causesand progression of the Messinian salinity crisis. Nature 400, 652–655.
Larkin, M.A., Blackshields, G., Brown, N.P., Chenna, R., McGettigan, P.A., McWilliam,H., Valentin, F., Wallace, I.M., Wilm, A., Lopez, R., Thompson, J.D., Gibson, T.J.,Higgins, D.G., 2007. Clustal W and Clustal X version 2.0. Bioinformatics 23,2947–2948.
Le Houérou, H.N., 1997. Climate, flora and fauna changes in the Sahara over the past500 million years. J. Arid Environ. 37, 619–647.
Levy, G., Amitai, P., 1980. Fauna Palaestina. Arachnida I: Scorpiones. Israel Academyof Sciences and Humanities, Jerusalem.
Lourenc o, W., Cuellar, O., 1995. Scorpions, scorpionism, life history strategies andparthenogenesis. J. Venom. Anim. Toxins 1, 51–62.
Lourenc o, W., Qi, J., 2007. A new species of Androctonus Ehrenberg, 1828 from Mau-ritania (Scorpiones, Buthidae). Bol. Soc. Entomol. Aragon. 1, 215–219.
Lourenc o, W.R., 2005. Nouvelles considérations taxonomiques sur les espèces dugenre Androctonus Ehrenberg, 1828 et description de deux nouvelles espèces(Scorpiones, Buthidae). Rev. Suisse Zool. 112, 145–171.
Lourenc o, W.R., 2008. A new species of Androctonus Ehrenberg, 1828 from Togo(Scorpiones, Buthidae). Entomol. Mitt. Zool Museum Hambg. 15, 37–44.
Miranda, F., Rochat, H., Rochat, C., Lissitzky, S., 1966. Complexes moléculairesprésentés par les neurotoxines animales—I, Neurotoxines des venins de scor-pions (Androctonus australis hector et Buthus occitanus tunetanus). Toxicon 4,123–136.
Newton, K.A., Clench, M.R., Deshmukh, R., Jeyaseelan, K., Strong, P.N., 2007. Massfingerprinting of toxic fractions from the venom of the Indian red scorpion,Mesobuthus tamulus: biotope-specific variation in the expression of venom pep-tides. Rapid Commun. Mass Spectrom. 21, 3467–3476.
Omran, M.A.A., McVean, A., 2000. Intraspecific variation in scorpion Leiurus quin-questriatus venom collected from Egypt (Sinai and Aswan Deserts). Toxin Rev.19, 247–264.
Pedroso, D., Sousa, P., Harris, D.J., Van der Meijden, A., 2013. Phylogeography ofButhus Leach, 1815 (Scorpiones: Buthidae): a multigene molecular approachreveals futher evolutionary history in the Maghreb. Afr. Invertebr. 48, 298–308,http://dx.doi.org/10.3377/004.048.0216.
Prasad, N., Uma, B., Bhatt, S., Gowda, V., 1999. Comparative characterisation of Rus-sell’s viper (Daboia/Vipera russelli) venoms from different regions of the Indianpeninsula. Biochim. Biophys. Acta 1428, 121–136.
Schuster, M., Duringer, P., Ghienne, J.-F., Vignaud, P., Mackaye, H.T., Likius, A., Brunet,M., 2006. The age of the Sahara desert. Science 311, 821.
Smertenko, A., Omran, M.A.A., Hussey, P.J., McVean, A., 2001. Toxin evolution inscorpion venom: evidence for toxin divergence under strong negative selectionin Leiurus quinquestriatus subspecies. Toxin Rev. 20, 229–244.
Simon, C., Frati, F., Beckenbach, A., Crespi, B., Liu, H., Flook, P., 1994. Evolution,weighting, and phylogenetic utility of mitochondrial gene sequences and a com-pilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am.87 (6), 651–700.
Sousa, P., Froufe, E., Alves, P., Harris, D.J., Alves, P.C., 2010. Genetic diversity withinscorpions of the genus Buthus from the Iberian Peninsula: mitochondrial DNAsequence data indicate additional distinct cryptic lineages. J. Arachnol. 28,206–211.
Sousa, P., Froufe, E., Harris, D.J., Alves, P.C., Van der Meijden, A., 2011. Geneticdiversity of Maghrebian Hottentotta (Scorpiones: Buthidae) scorpions based onCO1: new insights on the genus phylogeny and distribution. Afr. Invertebr. 52,135–143.
Sousa, P., Harris, D.J., Froufe, E., Meijden, A., 2012. Phylogeographic patterns of Buthusscorpions (Scorpiones: Buthidae) in the Maghreb and South-Western Europebased on CO1 mtDNA sequences. J. Zool. 288, 66–75.
Steininger, F.F., Rogl, F., 1984. Paleogeography and palinspastic reconstruction of theNeogene of the Mediterranean and Paratethys. Geological Society, London, Spe-cial Publications 17, 659–668, http://dx.doi.org/10.1144/GSL.SP.1984.017.01.52.
Stockmann, R., Ythier, E., 2010. Scorpions of the World. N.A.P. Editions, Verrières-le-Bruisson, France.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., Kumar, S., 2011. MEGA5:molecular evolutionary genetics analysis using maximum likelihood, evolution-ary distance, and maximum parsimony methods. Mol. Biol. Evol. 28, 2731–2739.
Tikader, B., Bastawade, D., 1983. The fauna of India. Scorpions (Scorpionida: Arach-nida), Arachnida, vol. 3. Zoological Survey of India, Calcutta.
Touloun, O., Boumezzough, A., Slimani, T., 2012. Scorpion envenomation in theregion of Marrakesh Tensift Alhaouz (Morocco): epidemiological characteriza-tion and therapeutic approaches. Serket 13, 38–50.
Vachon, M., 1952. Description des scorpions du Nord-Ouest de L’Afrique (Maroc,Algerie, Tunisie, Fezzan, Sahara et Sahel). A. Familie des Buthidae E. Simon,1879. [Part II: Genus Androctonus]. In: Etudes Sur Les Scorpions. Institut Pasteurd’Algérie, Alger, pp. 116–178.
Vachon, M., 1958. Scorpionidea (Chelicerata) de l’Afghanistan. The 3rd Danish Expe-dition to Central Asia. (Zoological Results 23). Vidensk. Meddelelser fra DanskNaturhistrorisk Foren. i Købehavn, vol. 120., pp. 121–187.
Van der Meijden, A., Kleinteich, T., Coelho, P., 2012. Packing a pinch: functional impli-cations of chela shapes in scorpions using finite element analysis. J. Anat. 220(5), 423–434, http://dx.doi.org/10.1111/j.1469-7580.2012.01485.x.
Watt, D.D., Simard, J.M., 1984. Neurotoxic proteins in scorpion venom. Toxin Rev. 3,181–221.
Copyright © 2022 FDOKUMEN




















