
Kinetics of endosome acidification detected by mutant and wild-type Semliki Forest virus

Combined effects of temperature and ocean acidification on the juvenile individualsof the mussel Mytilus chilensis
C. Duarte a,⁎, J.M. Navarro b, K. Acuña b, R. Torres c, P.H. Manríquez b,d, M.A. Lardies e,C.A. Vargas f, N.A. Lagos g, V. Aguilera f
a Departamento de Ecología y Biodiversidad, Facultad de Ecología y Recursos Naturales, Universidad Andres Bello, Santiago, Chileb Instituto de Ciencias Marinas y Limnológicas, Facultad de Ciencias, Universidad Austral de Chile, Valdivia, Chilec Centro de Investigación en Ecosistemas de la Patagonia (CIEP), Coyhaique, Chiled Laboratorio Costero de Recursos Acuáticos de Calfuco, Facultad de Ciencias, Universidad Austral de Chile, Valdivia, Chilee Facultad de Artes Liberales, Universidad Adolfo Ibáñez, Santiago, Chilef Laboratorio de Funcionamiento de Ecosistemas Acuáticos (LAFE), Unidad de Sistemas Acuáticos, Centro de Ciencias Ambientales EULA Chile, Universidad de Concepción, Concepción, Chileg Facultad de Ciencias, Universidad Santo Tomas, Ejército 146, Santiago, Chile
a b s t r a c ta r t i c l e i n f o
Article history:Received 15 February 2013Received in revised form 9 May 2013Accepted 1 June 2013Available online xxxx
Keywords:Global WarmingOcean AcidificationCalcification RateMussel
Anthropogenic CO2 emissions have led to increasing global mean temperatures (a process called globalwarming) and ocean acidification. Because both processes are occurring simultaneously, to better understandtheir consequences on marine species their combined effects must be experimentally evaluated. The aim ofthis study was to evaluate for the first time the combined effects of ocean acidification and water temperatureincrease on the total calcification rate, growth rate and survival of juvenile individuals of the mytilid musselMytilus chilensis (Hupe). Two temperature levels (12 and 16 °C) and three nominal CO2 concentrations (390,700 and 1000 ppm of CO2) were used. We found that the net rate of calcium deposition and total weightwere not significantly affected by temperature, but were negatively affected by the levels of CO2. Theinteractive effects of temperature and CO2 levels affected only the shell dissolution, but this process wasnot important for the animal's net calcification. These results suggest that individuals of M. chilensis areable to overcome increased temperatures, but not increments of CO2 levels. It is well known that musselsinfluence their physical and biological surroundings. Therefore, the negative effects of a CO2 increase couldhave significant ecological consequences, mainly in those habitats where this group is dominant in termsof abundance and biomass. Finally, taking into account that this species inhabit a wide geographic range,with contrasting environmental conditions (e.g., temperature, salinity and, pH), further studies are neededto evaluate the intraspecific variability in the responses of this species to different environmental stressors.
© 2013 Elsevier B.V. All rights reserved.
1. Introduction
Due to human activities, the content of atmospheric CO2 has in-creased by 100 ppm from the industrial revolution and today standsat 390 ppm (Solomon et al., 2007). It is expected that this concentra-tion will continue to rise (Caldeira and Wickett, 2003). This increasein atmospheric CO2 has led to increasing global mean temperatures(a process called global warming (GW)) and to ocean acidification(OA) (Caldeira and Wickett, 2003; Feely et al., 2008; Solomon et al.,2007).
The sea surface temperature has increased by approximately0.7 °C and it is expected that it will continue rising by between 1 °Cand 4 °C by the end of the century (IPCC, 2007). Several studies
have shown that elevated temperatures may significantly influencemarine biota, affecting the animal's physiology (specifically above or-ganisms' thermal optima or tolerance) and ecosystem processes (Hayet al., 2005; Hofmann and Todgham, 2010; Mieszkowska et al., 2006;Pörtner, 2010; Pörtner and Knust, 2007; Thomas et al., 2004). Forexample, mass coral bleaching events have been driven by tempera-ture increases in seawater (even with increases as small as 1 °C)(Hoegh-Guldberg, 2005). Peck et al. (2004) showed that small tem-perature increases (only a few degrees compared to the normal max-imum) negatively affected the locomotor activity (e.g., reburying) ofAntarctic marine mollusks (e.g., Nacella concinna).
Although less studied, OA has been also shown to affect marineorganisms (e.g., Gooding et al., 2009; Gutowska et al., 2008; Milleret al., 2009; Parker et al., 2011; Ries et al., 2009). OA is produced byan increase in CO2 in the seawater, which leads to a decrease inboth pH and the availability of carbonate ions (e.g., Orr et al., 2005;Wolf-Gladrow et al., 1999). The pH of the ocean surface is alreadynearly 0.1 units lower than the values registered in the preindustrial
Journal of Sea Research xxx (2013) xxx–xxx
⁎ Corresponding author at: Departamento de Ecología y Biodiversidad, Facultad deEcología y Recursos Naturales, Universidad Andres Bello, Republica 470, Santiago,Chile. Tel.: +56 2 27703650.
E-mail address: [email protected] (C. Duarte).
SEARES-01102; No of Pages 7
1385-1101/$ – see front matter © 2013 Elsevier B.V. All rights reserved.http://dx.doi.org/10.1016/j.seares.2013.06.002
Contents lists available at SciVerse ScienceDirect
Journal of Sea Research
j ourna l homepage: www.e lsev ie r .com/ locate /seares
Please cite this article as: Duarte, C., et al., Combined effects of temperature and ocean acidification on the juvenile individuals of the musselMytilus chilensis, Journal of Sea Research (2013), http://dx.doi.org/10.1016/j.seares.2013.06.002

epoch (Orr et al., 2005) and it is predicted to decrease by 0.4 units bythe end of the century and nearly 0.8 units within the next 300 years,while carbonate concentration could drop by about 50% by the end ofthe century (Caldeira and Wickett, 2003; Orr et al., 2005; Wolf-Gladrow et al., 1999). Both of these processes have the potential tonegatively affect marine calcifying organisms, complicating theformation of new shells or producing the disintegration of the alreadyformed shell (e.g., Caldeira and Wickett, 2003; Kleypas et al., 2006;Langdon and Atkinson, 2005; Langdon et al., 2000; Leclercq et al.,2000; Orr et al., 2005; Riebesell et al., 2000). Additionally, increasedCO2 produces hypercapnic conditions (or acidosis), which have beenshown to negatively affect the physiology, growth and reproductivesuccess of marine calcifier organisms (Arnold et al., 2009; Berge et al.,2006; Cummings et al., 2011; Kurihara, 2008; Michaelidis et al., 2005;Spicer et al., 2007; Widdicombe and Spicer, 2008).
There has been growing evidence that shows that many environ-mental stressors can act in a combinedmanner, thereby synergistically,antagonistically or additively affecting many physiological processes ofmarine organisms (Darling and Cote, 2008; Gooding et al., 2009;Schiedek et al., 2007). Although scarce, some recent studies haveshown that the effects of CO2 (the solubility of which depends on thetemperature) could be modified in environments with high tempera-tures (Findlay et al., 2010a; Gooding et al., 2009; Parker et al., 2009).For example, both temperature and CO2 increases affected shell degra-dation of the shelled pteropod Limacina helicina; however, while higherCO2 levels negatively affected shell degradation, higher temperaturevalues would help this species to counteract the loss of shell producedby the CO2 increase (Lischka et al., 2011). Combined effects of temper-ature and CO2 increases were also recorded by Wood et al. (2010),although with opposed results to those of Lischka et al. (2011). Theseauthors recorded that in higher temperatures, the CO2 increase reducedthe arm regeneration of the brittlestar Ophiura ophiura, a situation thatwas not observed when high CO2 levels were combined with lowtemperatures. Gooding et al. (2009) showed that increased tempera-ture and increased CO2 had positive and additive effects on the growthrate of the echinoderm Pisaster ochraceus; whilst these variables hadantagonistic effects on the calcified mass, since in high temperaturetreatments, the negative CO2 effects on the calcifiedmasswere reduced.Rosa and Seibel (2008) recorded that ocean acidification depressed themetabolic rate and activity levels of the squid Dosidicus gigas, and thatthis negative effect was exacerbated by high temperatures. The resultsof these studies highlight three important things: i) as global warmingand OA are occurring concomitantly, their combined effects must beevaluated, ii) the combined effects of global warming and OA are notonly the sum of their individual effects, iii) the responses of the organ-isms exposed to these environmental stressors are widely variable.Consequently, more studies are necessary to better understand howthe interaction between global warming and OA affects both theindividual organisms and the whole ecosystem.
The aim of this studywas to evaluate for the first time the combinedeffects of ocean acidification and the increase in water temperature onthe total calcification rate, growth rate and survival of the mytilidmussel Mytilus chilensis (Hupe). This species forms extensive subtidaland intertidal banks and has been shown to play an important ecologi-cal role (as do other mussel species, eg. Commito and Dankers, 2001;Gutiérrez et al., 2003), affecting significantly the community structureof the associated macrofauna and its physical surroundings (Duarte etal., 2006). In addition, M. chilensis represents an important species foraquaculture, since it is the most cultivated species in the south ofChile (over 200,000 tons. per year) under permanent submersion.Therefore, this species is a key organism for studying the effects pro-duced by ocean acidification and global warming, since their impactswill affect not only the ecosystem, but also the national economy. Wetherefore hypothesized that seawater concentrations of CO2 will nega-tively affect those processes and potentially that these effects could bemodulated by increments in seawater temperature.
2. Material and methods
2.1. Animal collection
Juvenile individuals of M. chilensis (~1.5 cm in size and ~350 mg infresh weight) were collected from culture ropes from Huelmo bay, atidal inlet located in southern Chile (ca. 40°S) during the austral winterof 2011 (water temperature: 11.5 °C; salinity: 33.4 ppm; and pH =8.01). The annual mean water temperature of the study area is 12 °C(±2.5 (Navarro and Jaramillo, 1994); meanwhile the water pH is8.09 ± 0.28 (unpublished data obtained in seasonal samplings during2011). The mussels were transported in chilled conditions to theLaboratorio Costero de Calfuco (LCC, ca. 39°S) and placed in five plasticbaskets (100 ! 100 ! 40 cm in height), where they were kept infiltered (0.1 μm) and aerated seawater (pH = 8.1 ± 0.01, tempera-ture = 13.1 °C ± 0.01 and salinity = ~33 ppm). To acclimate themto the mesocosm conditions, the mussels were kept for 2 weeks andfed daily with the microalgae Isochrysis galbana (~4 ! 106 cel/L) priorto the start of the experiments.
2.2. Experimental setup
After the acclimation period, the mussels were randomly assignedto one of six treatments: 12 °C and 390 ppm CO2, 12 °C and 700 ppmCO2, 12 °C and 1000 ppm CO2, 16 °C and 390 ppm CO2, 16 °C and700 ppm CO2 and 16 °C and 1000 ppm CO2. Each treatment was rep-licated five times (see Fig. 1). The experimental temperatures repre-sent the average surface value (12 °C) of the study area (Navarroand Jaramillo, 1994) and alternatively 3 °C above this average(approximate increase predicted by IPCC, 2007 to the end of thecentury). The CO2 concentrations were selected taking into accountthe rate of change projected by the years 2070–2110 (based on rateof change in pH predicted by the most extreme scenario (RCP8.5scenario) of atmospheric CO2 (see Meinshausen et al., 2011)). Thetemperatures were adjusted and maintained using external chillers.To obtain the different CO2 concentrations, the experimental con-tainers were adjusted following the methodology described byNavarro et al. (2013). Briefly, for 390 ppm, pure atmospheric airwas bubbled into experimental containers; for 700 and 1000 ppm,we blended dry air with pure CO2 to each target concentrationusing mass flow controllers (MFCs, www.aalborg.com) for air andCO2; then this blend was bubbled into the corresponding container.Dry and clean air was generated by compressing atmospheric air(117 psi) using an oil-free air compressor. The experimental con-tainers were constantly bubbled with the corresponding CO2 concen-tration. During the experiments, water pH and total alkalinity weremonitored every 3 d. pH measurements were done in a closed 25 mLcells thermostated at 25.0 °C using a Metrohm 713 pH meter (inputresistance > 1013 ! 0.1 mV sensitivity and nominal resolution0.001 pH units) and a glass combined double junction Ag/AgCl elec-trode (Metrohm model 6.0219.100) calibrated with 8.089 Tris–bufferat 25 °C; pH values are therefore reported on the total hydrogen ionscale (DOE, 1994). Temperature and salinity were measured using asmall CTD (Ocean Seven 305 Plus CTD, www.idronaut.it). Total alkalinitywas measured using the method of Haraldsson et al. (1997). The pH,total alkalinity (AT), phosphate (Strickland and Parsons, 1968) anddissolved silicate (Strickland and Parsons, 1968) were used to calculatethe rest of the carbonate system parameters and the saturation stage ofomega, aragonite and calcite using CO2SYS software (Lewis andWallace, 1998) set with Mehrbach solubility constants (Mehrbach etal., 1973) refitted by Dickson and Millero (1987).
During the experiments, the mussels were maintained accordingas suggested by Navarro et al. (2013). The seawater in each experi-mental container was replaced every day with seawater which hadpreviously been equilibrated in three large polyethylene header
2 C. Duarte et al. / Journal of Sea Research xxx (2013) xxx–xxx
Please cite this article as: Duarte, C., et al., Combined effects of temperature and ocean acidification on the juvenile individuals of the musselMytilus chilensis, Journal of Sea Research (2013), http://dx.doi.org/10.1016/j.seares.2013.06.002

tanks (250 L each one), following the methodology previouslyindicated.
2.3. Net shell deposition, shell dissolution and growth rate
Two experimental animals were randomly assigned to each repli-cate, being marked using bee-tags for individualization. In addition, ineach container, one previously weighted empty shell of M. chilensiswas placed. Empty shells were obtained by separating them from thesoft body of live mussels, similar in size to the experimental animals.These shells were cleaned with distilled water to eliminate any organicresidue and salt and then dried for 24 h (60 °C) before weighing. Theexperimental animals and empty shells were selected so that shelllengths, buoyant weights and shell weights did not show significantdifferences among the different CO2 treatments previous to theexperiment.
The net shell deposition rates of the organisms under the sixtreatments were estimated from changes in their buoyant weight(i.e., underwater weight), verified with the dry weight measure-ments of the shells after harvesting (Palmer, 1982). The measure-ments were made on day 0 (at the beginning of the experiment)and at the end of a 60-day period. Shell dissolution rates, in turn,were estimated by weighing in the air empty shells of M. chilensis,at the start and end of the experiment. Live animals and empty shellswere weighed to the nearest 0.0001 g. Growth rates were estimatedfrom changes in the total body weight (to the nearest 0.0001 g) ofthe mussels and expressed as percentage of increase. These mea-surements were made along with those for estimating shell deposi-tion and dissolution rate. For all measurements, each mussel wascarefully removed from the water, immediately weighed under thewater (buoyant weight), and then was gently blotted dry andweighed in the air (total weight).
2.4. Survival
The survival of M. chilensis under the experimental conditions wasevaluated at the end of the experimental period, however, nomortalitywas registered in any treatment.
2.5. Statistical analysis
To avoid pseudoreplication problems, the variables measured wereaveraged for the two mussels of each replicate. Two-way ANOVAwere used to evaluate whether temperature and CO2 levels and the in-teraction between factors affected net shell deposition and shell disso-lution rate, growth rate (increase in total weight with time) andsurvival (Zar, 1999).When the analysis showed significant interactions,a one-way ANOVA was carried out for each factor separately in eachlevel from the other factor, followed by Tukey's a posteriori HSD test.When the analysis did not show significant interactions, multiple com-parisons were carried out using Tukey's a posteriori HSD test on eachfactor that showed significant differences (Underwood, 1997). Netshell deposition and total weight increase were expressed in mg permussel per day, while shell dissolution was expressed in μg per musselper day. All analyses were carried out using Statistica version 7.0.Assumptions of normality and homoscedasticity of the one-wayANOVA were evaluated using the Kolmogorov–Smirnov and Burtletttests, respectively (Sokal and Rohlf, 1995).
3. Results
3.1. Water characteristics
The measured parameters of the seawater are shown in Table 1.Averaging all measurements, the mean (±SD) pH values for thedifferent CO2 treatments were: 8.057 (0.026) and 7.996 (0.026)
Fig. 1. Diagrammatic representation of the experimental design used to investigate the combine consequences of CO2 levels and temperature on Mytilus chilensis. The CO2 replicatetreatment (identified by the corresponding CO2 level) were assigned to one of the two temperature treatments (see Material and methods for further details).
Table 1Sea water characteristics (mean ± SD) used to maintain M. chilensis during the experimental period. See Material and methods section for details on the temperature and CO2
values used in the study. TA = total alkalinity; [CO32!] = carbonate ion concentration; !ca = omega calcite; !ar = omega aragonite.
CO2 system parameters Experimental treatments: temperature and CO2 concentrations
12 °C 16 °C 12 °C 16 °C 12 °C 16 °C
380 ppm (current) 700 (2050 year) 1000 (2100 year)
pH in situ (pH units) 8057 (0026) 7996 (0.026) 7835 (0.035) 7776 (0.034) 7673 (0.030) 7618 (0.030)Salinity (psu) 31.79 (1.88) 31.80(1.66) 31.81 (1.90) 31.83 (1.72) 31.87 (1.87) 31.88(1.66)TA (μmol kg!1) 2156.44 (86.24) 2148.63 (90.76) 2142.19 (93.23)pCO2 in situ (μatm) 371.35 (23.52) 439.54 (27.63) 656.91 (50.44) 772.64 (58.46) 977.56 (60.52) 1141.90 (69.19)[CO3
2!] in situ (μmol kg!1) 129.48 (14.65) 131.08 (14.74) 82.45 (11.24) 83.97 (11.32) 58.47 (7.92) 59.95 (8.00)!ca 3.14 (0.33) 319 (0.33) 1.99 (0.25) 2.04 (0.25) 1.42 (0.18) 1.46 (0.18)!ar 1.99 (0.22) 2.04 (0.22) 1.27 (0.17) 1.31 (0.17) 0.90 (0.12) 0.93 (0.12)
3C. Duarte et al. / Journal of Sea Research xxx (2013) xxx–xxx
Please cite this article as: Duarte, C., et al., Combined effects of temperature and ocean acidification on the juvenile individuals of the musselMytilus chilensis, Journal of Sea Research (2013), http://dx.doi.org/10.1016/j.seares.2013.06.002
(CO2 = 390 ppm and T° = 12 °C and 16 °C, respectively); 7.835(0.035) and 7.776 (0.034) (CO2 = 700 ppm and T° = 12 °C and16 °C, respectively); and 7.673 (0.030) and 7.618 (0.030) (CO2 =1000 ppm and T° = 12 °C and 16 °C, respectively). The seawaterwas undersaturated only with respect to aragonite in the treatmentCO2 = 1000 ppm (Table 1).
3.2. Net shell deposition and shell dissolution
The net shell deposition of mussels (estimated from changes in thebuoyant weight) in the different temperatures and CO2 treatments isshown in Fig. 2. The average net shell deposition rate over the studiedperiod was not affected by temperature; it was, however, negativelyaffected by CO2 concentration, so that the net shell deposition waslower in the highest CO2 concentration (i.e., 1000 ppm) comparedwith that registered in the control treatment (i.e., 390 ppm); while theshell deposition registered at intermediate CO2 concentration was notsignificantly different from the other CO2 treatments (two-wayANOVA; temperature: F1, 18 = 1.95, p > 0.05; CO2 concentration: F2,18 = 7.71, p b 0.05; temperature CO2 ! concentration: F2, 18 = 0.75,p > 0.05, Fig. 2).
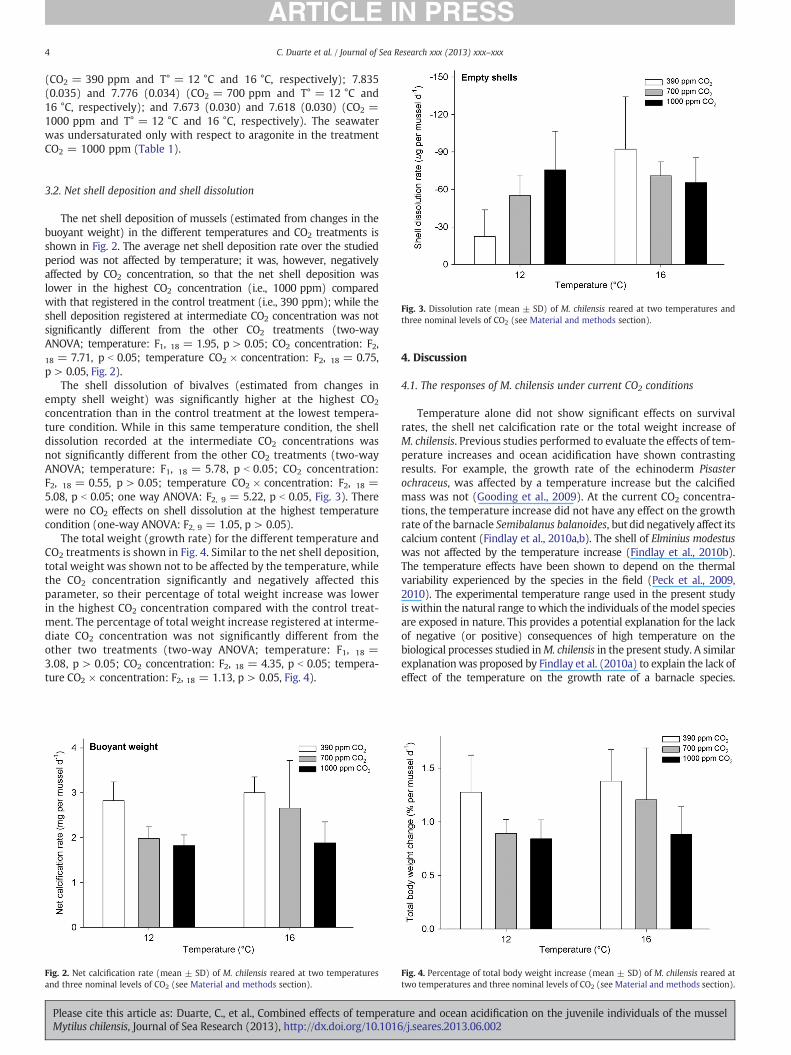
The shell dissolution of bivalves (estimated from changes inempty shell weight) was significantly higher at the highest CO2
concentration than in the control treatment at the lowest tempera-ture condition. While in this same temperature condition, the shelldissolution recorded at the intermediate CO2 concentrations wasnot significantly different from the other CO2 treatments (two-wayANOVA; temperature: F1, 18 = 5.78, p b 0.05; CO2 concentration:F2, 18 = 0.55, p > 0.05; temperature CO2 ! concentration: F2, 18 =5.08, p b 0.05; one way ANOVA: F2, 9 = 5.22, p b 0.05, Fig. 3). Therewere no CO2 effects on shell dissolution at the highest temperaturecondition (one-way ANOVA: F2, 9 = 1.05, p > 0.05).
The total weight (growth rate) for the different temperature andCO2 treatments is shown in Fig. 4. Similar to the net shell deposition,total weight was shown not to be affected by the temperature, whilethe CO2 concentration significantly and negatively affected thisparameter, so their percentage of total weight increase was lowerin the highest CO2 concentration compared with the control treat-ment. The percentage of total weight increase registered at interme-diate CO2 concentration was not significantly different from theother two treatments (two-way ANOVA; temperature: F1, 18 =3.08, p > 0.05; CO2 concentration: F2, 18 = 4.35, p b 0.05; tempera-ture CO2 ! concentration: F2, 18 = 1.13, p > 0.05, Fig. 4).
4. Discussion
4.1. The responses of M. chilensis under current CO2 conditions
Temperature alone did not show significant effects on survivalrates, the shell net calcification rate or the total weight increase ofM. chilensis. Previous studies performed to evaluate the effects of tem-perature increases and ocean acidification have shown contrastingresults. For example, the growth rate of the echinoderm Pisasterochraceus, was affected by a temperature increase but the calcifiedmass was not (Gooding et al., 2009). At the current CO2 concentra-tions, the temperature increase did not have any effect on the growthrate of the barnacle Semibalanus balanoides, but did negatively affect itscalcium content (Findlay et al., 2010a,b). The shell of Elminius modestuswas not affected by the temperature increase (Findlay et al., 2010b).The temperature effects have been shown to depend on the thermalvariability experienced by the species in the field (Peck et al., 2009,2010). The experimental temperature range used in the present studyis within the natural range to which the individuals of themodel speciesare exposed in nature. This provides a potential explanation for the lackof negative (or positive) consequences of high temperature on thebiological processes studied inM. chilensis in the present study. A similarexplanationwas proposed by Findlay et al. (2010a) to explain the lack ofeffect of the temperature on the growth rate of a barnacle species.
Fig. 2. Net calcification rate (mean ± SD) of M. chilensis reared at two temperaturesand three nominal levels of CO2 (see Material and methods section).
Fig. 3. Dissolution rate (mean ± SD) of M. chilensis reared at two temperatures andthree nominal levels of CO2 (see Material and methods section).
Fig. 4. Percentage of total body weight increase (mean ± SD) of M. chilensis reared attwo temperatures and three nominal levels of CO2 (see Material and methods section).
4 C. Duarte et al. / Journal of Sea Research xxx (2013) xxx–xxx
Please cite this article as: Duarte, C., et al., Combined effects of temperature and ocean acidification on the juvenile individuals of the musselMytilus chilensis, Journal of Sea Research (2013), http://dx.doi.org/10.1016/j.seares.2013.06.002

However, the highest average temperature used in the present study isjust recorded in a narrow temporal window during the austral summer(Navarro and Jaramillo, 1994). Therefore, given an unusual exposure ofindividuals of M. chilensis to high temperatures for more long periods(12 weeks), we would expect negative consequences on their biologicalprocesses as had been reported previously in a similar study (Martin andGattuso, 2009). Consequently, more data is required to find out thethermo-tolerance limit ofM. chilensis, whichwill help us to better under-stand the combined effects of global warming and ocean acidification onthis species. On the other hand, the temperature increase did negativelyaffect the shell degradation of M. chilensis. Contrasting results wereregistered in the juveniles of the shelled pteropod L. helicina, whereunder normocapnic conditions, temperature increase did not affectshell degradation (Lischka et al., 2011). We do not know themechanismthat explains this pattern, but it has been shown that temperature mayinfluence shell mineralogy significantly (Freitas et al., 2009).
4.2. Response of M. chilensis under acidic conditions
The results of this study showed that, only in the higher CO2 con-centrations, the responses of M. chilensis were negatively affected.This suggests that, within certain limits, this species can cope withsome CO2 increase.
The total net shell calcification of M. chilensis was significantly re-duced at 1000 ppm compared with those animals reared at 390 ppm.This coincides with studies carried out in other calcifying inverte-brates, including bivalves (Fabry et al., 2008; Gazeau et al., 2007,2010; Riebesell et al., 2000). Nevertheless, and as has been registeredpreviously in other species (e.g., Comeau et al., 2009; Gooding et al.,2009; Lischka et al., 2011), the M. chilensis individuals were able toincrease their shell mass at Ω b 1. The seawater in the highest CO2
levels was under-saturated with respect to aragonite, which could ex-plain the decrease in the net calcification rate shown by M. chilensis.However, previous studies on this species and other calcifyingorganisms have shown that net shell deposition may be affected athigh CO2 levels, even if seawater remains above the saturation state(e.g., Beniash et al., 2010; Ries et al., 2009; Talmage and Gobler,2009). For example, the net calcification rate of Mytilus edulis and theoyster Crassostrea gigas decreased when they were reared in seawaterenriched with CO2 but not undersaturated with respect to calciumcarbonate (Gazeau et al., 2007). This suggests that the carbonate avail-ability would not explain, or at least would not be the only factor toaccount for, the negative effects of CO2 increase on the net calcificationrate of this bivalve. The calcification problems registered in this speciescould therefore most likely be related to hypercapnia or acidosis, as hasbeen proposed in other studies (e.g., Arnold et al., 2009; Michaelidis etal., 2005). In mussels, for example, it has been shown that shell dis-solution may compensate acid–base disturbances (acidosis), affectingthe calcification process (Michaelidis et al., 2005). Consequently, futurestudies inM. chilensis are needed to evaluate specificallywhether or notthe calcification problems reported in the present study are associatedto acidosis.
There were no significant effects on the net calcification rate whenboth stressors were combined. Previous studies evaluating the effectsof both acidification and temperature on the net calcification rate arescarce and have shown contrasting results. For example, the tempera-ture increase diminished the negative effects of the CO2 increase onnet shell calcification of the foraminifera Neogloboquadrina pachyderma(Manno et al., 2012). Similar results have been recorded in studiescarried out in corals and echinoderms, where low increases in temper-ature have offset the negative effects of CO2 increase (Byrne, 2011;Kleypas and Yates, 2009; Sheppard-Brennand et al., 2010). On thecontrary, the shell content of S. balanoides was not affected by the CO2
increase alone, but decreased significantly when this occurred inconjunction with a temperature increase (Findlay et al., 2010b).The lack of similar interactive effects in M. chilensis could mean
that the temperature window used in this study was not enoughto detect effects in M. chilensis and consequently to detect negativeor synergic effects (see Gooding et al., 2009).
Shell deposition and shell dissolution have been shown to differ-entially affect the net carbonate production of animals (Nienhuis etal., 2010). In addition, previous studies have shown that an increasein the deposition rate to overcome shell dissolution can disguise theeffects of ocean acidification (e.g., Findlay et al., 2009; Lischka et al.,2011; McDonald et al., 2009; Wood et al., 2011). Consequently, it isrecommended that each of these processes be evaluated indepen-dently. In this study, the dissolution rate ofM. chilensiswas negative-ly affected by the CO2 increase, but this effect was counteracted bythe temperature increase. This is, at the higher temperature treat-ment, the rise in CO2 did not affect shell dissolution. Similar resultswere recorded by Lischka et al. (2011), who found that the negativeeffects of CO2 increase on the shell degradation of L. helicina werealso overcome by the temperature increase. However, in spite ofthe fact that the dissolution rate in M. chilensis was affected by theincrease in temperature and CO2, this process represented less than"8% of the net shell deposition of M. chilensis. This, therefore,suggests that the net calcification rate of this species was notexplained by the shell dissolution, but by the deposition process; aresult that does not agree with the findings of Nienhuis et al.(2010), namely that elevated CO2 concentration affected the shelldissolution rate but not the deposition rate in the intertidal snailNucella lamellose.
The growth rate of M. chilensis was negatively affected by the CO2
increase. However, these stressors did not show interactive effects onthe growth of this bivalve. Similar to the findings of this study, tem-perature and CO2 increase were not found to have antagonistic orsynergic effects on the growth rate of P. ochraceus (Gooding et al.,2009). No interactive effects of warming and acidification on thegrowth rate of S. balanoides were also registered (Findlay et al.,2010a). Opposite results were registered in the bivalve Mytilusgalloprovinciales, where the interaction between both stressorsresulted in a decrease in its growth rate (Berge et al., 2006). Althoughit has been proposed that low levels of temperature increase couldcounteract the negative effects of acidification (see Byrne, 2011 fora review), the results of the few studies carried out to date haveshown widely varying responses, both intra as interspecific and it istherefore not yet possible to make generalizations.
5. Conclusions
This study suggests that the juvenile M. chilensis individuals couldcope with some increase of temperature, but not with a CO2 increase.The absence of mortality in the individuals during the experimentindicated that the projected CO2 levels had chronic, but not lethaleffects on those animals. The interactive effects of temperature andCO2 affected only the shell dissolution, but this process was not im-portant to the animal's net calcification. The contrasting results ofthe scarce studies carried out until now, suggest that a substantialvariation in response to increments of CO2 and temperature occur.This illustrates the potential danger of generalizing from the resultsobtained in studies such us what is described here. In marine envi-ronments many species inhabits a wide geographic range, with con-trasting environmental conditions (e.g., temperature, salinity, pH).Therefore, future studies are needed to evaluate the responses ofthese species to different environmental stressors. In addition, futurestudies should consider individuals from different localities and thepotential for local adaptation to these global changes (see Sundayet al., 2011) to better understand their potential consequences.Finally, as mussels have been shown to influence their physical andbiological surroundings, the negative effects of CO2 increase couldhave significant ecological consequences, mainly in those habitatswhere this group is dominant in terms of abundance and biomass.
5C. Duarte et al. / Journal of Sea Research xxx (2013) xxx–xxx
Please cite this article as: Duarte, C., et al., Combined effects of temperature and ocean acidification on the juvenile individuals of the musselMytilus chilensis, Journal of Sea Research (2013), http://dx.doi.org/10.1016/j.seares.2013.06.002

Acknowledgments
The authors grateful to Jorge López, Barbara Cisternas, Maria ElisaJara and LoretoMardones for their valuable assistance during the exper-iments and for the algae production. Financial support provided bythe Programa de Investigación Asociativa, PIA-CONICYT-CHILE (GrantAnillos ACT-132) is gratefully acknowledged. This study was also,partially funded by FONDECYT Grant No. 11110407 (to CD). Duringthis research P.H.M. was under the tenure of FONDECYT Grant No.1090624. This is ANILLOS ACT-132 Publication No. 3.
References
Arnold, K.E., Findlay, H.S., Spicer, J.I., Daniels, C.L., Boothroyd, D., 2009. Effect of CO2-related acidification on aspects of the larval development of the European lobster,Homarus gammarus (L.). Biogeosciences Discussions 6, 3087–3107.
Beniash, E., Ivanina, A., Lieb, N.S., Kurochkin, I., Sokolova, I.M., 2010. Elevated level ofcarbon dioxide affects metabolism and shell formation in oysters Crassostreavirginica. Marine Ecology Progress Series 419, 95–108.
Berge, J.A., Bjerkeng, B., Pettersen, O., Schaanning, M.T., Oxnevad, S., 2006. Effects ofincreased sea water concentrations of CO2 on growth of the bivalve Mytilus edulisL. Chemosphere 62, 681–687.
Byrne, M., 2011. Impact of ocean warming and ocean acidification on marine inverte-brate life history stages: vulnerabilities and potential for persistence in a changingocean. Oceanography and Marine Biology: An Annual Review 49, 1–42.
Caldeira, K., Wickett, M.E., 2003. Anthropogenic carbon and ocean pH. Nature 425, 365.Comeau, S., Gorsky, G., Jeffree, R., Teyssié, J.L., Gattuso, J.P., 2009. Impact of ocean
acidification on a key Arctic pelagic mollusc (Limacina helicina). Biogeosciences 6,1877–1882.
Commito, J., Dankers, N., 2001. Dynamics of spatial and temporal complexity inEuropean and North American soft-bottom mussel beds. In: Reise, K. (Ed.),Ecological Comparisons of Sedimentary Shores. Ecological Studies, 151. Springer-Verlag, Berlin, Heidelberg, pp. 39–59.
Cummings, V., Hewitt, J., Van Rooyen, A., Currie, K., Beard, S., Thrush, S., Norkko, J., Barr,N., Heath, P., Halliday, N.J., Sedcole, R., Gomez, A., McGraw, C., Metcalf, V., 2011.Ocean acidification at high latitudes: potential effects on functioning of theAntarctic bivalve Laternula elliptica. PLoS One 6 (1), e16069.
Darling, E.S., Cote, I.M., 2008. Quantifying the evidence for ecological synergies. EcologyLetters 11, 1278–1286.
Dickson, A.G., Millero, F.J., 1987. A comparison of the equilibrium constants for the dis-sociation of carbonic acid in seawater media. Deep-Sea Research 34 (10A),1733–1743.
DOE, 1994. Handbook of methods for the analysis of the various parameters of thecarbon dioxide system in sea water; version 2. In: Dickson, A.G., Goyet, C. (Eds.),ORNL/CDIAC, 74.
Duarte, C., Jaramillo, E., Contreras, H., Figueroa, L., 2006. Community structure of themacroinfauna in the sediments below and intertidal mussel bed (Mytilus chilensis)(Hupe) of southern Chile. Revista Chilena de Historia Natural 79, 353–368.
Fabry, V.J., Seibel, B.A., Feely, R.A., Orr, J.C., 2008. Impacts of ocean acidification onmarine fauna and ecosystem processes. ICES Journal of Marine Science 65,414–432.
Feely, R.A., Sabine, C.L., Hernandez-Ayon, J.M., Ianson, D., Hales, B., 2008. Evidence forupwelling of corrosive “acidified” water onto the Continental Shelf. Science 320,1490.
Findlay, H.S., Kendall, M.A., Spicer, J.I., Widdicombe, S., 2009. Future high CO2 in the in-tertidal may compromise adult barnacle (Semibalanus balanoides) survival andembryo development rate. Marine Ecology Progress Series 398, 193–202.
Findlay, H.S., Kendall, M.A., Spicer, J.I., Widdicombe, S., 2010a. Relative influences ofocean acidification and temperature on intertidal barnacle post-larvae at thenorthern edge of their geographic distribution. Estuarine, Coastal and Shelf Science86, 675–682.
Findlay, H.S., Kendall, M.A., Spicer, J.I., Widdicombe, S., 2010b. Post-larval developmentof two intertidal barnacles at elevated CO2 and temperature. Marine Biology 157,725–735.
Freitas, P.S., Clarke, L.J., Kennedy, H., Richardson, C.A., 2009. Ion microprobe assessmentof the heterogeneity of Mg/Ca, Sr/Ca and Mn/Ca ratios in Pecten maximus andMytilus edulis (bivalvia) shell calcite precipitated at constant temperature.Biogeosciences Discussions 6, 1267–1316.
Gazeau, F., Quiblier, C., Jansen, J., Gattuso, J.P., Middleburg, J., Heip, C., 2007. Impact ofelevated CO2 on shellfish calcification. Geophysical Research Letters 34, 1–15.
Gazeau, F., Gattuso, J.-P., Dawber, C., Pronker, A.E., Peene, F., Peene, J., Heip, C.H.R.,Middelburg, J.J., 2010. Effect of ocean acidification on the early life stages of theblue mussel (Mytilus edulis). Biogeosciences Discussions 7, 2927–2947.
Gooding, R.A., Harley, C.D.G., Tang, E., 2009. Elevated water temperature and carbondioxide concentration increase the growth of a keystone echinoderm. Proceedingsof the National Academy of Sciences 106, 9316–9321.
Gutiérrez, J.L., Jones, C.G., Strayer, D.L., Iribane, O.O., 2003. Mollusk as ecosystemengineers: the role of shell production in aquatic habitats. Oikos 101, 79–90.
Gutowska, M.A., Pörtner, H.O., Melzner, F., 2008. Growth and calcification in thecephalopod Sepia officinalis under elevated seawater pCO2. Marine EcologyProgress Series 373, 303–309.
Haraldsson, C., Anderson, L.G., Hassellöv, M., Hulth, S., Olsson, K., 1997. Rapid, high-precision potentiometric titration of alkalinity in ocean and sediment pore waters.Deep-Sea Research Part I: Oceanographic Research Papers 44, 2031–2044.
Hay, G., Richardson, A.J., Robinson, C., 2005. Climate change and marine plankton.Trends in Ecology & Evolution 20, 338–344.
Hoegh-Guldberg, O., 2005. Low coral cover in a high-CO2 world. Journal of GeophysicalResearch. http://dx.doi.org/10.1029/2004JC002528.
Hofmann, G.E., Todgham, A.E., 2010. Living in the now: physiological mechanisms totolerate a rapidly changing environment. Annual Review of Physiology 72,127–145.
IPCC: Climate Change, 2007. The Physical Science Basis. Contribution of Working GroupI to the Fourth Assessment, Report of the Intergovernmental Panel on ClimateChange. In: Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B.,Tignor, M., Miller, H.L. (Eds.), Cambridge University Press, Cambridge, United Kingdomand New York, NY, USA, p. 996.
Kleypas, J.A., Yates, K.K., 2009. Coral reefs and ocean acidification. Oceanography 22,108–117.
Kleypas, J.A., Feely, R.A., Fabry, V.J., Langdon, C., Sabine, C.L., Robbins, L.L., 2006. Impactsof ocean acidification on coral reefs and other marine calcifiers: a guide for futureresearch. Report of a Workshop Held 18–20 April 2005, St. Petersburg, FL,Sponsored by NSF, NOAA, and the US Geological Survey.
Kurihara, H., 2008. Effects of CO2-driven ocean acidification on the early developmentalstages of invertebrates. Marine Ecology Progress Series 373, 275–284.
Langdon, C., Atkinson, M.J., 2005. Effect of elevated pCO2 on photosynthesis andcalcification of corals and interactions with seasonal change in temperature/irradiance and nutrient enrichment. Journal of Geophysical Research 110, C09S07.
Langdon, C., Takahashi, T., Sweeney, C., Chipman, D., Goddard, J., Marubini, F., Aceves,H., Barnett, H., Atkinson, M.J., 2000. Effect of calcium carbonate saturation stateon the calcification rate of an experimental reef. Global Biogeochemical Cycles14, 639–654.
Leclercq, N., Gattuso, J.P., Jaubert, J., 2000. CO2 partial pressure controls the calcificationrate of a coral community. Global Change Biology 6, 329–334.
Lewis, E., Wallace, D., 1998. Program Developed for CO2 System Calculations. CarbonDioxide Information Analysis Center. Oak Ridge, Tennessee, U.S.A.
Lischka, S., Büdenbender, J., Boxhammer, T., Riebesell, U., 2011. Impact of oceanacidification and elevated temperatures on early juveniles of the polar shelledpteropod Limacina helicina: mortality, shell degradation, and shell growth.Biogeosciences 8, 919–932.
Manno, C., Morata, N., Bellerby, R., 2012. Effect of ocean acidification and temperatureincrease on the planktonic foraminifer Neogloboquadrina pachyderma (sinistral).Polar Biology 35, 1311–1319.
Martin, S., Gattuso, J.P., 2009. Response of Mediterranean coralline algae to oceanacidification and elevated temperature. Global Change Biology 15, 2089–2100.
McDonald, M.R., McClintock, J.B., Amsler, C.D., Rittschof, D., Angus, R.A., Orihuela, B.,Lutostanski, K., 2009. Effects of ocean cidification over the life history of thebarnacle Amphibalanus amphitrite. Marine Ecology Progress Series 385, 179–187.
Mehrbach, C., Culberson, C.H., Hawley, J.E., Pytkowicz, R.N., 1973. Measurement of theapparent dissociation constants of carbonic acid in seawater at atmosphericpressure. Limnology and Oceanography 18, 897–907.
Meinshausen, M., Smith, S.J., Calvin, K., Daniel, J.S., Kainuma, M.L.T., Lamarque, J.-F.,Matsumoto, K., Montzka, S.A., Raper, S.C.B., Riahi, K., Thomson, A., Velders, G.J.M.,van Vuuren, D.P.P., 2011. The RCP greenhouse gas concentrations and theirextensions from 1765 to 2300. Climatic Change 109, 213–241.
Michaelidis, B., Ouzounis, C., Paleras, A., Pörtner, H.O., 2005. Effects of long-term mod-erate hypercapnia on acid–base balance and growth rate in marine musselsMytilusgalloprovincialis. Marine Ecology Progress Series 293, 109–118.
Mieszkowska, N., Kendall, M.A., Hawkins, S.J., Leaper, R., Williamson, P., Hardman-Mountford, N.J., Southward, A.J., 2006. Changes in the range of some commonrocky shore species in Britain — a response to climate change? Hydrobiologia555, 241–251.
Miller, A.W., Reynolds, A.C., Sobrino, C., Riedel, G.F., 2009. Shellfish face uncertainfuture in high CO2 world: influence of acidification on oyster larvae calcificationand growth in estuaries. PLoS One 4 (5), e5661. http://dx.doi.org/10.1371/journal.pone.0005661.
Navarro, J.M., Jaramillo, R., 1994. Evaluación de la oferta alimentaria natural disponiblea organismos filtrados de la bahía de Yaldad (43-08;73-44′), sur de Chile. Revistade Biologia Marina 29 (1), 57–75.
Navarro, J.M., Torres, R., Acuña, K., Duarte, C., Manríquez, P.H., Lardies, M., Lagos, N.A.,Vargas, C., Aguilera, V., 2013. Impact of medium-term exposure to elevated pCO2levels on the physiological energetics of the mussel Mytilus chilensis. Chemosphere90, 1242–1248.
Nienhuis, S., Palmer, A.R., Harley, C.D.G., 2010. Elevated CO2 affects shell dissolutionrate but not calcification rate in a marine snail. Proceedings of the Royal SocietyB: Biological Sciences 277, 2553–2558.
Orr, J.C., Fabry, V.J., Aumont, O., Bopp, L., Doney, S.C., Feely, R.A., Gnanadesikan, A.,Gruber, N., Ishida, A., Joos, F., Key, R.M., Lindsay, K., Maier-Reimer, E., Matear, R.,Monfray, P., Mouchet, A., Najjar, R.G., Plattner, G.-K., Rodgers, K.B., Sabine, C.L.,Sarmiento, J.L., Schlitzer, R., Slater, R.D., Totterdell, I.J., Weirig, M.-F., Yamanaka,Y., Yool, A., 2005. Anthropogenic ocean acidification over the twenty-first centuryand its impact on calcifying organisms. Nature 437, 681–686.
Palmer, A.R., 1982. Growth in marine gastropods: a non-destructive technique forindependently measuring shell and body weight. Malacologia 23, 63–73.
Parker, M., Ross, P.M., O'Connor, W.A., 2009. The effect of ocean acidification andtemperature on the fertilization and embryonic development of the Sydneyrock oyster Saccostrea glomerata (Gould 1850). Global Change Biology 15,2123–2136.
6 C. Duarte et al. / Journal of Sea Research xxx (2013) xxx–xxx
Please cite this article as: Duarte, C., et al., Combined effects of temperature and ocean acidification on the juvenile individuals of the musselMytilus chilensis, Journal of Sea Research (2013), http://dx.doi.org/10.1016/j.seares.2013.06.002

Parker, L.M., Ross, P.M., O'Connor, W.A., 2011. Populations of the Sydney rock oyster,Saccostrea glomerata, vary in response to ocean acidification. Marine Biology.http://dx.doi.org/10.1007/s00227-010-1592-4.
Peck, L.S., Webb, K.E., Bailey, D.M., 2004. Extreme sensitivity of biological function totemperature in Antarctic marine species. Functional Ecology 18, 625–630.
Peck, L.S., Massey, A., Thorne, M.A., Clark, M.S., 2009. Lack of acclimation in Ophionotusvictoriae: brittle stars are not fish. Polar Biology 32, 399–402.
Peck, L.S., Morley, S.A., Clark, M.S., 2010. Poor acclimation capacities in Antarcticmarine ectotherms. Marine Biology 157, 2051–2059.
Pörtner, H.O., 2010. Oxygen- and capacity-limitation of thermal tolerance: a matrix forintegrating climate-related stressor effects in marine ecosystems. Journal ofExperimental Biology 213, 881–893.
Pörtner, H.O., Knust, R., 2007. Climate change affects marine fishes through the oxygenlimitation of thermal tolerance. Science 315, 95–97.
Riebesell, U., Zondervan, I., Rost, B., Tortell, P.D., Zeebe, R.E., Morel, F.M.M., 2000.Reduced calcification of marine plankton in response to increased atmosphericCO2. Nature 407, 364–367.
Ries, J.B., Cohen, A.L., McCorkle, D.C., 2009. Marine calcifiers exhibit mixed responses toCO2-induced ocean acidification. Geology 37 (12), 1131–1134.
Rosa, R., Seibel, B.A., 2008. Synergistic effects of climate-related variables suggestfuture physiological impairment in a top oceanic predator. Proceedings of theNational Academy of Sciences 105, 20776–20780.
Schiedek, D., Sundelin, B., Readman, J.W., McDonald, R.W., 2007. Interactions betweenclimate change and contaminants (review). Marine Pollution Bulletin 54,1845–1856.
Sheppard-Brennand, H., Soars, N., Dworjanyn, S.A., Davis, A.R., Byrne, M., 2010. Impactof ocean warming and ocean acidification on larval development and calcificationin the sea urchin Tripneustes gratilla. PLoS One 5, e11372.
Sokal, R.R., Rohlf, F.J., 1995. Biometry: The Principles and Practice of Statistics inBiological Research. Freeman, W.H., New York.
Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller,H.L., 2007. Climate change 2007: the physical science basis. Contribution ofWorking Group I to the Fourth Assessment Report of the IntergovernmentalPanel on Climate Change. Cambridge University Press, Cambridge, United Kingdomand New York, NY, USA.
Spicer, J.I., Raffo, A., Widdicombe, S., 2007. Influence of CO2-related seawater acidifica-tion on extracellular acid–base balance in the velvet swimming crab Necora puber.Marine Biology 151, 1117–1125.
Strickland, J.D.H., Parsons, T.R., 1968. A practical handbook of seawater analysis.Fisheries Research Board of Canada, Bulletin 167, 311.
Sunday, J.M., Crim, R.N., Harley, C.D.G., Hart, M.W., 2011. Quantifying rates of evolu-tionary adaptation in response to ocean acidification. PLoS ONE 6, e22881.
Talmage, S.C., Gobler, C.J., 2009. The effects of elevated carbon dioxide concentrationson the metamorphosis, size and survivial of larval hard clams (Mercenariamercenaria), bay scallops (Argopecten irradians), and Eastern oysters (Crassostreavirginica). Limnology and Oceanography 54, 2072–2080.
Thomas, C.D., Cameron, A., Green, R.E., Bakkenes, M., Beaumont, L.J., Collingham, Y.C.,Erasmus, B.F.N., de Siqueira, M.F., Grainger, A., Hannah, L., Hughes, L., Huntley, B.,van Jaarsveld, A.S., Midgley, G.F., Miles, L., Ortega-Huerta, M.A., Peterson, A.T.,Phillips, O.L., Williams, S.E., 2004. Extinction risk from climate change. Nature427, 145–148.
Underwood, A.J., 1997. Experiments in Ecology: Their Logical Design and InterpretationUsing Analysis of Variance. Cambridge University Press, Cambridge.
Widdicombe, S., Spicer, J.L., 2008. Predicting the impact of ocean acidification onbenthic biodiversity: what can animal physiology tell us? Journal of ExperimentalMarine Biology and Ecology 366, 187–197.
Wolf-Gladrow, D.A., Riebesell, U., Burkhardt, S., Bijma, J., 1999. Direct effects of CO2
concentration on growth and isotopic composition of marine plankton. Tellus B51, 461–476.
Wood, H.L., Spicer, J.I., Lowe, D.M., Widdicombe, S., 2010. Interaction of ocean acidifica-tion and temperature; the high cost of survival in the brittlestar Ophiura ophiura.Marine Biology 157, 2001–2013.
Wood, H.L., Spicer, J.I., Kendall, M.A., Lowe, D.M., Widdicombe, S., 2011. Ocean warmingand acidification; implications for the Arctic brittlestar Ophiocten sericeum. PolarBiology 34, 1033–1044.
Zar, J.H., 1999. Biostatistical Analysis, 4th ed. Prentice Hall, Upper Saddle River.
7C. Duarte et al. / Journal of Sea Research xxx (2013) xxx–xxx
Please cite this article as: Duarte, C., et al., Combined effects of temperature and ocean acidification on the juvenile individuals of the musselMytilus chilensis, Journal of Sea Research (2013), http://dx.doi.org/10.1016/j.seares.2013.06.002
Copyright © 2022 FDOKUMEN




















