
Innervation of the rat trachea by bilateral cholinergic projections from the nucleus ambiguus and...
11
Research report Innervation of the rat trachea by bilateral cholinergic projections from the nucleus ambiguus and direct motor fibers from the cervical spinal cord: a retrograde and anterograde tracer study Yasuro Atoji a, * , Dwi Liliek Kusindarta a,b , Nao Hamazaki a , Akihisa Kaneko a a Laboratory of Veterinary Anatomy, Faculty of Applied Biological Sciences, Gifu University, Yanagido 1-1, Gifu 501-1193, Japan b Department of Anatomy, Faculty of Veterinary Medicine, Gadjah Mada University, Yogyakarta 55281, Indonesia Accepted 19 October 2004 Available online 2 December 2004 Abstract A tract–tracer method was employed to examine the innervation of the rat trachea. Cholera toxin h subunit (CTB) was injected into the following locations in separate groups of rats: (1) ventral trachea, (2) lateral trachea, (3) ventral trachea after the excision of the nodose ganglion, and (4) ventral trachea after the transection of C1–C2 spinal nerves. CTB injection in the ventral trachea showed bilateral labeling of neurons in the nucleus ambiguus (NA), medial subnucleus of the nucleus of the solitary nucleus, dorsal motor nucleus of the vagus (DMV), and lamina IX of C1–C6. CTB injection in the lateral trachea showed significant ipsilateral predominance of neuronal labeling in the NA and lamina IX of C1–C2 segments. CTB injection in rats after the excision of the nodose ganglion revealed no labeling in the ipsilateral DMV and NA and a significant reduction of neuronal labeling in C1. CTB injection in rats after the transection of C1–C2 spinal nerves showed a significant decrease in the number of labeled neurons in ipsilateral NA, C1, and C2 and no labeling of fibers in C1–C2. The combination of retrograde fluorogold labeling and choline acetyltransferase (ChAT) immunostaining revealed that all fluorogold-labeled neurons in the NA and lamina IX of C1–C2 colocalized with ChAT. The injection of biotinylated dextran amine in NA produced labeling in axonal terminals on postganglionic neurons, but not in other regions of the trachea. Our findings indicate that the rat trachea is innervated bilaterally by cholinergic motor neurons in NA and C1–C2, while those traveling through the spinal nerves project directly to the trachea. D 2004 Elsevier B.V. All rights reserved. Theme: Motor systems and sensorimotor integration Topic: Spinal cord and brainstem Keywords: Airway; Nerve transection; Sensory innervation; Cholera toxin; Dextran amine 1. Introduction Autonomic innervation of the airway includes the para- sympathetic and sympathetic nervous systems, which function to control smooth muscle tension, secretion of mucosal glands, vascular tone, and tracheobronchial reflexes [3,7]. The preganglionic parasympathetic neurons innervating the trachea originate from the dorsal motor nucleus of the vagus (DMV) and nucleus ambiguus (NA) [5,8]. Postganglionic parasympathetic neurons are located in the peritracheal ganglion or submucosal plexus [1,9,11,14]. Sensory fibers that are distributed in the tracheal epithelium project to the nucleus of the solitary tract (NTS) through neurons located in the nodose or jugular ganglion. The innervation pattern of vagal motor neurons to the lung shows species differences: NA in the cat innervates the lung contralaterally [8], whereas in the rat, it innervates the lung bilaterally [12]. The contralateral pathway in bilateral 0006-8993/$ - see front matter D 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.brainres.2004.10.032 * Corresponding author. Tel.: +81 58 293 2936; fax: +81 58 293 2840. E-mail address: [email protected] (Y. Atoji). Brain Research 1031 (2005) 90 – 100 www.elsevier.com/locate/brainres
Transcript of Innervation of the rat trachea by bilateral cholinergic projections from the nucleus ambiguus and...

www.elsevier.com/locate/brainres
Brain Research 1031
Research report
Innervation of the rat trachea by bilateral cholinergic projections from the
nucleus ambiguus and direct motor fibers from the cervical spinal cord:
a retrograde and anterograde tracer study
Yasuro Atojia,*, Dwi Liliek Kusindartaa,b, Nao Hamazakia, Akihisa Kanekoa
aLaboratory of Veterinary Anatomy, Faculty of Applied Biological Sciences, Gifu University, Yanagido 1-1, Gifu 501-1193, JapanbDepartment of Anatomy, Faculty of Veterinary Medicine, Gadjah Mada University, Yogyakarta 55281, Indonesia
Accepted 19 October 2004
Available online 2 December 2004
Abstract
A tract–tracer method was employed to examine the innervation of the rat trachea. Cholera toxin h subunit (CTB) was injected into the
following locations in separate groups of rats: (1) ventral trachea, (2) lateral trachea, (3) ventral trachea after the excision of the nodose
ganglion, and (4) ventral trachea after the transection of C1–C2 spinal nerves. CTB injection in the ventral trachea showed bilateral
labeling of neurons in the nucleus ambiguus (NA), medial subnucleus of the nucleus of the solitary nucleus, dorsal motor nucleus of the
vagus (DMV), and lamina IX of C1–C6. CTB injection in the lateral trachea showed significant ipsilateral predominance of neuronal
labeling in the NA and lamina IX of C1–C2 segments. CTB injection in rats after the excision of the nodose ganglion revealed no
labeling in the ipsilateral DMV and NA and a significant reduction of neuronal labeling in C1. CTB injection in rats after the transection
of C1–C2 spinal nerves showed a significant decrease in the number of labeled neurons in ipsilateral NA, C1, and C2 and no labeling of
fibers in C1–C2. The combination of retrograde fluorogold labeling and choline acetyltransferase (ChAT) immunostaining revealed that all
fluorogold-labeled neurons in the NA and lamina IX of C1–C2 colocalized with ChAT. The injection of biotinylated dextran amine in NA
produced labeling in axonal terminals on postganglionic neurons, but not in other regions of the trachea. Our findings indicate that the rat
trachea is innervated bilaterally by cholinergic motor neurons in NA and C1–C2, while those traveling through the spinal nerves project
directly to the trachea.
D 2004 Elsevier B.V. All rights reserved.
Theme: Motor systems and sensorimotor integration
Topic: Spinal cord and brainstem
Keywords: Airway; Nerve transection; Sensory innervation; Cholera toxin; Dextran amine
1. Introduction
Autonomic innervation of the airway includes the para-
sympathetic and sympathetic nervous systems, which
function to control smooth muscle tension, secretion of
mucosal glands, vascular tone, and tracheobronchial
reflexes [3,7]. The preganglionic parasympathetic neurons
0006-8993/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.brainres.2004.10.032
* Corresponding author. Tel.: +81 58 293 2936; fax: +81 58 293 2840.
E-mail address: [email protected] (Y. Atoji).
innervating the trachea originate from the dorsal motor
nucleus of the vagus (DMV) and nucleus ambiguus (NA)
[5,8]. Postganglionic parasympathetic neurons are located in
the peritracheal ganglion or submucosal plexus [1,9,11,14].
Sensory fibers that are distributed in the tracheal epithelium
project to the nucleus of the solitary tract (NTS) through
neurons located in the nodose or jugular ganglion.
The innervation pattern of vagal motor neurons to the
lung shows species differences: NA in the cat innervates the
lung contralaterally [8], whereas in the rat, it innervates the
lung bilaterally [12]. The contralateral pathway in bilateral
(2005) 90–100

Y. Atoji et al. / Brain Research 1031 (2005) 90–100 91
innervation undergoes decussation inside the thorax in the
rat [12]. In the trachea, however, the issue of whether the
trachea is innervated bilaterally or contralaterally remains to
be resolved, although both DMV and NA participate in
tracheal innervation [5].
A tract-tracing study using cholera toxin h subunit
(CTB) showed that neurons in the dorsomedial part of the
ventral horn in C1 and C2 participate in the innervation of
the trachea in the dog, ferret, and rat, in addition to NA and
DMV [5]. Furthermore, the authors found CTB-labeled
fibers in the ventral funiculus of the upper cervical segments
in the dog and in the cervical dorsal horn in the ferret. The
study suggested that the upper cervical spinal cord
participates partially in the control of the trachea.
Sensory or parasympathetic nerve fibers are distributed
in the epithelium or lamina submucosa of the trachea of
the ferret and rat [4,9,13]. Perez-Fontan and Velloff [11]
reported that the injection of CTB in the tracheal lumen
resulted in labeling neurons in NA and DMV by trans-
epithelial transport to intra- or subepithelial space, where
labeled vagal motor neurons project their axons in the rat.
The authors hypothesized that vagal motor neurons in
DMV and NA project their axons directly to the
epithelium or submucosa without interposition of intrinsic
neurons, which are located in the peritracheal para-
sympathetic ganglion. This hypothesis is contradictory to
the classic concept of the airway parasympathetic trans-
mission; preganglionic vagal neurons synapse on post-
ganglionic neurons in the local ganglion, and, in turn, the
latter exert their effects on smooth muscles, glandular
cells, and epithelial cells [7,10]. However, their experiment
was carried out under the presence of the classic
parasympathetic rely [11]. Therefore, the hypothesis of a
direct vagal motor innervation to the trachea remains to be
confirmed.
The aims of the present study were to determine whether
NA and DMV innervate trachea bilaterally and whether the
cervical motor neurons directly innervate the trachea.
Experiments involved tract tracing using CTB and denerva-
tion of the vagus nerve and spinal nerves. In the latter
experiment, we chose the cervical spinal cord, where
autonomic innervation of both sympathetic and parasympa-
thetic nerves is absent [2]. CTB was injected in the tracheal
wall because previous studies reported that retrograde-
labeled neurons were more numerous in NA and DMV
than those obtained by injection into the tracheal lumen
[5,11].
2. Materials and methods
A total of 52 Wistar rats (weight 250–300 g, 10–16
weeks, of both sexes) was used in the present study. The
experimental protocol was approved by the Ethics Review
Committee for Animal Experimentation of Faculty of
Agriculture, Gifu University.
2.1. Injection of retrograde tracers in the trachea
Experiments consisted of five groups to clarify bilateral
innervation of the medulla to the trachea and innervation of
motor neurons in the cervical spinal cord to the trachea. Rats
were anesthetized by intraperitoneal injection of sodium
pentobarbital (25 mg/kg body weight). Under sterile
conditions, the cervical trachea was exposed. In rats of
group 1 (n=5), 0.5% CTB (low salt, List Biological
Laboratories, USA) in phosphate-buffered saline (PBS)
was injected in the ventral midline of the trachea in the
5th to 14th intercartilagenous spaces, using a glass micro-
pipette (a total of 10 injections of 400 nl each, total
volume=4 Al). In rats of group 2 (n=5), the injection
protocol was similar to that used for group 1, except that the
injections were made in the left or right side of the trachea.
In rats of group 3 (n=5), the left or right nodose ganglion
was excised, followed 7–10 days later by tracheal injections
under anesthesia of the same toxin solution used for group
1, at the same dose and in the same anatomical site. In rats
of group 4 (n=5), 2 mm of the right C1 and C2 spinal
nerves, just lateral to the intervertebrate foramen, was cut,
followed 7–10 days later by tracheal injections under
anesthesia of the same toxin solution used for group 1, at
the same dose and in the same anatomical site. In rats of
group 5, 0.5% fluorogold (Biotum, USA) in distilled water
was injected in the ventral midline of the trachea in the 5th
to 14th intercartilagenous spaces using a glass micropipette
(a total of 10 injections of 400 nl each, total volume=4 Al) todetect colocalization with choline acetyltransferase (ChAT),
similar to group 1.
Three controls of injections were done to examine
whether tracers injected in the trachea were leaked into
neighboring tissues, i.e., the m. sternohyoideus, which
covers the ventral surface of the trachea. In rats of control
1 (n=3), after the m. sternohyoideus of both sides were
removed between near origin and insertion, CTB was
injected in the ventral midline of the trachea, in the 5th to
14th intercartilagenous spaces (a total of 10 injections of
400 nl each, total volume=4 Al). In rats of control 2 (n=3),
CTB was injected in the m. sternohyoideus of both sides
(each side received a total of 10 injections of 400 nl each,
total volume=4 Al). In rats of control 3 (n=3), fluorogold
was injected in the ventral midline of the trachea in the 5th
to 14th intercartilagenous spaces (a total of 10 injections
of 400 nl each, total volume=4 Al), while CTB was
injected in the m. sternohyoideus of both sides (each side
received a total of 10 injections of 400 nl each, total
volume=4 Al).On the 4th day after the injection of CTB or fluorogold,
all rats were anesthetized with sodium pentobarbital (50 mg/
kg, intraperitoneal injection), perfused with Ringer’s sol-
ution, followed by 4% formaldehyde in 0.1 M phosphate
buffer at pH 7.4. The brainstem and cervical spinal cord
were dissected out and postfixed in the same fixative for 2–3
days. Specimens were transferred to 30% sucrose in PBS at
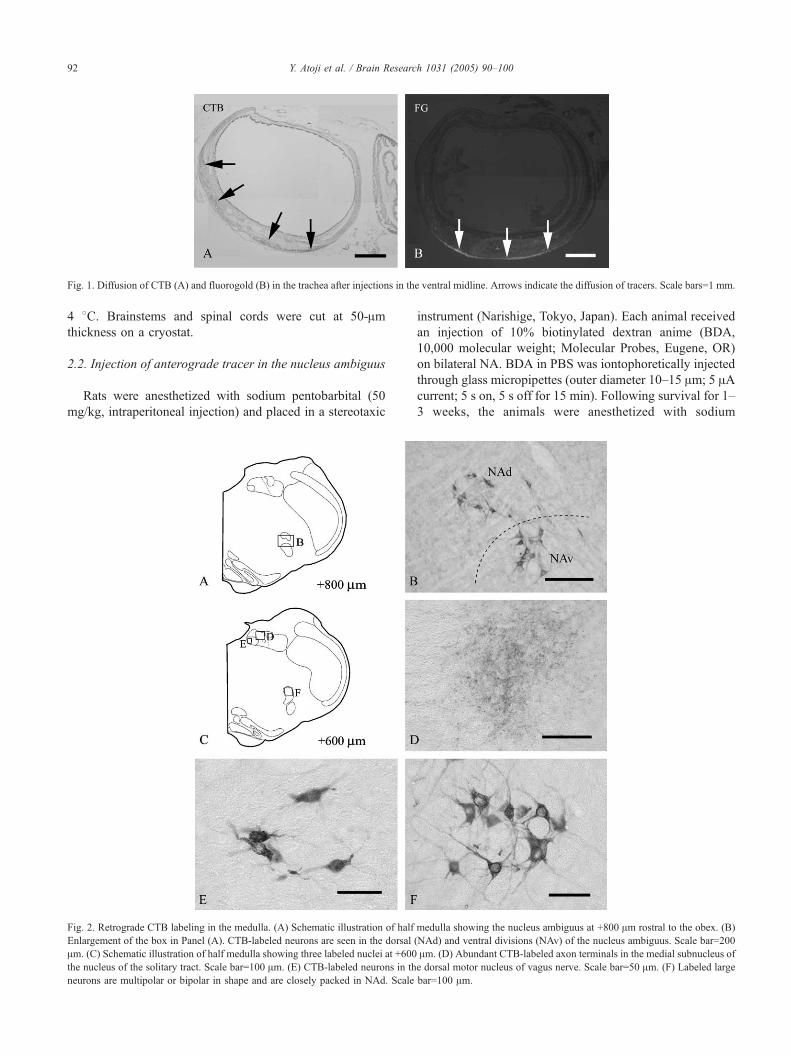
Fig. 1. Diffusion of CTB (A) and fluorogold (B) in the trachea after injections in the ventral midline. Arrows indicate the diffusion of tracers. Scale bars=1 mm.
Y. Atoji et al. / Brain Research 1031 (2005) 90–10092
4 8C. Brainstems and spinal cords were cut at 50-Amthickness on a cryostat.
2.2. Injection of anterograde tracer in the nucleus ambiguus
Rats were anesthetized with sodium pentobarbital (50
mg/kg, intraperitoneal injection) and placed in a stereotaxic
Fig. 2. Retrograde CTB labeling in the medulla. (A) Schematic illustration of half
Enlargement of the box in Panel (A). CTB-labeled neurons are seen in the dorsal
Am. (C) Schematic illustration of half medulla showing three labeled nuclei at +600
the nucleus of the solitary tract. Scale bar=100 Am. (E) CTB-labeled neurons in th
neurons are multipolar or bipolar in shape and are closely packed in NAd. Scale
instrument (Narishige, Tokyo, Japan). Each animal received
an injection of 10% biotinylated dextran anime (BDA,
10,000 molecular weight; Molecular Probes, Eugene, OR)
on bilateral NA. BDA in PBS was iontophoretically injected
through glass micropipettes (outer diameter 10–15 Am; 5 AAcurrent; 5 s on, 5 s off for 15 min). Following survival for 1–
3 weeks, the animals were anesthetized with sodium
medulla showing the nucleus ambiguus at +800 Am rostral to the obex. (B)
(NAd) and ventral divisions (NAv) of the nucleus ambiguus. Scale bar=200
Am. (D) Abundant CTB-labeled axon terminals in the medial subnucleus of
e dorsal motor nucleus of vagus nerve. Scale bar=50 Am. (F) Labeled large
bar=100 Am.

Y. Atoji et al. / Brain Research 1031 (2005) 90–100 93
pentobarbital (50 mg/kg) and perfused with Ringer’s
solution, followed by 4% paraformaldehyde in 0.1 M
phosphate buffer at pH 7.4. Brains and tracheas were
removed, postfixed in the same fixative for 2–3 days, and
transferred to 30% sucrose in PBS at 4 8C. The medulla was
transversely sectioned at 50 Am on a cryostat, whereas the
trachea was sectioned at 50 Am, and the sections were
mounted on glass slides.
2.3. Immunohistochemical staining
Sections of rats of groups 1–4 were stained with an
antibody against CTB. Sections of the medulla and cervical
spinal cord were rinsed in PBS and were treated with 50%
methanol containing 0.3% H2O2 for 30 min. After rinsing in
PBS, they were incubated with 1% nonimmune rabbit serum
for 1 h. The specimens were then incubated with a solution
containing goat anti-CTB antibody (1:60,000; List Bio-
logical Laboratories), 0.3% Triton X-100, and 1% rabbit
serum for 2 days at 4 8C, followed by incubation with
biotinylated rabbit antigoat IgG (1:500; Sigma, St. Louis,
Fig. 3. Histograms showing the distribution of labeled neurons in the nucleus ambi
solitary tract (NTS), and dorsal motor nucleus of the vagus nerve (DMV) after
continuity of two regions of NA (+1800 to �1800 Am) and cervical spinal cord (C
and ventral divisions. (B) Histogram shows continuity of two nuclei of NTS (+1800
of labeled neurons in four consecutive sections (n=5). The numbers of labeled ne
labeled neurons was enumerated in all sections (40–50 sections) of each segmen
(n=5). Data are meanFS.D. SC: spinal cord.
MO) for 1 h at room temperature, and finally incubated in
avidin–biotin–peroxidase complex (ABC, Elite ABC kit;
Vector Laboratories, Burlingame, CA) for 1 h at room
temperature. ABC was visualized by incubation in 0.05 M
Tris–HCl buffer (pH 7.4) containing 3, 3V-diaminobenzidine
tetrahydrochloride (DAB) and H2O2. Sections were
mounted on glass slides precoated with gelatin and chrome
alum, dehydrated in a series of ethanol solutions, cleared in
xylene, and coverslipped.
Sections of the brainstem and spinal cord of group 5 were
stained with goat anti-ChAT. Sections were first treated with
50% methanol containing 0.3% H2O2 for 30 min. Then,
they were incubated with 1% nonimmune donkey serum for
30 min. After washing in PBS, sections were incubated with
goat anti-ChAT antibody (1:400; Chemicon International,
Temecula, CA) overnight at 4 8C. The sections were
incubated with antigoat IgG conjugated with tetramethylr-
hodamine isothiocyanate (TRITC, 1:100; Jackson Immu-
noresearch Laboratories, West Grove, PA) for 2 h at room
temperature in the dark. After washing in PBS, the sections
were mounted on glass slides, coverslipped with carbonate-
guus (NA), cervical spinal cord, the medial subnucleus of the nucleus of the
CTB injection in the ventral midline of the trachea. (A) Histogram shows
1 to C6). The number of labeled neurons in NA indicates the sum of dorsal
to +1000 Am) and DMV (+800 to �2000 Am). Each bar expresses the total
urons in C1–C6 bars were calculated as follows: First, the total number of
t, divided as equivalent to four sections, and mean values were calculated

Y. Atoji et al. / Brain Research 1031 (2005) 90–10094
buffered glycerol, and examined with a fluorescence
microscope.
Sections of the medulla and trachea in the BDA-injected
rats were treated with 50% methanol containing 0.3% H2O2
for 30 min. After washing in PBS, they were incubated with
ABC (Vector Laboratories) in PBS containing 0.3% Triton
X-100 for 2 h at room temperature, followed by incubation
in 0.05 M Tris–HCl buffer (pH 7.4) containing DAB and
H2O2 in 0.1 M Tris–HCl buffer at pH 7.4. Tracheal sections
mounted on glass slides and floating sections were stained
as described above.
2.4. Data analysis
The numbers of labeled neurons in the medulla and
cervical spinal cord of groups 1–4 (n=5) were counted.
Differences between groups were examined for statistical
significance using Student’s t-test. Statistical significance
was established as Pb0.05 or 0.01, and all data were
expressed as meanFS.D.
3. Results
3.1. Spread of tracers in the trachea
In cases of CTB injections, the tracer was not found in
the trachea 4 days after the injections. Therefore, the
diffusion of the tracer was examined in 30 min and 2, 24,
and 48 h after the injections in the ventral midline of the
trachea. Thirty minutes after injection, CTB was seen in the
Fig. 4. Retrograde CTB labeling in the cervical spinal cord. (A) Schematic ill
Enlargement of the box in (A). Abundant labeled axons or terminals in the central c
nucleus. Scale bar=50 Am. (D) Many labeled neurons in the medial part of lamin
ventral part of the trachea (Fig. 1A). Two and 24 h after
injections, CTB extended into the ventral half of the trachea,
but 48 h after the injections, it was not yet detected in the
trachea. In cases of fluorogold injections, the tracer could be
still seen in the trachea 4 days after injections. In the midline
injection, fluorogold was found mainly in the ventral part
(Fig. 1B), while in the lateral injection, the tracer was
restricted to the ipsilateral side.
3.2. Injection of CTB and fluorogold
The injection of CTB in the ventral midline of the
tracheal wall (group 1) showed labeling neurons in the
dorsal (NAd) and ventral (NAv) divisions of the nucleus
ambiguus, DMV, and rostral part of the medial subnucleus
of NTS in the medulla oblongata (Fig. 2A–C, E, and F). The
number of labeled neurons was largest in NA among the
three nuclei (Fig. 3A and B). Labeled neurons in NAd were
largely multipolar and extended rostrocaudally from +1.8
(the obex is 0 mm) to �0.4 mm, whereas multipolar labeled
neurons in NAv were found at the levels of +1.4 to +0.8
mm. The densest of labeled neurons in the whole NA was
found in +1.20 mm rostral to the obex (Fig. 3A). Labeled
neurons were found in the rostral part of the medial
subnucleus of NTS, at +1.8 to +1.0 mm. Round or bipolar
small labeled neurons were found in the DMV, which was
localized in +0.8 to �2.0 mm. Labeled fibers in the NTS
were seen in the medial, ventrolateral, and commissural
subnuclei at +1.6 to �2.2 mm (Fig. 2D). The densest
labeled fibers in the NTS were localized in the medial
subnucleus at levels of +0.4 to �0.4 mm. In the cervical
ustration of half spinal cord showing the orientation of B–D at C1. (B)
ervical nucleus. Scale bar=100 Am. (C) A labeled neuron in the intermediate
a IX. Scale bar=100 Am. CC: central canal.

Fig. 5. Histograms showing the numbers of labeled neurons. (A) Number of labeled neurons when CTB was injected in the left lateral side of the trachea. (B)
Number of labeled neurons when CTB was injected after the excision of the nodose ganglion. (C) Number of labeled neurons when CTB was injected after
transection of C1–C2 spinal nerves. Each bar in the dorsal motor nucleus of the vagus nerve (DMV), nucleus ambiguus (NA), and C1–C6 expresses the total
number of labeled neurons in the nuclei and segments (n=5). Statistical analysis was not performed from C3 to C6 because of the low number of labeled
neurons. Data are meanFS.D. **Pb0.01, *Pb0.05.
Y. Atoji et al. / Brain Research 1031 (2005) 90–100 95

Fig. 6. CTB labeling after the excision of the right nodose ganglion (B–E) and transaction of the right C1–C2 spinal nerves (G–J). (A) Schematic illustration of
the medulla showing the orientation of B–E at +800 Am. (B and C) Enlargement of the respectively labeled boxes in Panel (A). Many CTB-labeled neurons are
seen in the left nucleus ambiguus (NA; B), but not in the right NA (Panel C). (D and E) CTB-labeled fibers are numerous in the left nucleus of the solitary tract
(NTS; D), but not in the right side of NTS (E). (F) Schematic illustration showing the orientation of G–J in C1. (G and H) Enlargement of the respectively
labeled boxes in Panel (F). CTB-labeled neurons are seen in the left lamina IX (G), but not in the right lamina IX (H). CTB-labeled fibers are found in left
central cervical nucleus (CeC; I), but not in the right CeC (J). Scale bars=100 Am in Panels (B) and (C); 50 Am in Panels (D), (E), (I), and (J); 25 Am in Panels
(G) and (H).
Y. Atoji et al. / Brain Research 1031 (2005) 90–10096
spinal cord, CTB-labeled neurons were detected mainly in
the medial part of lamina IX, from C1 to C6, and a few in
the lateral part of lamina IX (Fig. 4A and D). In some cases,
one or two labeled neurons were seen in C7. Labeled
neurons were also seen in the intermediate nucleus (Fig. 4C)
Fig. 7. Colocalization of retrogradely labeled fluorogold (FG) and immunohisto
fluorogold-labeled neurons in the nucleus ambiguus are ChAT immunoreactive (arr
of the vagus nerve are ChAT immunoreactive (arrows), and the others are negativ
medial part of lamina IX are ChAT immunoreactive (arrows). C1. Scale bar=50 A
and medial edge of lamina VII in C1. The number of labeled
neurons was densest in C1 and decreased as segments ran
caudally. Labeled varicose fibers were seen in an area
covering the central cervical nucleus and medial edge of
lamina IV in C1 to C3 (Fig. 4B).
chemical staining of choline acetyltransferase (ChAT). (A and B) Three
ows). (C and D) Two fluorogold-labeled neurons in the dorsal motor neuron
e for ChAT (arrowheads). (E and F) Two fluorogold-labeled neurons in the
m.

Y. Atoji et al. / Brain Research 1031 (2005) 90–100 97
When CBT was injected in the ventral midline of the
tracheal wall (group 1), the distribution pattern and
number of labeled neurons in NA, DMV, and C1–C6
Fig. 8. Schematic illustration of the rostrocaudal extent (C1 to C6) of labeling
column, A–F), in the ventral midline of the trachea after removing the m. sternohyoi
R), respectively. Dots indicate the relative differences in distribution but do not rep
labeled axons. Scale bar=1 mm.
were almost identical on both sides (Fig. 3A and B).
However, when CTB was injected in the left side of the
tracheal wall (group 2), the number of labeled neurons in
following injections of CTB in the ventral midline of the trachea (left
deus (middle column, G–L), and in the m. sternohyoidues (right column, M–
resent real numbers. Hatched area indicates the distribution of anterogradely

Fig. 9. Labeled neurons in the medial part of lamina IX of C1 after fluorogold injection in the trachea and CTB injection in the m. sternohyoideus. Arrows
indicate colocalization of fluorogold and CTB. Arrowheads show a fluorogold-positive, CTB-negative neuron. Scale bar=25 Am.
Y. Atoji et al. / Brain Research 1031 (2005) 90–10098
the left side of the NA (249.3F67.4) was significantly
higher than in the right side (141.3F63.9, n=5, Pb0.01),
in the left side of C1 (170.2F57.7) than in the right
(87.5F33.4, n=5, Pb0.01), and in the left side of C2
(39.8F27.9) than in the right (15.7F14.6, n=5, Pb0.05),
but not in DMV (Fig. 5A). Similar differences were noted
when CTB was injected in the right side of the trachea
(data not shown). When CTB was injected in the ventral
midline of the trachea wall in rats, after the removal of the
right nodose ganglion (group 3), no labeled neurons were
found in the NA and DMV of the ipsilateral side (Figs. 5B
and 6A–C), and the number of neurons in C1 decreased
significantly in the transected side (left: 117.8F57.5; right:
57.0F25.7; n=5, Pb0.05). There were also no labeled
terminal fibers in the right NTS (Fig. 6D and E).
Differences were also detected in rats with excised left
nodose ganglion (data not shown). When CTB was
injected in the ventral midline of the tracheal wall, after
the transection of the right C1–C2 spinal nerves (group 4),
labeled neurons were significantly fewer in the ipsilateral
NA (left: 135.6F26.9; right: 81.2F28.7; n=5, Pb0.01), C1
(left: 117.2F42.5; right: 29.6F24.0; n=5, Pb0.01), and C2
(left: 20.8F8.3; right: 2.0F1.2; n=5, Pb0.01) than in the
contralateral sides (Figs. 5C and 6F–H). Labeled fibers in
the right central cervical nucleus disappeared in C1–C2
(Fig. 6I and J).
In group 5, fluorogold-labeled neurons were only found
in the NA, DMV, and lamina IX of the cervical spinal
cord in the central nervous system (data not shown).
Fig. 10. BDA labeling in the nucleus ambiguus (NA) and trachea. (A) Injection si
bar=1 mm. (B) BDA-labeled axon terminals are seen on postganglionic neurons
ChAT was colocalized in all fluorogold-labeled neurons in
the NA and lamina IX of C1–C2 and some neurons in
DMV (Fig. 7A–F).
3.3. Control injections
In control 1, removing the m. sternohyoideus, the
distribution of labeled neurons and varicose fibers (Fig.
8A–F) did not show a difference compared with those of
group 1 (Fig. 8G–L). In control 2, injecting in the m.
sternohyoideus, no labeling was seen in the medulla
oblongata, including NA, DMV, or NTS. In the cervical
spinal cord, labeled neurons were found in most lamina IX
from C1 to C6 (Fig. 8M–R), but labeled varicose fibers were
not seen in all segments. In control 3, the percentage of
colocalization with CTB and fluorogold was 37.2% of
labeled neurons in the medial part of lamina IX of C1 (a
total 239 labeled neurons in three rats; Fig. 9A and B), and
the single labeling of fluorogold and CTB showed 41.9%
and 20.9%, respectively.
3.4. Injection of BDA in NA
When BDA was injected in the rostral part of NA,
labeled axon terminals were frequently found on cell bodies
in the peritracheal ganglion adjoining the smooth muscle
layer (Fig. 10A and B). However, no labeled axon terminals
or axons were detected in the lateral or ventral wall of the
trachea.
te of BDA in the right NA is at +1.2 mm rostral to the obex (arrow). Scale
in a peritracheal ganglion. SM: smooth muscle layer. Scale bar=25 Am.

Fig. 11. Schematic representation of the neural pathways involved in the
control of the trachea. The classic routes of parasympathetic and sensory
innervation are drawn by thin lines. In the current study, two distinct
pathways (thick lines) of motor neurons innervating the trachea originate in
NA and C1–C2. Nerve terminals of sensory neurons in nodose and C1–C2
dorsal root ganglia (DRG) project to C1 and C2 segments (thick lines).
DMV: dorsal motor nucleus of the vagus nerve; NA: nucleus ambiguus;
NTS: nucleus of the solitary tract; C1–C2: first and second segments of the
cervical spinal cord.
Y. Atoji et al. / Brain Research 1031 (2005) 90–100 99
4. Discussion
The present study demonstrated the bilateral innervation
of NA and C1–C2 to the trachea and the direct innervation
of motor neurons of the cervical spinal cord to the trachea in
the rat. The retrograde labeling of motor neurons in DMV
and NA and the axonal labeling in NTS in the medulla is in
good agreement with those of CTB tract-tracing studies
[5,6]. The combination of fluorogold labeling and ChAT
immunostaining showed that ChAT coexists on all fluo-
rogold-labeled neurons in the NA and lamina IX of the
spinal cord and some neurons in DMV. At least all motor
neurons innervating the trachea in NA and cervical spinal
cord are cholinergic.
4.1. Bilateral innervation of the trachea
Using cervical vagotomy and CTB or fluorogold injec-
tion, Perez-Fontan et al. [12] found that each NA innervates
the lung bilaterally in the rat, and the fiber pathway from the
NA to a contralateral lung crosses midline inside the thorax.
In the present study, CTB injection in the ventral midline of
the tracheal wall showed no significant difference in the
distribution of labeled neurons in the NA, DMV, and cervical
spinal cord, but CTB injection in the lateral wall showed not
only bilateral labeling but also significant predominance of
neuronal labeling in the injected side of NA, C1, and C2. The
evidence indicates that a single side of NA, C1, and C2
innervates both sides of the tracheal wall, as well as the lung.
CTB injection in rats after the removal of the nodose ganglion
(group 3) revealed no labeling of neurons in the vagotomized
side of NA and DMV. This finding indicates that CTB-
labeled neurons in the NA and DMV do not decussate in the
central region proximal to the nodose ganglion, but do in the
peripheral region of the vagus nerve. Therefore, it is likely
that the recurrent nerve containing neurons in the NA and
DMV, which innervate contralateral region of the trachea,
decussate inside the thorax as those in NA reach the lung
[12]. On the other hand, the number of labeled neurons in
vagotomized C1 was reduced to approximately half of that on
the intact side. This indicates that C1 neurons innervating the
trachea through the vagus nerve bilaterally decussate in the
central nervous region, i.e., in C1.
4.2. Direct, central innervation of the trachea
In the present study, the injection of CTB in the ventral
wall of the trachea resulted in the labeling of neurons of
lamina IX of C1–C6 and nerve fibers in the central cervical
nucleus of C1–C3. Labeled neurons in the cervical spinal
cord exhibited a wider distribution in the present study than
did that of Haxhiu and Loewy [5], who found neuronal
labeling in C1 and C2 of the dog, ferret, and rat. In control
3, the medial part of lamina IX of C1 contained three types
of labeled neurons: fluorogold-labeled neurons to the
trachea (41.9%), CTB-labeled neurons to the m. sterno-
hyoideus (20.9%), and double-labeled neurons to both the
trachea and m. sternohyoideus (37.2%). The percentage of
neurons innervating the trachea was 79.1% of all labeled
neurons. This shows the presence of a direct projection from
the medial part of lamina IX of C1 to the trachea, in addition
to the motor neurons to the m. sternohyoideus. Labeled
fibers were seen in the ventral funiculus of the upper
cervical segments in the dog and in the cervical dorsal horn
of the ferret, but no labeling was observed in the cervical
dorsal horn in the rat [5]. However, these fiber labeling
patterns are in sharp contrast to those in the present study, in
which CTB-labeled fibers were located mainly in the central
cervical nucleus but not in the dorsal horn or ventral
funiculus. The evidence that CTB-labeled fibers in the
central cervical nucleus of C1–C2 disappeared ipsilaterally
following the severance of C1–C2 spinal nerves suggests
that CTB-labeled fibers in the central cervical nucleus are
centrifugal axons of sensory neurons in C1–C2 dorsal root
ganglia, with projections peripherally to the trachea (Fig. 8).
The injection of CTB in the intercartilagenous space
labeled the neurons in the NA and DMV of the dog, ferret,
and rat [5]. The intraluminal injection of CTB into the
trachea also produced the labeling of neurons in the NA and
DMV in the rat [11]. The authors in the latter study suggested
the presence of direct innervation of the trachea by vagal
motor neurons. However, in these studies, possible labeling
by transganglionic transport via the classic parasympathetic
pathway could not be excluded because the classic para-
sympathetic pathway was still intact anatomically and
functionally when CTB was injected in the tracheal wall or
tracheal lumen. In the present study, the injection of CTB in

Y. Atoji et al. / Brain Research 1031 (2005) 90–100100
the ventral wall of the trachea after cutting the right spinal
nerves of C1 and C2 resulted in a significant reduction of the
numbers of labeled neurons in the right lamina IX. The
reduction of the number of labeled neurons in the cut side
indicates the participation of cervical motor neurons in direct
innervation of the trachea (Fig. 8) because no preganglionic
neurons of either parasympathetic or sympathetic nerve are
present in the cervical spinal cord [2]. The transection of the
spinal nerves in the present study also revealed significant
reduction of the number of CTB-labeled neurons in ipsi-
lateral NA. The evidence indicates the presence of descend-
ing neurons that run caudally in the lower medulla, enter the
upper cervical spinal cord, and go through C1–C2 spinal
nerves. In contrast, a significant reduction of the number of
CTB-labeled neurons in C1 was detected following the
excision of the nodose ganglion. The two experiments of
nerve transection indicate that a small population of motor
neurons in the NA reaches the trachea via the C1 and C2
spinal nerves, and inversely, those in C1 travel to the trachea
via the vagus nerve. That is, motor neurons located in the
transitional region between the medulla and upper cervical
spinal cord have two routes to reach the trachea. Further-
more, neurons in the NA, C1, and C2 that innervate the
trachea bilaterally via the spinal nerves decussate in a central
region, but not in a peripheral one (Fig. 11).
In the present study, BDA injection in the rostral NA,
where many neurons projected to the trachea (group 1),
revealed the labeling of axon terminals on postganglionic
neurons in the peritracheal ganglion, but no labeling in the
nerve fibers or axon terminals of other regions of the
tracheal wall. These results indicate that motor neurons in
the NA terminate on ganglionic neurons and suggest the
absence of direct innervation of the trachea without the
interposition of parasympathetic postganglionic neurons.
Therefore, it is likely that retrograde neuronal labeling in the
NA, DMV, and NTS after CTB injection in the trachea is
due to transganglionic transport. This is probably supported
by the finding that intrinsic tracheal ganglionic neurons
were labeled in addition to the NA, DMV, and cervical
spinal cord when fluorogold was injected in the ventral wall
in the present study (data not shown).
References
[1] D.G. Baker, D.M. McDonald, C.B. Basbaum, R.A. Mitchell, The
architecture of nerves and ganglia of the ferret trachea as revealed by
acetylcholinesterase histochemistry, J. Comp. Neurol. 246 (1986)
513–526.
[2] P. Brodal, Peripheral autonomic nervous system, The Central
Nervous System, 2nd edition, Oxford University Press, New York,
1998, pp. 493–528.
[3] B.J. Canning, A. Fischer, Neural regulation of airway smooth muscle
tone, Respir. Physiol. 125 (2001) 113–127.
[4] R.D. Dey, B. Satterfield, J.B. Altemus, Innervation of tracheal
epithelium and smooth muscle by neurons in airway ganglia, Anat.
Rec. 254 (1999) 166–172.
[5] M.A. Haxhiu, A.D. Loewy, Central connections of the motor and
sensory vagal systems innervating the trachea, J. Auton. Nerv. Syst.
57 (1996) 49–56.
[6] M.A. Haxhiu, A.S.P. Jansen, N.S. Cherniack, A.D. Loewy, CNS
innervation of airway-related parasympathetic preganglionic neurons:
a transneuronal labeling study using pseudorabies virus, Brain Res.
618 (1993) 115–134.
[7] D. Jordan, Central nervous pathways and control of the airways,
Respir. Physiol. 125 (2001) 67–81.
[8] M. Kalia, M.M. Mesulam, Brain stem projections of sensory and
motor components of the vagus complex in the cat: II. Laryngeal,
tracheobronchial, pulmonary, cardiac, and gastrointestinal branches, J.
Comp. Neurol. 193 (1980) 467–508.
[9] D.W. Kusindarta, Y. Atoji, Y. Yamamoto, Nerve plexuses in the
trachea and extrapulmonary bronchi of the rat, Arch. Histol. Cytol. 67
(2004) 41–55.
[10] A.C. Myers, Transmission in autonomic ganglia, Respir. Physiol. 125
(2001) 99–111.
[11] J.J. Perez-Fontan, C.R. Velloff, Labeling of vagal motoneurons and
central afferents after injection of cholera toxin B into the airway
lumen, Am. J. Physiol., Lung Cell. Mol. Physiol. 280 (2001)
L152–L164.
[12] J.J. Perez-Fontan, C.T. Diec, C.R. Velloff, Bilateral distribution of
vagal motor and sensory nerve fibers in the rat’s lungs and
airways, Am. J. Physiol., Regul. Integr. Comp. Physiol. 279 (2000)
R713–R728.
[13] M. Terada, T. Iwanaga, H. Takahashi-Iwanaga, I. Adachi, M.
Arakawa, T. Fujita, Calcitonin gene-related peptide (CGRP)-immu-
noreactive nerves in the tracheal epithelium of rats: an immunohis-
tochemical study by means of whole mount preparations, Arch.
Histol. Cytol. 55 (1992) 219–233.
[14] Y. Yamamoto, T. Ootsuka, Y. Atoji, Y. Suzuki, Morphological and
quantitative study of the intrinsic nerve plexuses of the canine trachea
as revealed by immunohistochemical staining of protein gene product
9.5, Anat. Rec. 250 (1998) 438–447.




















