
Neuropeptides in interscapular and perirenal brown adipose tissue in the rat: a plurality of...
9
Journal of Neurocytology 17, 305-311 (1988) Neuropeptides in interscapular and perirenal brown adipose tissue in the rat: a plurality of innervation D. NORMAN 1, S. MUKHERJEE 1, D. SYMONS 1, R. T. JUNG 2 and J. D. LEVER 1 1Department of Anatomy, University College, PO Box 78, Cardiff CF1 1XL, UK 2Department of Medicine, Ninewells Hospital, Dundee, DDI 9SY, UK Received 21 October 1987; revised 21 January 1988; accepted 28 January 1988 Summary The occurrence of neuropeptides in rat brown adipose tissue has been investigated. Immunohistochemical studies on interscapular and perirenal brown fat have demonstrated unequivocally the presence of substance-P (SP)-like, neuropeptide-Y (NPY)-like and calcitonin gene related-peptide (CGRP)-like immunoreactive elements (putative nerves) in adventitial distribution on inter- and intralobular supply arteries and in accompanying nerve bundles. At a more peripheral level, some NPY-like immunoreactive elements and a greater number of CGRP-like immunoreactive elements were observed in the parenchymal field. Somatostatin, bombesin, neurotensin, enkephalin, and vasoactive-intestinal- polypeptide immunoreactivities were not detected. No differences in neuropeptide distribution were noted between interscapular and perirenal brown fat. There is a degree of coincident distribution of SP, NPY and CGRP with that of noradrenergic nervous elements as visualized by condensation histochemistry. Since after 6-hydroxydopamine treatment not all the nerve terminals in rat brown adipose tissue are stigmatized (earlier report), the present results have been discussed in the light of a possible pluralism in innervation of brown adipose tissue. Introduction There is well-documented evidence to suggest that to a very large extent the functions of brown adipose tissue (BAT) are neurally controlled: that the sym- pathetic nervous system is involved in adaptational responses of BAT in rodents was indicated by Cottle & Cottle (1970), who demonstrated (by formaldehyde- induced fluorescence histochemistry) an increase in catecholamine levels in interscapular brown adipose tissue (ISBAT) during cold acclimation. Again BAT has been identified as the site of regulatory non- shivering thermogenesis in many mammalian species (Foster & Frydman, 1978) and of diet-induced thermogenesis in the rat during cafeteria-feeding (Rothwell & Stock, 1979, 1982). Moreover, the production of the essential uncoupling protein thermogenin, tissue-specific to BAT, has been shown to be noradrenaline dependent (Mory et al., 1984). In this connection it is generally agreed that the brown adipocyte is directly innervated by adrenergic nerves (Girardier & Seydoux, 1986). Evidence of the trophic significance of adrenergic nerves in BAT is suggested by the absence of non-shivering thermogenesis in cold-exposed rats shortly after treatment with 6- hydroxydopamine (Smith & Horwitz, 1969). Further, 0300--4864/88 $03.00 + .12 1988 Chapman and Hall Ltd. it has been shown that infusion of exogenous noradrenaline increases the regional blood flow through BAT in the intact rat (Foster et al., i980) and rabbit (Hardman & Hull, 1970). Although the thermogenic response of BAT to infused noradrena- line is mediated via ~3 receptors (being: propranalol blockable; Bukowiecki et al., 1980) and there is evidence (Flaim et al., 1976) of a noradrenaline or vasoconstrictive response, to date an a- or B-mediated vasodilator receptor mechanism in BAT has not been reported. However, Foster & Depocas (1980) have suggested the possible release of an as yet unidentified vasodilator agent during thermogenesis. Plurality of the innervation to rat ISBAT has also been suggested by the finding that after 6- hydroxydopamine treatment not all nerve terminals in the tissue are stigmatized (Lever et al., 1986). In another sympathetic projection, namely bovine sple- nic nerves, Fried et al., (1986) have demonstrated the coexistence of neuropeptide-tyrosine (NPY) and enkephalin (ENK) with noradrenaline. Again ENK- like immunoreactivity (i.r.) has been reported in association with dopamine-~-hydroxylase i.r. in
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Neuropeptides in interscapular and perirenal brown adipose tissue in the rat: a plurality of...

Journal of Neurocytology 17, 305-311 (1988)
Neuropeptides in interscapular and perirenal brown adipose tissue in the rat: a plurality of innervation
D. N O R M A N 1, S. M U K H E R J E E 1, D. S Y M O N S 1, R. T. J U N G 2 and J. D. L E V E R 1
1Department of Anatomy, University College, PO Box 78, Cardiff CF1 1XL, UK 2Department of Medicine, Ninewells Hospital, Dundee, DDI 9SY, UK
Received 21 October 1987; revised 21 January 1988; accepted 28 January 1988
Summary
The occurrence of neuropeptides in rat brown adipose tissue has been investigated. Immunohistochemical studies on interscapular and perirenal brown fat have demonstrated unequivocally the presence of substance-P (SP)-like, neuropeptide-Y (NPY)-like and calcitonin gene related-peptide (CGRP)-like immunoreactive elements (putative nerves) in adventitial distribution on inter- and intralobular supply arteries and in accompanying nerve bundles. At a more peripheral level, some NPY-like immunoreactive elements and a greater number of CGRP-like immunoreactive elements were observed in the parenchymal field. Somatostatin, bombesin, neurotensin, enkephalin, and vasoactive-intestinal- polypeptide immunoreactivities were not detected. No differences in neuropeptide distribution were noted between interscapular and perirenal brown fat.
There is a degree of coincident distribution of SP, NPY and CGRP with that of noradrenergic nervous elements as visualized by condensation histochemistry. Since after 6-hydroxydopamine treatment not all the nerve terminals in rat brown adipose tissue are stigmatized (earlier report), the present results have been discussed in the light of a possible pluralism in innervation of brown adipose tissue.
Introduction
There is well-documented evidence to suggest that to a very large extent the functions of brown adipose tissue (BAT) are neurally controlled: that the sym- pathetic nervous system is involved in adaptational responses of BAT in rodents was indicated by Cottle & Cottle (1970), who demonstrated (by formaldehyde- induced fluorescence histochemistry) an increase in catecholamine levels in interscapular brown adipose tissue (ISBAT) during cold acclimation. Again BAT has been identified as the site of regulatory non- shivering thermogenesis in many mammalian species (Foster & Frydman, 1978) and of diet-induced thermogenesis in the rat during cafeteria-feeding (Rothwell & Stock, 1979, 1982). Moreover, the production of the essential uncoupling protein thermogenin, tissue-specific to BAT, has been shown to be noradrenaline dependent (Mory et al., 1984). In this connection it is generally agreed that the brown adipocyte is directly innervated by adrenergic nerves (Girardier & Seydoux, 1986). Evidence of the trophic significance of adrenergic nerves in BAT is suggested by the absence of non-shivering thermogenesis in cold-exposed rats shortly after treatment with 6- hydroxydopamine (Smith & Horwitz, 1969). Further,
0300--4864/88 $03.00 + .12 �9 1988 Chapman and Hall Ltd.
it has been shown that infusion of exogenous noradrenaline increases the regional blood flow through BAT in the intact rat (Foster et al., i980) and rabbit (Hardman & Hull, 1970). Although the thermogenic response of BAT to infused noradrena- line is mediated via ~3 receptors (being: propranalol blockable; Bukowiecki et al., 1980) and there is evidence (Flaim et al., 1976) of a noradrenaline or vasoconstrictive response, to date an a- or B-mediated vasodilator receptor mechanism in BAT has not been reported. However, Foster & Depocas (1980) have suggested the possible release of an as yet unidentified vasodilator agent during thermogenesis.
Plurality of the innervation to rat ISBAT has also been suggested by the finding that after 6- hydroxydopamine treatment not all nerve terminals in the tissue are stigmatized (Lever et al., 1986). In another sympathetic projection, namely bovine sple- nic nerves, Fried et al., (1986) have demonstrated the coexistence of neuropeptide-tyrosine (NPY) and enkephalin (ENK) with noradrenaline. Again ENK- like immunoreactivity (i.r.) has been reported in association with dopamine-~-hydroxylase i.r. in

306 NORMAN, MUKHERJEE, SYMONS, JUNG and LEVER
membrane-bounded secretory granules of cat carotid body, h u m a n adrenal medul la and phaeochromocy- toma (Varndell et al., 1982).
The present investigation is aimed at the identifica- tion of neuropept ides in relation to known autonomic (noradrenergic) nervous projections in paravascular and parenchymal distributions in rat ISBAT and PRBAT. This is addit ionally pert inent in the light of recent comment by Girardier & Seydoux (1986) that there may be dissimilarities in the neural regulation of ISBAT and perirenal BAT (PRBAT) becuase of differences in their location and in the environmental influences upon them.
Materials and methods
Samples of ISBAT and PRBAT were obtained under ether anaesthesia from young (250g) warm-acclimated adult male Wistar rats. Material for the demonstration of noradrenergic nerves was frozen by dry ice onto chucks and 12-15-~m cryostat sections cut at -30~ were thawed onto glass slides and processed according to a modification (Cottle et al., 1985) of the sucrose-potassium phosphate- glyoxylic acid (SPG) technique of de la Torre & Surgeon (1976), employing the following sequences: specimen immersion for 20 s in SPG solution (excess of which was then drained from the slide); cool-air drying for 5min; coverslip mounting with liquid paraffin prior to oven incubation at 60 ~ C for 20 min. Slides were viewed in a Leitz Laboflux 12 microscope with Pleopak 2-5 fluorescence vertical illuminator employing BP 350-380 nm and suppres- sor LP 430 nm filters.
Material for immunohistochemical neuropeptide identi- fication was immersion-fixed for 90min at 4~ in 0.4% parabenzoquinone in phosphate-buffered saline (PBS) or for 24h at 4~ in Zamboni's fixative (Zamboni & de Martino, 1967~, followed by a 24-h rinse in 5% sucrose in PBS at 4~ Cryostat sections (10 ~m) were cut and mounted on poly-L-lysine-coated slides. Sections were separately reacted successively with (1) rabbit (primary) antisera (Amersham) prepared against substance P (SP), neuropeptide Y (NPY), somatostatin (SOM), vasoactive intestinal polypeptide (VIP), enkephalin (ENK), neurotensin (NT) (Amersham), calcitonin gene-related- peptide (CGRP) and bombesin (BOM) (Cambridge Re- search Biochemicals); (2) donkey biotinylated anti-rabbit IgG F(ab)2 fragment (secondary antibody) (Amersham); (3) fluorescein-streptavidin conjugate (Amersham). The fol- lowing protocol was employed in respect of all the above investigations: slides were rinsed in three changes of PBSFFriton X; incubated for 20rain in normal donkey serum; incubated overnight at high humidity at room
temperature in primary antibody) rinsed briefly in three changes of PBSFFriton X; incubated for 30min in the secondary antibody (1:100 dilution); rinsed again with three changes of PBSFFriton X; incubated at room temperature irf the fluorescent marker for 15min (1:400 dilution); rinsed finally in three changes of PBS/Triton X and mounted in PBS/glycerine (9:1) containing 0.1% p-phenylenediamine.
A range of primary antibody dilutions was investigated for each neuropeptide. Optimal dilutions (giving maximum contrast between background and reaction product) were as follows: SP, 1:2000; NPY, 1:3000; SOM, 1:4000; CGRP, 1:2000; VIP, 1:10000; ENK, 1:4000; NT, 1:3000; BOM, 1:4000.
Methodological control was insured in each instance by employing indentical procedural protocol, on the one hand omitting primary antibody and on the other including primary antibody preabsorbed with antigen. In Figs 2 and 3 such controls are exemplified in respect of NYP i.r. Similar controls were obtained for all the other neuropeptides studied, but for reasons of economy are not portrayed. Antibody specificity was strongly suggested by in-parallel demonstration of specific antigen at (other) authenticated distributions : SP and BOM in nerve fibres in posterior horn of spinal cord; NPY in myometrial nerves; SOM in D cells of islets of Langerhans; VIP and NT in paravascular nerves of the gut; ENK in ileal myenteric nerves: CGRP in thyroid C cells (depicted in Fig. 6).
Results
Distributions of SPG-positive elements (norad- renergic nerves) were similar in ISBAT and PRBAT in both periarterial and parenchymal deployment .
NPY i.r. elements (putative nerves) were discerned in adventitial distribution on inter- and intralobular arteries in both ISBAT and PRBAT (Fig. 1) but were additionally seen at more peripheral levels around much smaller blood vessels in the parenchymal field (Fig. 4). The NPY-positive i.r. periarterial elements presented as prominent dense plexuses of beaded fibres.
SP-like i.r. elements (putative nerves) were dis- cerned in adventitial distribution on inter- and intralobular arteries in both ISBAT and PRBAT. Within this distribution these presumptive SP- positive nerves were sparsely deployed in fine beaded plexuses (Figs 5, 7).
CGRP-like i.r. elements. (Figs 8-10) showed a similar periarterial distribution to NPY i.r. elements (Figs 1, 4) and to SP i.r. elements (Figs 5, 7) in both ISBAT and PRBAT. Again at more peripheral levels,
Fig. 1. NPY-like i.r. elements (large arrows) in putative nerve plexuses in adventitial distribution on walls of interlobular arteries (A) in rat ISBAT a lobule of which is at L. Note fine beaded NPY-like i.r. elements in accompanying nerve bundle (small arrows) and elastic laminae (e). x 400.
Figs 2 and 3 Sequential sections of the interlobular structures depicted in Fig. 1. used for methodological control. Fig. 2, identical methodological protocol minus primary antibody. Fig. 3, identical protocol but using antigen-absorbed primary antibody. Note absence of NPY-like i.r. fluorescence in Figs 2 and 3 x 400. Fig. 4. -NP~'-like i.r. (putative nerve) plexuses (large arrows) around small intralobular arteries in PRBAT. x 400.
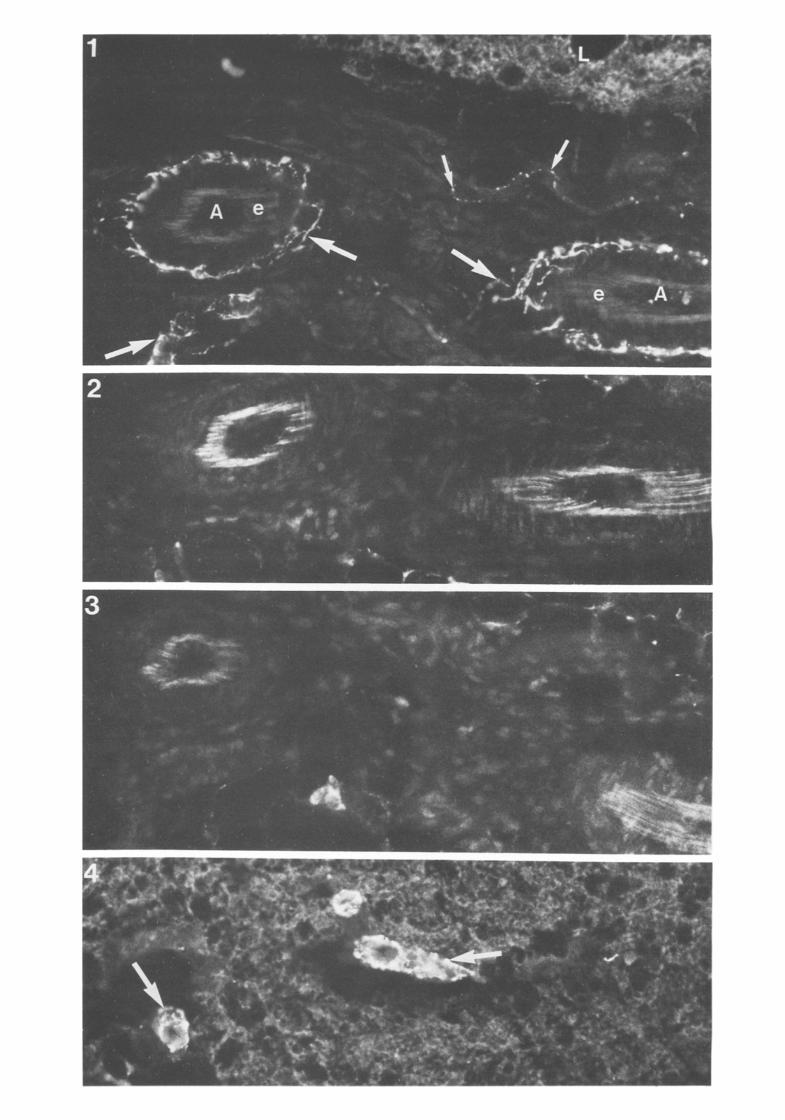

308 NORMAN, MUKHERJEE, SYMONS, JUNG and LEVER
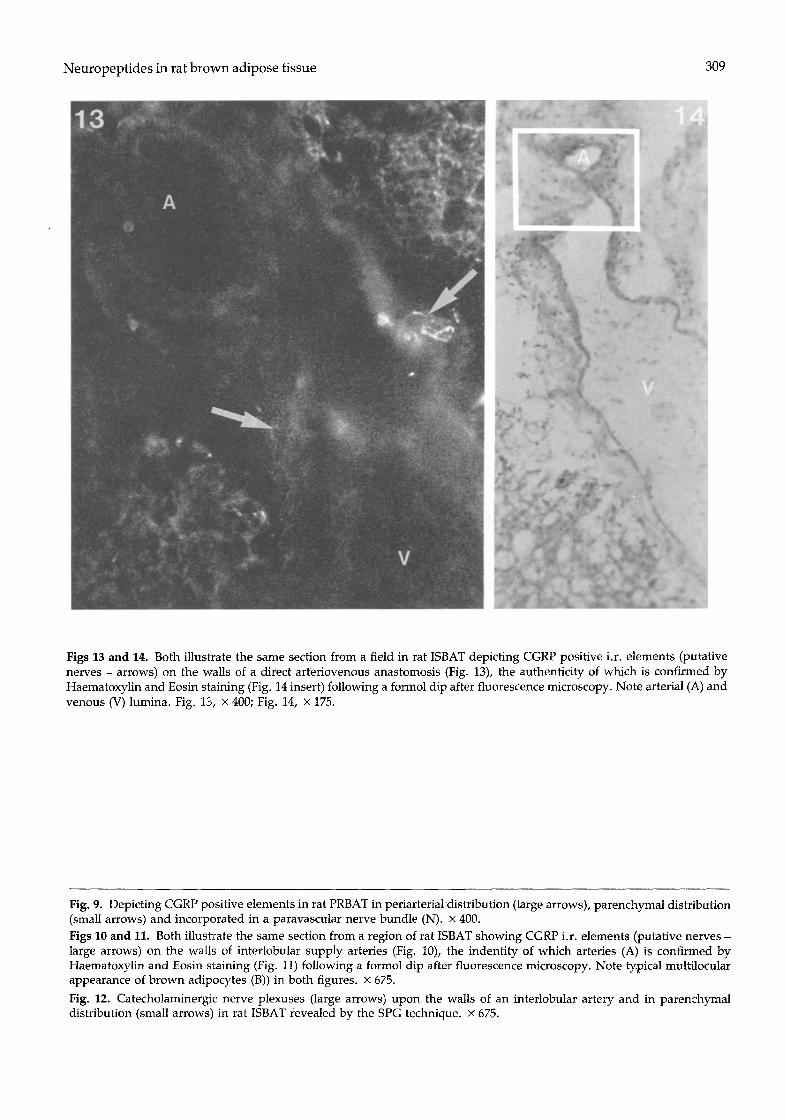
most probably in a paracapillary location, CGRP i.r. beaded elements were discerned in BAT parenchym- al fields and this distribution (Fig. 9) was more extensive than the comparable NPY i.r. projection. In what was certainly a fortunate section through an arteriovenous anastomosis (AVA) between interlobu- lar vessels (Fig. 13), CGRP i.r. beaded elements (putative nerves) were located in the arterial adventi- tia and on the wall of the neck-like AVA connecting channel but not upon venous walls (Fig. 14). The authenticity of the AVA was confirmed by haema- toxylin and eosin staining of the same section and light microscopy (Fig. 13) following examination by fluorescence microscopy.
In nerve bundles adjacent to interlobular vessels SP-positive, CGRP-positive (Fig. 9) and NPY-positive (Fig. 1) elements were discerned.
Neither SOM-positive, BOM-positive, ENK- positive, NT-positive nor VIP-positive i.r. elements were observed in either ISBAT or PRBAT.
In the case of all neuropeptides tested on BAT, satisfactory positive i.r. was demonstrated at aut- henticated sites: SP in nerve fibres in the substantia gelatinosa of spinal cord; NPY in myometrial and cardiac nerves; CGRP in thyroid C cells (Fig. 6); SOM in D cells of islets of Langerhans; NT and ENK in ileal myenteric nerve plexuses; VIP in paravascular nerves of the gut; BOM in nerve fibres in the substantia gelatinosa and ventral horn of spinal cord.
Methodological specificity was obtained in respect of all the above neuropeptides by means of protocol controls [i.e. minus primary antibody (cf. Figs 1, 2) and antigen-absorption controls (cf. Figs. 1, 3) (see Materials and methods)] whether or not positive neuropeptide distribution was found in BAT.
D i s c u s s i o n
From the present immunohistochemical study on the distribution of certain neuropeptides within rat brown adipose tissue (ISBAT and PRBAT), SP-, NPY- and CGRP- like i.r. elements (putative nerves) were located while no i.r. activity for SOM, VIP, BOM, ENK and NT was found. There was a varying degree of coincidence in the distribution within both ISBAT and PRBAT of the above three neuropeptides, with that of noradrenergic nerve plexuses as demonstrated
by formaldehyde-induced fluorescent and SPG tech- nology in rodent BAT (Cottle & Cottle, 1970; Lever et al., 1981). Thus SP-like i.r. elements were found only in periarterial distribution, while both NPY- and CGRP-like i.r. elements were located not on ly at periarterial sites, but also further peripherally in the parenchymal field. Furthermore, similar patterns of SP and CGRP i.r. distribution were observed at periarterial sites, although of the two peptides CGRP distribution was invariably more widespread. Fine structural studies on rat BAT have previously indicated a paracapillary location for nerve terminals related to brown adipocytes (Lever et al., 1981).
Recent observations based on three-dimensional studies of arterial and venous pathways in rat brown fat have revealed the presence of AVAs at peripheral and perilobular sites (Lever et al., 1985) in ISBAT. While the microscopic recognition of AVAs is quite feasible, their location within a tissue block requires serialization, a process which is laborious on the cryostat. However, in a good representative section in the present investigation of CGRP antigenicity in ISBAT, appearances suggestive of a CGRP-positive innervation of an AVA were encountered. It remains to investigate the distribution of SP and NYP in relation to the possible innervation of AVAs within BAT and in this context to confirm or otherwise the presence of an adrenergic innervation on the walls of these anastomoses.
Present and projected studies on the innervation of BAT must be viewed in both vasomotor and metabolic contexts. Namely the control of the linear blood supply (via the peri-interlobular and intralobu- lar arterial tree) and of diversionary arteriovenous anastomotic pathways; and in relation to metabolic trophism at a parenchymal level. In the latter connection the role of noradrenaline in promoting non-shivering and diet-induced thermogenesis is widely accepted (Foster & Frydman, 1978; Rothwell & Stock, 1979, 1982). However, the functional signifi- cance of peptidergic (nervous) elements within BAT remains to be established by experimental means and possibly by clinical trials.
Coincident distributions of neuropeptides per se and neuropeptides with classical transmitters have been reported in various tissues. Thus Cannon et al. (1986) have described co-localization of NPY and
Fig. 5. Substance P-like i.r.beaded elements (putative nerves) (arrows) on the walls of small supply arteries in rat ISBAT. x 675. Fig. 6 Rat thyroid gland showing CGRP immunoreactivity in the thyroid parafollicular C cells (arrows). x 400.
Figs 7 and 8 Consecutive sections illustrating arterial (A) and venous (V) profiles in rat ISBAT. Similar patterns of SP (Fig. 7) and CGRP (Fig. 8) positive elements (putative nerves) are depicted at periarterial sites (large arrows); the distribution of CGRP elements being more widespread. Note CGRP positive elements in parenchymal distribution (small arrow), x 400.
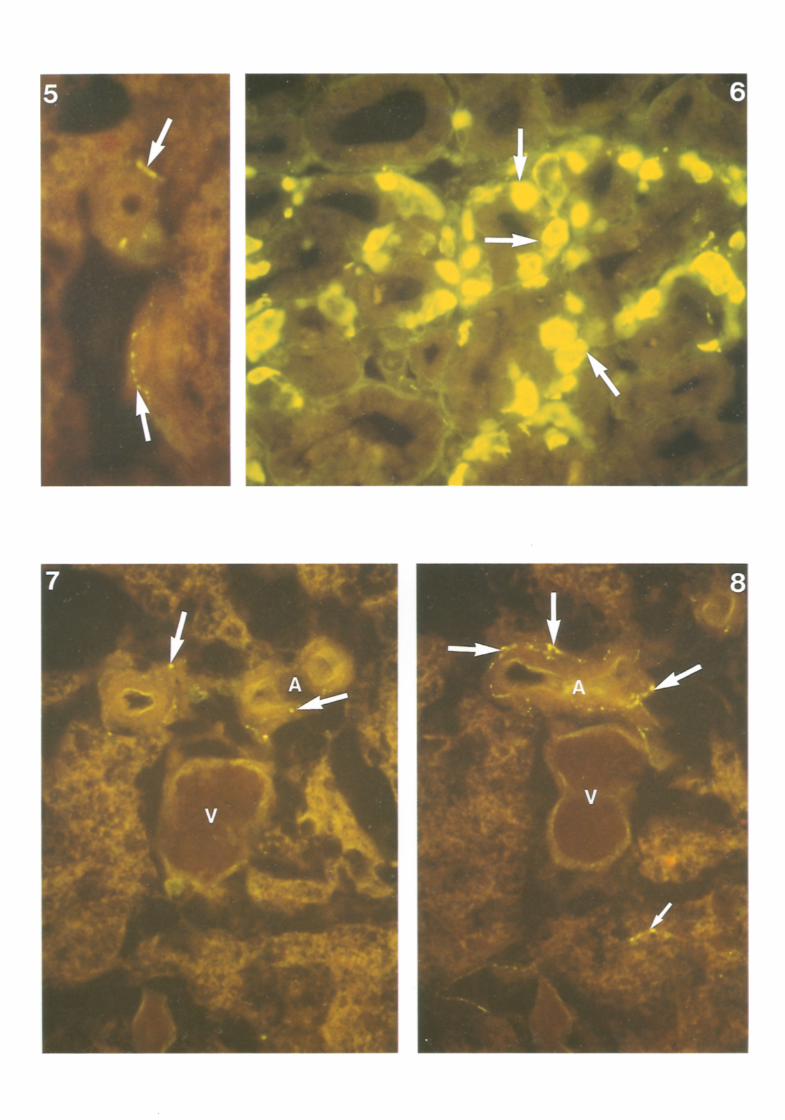

N e u r o p e p t i d e s in rat b r o w n ad ipose t issue 309
Figs 13 and 14. Both illustrate the same section from a field in rat ISBAT depicting CGRP positive i.r. elements (putative nerves - arrows) on the walls of a direct arteriovenous anastomosis (Fig. 13), the authenticity of which is confirmed by Haematoxylin and Eosin staining (Fig. 14 insert) following a formol dip after fluorescence microscopy. Note arterial (A) and venous (V) lumina. Fig. 13, x 400; Fig. 14, x 175.
Fig. 9. Depicting CGRP positive elements in rat PRBAT in periarterial distribution (large arrows), parenchymal distribution (small arrows) and incorporated in a paravascular nerve bundle (N). x 400. Figs 10 and 11. Both illustrate the same section from a region of rat ISBAT showing CGRP i.r. elements (putative nerves - large arrows) on the walls of interlobular supply arteries (Fig. 10), the indentity of which arteries (A) is confirmed by Haematoxylin and Eosin staining (Fig. 11) following a formol dip after fluorescence microscopy. Note typical multilocular appearance of brown adipocytes (B)) in both figures, x 675.
Fig. 12. Catecholaminergic nerve plexuses (large arrows) upon the walls of an interlobular artery and in parenchymal distribution (small arrows) in rat ISBAT revealed by the SPG technique, x 675.

310 NORMAN, MUKHERJEE, SYMONS, JUNG and LEVER
tryosine hydroxylase within sympathetic vasomotor nerves in rat ISBAT. Similarly the co-existence of NPY and noradrenaline has been recorded in sympathetic neurons innervating bovine spleen (Fried et al., 1986), and Sundler et al. (1986) have described NPY in peripheral adrenergic and enteric nervous systems. Available evidence suggests a vasoconstrictive role for NPY in relation to noradrenaline (Edvinsson et al., 1987).
We are currently engaged in immunofluorescence and EM immunogold studies on autonomic nervous projections in BAT in an investigation of the possible coincidence of neuropeptides and noradrenaline in animals pretreated with the false transmitters 5- hydroxy and 6-hydroxy dopamine.
Evidence that SP is distributed inter alia in primary sensory neurons has been afforded by H6kfelt et al.
(1975) and its co-distribution with CGRP has been
demonstrated ultrastructurally by immunogold tech- nology within vesicles in dorsal root and trigeminal ganglion cells and in peripheral sensory nerves by Gulbenkian et al. (1986). Pharmacological evidence indicates a likely vasodilatory role for both CGRP and SP in the cardiovascular system (Edvinsson et al., 1982; Brain et al., 1985)
In the present study no differences were observed in either the catecholamine or neuropeptide distribu- tions between ISBAT and PRBAT in spite of their different locations and environment.
Acknowledgements
We are grateful to Mr Peter Hire for photographic help and to Miss Adriana Filice for typing this manuscript. This work was supported by MRC grant 8611634N.
References
BRAIN, S. D., WILLIAMS, T. J., TIPPINS, J. R., MORRIS t H. R., & MACINTYRE, I. (1985) Ca lc i ton in g e n e - r e l a t e d peptide is a potent vasodilator. Nature 313, 54-6.
BUKOWIECKIt L., FOLLEA, N., PARAD1S, A. & COLLET, A. (1980) Stereospecific stimulation of brown adipocyte respiration by catecholamines via adrenoreceptors. American Journal of Physiology 238, E552-63.
CANNON, B., NEDERGAARD, J., LUNDBERG, J. M., H(gKFELT, T., TERENIUS, L. & GOLDSTEIN, M. (1986) Neuropeptide tryosine (NPY) is co-stored with noradre- naline in vascular but not in parenchymal sympathetic nerves of brown adipose tissue. Experimental Cell Research 164, 546-50.
COTTLE, M. K. W. & COTTLE, W. H. (1970) Adrenergic fibres in brown fat and cold-acclimated rats. Journal of Histochemistry and Cytochemistry 18, 116-9.
COTTLE, M. K. W., COTTLE, W. H., PERUSSE, F. & BUKOWIECKI, L. I. (1985) An improved glyoxylic acid technique for the histochemical localisation of catechola- mines in brown adipose tissue. Histochemical Journal 17, 1279-88.
DE LA TORRE, J. C. & SURGEON, J. W. (1976) A methodological approach to rapid and sensitive monoamine histofluorescence using a modified glyox- ylic acid technique. The SPG method. Histochemistry 49, 81-93,
EDVINSON, L., McCULLOCH, J., ROSELL, S. & UDDMAN, R. (1982) Antagonism by (D-Pro 2, D-Trp 7-9) --subst- ance P of the cerebrovascular dilatation induced by substance P. Acta Physiologica Scandinavica 116, 411-6.
EDVINSSON, L., HAKANSON, R., WAHLESTEDTr C. & UDDMAN, R. (1987) Effects of neuropeptide Y on the cardiovascular system. TIPS 8, 231-5.
FLAIMr K. E., HOROWITZ, J. M. & HORWITZ, B. (t976) Functional and anatomical characteristics of the nerve- brown adipose interaction in the rat. Pfliigers Arch. 365, 9-14.
FOSTER, D. O., & DEPOCAS, F. (1980) Ev idence aga ins t noradrenergic regulation of vasodilation in rat brown adipose tissue. Canadian Journal of Physiology and Pharmacology 58, 1418-25.
FOSTER, D. O. & FRYDMAN, M. L. (1978) Non-shivering thermogenesis in the rat. II. Measurements of blood flow with microspheres point to brown adipose tissue as the dominant site of the calorigenesis induced by no,adrenaline. Canadian Journal of Physiology and Pharma- cology 56, 110-2.
FOSTER, D. O., DEPOCAS, F. & FRYDMAN, L. (1980) Noradrenaline-induced calorigenesis in warm- and cold-acclimated rats: relations between concentration of no,adrenaline in arterial plasma blood flow to differently located masses of brown adipose tissue and calorigenic response. Canadian Journal of Physiology and Pharmacology 58, 915-24.
FRIED, G., TERENIUS, L., BRODIN, E., EFENDiC, S., DOCKRAY, G., FAHRENKRUG, J., GOLDSTEIN, M. & HOKFELT, T. (1986) Neuropeptide Y, enkephalin and nordrenaline coexist in sympathetic neurons inner- vating the bovine spleen. Biochemical and immuno- histochemical evidence. Cell and Tissue Research 243, 495-508.
GIRARDIER, L. & SEYDOUX, J. (1986) N e u r a l con t ro l of brown adipose tissue. In Brown Adipose Tissue (edited by TRAYHURN, P. & NICHOLLS, D. G..), pp. 122-51. London: E. Arnold.
GULBENKIAN, S., MERINGHI, A., WHARTON, J., VARNDE LL, I. M. & POLAK, I. M. (1986) Ultrastructural evidence for the coexistence of calcitonin gene-related peptide and substance P in secretory vesicles of peripheral nerves in the guinea pig. Journal of Neurocytology 15, 535-42.
HARDMAN, M. J., & HULL, D. (1970) Fat metabolism in brown adipose tissue in vivo. Journal of Physiology (London) 206, 263-73.

Neuropeptides in rat brown adipose tissue 311
HOKFELT, T., KELLERTH, J. O., N1LSSON, G. & PERNOW, B. (1975) Substance P. Localisation in the central sensory neurons. Science 190, 889-90.
LEVER, J. D., NNODIM, J. O., & SYMONS, D. (1985) Arter iovenous anastomoses in interscapular brown adipose tissue in the rat. Journal of Anatomy 143, 207-10.
LEVER, J. D. NORMAN, D., PALMER, S. A. & SANTER, R. M. (1981) Nerve distr ibution within brown adipose tissue of the rat. Journal of Anatomy 133, 661.
LEVER, J. D. NORMAN, D., SYMONS, D. & JUNG, R. T~ (1986) Evidence of pept idergic nerves within rat human brown adipose tissue. Journal of Anatomy 149, 230.
MORY, G., BOUILLAUD, F., COMBES-GEORGE, M. & RICQUIER, D. (1984) Noradrenal ine controls the concen- tration of the uncoupl ing protein in brown adipose tissue. FEBS Letters 16G 393-6.
ROTHWELL, N. J. & STOCK. M. J. (1979) The role of brown adipose tissue in diet- induced thermogenesis . Nature 281, 31-5.
ROTHWELL, N. J. & STOCK, M. J. (1982) Nerual regulation of thermogenesis . Trends in Neuroscience 5, 124-6.
SMITH, R. E. & HORWITZ, B. A. (1969) Brown fat and thermogenesis . Physiological Reviews 49, 330-425.
SUNDLER, F., H~KANSON, R., EKBLAD, E., UDDMANr R. & WAHLESTEDT, Co (1986) Neuropept ide Y in the per ipheral adrenergic and enteric nervous systems. International Review of Cytology 102, 243-69.
VARNDELL, I. M. TAPIA, F. J., DE MEY, J., RUSH, R. A., BLOOM S. R. & POLAK, J. M. (1982) Electron immunocytochemical localisation of enkephalin-l ike material in catecholamine-containing cells of the carotid body, the adrenal medulla, and in phaeochromocytomas of man and other mammals . Journal of Histochemistry and Cytochemistry 30, 682-90.
ZAMBONI, L. & DE MARTINO, C. (1967) Buffered picric acid formaldehyde: a new rapid fixative for electron microscopy Journal of Cell Biology 35, 148A.




















