
Expression of neuropeptides and anoctamin 1 in the embryonic and adult zebrafish intestine,...
18
1 23 Cell and Tissue Research ISSN 0302-766X Volume 354 Number 2 Cell Tissue Res (2013) 354:355-370 DOI 10.1007/s00441-013-1685-8 Expression of neuropeptides and anoctamin 1 in the embryonic and adult zebrafish intestine, revealing neuronal subpopulations and ICC-like cells Leen Uyttebroek, Iain T. Shepherd, Guy Hubens, Jean-Pierre Timmermans & Luc Van Nassauw
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Expression of neuropeptides and anoctamin 1 in the embryonic and adult zebrafish intestine,...

1 23
Cell and Tissue Research ISSN 0302-766XVolume 354Number 2 Cell Tissue Res (2013) 354:355-370DOI 10.1007/s00441-013-1685-8
Expression of neuropeptides andanoctamin 1 in the embryonic and adultzebrafish intestine, revealing neuronalsubpopulations and ICC-like cells
Leen Uyttebroek, Iain T. Shepherd, GuyHubens, Jean-Pierre Timmermans & LucVan Nassauw

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag Berlin Heidelberg. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

REGULAR ARTICLE
Expression of neuropeptides and anoctamin 1in the embryonic and adult zebrafish intestine, revealingneuronal subpopulations and ICC-like cells
Leen Uyttebroek & Iain T. Shepherd & Guy Hubens &
Jean-Pierre Timmermans & Luc Van Nassauw
Received: 22 November 2012 /Accepted: 17 June 2013 /Published online: 24 July 2013# Springer-Verlag Berlin Heidelberg 2013
Abstract This immunohistochemical study in zebrafish aimsto extend the neurochemical characterization of enteric neuro-nal subpopulations and to validate a marker for identification ofinterstitial cells of Cajal (ICC). The expression of neuropeptidesand anoctamin 1 (Ano1), a selective ICC marker in mammals,was analyzed in both embryonic and adult intestine.Neuropeptides were present from 3 days postfertilization(dpf). At 3 dpf, galanin-positive nerve fibers were found inthe proximal intestine, while calcitonin gene-related peptide(CGRP)- and substance P-expressing fibers appeared in thedistal intestine. At 5 dpf, immunoreactive fibers were presentalong the entire intestinal length, indicating a well-developedpeptidergic innervation at the onset of feeding. In the adultintestine, vasoactive intestinal peptide (VIP), pituitary adenylatecyclase-activating peptide (PACAP), galanin, CGRP and sub-stance P were detected in nerve fibers. Colchicine pretreatmentenhanced only VIP and PACAP immunoreactivity. VIP andPACAP were coexpressed in enteric neurons. Colocalizationstainings revealed three neuronal subpopulations expressingVIP and PACAP: a nitrergic noncholinergic subpopulation, aserotonergic subpopulation and a subpopulation expressing noother markers. Ano1-immunostaining revealed a 3-dimensionalnetwork in the adult intestine containing multipolar cells at themyenteric plexus and bipolar cells interspersed between circularsmoothmuscle cells. Ano1 immunoreactivity first appeared at 3
dpf, indicative of the onset of proliferation of ICC-like cells. Itis shown that the Ano1 antiserum is a selective marker of ICC-like cells in the zebrafish intestine. Finally, it is hypothesizedthat ICC-like cells mediate the spontaneous regular activity ofthe embryonic intestine.
Keywords Zebrafish . Enteric nervous system .
Neuropeptides . Anoctamin 1 . Interstitial cells of Cajal
Introduction
Over the past two decades, the zebrafish, Danio rerio, hasemerged as a leading vertebrate model system because ofseveral interesting characteristics. Zebrafish are easy to raiseand have a generation time of 3 months. Females lay hundredsof eggs at weekly intervals. Fertilization and development ofthe transparent embryos are external, allowing easy observa-tion and manipulation. Molecular pathways, organ systemsand physiology are well conserved between zebrafish andmammals including man. The zebrafish genome has beensequenced and techniques have been developed that make itpossible to produce genetically modified animals cheaply andrapidly (Grunwald and Eisen 2002; Matthews et al. 2002).Throughout the last decade, the zebrafish has also emerged asan important model for the development of gastrointestinal(GI) activities and their control mechanisms (Olsson 2009;Olsson et al. 2008; Olsson and Holmgren 2001; Rich et al.2007; Uyttebroek et al. 2010). In vertebrates, GI functions,including motility, absorption, secretion, immune responsesand blood flow, are controlled by the autonomic nervoussystem, including the enteric nervous system (ENS) (Furness2012). Although the general morphology and development ofthe ENS have been studied extensively in zebrafish (Shepherdand Eisen 2011), specific knowledge about the functional andmorphological characteristics of enteric neurons is still incom-plete. Characterization of enteric neurons into different
L. Uyttebroek :G. Hubens : L. Van Nassauw (*)Laboratory of Human Anatomy and Embryology, Faculty ofMedicine and Health Sciences, University of Antwerp,Groenenborgerlaan 171, 2020 Antwerpen, Belgiume-mail: [email protected]
I. T. ShepherdDepartment of Biology, Emory University, Atlanta, GA, USA
J.<P. TimmermansLaboratory of Cell Biology and Histology, Department ofVeterinary Sciences, University of Antwerp, Antwerpen, Belgium
Cell Tissue Res (2013) 354:355–370DOI 10.1007/s00441-013-1685-8
Author's personal copy

neuronal subpopulations allows linking to different functionalclasses that control GI processes (Brookes 2001; Furness 2012;Timmermans et al. 1997). The zebrafish ENS is less complexcompared to mammals: no submucosal plexus is present andsingle enteric neurons form a myenteric plexus (Wallace et al.2005). Based on the expression of serotonin (5HT), calcium-binding proteins—calretinin (CRT) and calbindin (CB)—andenzymes—neuronal nitric oxide synthase (nNOS) and cholineacetyltransferase (ChAT)—five neuronal subpopulations havebeen identified to date, i.e., a serotonergic, a nitrergicnoncholinergic, a nitrergic cholinergic and two cholinergicnonnitrergic subpopulations (Uyttebroek et al. 2010). Otherstudies have examined the expression of several neuropeptides,including vasoactive intestinal peptide (VIP) and pituitary ade-nylate cyclase-activating peptide (PACAP), in the zebrafish ENSbut their presence was detected in neuronal fibers, mainly invaricosities and not in neuronal somata (Olsson et al. 2008;Uyttebroek et al. 2010). These results have hampered the char-acterization of the neuronal subpopulations in the zebrafishENS. To overcome this lack of neuropeptide immunoreactivity(IR) in somata of enteric neurons, GI tissue has been pretreatedin vitro with colchicine before immunostaining in mammalianspecies (e.g., mice, rat, horse and guinea pig) (Chino et al. 2002;Freytag et al. 2008; Furness et al. 1989; Kapp et al. 2006; Mitsui2010; Portbury et al. 1995; Qu et al. 2008). Colchicine is a toxinknown to inhibit the polymerization of microtubules by bindingto tubulin (Terkeltaub 2009). It has been shown to preventaxoplasmic transport in neurons and, as a result, has been usedto increase the IR of neuropeptides in cell bodies of the periph-eral nervous system, including the ENS (Ekblad et al. 1996).
GI motility requires the interaction of enteric neurons,smooth muscle cells and interstitial cells of Cajal (ICC).ICC are mesenchymal cells distributed in specific locationswithin the tunica muscularis of the GI tract. Their presencehas been reported in the gut of a wide variety of mammalianand avian species. ICC serve as electrical pacemakers in thatthey generate and actively propagate spontaneous electricalactivity, i.e., slow waves, in the GI tract. In most vertebratespecies, ICC have been identified by the expression of thereceptor tyrosine kinase, kit (Burns et al. 2009; Ward et al.2004). Anoctamin 1 (Ano1) has been recently described as aselective ICC marker in the human and murine GI tract(Gomez-Pinilla et al. 2009). Ano1 belongs to the anoctaminfamily of chloride transporters and has been suggested toregulate ICC proliferation and to produce calcium-activatedcurrents generating ICC slow waves (Hwang et al. 2009;Sanders et al. 2011; Stanich et al. 2011). The presence ofICC in the zebrafish gut is still under debate (Burns et al.2009), though putative ICC-like cells expressing kit havebeen described in the zebrafish intestine (Ball et al. 2012;Rich et al. 2007).
The first main aim of the present study was to extendthe neurochemical characterization of enteric neuronal
subpopulations by revealing the expression of several neu-ropeptides. To this end, we used the in vitro colchicine pre-treatment method of Furness et al. (1989), which was appliedfor the first time on fish intestine. Additionally, we investigat-ed the spatiotemporal expression of these neuropeptidesthroughout the development of the zebrafish ENS, in particu-lar in embryos at 3 days post-fertilization (dpf; first entericneuronal progenitors start to differentiate), 4 dpf (neuronalprogenitors have colonized the entire GI tract) and 5 dpf (onsetof feeding). Our second aim was to examine the distributionand expression of Ano1 in both embryonic and adult zebrafishintestine to reveal the presence of ICC in the zebrafishintestine.
Materials and methods
Tissue preparation
Zebrafish were purchased from a local supplier and kept at theLaboratory of Human Anatomy& Embryology (University ofAntwerp). Zebrafish maintenance and breeding wereperformed as previously described (Matthews et al. 2002;Westerfield 2007). Eggs were collected after spawning andfertilization (0 dpf) and transferred to small containers con-taining embryo medium (0.99 mM MgSO4.7H2O, 0.15 mMKH2PO4, 0.04 mM Na2HPO4.2H2O, 0.50 mM KCl,14.97 mM NaCl, 7.14 mM NaHCO3 in dH2O) supplementedwith 0.003 % 1-phenyl-2-thiourea (Sigma-Aldrich, St. Louis,MO, USA; #P5272) to inhibit melanin pigment formation.The embryos were kept at 28.5 °C and staged by age andmorphological criteria (Kimmel et al. 1995). All experimentalprocedures were approved by the ethical committee of theUniversity of Antwerp (ECD2009-34).
Embryos were collected at 3, 4 and 5 dpf and euthanized with0.04 % MS222 (ethyl-m-aminobenzoate methanesulphonate;Western Chemical, Ferndale, WA, USA) in embryo medium.They were fixed in 4 % paraformaldehyde (PFA) in 10 mMphosphate-buffered saline (PBS; pH 7.4) for 1 h (3 and 4 dpf) or4 h (5 dpf) at room temperature, rinsed in PBS containing 0.5 %Triton X-100 (Sigma-Aldrich; #T8787) and stored at 4 °C inPBS containing 0.01 % sodium azide until further use.
Adult zebrafish (6–8 months old) were euthanized in0.04 % MS222 and decapitated. Next, the entire intestinewas dissected out. The intestine was opened along the mes-enteric border and pinned out in a Sylgard-lined Petri dishfilled with 4 % PFA in PBS for 2 h at room temperature. Afterfixation, the whole mounts were washed in PBS and furtherprocessed for immunohistochemical staining as previouslydescribed (Uyttebroek et al. 2010). To enhance IR for neuro-peptides in the neuronal somata, some whole mounts were,prior to fixation, maintained in organotypic culture inDulbecco’s Modified Eagle’s Medium supplemented with
356 Cell Tissue Res (2013) 354:355–370
Author's personal copy

Ham’s F12 (Gibco, Life Technologies, Paisley, UK; #31330-038), 10 % fetal calf serum (Gibco; #16000-044), 10 mg/mLantibiotic antimycotic (Sigma-Aldrich; #A5955), 2.5 μg/mLamphotericin B (Sigma-Aldrich; #A2942), 50 μg/mL genta-micin (Sigma-Aldrich; #48755), 1 μM nifedipine (Sigma-Aldrich; #7634), and the appropriate concentration of colchi-cine (Sigma-Aldrich; #C9754) (Furness et al. 1989).Furthermore, to optimize the colchicine pretreatment forzebrafish intestine, we tested different conditions of threeparameters, i.e., incubation time, incubation temperature andconcentration of colchicine (Table 1). For each change of aparameter, and for each neuropeptide, four whole mountswere immunostained. Following pretreatment with the colchi-cine solution, the tissues were fixed and further processed forimmunohistochemical staining.
In addition, the intestine of some animals was isolated, fixedand further processed by embedding in Tissue-Tek OCTCompound (Sakura Finetek, Zoeterwoude, The Netherlands)and cryosectioning at 10 μm, as previously described(Uyttebroek et al. 2010).
Immunohistochemistry
All antibodies and their dilutions are listed in Table 2. Thespecificity of most of the antibodies had been determined inprevious studies performed in zebrafish (Uyttebroek et al.2010). Since neuropeptides are small evolutionarily conservedmolecules, anti-neuropeptide antibodies are often able to spe-cifically cross-react in different species (Holmgren and Jensen2001). In the present study, we used an anti-calcitonin gene-related peptide (CGRP) antibody directed against rat αCGRP.Antibodies directed against the same epitope of rat αCGRPhave previously been used to detect CGRP in the intestine ofseveral fish species, including zebrafish (Dezfuli et al. 2002;Olsson et al. 2008; Vigliano et al. 2011). We also used an anti-galanin (GAL) antibody directed against unconjugated por-cine GAL, which has been used to detect GAL expression inthe GI tract of several fish species (Bosi et al. 2004, 2006,2007). A similar anti-GAL antibody has been previously usedin a study of the zebrafish brain (Castro et al. 2006). The anti-substance P (SP) antibody we used, just like all commercialanti-SP antisera, was directed against a synthetic peptide. Thisantiserum has been previously used in several vertebrate spe-cies, including several fish species (Bosi et al. 2005; Dezfuli
et al. 2002; Jensen and Holmgren 1985; Jensen et al. 1987) todetect SP in enteric nerve fibers and neurons. Since cross-reactivity with the tachykinergic neuropeptide neurokinin A(NKA) cannot be ruled out (Karila et al. 1998), immunostain-ing resulting from incubation with the anti-SP antibody, wastermed tachykinin IR. Ano1 was detected by a commercialantibody directed against a synthetic peptide of human Ano1.To date, this antibody has only been used on mammalian tissues(Gomez-Pinilla et al. 2009). Zebrafish Ano1 (NP_001155062.1)exhibits a protein sequence homology of 63 % with humanAno1 (NP_060513.5). Furthermore, we used two commerciallyavailable antibodies, both directed against a synthetic peptidecorresponding to human kit. Both antibodies have previouslybeen used to locate ICC in zebrafish intestine (Ball et al. 2012;Rich et al. 2007). The specificity of all these antibodies wasdeterminedwith preabsorption tests, revealing the absence of IR.Preabsorption of each anti-VIP antibody with 20 μg/ml synthet-ic VIP (Sigma-Aldrich) invariably eliminated IR, whereaspreabsorption with 20 μg/ml bovine thyroglobulin (AbD;Serotec, Oxford, UK) had no effect (Uyttebroek et al. 2010).Preabsorption of the anti-PACAP antibody with synthetic VIPdid not influence the immunolocalization of PACAP.Furthermore, the specificity of the known sequences of epitopeswas verifiedwith the BLAST program (http://blast.ncbi.nlm.nih.gov/Blast.cgi).
All immunolabelling experiments occurred at room temper-ature. Double-staining on whole embryos and whole mounts ofadult zebrafish intestines were performed as previously de-scribed (Uyttebroek et al. 2010). The antigens were simulta-neously detected by means of indirect immunofluorescenceusing primary antisera raised in different species. Briefly, em-bryos were rinsed three times with double-distilled water for1 h and subsequently immersed for 1 h in a blocking solution[PBS containing 0.5 % Triton X-100, 0.2 % bovine serumalbumin (BSA; Jackson Immunoresearch Laboratories, WestGrove, PA, USA), 1 % dimethyl sulphoxide, 0.02 % sodiumazide, and 5 % normal horse serum (NHS; JacksonImmunoresearch Laboratories)]. The embryos were incubatedovernight with the primary antibodies diluted in the blockingsolution. Next, the embryos were rinsed and immersed over-night in the appropriate secondary antibodies diluted in theblocking solution. Finally, the embryos were rinsed and theirintestines were dissected out and embedded in a glycerol-PBSsolution, Citifluor AF1 (Citifluor, London, UK).
The whole mounts of the adult zebrafish intestines wererinsed in PBS and incubated in PBS containing 50 mMglycine. Next, the tissues were transferred to PBS containing5 % NHS, 5 % BSA and 5 % Triton X-100 before incubationfor 18 h in a mix of two primary antibodies diluted in PBScontaining 1 % NHS, 0.1 % BSA and 1 % Triton X-100.After rinsing with PBS, the tissues were immersed for 2 h inthe appropriate secondary antibodies diluted in PBS. Afterwashing in PBS, the tissues were mounted in CitiFluorTM
Table 1 Parameters tested to optimize conditions for the in vitro col-chicine pretreatment
Parameter Conditions
Incubation temperature 28 °C, 37 °C
Concentration of colchicine 0.5 mg/mL, 1 mg/mL
Incubation time 4 h, 8 h, 10 h 12 h, 16 h, 24 h
Cell Tissue Res (2013) 354:355–370 357
Author's personal copy

AF1 (CitiFluor). A sequential protocol was used for thetriple-labeling experiments in which primary antisera raisedin different species were used. In the first step, two antigenswere simultaneously detected using standard fluorochrome-labeled secondary antibodies, while in a second step, thethird antigen was detected using a biotinylated secondaryantibody and a fluorochrome-conjugated streptavidin.Simultaneous detection of three antigens using primaryantisera raised in different species, produced less IR com-pared with the results of single-labeling experiments, whilethe results of the described sequential method did not differfrom those of the single-labeling procedures. The observeddifference is probably due to steric hindrance occurringduring the incubation step with primary or secondaryantisera. The double-labeling experiments using primaryantisera raised in the same species were performed using asequential immunostaining procedure as described byNegoescu et al. (1994), in which the first antibody is visual-ized with a fluorochrome-conjugated polyclonal monovalent
Fab fragment and the second with a standard fluorophore-labeled secondary antibody.
Cryosections were processed and immunostained as pre-viously described (Uyttebroek et al. 2010). Briefly, sectionswere dried at 37 °C, washed in PBS and pretreated with PBScontaining 10 % NHS, 0.1 % BSA, 0.5 % Triton X-100,0.01 % sodium azide (PBS*) and 0.05 % thimerosal, beforeincubation for 18 h in primary antibodies diluted in PBS*.Next, the sections were washed in PBS and visualization wasperformed using the appropriate secondary antibodies dilut-ed in PBS. After rinsing with PBS and staining of the nucleiwith DAPI nucleic acid stain (Invitrogen, Carlsbad, CA,USA), they were mounted in CitiFluorTM AF1.
The specificity of the secondary antibodies was tested byomitting the primary antibodies separately or together. Nonon-specific staining was observed, except in the embryonicand larval intestine where aspecific stained blood cells wereoccasionally observed and aspecific-stained fat cells wereseen at the distal end of the DI. Interference control stainings
Table 2 List of antisera and their respective dilutions used for immunohistochemistry
Primary antisera
Antigen Host Dilution Source
Serotonin (5HT) Rabbit 1/10,000 Immunostar, Hudson, WI, USA (20080)
Serotonin (5HT) Goat 1/2,500 Immunostar (20079)
Acetylated tubulin (α-tub) Mouse 1/2,000 Sigma-Aldrich (T6793)
Anoctamin 1 (Ano1) Rabbit 1/1,000 Abcam, Cambridge, U.K. (ab53212)
c-kit Rabbit 1/50 Calbiochem, Merck Millipore, Nothingham, U.K. (961–976)
c-kit Rabbit 1/50 Abcam (ab16832)
Calbindin (CB) Rabbit 1/2,000 SWant, Bellinzona, Switzerland (CB-38a)
Calcitonin gene-related peptide (CGRP) Mouse 1/,5000 Sigma-Aldrich (C7113)
Choline acetyltransferase (ChAT) Goat 1/100 Chemicon, Temecula, CA, USA (AB114P)
Calretinin (CRT) Rabbit 1/2,000 Chemicon (AB5054)
Calretinin (CRT) Goat 1/2,000 Swant (CG1)
Galanin (GAL) Rabbit 1/2,000 Biogenesis
Neuron-specific HuC/HuD proteins (Hu) Mouse 1/500 Molecular Probes, Eugene, OR, USA (A-21271)
Neuronal nitric oxide synthase (nNOS) Rabbit 1/100 Santa Cruz Biotechnology, Santa Cruz, CA, USA (sc-1025)
Substance P (SP) Rabbit 1/400 Sigma-Aldrich (S1542)
Pituitary adenylate cyclase-activating peptide (PACAP) Rabbit 1/5,000 Bachem, Torrance, CA, USA (T-4473)
Vasoactive intestinal peptide (VIP) Rabbit 1/500 Immunostar (20077)
Vasoactive intestinal peptide (VIP) Goat 1/100 Santa Cruz Biotechnology (sc-7841)
Secondary antisera and streptavidin complexes
Complex Dilution Company
Donkey anti-rabbit Cy3 1/800 Jackson Immunoresearch Laboratories, West Grove, PA, USADonkey anti-mouse DL488 1/200
Donkey anti-goat Cy3 1/800
Donkey anti-goat DL488 1/200
Donkey anti-mouse biotin 1/200
Donkey anti-rabbit Fab 1/50
Donkey anti-rabbit Fab Cy3 1/200
Cy5-conjugated Streptavidin 1/1,000
358 Cell Tissue Res (2013) 354:355–370
Author's personal copy

were also performed as described (Negoescu et al. 1994),showing no linking of secondary antibodies with primaryantibodies used in a previous step.
All preparations were analyzed by confocal microscopyon a laser scanning microscope LSM510 META equippedwith an argon-ion and helium-neon laser system (Carl ZeissMicroimaging, Göttingen, Germany). Average intensity pro-jection images of stacks along the z-axis were made.Contrast and brightness were adjusted in LSM ImageBrowser Release 4.2 (Carl Zeiss Microimaging) and mon-tages were made using Adobe Photoshop 7.0 (AdobeSystems, San Jose, CA, USA). Coexpression of the differentmarkers was analyzed by careful examination of all opticalsections collected in multiple-wavelength illumination modeand by Pearson’s Correlation tests using the VolocityQuantitation software (Perkin Elmer, Waltham, MA, USA).
For the quantitative analysis in adult zebrafish, the immu-noreactive nerve cell bodies were counted as previouslydescribed by Uyttebroek et al. (2010) in the three functionalregions of the intestine: the proximal (PI), mid (MI) anddistal intestine (DI) (Holmberg et al. 2003; Ng et al. 2005).The number of immunoreactive neurons was calculated in atleast four different animals and the proportional expressionof the neurochemical markers was estimated based on thenumber of neurons expressing the pan-neuronal marker Hu(neuron-specific HuC/HuD proteins) present in every visualframe. In addition, the proportional coexpression of differentmarkers was determined. The results are expressed as meanvalues ± standard errors of the mean and were analyzed byANOVA followed by a Student’s–Newman–Keuls test whenappropriate. Significance was assumed at P<0.05.
Results
Colchicine
Pretreatment with colchicine did not result in significantlyimproved IR of neuronal somata. The best results wereobtained with an in vitro pretreatment at 28 °C for 16 h in aconcentration of 1 mg/mL colchicine. Lower incubation timesand colchicine concentrations did not improve the results,whereas higher incubation times and/or temperatures resultedin very brittle tissue that was not suitable for immunostaining.Furthermore, colchicine pretreatment only enhanced the IR ofVIP and PACAP in somata of enteric neurons (Fig. 1a–b),whereas GAL, CGRP and SP were never detected in neuronalsomata—with or without colchicine pretreatment. These threeneuropeptides were only detected in enteric nerve fibers.Moreover, pretreatment of whole mounts with colchicine ledto the absence of IR for ChAT in neuronal somata as comparedwith untreated immunostained preparations. The proportion ofenteric neurons expressing the calcium-binding proteins CB
and CRT was also significantly decreased compared to previ-ous reports (35±3 and 38±5 % for colchicine-treated and43±6 and 45±7 % for untreated tissue) and were thereforeexcluded from proportional analysis (Uyttebroek et al. 2010).On the other hand, colocalization of nNOS or 5HT with thepan-neuronal marker Hu in pretreated tissue showed propor-tions and distributions of immunostained neurons that weresimilar to previously reported results (Uyttebroek et al. 2010):27±3 % were nitrergic and 18±1 % serotonergic. No signifi-cant variation was observed in the proportional distribution ofthe nitrergic neurons among the three intestinal regions, whilethat of serotonergic neurons significantly decreased in the distalpart of the intestine as compared to the proximal intestine (PI:23±2 %; MI: 18±1 %; DI: 13±2 %).
In general, no regional differences in the expression of thedifferent neuropeptides was observed, except for GAL (seebelow). Furthermore, as reported in a previous study(Uyttebroek et al. 2010), colocalization stainings in pre- anduntreated tissue revealed that VIP and PACAP werecolocalized in the same neuronal cell bodies and nerve fibers(Fig. 1c; i.e., colocalization of VIP and PACAP was 100 %).Colocalization of VIP/PACAPwith Hu showed that 22±1% ofthe enteric neurons expressed VIP/PACAP (VIP: PI 23±2 %;MI 20±1 %; DI 22±2 % // PACAP: PI 20±2 %; MI 22±2 %;DI 22±2 %). We named these neurons, VIPergic neurons.
VIP and PACAP
Coexpression of VIP/PACAPwith other neurochemical markerswas performed using the triple-immunolabelling method includ-ing the detection of Hu. Of the VIPergic neurons, 68±4 %expressed nNOS IR, while VIP/PACAP IR was observed in44±3 % of the nitrergic neurons (Fig. 2a). The proportion ofenteric neurons expressing nNOS/VIP/PACAP amounted to13±6 % of the total neuronal population. 5HT IR was found in16±2 % of the VIP/PACAP-positive neurons, while 23±4 % ofthe serotonergic neurons exhibited VIP/PACAP IR (Fig. 2b, c).Consequently, 4±1 % of the total neuronal population expressed5HT/VIP/PACAP. In the untreated whole mounts, VIP/PACAPIR was not detected in neuronal somata but only in nerve fiberspresent in the lamina propria and tunica muscularis of the threeintestinal regions. Some nerve fibers and neuronal cell bodiesexpressing CB and CRT coexpressed VIP/PACAP. Double-labelling of VIP/PACAP with ChAT in untreated tissue revealedno expression of VIP/PACAP in cholinergic nerve fibers.
Galanin
GAL IR was observed in myenteric nerve fibers in the tunicamuscularis. The number of GAL-expressing neuronal fiberswas clearly highest in the PI and lowest in the DI (Fig. 3a–c).No colocalization was observed with VIP/PACAP, 5HT,CRT/CB, nNOS, CGRP, or SP, while coexpression of GAL
Cell Tissue Res (2013) 354:355–370 359
Author's personal copy

and ChAT was detected in a few nerve fibers (Fig. 4a–c).Furthermore, GAL IR was also found in some epithelial cellsof the MI and DI region.
CGRP
In the tunica muscularis, CGRP IR was exclusively found inmyenteric nerve fibers throughout the whole intestinal wall.Some of these CGRP-positive nerve fibers coexpressedCRT/CB but not all CRT/CB immunoreactive fibersexpressed CGRP (Fig. 5a). Moreover, a partial overlap ofCGRP IR with nNOS, tachykinin, or ChAT IR in nerve fiberswas also demonstrated (Fig. 5b–d). A few CGRP immuno-reactive nerve fibers coexpressed VIP/PACAP and viceversa but no colocalization between CGRP immunoreactivefibers and 5HT was observed (Fig. 5e).
Substance P
Tachykinin IR was detected in nerve fibers located in thelamina propria and tunica muscularis of all three intestinal
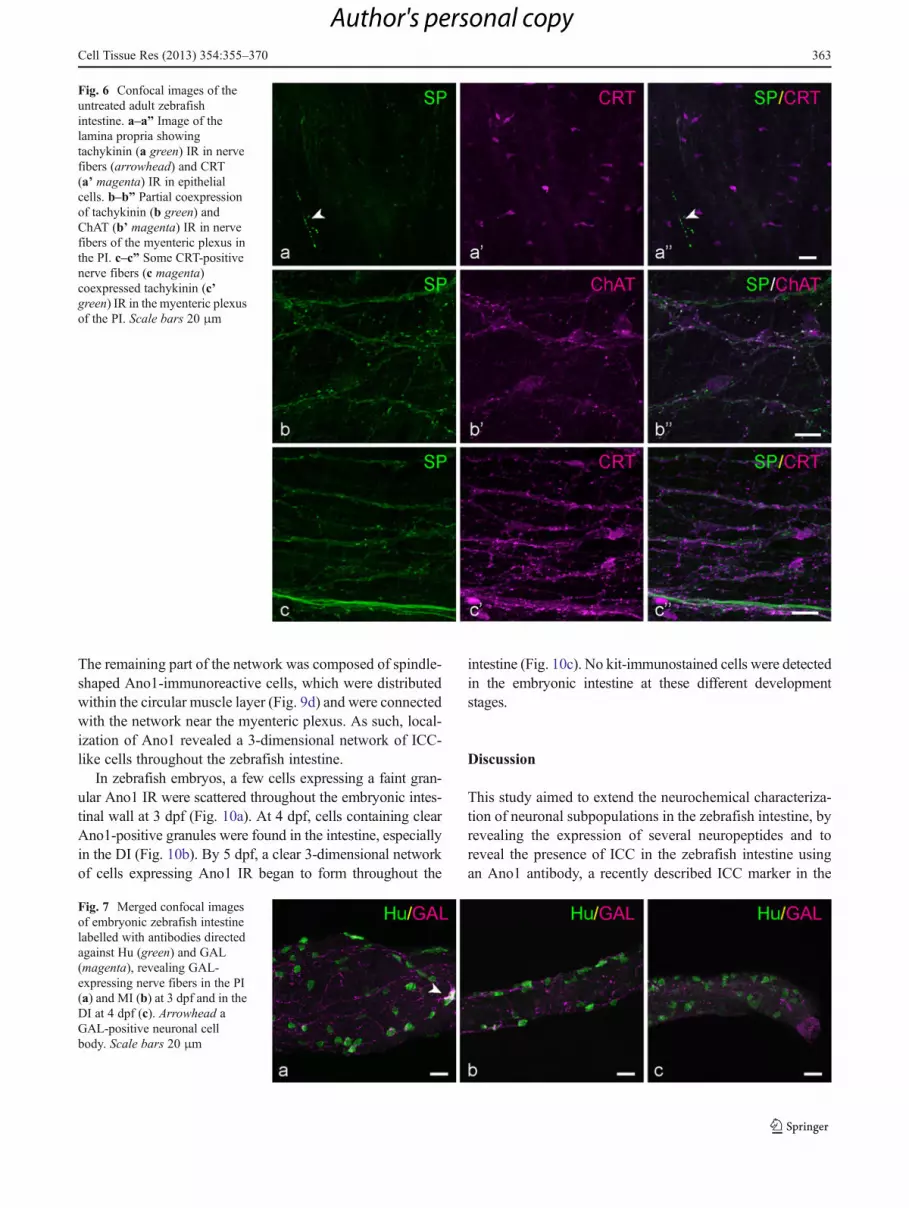
regions (Fig. 6a). Visual inspection of the preparations al-ready clearly showed that the tachykinergic nerve fibersoutnumber the CGRP immunoreactive fibers. As mentionedabove, some tachykinergic nerve fibers also expressedCGRP (Fig. 5c). Furthermore, a partial overlap of tachykininIR with ChAT and CRT/CB IR was observed (Fig. 6b, c).
Embryonic intestine
At 3 dpf, GAL was clearly expressed in nerve fibers andneuronal varicosities in the PI and MI (Fig. 7a, b). At 4 dpf,GAL-positive fibers were also observed in the DI (Fig. 7c).The number of GAL immunoreactive nerve fibers increasedwith time and from 4 dpf onwards, no obvious differences inthe number and the distribution of GAL-positive fibers wereobserved in the embryonic intestine. GAL immunoreactivesomata were occasionally detected, especially in the PI at 5dpf. Additionally, at 5 dpf, GAL-positive epithelial cells wereobserved in the second part of the MI and in the DI. At 3 dpf,tachykinin and CGRP IR were detected in a few neuronalvaricosities of the DI (Fig. 8a). At 4 dpf, tachykinergic and
Fig. 1 Confocal images of adultzebrafish intestine pretreated withcolchicine. Doubleimmunolabelling for Hu (a green),VIP (a’ magenta) or for Hu (bgreen) PACAP (b’ magenta)shows VIP (a”) and PACAP-immunoreactive (b”) varicositiesand neuronal cell bodies in the MI(a–a”) and PI (b–b”), respectively.c Coexpression of VIP (c green)and PACAP (c’ magenta) inneuronal cell bodies expressingHu (c” blue) in the MI. Scale bars20 μm
360 Cell Tissue Res (2013) 354:355–370
Author's personal copy

CGRP immunoreactive nerve fibers were also found in the PIand MI (Fig. 8b). The number of tachykinergic and CGRPimmunoreactive varicosities in the embryonic intestine in-creased with time and no regional differences were observedat 4 and 5 dpf.
Interstitial cells of Cajal
Using the antibodies directed against kit, immunopositivecells, were observed, in the epithelium and lamina propria ofthe adult zebrafish intestine. These cells appeared to be mastcells. A single immunostained cell containing granular kit
IR, was occasionally detected in the circular muscle layer ornear the myenteric plexus. No improvement in staining re-sults was obtained after the staining protocol was adapted tothat previously described by Ball et al. (2012) and Rich et al.(2007). So, no network of kit-expressing cells was observedin the tunica muscularis as previously reported (Ball et al.2012; Rich et al. 2007). As a result, no colocalizationimmunostainings with the anti-Ano1and anti-kit antibodieswere performed.
Cells displaying a granular IR for Ano1 were observed inthe adult zebrafish intestine (Fig. 9a). The distribution ofimmunostained cells did not differ among the three intestinal
Fig. 2 Confocal images of adultzebrafish intestine pretreated withcolchicine. a–a”Immunolocalization of nNOS(a green) and VIP (a’magenta) inthe PI. Arrowhead: nNOS/VIP-expressing neuronal cell body.b–b” Immunolocalization of 5HT(b green) and PACAP (b’magenta) in the PI. Arrowhead:5HT/PACAP-immunoreactiveneuronal cell body. c–c”Immunolocalization of 5HT(c green) and VIP (c’ magenta)in the PI. Arrowhead 5HT/VIP-immunoreactive neuronal cellbody. Scale bars 20 μm
Fig. 3 Confocal images of GALIR in the adult zebrafish intestinerevealing GAL-immunoreactivenerve fibers in the myentericplexus of the PI (a), MI (b) andDI (c). Scale bars 20 μm
Cell Tissue Res (2013) 354:355–370 361
Author's personal copy

regions. Ano1 IR revealed a network of ICC-like cells in twodistinct layers. Part of this network consisted in multipolarAno1-immunoreactive cells that were found between the
circular and longitudinal muscle layer, i.e., near themyenteric plexus, as clearly shown in colocalization studieswith the pan-neuronal markers Hu and α-tub (Fig. 9b, c).
Fig. 4 Confocal image showingcoexpression of GAL (a green)and ChAT (b magenta) in the PIof the adult zebrafish intestine.c Overlay. Scale bars 20 μm
Fig. 5 Confocal images showingcoexpression of CGRP withdifferent neurochemical markersin the PI of the adult zebrafish.a–a” CGRP (a green) partiallycoexpressed with CRT (a’magenta) in nerve fibers. b–b”Partial coexpression of CGRP(b green) and nNOS (b’magenta)in nerve fibers. c–c”Coexpression of CGRP (c green)and tachykinin (c’ magenta) innerve fibers. d–d” Somecoexpression of CGRP (d green)and ChAT (d’ magenta) in nervefibers. e–e” No coexpression ofCGRP (e green) and 5HT(e’ magenta) was observed innerve fibers. Scale bars 20 μm
362 Cell Tissue Res (2013) 354:355–370
Author's personal copy
The remaining part of the network was composed of spindle-shaped Ano1-immunoreactive cells, which were distributedwithin the circular muscle layer (Fig. 9d) and were connectedwith the network near the myenteric plexus. As such, local-ization of Ano1 revealed a 3-dimensional network of ICC-like cells throughout the zebrafish intestine.
In zebrafish embryos, a few cells expressing a faint gran-ular Ano1 IR were scattered throughout the embryonic intes-tinal wall at 3 dpf (Fig. 10a). At 4 dpf, cells containing clearAno1-positive granules were found in the intestine, especiallyin the DI (Fig. 10b). By 5 dpf, a clear 3-dimensional networkof cells expressing Ano1 IR began to form throughout the
intestine (Fig. 10c). No kit-immunostained cells were detectedin the embryonic intestine at these different developmentstages.
Discussion
This study aimed to extend the neurochemical characteriza-tion of neuronal subpopulations in the zebrafish intestine, byrevealing the expression of several neuropeptides and toreveal the presence of ICC in the zebrafish intestine usingan Ano1 antibody, a recently described ICC marker in the
Fig. 6 Confocal images of theuntreated adult zebrafishintestine. a–a” Image of thelamina propria showingtachykinin (a green) IR in nervefibers (arrowhead) and CRT(a’ magenta) IR in epithelialcells. b–b” Partial coexpressionof tachykinin (b green) andChAT (b’ magenta) IR in nervefibers of the myenteric plexus inthe PI. c–c” Some CRT-positivenerve fibers (c magenta)coexpressed tachykinin (c’green) IR in the myenteric plexusof the PI. Scale bars 20 μm
Fig. 7 Merged confocal imagesof embryonic zebrafish intestinelabelled with antibodies directedagainst Hu (green) and GAL(magenta), revealing GAL-expressing nerve fibers in the PI(a) and MI (b) at 3 dpf and in theDI at 4 dpf (c). Arrowhead aGAL-positive neuronal cellbody. Scale bars 20 μm
Cell Tissue Res (2013) 354:355–370 363
Author's personal copy

mammalian ENS. Furthermore, the putative beneficial effectof colchicine pretreatment on neuropeptide IR was exam-ined. Colchicine pretreatment was found to be less beneficial
in zebrafish than in mammalian tissues (Chino et al. 2002;Freytag et al. 2008; Kapp et al. 2006; Mitsui 2010; Portburyet al. 1995; Qu et al. 2008), since it only enhanced VIP andPACAP IR in neuronal cell bodies but not CGRP, tachykininand GAL IR. IR for these markers was only observed innerve fibers. In a previous study in rainbow trout, colchicinewas used to enhance CGRP IR in enteric neuronal somata(Ohtani et al. 1989). In this study, animals were treatedin vivo with intraperitoneally administered colchicine andno immunoreactive somata were observed in the GI tract ofthe rainbow trout. In the same study, similar results wereobtained in other colchicine-treated non-mammalian species,while CGRP IR was observed in enteric neuronal somata ofcolchicine-treated mammalian species. In another study,toads were treated in vivo with colchicine that was injectedinto the iliac lymph sac (Li et al. 1993). VIP IR but not SP IR,was found to be enhanced in neuronal somata in the GI tract
Fig. 9 a–a” Confocal image ofthe tunica muscularis of the adultzebrafish intestine showing α-tub (a green) and Ano1 (a’magenta) IR. b–b” Cryosectionof the wall of the adult zebrafishintestine labelled with antibodiesdirected against α-tub (b green)and Ano1 (b’ magenta) (nucleistained with DAPI (b” blue))revealing two interconnectedlayers of ICC-like cells located inthe circular muscle layer (M) andaround the myenteric plexus. L:lamina propria. cConfocal imageof the myenteric plexus revealinga network of multipolar Ano1-expressing ICC-like cells. dConfocal image of the circularmuscle layer displaying a loosenetwork of bipolar Ano1-positive ICC-like cellsinterspersed between the smoothmuscle cells. Scale bars 20 μm
Fig. 8 Confocal images of embryonic zebrafish intestine. a CGRP IRin neuronal varicosities in the DI at 3 dpf; * aspecific-stained fat cells atthe distal end of the DI. b Tachykinin IR in nerve fibers of the MI at 4dpf. Arrowheads aspecific-stained blood cells. Scale bars 20 μm
364 Cell Tissue Res (2013) 354:355–370
Author's personal copy

of treated toads. Although we used the in vitro colchicinemethod described by Furness et al. (1989), our results arecomparable to these previously obtained data in the intestineof non-mammalian species (Li et al. 1993; Ohtani et al.1989).
Consistent with previous studies, the present study showedthat VIP and PACAP, were coexpressed in enteric neuronal cellbodies of the zebrafish intestine and that 22±1 % of the totalneuronal population expressed VIP and PACAP (Olsson et al.2008; Uyttebroek et al. 2010). This finding is in agreement withthe results of previous studies revealing VIPergic neurons in themyenteric plexus of the small intestine in rat (20–25%; Johnsonet al. 1998), mouse (26%; Qu et al. 2008) and guinea pig (23%;Furness 2006) and of the human colon (22 %; Wattchow et al.1997). Colocalization stainings revealed that a major part ofthese VIPergic neurons in zebrafish was nitrergic (±13 % ofthe total neuronal population), while a minor part was seroto-nergic (±4% of the total neuronal population). Based on the datapreviously and presently observed in zebrafish that nitrergicneurons express CB and CRT (Uyttebroek et al. 2010), that5HT is not coexpressed with CB, CRT, nNOS, or ChAT(Uyttebroek et al. 2010) and that no coexpression ofVIP/PACAP with ChAT is observed, three neuronal subpopula-tions are deduced and added to the list of subpopulations ob-served in our previous work (Uyttebroek et al. 2010): a nitrergicnoncholinergic VIPergic (CB/CRT/nNOS/VIP/PACAP) sub-population, a serotonergicVIPergic (5HT/VIP/PACAP) subpop-ulation and a VIPergic (VIP/PACAP) subpopulation (Table 3).
VIP and PACAP, which belong to the secretin superfamilyof regulatory peptides, are highly conserved during evolution.In zebrafish, as in other teleost species, VIP and PACAP areencoded by duplicate genes (Roch et al. 2009) and two sepa-rated isoforms of PACAP-38 have been cloned (Wang et al.2003). Previous studies have reported on the coexpression ofVIP and PACAP in enteric neurons in several non-mammalianandmammalian species, such as the spiny dogfish, Atlantic codand trout (Olsson and Holmgren 1994; Olsson and Karila1995), axolotl (Badawy and Reinecke 2003), African clawedfrog (Holmberg et al. 2001; Olsson 2002), chicken (Salvi et al.2000; Sundler et al. 1992), duck (Mirabella et al. 2002) andpigeon (Mirabella et al. 2000) and also in the mouse (Grider
2003), rat (Hannibal et al. 1998) and human GI tract (Shenet al. 1992; Sundler et al. 1992), as well as in the guinea pigcaecum (McConalogue et al. 1995). Several of these studiesalso revealed that a major part of the VIP/PACAP-expressingneurons were nitrergic, suggesting that these neurons areinvolved in the inhibitory control of GI motility (Badawyand Reinecke 2003; Grider 2003; Hannibal et al. 1998;Holmberg et al. 2001; Mirabella et al. 2000, 2002; Olsson2002; Olsson and Karila 1995). The coexpression of VIP,PACAP and nNOS is a neurochemical property that is char-acteristic of inhibitory motor neurons in the mammalian GItract, while noncholinergic, nonnitrergic neurons expressingVIP and PACAP are secretomotor neurons (Furness 2012;Grider 2003; Holzer et al. 2001). This suggests that theCB/CRT/nNOS/VIP/PACAP-expressing neurons in thezebrafish intestine are inhibitory motor neurons.
Tachykinin, CGRP and GAL IR were only observed inenteric nerve fibers. Given this pattern of IR, we cannotexclude the possibility that some of these immunoreactivenerve fibers are from an extrinsic origin. Previous studies inzebrafish and Atlantic cod have reported on the expression ofVIP, PACAP, 5HT, CGRP, GAL and SP in the vagal nerve(Jensen et al. 1993a; Karila et al. 1993; Olsson and Holmgren
Fig. 10 Confocal images of theembryonic zebrafish MI labelledwith antibodies against Hu(green) and Ano1 (magenta) at 3dpf (a), 4 dpf (b) and 5 dpf (c).Scale bars 20 μm
Table 3 Summary of the known neuronal subpopulations detected inthe zebrafish intestine
Neuronal subpopulation Proportional expression
5HT ±14 %a
5HT/VIP/PACAP ±4 %
CB/CRT/ChAT/nNOS ±3 %a
CB/CRT/nNOS ±2 %a
CB/CRT/nNOS/VIP/PACAP ±13 %
CB/CRT/ChAT ±35 %a
VIP/PACAP ±5 %
ChAT ±5 %a
a Previously observed (Uyttebroek et al. 2010)
5HT serotonin, CB calbindin, ChAT choline acetyl transferase, CRTcalretinin, nNOS neuronal nitric oxide synthase, PACAP pituitary ade-nylate cyclase-activating peptide, VIP vasoactive intestinal peptide
Cell Tissue Res (2013) 354:355–370 365
Author's personal copy

1994; Olsson et al. 2008; Shahbazi et al. 1998; Uyttebroeket al. 2010). Colocalization stainings showed thattachykinergic and CGRP-positive nerve fibers partiallycoexpressed several other markers, while no colocalizationwas observed in GAL immunoreactive nerve fibers, exceptfor a few GAL-positive fibers being cholinergic. These resultslead us to suggest that the zebrafish ENS contains anoncholinergic, nonnitrergic subpopulation and a cholinergicsubpopulation of enteric neurons expressing GAL. However,it cannot be excluded that some of these fibers are extrinsic inorigin. Furthermore, GAL- and CGRP-positive fibers werefound in the tunica muscularis, while tachykinergic fiberswere present in the lamina propria and the tunica muscularis,indicating that the GAL- and CGRP-positive fibers predomi-nantly innervate the enteric smooth muscle, whiletachykinergic-positive fibers innervate the enteric smoothmuscle, lamina propria and epithelium. No regional differ-ences in the distribution of tachykinergic and CGRP-positivenerve fibers were observed, while the number of GAL-positive fibers decreased from the PI to the DI.
The expression of GAL in the zebrafish ENS is consistentwith previous studies in other teleost species (Bosi et al. 2004,2005, 2006, 2007; Dezfuli et al. 2007; Karila and Holmgren1997; Karila et al. 1993) in which GAL IR was predominantlyfound in the proximal part of the GI tract. In the ENS offlatfishes, GAL and ChATwere found to be colocalized withinthe same neurons and it was suggested that GAL functions as acholinergic modulator in the teleost GI tract (Bosi et al. 2007).Studies in the Atlantic cod have suggested that GAL, whilehaving a contractile effect on GI smoothmuscle, is expressed ininterneurons rather than in motor neurons (Karila andHolmgren 1997). In the mammalian gut, GAL is predominant-ly found in noncholinergic secretomotor and vasodilator neu-rons (Brookes 2001; Holzer et al. 2001; Timmermans et al.1997). During embryonic development, GAL immunoreactivenerve fibers have already been observed in the PI and the MI at3 dpf and in all intestinal regions at 4 dpf. In the zebrafish,spontaneous regular gut motility is known to begin at 3.5 dpf(Kuhlman and Eisen 2007). Although these propagating con-tractions can be modulated by substances like nitric oxide andacetylcholine, the intrinsic and extrinsic innervation is notnecessary to initiate these contractions in zebrafish embryos(Holmberg et al. 2007). It is assumed that, apart frommaintaining the propagating contractions, neurotransmittersand neuropeptides perform additional functions in the embry-onic zebrafish intestine. In chicken embryo, it has been sug-gested that the GAL present in the ENS also acts as a differen-tiating or growth factor during embryonic development of thegut (Salvi et al. 1999).
CGRP, a 37 amino acid peptide, is highly conservedthroughout vertebrate evolution. The amino acid sequenceof zebrafish CGRP shares 76–81 % identity with humanCGRP (Martinez-Alvarez et al. 2008). The presence of
CGRP in enteric neurons has been reported in several teleostspecies including zebrafish (Bermudez et al. 2007; Dezfuliet al. 2002; Domeneghini et al. 2000; Ohtani et al. 1989;Olsson et al. 2008; Shahbazi et al. 1998; Vigliano et al.2011). In the teleost intestine, CGRP has been suggested toact as a vasodilator (Kågström and Holmgren 1998), or as atransmitter in the descending inhibitory nerve pathways(Shahbazi et al. 1998). In mammals, CGRP has been foundto be coexpressed with acetylcholine and tachykinins in in-trinsic sensory neurons (Furness 2012; Grider 2003; Qu et al.2008) and to act as a transmitter in intrinsic sensory pathwaysinvolved in peristalsis (Grider 1994). However, CGRP hasalso been found to be coexpressed with SP in nerve fibers ofextrinsic sensory neurons (Timmermans et al. 1997).Furthermore, in the guinea pig ileum, CGRP has also beendetected in secretomotor and viscerofugal neurons (Brookes2001; Holzer et al. 2001; Timmermans et al. 1997). CGRP-positive nerve fibers appeared first in the DI of the zebrafishembryo at 3 dpf and also in the PI andMI from 4 dpf onwards.This observation is not in complete agreement with the resultsof a previous study in which the first CGRP immunoreactivenerve fibers were detected at 4 dpf in all intestinal regions(Olsson et al. 2008), which may be due to differences instaging occuring between laboratories or to different experi-mental techniques. A study of 1-day-old Xenopus nerve–mus-cle cultures has indicated that a major function of CGRP at theembryonic stage is to potentiate synaptic development duringsynaptogenesis (Lu et al. 1993).
The tachykinergic neuropeptide SP has been previouslyreported in the ENS of several fish species (Bermudez et al.2007; Bjenning and Holmgren 1988; Bosi et al. 2005; Dezfuliet al. 2002; Domeneghini et al. 2000; Holmgren et al. 1982;Jensen and Holmgren 1985; Jensen et al. 1987, 1993a; Karilaet al. 1998; Pederzoli et al. 2004). SP isolated from teleost brainhas three substitutions in comparison with mammalian SP(Jensen and Conlon 1992). Studies in teleosts revealed thatSP acts as a vasoconstrictor of intestinal blood vessels(Kågström and Holmgren 1998) and is responsible for contrac-tions of GI smooth muscle (Johansson and Holmgren 2003;Jensen and Holmgren 1985; Jensen et al. 1993a, b; Kitazawaet al. 2012). In the Atlantic cod intestine, SP has been shown tobe expressed in cholinergic enteric neurons (Karila et al. 1998).In themammalian GI tract, SP has been predominantly found incholinergic neurons, i.e., excitatorymotor neurons, intrinsic andextrinsic sensory neurons and ascending interneurons (Brookes2001; Furness 2012; Holzer et al. 2001; Qu et al. 2008;Timmermans et al. 1997). Furthermore, SP is known to exertimmunostimulatory and pro-inflammatory effects in the mam-malian gut (Collins et al. 1997). Tachykinergic nerve fibersappeared first in the DI of the zebrafish embryo at 3 dpf andin the other intestinal regions at 4 dpf. Accordingly,tackykinergic fibers also appeared first in the distal part of thedeveloping gut in the sea bass. Moreover, it has been
366 Cell Tissue Res (2013) 354:355–370
Author's personal copy

hypothesized in the sea bass that SP, like VIP, is involved in thephysiological maturation of the epithelium (Pederzoli et al.2004). In zebrafish embryos, NKA-like immunoreactive nervefibers were first observed in the DI at 3 dpf and in all threeintestinal regions from 4 dpf onwards (Olsson et al. 2008),whereas in a previous study with zebrafish embryos, NKA-like IR was first encountered in nerve fibers of the PI at 2 dpf(Holmberg et al. 2004). The same authors also showed thatNKA was responsible for an increase in the frequency ofcontraction waves in the zebrafish intestine from 5 dpf(Holmberg et al. 2004).
Kit IR has been used extensively in other species to studyand visualize the ICC network in the GI tract. Several physio-logical functions have been suggested for ICC: (1) ICC arepacemakers generating electrical slow waves that organize thephasic contractile behavior of the GI tract; (2) ICC provide apropagation pathway to regenerate slow waves; (3) intramus-cular ICC (ICC-IM) participate in excitatory and inhibitoryneurotransmission to enteric smooth muscle; and (4) ICC-IMact as stretch sensors (Garcia-Lopez et al. 2009). In zebrafish, ithas been reported that kit IR reveals a network of ICC-like cellsin the myenteric plexus region of the adult zebrafish intestine(Ball et al. 2012; Rich et al. 2007). Two layers of kit-expressingcells were observed: a layer of multipolar cells at the level ofthe myenteric plexus and a layer of bipolar cells in the circularmuscle layer. Kit-positive cells first appeared in zebrafish lar-vae at 7 dpf, i.e., after the onset of feeding (Rich et al. 2007).We attempted to reproduce these immunofluorescent stainingsbut were unable to do so. However, it has recently beenreported that antibodies directed against Ano1 constitute a goodmarker for all classes of ICC in human and mouse intestine(Gomez-Pinilla et al. 2009). Moreover, the Ano1 antiserumwas shown to be a more selective marker of ICC than the kitantiserum, in that the former only labels ICC, while the latter isalso expressed onmast cells in the GI tract (Gomez-Pinilla et al.2009). Ano1 has also been suggested to be essential in thegeneration of slow waves (Hwang et al. 2009), while kitexpression is necessary for the postnatal development andproliferation of ICC (Klüppel et al. 1998). Recent researchhas indicated that Ano1 regulates ICC proliferation (Mazzoneet al. 2012; Stanich et al. 2011). In the present study, Ano1-positive cells formed a 3-dimensional network in the zebrafishintestine. A network of multipolar Ano1-positive cells wasobserved near the myenteric plexus. This is similar to the ICCof the myenteric plexus (ICC-MY) seen in mammals (Komuro2006). This network was interconnected with a network ofspindle-shaped Ano1 immunoreactive cells, which again issimilar to ICC-IM in mammals (Komuro 2006). In mammals,ICC-IM are interspersed between the smooth muscle cells ofthe circular muscle layer. Ano1 IR was first detected at 3 dpf asscattered points of IR throughout the embryonic intestine. At 4dpf, immunoreactive cells were observed mainly in the DI,while a network of Ano1-positive cells was clearly visible at
5 dpf. Based on these observations, we conclude that the anti-Ano1 antiserum, just like in mammals, is a selective marker ofICC-like cells in the zebrafish intestine. Furthermore, we hy-pothesized that ICC-like progenitor cells start to proliferate inthe embryonic intestine of the zebrafish at 3 dpf, when the firstenteric neuron progenitors start to differentiate and that the firstICC-like cells appear at 4 dpf when the entire GI tract iscolonized by neuron progenitors. Likewise in humans, the firstICC also appear when the entire embryonic gut is colonizedwith neuron progenitor cells (Wallace and Burns 2005).Furthermore, it has been suggested that, in zebrafish, ICC-likecells are involved in spontaneous regular gut motility of theembryonic intestine, because the appearance of these cells co-incides with the onset of the propagating contractions, asobserved in the embryonic mouse intestine (Burns et al.2009; Holmberg et al. 2003; Kuhlman and Eisen 2007;Torihashi et al. 1997).
In conclusion, we have distinguished three new neuronal sub-populations in the zebrafish GI tract: a nitrergic noncholinergicVIPergic subpopulation (CB/CRT/nNOS/VIP/PACAP), whichis likely to consist in inhibitory motor neurons; a serotonergicVIPergic subpopulation (5HT/VIP/PACAP); and a VIPergicsubpopulation (VIP/PACAP). The present results also indicatethat the Ano1 antiserum is a selective marker of ICC-like cells inthe zebrafish intestine revealing a 3-dimensional network. Ano1IR first appears in the zebrafish intestine at 3 dpf, which isindicative of the onset of proliferation of ICC-like cells. Finally,it is hypothesized that ICC-like cells mediate the spontaneousregular activity in the embryonic intestine.
Acknowledgments The authors would like to thank Mrs. E. Goeman,Mrs. M. Van Geel and Mr. D. De Rijck for their technical assistance.
References
Badawy G, Reinecke M (2003) Ontogeny of the VIP system in thegastro-intestinal tract of the axolotl, Ambystoma mexicanum:successive appearance of co-existing PACAP and NOS. AnatEmbryol 206:319–325
Ball ER, Matsuda MM, Dye L, Hoffmann V, Zerfas PM, Szarek E, RichA, Chitnis AB, Stratakis CA (2012) Ultra-structural identificationof interstitial cells of Cajal in the zebrafishDanio rerio. Cell TissueRes 349:483–491
Bermudez R, Vigliano F, Quiroga MI, Nieto JM, Bosi G, DomeneghiniC (2007) Immunohistochemical study on the neuroendocrine sys-tem of the digestive tract of turbot, Scophthalmus maximus (L.),infected by Enteromyxum scophthalmi (Myxozoa). Fish ShellfishImmunol 22:252–263
Bjenning C, Holmgren S (1988) Neuropeptides in the fish gut. An immu-nohistochemical study of evolutionary patterns. Histochemistry88:155–163
Bosi G, Di Giancamillo A, Arrighi S, Domeneghini C (2004) Animmunohistochemical study on the neuroendocrine system in thealimentary canal of the brown trout, Salmo trutta, L., 1758. GenComp Endocrinol 138:166–181
Cell Tissue Res (2013) 354:355–370 367
Author's personal copy

Bosi G, Domeneghini C, Arrighi S, Giari L, Simoni E, Dezfuli BS(2005) Response of the gut neuroendocrine system of Leuciscuscephalus (L.) to the presence of Pomphorhynchus laevis Müller,1776 (Acanthocephala). Histol Histopathol 20:509–518
Bosi G, Shinn AP, Simoni E, Arrighi S, Domeneghini C (2006) Acomparative immunohistochemical study on a galanin-like peptidein the neuroendocrine system of the alimentary canal of threespecies of siluriform catfishes. J Fish Biol 68:86–100
Bosi G, Bermudez R, Domeneghini C (2007) The galaninergic entericnervous system of pleuronectiformes (Pisces, Osteichthyes): animmunohistochemical and confocal laser scanning immunofluo-rescence study. Gen Comp Endocrinol 152:22–29
Brookes SJ (2001) Classes of enteric nerve cells in the guinea-pig smallintestine. Anat Rec 262:58–70
Burns AJ, Roberts RR, Bornstein JC, Young HM (2009) Developmentof the enteric nervous system and its role in intestinal motilityduring fetal and early postnatal stages. Semin Pediatr Surg18:196–205
Castro A, Becerra M, Manso MJ, Anadon R (2006) Calretinin immu-noreactivity in the brain of the zebrafish, Danio rerio: distributionand comparison with some neuropeptides and neurotransmitter-synthesizing enzymes. II. Midbrain, hindbrain, and rostral spinalcord. J Comp Neurol 494:792–814
Chino Y, Fujimura M, Kitahama K, Fujimiya M (2002) Colocalizationof NO and VIP in neurons of the submucous plexus in the ratintestine. Peptides 23:2245–2250
Collins SM, Van Assche G, Hogaboam C (1997) Alterations in entericnerve and smooth-muscle function in inflammatory bowel dis-eases. Inflam Bowel Dis 3:38–48
Dezfuli BS, Pironi F, Giari L, Domeneghini C, Bosi G (2002) Effect ofPomphorhynchus laevis (Acanthocephala) on putative neuromodulatorsin the intestine of naturally infected Salmo trutta. Dis Aquat Org 51:27–35
Dezfuli BS, Pironi F, Shinn AP, Manera M, Giari L (2007) Histopathologyand ultrastructure of Platichthys flesus naturally infected withAnisakis simplex s.l. larvae (Nematoda: Anisakidae). J Parasitol93:1416–1423
Domeneghini C, Radaelli G, Arrighi S, Mascarello F, Veggetti A (2000)Neurotransmitters and putative neuromodulators in the gut ofAnguilla anguilla (L.). Localizations in the enteric nervous andendocrine systems. Eur J Histochem 44:295–306
Ekblad E, Mulder H, Sundler F (1996) Vasoactive intestinal peptideexpression in enteric neurons is upregulated by both colchicineand axotomy. Regul Pept 63:113–121
Freytag C, Seeger J, Siegemund T, Grosche J, Grosche A, Freeman DE,Schusser GF, Hartig W (2008) Immunohistochemical characteri-zation and quantitative analysis of neurons in the myenteric plexusof the equine intestine. Brain Res 1244:53–64
Furness JB (2006) The enteric nervous system. Blackwell, OxfordFurness JB (2012) The enteric nervous system and neurogastroenterology.
Nat Rev Gastroenterol Hepatol 9:286–294Furness JB, Pompolo S, Murphy R, Giraud A (1989) Projections of
neurons with neuromedin U-like immunoreactivity in the smallintestine of the guinea-pig. Cell Tissue Res 257:415–422
Garcia-Lopez P, Garcia-Marin V, Martinez-Murillo R, Freire M (2009)Updating old ideas and recent advances regarding the interstitialcells of Cajal. Brain Res Rev 61:154–169
Gomez-Pinilla PJ, Gibbons SJ, Bardsley MR, Lorincz A, Pozo MJ,Pasricha PJ, Van de Rijn M, West RB, Sarr MG, Kendrick ML,Cima RR, Dozois EJ, Larson DW, Ordog T, Farrugia G (2009)Ano1 is a selective marker of interstitial cells of Cajal in the humanand mouse gastrointestinal tract. Am J Physiol Gastrointest LiverPhysiol 296:G1370–G1381
Grider JR (1994) CGRP as a transmitter in the sensory pathway medi-ating peristaltic reflex. Am J Physiol Gastrointest Liver Physiol266:G1139–G1145
Grider JR (2003) Neurotransmitters mediating the intestinal peristalticreflex in the mouse. J Pharmacol Exp Ther 307:460–467
Grunwald DJ, Eisen JS (2002) Headwaters of the zebrafish—emergenceof a new model vertebrate. Nat Rev Genet 3:717–724
Hannibal J, Ekblad E, Mulder H, Sundler F, Fahrenkrug J (1998)Pituitary adenylate cyclase activating polypeptide (PACAP) inthe gastrointestinal tract of the rat: distribution and effects ofcapsaicin or denervation. Cell Tissue Res 291:65–79
Holmberg A, Hägg U, Fritsche R, Holmgren S (2001) Occurrence ofneurotrophin receptors and transmitters in the developing Xenopusgut. Cell Tissue Res 306:35–47
Holmberg A, Schwerte T, Fritsche R, Pelster B, Holmgren S (2003)Ontogeny of intestinal motility in correlation to neuronal develop-ment in zebrafish embryo and larvae. J Fish Biol 63:318–331
Holmberg A, Schwerte T, Pelster B, Holmgren S (2004) Ontogeny ofthe gut motility control system in zebrafish Danio rerio embryosand larvae. J Exp Biol 207:4085–4094
Holmberg A, Olsson C, Hennig GW (2007) TTX-sensitive and TTX-insensitive control of spontaneous gut motility in the developingzebrafish (Danio rerio) larvae. J Exp Biol 210:1084–1091
Holmgren S, Jensen J (2001) Evolution of vertebrate neuropeptides.Brain Res Bull 55:723–735
Holmgren S, Vaillant C, Dimaline R (1982) VIP-, substance P-, gastrin/CCK-, bombesin-, somatostatin- and glucagon-like immunoreac-tivities in the gut of the rainbow trout, Salmo gairdneri. Cell TissueRes 223:141–153
Holzer P, Schicho R, Holzer-Petsche U, Lippe IT (2001) The gut as aneurological organ. Wien Klin Wochenschr 113:647–660
Hwang SJ, Blair PJ, Britton FC, O’Driscoll KE, Hennig G, BayguinovYR, Rock JR, Harfe BD, Sanders KM, Ward SM (2009)Expression of anoctamin 1/TMEM16A by interstitial cells ofCajal is fundamental for slow wave activity in gastrointestinalmuscles. J Physiol 587:4887–4904
Jensen J, Conlon JM (1992) Substance-P-related and neurokinin-A-related peptides from the brain of the cod and trout. Eur JBiochem 206:659–664
Jensen J, Holmgren S (1985) Neurotransmitters in the intestine of theAtlantic cod, Gadus morhua. Comp Biochem Physiol C 82:81–89
Jensen J, Holmgren S, Jonsson AC (1987) Substance P-like immuno-reactivity and the effects of tachykinins in the intestine of theAtlantic cod, Gadus morhua. J Auton Nerv Syst 20:25–33
Jensen J, Karila P, Jönsson A-C, Aldman G, Holmgren S (1993a)Effects of substance P and distribution of substance P-like immu-noreactivity in nerves supplying the stomach of the cod, Gadusmorhua. Fish Physiol Biochem 12:237–247
Jensen J, Olson KR, Conlon JM (1993b) Primary structures and effectson gastrointestinal motility of tachykinins from the rainbow trout.Am J Physiol Regul Integr Comp Physiol 265:R804–R810
Johansson Å, Holmgren S (2003) Ca2+-recruitment in tachykinin-induced contractions of gut smooth muscle from African clawedfrog, Xenopus laevis and rainbow trout, Oncorhynchus mykiss.Gen Comp Endocrinol 131:185–191
Johnson RJR, SchemannM, Santer RM, Cowen T (1998) The effects ofage on the overall population and on subpopulations of myentericneurons in the rat small intestine. J Anat 192:479–488
Kågström J, Holmgren S (1998) Calcitonin gene-related peptide(CGRP), but not tachykinins, causes relaxation of small arteriesfrom the rainbow trout gut. Peptides 19:577–584
Kapp S, Schrodl F, NeuhuberW, Brehmer A (2006) Chemical coding ofsubmucosal type V neurons in porcine ileum. Cells Tissues Organs184:31–41
Karila P, Holmgren S (1997) Anally projecting neurons exhibitingimmunoreactivity to galanin, nitric oxide synthase and vasoactiveintestinal peptide, detected by confocal laser scanning microscopy,in the intestine of the Atlantic cod,Gadus morhua. Cell Tissue Res287:525–533
368 Cell Tissue Res (2013) 354:355–370
Author's personal copy

Karila P, Jonsson AC, Jensen J, Holmgren S (1993) Galanin-like im-munoreactivity in extrinsic and intrinsic nerves to the gut of theAtlantic cod, Gadus morhua, and the effect of galanin on thesmooth muscle of the gut. Cell Tissue Res 271:537–544
Karila P, Shahbazi F, Jensen J, Holmgren S (1998) Projections andactions of tachykininergic, cholinergic, and serotonergic neuronesin the intestine of the atlantic cod. Cell Tissue Res 291:403–413
Kimmel CB, Ballard WW, Kimmel SR, Ullmann B, Schilling TF(1995) Stages of embryonic development of the zebrafish. DevDyn 203:253–310
KitazawaT, ItohK,YaosakaN,MaruyamaK,MatsudaK, TeraokaH,KaiyaH (2012) Ghrelin does not affect gastrointestinal contractility in rain-bow trout and goldfish in vitro. Gen Comp Endocrinol 178:539–545
Klüppel M, Huizinga JD, Malysz J, Bernstein A (1998) Developmentalorigin and Kit-dependent development of the interstitial cells ofCajal in the mammalian intestine. Dev Dyn 211:60–71
Komuro T (2006) Structure and organization of interstitial cells of Cajalin the gastrointestinal tract. J Physiol 576:653–658
Kuhlman J, Eisen JS (2007) Genetic screen for mutations affectingdevelopment and function of the enteric nervous system. DevDyn 236:118–127
Li ZS, Murphy S, Furness JB, Young HM, Campbell G (1993)Relationships between nitric oxide synthase, vasoactive intestinalpeptide and substance P immunoreactivities in neurons of theamphibian intestine. J Auton Nerv Syst 44:197–206
Lu B, Fu WM, Greengard P, Poo MM (1993) Calcitonin gene-relatedpeptide potentiates synaptic responses at developing neuromuscu-lar juntion. Nature 363:76–79
Martinez-Alvarez RM, Volkoff H, Munoz Cueto JA, Delgado MJ(2008) Molecular characterization of calcitonin gene-related pep-tide (CGRP) related peptides (CGRP, amylin, adrenomedullin andadrenomedullin-2/intermedin) in goldfish (Carassius auratus):cloning and distribution. Peptides 29:1534–1543
Matthews M, Trevarrow B, Matthews J (2002) A virtual tour of theGuide for zebrafish users. Lab Anim 31:34–40
Mazzone A, Eisenman ST, Yao Z, Ordog T, Gibbons SJ, Farrugia G (2012)Smooth muscle interstitial cells of Cajal pacing/electrical stimulationelectrogastrography. Gastroenterology 142(Suppl 1):S29
McConalogue K, Furness JB, Vremec MA, Holst JJ, Tornøe K, MarleyPD (1995) Histochemical, pharmacological, biochemical andchromatographic evidence that pituitary adenyl cyclase activatingpeptide is involved in inhibitory neurotransmission in the taenia ofthe guinea-pig caecum. J Auton Nerv Syst 50:311–322
Mirabella N, Lamanna C, Assisi L, Botte V, Cecio A (2000) Therelationships of nicotinamide adenine dinucleotide phosphate-dto nitric oxide synthase, vasoactive intestinal polypeptide, galaninand pituitary adenylate activating peptide (PACAP) in pigeon gutneurons. Neurosci Lett 293:147–151
Mirabella N, Squillacioti C, Colitti M, Germano G, Pelagalli A, Paino G(2002) Pituitary adenylate cyclase activating peptide (PACAP)immunoreactivity and mRNA expression in the duck gastrointes-tinal tract. Cell Tissue Res 308:347–359
Mitsui R (2010) Immunohistochemical characteristics of submucosalDogiel type II neurons in rat colon. Cell Tissue Res 340:257–265
Negoescu A, Labat-Moleur F, Lorimier P, Lamarcq L, Guillermet C,Chambaz E, Brambilla E (1994) F(ab) secondary antibodies: ageneral method for double immunolabeling with primary antiserafrom the same species. Efficiency control by chemiluminescence. JHistochem Cytochem 42:433–437
Ng ANY, de Jong-Curtain TA, Mawdsley DJ, White SJ, Shin J, AppelB, Dong PD, Stainier DY, Heatj JK (2005) Formation of thedigestive system in zebrafish: III. Intestinal epithelium morpho-genesis. Dev Biol 286:114–135
Ohtani R, Kaneko T, Kline LW, Labedz T, Tang Y, Pang PKT (1989)Localization of calcitonin gene-related peptide in the small intes-tine of various vertebrate species. Cell Tissue Res 258:35–42
Olsson C (2002) Distribution and effects of PACAP, VIP, nitric oxideand GABA in the gut of the African clawed frog Xenopus laevis. JExp Biol 205:1123–1134
Olsson C (2009) Autonomic innervation of the fish gut. Acta Histochem111:185–195
Olsson C, Holmgren S (1994) Distribution of PACAP (pituitary ade-nylate cyclase-activating polypeptide)-like and helospectin-likepeptides in the teleost gut. Cell Tissue Res 277:539–547
Olsson C, Holmgren S (2001) The control of gut motility. CompBiochem Physiol A 128:481–503
Olsson C, Karila P (1995) Coexistence of NAPDPH-diaphorase andvasoactive intestinal polypeptide in the enteric nervous system ofthe Atlantic cod (Gadus morhua) and the spiny dogfish (Squalusacanthias). Cell Tissue Res 280:297–305
Olsson C, Holmberg A, Holmgren S (2008) Development of enteric andvagal innervation of the zebrafish (Danio rerio) gut. J CompNeurol 508:756–770
Pederzoli A, Bertacchi I, Gambarelli A,Mola L (2004) Immunolocalisationof vasoactive intestinal peptide and substance P in the developing gutof Dicentrarchus labrax (L.). Eur J Histochem 48:179–184
Portbury AL, Pompolo S, Furness JB, StebbingMJ, KunzeWA, BornsteinJC, Hughes S (1995) Cholinergic, somatostatin-immunoreactive in-terneurons in the guinea pig intestine: morphology, ultrastructure,connections and projections. J Anat 187:303–321
Qu ZD, Thacker M, Castelucci P, Bagyanszki M, Epstein ML, FurnessJB (2008) Immunohistochemical analysis of neuron types in themouse small intestine. Cell Tissue Res 334:147–161
Rich A, Leddon SA, Hess SL, Gibbons SJ, Miller S, Xu X, Farrugia G(2007) Kit-like immunoreactivity in the zebrafish gastrointestinaltract reveals putative ICC. Dev Dyn 236:903–911
Roch GJ, Wu S, Sherwood NM (2009) Hormones and receptors in fish:do duplicates matter? Gen Comp Endocrinol 161:3–12
Salvi E, Vaccaro R, Rendag TG (1999)Ontogeny of galanin-immunoreactiveelements in the intrinsic nervous system of the chicken gut. Anat Rec254:28–38
Salvi EP, Vaccaro R, Renda TG (2000) Ontogeny of PACAP immuno-reactivity in extrinsic and intrinsic innervation of chicken gut.Peptides 21:1703–1709
Sanders KM, Zhu MH, Britton F, Koh SD, Ward SM (2011)Anoctamins and gastrointestinal smooth muscle excitability. ExpPhysiol 97:200–206
Shahbazi F, Karila P, Olsson C, Holmgren S, Conlon JM, Jensen J(1998) Primary structure, distribution, and effects on motility ofCGRP in the intestine of the cod, Gadus morhua. Am J PhysiolRegul Integr Comp Physiol 275:R19–R28
Shen Z, Larsson LT, Malmfors G, Absood A, Håkanson R, Sundler F(1992) A novel neuropeptide, pituitary adenylate cyclase-activatingpolypeptide (PACAP), in human intestine: evidence for re-duced content in Hirschsprung’s disease. Cell Tissue Res269:369–374
Shepherd I, Eisen J (2011) Development of the zebrafish enteric ner-vous system. Methods Cell Biol 101:143–160
Stanich JE, Gibbons SJ, Eisenman ST, Bardsley MR, Rock JR, HarfeBD, Ordog T, Farugia G (2011) Ano1 as a regulator of prolifera-tion. Am J Physiol Gastrointest Liver Physiol 301:G1044–G1051
Sundler F, Ekblad E, Absood A, Håkanson R, Köves K, Arimura A(1992) Pituitary adenylate cyclase activating peptide: a novel vaso-active intestinal peptide-like neuropeptide in the gut. Neuroscience46:439–454
Terkeltaub RA (2009) Colchicine update: 2008. Semin Arthritis Rheum38:411–419
Timmermans JP, Adriaensen D, Cornelissen W, Scheuermann DW(1997) Structural organization and neuropeptide distribution inthe mammalian enteric nervous system, with special attention tothose components involved in mucosal reflexes. Comp BiochemPhysiol A 118:331–340
Cell Tissue Res (2013) 354:355–370 369
Author's personal copy

Torihashi S, Ward SM, Sanders KM (1997) Development of c-Kit-positive cells and the onset of electrical rhythmicity in murinesmall intestine. Gastroenterology 112:144–155
Uyttebroek L, Shepherd IT, Harrisson F, Hubens G, Blust R,Timmermans JP, Van Nassauw L (2010) Neurochemical codingof enteric neurons in adult and embryonic zebrafish (Danio rerio).J Comp Neurol 518:4419–4438
Vigliano FA, Munoz L, Hernandez D, Cerutti P, Bermudez R, QuirogaMI (2011) An immunohistochemical study of the gut neuroendo-crine system in juvenile pejerrey Odontesthes bonariensis(Valenciennes). J Fish Biol 78:901–911
Wallace AS, Burns AJ (2005) Development of the enteric nervoussystem, smooth muscle and interstitial cells of Cajal in the humangastrointestinal tract. Cell Tissue Res 319:367–382
Wallace KN, Akhter S, Smith EM, Lorent K, Pack M (2005) Intestinalgrowth and differentiation in zebrafish. Mech Dev 122:157–173
Wang Y, Wong AO, Ge W (2003) Cloning, regulation of messengerribonucleic acid expression, and function of a new isoform ofpituitary adenylate cyclase-activating polypeptide in the zebrafishovary. Endocrinology 144:4799–4810
Ward SM, Sanders KM, Hirst GD (2004) Role of interstitial cells ofCajal in neural control of gastrointestinal smooth muscles.Neurogastroenterol Motil 16(Suppl 1):112–117
Wattchow DA, Porter AJ, Brookes SJH, Costa M (1997) The polarity ofneurochemically defined myenteric neurons in the human colon.Gastroenterology 113:497–506
Westerfield M (2007) The zebrafish book a guide for the laboratory useof zebrafish Danio Rerio. Univiversity of Oregon Press, Eugene
370 Cell Tissue Res (2013) 354:355–370
Author's personal copy




















