
Hippocampal damage produces retrograde but not anterograde amnesia for a cued location in a...
10
Hippocampal Damage Produces Retrograde but Not Anterograde Amnesia for a Cued Location in a Spontaneous Exploratory Task in Rats Scott G. Travis, Fraser T. Sparks, Tyrell Arnold, Hugo Lehmann, Robert J. Sutherland, and Ian Q. Whishaw * ABSTRACT: Performance in several memory tasks is known to be unaffected by hippocampal damage sustained before learning, but is severely disrupted if the same damage occurs after learning. Memories for preferred locations, or home bases, in exploratory tasks can be formed by rats with hippocampal damage, but it is unknown if the memory for a home base survives hippocampal damage. To examine this question, for 30 min each day for five consecutive days, rats explored a circular open field containing one local cue. By Day 5 the rats preferentially went directly to that location, spent the majority of their time at that location, made rapid direct trips to that location when returning from an excursion and so demonstrated that the location was a home base. Memory for the cued location was examined after a 24 h or 14-day interval with the cue removed. In Experiments 1 and 2, con- trol rats and rats with prior N-methyl-D-aspartic acid hippocampal lesions demonstrated memory of the home base location by making direct trips to that location. In Experiment 3, rats that had first explored the open field and cue and then received hippocampal lesions showed no memory for the cued location. The absence of anterograde impair- ment vs. the presence of retrograde impairment for memory of a spatial home base confirms a role for the hippocampus in the retention of spa- tial memory acquired during exploration. V V C 2009 Wiley-Liss, Inc. KEY WORDS: hippocampus; lesion; home base; open field; spatial navigation INTRODUCTION The hippocampus is thought to be essential to exploration and the recall of visited locations (O’Keefe and Nadel, 1978). Experimental evi- dence suggests an important role for the hippocampus (HPC) in place memory in humans (Bohbot et al., 1998; Rosenbaum et al., 2000; Wors- ley et al., 2001; Astur et al., 2002) and rodents (Morris et al., 1982; Sutherland et al., 1982; Mundy and Tilson, 1988; McDonald and White, 1995; Richmond et al., 1999; Whishaw et al., 2001; Mumby et al., 2002; Wallace and Whishaw, 2003; Clark et al., 2005a,b; Martin et al., 2005). HPC lesions can also impair subsequent acquisition of many spa- tial navigation tasks (anterograde deficits) (Morris et al., 1982; Sutherland et al., 1982; Whishaw, 1987; Bohbot et al., 1998; Astur et al., 2002; Martin et al., 2005). Finally, hippocampal lesions produce impair- ments in spatial tasks in which information was acquired before hippocampal injury (retrograde impairments) (Mumby et al., 1999; Sutherland et al., 2001; Clark et al., 2005b; Martin et al., 2005; Wino- cur et al., 2005). Nevertheless, under certain testing/training condi- tions, HPC-damaged rats can learn spatial locations in formal tests of spatial ability (Whishaw and Jarrard, 1996; Whishaw and Tomie, 1997; Hannesson and Skelton, 1998; Day et al., 1999; Hines and Whishaw, 2005; Lehmann et al., 2007a). They can also display memory for incidentally learned places. When placed in an open field, intact rats display organized behavior (Eilam and Golani, 1989; Techernichovski et al., 1998; Wallace and Whishaw, 2003; Whishaw et al., 2006; Martin et al., 2007). They form home bases— areas of preference where they will dwell, rear and groom, and from which they make slow, halting ex- ploratory excursions, followed by rapid, direct return trips. Rats with HPC damage prior to exploration ex- perience display normal home base behavior and use distal visual cues to remember home base locations (Hines and Whishaw, 2005; Lehmann et al., 2007a). There are several memory tasks for rats that do not involve spatial navigation in which hippocampal dam- age prior to experience had no effect, but the same damage after the learning experience causes severe ret- rograde disruption. For example, contextual fear condi- tioning, picture memory, and visual cue discrimination fall into this category (Maren et al., 1997; Sutherland et al., 2001, 2008; Gaskin et al., 2003; Driscoll et al., 2005; Lehmann et al., 2005, 2006, 2007b; Wiltgen et al., 2006; Broadbent et al., 2007; Epp et al., 2008). The finding that rats with prior hippocampal damage can display anterograde memory for spatial locations acquired during spontaneous exploration raises the question of whether they can display similar intact ret- rograde memory. In the present study, a previously described ‘‘home base’’ task was used to evaluate retrograde memory (Hines and Whishaw, 2005; Lehmann et al., 2007a). In three experiments, rats were exposed to a small box Department of Neuroscience, Canadian Centre for Behavioural Neuro- science, University of Lethbridge, Lethbridge, Alberta, Canada Grant sponsor: Natural Sciences and Engineering Research Council of Canada; Grant number: 746; Grant sponsors: Canadian Institute of Health Research, Alberta Heritage Fund for Medical Research. *Correspondence to: Ian Q. Whishaw, Department of Neuroscience, Ca- nadian Centre for Behavioural Neuroscience, University of Lethbridge, Lethbridge, Alberta, Canada T1K 3M4. E-mail: [email protected] Accepted for publication 20 August 2009 DOI 10.1002/hipo.20710 Published online 2 December 2009 in Wiley Online Library (wileyonlinelibrary.com). HIPPOCAMPUS 20:1095–1104 (2010) V V C 2009 WILEY-LISS, INC.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Hippocampal damage produces retrograde but not anterograde amnesia for a cued location in a...

Hippocampal Damage Produces Retrograde but Not AnterogradeAmnesia for a Cued Location in a Spontaneous Exploratory
Task in Rats
Scott G. Travis, Fraser T. Sparks, Tyrell Arnold, Hugo Lehmann,Robert J. Sutherland, and Ian Q. Whishaw*
ABSTRACT: Performance in several memory tasks is known to beunaffected by hippocampal damage sustained before learning, but isseverely disrupted if the same damage occurs after learning. Memoriesfor preferred locations, or home bases, in exploratory tasks can beformed by rats with hippocampal damage, but it is unknown if thememory for a home base survives hippocampal damage. To examinethis question, for 30 min each day for five consecutive days, ratsexplored a circular open field containing one local cue. By Day 5 therats preferentially went directly to that location, spent the majority oftheir time at that location, made rapid direct trips to that location whenreturning from an excursion and so demonstrated that the location wasa home base. Memory for the cued location was examined after a 24 hor 14-day interval with the cue removed. In Experiments 1 and 2, con-trol rats and rats with prior N-methyl-D-aspartic acid hippocampallesions demonstrated memory of the home base location by makingdirect trips to that location. In Experiment 3, rats that had first exploredthe open field and cue and then received hippocampal lesions showedno memory for the cued location. The absence of anterograde impair-ment vs. the presence of retrograde impairment for memory of a spatialhome base confirms a role for the hippocampus in the retention of spa-tial memory acquired during exploration. VVC 2009 Wiley-Liss, Inc.
KEY WORDS: hippocampus; lesion; home base; open field; spatialnavigation
INTRODUCTION
The hippocampus is thought to be essential to exploration and therecall of visited locations (O’Keefe and Nadel, 1978). Experimental evi-dence suggests an important role for the hippocampus (HPC) in placememory in humans (Bohbot et al., 1998; Rosenbaum et al., 2000; Wors-ley et al., 2001; Astur et al., 2002) and rodents (Morris et al., 1982;Sutherland et al., 1982; Mundy and Tilson, 1988; McDonald and White,1995; Richmond et al., 1999; Whishaw et al., 2001; Mumby et al.,2002; Wallace and Whishaw, 2003; Clark et al., 2005a,b; Martin et al.,2005). HPC lesions can also impair subsequent acquisition of many spa-
tial navigation tasks (anterograde deficits) (Morriset al., 1982; Sutherland et al., 1982; Whishaw, 1987;Bohbot et al., 1998; Astur et al., 2002; Martin et al.,2005). Finally, hippocampal lesions produce impair-ments in spatial tasks in which information wasacquired before hippocampal injury (retrogradeimpairments) (Mumby et al., 1999; Sutherland et al.,2001; Clark et al., 2005b; Martin et al., 2005; Wino-cur et al., 2005).
Nevertheless, under certain testing/training condi-tions, HPC-damaged rats can learn spatial locations informal tests of spatial ability (Whishaw and Jarrard,1996; Whishaw and Tomie, 1997; Hannesson andSkelton, 1998; Day et al., 1999; Hines and Whishaw,2005; Lehmann et al., 2007a). They can also displaymemory for incidentally learned places. When placedin an open field, intact rats display organized behavior(Eilam and Golani, 1989; Techernichovski et al.,1998; Wallace and Whishaw, 2003; Whishaw et al.,2006; Martin et al., 2007). They form home bases—areas of preference where they will dwell, rear andgroom, and from which they make slow, halting ex-ploratory excursions, followed by rapid, direct returntrips. Rats with HPC damage prior to exploration ex-perience display normal home base behavior and usedistal visual cues to remember home base locations(Hines and Whishaw, 2005; Lehmann et al., 2007a).There are several memory tasks for rats that do notinvolve spatial navigation in which hippocampal dam-age prior to experience had no effect, but the samedamage after the learning experience causes severe ret-rograde disruption. For example, contextual fear condi-tioning, picture memory, and visual cue discriminationfall into this category (Maren et al., 1997; Sutherlandet al., 2001, 2008; Gaskin et al., 2003; Driscoll et al.,2005; Lehmann et al., 2005, 2006, 2007b; Wiltgenet al., 2006; Broadbent et al., 2007; Epp et al., 2008).The finding that rats with prior hippocampal damagecan display anterograde memory for spatial locationsacquired during spontaneous exploration raises thequestion of whether they can display similar intact ret-rograde memory.
In the present study, a previously described ‘‘homebase’’ task was used to evaluate retrograde memory(Hines and Whishaw, 2005; Lehmann et al., 2007a).In three experiments, rats were exposed to a small box
Department of Neuroscience, Canadian Centre for Behavioural Neuro-science, University of Lethbridge, Lethbridge, Alberta, CanadaGrant sponsor: Natural Sciences and Engineering Research Council ofCanada; Grant number: 746; Grant sponsors: Canadian Institute of HealthResearch, Alberta Heritage Fund for Medical Research.*Correspondence to: Ian Q. Whishaw, Department of Neuroscience, Ca-nadian Centre for Behavioural Neuroscience, University of Lethbridge,Lethbridge, Alberta, Canada T1K 3M4. E-mail: [email protected] for publication 20 August 2009DOI 10.1002/hipo.20710Published online 2 December 2009 in Wiley Online Library(wileyonlinelibrary.com).
HIPPOCAMPUS 20:1095–1104 (2010)
VVC 2009 WILEY-LISS, INC.

in a fixed location on an open field for five consecutive days.During these sessions, the box biased the formation of a homebase at that location. A probe test, with the cue removed, wasgiven either 24 h or 2 weeks after the last day of familiariza-tion. This test was given to assess spatial memory of the homebase. Experiments 1 and 2 examined whether control rats andHPC lesion rats remembered the location of the removed cuefor both short and long durations. Experiment 3 examinedwhether rats that were first familiarized in the task and thenunderwent HPC lesions could retain a memory for the locationof the removed cue.
MATERIALS AND METHODS
Animals
Forty-seven female Long-Evans rats taken from the Univer-sity of Lethbridge vivarium were used. All were mature adults(3- to 4-months-old), and weighed 250–300 g at the beginningof the experiments. Rat housing consisted of Plexiglas cageswith sawdust bedding shared with two or three individuals.The colony room was temperature-controlled (20–218C) witha 12 h light/12 h dark cycle, beginning each day at 07:30.Food and water were provided ad libitum. In Experiments 2and 3, randomly selected rats received HPC lesions. All proce-dures were in accordance with the University of LethbridgeAnimal Care Committee guidelines, which follow the standardsset by the Canadian Council on Animal Care.
Lesion Surgeries
Rats were anesthetized with an isoflurane/oxygen mixturegiven at a rate of 0.8 L/min (Janssen, Toronto, ON), andreceived buprenorphine (Schering-Plough, Hertfordshire, UK)as an analgesic. Once anesthetized, they were secured in a ste-reotaxic frame (Kopf Instruments, Tujunga, CA) and a midlinescalpel incision was made. The lesions were made by N-methyl-D-aspartic acid (NMDA) infusions at 10 sites bilaterallyalong the hippocampus [see Lehmann et al. (2007) for coordi-nates]. A 30-gauge injection needle delivered perfusions sequen-tially, delivering 0.4 lL of NMDA at each site at a rate of0.15 lL/min. After the initial injections, the needle was left inplace for two to three additional minutes to allow for adequatediffusion of the toxin. Following the lesions, the scalp incisionwas closed using wound clips. The same surgical procedureswere used for the control rats in Experiments 2 and 3, exceptthat no damage was done to the skull or brain of these rats. Allrats received a minimum recovery period of 10 days before be-havioral testing began/resumed.
Open Field Environment
The open field apparatus shown in Figure 1 consisted of alatex-painted, circular wooden Table 155 cm in diameter, raised64 cm above the floor (Hines and Whishaw, 2005). To avoid
scented residue confounds, a ball bearing base allowed the tableto be rotated between trials. The table was also wiped downwith soapy water after each trial. The table was located in alarge room, painted white, which contained several visual cuesincluding light switches, electrical outlets, a paper towel dis-penser, a door, and two posters. Two large objects, a bench anda shelf, were covered with white bed sheets to avoid excessivesalient cue presence and decreasing chances of the rats estab-lishing home bases in proximity of these cues rather than thebox placed on the table. An opaque Plexiglas box, measuring25 cm 3 18 cm 3 12 cm served as the cue, and was placedon the edge of the table with an overhang of 1 cm. The boxwas wiped down with a soapy solution between trials, and didnot move between testing sessions, relative to the room. Thecue location, however, was balanced in each experiment. Forone half of the rats in each group it was located in SE quadrantof the table and for the other half of the rats it was in the NWquadrant of the table, with randomly selected rats exposed toopposed positions. It remained in the same location for eachrat during all sessions until it was removed for the probe trial.
Movement Analysis
A ceiling-mounted, wide-angle lens video camera recordedeach trial in Standard Play format onto Mini Digital Videocas-settes for later analysis. During open field sessions, each rat wasbrought into the testing room from its home cage and placedin the center of the open field, away from the edges of the ta-ble to avoid the influence of point of entry on location prefer-ence (Nemati and Whishaw, 2007). The experimenter then leftthe room and initiated video recording. Sessions lasted 30 min,after which the rat was removed from the table and returned toits home cage. Each rat was exposed to the open field with thecue present for five consecutive days of ‘‘cue exposure.’’ After a
FIGURE 1. The open field situation for a cue exposure trial.The cue (Plexiglas box) was removed for the probe trials. Theroom contains a number of distal cues that are clearly visible tothe rats, but dominant cues, including a sink and bookcase, arecovered by a white sheet to make them less prominent.
1096 TRAVIS ET AL.
Hippocampus

designated number of days, rats were re-exposed to the openfield without the cue in a probe test.
Time Measures
To examine the amount of time the rats spend in the variousquadrants of the table, an AccuTrak software program trans-formed the position of the rat on the Table into a series ofCartesian coordinates sampled at a rate of 30 Hz. A customwritten C11 computer program divided the open field Tableinto four quadrants, and used the AccuTrak output coordinatesto determine the amount of time the rat spent in each quad-rant over the course of the trial (Clark et al., 2005, 2006; Leh-mann et al., 2007a). Time accrued within these quadrants wasmeasured as a sum for the entire 30-min trial, and as experi-menter-assigned 3-min bins, for a total of 10 time-bins.
Direct Trips
Direct trips, defined as distinctive rapid crossing of the tabletoward the cue location (Wallace et al., 2006), were also usedas a behavioral measure. On an open field, rats will make wan-dering excursions from a cued home base, after which they willreturn to the home base in a direct and rapid manner (Wallaceet al., 2006). Over the course of an entire trial, rats may makeseveral of these exploratory movements, each followed by adirect return trip. These were identified and drawn onto a mapof the Table in a real-time review of the video recording. Direct
trips were also confirmed from the AccuTrak records of the ex-ploration of the rats, and were entered as a sum per 3-mintimebins for the entire 30-min trial, giving ten 3-min time-bins.
Histology and Lesion Analysis
Following behavioral testing, all rats were euthanized via anoverdose of sodium pentobarbital (i.p. injection). Subjects inExperiments 2 and 3 were then perfused intracardially with0.9% saline, followed by formalin (10%). Removed brainswere preserved in a 30% sucrose/10% formalin solution, untilthey were frozen and cut on a cryostat into 40-lm coronal sec-tions. Once mounted onto glass slides, they were stained usingcresyl violet. Digital images were recorded at the following valuesrelative to bregma: 22.3 mm, 23.3 mm, 24.3 mm, 25.3 mm,and 26.3 mm. Using the Image-J computer program, the tis-sue remaining within hippocampal structures at these planeswas measured, and was calculated as a percent damage com-pared to three control rats.
Procedure
A summary of the procedures used for the experiments isgiven in Figure 2. This illustrates the timeline for the experi-ments, training and testing conditions, and number of subjectstested in each group.
FIGURE 2. Experimental design and time line. In each experi-ment, the rats were given exposure to the cue (cue exposure) fol-lowed by a probe trial given either on the Day 6 or Day 19.Experiment 1 compared the performance of control rats given theprobe trial on Day 6 vs. Day 19. In Experiment 2, the rats with
either hippocampal lesions or sham surgery were given cue expo-sure followed by a probe trial on Day 19. In Experiment 3, therats received hippocampal or sham lesions following cue exposureon Day 6 and were run in a probe trial on Day 19.
HIPPOCAMPAL DAMAGE PRODUCES RETROGRADE AMNESIA 1097
Hippocampus

Experiment 1: Control Retention
Sixteen control rats were each placed on the open field for a30-min trial for 6 days, comprising of cue exposure sessions(Days 1–5), and probe trial sessions (Day 6 or Day 19). Forcue exposure sessions, the Plexiglas box was present on thetable. The rats were then randomly divided into two groups ofeight to receive either a short-delay probe trial (Day 6 probe)or long-delay probe trial (Day 19 probe). The probe trials con-sisted of a 30-min exposure to the table that was identical tothe cue exposure trials except that the cue was not present. Thelast day of cue exposure provided data for baseline, cue-presentperformance, which was contrasted with performance on theprobe trials.
Experiment 2: Anterograde Retention
Eight rats underwent HPC lesion surgery (Lesion Sx), andseven received sham surgery (Sham Sx). Two weeks later, therats were trained and tested according to the proceduredescribed in Experiment 1 (i.e., Day 19 protocol).
Experiment 3: Retrograde Retention
Similar to the procedure described in Experiment 1, 16 ratswere placed on the open field for a 30-min trial each day forfive consecutive days, comprising of cue exposure sessions(Days 1–5). On the day following the final cue exposure ses-sions (Day 6), however, eight rats underwent HPC lesion sur-gery (Lesion Sx), and eight received sham surgery (Sham Sx).Each rat was then tested on a probe trial as described inExperiment 1 on Day 19.
RESULTS
Histological Results
The NMDA lesions resulted in extensive cell loss in the areasof Ammon’s horn and the dentate gyrus. Figure 3 illustrates thelesions to the HPC by comparing a hemisphere of a control ani-mal to that of a representative lesion animal, at rostrocaudalcoronal sections 23.3 mm, 24.3 mm, and 25.3 mm frombregma. The damage to the dorsal HPC was pronounced in allrats. Grid analysis performed on coronal sections beginning at22.3 mm and ending at 26.3 mm from bregma revealed thatonly 3.56% (61.54%) of dorsal hippocampal tissue remainedin the HPC-damaged rats. The minimal amounts remaining inthe dorsal hippocampus were generally limited to the dentategranule cells and CA1-CA3 pyramidal neurons in the most lat-eral portions of the HPC. The extent of damage to the ventralHPC was also substantial with an average of 12.6% (64.14%)of tissue remaining across lesion rats. The ventral sparing was ofthe most temporal part of the HPC bilaterally in seven rats andunilaterally in three rats. The amount of damage to the subicu-lum was variable across rats and limited to the anterior region.
The lowering of the injection cannulae caused minor damage tothe parietal cortex in all lesion rats, but it was more pronouncedin one rat. The behavioral performance of that rat, however, didnot differ from the rest of the group. No noticeable damage wasfound in the thalamus, amygdala, or rhinal cortex of the hippo-campal-damaged rats. Overall, the lesion coverage is within pa-rameters of previous hippocampal lesion studies (Jarrard andMeldrum, 1993; Wallace and Whishaw, 2003).
Behavioral Results
Figure 4 illustrates direct trips made by a control rat on Day5 of cue exposures. Measures of direct trips and time spent ineach quadrant successfully characterized the behavior of the ratson Day 5 of cue exposure sessions. The measure of direct tripsindicated the organization of the rats’ behavior in response tothe cue as well as its location when it was absent. On the probetrial sessions, for which the cue was absent, the behavior ofeach rat with respect to the selection of an alternative homebase was idiosyncratic, and the rats spent very little time in thepreviously preferred quadrant. Consequently, the first directtrip and subsequent direct trips were analyzed in order to deter-mine the rats’ response to the cue and their memory for thecue’s former location.
FIGURE 3. Photomicrographs of a representative sham lesionbrain (left) and a brain that had received a hippocampal lesion(right). (a) coronal section 23.3 mm from bregma; (b) 24.3 mm;(c) 25.3 mm. Note: the lesion extensively removes the hippocampus.
1098 TRAVIS ET AL.
Hippocampus

Experiment 1: Control Retention
A summary of the results of Experiment 1 is shown in Fig-ure 5. On Day 5 of Cue Exposure the rats made more directtrips to the cue in the early timebins of the testing session;they spent the majority of their time in the quadrant of theopen field table containing the cue. Their first direct trip wasdirected to the cue location and this was also the most frequentdestination of subsequent direct trips. On the probe trials givenon Day 6 and Day 19, the rats made most visits to the previ-ous cue location in the first few time bins of the session. Withthe exception of one rat, their first direct trip was also directedto this location. The time spent in the cue quadrant decreasedfrom the cue exposure day, and the previously cued quadrantwas no longer the preferred quadrant with respect to the mea-sure of time. There were fewer direct trips made to the formercue location on Day 19 than on Day 6, but the rats also mademany fewer direct trips on Day 19.
These main findings were confirmed statistically. A 2 3 2 310 repeated measures ANOVA compared the number of directtrips to the cue location using GROUP (Day 6 Probe, Day 19Probe), DAY (cue exposure and probe testing), and TIME-BINS (10 3 3 min) as factors. There was no significant effectof GROUP (F(1,14) 5 2.16, P > 0.05), indicating that the
FIGURE 4. Representative organization of a 30-min explora-tory session on the final day of training (Day 5). After beingplaced in the center, this representative rat made an immediaterapid direct trip (1) to the cued location. Following each subse-quent exploratory excursion, the rat made a direct trip to thehome base (2–5).
FIGURE 5. Performance of control rats on Day 5 of cue expo-sure (top) and on probe trials given on Day 6 and Day 19(bottom). Left: number of direct trips per 3-min time bin to thecue location. Center: time per 30-min test spent in the quadrantcontaining the cue. Right: total number of direct trips to the table
edge in each of the four quadrants of the table (open circles—firsttrip of the session, dark circles—subsequent trips). Note: controlrats indicated that they remembered the location of the cue bydirecting their first trip, and subsequent few trips, to the cue loca-tion during the first few minutes of the test.
HIPPOCAMPAL DAMAGE PRODUCES RETROGRADE AMNESIA 1099
Hippocampus
performance of the two groups was similar on the cue exposureand probe tests. There was no significant effect of DAY(F(1,14) 5 0.09, P > 0.05), indicating that the rats made asmany direct trips with cue present as they did with the cueabsent. There was a significant effect of TIMEBINS (F(9,126)5 16.67, P < 0.001), indicating that rats made more directtrips in the initial time bins. There was a GROUP 3 DAYinteraction (F(1,14) 5 14.10, P < 0.01), caused mainly by theincreased number of direct trips made by the rats on the Day6 probe trial. Finally, there was a GROUP 3 DAY 3 TIME-BINS interaction (F(9,126) 5 2.41, P < 0.01), reflecting thereduced number of direct trips made by the Day 6 ProbeGroup in the initial timebins.
A repeated measures ANOVA comparing the time spent in thecued location using DAY (cue exposure and probe testing) andGROUP (Day 6 Probe, Day 19 Probe) revealed an effect of DAY(F(1,14) 5 158.10, P < 0.001), but no significant GROUPeffect (F(1,14) 5 0.045, P > 0.05). Hence, regardless on thelength of the retention interval, the rats spent less time at the cuelocation on the probe trials than on the cue exposure days.
An analysis of total direct trips gave no GROUP (F(1,14) 52.32, P > 0.05) difference, no significant effect of DAY(F(1,14) 5 0.716, P > 0.05), but a significant effect ofQUADRANT (F(3,42) 5 42.08, P < 0.001), and a significant
GROUP by DAY effect (F(1,14) 5 15.91, P < 0.01). Therewas no significant DAY 3 QUADRANT effect (F(3,42) 50.86, P > 0.05), but there was a significant effect of GROUP3 DAY 3 QUADRANT (F(3.42)4.49, P < 0.01). This three-way interaction reflects the fewer direct trips made to the cuedarea by the Day 19 Probe Group rats on their probe trial.
Experiment 2: Anterograde Retention
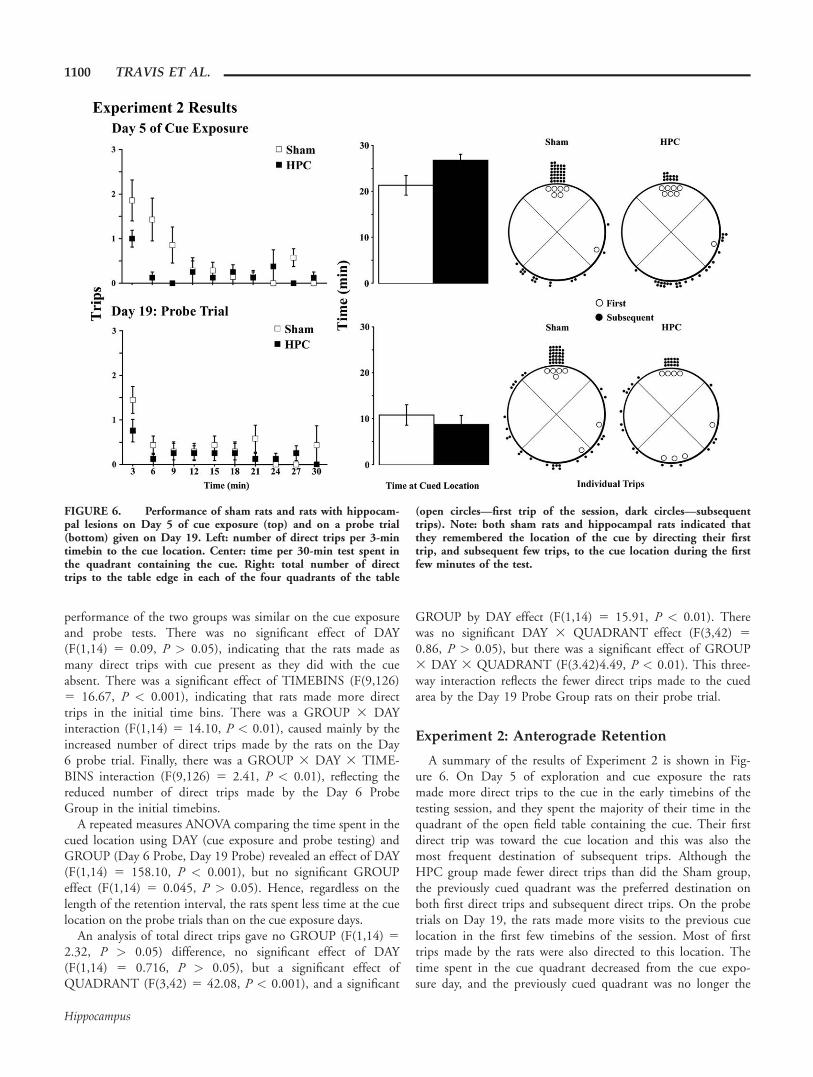
A summary of the results of Experiment 2 is shown in Fig-ure 6. On Day 5 of exploration and cue exposure the ratsmade more direct trips to the cue in the early timebins of thetesting session, and they spent the majority of their time in thequadrant of the open field table containing the cue. Their firstdirect trip was toward the cue location and this was also themost frequent destination of subsequent trips. Although theHPC group made fewer direct trips than did the Sham group,the previously cued quadrant was the preferred destination onboth first direct trips and subsequent direct trips. On the probetrials on Day 19, the rats made more visits to the previous cuelocation in the first few timebins of the session. Most of firsttrips made by the rats were also directed to this location. Thetime spent in the cue quadrant decreased from the cue expo-sure day, and the previously cued quadrant was no longer the
FIGURE 6. Performance of sham rats and rats with hippocam-pal lesions on Day 5 of cue exposure (top) and on a probe trial(bottom) given on Day 19. Left: number of direct trips per 3-mintimebin to the cue location. Center: time per 30-min test spent inthe quadrant containing the cue. Right: total number of directtrips to the table edge in each of the four quadrants of the table
(open circles—first trip of the session, dark circles—subsequenttrips). Note: both sham rats and hippocampal rats indicated thatthey remembered the location of the cue by directing their firsttrip, and subsequent few trips, to the cue location during the firstfew minutes of the test.
1100 TRAVIS ET AL.
Hippocampus

preferred quadrant with respect to the measure of time. Inde-pendent t-tests revealed that HPC animals made significantlyfewer total direct trips to the cue location than Shams on thelast day of cue exposure (P < 0.05).
These main finding were confirmed statistically. A 2 3 2 310 repeated measures ANOVA compared the number of tripsto the cue location using GROUP (Sham, HPC), DAY (cue ex-posure and probe testing), and TIMEBINS (10 3 3 min) asfactors. There was no significant GROUP effect (F(1,13) 52.98, P > 0.05). There was also no DAY effect (F(1,13) 51.55, P > 0.05). There was a significant effect of TIMEBINS(F(4.29, 55.71) 5 12.46, P < 0.001), and a GROUP 3TIMEBINS interaction (F(4.29, 55.71) 5 3.13, P < 0.01).Finally, there was no GROUP 3 DAY 3 TIMEBINS interac-tion (F(9,117) 5 1.76, P > 0.05).
A repeated measures ANOVA comparing the time spent inthe cued location using DAY (cue exposure and probe testing)and GROUP (HPC lesion, Sham) revealed an effect of DAY(F(1,13) 5 54.51, P < 0.001), but no significant GROUPeffect (F(1,13) 5 0.78, P > 0.05). This reflects the reducedtime spent at the cue location on the probe trial by bothgroups.
An analysis of direct trips gave no GROUP difference(F(1,13) 5 2.70, P < 0.05), no significant effect of DAY
(F(1,13) 5 1.6, P > 0.05), but a significant effect of QUAD-RANT, (F(3,39) 5 19.28, P < 0.001). There was no signifi-cant GROUP by DAY effect (F(1,13) 5 0.7, P > 0.05). Therewas no GROUP 3 DAY 3 QUADRANT effect (F(3,13) 50.74, P > 0.05).
Experiment 3: Retrograde Retention
A summary of the results of Experiment 3 is shown in Fig-ure 7. On Day 5 of cue exposure the rats made more directreturns to the cue in the early timebins of the testing session,and they spent the majority of their time in the quadrant ofthe open field table containing the cue. For most rats, their firstdirect trip was toward the cue location and this was also themost frequent destination of subsequent trips. On the probetrials on Day 19, the behavior of the Sham Group and theHPC group was different. The control group directed mosttrips to the previously cued location and all rats directed theirfirst trip to this location. The rats in the HPC group displayedno preference for the previously cued location on either theirfirst or subsequent trips. For both groups, the time spent in thecue quadrant decreased from the cue exposure day, and the pre-viously cued quadrant was no longer the preferred quadrantwith respect to the measure of time.
FIGURE 7. Performance of sham rats and rats with hippocam-pal lesions, given after cue exposure, on Day 5 of cue exposure(top) and on a probe trial (bottom) given on Day 19. Left: num-ber of direct trips per 3-min time bin to the cue location. Center:time per 30-min test spent in the quadrant containing the cue.Right: total number of direct trips to the table edge in each of the
four quadrants of the table (open circles—first trip of the session,dark circles—subsequent trips). Note: the sham rats, but not thehippocampal rats, indicated that they remembered the location ofthe cue by directing their first trip, and subsequent few trips, tothe cue location during the first few minutes of the test.
HIPPOCAMPAL DAMAGE PRODUCES RETROGRADE AMNESIA 1101
Hippocampus

These main finding were confirmed statistically. A 2 3 2 310 repeated measures ANOVA compared the number of tripsto the cue location using GROUP (Sham, HPC), DAY (cue ex-posure and probe testing), and TIMEBINS (10 3 3 min) asfactors. There was a significant GROUP effect (F(1,14) 56.89, P < 0.001). There was no DAY effect (F(1,14) 5 3.76,P > 0.07). There was a significant effect of TIMEBINS(F(9,126) 5 20.46 P < 0.001) and a GROUP 3 TIMEBINSinteraction (F(9,126) 5 4.07, P < 0.01). Finally, there was aGROUP 3 DAY 3 TIMEBINS interaction (F(9,126) 5 2.95,P < 0.05).
A repeated measures ANOVA comparing the time spent inthe cued location using DAY (cue exposure and probe testing)and GROUP (HPC lesion, Sham) revealed an effect of DAY(F(1,14) 5 25.30, P < 0.001), but no significant GROUPeffect (F(1,14) 5 0.21, P > 0.05). This reflects the reducedtime spent at the cue location on the probe trial by bothgroups.
An analysis of direct trips gave no GROUP effect (F(1,14)5 2/59, P > 0.05), or DAY effect (F(1,14) 5 1.03, P >0.05), but there was a significant effect of QUADRANT(F(3,42) 5 27.28, P < 0.01). There no significant effect ofGROUP 3 DAY effect (F(1,14) 5 1.03, P > 0.05), but therewas a significant GROUP 3 DAY 3 QUADRANT effect(F(3,42) 5 2.69, P < 0.05).
DISCUSSION
The present study is the first to demonstrate retrograde lossof memory for the location of a ‘‘home base’’ in an open field.The primary methods were to examine the organization of ex-ploratory behavior in order to evaluate the properties of spatialmemory. Rats were allowed to explore an open field environ-ment containing a single salient cue. At the end of this explora-tion and cue exposure, all of the rats had established a ‘‘homebase’’ near the cue in that they spent the majority of theirtime near the cue and it was the center of their exploratoryexcursions. Experiment 1 demonstrated that at both 24 h and2 weeks following exploration and cue exposure, control ratsremembered the location of the cue, as evidenced by rapidlylocomoting to that location even with the cue absent. However,the memory is weaker with the 14-day, than the 1-day, interval,suggesting that home base memory is subject to forgetting.Experiment 2 demonstrated that rats with extensive hippocam-pal damage sustained before exploration also remembered thecue location and so displayed anterograde memory. Experiment3 revealed that extensive hippocampal damage made after ex-ploration and cue exposure resulted in retrograde amnesia forthe cue location. The results extend previous findings that hip-pocampal damage produces retrograde spatial memory deficits.
The design of the present study was based on a number ofprevious studies on the organization of spontaneous exploratorybehavior in rats. Animals were allowed to explore an open fieldin which they might be expected to establish a ‘‘home base.’’ A
home base is defined as a region of the apparatus in which theanimals spend a disproportionate amount of time and fromwhich they make excursions (Eilam and Golani, 1989; Hinesand Whishaw, 2005; Whishaw et al., 2006; Lehmann et al.,2007a; Nemati and Whishaw, 2007). To bias the animals to-ward forming a home base at a given location, a salient cuewas placed on the table (Clark et al., 2005; Hines andWhishaw, 2005). Previous studies have demonstrated that bothcontrol and rats with hippocampal damage will establish ahome base near the cue (Hines and Whishaw, 2005). Thisfinding was confirmed in the present study. By the fifth day ofexposure to the open field, both control and rats that hadreceived HPC lesions ran quickly to the cue location whenplaced on the table, spent the majority of the time near thecue, and when they left the cue, returned to it with a distinc-tive direct rapid ‘‘run’’ (Hines and Whishaw, 2005).
Previous studies have also demonstrated that intact andHPC-damaged rats remember the location of a cued homebase. If they re-enter the test environment with the cueremoved, their first response is to run quickly and directly tolocation of the absent cue and repeat that behavior a numberof times during the first minutes of the exploratory test (Hinesand Whishaw, 2005; Lehmann et al., 2007a). It is likely thatthe rats remember the cue location in relation to distal roomcues, since there is no surface or other local cue that couldguide behavior (Lehmann et al., 2007a). This result was con-firmed for both intact rats and rats with HPC lesions in thepresent study. The intact control rats in all three experimentsand the HPC-damaged rats in Experiment 2, made many rapidexcursions to the quadrant of the table that had previously con-tained the cue. This behavior was prominent in the first fewminutes of the probe test. As described previously by Lehmannet al. (2007a), the control and hippocampal-damaged rats spentfar less time at the cue’s previous location later in the test.According to Whishaw et al. (2006), both the control and therats with HPC damage appeared to view the cue location as anarea of ‘‘safety’’ that they then avoid in favor of an alternateregion of the environment when the preferred cue is no longerthere.
A novel finding of the present study is that both control ratsand rats with HPC lesions can remember a cue’s previous loca-tion for an extended period of time. When the rats re-enteredthe explored environment after an interval of 14 days, with thecue absent, their behavior was similar to that observed in nor-mal rats tested after a 24 h interval (Experiment 1). Their ini-tial response was to go to the cue’s previous location and thatlocation was a preferred destination for subsequent direct trips.One difference in the behavior of the rats tested at the 14-day vs.24-h interval was that they showed less persistence in returningto the cue’s previous location. That is, control and HPC ratstested after a 14-day interval made only a few excursions in thefirst few minutes to the cue’s previous location, whereas normalrats tested after a 24-h interval in Experiment 1 investigatedthat location more frequently and for longer. It is noteworthy,however, that the response of the hippocampal-damaged ratsto the cue’s location resembled the control rats, but was not as
1102 TRAVIS ET AL.
Hippocampus

robust. In part, this difference may reflect another behavioraleffect of hippocampal damage. In tests with the cue present,the rats with HPC lesions tend to spend more time near thecue than do the control rats and make fewer excursions fromthat location. This behavior in the cue-absent tests may facili-tate more quickly establishing alternate preferred home basesand so the HPC-damaged rats returned to the cue’s previouslocation less frequently (Lehmann et al., 2007a).
The absence of anterograde amnesia in this task contrastswith extensive evidence, suggesting that the hippocampus isinvolved in acquiring and retaining new spatial information.For instance, pretraining hippocampal damage typically impairslearning and memory in the Morris Water Task, in which ratsare required to swim to a platform hidden below the surface ofthe water (Morris et al., 1982; Sutherland et al., 1982;Whishaw, 1987; Bohbot et al., 1998; Astur et al., 2002; Martinet al., 2005). However, under some circumstances, rats withHPC-damage are able to learn and remember the location ofthe hidden platform in the Morris Water task (Whishaw et al.,1995; Whishaw and Jarrard, 1996; Whishaw and Tomie, 1997;Hannesson and Skelton, 1998; Day et al., 1999; Bast et al.,2009). The latter can be achieved by additional learning ses-sions, which is believed to incrementally establish a spatial rep-resentation in nonhippocampal memory systems that acquiresinformation at a slower rate than the hippocampus (Bast et al.,2009). Given that home base memory was acquired over severallengthy sessions, it thus seems that the experience was sufficientto incrementally establish a representation in the nonhippocam-pal systems, accounting for the absence of anterograde amnesia.
The stability of long-term memory for the cue location madeit possible to first expose the rats to the exploration procedure,perform a HPC lesion, and then test the animals for retrogradememory of the cue’s location. The results of Experiment 3showed that all rats, before surgery, displayed a strong prefer-ence for the cue’s location on the fifth day of cue exposure.Their initial and subsequent direct trips were mainly to thatlocation, and they spent the majority of their exploratory sessionin the quadrant of the table that contained the cue. Whenretested 2 weeks later, and 2 weeks after receiving sham or HPCsurgery, only the rats that did not sustain brain damage rats(i.e., control) displayed memory for the cue’s previous location.All of the control rats directed their first direct trip to that loca-tion and that location was a preferred target for subsequentdirect trips. A similar preference was not observed for the ratswith HPC lesions. In fact, the first and subsequent homing tripswere idiosyncratic with respect to locations on the table.
Retrograde amnesia for spatial locations has been reported inthe Morris water maze (Gasbarri et al., 1996; Mumby et al.,1999; Sutherland et al., 2001; Clark et al., 2005a,b), the annu-lar water maze, and the Oasis dry-land task (Clark et al.,2005a). A retrograde loss of spatial memory following HPClesions suggests to us two things: first, the HPC is involved informing place memories, and second, while HPC is intact,place memories are not learned by other systems. This oversha-dowing effect would reconcile the finding of retrograde but notanterograde amnesia in the open field task, since an absence of
the HPC would allow for place memory to be learned else-where. It is possible that the network of grid cells in the ento-rhinal cortex (EC), in particular the dorsolateral EC (Steffenachet al., 2005), might, in the absence of the hippocampus be ableto store sufficient spatial information to navigate some placetasks (Hargreaves et al., 2005; Knierim, 2006; Sargolini et al.,2006), even though it does not normally do so independentlyof HPC connections. A more selective lesion study would elu-cidate these compensatory structures.
In conclusion, the finding that place memories are lost fol-lowing HPC lesions in the open field provides a new interpre-tation of the role of the HPC in learning place memory inspontaneous exploration. This loss of memory followed consid-erable exploration in the task and the full establishment of ahome base proximal to a salient cue. That there was no antero-grade memory loss suggests that the HPC normally overshad-ows other structures that are otherwise capable of acquiringplace memories.
Acknowledgments
The authors thank Lori-Ann Sacrey, Boguslaw Gorny, andMariam Alaverdashvili for their assistance with the project.
REFERENCES
Astur RS, Taylor LB, Mamelak AN, Philpott L, Sutherland RJ. 2002.Humans with hippocampus damage display severe spatial memoryimpairments in a virtual Morris water task. Behav Brain Res132:77–84.
Bast T, Wilson IA, Witter MP, Morris RG. 2009. From rapid placelearning to behavioral performance: a key role for the intermediatehippocampus. PLoS Biol 7:e1000089.
Bohbot VD, Kalina M, Stepankova K, Spackova N, Petrides M, NadelL. 1998. Spatial memory deficits in patients with lesions to theright hippocampus and to the right parahippocampal cortex. Neu-ropsychologia 36:1217–1238.
Broadbent NJ, Squire LR, Clark RE. 2007. Rats depend on habitmemory for discrimination learning and retention. Learn Mem14:145–151.
Clark BJ, Hines DJ, Hamilton DA, Whishaw IQ. 2005. Movementsof exploration intact in rats with hippocampal lesions. Behav BrainRes 163:91–99.
Clark BJ, Hines DJ, Hamilton DA, Whishaw IQ. 2006. Motor activ-ity (exploration) and formation of home bases in mice (C57BL/6)influenced by visual and tactile cues: Modification of movementdistribution, distance, location and speed. Physiol Behav 87:805–816.
Clark RE, Broadbent NJ, Squire LR. 2005a. Hippocampus andremote spatial memory in rats. Hippocampus 15:260–272.
Clark RE, Broadbent NJ, Squire LR. 2005b. Impaired remote spatialmemory after hippocampal lesions despite extensive training begin-ning early in life. Hippocampus 15:340–346.
Day LB, Weisand M, Sutherland RJ, Schallert T. 1999. The hippo-campus is not necessary for a place response but may be necessaryfor pliancy. Behav Neurosci 113:914–924.
Driscoll I, Howard SR, Prusky GT, Rudy JW, Sutherland RJ. 2005.Seahorse wins all races: Hippocampus participates in both linearand non-linear visual discrimination learning. Behav Brain Res164:29–35.
HIPPOCAMPAL DAMAGE PRODUCES RETROGRADE AMNESIA 1103
Hippocampus

Eilam D, Golani I. 1989. Home base behaviour of rats (Rattus norve-gicus) exploring a novel environment. Behav Brain Res 34:199–211.
Epp J, Keith JR, Spanswick SC, Stone JC, Prusky GT, Sutherland RJ.2008. Retrograde amnesia for visual memories after hippocampaldamage in rats. Learn Mem 15:214–221.
Gasbarri A, Sulli A, Innocenzi R, Pacitti C, Brioni JD. 1996. Spatialmemory impairment induced by lesion of the mesohippocampaldopaminergic system in the rat. Neuroscience 74:1037–1044.
Gaskin S, Tremblay A, Mumby DG. 2003. Retrograde and antero-grade object recognition in rats with hippocampal lesions. Hippo-campus 13:962–969.
Hannesson DK, Skelton RW. 1998. Recovery of spatial performancein the Morris water maze following bilateral transaction of the fim-bria/fornix in rats. Behav Brain Res 90:35–56.
Hargreaves EL, Rao G, Lee I, Knierim JJ. 2005. Major dissociationbetween medial and lateral entorhinal input to dorsal hippocam-pus. Science 308:1792–1794.
Hines DJ, Whishaw IQ. 2005. Home bases formed to visual cues butnot to self-movement (dead reckoning) cues in exploring hippo-campectomized rats. Eur J Neurosci 22:2363–2375.
Jarrard LE, Meldrum BS. 1993. Selective excitotoxic pathology in therat hippocampus. Neuropathol Appl Neurobiol 19:381–389.
Knierim JJ. 2006. Neural representations of location outside the hip-pocampus. Learn Mem 13:405–415.
Lehmann H, Carfagnini A, Yamin S, Mumby DG. 2005. Context-dependent effects of hippocampal damage on memory in theshock-probe test. Hippocampus 15:18–25.
Lehmann H, Lecluse V, Houle A, Mumby DG. 2006. Retrograde am-nesia following hippocampal lesions in the shock-probe condition-ing test. Hippocampus 16:379–387.
Lehmann H, Clark BJ, Whishaw IQ. 2007a. Similar development ofcued and learned home bases in control and hippocampal-damagedrats in an Open Field exploratory task. Hippocampus 17:370–380.
Lehmann H, Lacanilao S, Sutherland RJ. 2007b. Complete or partialhippocampal damage produces equivalent retrograde amnesia forremote contextual fear memories. Eur J Neurosci 25:1278–1286.
Maren S, Aharonov G, Fanselow MS. 1997. Neurotoxic lesions of thedorsal hippocampus and Pavlovian fear conditioning in rats. BehavBrain Res 88:261–274.
Martin MM, Horn KL, Kusman KJ, Wallace DG. 2007. Medial sep-tum lesions disrupt exploratory trip organization: Evidence for sep-tohippocampal involvement in dead reckoning. Physiol Behav90:412–424.
Martin SJ, de Hoz L, Morris RG. 2005. Retrograde amnesia: Neitherpartial nor complete hippocampal lesions in rats result in preferen-tial sparing of remote spatial memory, even after reminding. Neu-ropsychologia 43:609–624.
McDonald RJ, White NM. 1995. Hippocampal and nonhippocampalcontributions to place learning in rats. Behav Neurosci 109:579–593.
Morris RG, Garrud P, Rawlins JN, O’Keefe J. 1982. Place navigationimpaired in rats with hippocampal lesions. Nature 297:681–683.
Mumby DG, Astur RS, Weisend MP, Sutherland RJ. 1999. Retrogradeamnesia and selective damage to the hippocampal formation:Memory for places and object discriminations. Behav Brain Res106:97–107.
Mumby DG, Gaskin S, Glenn MJ, Schramek TE, Lehmann H. 2002.Hippocampal damage and exploratory preferences in rats: Memoryfor objects, places, and contexts. Learn Mem 9:49–57.
Mundy WR, Tilson HA. 1988. Behavioral impairment in the rat aftercolchicine lesions of the hippocampus and nucleus basalis. Neuro-toxicology 9:511–519.
Nemati F, Whishaw IQ. 2007. The point of entry contributes to theorganization of exploratory behaviour of rats on an open field: Anexample of spontaneous episodic memory. Behav Brain Res 182:119–128.
O’Keefe J, Nadel L. 1978. The Hippocampus As a Cognitive Map.Oxford, UK: Clarendon Press.
Richmond MA, Yee BK, Pouzet B, Veenman L, Rawlins JN, Feldon J,Bannerman DM. 1999. Dissociating context and space within thehippocampus: effects of complete, dorsal, and ventral excitotoxichippocampal lesions on conditioned freezing and spatial learning.Behav Neurosci 113:1189–1203.
Rosenbaum RS, Priselac S, Kohler S, Black SE, Gao F, Nadel L, Mosco-vitch M. 2000. Remote spatial memory in an amnesic person withextensive bilateral hippocampal lesions. Nat Neurosci 3:1044–1048.
Sargolini F, Fyhm M, Hafting T, McNaughton BL, Witter MP, MoserMB, Moser EI. 2006. Conjunctive representation of position,direction, and velocity in entorhinal cortex. Science 312:758–762.
Steffenach HA, Witter M, Moser MB, Moser EI. 2005. Spatial mem-ory in the rat requires the dorsolateral band of the entorhinal cor-tex. Neuron 45:301–313.
Sutherland RJ, Kolb B, Whishaw IQ. 1982. Spatial mapping: Defini-tive disruption by hippocampal or medial frontal cortical damagein the rat. Neurosci Lett 31:271–276.
Sutherland RJ, Weisend MP, Mumby D, Astur RS, Hanlon FM,Koerner A, Thomas MJ, Wu Y, Moses SN, Cole C, Hamilton DA,Hoesing JM. 2001. Retrograde amnesia after hippocampal damage:Recent vs. remote memories in two tasks. Hippocampus 11:27–42.
Sutherland RJ, O’Brien J, Lehmann H. 2008. Absence of systems con-solidation of fear memories after dorsal, ventral, or complete hip-pocampal damage. Hippocampus 18:710–718.
Techernichovski O, Benjamini Y, Golani I. 1998. The dynamics oflong-term exploration in the rat. I. A phase plane analysis of therelationship between location and velocity. Biol Cybern 78:423–432.
Wallace DG, Whishaw IQ. 2003. NMDA lesions of Ammon’s hornand the dentate gyrus disrupt the direct and temporally pacedhoming displayed by rats exploring a novel environment: Evidencefor a role of the hippocampus in dead reckoning. Eur J Neurosci18:513–523.
Wallace DG, Hamilton DA, Whishaw IQ. 2006. Movement character-istics support a role for dead reckoning in organizing exploratorybehavior. Anim Cogn 9:219–228.
Whishaw IQ. 1987. Hippcampal, granule cell and CA3-4 lesionsimpair formation of a place learning-set in the rat and induce reflexepilepsy. Behav Brain Res 24:59–72.
Whishaw IQ, Jarrard LE. 1996. Evidence for extrahippocampalinvolvement in place learning and hippocampal involvement inpath integration. Hippocampus 6:513–524.
Whishaw IQ, Tomie JA. 1997. Perseveration on place reversals in spa-tial swimming pool tasks: Further evidence for place learning inhippocampal rats. Hippocampus 7:361–370.
Whishaw IQ, Cassel JC, Jarrad LE. 1995. Rats with fimbria-fornixlesions display a place response in a swimming pool: A dissociationbetween getting there and knowing where. J Neurosci 15:5779–5788.
Whishaw IQ, Hines DJ, Wallace DG. 2001. Dead reckoning (pathintegration) requires the hippocampal formation: Evidence fromspontaneous exploration and spatial learning tasks in light (allo-thetic) and dark (idiothetic) tests. Behav Brain Res 127:49–69.
Whishaw IQ, Gharbawie OA, Clark BJ, Lehmann H. 2006. The ex-ploratory behaviour of rats in an open environment optimizes secu-rity. Behav Brain Res 171:230–239.
Wiltgen BJ, Sanders MJ, Anagnostaras SG, Sage JR, Fanselow MS.2006. Context fear learning in the absence of the hippocampus.J Neurosci 26:5484–5491.
Winocur G, Moscovitch M, Caruana DA, Binns MA. 2005. Retro-grade amnesia in rats with lesions to the hippocampus on a test ofspatial memory. Neuropsychologia 43:1580–1590.
Worsley CL, Recce M, Spiers HJ, Marley J, Polkey CE, Morris RG.2001. Path integration following temporal lobectomy in humans.Neuropsychologia 39:452–464.
1104 TRAVIS ET AL.
Hippocampus




















