
Differential cellular proliferation underlies heterochronic ...
Upload
independentCategory
view
5download
0
Inhibition of proliferation and induction of apoptosis in humanbreast cancer cells by lauryl gallate
Annarica Calcabrini1, Jose Manuel Garcıa-Martınez2,Lorena Gonzalez2, Mercedes Julian Tendero2,Marıa Teresa Agullo Ortuno2, Pasqualina Crateri1,Abelardo Lopez-Rivas3, Giuseppe Arancia1,Pedro Gonzalez-Porque4 and Jorge Martın-Perez2,�1Istituto Superiore di Sanita, Roma, 2Instituto de Investigaciones BiomedicasA. Sols (CSIC/UAM), Madrid, 3Centro Andaluz de Biologia del Desarrollo(CSIC/UPO), Sevilla and 4Servicio de Inmunologıa, Hospital Ramon y Cajal,Madrid
�To whom correspondence should be addressed at: Instituto de InvestigacionesBiomedicas A. Sols (CSIC/UAM), Arturo Duperier 4, 28029 Madrid, Spain.Tel: +34915854416; Fax: +34915854401;E-mail: [email protected]
Lauryl gallate is an antioxidant food additive showing lowtoxicity to normal cells. Here, its antiproliferative effecthas been studied on three human breast cancer celllines: estrogen-dependent, wild-type p53, MCF7; estrogen-independent, non-functional p53, MDA-MB-231 andMCF7 ADR, which overexpresses P-glycoprotein (P-gp)and displays a multidrug-resistant phenotype. Laurylgallate inhibited proliferation and induced cell cyclealterations in all three cell lines without altering P-gpfunctionality in the drug-resistant cells. A stable arrestin G1 phase was observed in MCF7, while a slow-downof cell cycle progression was induced in the other twocell lines. Lauryl gallate increased p53 expression onlyin MCF7, and upregulated p21Cip1 and reduced cyclinD1 levels in all three cell lines. The induction of apoptosis,demonstrated by annexin V-FITC labeling, PARP cleavageand mitochondrial membrane depolarization and morpho-logical alterations, were clearly detected in MCF7 ADRand MDA-MB-231 and to a minor extent in MCF7. Over-expression of Bcl-2 in MCF7 ADR cells demonstrated itsprotective role against morphological alterations andapoptosis. Lauryl gallate induction of p21Cip1 and apopto-sis observed in all three cell lines was regulated by Erk1/2 activation. These findings suggest a potential use of laurylgallate against tumors harboring p53 mutations and drug-resistant phenotypes.
Introduction
Breast cancer is the malignancy with the highest incidence anddeath rate for women (1), and chemotherapy is the treatment ofchoice at various stages of the disease. Although consideredone of the most chemosensitive solid tumors (2), usually after apositive response, they often relapse and develop resistance to
a broad spectrum of drugs. Consequently, metastatic breastcancer finally becomes refractory to cytotoxic drugs (3).Multidrug-resistance (MDR) represents one of the major obsta-cles for successful cancer treatment. Among other mecha-nisms, it is often associated with the overexpression of themembrane-bound drug-transporter P-glycoprotein (P-gp,ATP-dependent excretory pump), product of mdr-1 gene (4)and responsible for reducing intracellular accumulation of thedrugs and/or modifying their distribution in tumor cells (5,6).
The inability to activate apoptotic responses also representsa mechanism of drug resistance: altered expression of proteinsinvolved in apoptosis and/or cell cycle regulation (p53, Bcl-2 family proteins, cyclins, NF-kB) has been proposed toexplain drug resistance in different cell lines (7,8). Theinvolvement of multiple MDR mechanisms makes the devel-opment of new agents, less toxic to normal tissues and moreeffective against multidrug-resistant tumors than conventionalantitumoral drugs, a pivotal task.
Apoptosis is generally triggered by a number of chemicaland physical agents, such as oxidants, while antioxidant com-pounds have been reported to protect cells. However, gallicacid, a natural plant triphenol with well-known antioxidantproperties, has been demonstrated to induce apoptosis in sev-eral human cell lines (9,10). More recent studies have demon-strated that octyl and lauryl esters of gallic acid are between 50and 250 times more effective than gallic acid itself in inhibit-ing proliferation and inducing apoptosis in lymphoma andleukemia cell lines and in HT29 colon carcinoma (11).
Laurylgallatehasbeenusedasanantioxidantfoodadditiveforover 50 years with the code E312. This compound shows verylow toxicity to normal cells (12) and good specificity to tumorcells (11,13,14). Also, most of the toxicological and pharma-cokinetic studies have already been performed (12). These prop-erties support lauryl gallate as a good candidate for furtherstudies not only to determine its mechanism of action but alsofor testing its efficacy against drug-resistant tumor cell lines.
Here, we report the effect that lauryl gallate has on threehuman breast cancer epithelial cell lines: the estrogen-dependent MCF7, expressing wild-type p53, and estrogen-independent MCF7 ADR and MDA-MB-231 cell lines thatexpress non-functional p53 (15). MDA-MB-231 is character-ized by an aggressive metastatic behavior (16), while MCF7ADR was derived from its sensitive counterpart and displaysa MDR phenotype involving P-gp overexpression (17).
Our results demonstrate that lauryl gallate reduced cell via-bility, induced p21Cip1 upregulation, cell cycle alterations andapoptosis in all of these breast cancer cell lines. Also, a role ofextracellular signal-regulated kinase 1/2 (Erk1/2) on laurylgallate-induced cytotoxicity was ascertained.
Materials and methods
Reagents
Lauryl gallate was obtained from Fluka Chemie GmbH (Buchs, Switzerland).Monoclonal antibody (MAb) anti-p21Cip1 (clone SXM30) and annexin
Abbreviations: Dox, doxorubicin; Erk1/2, extracellular signal-regulatedkinase 1/2; JC-1, 5,50,6,60-tetrachloro-1,10,3,30-tetraethyl-benzimidazolcarbo-cyanine iodide; JNK; c-jun N-terminal kinase; MAb, Monoclonal antibody;MAPK, Mitogen-activated protein kinase; MDR, Multidrug-resistance; MFC,mean fluorescence channel; Mek1/2, mitogen-activated protein kinase kinase;PARP, poly (ADP-ribose) polymerase; PI, Propidium iodide; P-gp, P-glycoprotein.
Carcinogenesis vol.27 no.8 pp.1699–1712, 2006doi:10.1093/carcin/bgl044Advance Access publication April 19, 2006
# The Author 2006. Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected] 1699
by guest on March 31, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

V-FITC-conjugated were from BD Pharmingen (San Diego, CA). MAbs anti-p53 (clone DO-1, which recognizes wild-type and mutant p53) and anti-Bcl-2 were obtained from BD Transduction Laboratories. MAb anti-poly (ADP-ribose) polymerase (PARP) was from Biomol (Biomolecules for ResearchSuccess, Plymouth Meeting, PA). MAb anti-P-gp (clone F4) was from KamiyaBiomedical Company (Seattle, WA). MAb UIC2 was purchased from Chemi-con (Chemicon International, Temecula, CA). MAb anti-a-tubulin was fromSigma Chemical (St Louis, MO). Polyclonal antibodies, anti-cyclin D1 andanti-Erk2, were obtained from Santa Cruz Biotechnology (Santa Cruz, CA).Secondary horseradish peroxidase-conjugated antibodies were purchased fromBiosource International (Camarillo, CA). Secondary rabbit anti-mouse IgGFITC-conjugated antibody was obtained from Dako (Dako A/S, Glostrup,Denmark). The enhanced chemiluminescence (ECL) kit was from AmershamPharmacia Biotech (Buckinghamshire, UK). Propidium iodide (PI), 0.4% solu-tion trypan blue, ribonuclease A (RNAse), verapamil and nocodazole wereobtained from Sigma. Anti-phosphorylated-Erk1/2 (P-Erk1/2) and Mek1/2inhibitor PD98059 were purchased from Cell Signaling Technology(Beverly, MA). Doxorubicin (Adriblastina) was obtained from Pharmacia& Upjohn (Milan, Italy). Vinblastine was purchased from Eli Lilly (Paris,France). JC-1 (5,50,6,60-tetrachloro-1,10,3,30-tetraethyl-benzimidazolcarbocya-nine iodide) was from Molecular Probes (Eugene, OR).
Cell cultures
Human breast cancer cell line MCF7 and its multidrug-resistant derivativeMCF7 ADR were kindly provided by Dr K. Cowan (Nebraska Medical Center,Omaha, NE). MDA-MB-231 cell line was from American Type CultureCollection. Cells were grown in Dulbecco’s modified Eagle’s medium(DMEM) supplemented with 5% fetal calf serum (FCS), 2 mM glutamine,100 IU/ml penicillin and 100 mg/ml streptomycin in a humidified atmospherecontaining 5% CO2 and 95% air at 37�C. MCF7 ADR cells were maintained inmedium supplemented with 10 mM doxorubicin. Prior to each experiment,cells were grown in drug-free medium for 1 week. Bcl-2 overexpressing MCF7ADR clone (MAB25) and its control transfectant clone (MAN9) were providedby Dr Del Bufalo (Regina Elena Cancer Institute, Rome, Italy) (18). MCF7-E6cell line (expressing the human papillomavirus type 16 E6 protein) and MCF7-C4 (control) have been described previously (19). To generate MDA-MB-231cells expressing a mouse mutant temperature-sensitive p53 (Val135), cellswere cotransfected with the plasmid pBabe-puro (2 mg), which confers resis-tance to puromycin, and 10 mg of either empty vector or pLTRcGp53 (Val135)(20). Resistant clones were selected in 1 mg/ml puromycin (Sigma). Clones1 and 8 were chosen after analysis of p53 and p21 expression by western blot.
Cell proliferation assay
MCF7, MCF7 ADR and MDA-MB-231 cells were seeded (5 · 105 per 60-mmplate) in complete medium. After 24 h, cultures were treated with differentconcentrations of lauryl gallate (0.5, 1, 5 and 10 mM) for 24, 48 and 72 h.MCF7 C4 and E6, Bcl-2 overexpressing MCF7 ADR clone (MAB25) and itscontrol transfectant clone (MAN9), MDA-MB-231 clones 1 and 8, expressingtemperature-sensitive p53, were treated with 5 and 10 mM lauryl gallate for 72h. Lauryl gallate stock solution (10 mM) was prepared in dimethyl sulfoxide(DMSO) and added to the medium to achieve the desired final concentration.Control samples were treated with DMSO vehicle alone. To assess cell via-bility, after incubation for specified time at 37�C, aliquots from both floatingand adherent cells were mixed with a 0.4% trypan blue solution (1:1) andloaded on to a hemocytometer. The percentage of viable and dead cells persample was calculated to determine the effect of lauryl gallate on cell growth.
Analysis of P-gp functionality
MCF7 and MCF7 ADR cells were seeded (5 · 105 per 60-mm plate) incomplete medium. After 24 h, cultures were treated with 5 mM doxorubicin(Dox) for 1 h in the presence or in absence of 50 mM verapamil or 5 mM laurylgallate. At the end of treatment, samples were washed with cold phosphate-buffered saline (PBS), harvested and submitted to P-gp labeling. To this pur-pose, cells were fixed with 3.7% paraformaldehyde in PBS for 10 min at 4�C,washed twice with cold PBS and incubated with anti-P-gp monoclonal anti-body (clone F4, 2 mg/ml in PBS containing 1% BSA) for 30 min at 4�C. Aftertwo washes in cold PBS, cells were incubated with a secondary rabbit anti-mouse IgG FITC conjugate antibody for 30 min at 4�C, washed twice in coldPBS and analyzed on the FACScan flow cytometer (Becton Dickinson, Moun-tain View, CA). For UIC2 reactivity shift assay, MCF7 ADR cells (106 cellsper tube) were harvested and resuspended in 1 ml of PBS supplemented with2% FCS. The cells were warmed up to 37�C in a water bath for 10 min. Thensamples were treated with lauryl gallate (final concentrations 5 and 10 mM) orvinblastine (10 mg/ml) and incubated at 37�C for 10 min with periodic agita-tion. Vinblastine was used, as it is a well-known UIC2 shift agent (21). MAbUIC2 (final concentration 12.5 mg/ml) was added to cell suspensions and, aftermixing, incubated for an additional 15 min at 37�C. Cells were then washed
twice in ice-cold PBS supplemented with 2% FCS and 0.01% sodium azide(Shift Stop Buffer, SSB), stained on ice in SSB for an additional 15 min with asecondary rabbit anti-mouse IgG FITC-conjugated antibody. Dead cells wereexcluded from the analysis by adding PI to cell suspensions immediatelybefore acquisition. Samples were analyzed on a FACScan flow cytometerequipped with a 15 mW, 488 nm, air-cooled argon ion laser. The fluorescenceemissions were collected through a 530 nm band-pass filter for fluorescein anda 575 nm band-pass filter for doxorubicin and PI. At least 10 000 events persample were acquired in log mode.
Cell cycle analysis
MCF7, MCF7 ADR and MDA-MB-231 cells were seeded in complete medium(5 · 105 per 60-mm plate). After 24 h, they were treated for 16 h with 0.5 mg/mlnocodazole to synchronize them in G2/M phase. Then samples were washedand placed in complete medium with DMSO (control) or lauryl gallate (5 mM)for 24 and 48 h to assess the effects on cell cycle progression. For experimentsof mitogen-activated protein kinase kinase (Mek1/2) activity inhibition, cellswere treated with 10 mM PD98059 for 2 h prior to addition of 5 mM laurylgallate for 48 h. Both floating and adherent cells were collected, washed twicewith cold PBS and centrifuged. The pellet was fixed in 70% ethanol in PBS at4�C for 1 h, washed twice with cold PBS and then resuspended in PBScontaining 40 mg/ml PI and 100 mg/ml RNAse, at 37�C for 30 min. Sampleswere then analyzed on the FACScan flow cytometer. After activation of the’doublet discrimination module’ and exclusion of cell debris defining a gate inthe side and forward scatter dot-plot, at least 10 000 events per sample wereacquired in linear mode. Percentage of cells in subG1, G1, S and G2/M phaseswere calculated using the CellQuest software (Becton Dickinson).
Western blotting
Cells (1–2 · 106 cells per dish) were washed twice in ice-cold Tris-bufferedsaline [TBS; 20 mM Tris–HCl (pH 7.6) and 140 mM NaCl] and lysed at 4�C in200 ml of lysis buffer [10 mM Tris–HCl ( pH 7.6), 50 mM NaCl, 30 mMsodium pyrophosphate, 5 mM EDTA, 0.5% Nonidet P40, 1% Triton X-100, 50mM NaF, 0.1 mM Na3VO4, 1 mM PMSF, 1 mM benzamidine, 1 mM iodoac-etamide and 1 mM phenantroline]. Cell lysates were obtained by centrifugationat 17 000· g for 30 min at 4�C; protein concentration in the supernatant wasdetermined by BCA protein assay (Pierce, Rockford, IL), and lysates wereadjusted to equivalent concentrations with lysis buffer. Aliquots of 10–40 mg oftotal cell lysate were then separated on SDS–PAGE. Proteins were transferredto PVDF membranes that were blocked overnight at 4�C with 5% non-fat milkin TTBS (TBS with 0.05% Tween-20). Incubation with primary specific anti-bodies and horseradish peroxidase-conjugated secondary antibodies was per-formed in blocking solution for 1 h at room temperature. Immunoreactivebands were visualized by ECL kit. Membrane stripping was performed incu-bating for 30 min at 65�C in 62.5 mM Tris–HCl (pH 6.8) containing 2% SDSand 100 mM 2-mercaptoethanol, and extensively washed with TTBS at roomtemperature. Stripping was checked by re-exposure to enhanced chemilumi-nescence (ECL), and membranes were subsequently blocked and reproved asdescribed above.
Quantification of apoptosis by annexin V-FITC labeling
To study cell surface phosphatidylserine exposure, an early marker of apop-totic cell death, MCF7, MCF7 ADR and MDA-MB-231 cells were treated with5 mM lauryl gallate for 0.5, 1, 6 and 24 h. MCF7 C4 and E6, Bcl-2-overexpressing MCF7 ADR clone (MAB25) and its control transfectantclone (MAN9), MDA-MB-231 clones 1 and 8, expressing temperature-sensitive p53, were treated with 5 mM lauryl gallate for 6 and 24 h. ForMek1/2 activity inhibition experiments, MCF7, MCF7 ADR and MDA-MB-231 cells were treated with 10 mM PD98059 for 2 h prior to additionof 5 mM lauryl gallate for 48 h. Detached and adherent cells were collected andlabeled for 10 min at room temperature with annexin V-FITC-conjugated (1mg/ml, final concentration) and with PI (40 mg/ml, final concentration) andimmediately acquired on flow cytometer.
Ultrastructural analysis by transmission electron microscopy
Cells were seeded in complete medium (1.5 · 106 per 90-mm plate). After24 h, samples were treated with 5 mM lauryl gallate for 48 h. After thistreatment, both detached and adherent cells were collected and washed withPBS. The pellet was fixed with 2.5% glutaraldehyde in 0.1 M cacodylate buffer(pH 7.2) at room temperature for 1 h. After post-fixation with 1% OsO4 incacodylate buffer (room temperature, 1 h), cells were dehydrated throughgraded ethanol concentrations with a final propylene oxide dehydration. Sam-ples were embedded in Epon 812 resin (Electron Microscopy Science, FortWashington, PA). Ultrathin sections, obtained with a LKB ultramicrotome(LKB, Bromma, Sweden), were stained with uranyl acetate and lead citrate andexamined with a Philips 208S electron microscope (Philips Electron OpticsB.V., Eindhoven, The Netherlands). To evaluate cellular and mitochondrial
A.Calcabrini et al.
1700
by guest on March 31, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
morphological alterations, �50 cells per sample were observed from twoindependent experiments.
Analysis of mitochondrial membrane potential (MMP)
MMP was evaluated using the lipophilic cationic probe JC-1, stocked as 1 mg/ml in DMSO and freshly diluted in complete medium. Cells were treated with
5 mM lauryl gallate for 6 and 24 h. Detached and adherent cells were collectedand incubated at 37�C with medium containing JC-1 (10 ng/ml) for 10 min.Finally, cells were washed and resuspended in cold PBS for flow cytometricanalysis. JC-1 accumulates into the mitochondria as both monomers (respon-sible for green fluorescence emission, FL1) and aggregates (red fluorescenceemission, FL2) depending on MMP. Depolarization of the mitochondrial
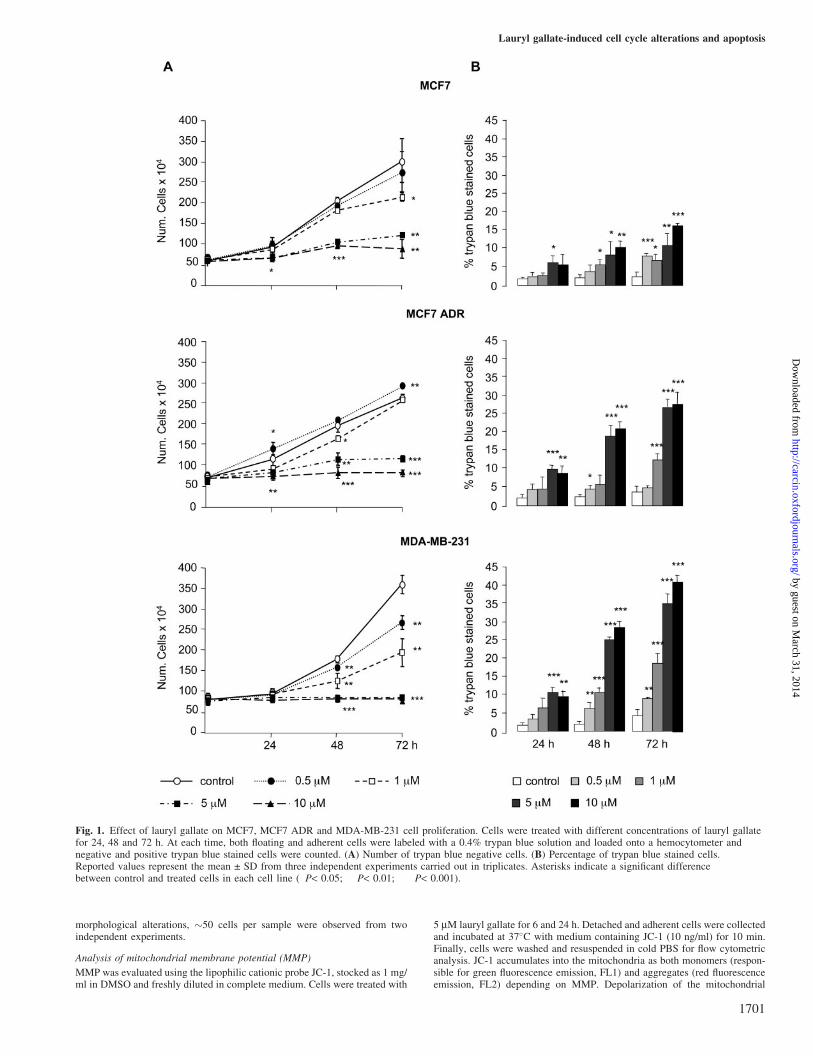
Fig. 1. Effect of lauryl gallate on MCF7, MCF7 ADR and MDA-MB-231 cell proliferation. Cells were treated with different concentrations of lauryl gallatefor 24, 48 and 72 h. At each time, both floating and adherent cells were labeled with a 0.4% trypan blue solution and loaded onto a hemocytometer andnegative and positive trypan blue stained cells were counted. (A) Number of trypan blue negative cells. (B) Percentage of trypan blue stained cells.Reported values represent the mean ± SD from three independent experiments carried out in triplicates. Asterisks indicate a significant differencebetween control and treated cells in each cell line (�P< 0.05; ��P< 0.01; ���P< 0.001).
Lauryl gallate-induced cell cycle alterations and apoptosis
1701
by guest on March 31, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
membrane is represented by an increase of FL1 and a decrease of FL2 signals,collected through a 530 and 575 nm band-pass filters, respectively. Results arepresented as mean fluorescence channel (MFC) values, calculated by theCellQuest software.
Statistical analysis
Data were expressed as means ± SD. For comparison between two groups, at-test for independent samples was used and considered significant at*P < 0.05; ��P < 0.01; or ���P < 0.001 levels.
Results
Cell proliferation
The effect of lauryl gallate treatment on the growth of threedifferent human breast cancer cell lines (MCF7, MCF7 ADRand MDA-MB-231) is shown in Figure 1. Concentrationsbetween 0.5 and 10 mM lauryl gallate reduced the numberof viable cells in all three cell lines (Figure 1A). Lauryl gallatewas more effective in MDA-MB-231 than in the other two celllines, since 50% inhibition of cell proliferation was inducedwith 1 mM and complete inhibition attained with 5 mM. Asshown in Figure 1B, MCF7 ADR and MDA-MB-231 weremore sensitive to lauryl gallate than MCF7, since a higher
percentage of trypan blue stained cells were found after treat-ment with increasing concentrations of this compound. Highersensitivity to lauryl gallate, versus MCF7, was confirmed bycounting detached cells or direct observation by phase-contrastmicroscopy (data not shown).
Functionality and expression of P-gp in MCF7 ADR cell line
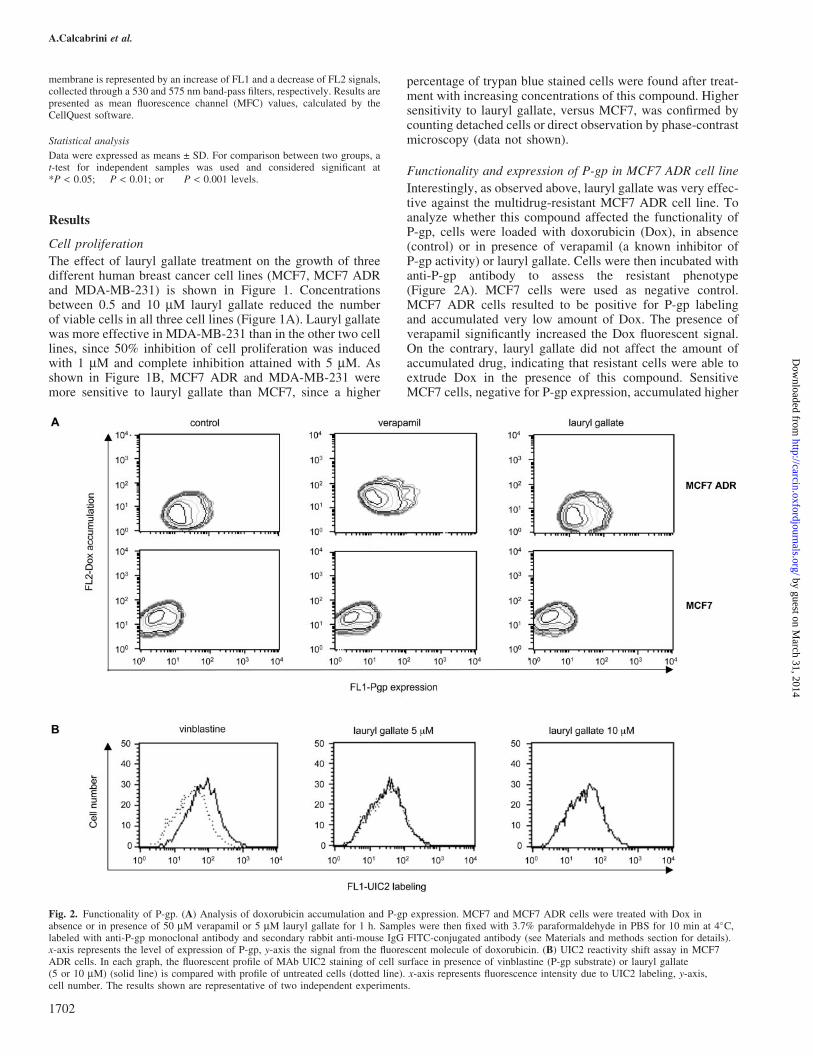
Interestingly, as observed above, lauryl gallate was very effec-tive against the multidrug-resistant MCF7 ADR cell line. Toanalyze whether this compound affected the functionality ofP-gp, cells were loaded with doxorubicin (Dox), in absence(control) or in presence of verapamil (a known inhibitor ofP-gp activity) or lauryl gallate. Cells were then incubated withanti-P-gp antibody to assess the resistant phenotype(Figure 2A). MCF7 cells were used as negative control.MCF7 ADR cells resulted to be positive for P-gp labelingand accumulated very low amount of Dox. The presence ofverapamil significantly increased the Dox fluorescent signal.On the contrary, lauryl gallate did not affect the amount ofaccumulated drug, indicating that resistant cells were able toextrude Dox in the presence of this compound. SensitiveMCF7 cells, negative for P-gp expression, accumulated higher
Fig. 2. Functionality of P-gp. (A) Analysis of doxorubicin accumulation and P-gp expression. MCF7 and MCF7 ADR cells were treated with Dox inabsence or in presence of 50 mM verapamil or 5 mM lauryl gallate for 1 h. Samples were then fixed with 3.7% paraformaldehyde in PBS for 10 min at 4�C,labeled with anti-P-gp monoclonal antibody and secondary rabbit anti-mouse IgG FITC-conjugated antibody (see Materials and methods section for details).x-axis represents the level of expression of P-gp, y-axis the signal from the fluorescent molecule of doxorubicin. (B) UIC2 reactivity shift assay in MCF7ADR cells. In each graph, the fluorescent profile of MAb UIC2 staining of cell surface in presence of vinblastine (P-gp substrate) or lauryl gallate(5 or 10 mM) (solid line) is compared with profile of untreated cells (dotted line). x-axis represents fluorescence intensity due to UIC2 labeling, y-axis,cell number. The results shown are representative of two independent experiments.
A.Calcabrini et al.
1702
by guest on March 31, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

amount of Dox, both in the absence and in the presence ofverapamil or lauryl gallate. To confirm this result, UIC2 reac-tivity shift assay was performed (21). As shown in Figure 2B,UIC2 labeling was increased in the presence of vinblastine (aknown P-gp substrate), while lauryl gallate treatment producedno shift in UIC2 reactivity even when used at higher concen-tration (10 mM), indicating that it was unable to bind to P-gpmolecules and alter their conformation.
Cell cycle analysis and expression of signaling molecules
As observed above, lauryl gallate caused cell growth inhibi-tion. Therefore, we studied the effect of lauryl gallate on cellcycle progression and on expression of proteins involved in itsregulation (Figure 3). Cell cycle studies were performed onsynchronized cells to emphasize the effect of lauryl gallate oncycle progression. Samples were treated for 16 h with 0.5 mg/ml nocodazole to synchronize cells in G2/M. Then, they werewashed and placed in complete medium with DMSO (control)or 5 mM lauryl gallate for 24 and 48 h. As shown in Figure 3A,treatment with nocodazole induced G2/M accumulation in allthree cell lines, and recovered progressively to a normal cycleafter washing-out nocodazole. In contrast, in the presence oflauryl gallate (5 mM), MCF7 accumulated in G1 phase. AlsoMCF7 ADR seemed to accumulate in G1 in the presence oflauryl gallate at 24 h but an evident shift to G2/M phase wasobserved at 48 h. Similarly, in MDA-MB-231 5 mM laurylgallate increased the percentage of cells in G1–S phases at 24 hand S–G2/M phases at 48 h. These findings suggested thatlauryl gallate induced cell cycle alterations consisting ofblocking MCF7 in G1 and delaying cell cycle progressionin MDA-MB-231 and MCF7 ADR cells.
To characterize the molecular mechanisms underlying thesedifferent behaviors, we determined p53 expression, whichplays an important role in cell cycle regulation and sensitivityto cytotoxic drugs (22). Most p53 mutations result in proteinstabilization (15,23). Therefore, as expected, the levels of p53detected in MCF7 ADR and MDA-MB-231 are higher than inp53 wild-type expressing MCF7 cells (Figure 3B). As shownin Figure 3C, treatment of MCF7 with 5 mM lauryl gallate for24, 48 and 72 h increased p53 protein, while in MCF7 ADRand MDA-MB-231 cells, lauryl gallate did not modify p53expression.
As p53 activates expression of several endogenous genes,including p21Cip1 (termed p21 hereafter), a known inhibitor ofcell cycle progression (24), we analyzed its expression in bothcontrol and lauryl gallate-treated cells (5 mM up to 72 h).Interestingly, lauryl gallate caused a time-dependent inductionof p21 in all cell lines (Figure 3C). As MCF7 ADR and MDA-MB-231 are p53 mutant cell lines, the upregulation of p21expression was likely p53-independent. Since lauryl gallateinduced alterations in cell cycle progression, we studied theexpression of cyclin D1, involved in G1 to S transition(Figure 3C). Treatment with 5 mM lauryl gallate reducedthe expression of cyclin D1, when compared with controls.
Apoptosis induction
Cell cycle modifications and alteration of its regulatory pro-teins are frequently associated with induction of apoptosis. Todetermine whether lauryl gallate activated apoptosis, somebiochemical and morphological parameters were evaluated(Figure 4). First, surface exposure of phosphatidylserine, anearly event of apoptotic program, was analyzed. Cells weretreated with 5 mM lauryl gallate for 0.5, 1, 6 and 24 h and then
labeled with annexin V-FITC-conjugated. As shown inFigure 4A, lauryl gallate significantly induced apoptosis.Early appearance (0.5 h) of apoptotic cells (defined as annexinV-positive/PI-negative) increased until 6 h in MCF7 andMCF7 ADR, while in MDA-MB-231 reached maximumafter 1 h, as compared with control (DMSO-treated) cells.MCF7 cells appeared to be more resistant to apoptosis thanthe other cell lines, even though lauryl gallate was able toinhibit cell proliferation in this cell line. These results werealso confirmed by analysis of chromatin condensation andfragmented nuclei after Hoechst labeling (data not shown).
Induction of apoptosis by lauryl gallate was also demon-strated by PARP cleavage, a commonly late marker of apop-tosis (25). In MCF7 cells, only after treatment with 10 mMlauryl gallate for 72 h the 85 kDa fragment was detected(Figure 4B), indicating that the lower dose produced verylow percentage of apoptotic cells. Since MCF7 cell linedoes not express a functional caspase 3 (26), other caspasesmust be responsible for PARP cleavage (27). In contrast, treat-ment with 5 mM lauryl gallate clearly induced the 85 kDafragment in MCF7 ADR and MDA-MB-231 at 48 and 72 h(Figure 4B). Similar results were found at 10 mM (data notshown). Consistent with these observations, lauryl gallateinduced internucleosomal DNA fragmentation in MDA-MB-231 and MCF7 ADR cells after 72 h treatment, as they showedthe typical DNA ladder even at 5 mM, while in MCF7 cells thiseffect was not observed at any concentration (data not shown).
To gain insight into morphological alterations caused bylauryl gallate, we performed transmission electron microscopystudies (Figure 4C). Exposure to 5 mM lauryl gallate for 48 hcaused ultrastructural alterations typical of apoptotic cell deaththat resulted more evident in MDA-MB-231 and MCF7 ADRthan in MCF7 cell line, as previously demonstrated by differ-ent techniques. Electron micrographs in Figure 4C refer tomorphological features of MDA-MB-231 cell line. Untreatedcells showed a nucleus characterized by a homogeneouslydistributed chromatin, a cytoplasm with numerous mitochon-dria and lysosomes and a cell surface covered by shortmicrovilli (Figure 4C1). Lauryl gallate exposure led to dra-matic alterations of the normal cellular architecture. Theseincluded an evident condensation and marginalization of chro-matin, a diffuse vacuolization of cytoplasm and loss ofmicrovilli with subsequent smoothing of cell surface(Figure 4C2). When observed at higher magnification, othertypical markers of cell damage, such as the highly condensedmitochondria, the dilation of the nuclear envelope and therough endoplasmic reticulum could be observed (Figure 4C3).
It has been demonstrated that Bcl-2 protein protects fromapoptosis (28). Since lauryl gallate treatment induced apopto-sis, but to a different extent, in MCF7, MCF7 ADR and MDA-MB-231, we analyzed possible alterations of Bcl-2 expression.As shown in Figure 5A, MCF7 expressed higher Bcl-2 levelsthan the other two cell lines. Lauryl gallate decreased Bcl-2-expression in MCF7 ADR and MDA-MB-231 cells, but did notaffect expression of Bcl-2 in MCF7 (Figure 5B). Among dif-ferent mechanisms through which Bcl-2 inhibits apoptosis, pre-vention of mitochondrial membrane depolarization due to acytotoxic insult plays a central role (29). To verify whetherlauryl gallate induced alteration of mitochondrial functionalityand that differential Bcl-2 expression could affect this response,a flow cytometric study was performed after loading with JC-1probe. In MCF7 only light alterations in FL1 and FL2 emissionswere observed in the presence of lauryl gallate for 6 and 24 h
Lauryl gallate-induced cell cycle alterations and apoptosis
1703
by guest on March 31, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
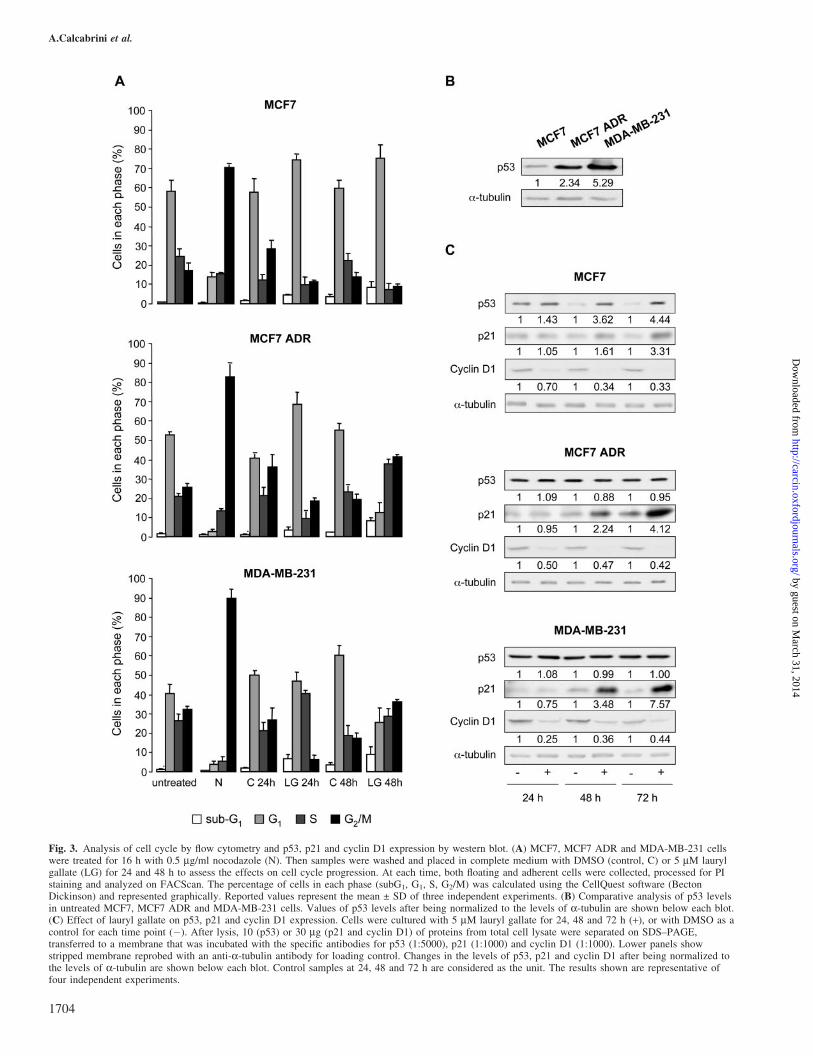
Fig. 3. Analysis of cell cycle by flow cytometry and p53, p21 and cyclin D1 expression by western blot. (A) MCF7, MCF7 ADR and MDA-MB-231 cellswere treated for 16 h with 0.5 mg/ml nocodazole (N). Then samples were washed and placed in complete medium with DMSO (control, C) or 5 mM laurylgallate (LG) for 24 and 48 h to assess the effects on cell cycle progression. At each time, both floating and adherent cells were collected, processed for PIstaining and analyzed on FACScan. The percentage of cells in each phase (subG1, G1, S, G2/M) was calculated using the CellQuest software (BectonDickinson) and represented graphically. Reported values represent the mean ± SD of three independent experiments. (B) Comparative analysis of p53 levelsin untreated MCF7, MCF7 ADR and MDA-MB-231 cells. Values of p53 levels after being normalized to the levels of a-tubulin are shown below each blot.(C) Effect of lauryl gallate on p53, p21 and cyclin D1 expression. Cells were cultured with 5 mM lauryl gallate for 24, 48 and 72 h (+), or with DMSO as acontrol for each time point (�). After lysis, 10 (p53) or 30 mg (p21 and cyclin D1) of proteins from total cell lysate were separated on SDS–PAGE,transferred to a membrane that was incubated with the specific antibodies for p53 (1:5000), p21 (1:1000) and cyclin D1 (1:1000). Lower panels showstripped membrane reprobed with an anti-a-tubulin antibody for loading control. Changes in the levels of p53, p21 and cyclin D1 after being normalized tothe levels of a-tubulin are shown below each blot. Control samples at 24, 48 and 72 h are considered as the unit. The results shown are representative offour independent experiments.
A.Calcabrini et al.
1704
by guest on March 31, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
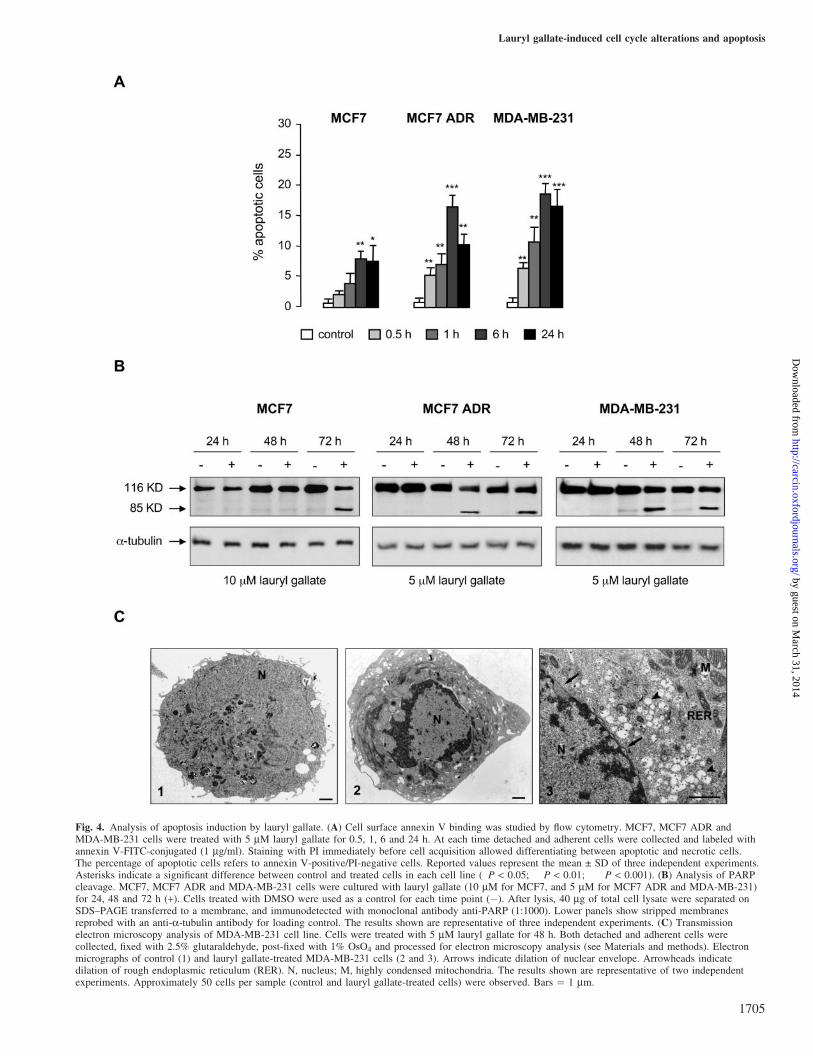
Fig. 4. Analysis of apoptosis induction by lauryl gallate. (A) Cell surface annexin V binding was studied by flow cytometry. MCF7, MCF7 ADR andMDA-MB-231 cells were treated with 5 mM lauryl gallate for 0.5, 1, 6 and 24 h. At each time detached and adherent cells were collected and labeled withannexin V-FITC-conjugated (1 mg/ml). Staining with PI immediately before cell acquisition allowed differentiating between apoptotic and necrotic cells.The percentage of apoptotic cells refers to annexin V-positive/PI-negative cells. Reported values represent the mean ± SD of three independent experiments.Asterisks indicate a significant difference between control and treated cells in each cell line (�P < 0.05; ��P < 0.01; ���P < 0.001). (B) Analysis of PARPcleavage. MCF7, MCF7 ADR and MDA-MB-231 cells were cultured with lauryl gallate (10 mM for MCF7, and 5 mM for MCF7 ADR and MDA-MB-231)for 24, 48 and 72 h (+). Cells treated with DMSO were used as a control for each time point (�). After lysis, 40 mg of total cell lysate were separated onSDS–PAGE transferred to a membrane, and immunodetected with monoclonal antibody anti-PARP (1:1000). Lower panels show stripped membranesreprobed with an anti-a-tubulin antibody for loading control. The results shown are representative of three independent experiments. (C) Transmissionelectron microscopy analysis of MDA-MB-231 cell line. Cells were treated with 5 mM lauryl gallate for 48 h. Both detached and adherent cells werecollected, fixed with 2.5% glutaraldehyde, post-fixed with 1% OsO4 and processed for electron microscopy analysis (see Materials and methods). Electronmicrographs of control (1) and lauryl gallate-treated MDA-MB-231 cells (2 and 3). Arrows indicate dilation of nuclear envelope. Arrowheads indicatedilation of rough endoplasmic reticulum (RER). N, nucleus; M, highly condensed mitochondria. The results shown are representative of two independentexperiments. Approximately 50 cells per sample (control and lauryl gallate-treated cells) were observed. Bars ¼ 1 mm.
Lauryl gallate-induced cell cycle alterations and apoptosis
1705
by guest on March 31, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
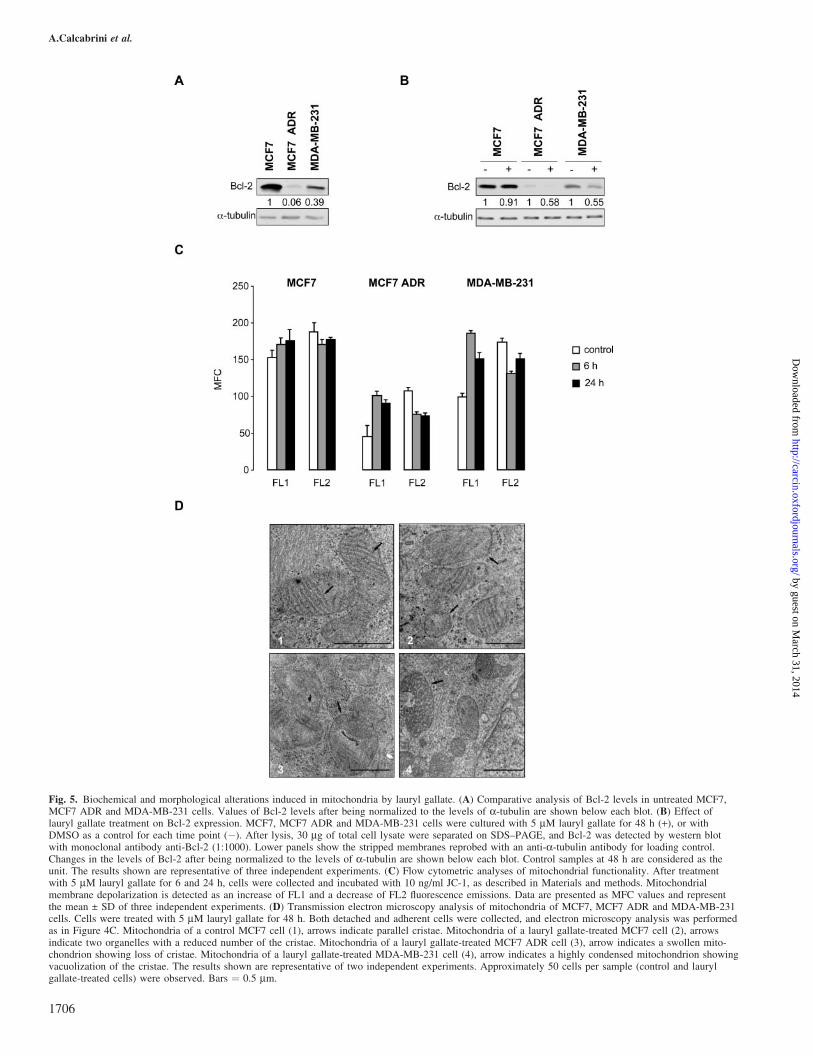
Fig. 5. Biochemical and morphological alterations induced in mitochondria by lauryl gallate. (A) Comparative analysis of Bcl-2 levels in untreated MCF7,MCF7 ADR and MDA-MB-231 cells. Values of Bcl-2 levels after being normalized to the levels of a-tubulin are shown below each blot. (B) Effect oflauryl gallate treatment on Bcl-2 expression. MCF7, MCF7 ADR and MDA-MB-231 cells were cultured with 5 mM lauryl gallate for 48 h (+), or withDMSO as a control for each time point (�). After lysis, 30 mg of total cell lysate were separated on SDS–PAGE, and Bcl-2 was detected by western blotwith monoclonal antibody anti-Bcl-2 (1:1000). Lower panels show the stripped membranes reprobed with an anti-a-tubulin antibody for loading control.Changes in the levels of Bcl-2 after being normalized to the levels of a-tubulin are shown below each blot. Control samples at 48 h are considered as theunit. The results shown are representative of three independent experiments. (C) Flow cytometric analyses of mitochondrial functionality. After treatmentwith 5 mM lauryl gallate for 6 and 24 h, cells were collected and incubated with 10 ng/ml JC-1, as described in Materials and methods. Mitochondrialmembrane depolarization is detected as an increase of FL1 and a decrease of FL2 fluorescence emissions. Data are presented as MFC values and representthe mean ± SD of three independent experiments. (D) Transmission electron microscopy analysis of mitochondria of MCF7, MCF7 ADR and MDA-MB-231cells. Cells were treated with 5 mM lauryl gallate for 48 h. Both detached and adherent cells were collected, and electron microscopy analysis was performedas in Figure 4C. Mitochondria of a control MCF7 cell (1), arrows indicate parallel cristae. Mitochondria of a lauryl gallate-treated MCF7 cell (2), arrowsindicate two organelles with a reduced number of the cristae. Mitochondria of a lauryl gallate-treated MCF7 ADR cell (3), arrow indicates a swollen mito-chondrion showing loss of cristae. Mitochondria of a lauryl gallate-treated MDA-MB-231 cell (4), arrow indicates a highly condensed mitochondrion showingvacuolization of the cristae. The results shown are representative of two independent experiments. Approximately 50 cells per sample (control and laurylgallate-treated cells) were observed. Bars ¼ 0.5 mm.
A.Calcabrini et al.
1706
by guest on March 31, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

(Figure 5C). In MCF7 ADR and MDA-MB-231 the treatmentinduced an evident increase of FL1 and a parallel decrease ofFL2 signals, indicating mitochondrial membrane depolariza-tion. As compared to sensitive MCF7 and MDA-MB-231 cells,MCF7 ADR cells showed a lower labeling pattern. This findingcould be ascribed to P-gp activity responsible for JC-1 extrusionfrom cell (30). Nevertheless, modifications of membrane poten-tial due to treatment were clearly detectable. Transmissionelectron microscopy observations of mitochondrial ultrastruc-ture were in agreement with higher alterations of mitochondrialfunctionality observed in MCF7 ADR and in MDA-MB-231than in MCF7 (Figure 5D). Control MCF7 cells showed typicalmitochondrial morphology, characterized by a double mem-brane containing a homogeneous matrix and a system of parallelcristae (Figure 5D1). Mitochondria of untreated MCF7 ADRand MDA-MB-231 cells displayed very similar features (datanot shown). Treatment with 5mM lauryl gallate for 48 h inducedin MCF7 cells only modest structural alterations consisting in areduced number of cristae (Figure 5D2). However, in MCF7ADR, swollen mitochondria showing an altered morphology, ashortening and reduction in number of cristae were clearlydetected (Figure 5D3). Also in MDA-MB-231 mitochondrialstructural modifications were more evident than in MCF7 ADR.In fact, numerous organelles with increased matrix density andsevere vacuolization of the cristae were detected (Figure 5D4).
Effect of Bcl-2 and p53 expression on apoptosis induction bylauryl gallate
As described above, lauryl gallate-induced apoptosis involvedreduction of Bcl-2 expression and mitochondrial membranedepolarization and structural modifications. Furthermore,Bcl-2 expression was higher in MCF7 than in the other two celllines, suggesting that Bcl-2 could account for the lower sen-sitivity of MCF7 to lauryl gallate. To examine this hypothesis,a Bcl-2 overexpressing clone (MAB25) derived from trans-fected MCF7 ADR cells was used (18). Cell viability andannexin V-FITC labeling analyses showed that overexpressionof Bcl-2 protected cells from apoptosis. Treatment with laurylgallate significantly reduced cell viability (Figure 6A) andinduced apoptosis (Figure 6B) in MAN9 (control transfectantclone) when compared to MAB25 (Bcl-2 overexpressingclone). When analyzed by transmission electron microscopy,MAN9 showed evident morphological alterations (condensedchromatin, vacuolization of cytoplasm) after treatment withlauryl gallate (5 mM for 48 h, Figure 6C2) as compared tocontrol cells (Figure 6C1). At higher magnification, dilation ofendoplasmic reticulum and nuclear envelope were clearlydetectable and mitochondria appeared strongly altered, show-ing swelling and loss of cristae (Figure 6C3). On the contrary,few treated MAB25 cells showed morphological featuresof apoptosis, as most of them displayed nuclear and cyto-plasmic morphology (Figure 6C5) similar to control(Figure 6C4). Furthermore, most of mitochondria preservedtheir ultrastructure despite 48 h of treatment (Figure 6C6).
Since the most sensitive cells were those expressing mutantinactive forms of p53, MCF7 ADR and MDA-MB-231, wenext analyzed the possible contribution of p53 as a target forlauryl gallate effects by two independent approaches. First, weused a MCF7 clone (MCF7-E6) expressing E6 protein fromhuman papilloma virus type 16, which induces p53 degrada-tion, therefore expressing lower levels of wild-type p53 thancontrol clone C4 (data not shown) (19). Second, we generatedclones of MDA-MB-231 expressing temperature-sensitive
mutant p53-Val135, which acquires active wild-type con-formation at 32�C (20). Clones were tested for increasedexpression of p21 at 32�C versus 37�C by western blot(data not shown), and clones 1 and 8 were selected. MCF7and MDA-MB-231 clones were treated as described above tostudy cytotoxic responses (cell viability and apoptosis) to lau-ryl gallate. If wild-type p53 confers a certain degree of resis-tance, decreasing its expression in MCF7-E6 should makethem more sensitive to lauryl gallate than control MCF7-C4. Although we observed a slightly higher sensitivity tolauryl gallate (decreased cell viability and increased apopto-sis), when calculations were made adjusting basal values, therewere not significant differences between control MCF7-C4clone and MCF7-E6 (Figure 6D). Similarly, the expressionof functional p53 in MDA-MB-231 was supposed to reducesensitivity to lauryl gallate. However, no significant differ-ences were found between clones expressing p53-Val135 at32 and 37�C (Figure 6D).
Role of Erk1/2 on lauryl gallate induction of apoptosis
Mitogen-activated protein kinase (MAPK) signaling pathwaysare activated in response to several cytotoxic compounds (31).To explore whether lauryl gallate could induce MAPK activa-tion, lysates from cells treated with 5 mM lauryl gallate for 24,48 and 72 h were subjected to western blot analysis using anti-phospho-MAPKs antibodies to detect their activation. Asshown in Figure 7A, lauryl gallate induced phosphorylationof Erk1/2 at 48 and 72 h in all three cell lines. Concerning othermembers of the MAPK family, no phosphorylation of p38kinase was observed, while only a light c-jun N-terminalkinase (JNK) activation was detected (data not shown). Anumber of studies have demonstrated a correlation betweenErk1/2 activation and p21 induction following cellular stress(32–34). To verify whether Erk1/2 activation was required forlauryl gallate-induced p21 upregulation, we inhibited theiractivation with PD98059. Treatment with 10 mM PD98059for 2 h prior to addition of 5 mM lauryl gallate for 48 h partiallyblocked both activation of Erk1/2 and upregulation of p21(Figure 7B) in all three cell lines. These results suggest thatErk1/2 activation was required for p21 upregulation. As thisprotein plays an important role in cell cycle, we then analyzedthe effects of Erk1/2 activation on cell cycle progression. Aspreviously shown (Figure 3) lauryl gallate caused MCF7 accu-mulation in G1, while in MCF7 ADR and MDA-MB-231 itinduced slow-down of cell cycle progression. Pretreatmentwith 10 mM PD98059 for 2 h partially diminished G1 accu-mulation in MCF7 cells and also modified the temporary accu-mulation in G2/M observed in the other two cell lines, wherean increased G1 was clearly detected (Figure 7C, upper panel).These results suggested that Erk1/2 activation may be involvedin cell cycle alterations induced by lauryl gallate.
Since it has been previously shown regarding the involve-ment of Erk1/2 pathway in apoptosis (35–38), we analyzed therole of Erk1/2 activation in lauryl gallate-induced apoptosis.As observed in Figure 7C, lower panel, cells treated with 10mM PD98059 for 2 h prior to addition of 5 mM lauryl gallatefor 48 h, showed a significant reduction of the percentage ofapoptotic cells as compared with lauryl gallate-treated cells.
Discussion
The effect of lauryl gallate, an antioxidant gallic acid deriva-tive, has been studied on three human breast cancer cell
Lauryl gallate-induced cell cycle alterations and apoptosis
1707
by guest on March 31, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
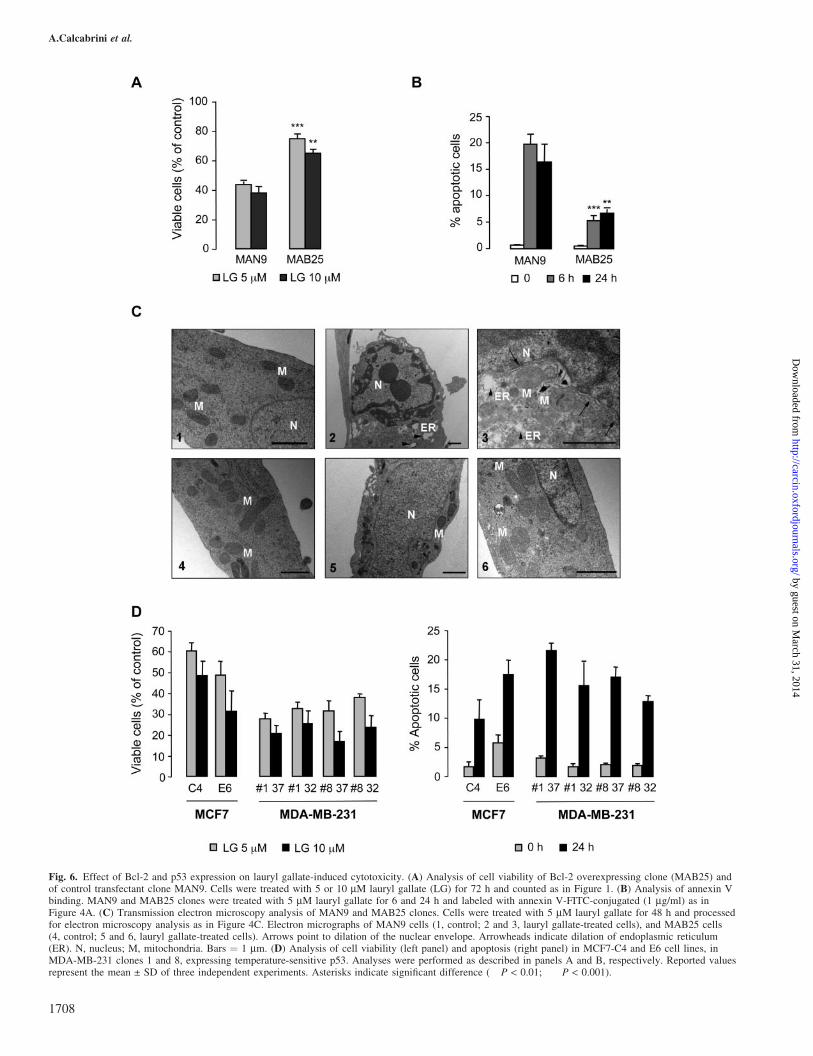
Fig. 6. Effect of Bcl-2 and p53 expression on lauryl gallate-induced cytotoxicity. (A) Analysis of cell viability of Bcl-2 overexpressing clone (MAB25) andof control transfectant clone MAN9. Cells were treated with 5 or 10 mM lauryl gallate (LG) for 72 h and counted as in Figure 1. (B) Analysis of annexin Vbinding. MAN9 and MAB25 clones were treated with 5 mM lauryl gallate for 6 and 24 h and labeled with annexin V-FITC-conjugated (1 mg/ml) as inFigure 4A. (C) Transmission electron microscopy analysis of MAN9 and MAB25 clones. Cells were treated with 5 mM lauryl gallate for 48 h and processedfor electron microscopy analysis as in Figure 4C. Electron micrographs of MAN9 cells (1, control; 2 and 3, lauryl gallate-treated cells), and MAB25 cells(4, control; 5 and 6, lauryl gallate-treated cells). Arrows point to dilation of the nuclear envelope. Arrowheads indicate dilation of endoplasmic reticulum(ER). N, nucleus; M, mitochondria. Bars ¼ 1 mm. (D) Analysis of cell viability (left panel) and apoptosis (right panel) in MCF7-C4 and E6 cell lines, inMDA-MB-231 clones 1 and 8, expressing temperature-sensitive p53. Analyses were performed as described in panels A and B, respectively. Reported valuesrepresent the mean ± SD of three independent experiments. Asterisks indicate significant difference (��P < 0.01; ���P < 0.001).
A.Calcabrini et al.
1708
by guest on March 31, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

Fig. 7. Role of Erk1/2 in lauryl gallate-induced cytotoxicity. (A) Effect of lauryl gallate on Erk1/2 activation. MCF7, MCF7 ADR and MDA-MB-231 cellswere cultured with 5 mM lauryl gallate (LG) or DMSO, as a control, for 24, 48 and 72 h. After lysis, 30 mg of total cell lysate were separated on SDS–PAGE, transferred to a membrane that was then incubated with polyclonal antibody anti-phosphorylated-Erk1/2 (P-Erk1/2) (1:2000). Lower panels show thestripped membranes reprobed with anti-Erk2 antibody for loading control. (B) Role of lauryl gallate on Erk1/2 induction of p21 expression. Cells weretreated with 10 mM PD98059 for 2 h prior to addition of 5 mM lauryl gallate (LG) for 48 h. Analysis of P-Erk1/2 and Erk2 proteins were performed asabove. For p21 detection, 40 mg of total cell lysate were separated on SDS–PAGE, transferred to a membrane that was then incubated with monoclonalantibody anti-p21 (1:1000). Lower panels show stripped membranes reprobed with an anti-a-tubulin antibody for loading control. Changes in the levels ofP-Erk1/2 and p21 after being normalized to the levels of Erk2 and a-tubulin, respectively, are shown below each blot. Control samples are considered as theunit. Results shown are representative of three independent experiments. (C) Role of Erk1/2 on cell cycle alterations (upper panel) and apoptosis (lowerpanel) induced by lauryl gallate. Cells were treated with 10 mM PD98059 for 2 h prior to addition of 5 mM lauryl gallate (LG) for 48 h. Cell cycle analyseswere performed as in Figure 3A. Apoptosis was determined with annexin V-FITC-conjugated (1 mg/ml) as in Figure 4A. Reported values represent themean ± SD of three independent experiments. Asterisks indicate a significant difference between samples treated with PD98059+LG and those treated withLG alone in each cell line (*P < 0.05; ��P < 0.01).
Lauryl gallate-induced cell cycle alterations and apoptosis
1709
by guest on March 31, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

lines: wild-type p53 MCF7, and two non-functional p53 celllines, MCF7 ADR (multidrug-resistant) and the highly prolif-erative and invasive MDA-MB-231. Our results show thatlauryl gallate behaved as a potent antiproliferative agent induc-ing cell cycle alterations and apoptosis in all cell lines tested.
A wide variety of cytotoxic insults result in activation ofmechanisms that arrest cell cycle progression at specificcheckpoints, presumably to allow cells to repair DNA damage.This is consistent with G1 arrest of MCF7 induced by laurylgallate. However, in MCF7 ADR and MDA-MB-231, a stableblock in G1 phase was not achieved and cell cycle progressedallowing cells to accumulate damages.
Cell cycle-mediated apoptosis is a common way for celldeath upon treatment with anti-tumor agents and inability toactivate the apoptotic program confers drug resistance.Another mechanism to evade the action of cytotoxic agentsis to express MDR genes, such as mdr-1 encoding for P-gp (4).For that reason, one of the strategies employed to overcomedrug resistance is to inhibit the function of P-gp, allowingresistant cells to accumulate drugs to levels similar to thosefound in sensitive cells (39). Therefore, it is of an utmostimportance to obtain compounds that will not be affectedby P-gp excretory action. Our results demonstrate that laurylgallate induces strong effects in the multidrug-resistant MCF7ADR cells, characterized by P-gp overexpression. Similarly,cytofluorimetric assays by doxorubicin accumulation andUIC2 shift demonstrated that lauryl gallate did not interferewith the proper functionality of P-gp when analyzed towardsother compounds. This result is in contrast with recent studiesdemonstrating that polyphenols from green tea, such as epi-gallocatechin gallate, inhibited activity of P-gp, increasingintracellular accumulation of P-gp substrates, acting as poten-tial MDR reversal agents when used in combination with anti-tumoral compounds (40,41). The higher complexity of thesepolyphenols may explain a different behavior from the one wereport for lauryl gallate.
Among proteins involved in control of apoptotic pathway,Bcl-2 has been demonstrated to protect cells from both normaland experimentally induced apoptosis (28). Bcl-2 is located inthe outer mitochondrial membrane and its apoptosis-regulatory effects are mainly related with mitochondrial struc-ture and function. This protein prevents opening of mitochon-drial permeability transition pores and, subsequently, inhibitsmitochondrial membrane depolarization and release of apop-togenic factors to cytosol (29,42).
The higher Bcl-2 protein levels detected in MCF7 as com-pared to the other two cell lines could result in a lower sen-sitivity to lauryl gallate. This hypothesis was confirmed byelectron microscopy and flow cytometry analyses, showinglower functional and morphological alterations in mitochon-dria of MCF7 cells as compared to those observed in MDA-MB-231 and MCF7 ADR cells. Furthermore, the protectiverole of Bcl-2 against lauryl gallate-induced apoptosis wasclearly demonstrated by higher viability and lower numberof cells with morphological apoptotic features shown byMAB25 (Bcl-2 overexpressing MCF7 ADR clone) as com-pared to MAN9 control clone. Similar protective effects ofBcl-2 have been reported in murine B cell lymphoma lineWehi 231 (13). Other studies have demonstrated that a partialresistance of MCF7 against camptothecin and UCN-01 isrelated to Bcl-2 expression (23).
Activation of G1 checkpoint requires functional p53, whichtranscriptionally activates downstream genes to induce G1 cell
cycle arrest and, in some cases, apoptosis (22). Mutation ofp53 tumor suppressor gene is one of the most common geneticalterations in human cancers, including breast cancer, whichinduces uncontrolled cell proliferation (43). Wild-type p53protein has been shown to mediate apoptosis, while mutationsor deletions within p53 gene have been associated, in vivo andin vitro, both with resistance to apoptosis by differentchemotherapeutic agents in some cell lines and with increasedsensitivity in others (44,45). Results obtained in MCF7-E6cells and in MDA-MB-231 p53-Val135 suggest that p53 isnot a major target for lauryl gallate-induced cytotoxicity.
Among p53-activated genes, p21 is a potent inhibitor ofcyclin-dependent kinase activities required for progressionfrom G1 to S phase of the cell cycle (24). Surprisingly, laurylgallate induced p21 expression in all three cell lines tested.Increased p21 expression in MCF7 correlated with G1 arrest,and a delayed cell cycle progression in the other two cell lines.As MCF7 ADR and MDA-MB-231 cells express a mutatedp53, induction of p21 by lauryl gallate could be attributed toactivation of a p53-independent pathway.
A number of studies demonstrated p53-independent mecha-nisms able to regulate p21 expression through Erk1/2 activa-tion (32,46). Cell cycle perturbations and p21 upregulation bysynthetic compounds occurred through activation of ATM/ATR-Erk1/2 pathway (33). Also, inhibition of Mek1/2 activityclearly reduced the induction of p21 promoter, mRNA andprotein expression (34). In our cell model, lauryl gallate causedsustained activation of Erk1/2, clearly detected in both wild-type and mutated p53 cell lines, indicating a p53-independentmechanism. Abrogation of Erk1/2 activation by Mek1/2 inhibitor PD98059 reduced p21 induction and cell cyclealterations (G1 block in MCF7 and delayed progressionin MCF7 ADR and MDA-MB-231) due to lauryl gallate.Furthermore, apoptosis induction was significantly reducedby Erk1/2 inhibition. The involvement of Mek1/2–Erk1/2-pathway in the induction of apoptosis by different cytotoxiccompounds has been recently demonstrated (32,35,37,38).Also, its contribution to cell proliferation and survival iswell established (47). Erk1/2 has been identified as kinaseresponsible for Bcl-2 phosphorylation at Ser87 residue (48).Phosphorylation of Bcl-2, usually distributed in hypophospho-rylated form on the mitochondrial membrane, results in reduc-tion of its anti-apoptotic activity and redistribution frommitochondria to cytoplasm. We could hypothesize that laurylgallate induced apoptosis by activation of Erk1/2 and subse-quent phosphorylation of Bcl-2. The ability of Mek1/2 inhibitor PD98059 to reduce apoptosis could account forthis hypothesis. In addition, the lower cytotoxicity observedin MCF7 and MAB25 could be in agreement with this theory.Given that these cells expressed elevated levels of Bcl-2, ahigher lauryl gallate concentration may be required to obtainthe same effect as those observed in MCF7 ADR andMDA-MB-231 with lower levels of Bcl-2. As for activationof other MAPKs, lauryl gallate did not induce p38 phospho-rylation and only slightly activated JNK (data not shown).Studies on the mechanism of action of other molecules withanticancer properties showed that, in addition to Erk1/2, p38and/or JNK were also activated (35,37). The fact that theMek1/2–Erk1/2 pathway has been shown to contribute toboth cell proliferation and survival and also to mediate cellcycle alteration and apoptosis (47), agrees with the commonbelief that no general mechanism of action for cytotoxiccompounds can be postulated.
A.Calcabrini et al.
1710
by guest on March 31, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

In conclusion, our data indicate that lauryl gallate inducedreduction of cell survival and cell cycle alterations in MCF7,MCF7 ADR and MDA-MB-231 cells. An evident p21 upregu-lation was observed in all three breast cancer cell lines, fol-lowed by apoptotic cell death. Both p21 upregulation andapoptosis induction were regulated, at least in part, byErk1/2 activation. Interestingly, a stronger cytotoxic effectwas observed on multidrug-resistant MCF7 ADR cells aswell as on MDA-MB-231 carrying mutations in the p53gene, which, in theory, should be more resistant to cytotoxictreatments and apoptosis. These findings suggest that laurylgallate might be a good candidate for innovative therapeuticstrategies against tumors carrying p53 mutations and resistantto conventional chemotherapeutic agents.
Acknowledgements
We thank J. Perez and L. Camilli for the artwork. Authors are grateful to D. DelBulafo and D. Trisciuoglio for donation of MCF7 ADR MAN9 and MAB25clones, and together with I. Lazaro-Trueba for their constructive comments.This work was supported by grants to JM-P from MCyT (SAF2003-02188),and FIS (01/1316, 03C03/10 and PI040682) and a grant from MCyT to AL-R(SAF2003-00402). JMGM was supported by a fellowship from FIS, LG wassupported by a fellowship from Fundacion Carolina, MTAO was supported bythe FIS 03C03/10 grant, and MJT by a FINNOVA fellowship. JMP is amember of GEICAM.
Conflict of Interest Statement: None declared.
References
1.Edwards,B.K., Brown,M.L., Wingo,P.A. et al. (2005) Annual report to thenation on the status of cancer, 1975-2002, featuring population-basedtrends in cancer treatment. J. Natl Cancer Inst., 97, 1407–1427.
2.Cristofanilli,M. and Hortobagyi,G.N. (2004) Breast cancer highlights: keyfindings from the San Antonio Breast Cancer Symposium: a U.S.perspective. Oncologist, 9, 471–478.
3.Mariani,G. (2005) New developments in the treatment of metastatic breastcancer: from chemotherapy to biological therapy. Ann. Oncol., 16(Suppl 2), ii191–ii194.
4.Ambudkar,S.V., Kimchi-Sarfaty,C., Sauna,Z.E. and Gottesman,M.M.(2003) P-glycoprotein: from genomics to mechanism. Oncogene, 22,7468–7485.
5.Larsen,A.K., Escargueil,A.E. and Skladanowski,A. (2000) Resistancemechanisms associated with altered intracellular distribution ofanticancer agents. Pharmacol. Ther., 85, 217–229.
6.Molinari,A., Calcabrini,A., Meschini,S., Stringaro,A., Crateri,P.,Toccacieli,L., Marra,M., Colone,M., Cianfriglia,M. and Arancia,G.(2002) Subcellular detection and localization of the drug transporter P-glycoprotein in cultured tumor cells. Curr. Protein Pept. Sci., 3, 653–670.
7.Herr,I. and Debatin,K.M. (2001) Cellular stress response and apoptosis incancer therapy. Blood, 98, 2603–2614.
8.Pommier,Y., Sordet,O., Antony,S., Hayward,R.L. and Kohn,K.W. (2004)Apoptosis defects and chemotherapy resistance: molecular interactionmaps and networks. Oncogene, 23, 2934–2949.
9. Inoue,M., Suzuki,R., Koide,T., Sakaguchi,N., Ogihara,Y. and Yabu,Y.(1994) Antioxidant, gallic acid, induces apoptosis in HL-60RG cells.Biochem. Biophys. Res. Commun., 204, 898–904.
10. Inoue,M., Suzuki,R., Sakaguchi,N., Li,Z., Takeda,T., Ogihara,Y.,Jiang,B.Y. and Chen,Y. (1995) Selective induction of cell death incancer cells by gallic acid. Biol. Pharm. Bull., 18, 1526–1530.
11.Serrano,A., Palacios,C., Roy,G., Cespon,C., Villar,M.L., Nocito,M. andGonzalez-Porque,P. (1998) Derivatives of gallic acid induce apoptosisin tumoral cell lines and inhibit lymphocyte proliferation. Arch.Biochem. Biophys., 350, 49–54.
12.van der Heijden,C.A., Janssen,P.J. and Strik,J.J. (1986) Toxicology ofgallates: a review and evaluation. Food Chem. Toxicol., 24, 1067–1070.
13.Roy,G., Lombardia,M., Palacios,C., Serrano,A., Cespon,C., Ortega,E.,Eiras,P., Lujan,S., Revilla,Y. and Gonzalez-Porque,P. (2000)Mechanistic aspects of the induction of apoptosis by lauryl gallate in themurine B-cell lymphoma line Wehi 231. Arch. Biochem. Biophys., 383,206–214.
14.Ortega,E., Sadaba,M.C., Ortiz,A.I., Cespon,C., Rocamora,A.,Escolano,J.M., Roy,G., Villar,L.M. and Gonzalez-Porque,P. (2003)Tumoricidal activity of lauryl gallate towards chemically induced skintumours in mice. Br. J. Cancer, 88, 940–943.
15.O’Connor,P.M., Jackman,J., Bae,I. et al. (1997) Characterization of the p53tumor suppressor pathway in cell lines of the National Cancer Instituteanticancer drug screen and correlations with the growth-inhibitory potencyof 123 anticancer agents. Cancer Res., 57, 4285–4300.
16.Korah,R.M., Sysounthone,V., Golowa,Y. and Wieder,R. (2000) Basicfibroblast growth factor confers a less malignant phenotype in MDA-MB-231 human breast cancer cells. Cancer Res., 60, 733–740.
17.Fairchild,C.R., Ivy,S.P., Kao-Shan,C.S., Whang-Peng,J., Rosen,N.,Israel,M.A., Melera,P.W., Cowan,K.H. and Goldsmith,M.E. (1987)Isolation of amplified and overexpressed DNA sequences fromadriamycin-resistant human breast cancer cells. Cancer Res., 47,5141–5148.
18.Del Bufalo,D., Biroccio,A., Leonetti,C. and Zupi,G. (1997) Bcl-2overexpression enhances the metastatic potential of a human breastcancer line. FASEB J., 11, 947–953.
19.Ruiz-Ruiz,M.C. and Lopez-Rivas,A. (1999) p53-mediated up-regulation ofCD95 is not involved in genotoxic drug-induced apoptosis of human breasttumor cells. Cell Death Differ., 6, 271–280.
20.Michalovitz,D., Halevy,O. and Oren,M. (1990) Conditional inhibition oftransformation and of cell proliferation by a temperature-sensitive mutantof p53. Cell, 62, 671–680.
21.Mechetner,E.B., Schott,B., Morse,B.S., Stein,W.D., Druley,T., Davis,K.A.,Tsuruo,T. and Roninson,I.B. (1997) P-glycoprotein function involvesconformational transitions detectable by differential immunoreactivity.Proc. Natl Acad. Sci. USA, 94, 12908–12913.
22.Ryan,K.M., Phillips,A.C. and Vousden,K.H. (2001) Regulation and functionof the p53 tumor suppressor protein. Curr. Opin. Cell Biol., 13, 332–337.
23.Nieves-Neira,W. and Pommier,Y. (1999) Apoptotic response tocamptothecin and 7-hydroxystaurosporine (UCN-01) in the 8 humanbreast cancer cell lines of the NCI Anticancer Drug Screen:multifactorial relationships with topoisomerase I, protein kinase C,Bcl-2, p53, MDM-2 and caspase pathways. Int. J. Cancer, 82, 396–404.
24.Gartel,A.L. and Tyner,A.L. (2002) The role of the cyclin-dependent kinaseinhibitor p21 in apoptosis. Mol. Cancer Ther., 1, 639–649.
25.Patel,T., Gores,G.J. and Kaufmann,S.H. (1996) The role of proteasesduring apoptosis. FASEB J., 10, 587–597.
26. Janicke,R.U., Sprengart,M.L., Wati,M.R. and Porter,A.G. (1998)Caspase-3 is required for DNA fragmentation and morphologicalchanges associated with apoptosis. J. Biol. Chem., 273, 9357–9360.
27. Janicke,R.U., Ng,P., Sprengart,M.L. and Porter,A.G. (1998) Caspase-3 isrequired for alpha-fodrin cleavage but dispensable for cleavage of otherdeath substrates in apoptosis. J. Biol. Chem., 273, 15540–15545.
28.Sharpe,J.C., Arnoult,D. and Youle,R.J. (2004) Control of mitochondrialpermeability by Bcl-2 family members. Biochim. Biophys. Acta, 1644,107–113.
29.Zamzami,N., Brenner,C., Marzo,I., Susin,S.A. and Kroemer,G. (1998)Subcellular and submitochondrial mode of action of Bcl-2-likeoncoproteins. Oncogene, 16, 2265–2282.
30.Calcabrini,A., Arancia,G., Marra,M., Crateri,P., Befani,O., Martone,A. andAgostinelli,E. (2002) Enzymatic oxidation products of spermine inducegreater cytotoxic effects on human multidrug-resistant colon carcinomacells (LoVo) than on their wild-type counterparts. Int. J. Cancer, 99, 43–52.
31.Wada,T. and Penninger,J.M. (2004) Mitogen-activated protein kinases inapoptosis regulation. Oncogene, 23, 2838–2849.
32.Tang,D., Wu,D., Hirao,A., Lahti,J.M., Liu,L., Mazza,B., Kidd,V.J.,Mak,T.W. and Ingram,A.J. (2002) ERK activation mediates cell cyclearrest and apoptosis after DNA damage independently of p53. J. Biol.Chem., 277, 12710–12717.
33.Zhu,H., Zhang,L., Wu,S., Teraishi,F., Davis,J.J., Jacob,D. and Fang,B.(2004) Induction of S-phase arrest and p21 overexpression by a smallmolecule 2[[3-(2,3-dichlorophenoxy)propyl] amino]ethanol incorrelation with activation of ERK. Oncogene, 23, 4984–4992.
34.Facchinetti,M.M., De Siervi,A., Toskos,D. and Senderowicz,A.M. (2004)UCN-01-induced cell cycle arrest requires the transcriptional induction ofp21(waf1/cip1) by activation of mitogen-activated protein/extracellularsignal-regulated kinase kinase/extracellular signal-regulated kinasepathway. Cancer Res., 64, 3629–3637.
35.Hsu,Y.L., Kuo,P.L., Lin,L.T. and Lin,C.C. (2005) Asiatic acid, atriterpene, induces apoptosis and cell cycle arrest through activation ofextracellular signal-regulated kinase and p38 mitogen-activated proteinkinase pathways in human breast cancer cells. J. Pharmacol. Exp.Ther., 313, 333–344.
Lauryl gallate-induced cell cycle alterations and apoptosis
1711
by guest on March 31, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

36.He,X., Wang,J., Guo,Z., Liu,Q., Chen,T., Wang,X. and Cao,X. (2005)Requirement for ERK activation in sinomenine-induced apoptosis ofmacrophages. Immunol. Lett., 98, 91–96.
37.Nguyen,T.T., Tran,E., Nguyen,T.H., Do,P.T., Huynh,T.H. and Huynh,H.(2004) The role of activated MEK-ERK pathway in quercetin-inducedgrowth inhibition and apoptosis in A549 lung cancer cells.Carcinogenesis, 25, 647–659.
38.Chang,G.C., Hsu,S.L., Tsai,J.R., Wu,W.J., Chen,C.Y. and Sheu,G.T.(2004) Extracellular signal-regulated kinase activation and Bcl-2downregulation mediate apoptosis after gemcitabine treatment partly viaa p53-independent pathway. Eur. J. Pharmacol., 502, 169–183.
39.Leonard,G.D., Fojo,T. and Bates,S.E. (2003) The role of ABC transportersin clinical practice. Oncologist, 8, 411–424.
40. Jodoin,J., Demeule,M. and Beliveau,R. (2002) Inhibition of the multidrugresistance P-glycoprotein activity by green tea polyphenols. Biochim.Biophys. Acta, 1542, 149–159.
41.Qian,F., Wei,D., Zhang,Q. and Yang,S. (2005) Modulation of P-glycoprotein function and reversal of multidrug resistance by (-)-epigallocatechin gallate in human cancer cells. Biomed. Pharmacother.,59, 64–69.
42.Kowaltowski,A.J., Cosso,R.G., Campos,C.B. and Fiskum,G. (2002)Effect of Bcl-2 overexpression on mitochondrial structure and function.J. Biol. Chem., 277, 42802–42807.
43.Sigal,A. and Rotter,V. (2000) Oncogenic mutations of the p53 tumorsuppressor: the demons of the guardian of the genome. Cancer Res., 60,6788–6793.
44.Oren,M., Damalas,A., Gottlieb,T., Michael,D., Taplick,J., Leal,J.F.,Maya,R., Moas,M., Seger,R., Taya,Y. and Ben-Ze’ev,A. (2002)Regulation of p53: intricate loops and delicate balances. Biochem.Pharmacol., 64, 865–871.
45.El-Deiry,W.S. (2003) The role of p53 in chemosensitivity andradiosensitivity. Oncogene, 22, 7486–7495.
46.Russo,T., Zambrano,N., Esposito,F., Ammendola,R., Cimino,F.,Fiscella,M., Jackman,J., O’Connor,P.M., Anderson,C.W. and Appella,E.(1995) A p53-independent pathway for activation of WAF1/CIP1 expression following oxidative stress. J. Biol. Chem., 270,29386–29391.
47.Chen,Z., Gibson,T.B., Robinson,F., Silvestro,L., Pearson,G., Xu,B.,Wright,A., Vanderbilt,C. and Cobb,M.H. (2001) MAP kinases. Chem.Rev., 101, 2449–2476.
48.Tamura,Y., Simizu,S. and Osada,H. (2004) The phosphorylation statusand anti-apoptotic activity of Bcl-2 are regulated by ERK and proteinphosphatase 2A on the mitochondria. FEBS Lett., 569, 249–255.
Received December 19, 2005; revised March 24, 2006;accepted March 31, 2006
A.Calcabrini et al.
1712
by guest on March 31, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















