
Inhibition of photosynthetic oxygen evolution by protonophoric uncouplers
11
Photosynthesis Research 46: 455-465, 1995. O 1995 KluwerAcademic Publishers. Printed in theNetherlands. Regular paper Inhibition of photosynthetic oxygen evolution by protonophoric uncouplers Vitaly D. Samuilov 1, Gernot Renger2,*, Vladimir Z. Paschenko 3, Alexander V. Oleskin 1, Michail V. Gusev 1, Olga N. Gubanova 1, Sergei S. Vasil'ev 3 & Eugene L. Barsky 1 IDepartment of Cell Physiology and Immunology, School of Biology, Moscow State University, Moscow 119899, Russian Federation; 2Max- Volmer-lnstitute for Biophysical and Physical Chemistry, Technical University Berlin, Strasse des 17. Juni 135, D-10623 Berlin, Germany; 3Department of Biophysics, School of Biology, Moscow State University, Moscow 119899, Russian Federation; *Author for correspondence Received 11 April 1995;accepted in revisedform 10 October 1995 Key words: Photosystem II, water oxidation, ADRY-agents, uncouplers, CCCP, PCP, TI'FB Abstract The protonophoric uncouplers carbonyl cyanide m-chlorophenylhydrazone (CCCP), 2,3,4,5,6-pentachlorophenol (PCP) and 4,5,6,7-tetrachloro-2-trifluoromethylbenzimidazole(TFFB) inhibited the Hill reaction with K3 [Fe(CN)6] (but not with SiMo) in chloroplast and cyanobacterial membranes (the 150 values were approx. 1-2, 4-6 and 0.04- 0.10 #M, respectively). The inhibition is due to oxidation of the uncouplers on the Photosystem II donor side (ADRY effect) and their subsequent reduction on the acceptor side, ie. to the formation of a cyclic electron transfer chain around Photosystem II involving the uncouplers as redox carriers. The relative amplitude of nanosecond chlorophyll fluorescence in chloroplasts was increased by DCMU or HQNO and did not change upon addition of uncouplers, DBMIB or DNP-INT; the HQNO effect was not removed by the uncouplers. The uncouplers did not inhibit the electron transfer from reduced TMPD or duroquinol to methylviologen which is driven by Photosystem I. These data show that CCCP, PCP and TI"FB oxidized on the Photosystem II donor side are reduced by the membrane pool of plastoquinone (Qp) which is also the electron donor for K3 [Fe(CN)6] in the Hill reaction as deduced from the data obtained in the presence of inhibitors. Inhibition of the Hill reaction by the uncouplers was maximum at the pH values corresponding to the pK of these compounds. It is suggested that the tested uncouplers serve as proton donors, and not merely as electron donors on the oxidizing side of Photosystem II. Abbreviations: ADRY- acceleration of the deactivation reactions of the water-splitting enzyme system Y; ANT2p- 2-(3-chloro-4-trifluoromethyl) anilino-3,5-dinitrothiophene; CCCP-carbonyl cyanide m-chlorophenylhydrazone; DBMIB - 2,5-dibromo-3-methyl 6-isopropyl-p-benzoquinone; DCMU - 3-(3,4-dichlorophenyl)- 1,1-dimethylurea; DNP-INT - 2-iodo-6-isopropyl-3-methyl 2' ,4,4'-trinitrodiphenyl ether; DPC - 1,5-diphenylcarbazide; DPIP - 2,6- dichlorophenolindophenol; FCCP-carbonyl cyanide p-trifuoromethoxyphenylhydrazone;FeCy-potassium ferri- cyanide; HQNO- 2-n-heptyl-4-hydroxyquinolineN-oxide; (MN)4 - the tetranuclear Mn cluster of water oxidizing complex; P680-photoactive Chl of the reaction center of Photosystem II; PCP-2,3,4,5,6-pentachlorophenol; PS-photosystem; QA and QB-primary and secondary plastoquinones of PS II; Qc and Qz-plastoquinone binding sites in the cytochrome b/f complex; Qp-membrane pool of plastoquinone; SiMo-sodium silico- molybdate; TMPD - N,N ~,N~-tetramethyl-p-phenylenediamine;T/'FB - 4,5,6,7-tetrachloro-2-trifluoromethylbenz- imidazole; WOC- water oxidizing complex; Yz - tyrosine- 161 of the Photosystem II DI polypeptide
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Inhibition of photosynthetic oxygen evolution by protonophoric uncouplers

Photosynthesis Research 46: 455-465, 1995. O 1995 KluwerAcademic Publishers. Printed in the Netherlands.
Regular paper
Inhibi t ion of photosynthet ic oxygen evolut ion by protonophor ic uncouplers
Vitaly D. Samuilov 1, Gernot Renger 2,*, Vladimir Z. Paschenko 3, Alexander V. Oleskin 1, Michail V. Gusev 1, Olga N. Gubanova 1, Sergei S. Vasil'ev 3 & Eugene L. Barsky 1 I Department of Cell Physiology and Immunology, School of Biology, Moscow State University, Moscow 119899, Russian Federation; 2Max- Volmer-lnstitute for Biophysical and Physical Chemistry, Technical University Berlin, Strasse des 17. Juni 135, D-10623 Berlin, Germany; 3Department of Biophysics, School of Biology, Moscow State University, Moscow 119899, Russian Federation; *Author for correspondence
Received 11 April 1995; accepted in revised form 10 October 1995
Key words: Photosystem II, water oxidation, ADRY-agents, uncouplers, CCCP, PCP, TI'FB
A b s t r a c t
The protonophoric uncouplers carbonyl cyanide m-chlorophenylhydrazone (CCCP), 2,3,4,5,6-pentachlorophenol (PCP) and 4,5,6,7-tetrachloro-2-trifluoromethylbenzimidazole (TFFB) inhibited the Hill reaction with K3 [Fe(CN)6] (but not with SiMo) in chloroplast and cyanobacterial membranes (the 150 values were approx. 1-2, 4-6 and 0.04- 0.10 #M, respectively). The inhibition is due to oxidation of the uncouplers on the Photosystem II donor side (ADRY effect) and their subsequent reduction on the acceptor side, ie. to the formation of a cyclic electron transfer chain around Photosystem II involving the uncouplers as redox carriers. The relative amplitude of nanosecond chlorophyll fluorescence in chloroplasts was increased by DCMU or HQNO and did not change upon addition of uncouplers, DBMIB or DNP-INT; the HQNO effect was not removed by the uncouplers. The uncouplers did not inhibit the electron transfer from reduced TMPD or duroquinol to methylviologen which is driven by Photosystem I. These data show that CCCP, PCP and TI"FB oxidized on the Photosystem II donor side are reduced by the membrane pool of plastoquinone (Qp) which is also the electron donor for K3 [Fe(CN)6] in the Hill reaction as deduced from the data obtained in the presence of inhibitors. Inhibition of the Hill reaction by the uncouplers was maximum at the pH values corresponding to the pK of these compounds. It is suggested that the tested uncouplers serve as proton donors, and not merely as electron donors on the oxidizing side of Photosystem II.
Abbreviations: ADRY- acceleration of the deactivation reactions of the water-splitting enzyme system Y; ANT2p- 2-(3-chloro-4-trifluoromethyl) anilino-3,5-dinitrothiophene; CCCP-carbonyl cyanide m-chlorophenylhydrazone; DBMIB - 2,5-dibromo-3-methyl 6-isopropyl-p-benzoquinone; DCMU - 3-(3,4-dichlorophenyl)- 1,1-dimethylurea; DNP-INT - 2-iodo-6-isopropyl-3-methyl 2' ,4,4'-trinitrodiphenyl ether; DPC - 1,5-diphenylcarbazide; DPIP - 2,6- dichlorophenolindophenol; FCCP-carbonyl cyanide p-trifuoromethoxyphenylhydrazone; FeCy-potassium ferri- cyanide; HQNO- 2-n-heptyl-4-hydroxyquinoline N-oxide; (MN)4 - the tetranuclear Mn cluster of water oxidizing complex; P680-photoactive Chl of the reaction center of Photosystem II; PCP-2,3,4,5,6-pentachlorophenol; PS-photosystem; QA and QB-primary and secondary plastoquinones of PS II; Qc and Qz-plastoquinone binding sites in the cytochrome b/f complex; Qp-membrane pool of plastoquinone; SiMo-sodium silico- molybdate; TMPD - N,N ~,N~-tetramethyl-p-phenylenediamine; T/'FB - 4,5,6,7-tetrachloro-2-trifluoromethylbenz- imidazole; W O C - water oxidizing complex; Yz - tyrosine- 161 of the Photosystem II DI polypeptide

456
Introduction
In all O2-evolving photosynthesizing organisms the key steps of water cleavage take place within the PS II complex that acts as a water-plastoquinone oxido- reductase (for recent reviews, see Debus 1992; Renger 1993). The overall reaction comprises a sequence of univalent oxidation steps at a manganese containing functional unit referred to as the water oxidizing com- plex (WOC). A redox active tyrosine of polypeptide D 1 (symbolized by Yz and identified as Tyr 161) mediates the electron transfer from the WOC to P680 +. The dif- ferent redox states of the WOC are symbolized by Si, where the index i represents the number of stored oxi- dizing equivalents. 02 is released after attaining redox state $4 which rapidly converts into So. The redox states $2 and $3 relax to $1 via reduction by endoge- nous reductants in the time domain from seconds to a few minutes, whereas So becomes slowly oxidized to $1 by the oxidized form yOX of the redox active tyro- sine of polypeptide D2 (for a recent detailed analysis of these reactions, see Messinger et al. 1993).
The reaction pattern can be modified selective- ly by different types of exogenous agents: (i) small hydrophilic reductants like NH2NH2 and NH2OH leading to 'super-reduced' states S_1 and S-2 (see Messinger and Renger 1993 and refs. therein), (ii) lipophilic electron donors like tetraphenylboron that compete effectively with water for oxidizing redox equivalents (Erixon and Renger 1974), and (iii) ADRY substances causing an accelerated decay ofredox states $2 and $3 of the WOC (Renger et al. 1973b; Hanssum et al. 1985).
The latter type reactants comprise a number of chemicals with different structures like anilinothio- phenes (Renger et al. 1973b; Hanssum et al. 1985), phenylhydrazone derivatives (Etienne 1974), nitro- phenols (Renger 1972), indophenols (Vater 1973) and diphenyl amines (Oettmeier and Renger 1980). As a common structural element, all ADRY agents so far identified contain an acidic OH- or NH-group and act as potent protonophoric uncouplers. In general, ADRY agents exert different effects depending on their con- centration used. At sufficiently low concentrations, these substances specifically accelerate the $2 and $3 decay without any effect on the electron transfer from Yz to P680 (Renger et al. 1989). Stoichiometric con- siderations based on measurements of the average oxy- gen yield under repetitive flash excitation led to the conclusion that ADRY agents act as mobile catalysts of a cyclic electron flow giving rise to $2 and $3 reduction
by endogenous electron carriers (Renger et al. 1973b, Hanssum et al. 1985). Interestingly, the most powerful species even act at substoichiometric amounts of about 1 ADRY molecule per 10 PS II complexes (Hanssum et al. 1985). The idea of an ADRY agent induced cycle is supported by the finding that the electron flow to exogenous electron acceptors like methylviologen (Renger et al. 1973b) or DPIP (Packham and Bar- ber 1984) becomes diminished. At higher concentra- tions additional effects arise as reflected by several phenomena: (a) light-induced oxidation of carotenoids (Velthyus 1981) and of chlorophyll a (Yamashita et al. 1969), (b) transformation of cytochrome b-559 from its high to its low potential form (Cramer and Whitmarsh 1977; Maroc and Garnier 1979), and (c) inhibition of electron flow in chloroplasts and cyanobacterial mem- branes under light-saturating conditions (Renger 1975; Barsky et al. 1991a).
It has been shown that ADRY agents are redox- active reducing $2 and $3 in normal chloroplasts (Renger et al. 1973b; Hanssum et al. 1985) and Yz °x in samples deprived of their oxygen evolution capacity (Renger and Reuter 1982; Ghanotakis et al. 1982). The oxidized forms of ADRY agents become rereduced by endogenous redox components probably by plastohy- droquinone at the level of Qp (Samuilov and Barsky 1993).
The goal of this work is to study inhibitory effects of the phenylhydrazone derivative CCCP on the electron transport and to compare them with those elicited by two other protonophoric uncouplers of benzimidazole and phenolic-type compounds.
Materials and methods
Chloroplasts from the leaves of 12 to 15 day old pea or wheat seedlings were isolated as reported previously (Barsky et al. 1991a,b). Thylakoid membranes from cyanobacterium Anacystis nidulans were prepared as in Barsky et al. (1988). Chromatophores were isolated from Rhodospirillum rubrum as described by Isaev et al. (1970). In the experiments, the chloroplasts were incubated in medium I containing 0.4 M sucrose, 35 mM NaC1, 50 mM Tricine-KOH (pH 7.8), or medium II containing 0.5 M sucrose, 0.5 M sodium citrate, 50 mM NaH2PO4 (pH 7.0), 20 mM KC1 and 2 mM MgC12; thylakoids - in medium II without MgC12; the chromatophores - in a medium containing 0.25 M sucrose and 50 mM Tris-HC1 (pH 7.6).

Light-dependent oxygen evolution or uptake by chloroplasts (Cchl = 20--30/~g/ml) incubated in medi- um I or by cyanobacterial membranes (Cchl = 15-20 #g/ml) and oxygen uptake by chromatophores (Cchl = 20 #g/ml) was recorded polarographically with a Clark-type closed platinum electrode at a white light intensity of --~ 103 W m -2. 2 mM K3[Fe(CN)6], 0.05 mM SiMo + 2 mM K3 [Fe(CN)6] or 0.2 mM methylvio- logen were employed as electron acceptors. 5 mM sodi- um ascorbate + 0.1 mM TMPD or 0.2 mM duroquinol were used as electron donors. Duroquinol was prepared by reduction with sodium borohydride as described by Izawa and Pan (1978).
Light-induced reduction of K3[Fe(CN)6] (added at concentration 0.2 mM) was measured using the Uvidec-4 Jasco spectrophotometer at 420 nm (Barsky et al. 1988).
Generation of ApH in the illuminated chloro- plasts was followed by fluorescence changes of 9- aminoacridine as described by Barsky et al. (1981). The concentration of 9-aminoacridine was 20/~M.
The decay kinetics of the chlorophyll fluorescence from chloroplasts incubated in medium II at a chloro- phyll content of 200-300 #g/ml were measured with a laser picosecond fluorometer as described previously (Barsky et al. 1991b). The kinetics monitored in the spectral region with A > 650 nm were fitted by a sum of three exponential components with lifetimes of 100 ps, 400 ps and 1.2 ns. The relative amplitude of the 1.2 ns fluorescence component was analysed in the exper- iments of this study. The chloroplasts were exposed to constant white light of low non saturating intensity (1-2 W m -2) or high intensity (103 W m -2) which saturates photosynthetic electron-transfer.
Results
Figure 1A shows that CCCP (translocating H + across the membranes) and gramicidin D (forming ion- conducting channels) suppress the light-induced ApH generation in pea chloroplasts. Both compounds stimu- lated the light saturated rate of photosynthetic electron transfer from H20 to K3[Fe(CN)6]. The stimulatory effect of gramicidin reached its maximum at a con- centration of approx. 2 #M and remained virtually constant up to tenfold concentrations (20 ~tM, see Fig. 1). On the other hand CCCP inhibited the Hill reaction at elevated concentrations. Similar features were pre- viously observed with other protonophoric uncouplers like S 13 and 1799 (Renger 1975). A comparison of the
457
data in Fig. 1B also reveals that gramicidin was more efficient than CCCP in stimulating the light saturated rate of 02 evolution.
CCCP failed to stimulate 02 evolution in chloro- plasts where the A#H generation was suppressed by gramicidin D; in this case CCCP only exerted an inhibitory effect (Fig. 2A). Stoichiometric consider- ations readily show that the observed inhibition cannot be explained by a donor function of CCCP similar to that of tetraphenylboron (Erixon and Renger 1974) because at a concentration of 10 #M this agent would be exhausted after about 100 turnovers. Alternatively, CCCP could act as efficient electron donor provid- ed that the oxidized form becomes rapidly rereduced by endogenous components either from the accep- tor side or from other sources like chlorophyll a or caroteinoids. The latter alternative seems to be less attractive because only limited amounts of these com- ponents are available in the thylakoid membrane. It can be definitely excluded on the basis of the data presented in Fig. 2A showing that CCCP suppressed 02 evolution and K3[Fe(CN)6] photoreduction to the same extent. The CCCP concentration causing 50% inhibition (I50) was about 2 ~M in both cases. Pho- toreduction of K3 [Fe(CN)6] would be less suppressed by CCCP than O2 evolution, if endogenous electron donors distinct from water were involved in the reac- tion. In order to further characterize this possibility, experiments were performed with SiMo as electron acceptor.
The data presented in Fig. 2A show that CCCP was practically without effect on the SiMo mediated light- induced 02 evolution within the concentration range tested. This observation could be explained in two dif- ferent ways: (a) the ADRY agents becomes complete- ly oxidized and is no longer able to act as an electron donor or (b) SiMo interacts with ADRY-agent by form- ing an inactive complex. In an attempt to discriminate between these alternatives further experiments were performed.
It was found that addition of K3[Fe(CN)6] and DCMU at pH 4.7 also eliminates inhibitory effects of CCCP (Fig. 2A). It is known that under this con- ditions QA is directly reoxidized by K3[Fe(CN)6] in a DCMU insensitive reaction (Itoh 1978). Likewise, K3[Fe(CN)6] also leads to the oxidation of the non- heme iron center on the acceptor side of PS II but with an extremely low rate (Wraight 1985; Petrouleas and Diner 1986). Therefore, in this case CCCP can be completely oxidized. According to Schansker and Van Rensen (1993) SiMo-binding site is located between

458
50 ¸
e~0
o
¢o
25-
0
I1)
0
- < -<
300"
A
~ CCCP G r a m i e i d i n D
Lj~
1 10 CCCP or g r a m i e i d i n D (pdvl)
::L
.o
2 0 > ©
b~,
0
150"
0 0 0.1 0.1
\o
B
\% 1 10 100
CCCP or g r a m i c i d i n D (/uM)
Fig. I. Effect of CCCP and gramicidin D on light-induced 9-aminoacridine (9AA) fluorescence quenching (A) and oxygen evolution (B) by pea chloroplasts.
0
0
o
O
100
50-
o [] 02 (Hi0 FeCy)
• FeCy (H20, -FeCy) .°;2°Zi2/ [] x 02 (I{20------,-FeCy)
100
• $~ ~ H20 ~FeCy
~ 50 o
0 12 0 2 CCCP (/uM) PCP (~fl~{)
Fig. 2. Effect of CCCP (A) and PCP (B) on photoreduction of FeCy and on light-induced oxygen evolution by pea chloroplasts incubated with gramicidin D (4 ttM), FeCy, SiMo and FeCy + DCMU (10 /zM). In the experiment with FeCy + DCMU tricine was replaced by 4-morpholineetanesulfonic acid (50 mM, pH 4.7). The 100% rates of 02 evolution with FeCy and SiMo were 250-300 and with FeCy + DCMU (at pH = 4.7) 50-60/~mol mg- l Chl per h. The 100% rate of FeCy reduction was 900-1100/zmol mg- l Chl per h.
the parallel helices of the D1 and D2 proteins of PS II and the acceptor-side non heme iron donates the electrons to SiMo. Indeed, under the conditions of our study the rate of oxygen evolution in presence of SiMo remains insensitive to DCMU (data not shown).
The similar decrease of the inhibitory effect of CCCP found both with SiMo and K3[Fe(CN)6] sug- gests that it is caused by the oxidation of the ADRY agent due to withdrawal of electrons from the internal carriers before the QB site. Accordingly, an inactive complex of CCCP with SiMo cannot account for the

insensitive electron flow. In order to exclude the pos- sibility that the phenomena reported here are specific for CCCP, other protonophoric uncouples were ana- lyzed.
Figure 2B presents the data on the PCP effect on 02 evolution with K3[Fe(CN)6] and SiMo. Like CCCP, PCP suppressed the Hill reaction with K3[Fe(CN)6] as acceptor (the 15o value was 4 to 6/2M) but only insignificantly decreased the reaction rate with SiMo. Analogous data were obtained with TI'tVB: the I50 value of blocking the Hill reaction with K3 [Fe(CN)6] was about 40 nM in chloroplasts (data not shown).
Uncouplers inhibit the photosynthetic electron transfer not only in chloroplasts, but also in the mem- branes of cyanobacteria. With respect to the inhibitory effect of protonophoric uncouplers, thylakoids isolat- ed from A. nidulans are of particular interest for two reasons: (i) they were shown to be incapable to gen- erate a light-dependent A/tH and therefore need not to be de-energized with gramicidin D, (ii) electron trans- fer with consecutive involvement of Photosystems II and I in the membranes is disrupted, due to plastocya- nine/cytochrome c-553 depletion during the isolation procedure. This enables us to rule out any contribu- tion of PSI to the reduction of the exogenous electron acceptors.
The results obtained show that the tested protonophoric compounds produce similar effects on the Hill reaction in the chloroplasts and cyanobacterial thylakoids (data not shown). Since the electron transfer between Photosystems II and I is disrupted in isolated cyanobacterial membranes, the Hill reagents in these membranes are not PS 1-reducible. Accordingly, one target-site of protonophoric uncouplers is inferred to be located on the PS II acceptor side.
In another approach to study possible effects on the acceptor side, the relative extent of chlorophyll fluorescence in the nanosecond time domain of the overall decay was measured as a sensitive probe for electron transfer disruption in PS II (Barsky et al. 1991b). The redox level of electron transport com- ponents was poised by simultaneous background illu- mination with white light of either low or high inten- sity (see Materials and methods). The results obtained are compiled in Table 1. It shows that the relative amplitude of the nanosecond decay at low light inten- sity markedly increased in response to an addition of DCMU and HQNO (inhibiting plastoquinone QB oxi- dation by plastoquinone Qp; Barton et al. 1983; Cohen and Barton 1983) and plastoquinone Qc reduction by cytochrome b (Jones and Whitmarsh 1988), but not
459
due to DBMIB and DNP-INT (both inhibiting plas- toquinol Qz oxidation in the cytochrome b/fcomplex (Jones and Whitmarsh 1988; Rich et al. 1991). Like- wise, CCCP, TYFB or PCP have virtually no effect (actually under these conditions the limited sensitivity does not permit to resolve the possibility of fluores- cence quenching). The uncouplers have practically no effect on the nanosecond fluorescence of chloroplasts incubated with HQNO. These data suggest that the site of action of the tested uncouplers on the PS II acceptor side is located subsequent to plastoquinone QB.
Continuous background illumination with white light of high intensity (1000 W m -x) increased the relative amplitude of the nanosecond fluorescence to a similar extent as DCMU under low intensity illu- mination. Addition of DCMU, HQNO, DBMIB and DNP-INT had no appreciable effect on the level of the nanosecond fluorescence component(s) under con- ditions of high intensity light. This finding indicates that the strong background light keeps QA reduced in most of the PS II centers: A strikingly different feature is observed in the presence of CCCP, i.e. a marked decrease of the fluorescence amplitude in the ns-time domain. TTFB was less efficient in quenching the flu- orescence, and PCP had no effect on it.
In order to check that the comparatively high pho- ton density of the actinic laser pulses does not marked- ly affect the results presented in Table 1, compara- tive experiments were performed using a single photon counting technique as described by Liu et al. (1993). Qualitatively, the same results were obtained (data not shown) thus ruling out serious effects due to the high photon density of the laser flashes used in this study (Barsky et al. 1991b).
In principle, the effect of CCCP (TFFB) on the nanosecond component of the fluorescence decay can be explained by two different types of quenching mech- anisms: a) increase of the steady state level of oxidized QA (acting as photochemical quencher) under strong background illumination, or b) formation of nonpho- tochemical quenchers, e.g. chlorophyll cation radicals. The influence of the latter type of quenching can be definitely excluded (see Discussion).
Based on the conclusion that CCCP diminishes the nanosecond fluorescence decay component under strong background illumination predominantly via a decrease of the steady level of QA it remains to be shown whether this effect is mainly owing to a cyclic electron flow competing with the linear electron trans- port or due to an impairment of the donor side capacity of PS II. Both alternatives can be distinguished by

460
Table 1. Effect of uncouplers and other agents on nanosecond chlorophyll fluorescence in wheat chloroplasts
Agent Concentration Amplitude of the Concentration Amplitude of (uM) ns component of (~M) ns component
fluorescence of fluorescence decay at low at high light
light intensity a intentsity a
None - 3 - 97 DCMU 25 100 25 100 HQNO 25 79 30 94 HQNO 50 90 n.d. DBMIB 5 3 10 94 DNP-INT 5 3 20 100 CCCP 20 3 20 52 TTFB 2 3 2 88 PCP 40 3 40 100 HQNO + CCCP 50 + 20 100 n.d. HQNO + TrFB 50 + 5 90 n.d. HQNO + PCP 50 + 40 100 n.d. DCMU + FeCy 25 + 6000 93 25 + 6000 100 DCMU -t- SiMo + FeCy 25 + 200 + 6000 3 25 + 200 + 6000 3 FeCy n.d. 6000 97 SiMo + FeCy n.d. 200 + 6000 2
aValues are given as arbitrary units. n.d. = not determined.
determining the dependency of the 150 values on the actinic l ight intensity. In the former case of a com- peting electron cycle with an efficient rate strongly
depending on the ADRY agent concentration, the Is0 value is expected to decrease drastically at low light intensity where the rate of the linear electron flow is l imited by the photon flux density. In contrast to this behaviour, the 150 value should remain virtually inde- pendent of the actinic light intensity i f a specific block of the electron transport chain is induced by a certain compound (the small effect expected to originate from the coupling via a common plastoquinone pool, see Siggel et al, (1972) can be neglected under the condi- tions of this study).
Figure 3 shows the results obtained by using sub- stances with different modes of action. As expected for inhibitors, in the case of DCMU and HQNO the 150 value is practically invariant to the intensity of the actinic light. On the other hand, however, the Is0 val- ues of CCCP, T r F B and PCP exhibit a pronounced dependence on light intensity. The increase of 15o with increasing intensity generally matched the light satu- ration curve of the linear electron flow. These find-
125- 125
~v*lO0
o
75
50
o
25
/: /
x / x x x _ _ A
[] Oxygen evolution
+~ I50 for: • CCCP
• DCMU + PCP
f x HQNO ~ TTFB
m'o 100
75
50
25
d5 go 7's 1 bo 12E Light intensity (arbitrary units)
Fig. 3. Effect of light intensity on 02 evolution coupled with FeCy reduction and on the I5o values for DCMU, HQNO and uncouplers in pea chloroplasts incubated with 5/zM gramicidin D. The 100% rates of 02 evolution were 250-300 #mol mg -1 Chl per h. I50 for DCMU HQNO, CCCP, TTFB and PCP at maximum light intensity were 0.02, 2, 2, 0.11 and 5 tzM, respectively.
ings clearly show that the suppression of 02 evolution and K3 [Fe(CN)6] reduction rate by the protonophoric
O
125"
o ° , . .q
o b E)
02
o
100- /f× / /k ×\
75- d
5O
25 x Without addi t ions
[] CCCP • TTFB
0 4 5 6 7
pH
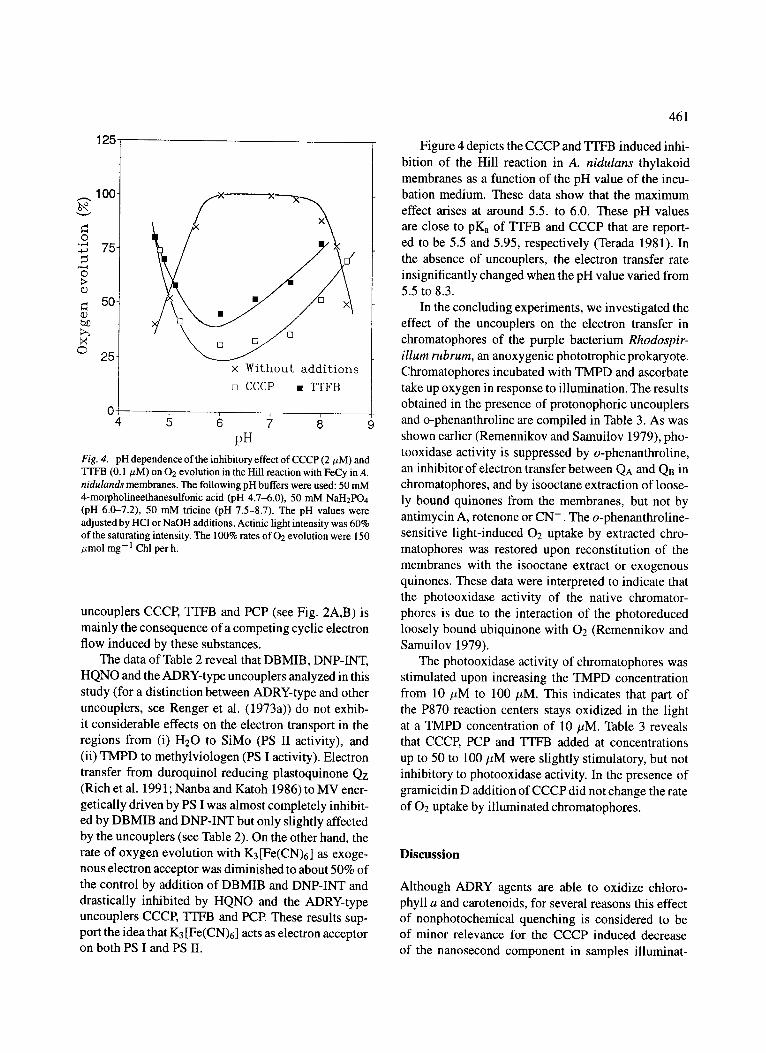
Fig. 4. pH dependence of the inhibitory effect of CCCP (2/~M) and TI'FB (0.1/zM) on 02 evolution in the Hill reaction with FeCy in A. nidulands membranes. The following pH buffers were used: 50 mM 4-morpholineethanesulfonic acid (pH 4.7-6.0), 50 mM NaH2PO4 (pH 6.0-7.2), 50 mM tricine (pH 7.5-8.7). The pH values were adjusted by HC1 or NaOH additions. Actinic light intensity was 60% of the saturating intensity. The 100% rates of 02 evolution were 150 tzmol mg- 1 Chl per h.
uncouplers CCCP, T1TB and PCP (see Fig. 2A,B) is mainly the consequence of a competing cyclic electron flow induced by these substances.
The data of Table 2 reveal that DBMIB, DNP-INT, HQNO and the ADRY-type uncouplers analyzed in this study (for a distinction between ADRY-type and other uncouplers, see Renger et al. (1973a)) do not exhib- it considerable effects on the electron transport in the regions from (i) H20 to SiMo (PS II activity), and (ii) TMPD to methylviologen (PS I activity). Electron transfer from duroquinol reducing plastoquinone Qz (Rich et al. 1991; Nanba and Katoh 1986) to MV ener- getically driven by PSI was almost completely inhibit- ed by DBMIB and DNP-INT but only slightly affected by the uncouplers (see Table 2). On the other hand, the rate of oxygen evolution with K3[Fe(CN)6] as exoge- nous electron acceptor was diminished to about 50% of the control by addition of DBMIB and DNP-INT and drastically inhibited by HQNO and the ADRY-type uncouplers CCCP, "Iq'FB and PCE These results sup- port the idea that K3 [Fe(CN)6] acts as electron acceptor on both PSI and PS II.
461
Figure 4 depicts the CCCP and TTFB induced inhi- bition of the Hill reaction in A. nidulans thylakoid membranes as a function of the pH value of the incu- bation medium. These data show that the maximum effect arises at around 5.5. to 6.0. These pH values are close to pKa of TrFB and CCCP that are report- ed to be 5.5 and 5.95, respectively (Terada 1981). In the absence of uncouplers, the electron transfer rate insignificantly changed when the pH value varied from 5.5 to 8.3.
In the concluding experiments, we investigated the effect of the uncouplers on the electron transfer in chromatophores of the purple bacterium Rhodospir- illum rubrum, an anoxygenic phototrophic prokaryote. Chromatophores incubated with TMPD and ascorbate take up oxygen in response to illumination. The results obtained in the presence of protonophoric uncouplers and o-phenanthroline are compiled in Table 3. As was shown earlier (Remennikov and Samuilov 1979), pho- tooxidase activity is suppressed by o-phenanthroline, an inhibitor of electron transfer between QA and QB in chromatophores, and by isooctane extraction of loose- ly bound quinones from the membranes, but not by antimycin A, rotenone or CN-. The o-phenanthroline- sensitive light-induced O2 uptake by extracted chro- matophores was restored upon reconstitution of the membranes with the isooctane extract or exogenous quinones. These data were interpreted to indicate that the photooxidase activity of the native chromator- phores is due to the interaction of the photoreduced loosely bound ubiquinone with 02 (Remennikov and Samuilov 1979).
The photooxidase activity of chromatophores was stimulated upon increasing the TMPD concentration from 10 #M to 100 #M. This indicates that part of the P870 reaction centers stays oxidized in the light at a TMPD concentration of 10 pM. Table 3 reveals that CCCP, PCP and TITB added at concentrations up to 50 to 100 pM were slightly stimulatory, but not inhibitory to photooxidase activity. In the presence of gramicidin D addition of CCCP did not change the rate of 02 uptake by illuminated chromatophores.
Discussion
Although ADRY agents are able to oxidize chloro- phyll a and carotenoids, for several reasons this effect of nonphotochemical quenching is considered to be of minor relevance for the CCCP induced decrease of the nanosecond component in samples illuminat-

462
Table 2. Effect of uncouplers and other agents on photosynthetic electron transport in pea chloroplasts
Agent Concentration
(#M) Rate of electron transport (%)
02 evolution 02 uptake 02 uptake 02 evolution
H20 TMPD + asc duroquinol 1-120
SiMo methylviologen methylviologen FeCy
(+ DCMU) (+DCMU)
DBMIB 1 96 98 5 45
DNP-INT 10 98 100 3 40
HQNO 10 96 95 90 10
CCCP 10 98 104 80 15 TTFB 1 89 98 97 7
PCP 30 92 94 101 8
Gramicidin D and DCMU concentrations were 4 and I #M, respectively. The 100% rate of electron transport in each column, starting from the left, was 100, 85, 75 and 115/~mol 02 mg -1 Chl per h, respectively.
Table 3. Photooxidase activity of R. rubrum chromatophores
Experiment No. Successive additions Final concentrations (mM) 02 uptake (%)
1 CCCP 0.01 105
CCCP 0.05 115
TMPD 0.1 280
CCCP 0.1 289
o-Phenanthroline 5.0 17
PCP 0.01 122
PCP 0.05 179
3 TTFB 0.01 108
TrFB 0.05 120
Gramicidin D 0.01 125
CCCP 0.01 125
Incubation medium contained 5 mM Tris-ascorbate and 10/zM TMPD. 100% of 02 uptake were 45 to 50 #mol nag -1 bacteriochlorophyll per h minus dark O2 uptake.
ed by strong white light. The quenching effect of the carotenoid radical is expected to be small because the absorption spectrum of this species (Yamashita et al. 1969; Schenk et al. 1982; Mathis and Rutherford 1984) permits only a rather poor overlapping with that of the chlorophyll emission. Furthermore, DCMU blocking of the Ka[Fe(CN)6] induced carotenoid cation radical formation in illuminated chloroplasts (Yamashita et al. 1969; Schenk et al. 1982) does not affect the extent of the nanosecond fluorescence. With respect to a chloro-
phyll cation radical it was previously reported (Renger et al. 1983) that its light induced formation is stimu- lated by K3[Fe(CN)6]. The data of Table 1, however, reveal that K3 [Fe(CN)6] does not cause any quenching effect. Simultaneously, SiMo+Ka[-Fe(CN)6] quenched nanosecond chlorophyll fluorescence both at low and high light intensity.
The results of this study show that the ADRY agents are capable to induce a cyclic electron flow in PS II. The idea of an ADRY agent-induced cycle is in line with

a previous report showing that the extent of ANT2p inhibition of methylviologen photoreduction (Renger et al. 1973b) and DCIP (Packham and Barber 1984), 02 evolution (Packham and Ford 1986) and photooxi- dation of the cytochrome b-559 (Packham and Barber 1984) depends on the intensity of the actinic light.
What redox components are involved in the K3 [Fe(CN)6] mediated electron transfer and the ADRY agent-induced cyclic electron flow? QpH2 is the most likely endogenous electron transport component that becomes oxidized by K3[Fe(CN)6] in PS II. The quinones QA and QB can be excluded as potent can- didates because nanosecond chlorophyll fluorescence was unaffected by K3[Fe(CN)6] (Table 1) and, further- more, the electron transfer reactions from QA and Q~ to K3 [Fe(CN)6] are rather slow probably due to shield- ing effects of the surrounding protein matrix. The rate of QA reoxidation can be stimulated by orders of mag- nitudes after modification of the protein environment by a mild trypsin treatment (Renger 1976). Taking into account that Qz is located closer to the inner lumenal side of the thylakoid membrane whereas Qp is more exposed to the outer (stromal) side and therefore more easily accessible to the impermeable K3[Fe(CN)6], it appears most likely that Qp is the target site of the K3 [Fe(CN)6] mediated electron transfer.
With respect to the ADRY type uncouplers, QA can be excluded as endogenous donor of the cycle as shown previously (Hanssum et al. 1985; Renger and Inoue 1983). Likewise, the QB site is probably also not the main site of electron funnelling into the uncou- pler induced cycle because CCCP (TITB, PCP) does not affect the nanosecond component of the fluores- cence decay in HQNO blocked chloroplasts under low intensity background illumination (see Table 1). On the other hand, the duroquinol oxidation was found to be resistant to the tested uncouplers (see Table 2), ie. these compounds act prior to Qz. It therefore appears most reasonable to assume that Qp also acts as endogenous electron source for the ADRY agent-induced cycle. The electron sink is located on the PS II donor side. The ADRY agent ANT2p is known to interact with the PS II donor side at two separate loci: (i) it catalyzes the $2 and $3 decay within the WOC (Renger 1973; Hanssum er al. 1985) (the classical ADRY effect), and (ii) it inhibits at higher concentrations the electron transfer between Yz and P680 + (Packham and Ford 1986).
Based on the above described considerations the following cycle of the ADRY-type uncoupler induced electron flow is proposed:
463
K3[Fe(CN)6]
T H20 "-~ (Mn)4 --~ Yz "-~ P680 --~ QA "-~QB --~ QP ~ Qz (b/f)
CCCP
TTFB
PCP
Likewise, the site of electron transfer to K3 [Fe(CN)6] is shown to be located at Q p .
It has been proposed that ADRY agents can mediate the electron transfer from the high potential cytochrome b-559, carotenoids and chlorophylls to $2 and S3 states or to yOX (Sayre and Homann 1979; Ghanotakis et al. 1982). However, the extent of the rereduction of the photooxidized cytochrome b-559 does not show a linear relationship with the inhibition degree of the DPIP reduction rate occurring with the ADRY agent (Packham and Barber 1984). In addition, the ANT2p inhibition of the electron transfer between Yz and P680 + was unaffected by changes of the actinic light intensity (Packham and Ford 1986). From these data they concluded that the ADRY agent-induced pho- tooxidation of carotenoids results from the inhibition of electron transfer between Yz and P680 + and not from the ADRY effect on the WOC. The same rea- soning applies to cytochrome b-559, an integral PS II component protecting it against photoinhibition, pos- sibly via a cyclic electron pathway around PS II (Heber et al. 1979; Buser et al. 1992). Thus, the reduced Qp seems to be the electron donor for the independent and competitive pathways of the electron transfer to the oxidized carotenoids, chlorophyll, cytochrome b-559 and ADRY agents.
The pH dependent inhibitory effect of CCCP and TTFB shown in Fig. 4 cannot be simply explained by a decrease of the 150 value of these compounds as was the case for the variation of 150 as a function of the actinic light intensity (see Fig. 3). The decline in the control rates of the electron transfer at pH 4.7 and 8.5 should enhance the inhibitory effect of CCCP and TITB. However, the experiment reveals just the opposite: the inhibitory effect of CCCP and TTFB is decreased at the indicated pH values. In this context, Fig. 4 appears to provide convincing evidence that the inhibition of the Hill reaction by uncouplers is maximal at pH values corresponding with their pKa values, i.e. to their maximum activity as proton conductors across the membranes (Terada 1981). The pH dependence of the protonophoric efficiency is therefore interpret- ed as evidence that both the neutral and the ionized

464
uncouplers forms are invo lved in induc ing membrane
conductance (Terada 1981). The slope at the left side
of the curve is steeper than that of the right side (Fig.
4), poss ib ly due to elevated access o f Q A to exogenous
K3[Fe(CN)6]. However, the rate of this reaction is low (cf. Fig. 2).
What are the impl icat ions of the correlation
be tween the properties of the tested compounds acting s imul taneous ly as A D R Y agents and H + conductors
(see also (Renger and Eckert (1980))? The redox tran-
si t ions S0--*SI, 81--+52, 52"+53 and 53---~(54)--+50 of the water oxid iz ing complex are accompanied by H + release with a s toichiometry that depends on the sam-
ple type and the experimental condi t ions (for review,
see Lavergne and Junge (1993). Most apparently, the
tested uncouplers , acting as A D R Y agents, br ing about the relaxat ion of the $2 and $3 states, operat ing not on ly as electron donors, but also as proton donors.
In contrast with PS II the tested uncouplers do not inhib i t photooxidase activity of the R. rubrum chro-
matophores. The probable reason for this phenomenon is that they cannot be oxidized by the redox chain com-
ponents of the chromatophores, inc luding P870 + with
an Em of about +0.5 V (Wil l iams et al. 1992).
Acknowledgements
This work was supported in part by a Grant from the
Internat ional Science Founda t ion and Russian Govern- men t (No. ML7300) and by a Grant from the Russian Founda t ion for Fundamenta l Research (No. 94-04- 12227a). S.S.V. wishes to acknowledge the financial support f rom Alexander von Humbold t Foundat ion.
The authors would like to thank A. Bergmann and A
Napiwotzki for per forming the single photon count ing
experiments .
References
Barsky EL, Gubanova ON and Samuilov VD (1988)The inhibitory effect of 2,6-di-tert-butyl-4-methylphenol on electron transfer in Photosystem II of isolated Anacystis nidulans membranes and pea chloroplasts. Biokhimiya 53:297-301
Barsky EL, Gubanova ON and Samuilov VD (1991a) Inhibition of photosynthetic electron transfer in chloroplasts by carbonyl- cyanide m-chlorophenylhydrazone. Biokhimiya 56:434-438
Barsky EL, Gusev MV, Nikitina KA and Samuilov VD (1981 ) Light- induced proton translocation through thylakoid and cytoplasmic membranes of Plectonema boryanum. Arch Microbiol 129:105- 108
Barsky EL, Vasil'ev SS, Paschenko VZ and Samuilov VD (1991b) Nanosecond fluorescence of chloroplasts as a probe for electron transfer disruption in Photosystem II. J Photochem Photobiol B: Biol 8:175-181
Barton JR, MacPeek WA and Cohen WS (1983) Interaction of 2-n-heptyl 4-hydroxyquinoline-N-oxide with Photosystem II in chloroplasts and subchloroplast particles. J Bioenerg Biomembr 15:93-104
Buser CA, Diner BA and Brudvig GW (1992) Photooxidation of cytochrome b559 in oxygen evolving Photosystem II. Biochem- istry 31:11449-11459
Cohen WS and Barton JR (1983) The use of 02-evolving subchloro- plast particles to study acceptor and inhibitor sites on the reducing side of Photosystem II. Z Naturforsch 38c: 793-798
Cramer WA and Whitmarsh J (1977) Photosynthetic cytochromes. Ann Rev Plant Physiol 28:133-172
Cheniae GM and Martin IF (1971) Effects of hydroxylamine on Photosystem II. I. Factors affecting the decay of 02 evolution. Plant Physiol. 47:568-575
Debus RJ (1992) The manganese and calciumions of photosynthetic oxygen evolution. Biochim Biophys Acta 1102:269-352
Erixon K and Renger G (1974) The action of tetraphenylboron as a system II electron donor and its effect on the decay of the electrical field across the thylakoid membrane. Biochim Biophys Acta 333:95-106
Etienne AL (1974) Effects of carbonyl cyanide m-chloro- phenylhydrazone and hydroxylamine on the Photosystem II electron exchange mechanism in 3-(3,4-dichlorophenyl)-l.1- dimethyhuea treated algae and chloroplasts. Biochim Biophys Acta 333:497-508
Ghanotakis DE Yerkes C and Babcock GT (1982) The role of reagents accelerating the deactivation reactions of water-splitting enzyme system Y (ADRY reagents) in destabilizing high- potential oxidizing equivalents generated in chloroplast Photo- system II. Biochim Biophys Acta 682:21-31
Hanssum B, Renger G and Weiss W (1985) Studies on the reaction mechanism of tetraphenylboron at the Photosystem II donor side in isolated spinach chloroplasts. Biochim Biophys Acta 808: 210- 220
Heber U, Kirk MR and Boardman NK (1979) Photoreactions of cytochrome b559 and cyclic electron flow in Photosystem II of intact chloroplasts. Biochim Biophys Acta 546:292-306
Isaev PI, Liberman EA, Samuilov VD, Skulachev VP and Tsofina LM (1970) Conversion of biomembrane-produced energy into electric form. III. Chromatophores of Rhodospirillum rubrum. Biochim Biophys Acta 216:22-29
Itoh S (1978) Membrane surface potential and the reactivity of the system II primary electron acceptor to charged electron carriers in the medium Biochim Biophys Acta 504:324-340
Izawa S and Pan RL (1978) Photosystem I electron transport and phosphorylation supported by electron donation to the plasto- quinone region. Biochem Biophys Res Comrnuns 83:1171-1177
Jones RW and Whitmarsh J (1988) Inhibition of electron transfer and the electrogenic reaction in the cytochrome b/fcomplex by 2-n- nonyl 4-hydroxyquinoline N-oxide (NQNO) and 2,5-dibromo- 3-methyl-6-isopropyl-p-benzoquinone (DBMIB). Biochim Bio- phys Acta 933:258-268
Lavergne J and Junge W (1993) Proton release during the redox cycle of the water oxidase. Photosynth Res 38:279-296
Liu B, Napiwotzki A, Eckert H J, Eichler HJ and Renger G (1993) Studies on the recombination kinetics of the radical pair P680 + Pheo- in isolated PS II core complexes from spinach. Biochim Biophys Acta 1142:129-138

Maroc J and Gamier J (1979) Photooxidation of the cytochrome b-559 in the presence of various substituted 2-anilinothiophenes and of some other compounds in Chlamydomonas reinhardtii. Biochim Biophys Acta 548:374--385
Mathis P and Rutherford AW (1984) Effect of phenolic herbicides on the oxygen-evolving side of Photosystem II. Formation of the carotenoid cation. Biochim Biophys Acta 767:217-222
Messinger J and Renger G (1993) Generation, oxidation by the oxidized form of the tyrosine of polypeptide D2, and possible electronic configuration of the mdox states So, S_ 1, and S-2 of the water oxidase in isolated spinach thylakoids. Biochemistry 32:9378-9386
Messinger J, Schr6der WP and Renger G (1993) Structure-function relations in Photosystem II. Effects of temperature and chaotropic agents on the period four oscillation of flash-induced oxygen evolution. Biochemistry 32:7658-7668
Nanba M and Katoh S (1986) The site and mechanism of duroquinol oxidation by the cytochrome b6-fcomplex in Synechococcus sp. Biochim Biophys Acta 851:484.-490
Oettmeier W and Renger G (1980) The function of diphenylammes as modifiers of Photosystem II electron transport in isolated spinach chloroplasts. Biochim Biophys Acta 593:113-124
Packham NK and Barber J (1984) The light-intensity depen- dence of the efficacy of 2-(3-chloro-4-trifluoromethyl)-anilino- 3,5-dmitrothiophene (Ant 2p) to inhibit the Photosystem 2 reac- tions of chloroplasts. Biochem J 221:513-520
Packham NK and Ford RC (1986) Deactivation of the Photosystem II oxidation (S) states by 2-(3-chloro-4-trifluoromethyl)anilino- 3,5-dinitrothiophene (ANT2p) and the putative role of carotenoid. Biochim Biophys Acta 852:183-190
Petrouleas V and Diner BA (1986) Identification of Q4oo, a high- potential electron acceptor of Photosystem II, with the iron of the quinone-iron acceptor complex. Biochim Biophys Acta 849: 264-275
Remennikov VG and Samuilov VD (1979) Photooxidase activity of Rhodospirillum rubrum chromatophores and reaction center complexes. The role of non-cyclic electron transfer in generation of the membrane potential. Biochim Biophys Acta 546: 220-235.
Renger G (1972) Requirement of an acidic proton in substances which act as accelerators of the deactivation reactions in the water-splitting enzyme system of photosynthesis. FEB S Lett 23: 321-324
Renger G (1975) The action of 5-chloro-3-tert.butyl 2'-chloro-4'- nitro-salicylanilide and ~,c~'-bis(hexafluoroacetonyl)aceton on the water-splitting enzyme system Y in spinach chloroplasts. FEBS Lett 52:30-32
Renger G (1976) The induction of a high resistance to 3-(3,4- dicblorophenyl t, 1-dimethylurea (DCMU) of oxygen evolution in spinach chloroplasts by trypsin treatment. FEBS Lett 69: 225- 230
Renger G (1993) Water cleavage by solar radiation - an inspiring challenge of photosynthesis research. Photosynth Res 38: 229- 247
Renger G, Bouges-Bocquet B and Biachel KH (1973 a) The mod- ification of the trapping properties of the photosynthetic water splitting enzyme system by a new class of biological effectors. Bioenerg 4:491-505
465
Renger G, Bouges-Bocquet B and Delosme R (1973b) Studies on the ADRY agent-induced mechanism of the discharge of the holes trapped in the photosynthetic watersplitting enzyme system Y. Biochim Biophys Acta 292:796-807
Renger G and Eckert H J (1980) Studies on the structural and func- tional organization of water cleavage by visible light in photo- synthesis. Bioelectrochem Bioenerg 7:101-124
Renger G, Eckert H J and V61ker M (1989) Studies on the electron transfer from Tyr-161 of polypeptide D-1 to P680 + in PS 1I membrane fragments from spinach. Photosynth Res 22:247-256
Renger G and Inoue Y (1983) Studies on the mechanism of ADRY agents (agents accelerating the deactivation reactions of water- spliting enzyme system Y) on thermoluminescence emission. Biochim Biophys Acta 725: 146-154.
Renger G, Koike H, Yuasa M and Inoue Y (1983) Studies on the mechanism of the fluorescence decline induced by strong actinic light in PS II particles under different redox conditions. FEBS Lett 163:89-93
Renger G and Reuter R ( 1982) The destabilization of oxidizing redox equivalents of system II by ADRY-reagents in normal and Tris- washed chloroplasts. Photobiochem Photobiophys 3:317-325
Renger G (1993) Water cleavage by solar radiation - an inspiring challenge of photosynthesis. Photosynth Res 38:229-247
Rich PR, Madgwick SA and Moss DA (1991) The interactions of duroquinol, DBMIB and NQNO with the chloroplast cytochrome bfcomplex. Biochim Biophys Acta 1058:312-328
Samuilov VD and Barsky EL (1993) Interaction of carbonyl cyanide m-chlorophenylhydrazone with the Phorosystem II acceptor side. FEBS Lett 320:118-120
Sayre RT and Homann PH (1979) A light dependent oxygen con- sumption induced by PS II of isolated chloroplasts. Arch Biochem Biophys 196: 525-533,
Schansker G and Van Rensen JJS (1993) Characterization of the complex interaction between the electron acceptor silicomolyb- date and Photosystem II. Photosynth Res 37:165-175
Schenk CC, Diner B, Mathis P and Satoh K (1982) Flash induced carotenoid radical cation formation in Photosystem II. Biochim Biophys Acta 680:216-227
Siggel U, Renger G, Stiehl HH and Rumberg B (1972) Evidence for electronic and ionic interaction between electron transport chains in chloroplasts. Biochim Biophys Acta 256:328-335
Terada H (1981) The interaction of highly active uncouplers with mitochondria. Biochim Biophys Acta 639:225-242
Vater J (1973) The action of indophenols and nitrophenols on the deactivation reactions in the water-splitting system of photosyn- thesis. Biochim Biophys Acta 292:786-795
Velthyus BR (1981) Carotenoid and cytochrome b-559 reactions in Photosystem II in the presence of tetraphenylboron. FEBS Lett 126:272-276
Williams JC, Alden RG, Murchison HA, Peloquin JM, Woodbury NN and Allen JP (1992) Biochemistry 31 : 11029-11037
Wraight CA (1985) Modulation of herbicide-binding by the redox state of Q400, an endogenous component of Photosystem II. Bioc'him Biophys Acta 809:320-330
Yamashita K, Konishi K, Itoh M and Shibata K (1969) Photo- bleaching of carotenoids related to the electron transport in chloroplasts. Biochim Biophys Acta 680:286-227




















