
Design of amino acid sequences to fold into Cα-model proteins
Upload
independentCategory
view
1download
0
Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 4 | Page 297
Survivin expression and subcellular translocation of Smac in apoptosis inhibitionActa Biochim Biophys Sin (2008): 297-303 | © 2008 Institute of Biochemistry and Cell Biology, SIBS, CAS | All Rights Reserved 1672-9145http://www.abbs.info; www.blackwellpublishing.com/abbs | DOI: 10.1111/j.1745-7270.2008.00401.x
Inhibition of fibroblast growth factor 2-induced apoptosis involves survivinexpression, protein kinase Cα activation and subcellular translocation of Smac inhuman small cell lung cancer cells
Desheng Xiao#, Kuansong Wang#, Jianhua Zhou*, Huiqiu Cao, Zhenghao Deng, Yongbin Hu, Xiahui Qu,and Jifang Wen
Department of Pathology, Xiangya School of Medicine, Central South University, Changsha 410013, China
Received: September 17, 2007 Accepted: February 13, 2008This work was supported by the Natural Science Foundation of HunanProvince (No. 06JJ2098)#These authors contributed equally to this work*Corresponding author: Tel, 86-731-2650406; Fax, 86-731-2650406;E-mail, [email protected]
To investigate the mechanism by which fibroblast growthfactor 2 (FGF-2) inhibits apoptosis in the human small celllung cancer cell line H446 subjected to serum starvation,apoptosis was evaluated by flow cytometry, Hoechst 33258staining, caspase-3 activity, and DNA fragmentation.Survivin expression induced by FGF-2 and protein kinaseCα (PKCα) translocation was detected by subcellular frac-tionation and Western blot analysis. In addition, FGF-2-in-duced release of Smac from mitochondria to the cytoplasmwas analyzed by Western blotting and immunofluorescence.FGF-2 reduced apoptosis induced by serum starvation andup-regulated survivin expression in H446 cells in a dose-dependent and time-dependent manner, and inhibited caspase-3 activity. FGF-2 also inhibited the release of Smac frommitochondria to the cytoplasm induced by serum starvationand increased PKCα translocation from the cytoplasm tothe cell membrane. In addition, PKC inhibitor inhibited theexpression of survivin. FGF-2 up-regulates the expressionof survivin protein in H446 cells and blocks the release ofSmac from mitochondria to the cytoplasm. PKCα regulatedFGF-2-induced survivin expression. Thus, survivin, Smac,and PKCα might play important roles in the inhibition ofapoptosis by FGF-2 in human small cell lung cancer cells.
Keywords FGF-2; survivin; Smac; PKC; apoptosis;lung cancer
Lung cancer remains one of the leading causes of cancer
death and, of all lung cancers, small cell lung cancer(SCLC) is the most malignant histological type [1].Currently, the primary therapy used for patients with SCLCis chemotherapy. Despite rapid progress in improving theefficacy of chemotherapy, it has a very poor prognosisas it is refractory to most chemotherapeutic agents. The5-year survival rate is the lowest among all primary lungcancers [1,2].
A high expression of fibroblast growth factor 2 (FGF-2) was associated with poor overall survival in SCLC [3].FGF-2 induces a broad spectrum of drug resistance byinhibiting apoptosis in various tumor types, but the mo-lecular mechanism is not clear [4]. Apoptosis is regulatedby a complex balance of signals, including the pro-apoptotic factors, such as p53 and Bax, and the anti-apoptotic factors, such as bcl-2 family members and mem-bers of the inhibitor of apoptosis protein (IAP) family [5−7]. Survivin, an IAP family member, is involved in bothinhibition of apoptosis and control of cell division. Its anti-apoptotic function is related to its ability to inhibit caspases[8−13]. Smac/DIABLO, a recently identified protein re-leased from mitochondria after apoptotic stimuli, bindsIAPs, allowing caspase activation and cell death [14−17].
In the present study, we investigated the effect of FGF-2 on the expression of survivin, and the subcellular loca-tion of Smac in the human SCLC cell line H446, to clarifythe molecular mechanisms involved in repression ofapoptosis by this agent.
Materials and Methods
ReagentsAntibodies against survivin, β-actin, and protein kinase C(PKCα) were purchased from Santa Cruz Biotechnology(Santa Cruz, USA), the neutralizing anti-FGF-2 antibody

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 4 | Page 298
Survivin expression and subcellular translocation of Smac in apoptosis inhibition
was from Upstate Biotechnology (Lake Placid, USA), andthe antibody against Smac was from R&D Systems(Minneapolis, USA). Recombinant human FGF-2 wasfrom PeproTech EC (London, UK). The SABC-Cy3 Im-munofluorescence reagent kit was from Boster BiologicalTechnology Company (Wuhan, China). The BCA ProteinAssay reagent kit was from Pierce (Rockford, USA) andthe In situ cell death detection kit was purchased fromRoche Diagnostics (Indianapolis, USA). The caspase-3activity assay kit was from R&D Systems, and calphostinC was from Sigma-Aldrich (St. Louis, USA). The mem-brane protein extraction kit was from Keygen (Nanjing,China).
Cell lines and cell cultureThe human SCLC cell line H446 was a generous gift fromProf. Zhiming He of the Tumor Research Institute of Cen-tral South University (Changsha, China). Cell cultures weremaintained in RPMI 1640 (Gibco Biocult, Paisley, UK)supplemented with 10% (V/V) fetal calf serum, 50 U/mlpenicillin, and 50 µg/ml streptomycin in an atmosphere of5% CO2/95% humidified air at 37 ºC. The incubation me-dium was changed every 1 or 2 d, and subculture wascarried out every 2 or 3 d according to the cell confluence.For experimental purposes, the cells were grown in me-dium contain 1% serum for 36 h then treated with FGF-2 for the indicated time.
Flow cytometryApproximately 5×106 cells were immediately fixed in 70%ethanol and stored at 4 ºC in phosphate-buffered saline(PBS) for fluorescence-activated cell sorting. Flowcytometry analysis was carried out on a FACStar flowcytometer (Becton Dickinson, Franklin Lakes, USA). His-tograms of the number of cells at each fluorescence in-tensity increment (on a logarithmic scale) were recordedfor 10,000 cells per sample.
Hoechst 33258 stainingHoechst 33258 staining was carried out as previously de-scribed [18,19]. Briefly, cells were collected by centrifu-gation and the pellets were washed twice with PBS. Pel-lets were then resuspended in PBS and stained with 10µg/ml Hoechst 33258 (Sigma) for 10 min at roomtemperature. Morphological evaluation of nuclear conden-sation and fragmentation was carried out immediately witha Nikon Microphot-FXA fluorescence microscope (Tokyo,Japan). The percentage of apoptotic nuclei was deter-mined by counting the number of apoptotic nuclei forevery 300 cells in a particular microscopic field, dividing
that number by 300, and multiplying it by 100.
Terminal deoxynucleotidyl transferase-mediateddigoxigenin-dUTP nick-end labeling (TUNEL) assayA modified TUNEL assay was carried out using the Insitu cell death detection kit according to the manufacturer’sprotocol. The morphological changes were observed withthe Nikon Microphot-FXA fluorescence microscope.TUNEL-positive cells revealed nuclei with light greenfluorescence. Positive cells per field were counted.
Caspase-3 activity assaySCLC cells were harvested and washed with cold PBSfollowed by centrifugation. The cell pellet was incubatedin buffer A [20 mM HEPES potassium salt (pH 7. 5), 10mM KCl, 1.5 mM MgCl2, 1.0 mM EDTA disodium salt,and 250 mM sucrose] for 10 min at 4 ºC. The cell lysateswere centrifuged at 12,000 g for 15 min and the proteinconcentration was determined by the Bradford method.Quantitative detection of caspase-3 activity in cellular ly-sates was carried out using a caspase-3 activity assay kitaccording to the manufacturer’s instructions.
Western blot analysisTotal cellular extracts were obtained by lysing the cells inlysis buffer and membrane protein was isolated as de-scribed by Li and colleagues [20]. Western blot analysiswas carried out as previously described [21]. Briefly, 60mg cell extract from each sample was separated by elec-trophoresis on a 10% sodium dodecyl sulfate-polyacryla-mide gel and transferred to an Immobilon P polyvinylidenedifluoride membrane (Millipore, Billerica, USA) with a semi-dry electroblotter (PatentStorm, Washington, USA). Af-ter blocking, the membrane was incubated overnight withprimary antibodies to PKCα, survivin, and Smac in 5%milk, followed by incubation with the corresponding horse-radish peroxidase-conjugated anti-goat or anti-rabbit IgG.The bands were detected by diaminobenzidine coloration.
Subcellular fractionH446 cells were centrifuged in 1.5 ml Eppendorf tubes,resuspended in 100 ml ice-cold RSB hypotonic buffer [10mM Tris-HCl (pH 7.4) containing 2.5 mM MgCl2, 10 mMNaCl, 1 µl/ml phenylmethylsulfonyl fluoride, and 1 µl/mldithiothreitol], and incubated on ice for 10 min. The cellsuspension was homogenized in a precooled Douncehomogenizer, then resuspended in 2.5× MS buffer [5 mMTris-HCl (pH 7.4) containing 210 mM mannitol, 70 mMsucrose, 1 mM EDTA, 1 µl/ml phenylmethylsulfonylfluoride, and 1 µl/ml dithiothreitol]. Cells were then cen-

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 4 | Page 299
Survivin expression and subcellular translocation of Smac in apoptosis inhibition
trifuged at 3750 g at 4 ºC for 5 min. The supernatant wasremoved and resuspended, then centrifuged at 15,000 gat 4 ºC for 20 min. The supernatant, containing the cyto-plasmic fraction, was separated from the pellet, contain-ing the mitochondrial fraction.
ImmunofluorescenceAfter treatment, SCLC cells were centrifuged in 1.5 mlEppendorf tubes. The cell pellet was resuspended in Ca2+-free and Mg2+-free PBS containing 4% formaldehyde for15 min at room temperature. Cells were washed twice inCa2+-free and Mg2+-free PBS and were resuspended inblocking buffer [PBS containing 3% bovine serum albu-min (BSA)] for 30 min. After three washes in PBS, thecorresponding primary antibody was added (in PBS con-taining 1% BSA) and incubated for 1 h at 37 ºC, thenovernight at 4 ºC. After three washes in PBS, thebiotinylated secondary antibody was added (in PBS con-taining 1% BSA) and incubated for 30 min at roomtemperature. After three washes in PBS, the Cy3-labeledstreptavidin was added (in PBS containing 1% BSA) andincubated for 30 min at room temperature in the dark.Samples were then washed four times in PBS, tiled ontoglass slides, and observed with the Nikon Microphot-FXAfluorescence microscope.
Statistical analysisThe results were statistically evaluated by the least sig-nificant difference, Student-Newman-Keuls’, and χ2-testsusing SPSS version 12.0 software (SPSS, Chicago, USA).A P value of less than 0.05 was regarded as significant.
Results
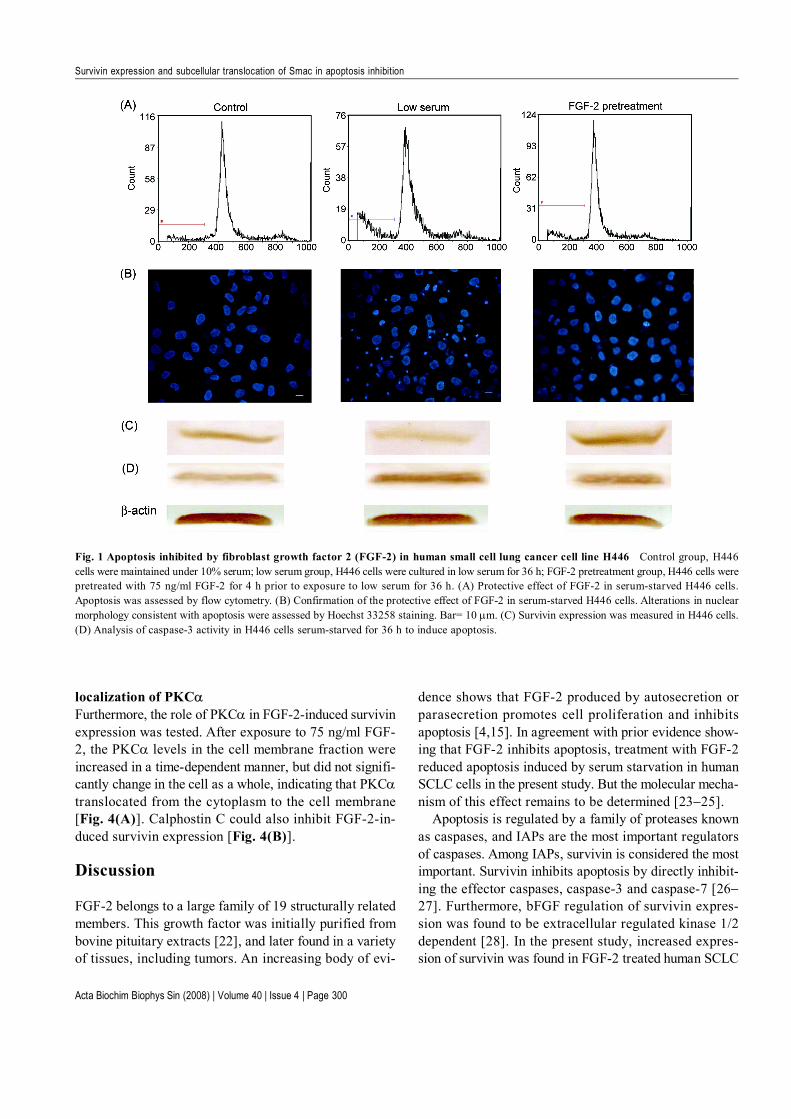
Influence of fibroblast growth factor 2 (FGF-2) onapoptosis, survivin expression and caspase-3 activityin human small cell lung cancer cell line H446To confirm the effect of FGF-2 on apoptosis, the cellswere examined by flow cytometry. As shown in Fig. 1(A),after serum starvation, the percentage of apoptotic cellsincreased from 7.29% to 22.6%. FGF-2 exposure reducedapoptosis to 12.5%. The effect of FGF-2 exposure onthe appearance of apoptotic morphology was further in-vestigated by nuclear staining and DNA fragmentationassay. After the cells were deprived of serum for 36 h,the typical morphological changes of apoptotic cells(condensed chromatin and fragmented nucleus) wereapparent, an effect that was attenuated by FGF-2 expo-sure [Fig. 1(B)]. Control cells had minimal apoptotic nu-clei (2.42%±0.85%), whereas the starved cells showed a
significant increase (14.51%±2.68%). FGF-2 exposuredecreased the percentage of apoptotic nuclei (6.13%±1.89%) in serum-starved cells, but not to the levels seen inthe control cells. The TUNEL assay, which assessed DNAfragmentation, showed that the number of apoptotic cells/high power was 2.52±1.15 in the control cultures, 7.65±2.21 in the serum-deprived cultures, and 5.32±1.25 in theserum-deprived cells pretreated with FGF-2. The differ-ence between the serum-starved group and the FGF-2stimulation group was significant (P<0.05). Therefore,FGF-2 could prevent apoptosis of H446 cells. After thecells were serum-starved for 36 h, caspase-3 activity wasincreased and survivin expression was decreased, butFGF-2 pretreatment inhibited the caspase-3 activity andupregulated the survivin expression [Fig. 1(C,D)].
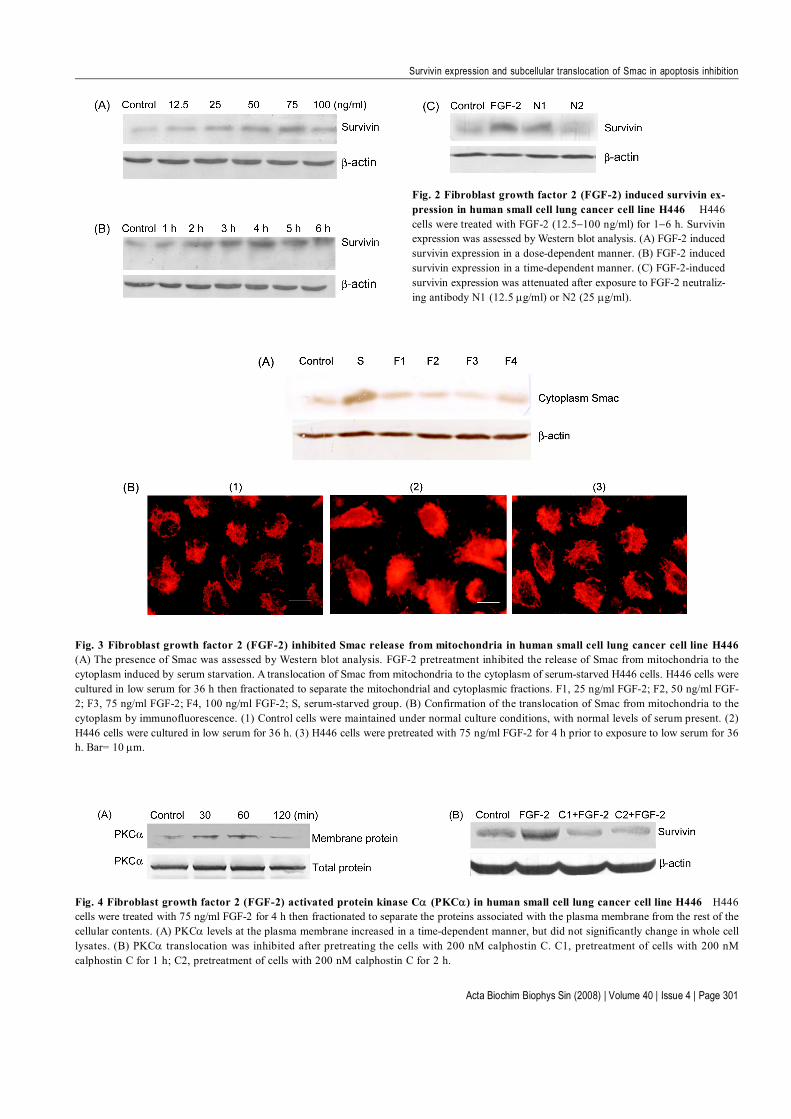
FGF-2 up-regulated expression of survivin protein butinhibited caspase-3 activityAlthough H446 cells express the basal level of survivin,the levels in FGF-2 treated groups were significantly higher.After treatment with FGF-2 for 4 h, survivin expressionwas induced in a dose-dependent manner [from 12.5 to75 ng/ml; Fig. 2(A)]. When the cells were exposed to 12.5 ng/ml FGF-2, the expression of survivin protein peakedat 4 h [Fig. 2(B)]. The FGF-2-induced survivin expres-sion was attenuated after treatment with FGF-2 neutraliz-ing antibody [Fig. 2(C)].
FGF-2 inhibited Smac release from mitochondriaNext, we investigated the effect of FGF-2 on Smac frommitochondria in H446 cells. In this study, we identifiedthat serum starvation for 36 h could induce Smac releasefrom mitochondria in H446 cells, compared with the con-trol in which cells were incubated in medium contain 1%serum. Furthermore, when the H446 cells werepreincubated with recombinant human FGF-2 (25–100ng/ml) for 4 h, the release of Smac protein from the mito-chondria to the cytoplasm was inhibited from approxi-mately 53% to approximately 62% [Fig. 3(A)]. The ex-pression level of Smac protein in the cytoplasm of theFGF-2 pretreated cells was lower than in the serum-starved cells (P<0.05).
Immunofluorescent detection of Smac in untreated cellsshowed a punctate staining consistent with mitochondriallocalization. This staining became diffuse on serumstarvation. However, Smac localization was not altered incells pretreated with 75 ng/ml FGF-2 [Fig. 3(B)], con-firming the Western blot results.
FGF-2-induced survivin expression involved in altered
Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 4 | Page 300
Survivin expression and subcellular translocation of Smac in apoptosis inhibition
Fig. 1 Apoptosis inhibited by fibroblast growth factor 2 (FGF-2) in human small cell lung cancer cell line H446 Control group, H446cells were maintained under 10% serum; low serum group, H446 cells were cultured in low serum for 36 h; FGF-2 pretreatment group, H446 cells werepretreated with 75 ng/ml FGF-2 for 4 h prior to exposure to low serum for 36 h. (A) Protective effect of FGF-2 in serum-starved H446 cells.Apoptosis was assessed by flow cytometry. (B) Confirmation of the protective effect of FGF-2 in serum-starved H446 cells. Alterations in nuclearmorphology consistent with apoptosis were assessed by Hoechst 33258 staining. Bar= 10 µm. (C) Survivin expression was measured in H446 cells.(D) Analysis of caspase-3 activity in H446 cells serum-starved for 36 h to induce apoptosis.
localization of PKCαFurthermore, the role of PKCα in FGF-2-induced survivinexpression was tested. After exposure to 75 ng/ml FGF-2, the PKCα levels in the cell membrane fraction wereincreased in a time-dependent manner, but did not signifi-cantly change in the cell as a whole, indicating that PKCαtranslocated from the cytoplasm to the cell membrane[Fig. 4(A)]. Calphostin C could also inhibit FGF-2-in-duced survivin expression [Fig. 4(B)].
Discussion
FGF-2 belongs to a large family of 19 structurally relatedmembers. This growth factor was initially purified frombovine pituitary extracts [22], and later found in a varietyof tissues, including tumors. An increasing body of evi-
dence shows that FGF-2 produced by autosecretion orparasecretion promotes cell proliferation and inhibitsapoptosis [4,15]. In agreement with prior evidence show-ing that FGF-2 inhibits apoptosis, treatment with FGF-2reduced apoptosis induced by serum starvation in humanSCLC cells in the present study. But the molecular mecha-nism of this effect remains to be determined [23−25].
Apoptosis is regulated by a family of proteases knownas caspases, and IAPs are the most important regulatorsof caspases. Among IAPs, survivin is considered the mostimportant. Survivin inhibits apoptosis by directly inhibit-ing the effector caspases, caspase-3 and caspase-7 [26−27]. Furthermore, bFGF regulation of survivin expres-sion was found to be extracellular regulated kinase 1/2dependent [28]. In the present study, increased expres-sion of survivin was found in FGF-2 treated human SCLC
Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 4 | Page 301
Survivin expression and subcellular translocation of Smac in apoptosis inhibition
Fig. 3 Fibroblast growth factor 2 (FGF-2) inhibited Smac release from mitochondria in human small cell lung cancer cell line H446(A) The presence of Smac was assessed by Western blot analysis. FGF-2 pretreatment inhibited the release of Smac from mitochondria to thecytoplasm induced by serum starvation. A translocation of Smac from mitochondria to the cytoplasm of serum-starved H446 cells. H446 cells werecultured in low serum for 36 h then fractionated to separate the mitochondrial and cytoplasmic fractions. F1, 25 ng/ml FGF-2; F2, 50 ng/ml FGF-2; F3, 75 ng/ml FGF-2; F4, 100 ng/ml FGF-2; S, serum-starved group. (B) Confirmation of the translocation of Smac from mitochondria to thecytoplasm by immunofluorescence. (1) Control cells were maintained under normal culture conditions, with normal levels of serum present. (2)H446 cells were cultured in low serum for 36 h. (3) H446 cells were pretreated with 75 ng/ml FGF-2 for 4 h prior to exposure to low serum for 36h. Bar= 10 µm.
Fig. 2 Fibroblast growth factor 2 (FGF-2) induced survivin ex-pression in human small cell lung cancer cell line H446 H446cells were treated with FGF-2 (12.5−100 ng/ml) for 1−6 h. Survivinexpression was assessed by Western blot analysis. (A) FGF-2 inducedsurvivin expression in a dose-dependent manner. (B) FGF-2 inducedsurvivin expression in a time-dependent manner. (C) FGF-2-inducedsurvivin expression was attenuated after exposure to FGF-2 neutraliz-ing antibody N1 (12.5 µg/ml) or N2 (25 µg/ml).
Fig. 4 Fibroblast growth factor 2 (FGF-2) activated protein kinase Cα (PKCα) in human small cell lung cancer cell line H446 H446cells were treated with 75 ng/ml FGF-2 for 4 h then fractionated to separate the proteins associated with the plasma membrane from the rest of thecellular contents. (A) PKCα levels at the plasma membrane increased in a time-dependent manner, but did not significantly change in whole celllysates. (B) PKCα translocation was inhibited after pretreating the cells with 200 nM calphostin C. C1, pretreatment of cells with 200 nMcalphostin C for 1 h; C2, pretreatment of cells with 200 nM calphostin C for 2 h.

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 4 | Page 302
Survivin expression and subcellular translocation of Smac in apoptosis inhibition
cells. Neutralizing FGF-2 with anti-FGF2 antibody attenu-ated the expression of survivin significantly. In addition,FGF-2 pretreatment inhibited caspase-3 activity, indicat-ing that FGF-2-mediated survivin expression inhibitsapoptosis through direct interaction with caspase-3 inH446 cells.FGF-2 functions through three signal transductionpathways, the mitogen-activated protein kinase (MAPK)pathway, the phosphatidylinositol 3-kinase pathway, andthe PKC pathway [29,30]. Sequence analysis of survivinsuggests that the survivin gene contains three PKC activesites (Thr21, Ser88, and Thr127). Therefore, we hypoth-esized that FGF-2 might up-regulate survivin protein ex-pression through the PKC signal transduction pathway.FGF-2 induces survivin expression in a mammary cancercell line by activating the MAPK signaling pathway andrecruitment of c-myc [21,22,31]. Other studies haveshown activation of the MAPK signaling pathway in FGF-2-induced proliferation of coronary artery smooth musclecells [29], an effect modulated by PKCδ. In oral carci-noma TSCCa cells, survivin expression was down-regu-lated and caspase-3 expression was up-regulated after in-hibition of the PKCα signaling pathway with staurosporine,a potent kinase inhibitor [26]. Thus, PKC is capable ofaffecting cell death in cancer cells, and could therefore beinvolved in FGF-2-related carcinogenesis and development.Our results indicate that FGF-2 induces translocation ofPKCα from the cytoplasm to the cell membrane of H446cells. Treatment with calphostin C, a specific PKCinhibitor, prevented this translocation. In addition, FGF-2-induced survivin expression was significantly inhibited.Our findings suggest that FGF-2 might regulate survivinexpression in SCLC cells by altering the localization ofPKCα. Pardo et al. reported that FGF-2 increased ex-pression of the anti-apoptotic proteins XIAP and Bcl-XLthrough the PKCε signaling pathway to inhibit apoptosisof lung cancer cells [32]. However, we are the first toreport that FGF-2 up-regulated survivin to inhibit apoptosisin SCLC cells.
There are a large number of known factors that canenhance apoptosis by eliminating the inhibitory effects ofIAPs such as survivin on caspases, such as Smac [19,33]. Smac, a direct IAP binding protein with a low iso-electric point (DIABLO), normally localizes within mito-chondria and is released from mitochondria into the cyto-sol during apoptosis. In the present study, treatment ofserum-starved H446 cells with FGF-2 inhibited Smac re-lease from mitochondria, indicating that Smac is involvedin FGF-2-prevented apoptosis.
In conclusion, to our knowledge, these data show for
References
1 Yip D, Harper PG. Predictive and prognostic factors in small celllung cancer: current status. Lung Cancer 2000, 28: 173−185
2 Debatin K. Activation of apoptosis pathways by anticancertreatment. Toxicol Lett 2000, 112−113: 41−48
3 Ruotsalainen T, Joensuu H, Mattson K, Salven P. High pretreat-ment serum concentration of basic fibroblast growth factor is apredictor of poor prognosis in small cell lung cancer. CancerEpidemiol Biomarkers Prev 2002, 11: 1492−1495
4 Song S, Wientjes MG, Gan Y, Au JL. Fibroblast growth factors: anepigenetic mechanism of broad spectrum resistance to anticancerdrugs. Proc Natl Acad Sci USA 2000, 97: 8658−8663
5 Shi YA. A structural view of mitochondria-mediated apoptosis.Nature Struct Biol 2001, 8: 394−401
6 Adams JM, Cory S. Life-or-death decisions by the Bcl-2 proteinfamily. Trends Biochem Sci 2001, 26: 61−66
7 Deng Y, Lin Y, Wu X. TRAIL-inducing apoptosis requires Bax-dependent mitochondrial release of Smac/DIABLO. Genes Dev2002, 16: 33−45
8 Giodini A, Kallio MJ, Wall NR, Gorbsky GJ, Tognin S, MarchisioPC, Symons M et al. Regulation of microtubule stability and mi-totic progression by survivin. Cancer Res 2002, 62: 2462−2467
9 Li F, Ambrosini G, Chu EY, Plescia J, Tognin S, Marchisio PC,Altieri DC. Control of apoptosis and mitotic spindle checkpoint bysurvivin. Nature 1998, 396: 580−584
1 0 Ambrosini, G, Adida C, Sirugo G, Altieri DC. Induction of apoptosisand inhibition of cell proliferation by survivin gene targeting. J BiolChem 1998, 273: 11177−11182
1 1 Liu JR, Opipari AW, Tan L, Jiang Y, Zhang Y, Tang H, Nuñez G.Dysfunctional apoptosome activation in ovarian cancer: implica-tions for chemoresistance. Cancer Res 2002, 62: 924−931
1 2 Shin S, Sung BJ, Cho YS, Kim HJ, Ha NC, Hwang JI, Chung CW etal. An anti-apoptotic protein human survivin is a direct inhibitor ofcaspase-3 and -7. Biochemistry 2001, 40: 1117−1123
1 3 Tamm, I, Wang Y, Sausville E, Scudiero DA, Vigna N, Oltersdorf T,Reed JC. IAP-family protein survivin inhibits caspase activity andapoptosis induced by Fas (CD95), Bax, caspases, and anticancerdrugs. Cancer Res 1998, 58: 5315−5320
1 4 Du C , Fang M, Li Y, Li L, Wang X. Smac, a mitochondrial protein
the first time that FGF-2 inhibits apoptosis in human SCLCcells. The underlying mechanism for the inhibition ofapoptosis is closely related to the up-regulation of thesurvivin protein and the reduced release of Smac frommitochondria to the cytoplasm. We conclude that survivin,Smac, and PKCα might play important roles in inhibitingapoptosis by FGF-2 treatment in human SCLC cells. Theseresults indicate new targets for adjuvant therapy to im-prove the effectiveness of conventional therapies for SCLC.
Acknowledgements
We thank Prof. Weijun Cai (Central South University,Changsha, China) for his comments and suggestions dur-ing the preparation of this manuscript.

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 4 | Page 303
Survivin expression and subcellular translocation of Smac in apoptosis inhibition
that promotes cytochrome c-dependent caspase activation by elimi-nating IAP inhibition. Cell 2000, 102: 33−42
1 5 Sekimura A, Konishi A, Mizuno K, Kobayashi Y, Sasaki H, Yano M,Fukai I et al. Expression of Smac/DIABLO is a novel prognosticmarker in lung cancer. Oncol Rep 2004, 11: 797−802
1 6 McNeish IA, Bell S, McKay T, Tenev T, Marani M, Lemoine NR.Expression of Smac/DIABLO in ovarian carcinoma cells inducesapoptosis via a caspase-9-mediated pathway. Exp Cell Res 2003,286: 186−198
1 7 Yang L, Mashima T, Sato S, Mochizuki M, Sakamoto H, Yamori T,Oh-Hara T et al. Predominant suppression of apoptosome by in-hibitor of apoptosis protein in non-small cell lung cancer H460cells: therapeutic effect of a novel polyarginine-conjugated Smacpeptide. Cancer Res 2003, 63: 831−837
1 8 Battaglia M, Pozzi D, Grimaldi S, Parasassi T. Hoechst 33258staining for detecting mycoplasma contamination in cell cultures: amethod for reducing fluorescence photobleaching. BiotechHistochem 1994, 69: 152−156
1 9 Pardo OE, Lesay A, Arcaro A, Lopes R, Ng BL, Warne PH, McNeishIA et al. Fibroblast growth factor 2-mediated translational controlof IAPs blocks mitochondria l release of Smac/DIABLO andapoptosis in small cell lung cancer cells. Mol Cell Biol 2003, 23:7600−7610
2 0 Li MC, Liang DS, Xu ZM, Lei LS, Wang QB, Yang SQ, Sun LS.Effects of Ganoderma polysaccharides on PKC and PKA activitiesin murine T cells. China Pharmacy 2001, 12: 78−79
2 1 Wang KS, Hu ZL, Li JH, Xiao DS, Wen JF. Enhancement of meta-static and invasive capacity of gastric cancer cells by transforminggrowth factor-beta1. Acta Biochim Biophys Sin 2006, 38: 179−186
2 2 Okada-Ban M, Thiery JP, Jouanneau J. Fibroblast growth factor-2.Int J Biochem Cell Biol 2000, 32: 263−267
2 3 Takanami I, Tanaka F, Hashizume T, Kikuchi K, Yamamoto Y,Yamamoto T, Kodaira S. The basic fibroblast growth factor and its
receptor in pulmonary adenocarcinomas: an investigation of their ex-pression as prognostic markers. Eur J Cancer 1996, 32A: 1504−1509
2 4 Takanami I, Tanaka F, Hashizume T, Kodaira S. Tumor angiogen-esis in pulmonary adenocarcinoma: relationship with basic fibro-blast growth factor, its receptor, and survival. Neoplasma 1997,44: 295−298
2 5 Lowe SW, Lin AW. Apoptosis in cancer. Carcinogenesis 2000, 21:485−495
2 6 Nakamura M, Tsuji N, Asanuma K, Kobayashi D, Yagihashi A,Hirata K, Torigoe T et a l . Survivin as a predictor of cis-diamminedichloroplatinum sensitivity in gastric cancer patients.Cancer Sci 2004, 95: 44−51
2 7 Altieri DC. Validating survivin as a cancer therapeutic target. NatRev Cancer 2003, 3: 46−54
2 8 Teh SH, Hill AK, Foley DA, McDermott EW, O’Higgins NJ, YoungLS. COX inhibitors modulate bFGF-induced cell survival in MCF-7breast cancer cells. J Cell Biochem 2004, 91: 796−807
2 9 Adida C, Crotty PL, McGrath J, Berrebi D, Diebold J, Altieri DC.Developmentally regulated expression of the novel cancer anti-apoptosis gene survivin in human and mouse differentiation. Am JPathol 1998, 152: 43−49
3 0 Verdecia M, Huang H, Dutil E, Kaiser DA, Hunter T, Noel JP.Structure of the human anti-apoptotic protein reveals a dimericarrangement. Nat Struct Biol 2000, 7: 602−608
3 1 Cosgrave N, Hill AD, Young LS. Growth factor-dependent regula-tion of survivin by c-myc in human breast cancer. J Mol Endocrinol2006, 37: 377−390
3 2 Pardo OE, Wellbrock C, Khanzada UK, Aubert M, Arozarena I,Davidson S, Bowen F et al. FGF-2 protects small cell lung cancercells from apoptosis through a complex involving PKCepsilon, B-Raf and S6K2. EMBO J 2006, 25: 3078−3088
3 3 Chai J, Du C, Wu JW, Kyin S, Wang X, Shi Y. Structural andbiochemical basis of apoptotic activation by Smac/DIABLO. Na-ture 2000, 406: 855−862
Copyright © 2022 FDOKUMEN




















