
Cell-Specific Upregulation of Survivin after Experimental Traumatic Brain Injury in Rats
13
JOURNAL OF NEUROTRAUMA Volume 21, Number 9, 2004 © Mary Ann Liebert, Inc. Pp. 1183–1195 Cell-Specific Upregulation of Survivin after Experimental Traumatic Brain Injury in Rats ERIK A. JOHNSON, 1,2 STANISLAV I. SVETLOV, 1,2 BRIAN R. PIKE, 1,2 PAUL J. TOLENTINO, 1,4 GERALD SHAW, 1,2,5 KEVIN K.W. WANG, 1,2,6 RONALD L. HAYES, 1,2,4,6 and JOSE A. PINEDA 1,3 ABSTRACT In this study, we examined the expression and cellular localization of survivin and proliferating cell nuclear antigen (PCNA) after controlled cortical impact traumatic brain injury (TBI) in rats. There was a remarkable and sustained induction of survivin mRNA and protein in the ipsilateral cortex and hippocampus of rats after TBI, peaking at five days post injury. In contrast, both survivin mRNA and protein were virtually undetectable in craniotomy control animals. Concomitantly, ex- pression of PCNA was also significantly enhanced in the ipsilateral cortex and hippocampus of these rats with similar temporal and spatial patterns. Immunohistochemistry revealed that survivin and PCNA were co-expressed in the same cells and had a focal distribution within the injured brain. Further analysis revealed a frequent co-localization of survivin and GFAP, an astrocytic marker, in both the ipsilateral cortex and hippocampus, while a much smaller subset of cells showed co-lo- calization of survivin and NeuN, a mature neuronal marker. Neuronal localization of survivin was observed predominantly in the ipsilateral cortex and contralateral hippocampus after TBI. PCNA protein expression was detected in both astrocytes and neurons of the ipsilateral cortex and hip- pocampus after TBI. Collectively these data demonstrate that the anti-apoptotic protein survivin, previously characterized in cancer cells, is abundantly expressed in brain tissues of adult rats sub- jected to TBI. We found survivin expression in both astrocytes and a sub-set of neurons. In addi- tion, the expression of survivin was co-incident with PCNA, a cell cycle protein. This suggests that survivin may be involved in regulation of neural cell proliferative responses after traumatic brain injury. Key words: astrocyte; neuron; PCNA; survivin; traumatic brain injury 1183 1 Center for Traumatic Brain Injury Studies, E.F and W.L. McKnight Brain Institute of the University of Florida, Gainesville, Florida. Departments of 2 Neuroscience, 3 Pediatrics, 4 Neurosurgery, 5 Anatomy and Cell Biology, and 6 Psychiatry, University of Florida, Gainesville, Florida. INTRODUCTION T RAUMATIC BRAIN INJURY (TBI) is a major health care issue that can lead to permanent motor, cognitive and behavioral deficits. These deficits are the result of neural tissue injury and cell death, most of which occurs within the first days after injury (Raghupathi et al., 2000). The ability of this tissue to resist injury and recover depends
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Cell-Specific Upregulation of Survivin after Experimental Traumatic Brain Injury in Rats

JOURNAL OF NEUROTRAUMAVolume 21 Number 9 2004copy Mary Ann Liebert IncPp 1183ndash1195
Cell-Specific Upregulation of Survivin after ExperimentalTraumatic Brain Injury in Rats
ERIK A JOHNSON12 STANISLAV I SVETLOV12 BRIAN R PIKE12
PAUL J TOLENTINO14 GERALD SHAW125 KEVIN KW WANG126
RONALD L HAYES1246 and JOSE A PINEDA13
ABSTRACT
In this study we examined the expression and cellular localization of survivin and proliferating cellnuclear antigen (PCNA) after controlled cortical impact traumatic brain injury (TBI) in rats Therewas a remarkable and sustained induction of survivin mRNA and protein in the ipsilateral cortexand hippocampus of rats after TBI peaking at five days post injury In contrast both survivinmRNA and protein were virtually undetectable in craniotomy control animals Concomitantly ex-pression of PCNA was also significantly enhanced in the ipsilateral cortex and hippocampus of theserats with similar temporal and spatial patterns Immunohistochemistry revealed that survivin andPCNA were co-expressed in the same cells and had a focal distribution within the injured brainFurther analysis revealed a frequent co-localization of survivin and GFAP an astrocytic markerin both the ipsilateral cortex and hippocampus while a much smaller subset of cells showed co-lo-calization of survivin and NeuN a mature neuronal marker Neuronal localization of survivin wasobserved predominantly in the ipsilateral cortex and contralateral hippocampus after TBI PCNAprotein expression was detected in both astrocytes and neurons of the ipsilateral cortex and hip-pocampus after TBI Collectively these data demonstrate that the anti-apoptotic protein survivinpreviously characterized in cancer cells is abundantly expressed in brain tissues of adult rats sub-jected to TBI We found survivin expression in both astrocytes and a sub-set of neurons In addi-tion the expression of survivin was co-incident with PCNA a cell cycle protein This suggests thatsurvivin may be involved in regulation of neural cell proliferative responses after traumatic braininjury
Key words astrocyte neuron PCNA survivin traumatic brain injury
1183
1Center for Traumatic Brain Injury Studies EF and WL McKnight Brain Institute of the University of Florida GainesvilleFlorida
Departments of 2Neuroscience 3Pediatrics 4Neurosurgery 5Anatomy and Cell Biology and 6Psychiatry University of FloridaGainesville Florida
INTRODUCTION
TRAUMATIC BRAIN INJURY (TBI) is a major health careissue that can lead to permanent motor cognitive and
behavioral deficits These deficits are the result of neuraltissue injury and cell death most of which occurs withinthe first days after injury (Raghupathi et al 2000) Theability of this tissue to resist injury and recover depends
largely on two factors the survival potential of the cellsand the proliferative ability of the cells in the affectedarea Therefore proliferation of cells in response to in-jury is important in the compensatoryreparative processAstrocytes multiply possibly to support surviving neu-rons and prevent further tissue damage through forma-tion of the glial scar (Ridet et al 1997 Bush et al 1999Smith et al 2001) Microglia increase to remove cellu-lar debris and promote recovery (Giulian 1991) Neu-rons may be replenished by neural stem cells in the den-tate gyrus and subventricular zones (Doetsch et al 1999Cameron and McKay 2001 Kernie et al 2001 Yagitaet al 2001 Peterson 2002) Consistent with these find-ings cell cycle protein expression has been shown afterTBI (Kaya et al 1999a) However studies have not in-vestigated the role of survivin a pro-mitotic and anti-apoptotic protein in the adult brain after TBI
Survivin is a novel member of the inhibitor of apopto-sis protein (IAP) family that can inhibit activated caspases(Ambrosini et al 1997 LaCasse et al 1998 Takahashiet al 1998 Tamm et al 1998 Deveraux and Reed 1999Li and Altieri 1999 Muchmore et al 2000 Jiang et al2001) Survivin is also an evolutionarily conserved chro-mosomal passenger protein that is required for propercompletion of mitosis Survivin is present during normaltissue development (Adida et al 1998 Kobayashi et al1999) but is absent in most adult tissues including thebrain (Ambrosini et al 1997 Kobayashi et al 1999)Many cancer cell lines and cancer tumors such as neuralderived neuroblastoma and glioblastoma which prolifer-ate at high rates exhibit survivin over-expression (Altieriet al 1999 Sasaki et al 2002) In addition blocking sur-vivin expression with anti-sense oligonucleotides in thesecell lines leads to cell death (Shankar et al 2001)
In this paper we demonstrate the induction of survivinexpression at the levels of mRNA and protein in the cor-tex and hippocampus of rats after traumatic brain injurySurvivin protein was primarily localized to astrocytes andin a small subset of neurons as indicated by its co-local-ization with GFAP and NeuN In addition a remarkable in-duction of proliferating cell nuclear antigen (PCNA) wasobserved after TBI and also localized to astrocytes and neu-rons Finally survivin and PCNA were co-expressed in sin-gle cells suggesting a possible role for survivin in regula-tion of cellular proliferative responses following TBI
MATERIALS AND METHODS
Induction of Controlled Cortical Impact Brain Injury
The surgical and cortical impact injury procedureswere conducted as previously described (Dixon et al1991 Pike et al 1998) Briefly adult male Sprague-
Dawley rats (250ndash300 g) were anesthetized with 4isoflurane (Halocarbon Laboratories River Edge NJ) in11 O2N2O for 4 min and maintained during surgery with25 isoflurane Core body temperature was continu-ously monitored using a rectal thermistor probe and main-tained at 365ndash375degC using an adjustable heating pad Aunilateral craniotomy (ipsilateral to injury) was performedover the right cortex between the sagittal suture bregmaand lambda while leaving the dura intact Traumatic in-sult was generated by impacting the exposed cortex witha 5-mm-diameter aluminum tip at a velocity of 4 mseca 150-msec dwell time and a 16-mm compression Cran-iotomy control animals received the cra-niotomy but notthe impact injury All procedures were performed ac-cording to guidelines established by the University ofFlorida Institutional Animal Care and Use Committee(IACUC) and the National Institutes of Health (NIH)
Quantitative Real-Time Polymerase ChainReaction (Q-PCR)
Survivin primers were generated using GeneBank lo-cus AF 276775 forward primer 5 TAAGC CACTTGTCCC AGCTT 3 and reverse primer 5 AGGATGGTAC CCCAT TACCT 3 GAPDH forward primer5 GGCTG CCTTC TCTTG TGAC 3 and the reverseprimer 5 CACCA CTTCG TCCGC CGG 3 Corticaland hippocampal tissues from the ipsilateral and con-tralateral hemispheres were rapidly excised at either 1 23 5 7 or 14 days and ldquosnap-frozenrdquo with liquid nitro-
JOHNSON ET AL
1184
FIG 1 Survivin mRNA induction in rat brain after traumaticbrain injury Rats were subjected to craniotomy followed bycontrolled cortical impact brain injury Total RNA was isolatedfrom injured (ipsilateral) cortex (ic) and hippocampus (ih) atindicated post-injury times cDNA was synthesized and quan-titative PCR using survivin primers was performed Data aregiven as percent of survivin expression over craniotomy con-trols (C Cont) each time point represents mean SEM offour independent measurements in craniotomy control or TBIgroup p 001 versus craniotomy control (one-wayANOVA test with post hoc Bonferroni analysis)
1185
FIG 2 Expression of survivin protein after TBI in rats Brain tissue homogenate proteins (40 g) were separated using SDS-PAGE immunoblotted with survivin antibody and visualized (A) Representative Western blot of survivin (17-kDa protein) inipsilateral cortex (ic) and hippocampus (ih) contralateral cortex (cc) and hippocampus (ch) obtained from injured rats and fromcraniotomy control rats without cortical impact (C Cont) Densitometry analysis representation of survivin-positive bands in ip-silateral (ic) and contralateral (cc) cortex (B) and ipsilateral (ih) and contralateral (ch) hippocampus (C) after TBI is shown aspercent of craniotomy control values Each data point represents the mean SEM of four to six independent experiments p 005 p 0001 versus craniotomy control (one-way ANOVA test with post hoc Bonferroni analysis)
SURVIVIN UPREGULATION AFTER TBI
gen Total RNA was isolated from the samples using TRI-zol reagent (Invitrogen Carlsbad CA) according to themanufacturerrsquos instructions cDNA synthesis was per-formed using 1 g of total RNA with the SuperScripttradeFirst-Strand Synthesis System for RT-PCR kit (Invitro-genLife Technologies Carlsbad CA) according to the
manufacturerrsquos instructions Q-PCR was performed aspreviously described (Tolentino et al 2002) using theLightCycler-FastStart DNA Master SYBR Green I reac-tion mix (Roche Diagnostics Indianapolis IN) in com-bination with 05 M primers 25 mM MgCl2 in theLight Cycler rapid thermal cycler system (Roche Diag-
JOHNSON ET AL
1186
FIG 3 Expression of PCNA after TBI in rats PVDF membranes visualized for survivin were stripped and re-probed withPCNA antibody Representative western blots showing PCNA (36 kDa) (A) and densitometry analysis of PCNA-positive bands(BC) are presented Experimental conditions sample size and abbreviations are identical to those in Figure 2 p 005 p 001 versus craniotomy control (one-way ANOVA test with post hoc Bonferroni analysis)
SURVIVIN UPREGULATION AFTER TBI
1187
FIG 4 Immunohistochemistry of survivin and PCNA Double fluorescent immunostaining for survivin (red) and PCNA (green)was performed in the ipsilateral cortex (A) and hippocampus (B) at 5 day post-injury Survivin is expressed in the cytoplasm (Cred) while PCNA is expressed in the nucleus (D green) The white arrow indicates the typical focal co-expression of survivinand PCNA as shown in merged survivin and PCNA images (E) PCNA expression was co-incident with DAPI staining (F bluewhite arrow) Original magnification 200 bar 50 m (AB) original magnification 400 bar 20 m (CndashF)
nostics Indianapolis IN) Briefly the primers were ampli-fied then quantified by online monitoring and identificationof the crossing point value (CPV) which is the exact timepoint at which the logarithmic linear phase could be dis-tinguished from the background The survivin primer setswere subjected to serial dilution and linear regression analy-sis of the logarithm of the dilution factor versus the CPVgenerated a standard curve for each transcript-speci-fic tem-plate Results are presented as percentage of cranio-tomycontrol Data were analyzed by ANOVA with a post-hocBonferroni-test and are given as mean SEM Differenceswere considered significant at the level of p 005
Western Blot Analyses
Brain tissue was removed as described above rinsedwith cold PBS snap frozen in liquid nitrogen and ho-
mogenized in ice-cold triple detergent lysis buffer con-taining a Completetrade protease inhibitor cocktail (RocheBiochemicals Indianapolis IN) Protein concentrationwas determined by bicinchoninic acid (BCA) micro pro-tein assays (Pierce Inc Rockford IL) Forty microgramsof protein per well was loaded and separated by SDS-PAGE transferred to PVDF membranes and probed withgoat-anti-rabbit survivin antibody (Novus BiologicalsLittleton CO 11000) After incubation with goat anti-rabbit HRP-labeled secondary antibody (Biorad Her-cules CA) the membranes were developed using En-hanced Chemiluminescence Plus reagents (ECL PlusAmersham Arlington Heights IL) For further PCNAanalysis developed PVDF membranes were incubated instripping buffer rinsed twice in TBST and incubated withPCNA antibody (Santa Cruz Biotech Santa Cruz CA
11000) as described above Semi-quantitative densito-metric analysis was performed using the AlphaImagertrade2000 Digital Imaging System (San Leandro CA) Trans-formed data (experimental densitometry value crani-otomy control densitometry value 100) was evaluatedby ANOVA and a post-hoc Bonferroni-test Values areexpressed as percentage of craniotomy controls and aregiven as mean SEM Differences were considered sig-nificant at the level of p 005
Characterization of Survivin Antibody (R51)
First we compared the specificity of the survivin an-tibody developed within our group (R51 Dr G Shaw)and a commercially available survivin antibody (Chemi-con Temecula CA) Our antibody showed characteris-tic staining of the cleavage furrow between dividingHeLa cells consistent with other reports (Li et al 19981999 Uren et al 2000) In addition double labeling withboth survivin antibodies showed co-localization at thecleavage furrow The peptides used to develop our sur-vivin antibody are specific to survivin and do not recog-nize other IAP family proteins according to SDSC Biol-ogy Workbench BLASTP (222) (Altschul et al 1997)and CLUSTAL W (181) analysis (Higgins et al 1992Thompson et al 1994) resulting in the survivin anti-bodyrsquos specificity
Immunohistochemistry (IHC)
Animals were transcardially perfused with 2 Heparin(Elkins-Sinn Inc Cherry Hill NJ) in 09 saline solu-tion (pH 74) followed by 4 paraformaldehyde in 01M phosphate buffer (pH 74) The brains were post-fixedin 4 paraformaldehyde and stored in 01M PBS or cryo-buffer Forty micron sections were fluorescent immuno-labeled with two primary antibodies in the following experiments survivin (1500)GFAP for astrocytes(Sternberger Lutherville MD 11000) survivinNeuNfor mature neurons (Chemicon Temecula CA 11000)survivinPCNA (Santa Cruz Biotech Santa Cruz CA1200) PCNAGFAP and PCNANeuN The nuclear dyeDAPI (in Vectashield H-1200 Vector LaboratoriesBurlingame CA) was used to label the nuclei The firstprimary antibody was incubated at 4degC for 24ndash48 h in a2 goat serum2 horse serum02 Triton-X 100 in01 M PBS (block) solution followed by the second pri-mary antibody at 4degC for 1 h in block solution Fluores-cent-tagged secondary antibody (Molecular Probes Eu-gene OR) was used for visualization For double-labelingusing same species antibodies we used the tyramide sig-nal amplification (TSA) kit (PerkinElmer Life SciencesBoston MA) according to the manufacturerrsquos instruc-tions and as previously described (Stone et al 2002) The
sections were viewed and digitally captured with a ZeissAxioplan 2 microscope (Zeiss Thornwood NY)equipped with a SPOT Real Time Slider high-resolutioncolor CCD digital camera (Diagnostic Instruments IncSterling Heights MI) The number of animals used fordual-labeling IHC is as follows survivin PCNA 4survivin GFAP 6 survivin NeuN 4 PCNA GFAP 4 and PCNA NeuN 4 Cell counts wereobtained by comparing the number of double-labeledcells to total single-labeled cells in the following groupssurvivinNeuN positive cells to total NeuN positive cellssurvivinPCNA positive cells to total PCNA positivecells PCNANeuN positive cells to total NeuN positivecells and survivinGFAP positive cells to total GFAPpositive cells Percentages were calculated by dividingthe number of double-labeled cells with the total numberof single-labeled cells For each group representativephotomicrographs were selected and counted Cells werecounted in a total area of at least 188000 m2 for eachgroup with no distinction made between cortical and hip-pocampal regions
RESULTS
Induction of Survivin Expression after TBI
Q-PCR analysis revealed an initial increase in survivinmRNA at 2 days post injury in the ipsilateral cortex andhippocampus These transcripts remained elevated inboth regions reached maximum levels at day 5 post-in-jury and declined at 7 days in the cortex and at 14 daysin the hippocampus All experimental animals remainedalive and exhibited slightly impaired motor and cogni-tive impairments (data not shown) Cortical mRNA lev-els reached a maximum of 448 100 whereas hip-pocampal mRNAs attained 606 100 compared tocraniotomy control values (Fig 1) To determine if theinduction of survivin mRNA resulted in correspondingincreases in survivin protein western blot analysis wasperformed Survivin (17-kDa protein) was readily de-tectable in the ipsilateral cortex and hippocampus of TBIrats while it was negligible in contralateral cortex andhippocampus (Fig 2A) Survivin was expressed in atime-dependent manner with a maximum increase at 5days after injury followed by a gradual decline by 14days Specifically the levels of survivin in cortical tis-sue were 616 257 at 3 days and 839 339 at 5days compared to craniotomy controls (Fig 2B) Similarincreases of survivin protein in the ipsilateral hippocam-pus were detected at 3 d and 5 days post injury 464 196 and 545 102 compared to craniotomy controlrespectively (Fig 2C)
JOHNSON ET AL
1188
FIG 5 Co-localization of survivin and GFAP in brain tissue after TBI Fluorescent immunohistochemistry for survivin (green)and GFAP (red) was performed in the ipsilateral and contralateral cortex (AB) and in the CA1 and dentate gyrus regions of thehippocampus (GH) at 5 day post-injury The injury has completely destroyed the cortex in G leaving only the hippocampus inthis picture Survivin was expressed in the cytoplasm (DJ green) of GFAP-positive astrocytes (CI red) of the ipsilateral cor-tex and hippocampus and was found to co-localize to these cells as shown in merged CD and IJ images (EK respectively yel-low) White arrows indicate typical survivin-positive astrocytes Nuclei are shown using DAPI (FL blue) Original magnifica-tion 100 bar 50 m (ABGH) 400 bar 20 m (CndashF and IndashL)
FIG 6 Co-localization of PCNA and GFAP in brain tissue after TBI Double immunostaining for PCNA (green) and GFAP(red) was performed in the ipsilateral and contralateral cortex (AB) and the CA1 and dentate gyrus regions of the hippocampus(GH) at 5 day post-injury PCNA is present in GFAP-positive cells of ipsilateral cortex (CD) and to a lesser extent hippocampus(IJ) (EK) Merged CD and IJ respectively White arrows indicate typical PCNA-positive astrocytes PCNA expression wasco-incident with DAPI staining (FL blue) Original magnification 100 bar 50 m (ABGH) 400 bar 20 m (CndashFand IndashL)
PCNA Expression after TBI
For detection of proliferating cell nuclear antigen(PCNA) PVDF membranes immunostained for survivinwere stripped and re-probed using a PCNA-specific an-tibody PCNA (36-kDa protein) was significantly de-tectable in the ipsilateral cortex and hippocampus of TBIrats but only negligible amounts were observed in thecontralateral cortex and hippocampus (Fig 3A) Thetemporal patterns exhibited by PCNA protein were sim-ilar to that of survivin protein Namely PCNA expressedin a time-dependant fashion with a maximum increase at5 days after injury followed by a gradual decline by 14days The levels of PCNA in ipsilateral cortical tissuewere raised over craniotomy control by 919 459 at3 days 2263 333 at 5 days and 1035 356 at 7days post injury (Fig 3B) Similar increases of PCNAprotein in ipsilateral hippocampus were detected at 5 dayspost injury with a maximum of 1006 229 comparedto craniotomy controls (Fig 3C) No significant increasewas found in the contralateral regions when compared tocraniotomy controls (Fig 3A)
Co-Expression of Survivin and PCNA following TBI
To examine spatial co-localization of survivin andPCNA double-label immunohistochemistry of brain tis-sue sections was performed on day 5 post injury whenpeak expression of these proteins was observed
At this time point a remarkable survivin and PCNAimmunoreactivity was found in the ipsilateral cortex (Fig4A) and ipsilateral hippocampus (Fig 4B) consistent withdata obtained using Western blot analyses Within bothregions focal co-expression patterns of survivin andPCNA in single cells were detected which was demon-strated by both separate fluorescent visualization of in-dividual proteins and by merging the images of double-stained slides (Fig 4CndashE) However the dual expressionof survivin and PCNA occurred infrequently as survivinand PCNA immunoreactivity could readily be found sep-arately (Fig 4CndashE) Approximately 12 of the total num-ber of PCNA-positive cells also labeled with survivin Thenuclear morphology of dual survivin and PCNA-positivecells was ambiguous as indicated by DAPI staining (Fig4F) Therefore DAPI staining was simply used for cellidentification in all subsequent experiments
Survivin and PCNA Are Expressed in Astrocytes after TBI
To determine the cell types expressing survivin andPCNA double-label immunohistochemistry for theseproteins and GFAP a marker of astrocytes was per-formed on day 5 post injury
In accordance with Western blot data remarkable sur-vivin-positive immunoreactivity was observed in the ip-silateral cortex and hippocampus proximal to the injurycavity (Fig 5AG green) but not in the contralateral ar-eas (Fig 5BH) Survivin was co-localized with GFAPin the cells of injured cortex and hippocampus whichstrongly suggested primary accumulation of survivin incells of astrocytic lineage (Fig 5CndashEIndashL) Survivin wasuniformly distributed in the cytoplasm and processes ofastrocytes in both cortex and hippocampus (Fig 5DJ)DAPI staining is shown in Figure 5FL3 Approximately88 of the total number of GFAP-positive cells also la-beled with survivin
PCNA-positive immunoreactivity staining was ob-served in the ipsilateral cortex (Fig 6A green) and hip-pocampus (Fig 6G green) of injured brain while con-tralateral cortex and hippocampus exhibited negligiblePCNA immunoreactivity (Fig 6BH) PCNA (Fig 6CI)was partially co-localized with GFAP (Fig 6DJ red) inboth regions and was characteristically distributed in thenucleus of the cells in both cortex and hippocampus (Fig6EK) DAPI staining is shown in Figure 6FL
Taken together double-label immunohistochemistrydata provides evidence that both survivin and PCNA canbe detected in GFAP-positive astrocytes following trau-matic insult Since survivin and PCNA immunoreactiv-ity was not exclusively localized in GFAP-positive cellswe next addressed a question what other cell type mightexpress survivin after TBI We suggested that a certainpopulation of mature neurons might express survivin inresponse to injury
Survivin and PCNA Are Expressed in a Sub-Set of Neurons after TBI
As can be seen in Figure 7 survivin and PCNA wereeach co-expressed with NeuN a marker of mature neu-rons NeuN-positive cells were found to express survivinin the ipsilateral cortex distal to the injury cavity (Fig7AndashD) and in the contralateral hippocampus (Fig 7EndashH)It should be noted however that NeuN-positive cells thatalso expressed survivin occurred infrequently For ex-ample we estimated the number of dual survivinNeuNpositive cells as 01ndash15 of the total number of NeuN-positive cells in these regions Survivin immunoreactiv-ity was negligible in either hemisphere of craniotomycontrol brains (Fig 7QR) No co-localization of survivinand NeuN was observed in ipsilateral hippocampus (datanot shown) As can be seen in Figure 7BF survivin waspredominantly localized to the cytoplasm and axons ofNeuN-positive neurons DAPI staining is shown in Fig-ure 7DH
PCNA-positive neurons were found in the ipsilateralcortex (Fig 7IndashL) and hippocampus after TBI (Fig
JOHNSON ET AL
1190
1191
FIG 7 A sub-set of NeuN-positive neurons express survivin and PCNA after TBI Double fluorescent immunohistochemistryfor survivin (green) and NeuN (red) in the ipsilateral cortex (AB) and the CA1 pyramidal layer of the contralateral hippocam-pus (EF) was performed Survivin is expressed in the cytoplasm and to a limited extent in the processes of NeuN-positive neu-rons (merged images CG) Dual staining for PCNA (green) and NeuN (red) is shown in the ipsilateral cortex (IJ) and the CA1pyramidal layer of the ipsilateral hippocampus (MN) The nuclei are shown using DAPI staining (DH blue) PCNA is expressedin the nucleus of NeuN-positive neurons (merged images KO) PCNA expression was co-incident with DAPI staining in theseexamples (LP blue) White arrows indicate focal co-localization of survivinNeuN and PCNANeuN SurvivinNeuN co-local-ization of survivin (green) and NeuN (red) was seen only in TBI rats as opposed to either hemisphere of craniotomy control(QR) Original magnification 400 bar 20 m (AndashP) original magnification 50 bar 100000 m (QR)
SURVIVIN UPREGULATION AFTER TBI
7MndashP) whereas craniotomy control tissue exhibited onlytrace amounts of PCNA (data not shown) Similar to thesurvivinNeuN co-localization data dual PCNANeuNimmunostaining was a rare event accounting for approx-imately 4 of the total number of NeuN positive cellsPCNA was distributed in the nuclei of these neurons (Fig7KO) although the nuclear morphology of these cellswas not clearly resolved by DAPI staining (Fig 7LP)
DISCUSSION
Traumatic brain injury (TBI) sets into motion variousbiochemical cascades that induce neural tissue injury andcell death Conversely several proteins expressed inneural cells after TBI are directed to resist cell death andpromote recovery in the injured CNS (Ridet et al 1997Chen and Swanson 2003) Survivin is a multi-functionalprotein that inhibits apoptosis and is also required for theproper completion of mitosis Anti-apoptotic and pro-mi-togenic roles for survivin have been documented in pro-liferating cells of neural origin in vitro such as in neu-roblastoma and glioma cells (LaCasse et al 1998 Tammet al 1998 Deveraux and Reed 1999 Conway et al2000 Shin et al 2001 Sasaki et al 2002) Howeverno studies have investigated the potential role of survivinin the adult brain after TBI when a sub-population ofCNS cells may initiate a cell cycle-related process in re-sponse to injury
In the present paper we demonstrate the induction ofsurvivin expression in rat brain subjected to TBI The ex-pression of survivin was time-dependent and cell-spe-cific and was present in astrocytes and to a much lesserextent in neurons in brain cortex and hippocampus In-duction of survivin in these cells was accompanied byoccasional expression of PCNA a cell cycle protein in-volved in mitotic G1S progression Our present data arethe first to show that survivin mRNA and protein are sig-nificantly up-regulated after traumatic brain injury in ratsPCNA expression after TBI has been described previously(Miyake et al 1992 Chen et al 2003) suggesting itsrole in mechanisms of brain recovery after injury Theconcurrent up-regulation of survivin with a similar tem-poral profile as PCNA shown herein further suggests thatsurvivin may play a role in cellular proliferation after TBI
Brain injury evoked the expression of survivin andPCNA in a time-dependent manner (Figs 2 and 3) West-ern blot analysis revealed maximal co-expression of bothsurvivin and PCNA at five days post injury Immuno-histochemistry at this time point demonstrated co-local-ization of these proteins (Fig 4) although most cells werelabeled separately with PCNA and survivin In fact only12 of the total number of PCNA-positive cells were
also survivin positive It has been reported that PCNA isexpressed predominantly in G1S (Bravo et al 1987)while survivin is found at the G2M phase of the cell cy-cle (Bravo et al 1987 Otaki et al 2000) Hence a lackof strict co-localization of survivin and PCNA in ourstudy may be explained by their expression at differentpoints in the cell cycle
In our experimental model we observed survivin- aswell as PCNA-positive astrocytes in the proximal area ofthe injury and in the ipsilateral hippocampus Prolifera-tion of astrocytes is well documented after TBI as shownby cell labeling with BrdU as well as expression of PCNA(Latov et al 1979 Dunn-Meynell and Levin 1997 Car-bonell and Grady 1999 Norton 1999 Csuka et al 2000Kernie et al 2001 Chen et al 2003) Because survivinand PCNA were expressed in astrocytes following TBI(Figs 5 and 6) it is possible that survivin plays an im-portant role linking astrocyte survival and proliferationafter traumatic insult Astrocyte proliferation has beenimplicated in the formation of the glial scar observed af-ter injury (Latov et al 1979) and creates a non-permis-sive environment for repair (Sykova et al 1999) How-ever glial proliferation may also enhance neuronalsurvival (Smith et al 2001 Wei et al 2001)
Of particular interest is a sub-set of NeuN-positive neu-rons found to express survivin only after TBI (Fig 7)These cells were much less abundant than survivin-pos-itive astrocytes and their functional significance is cur-rently unknown However both neurons and astrocyteshave been documented previously to express cell cycleproteins after various insults such as exposure to -amy-loidndashactivated microglia (Wu et al 2000) TBI (Kaya etal 1999ab) chlorine toxicity (Magavi et al 2000) oras a consequence of Alzheimerrsquos disease (Yang et al2001) These papers underscore the significant contro-versy that exists regarding the function of cell cycle pro-teins such as PCNA in neurons after different types ofinjury We are currently conducting further studies whichwill elucidate the roles for PCNA and survivin in neu-rons after TBI
It should be noted that dual staining of survivin andPCNA could not be directly attributed to a specific celltype due to the technical difficulties of triple labeling an-tibody-based IHC Therefore we cannot rule out the pos-sibility that other cell types such as endothelial (Conwayet al 2003) or inflammatory cells (Hill-Felberg et al1999) may also contribute to survivin and PCNA ex-pression after TBI The appearance of survivin and PCNAseparately in neurons (NeuN-positive) and astrocytes(GFAP-positive) along with co-localization of survivinwith PCNA in the same cells provide correlative data tosuggest an activation of cell cycle-like program in astro-cytes and possibly in a small subtype of neurons after
JOHNSON ET AL
1192
TBI In our experiments survivin co-localization withPCNA does suggest that survivin may be associated witha pro-mitotic process In an attempt to clarify these pro-teinrsquos roles after TBI we analyzed the nuclear morphol-ogy of survivin-positive cells to define the apoptotic ormitotic architecture of nuclei DAPI staining proved tooambiguous in identifying apoptotic versus mitotic phe-notypes likely due to the thickness of the brain sections(40 m) Further studies using direct markers of mitosissuch as BrdU incorporation as well as simultaneous la-beling with cell death related proteins is required to de-lineate anti-apoptotic and pro-mitotic activities of sur-vivin and PCNA in these cells
In conclusion our data demonstrate the induction ofsurvivin in the rat brain following TBI Expression of sur-vivin occurred predominantly in astrocytes as comparedto neurons in a time-dependant fashion and was accom-panied by expression of PCNA Taken together these re-sults suggest that survivin plays a role in neural cell re-sponses following traumatic brain injury in rats Futurestudies will investigate the implications of these findingsto the pathophysiology of TBI
ACKNOWLEDGMENTS
We thank Barbara OrsquoSteen and Tao Fan MD fortechnical assistance This work was funded by grants NIHRO1 NS39091 NIH RO1 NS40182 DAMD 17-99-1-9565 and DAMD 17-01-1-0765
REFERENCES
ADIDA C CROTTY PL MCGRATH J BERREBI DDIEBOLD J and ALTIERI DC (1998) Developmentallyregulated expression of the novel cancer anti-apoptosis genesurvivin in human and mouse differentiation Am J Pathol152 43ndash49
ALTIERI DC MARCHISIO PC and MARCHISIO C(1999) Survivin apoptosis an interloper between cell deathand cell proliferation in cancer Lab Invest 79 1327ndash1333
ALTSCHUL SF MADDEN TL SCHAFFER AA et al(1997) Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 253389ndash3402
AMBROSINI G ADIDA C and ALTIERI DC (1997) Anovel anti-apoptosis gene survivin expressed in cancer andlymphoma Nat Med 3 917ndash921
BRAVO R FRANK R BLUNDELL PA and MAC-DONALD-BRAVO H (1987) CyclinPCNA is the auxil-iary protein of DNA polymerase-delta Nature 326 515ndash517
BUSH TG PUVANACHANDRA N HORNER CH et al
(1999) Leukocyte infiltration neuronal degeneration andneurite outgrowth after ablation of scar-forming reactive as-trocytes in adult transgenic mice Neuron 23 297ndash308
CAMERON HA and MCKAY RD (2001) Adult neuroge-nesis produces a large pool of new granule cells in the den-tate gyrus J Comp Neurol 435 406ndash417
CARBONELL WS and GRADY MS (1999) Regional andtemporal characterization of neuronal glial and axonal re-sponse after traumatic brain injury in the mouse Acta Neu-ropathol (Berl) 98 396ndash406
CHEN XH IWATA A NONAKA M BROWNE KDand SMITH DH (2003) Neurogenesis and glial prolifera-tion persist for at least one year in the subventricular zonefollowing brain trauma in rats J Neurotrauma 20 623ndash631
CHEN Y and SWANSON RA (2003) Astrocytes and braininjury J Cereb Blood Flow Metab 23 137ndash149
CONWAY EM POLLEFEYT S CORNELISSEN J et al(2000) Three differentially expressed survivin cDNA vari-ants encode proteins with distinct antiapoptotic functionsBlood 95 1435ndash1442
CSUKA E HANS VH AMMANN E TRENTZ OKOSSMANN T and MORGANTI-KOSSMANN MC(2000) Cell activation and inflammatory response followingtraumatic axonal injury in the rat Neuroreport 112587ndash2590
DEVERAUX QL and REED JC (1999) IAP family pro-teinsmdashsuppressors of apoptosis Genes Dev 13 239ndash252
DIXON CE CLIFTON GL LIGHTHALL JW YAGH-MAI AA and HAYES RL (1991) A controlled corticalimpact model of traumatic brain injury in the rat J NeurosciMethods 39 253ndash262
DOETSCH F CAILLE I LIM DA GARCIA-VERDUGOJM and ALVAREZ-BUYLLA A (1999) Subventricularzone astrocytes are neural stem cells in the adult mammalianbrain Cell 97 703ndash716
DUNN-MEYNELL AA and LEVIN BE (1997) Histolog-ical markers of neuronal axonal and astrocytic changes af-ter lateral rigid impact traumatic brain injury Brain Res 76125ndash41
GIULIAN D (1991) Microgliamdashneuron interactions after in-jury to the central nervous system in Peripheral Signalingof the Brain Role in Neural-Immune Interactions Learningand Memory R Frederickson J McGaugh and D Felton(eds) Lewiston NY Hogrefe and Huber pps 73ndash82
HIGGINS DG BLEASBY AJ and FUCHS R (1992)CLUSTAL V improved software for multiple sequencealignment Comput Appl Biosci 8 189ndash191
HILL-FELBERG SJ MCINTOSH TK OLIVER DLRAGHUPATHI R and BARBARESE E (1999) Concur-rent loss and proliferation of astrocytes following lateral fluidpercussion brain injury in the adult rat J Neurosci Res 57271ndash279
1193
SURVIVIN UPREGULATION AFTER TBI
JIANG X WILFORD C DUENSING S MUNGER KJONES G and JONES D (2001) Participation of Survivinin mitotic and apoptotic activities of normal and tumor-de-rived cells J Cell Biochem 83 342ndash354
KAYA SS MAHMOOD A LI Y YAVUZ E andCHOPP M (1999a) Expression of cell cycle proteins (cy-clin D1 and cdk4) after controlled cortical impact in rat brainJ Neurotrauma 16 1187ndash1196
KAYA SS MAHMOOD A LI Y YAVUZ E GOKSELM and CHOPP M (1999b) Apoptosis and expression ofp53 response proteins and cyclin D1 after cortical impact inrat brain Brain Res 818 23ndash33
KERNIE SG ERWIN TM and PARADA LF (2001)Brain remodeling due to neuronal and astrocytic prolifera-tion after controlled cortical injury in mice J Neurosci Res66 317ndash326
KOBAYASHI K HATANO M OTAKI M OGA-SAWARA T and TOKUHISA T (1999) Expression of amurine homologue of the inhibitor of apoptosis protein is re-lated to cell proliferation Proc Natl Acad Sci USA 961457ndash1462
LACASSE EC BAIRD S KORNELUK RG andMACKENZIE AE (1998) The inhibitors of apoptosis(IAPs) and their emerging role in cancer Oncogene 173247ndash3259
LATOV N NILAVER G ZIMMERMAN EA et al(1979) Fibrillary astrocytes proliferate in response to braininjury a study combining immunoperoxidase technique forglial fibrillary acidic protein and radioautography of tritiatedthymidine Dev Biol 72 381ndash384
LI F and ALTIERI DC (1999) The cancer antiapoptosismouse survivin gene characterization of locus and tran-scriptional requirements of basal and cell cycle-dependentexpression Cancer Res 59 3143ndash3151
LI F AMBROSINI G CHU EY et al (1998) Control ofapoptosis and mitotic spindle checkpoint by survivin Nature396 580ndash584
LI F ACKERMANN EJ BENNETT CF et al (1999)Pleiotropic cell-division defects and apoptosis induced by in-terference with survivin function Nat Cell Biol 1 461ndash466
MAGAVI SS LEAVITT BR and MACKLIS JD (2000)Induction of neurogenesis in the neocortex of adult mice Na-ture 405 951ndash955
MIYAKE T OKADA M and KITAMURA T (1992) Re-active proliferation of astrocytes studied by immunohisto-chemistry for proliferating cell nuclear antigen Brain Res590 300ndash302
MUCHMORE SW CHEN J JAKOB C et al (2000)Crystal structure and mutagenic analysis of the inhibitor-of-apoptosis protein survivin Mol Cell 6 173ndash182
NORTON WT (1999) Cell reactions following acute braininjury a review Neurochem Res 24 213ndash218
OTAKI M HATANO M KOBAYASHI K et al (2000)Cell cyclendashdependent regulation of TIAPm-survivin expres-sion Biochim Biophys Acta 1493 188ndash194
PETERSON DA (2002) Stem cells in brain plasticity and re-pair Curr Opin Pharmacol 2 34ndash42
PIKE BR ZHAO X NEWCOMB JK POSMANTURRM WANG KK and Hayes RL (1998) Regional cal-pain and caspase-3 proteolysis of alpha-spectrin after trau-matic brain injury Neuroreport 9 2437ndash2442
RAGHUPATHI R GRAHAM DI MCINTOSH TK(2000) Apoptosis after traumatic brain injury J Neuro-trauma 17 927ndash938
RIDET JL MALHOTRA SK PRIVAT A and GAGEFH (1997) Reactive astrocytes cellular and molecular cuesto biological function Trends Neurosci 20 570ndash577
SASAKI T LOPES MB HANKINS GR and HELMGA (2002) Expression of survivin an inhibitor of apopto-sis protein in tumors of the nervous system Acta Neu-ropathol (Berl) 104 105ndash109
SHANKAR SL MANI S OrsquoGUIN KN KANDIMALLAER AGRAWAL S and SHAFIT-ZAGARDO B (2001)Survivin inhibition induces human neural tumor cell deaththrough caspase-independent and -dependent pathways JNeurochem 79 426ndash436
SHIN S SUNG BJ CHO YS et al (2001) An anti-apop-totic protein human survivin is a direct inhibitor of caspase-3 and -7 Biochemistry 40 1117ndash1123
SMITH C BERRY M CLARKE WE and LOGAN A(2001) Differential expression of fibroblast growth factorndash2and fibroblast growth factor receptor 1 in a scarring and non-scarring model of CNS injury in the rat Eur J Neurosci 13443ndash456
STONE JR OKONKWO DO SINGLETON RHMUTLU LK HELM GA and POVLISHOCK JT(2002) Caspase-3ndashmediated cleavage of amyloid precursorprotein and formation of amyloid Beta peptide in traumaticaxonal injury J Neurotrauma 19 601ndash614
SYKOVA E VARGOVA L PROKOPOVA S and SI-MONOVA Z (1999) Glial swelling and astrogliosis pro-duce diffusion barriers in the rat spinal cord Glia 25 56ndash70
TAKAHASHI R DEVERAUX Q TAMM I et al (1998)A single BIR domain of XIAP sufficient for inhibiting cas-pases J Biol Chem 273 7787ndash7790
TAMM I WANG Y SAUSVILLE E et al (1998) IAP-family protein survivin inhibits caspase activity and apopto-sis induced by Fas (CD95) Bax caspases and anticancerdrugs Cancer Res 58 5315ndash5320
THOMPSON JD HIGGINS DG and GIBSON TJ (1994)CLUSTAL W improving the sensitivity of progressive mul-tiple sequence alignment through sequence weighting posi-tion-specific gap penalties and weight matrix choice NucleicAcids Res 22 4673ndash4680
JOHNSON ET AL
1194
TOLENTINO PJ DEFORD SM NOTTERPEK L et al(2002) Up-regulation of tissue-type transglutaminase aftertraumatic brain injury J Neurochem 80 579ndash588
UREN AG WONG L PAKUSCH M et al (2000) Sur-vivin and the inner centromere protein INCENP show simi-lar cell-cycle localization and gene knockout phenotypeCurr Biol 10 1319ndash1328
WEI LH HUANG CY CHENG SP CHEN CA andHSIEH CY (2001) Carcinosarcoma of ovary associatedwith previous radiotherapy Int J Gynecol Cancer 11 81ndash84
WU Q COMBS C CANNADY SB GELDMACHERDS and HERRUP K (2000) Beta-amyloid activated mi-croglia induce cell cycling and cell death in cultured corticalneurons Neurobiol Aging 21 797ndash806
YAGITA Y KITAGAWA K OHTSUKI T et al (2001)
Neurogenesis by progenitor cells in the ischemic adult rathippocampus Stroke 32 1890ndash1896
YANG Y GELDMACHER DS and HERRUP K (2001)DNA replication precedes neuronal cell death in Alzheimerrsquosdisease J Neurosci 21 2661ndash2668
Address reprint requests toJose A Pineda PhD
EF and WL McKnight Brain InstituteUniversity of Florida
100 S Newell Dr Bldg 59L1-118 Dept of Pediatrics
Gainesville FL 32610
E-mail pinedjapedsufledu
SURVIVIN UPREGULATION AFTER TBI
1195

largely on two factors the survival potential of the cellsand the proliferative ability of the cells in the affectedarea Therefore proliferation of cells in response to in-jury is important in the compensatoryreparative processAstrocytes multiply possibly to support surviving neu-rons and prevent further tissue damage through forma-tion of the glial scar (Ridet et al 1997 Bush et al 1999Smith et al 2001) Microglia increase to remove cellu-lar debris and promote recovery (Giulian 1991) Neu-rons may be replenished by neural stem cells in the den-tate gyrus and subventricular zones (Doetsch et al 1999Cameron and McKay 2001 Kernie et al 2001 Yagitaet al 2001 Peterson 2002) Consistent with these find-ings cell cycle protein expression has been shown afterTBI (Kaya et al 1999a) However studies have not in-vestigated the role of survivin a pro-mitotic and anti-apoptotic protein in the adult brain after TBI
Survivin is a novel member of the inhibitor of apopto-sis protein (IAP) family that can inhibit activated caspases(Ambrosini et al 1997 LaCasse et al 1998 Takahashiet al 1998 Tamm et al 1998 Deveraux and Reed 1999Li and Altieri 1999 Muchmore et al 2000 Jiang et al2001) Survivin is also an evolutionarily conserved chro-mosomal passenger protein that is required for propercompletion of mitosis Survivin is present during normaltissue development (Adida et al 1998 Kobayashi et al1999) but is absent in most adult tissues including thebrain (Ambrosini et al 1997 Kobayashi et al 1999)Many cancer cell lines and cancer tumors such as neuralderived neuroblastoma and glioblastoma which prolifer-ate at high rates exhibit survivin over-expression (Altieriet al 1999 Sasaki et al 2002) In addition blocking sur-vivin expression with anti-sense oligonucleotides in thesecell lines leads to cell death (Shankar et al 2001)
In this paper we demonstrate the induction of survivinexpression at the levels of mRNA and protein in the cor-tex and hippocampus of rats after traumatic brain injurySurvivin protein was primarily localized to astrocytes andin a small subset of neurons as indicated by its co-local-ization with GFAP and NeuN In addition a remarkable in-duction of proliferating cell nuclear antigen (PCNA) wasobserved after TBI and also localized to astrocytes and neu-rons Finally survivin and PCNA were co-expressed in sin-gle cells suggesting a possible role for survivin in regula-tion of cellular proliferative responses following TBI
MATERIALS AND METHODS
Induction of Controlled Cortical Impact Brain Injury
The surgical and cortical impact injury procedureswere conducted as previously described (Dixon et al1991 Pike et al 1998) Briefly adult male Sprague-
Dawley rats (250ndash300 g) were anesthetized with 4isoflurane (Halocarbon Laboratories River Edge NJ) in11 O2N2O for 4 min and maintained during surgery with25 isoflurane Core body temperature was continu-ously monitored using a rectal thermistor probe and main-tained at 365ndash375degC using an adjustable heating pad Aunilateral craniotomy (ipsilateral to injury) was performedover the right cortex between the sagittal suture bregmaand lambda while leaving the dura intact Traumatic in-sult was generated by impacting the exposed cortex witha 5-mm-diameter aluminum tip at a velocity of 4 mseca 150-msec dwell time and a 16-mm compression Cran-iotomy control animals received the cra-niotomy but notthe impact injury All procedures were performed ac-cording to guidelines established by the University ofFlorida Institutional Animal Care and Use Committee(IACUC) and the National Institutes of Health (NIH)
Quantitative Real-Time Polymerase ChainReaction (Q-PCR)
Survivin primers were generated using GeneBank lo-cus AF 276775 forward primer 5 TAAGC CACTTGTCCC AGCTT 3 and reverse primer 5 AGGATGGTAC CCCAT TACCT 3 GAPDH forward primer5 GGCTG CCTTC TCTTG TGAC 3 and the reverseprimer 5 CACCA CTTCG TCCGC CGG 3 Corticaland hippocampal tissues from the ipsilateral and con-tralateral hemispheres were rapidly excised at either 1 23 5 7 or 14 days and ldquosnap-frozenrdquo with liquid nitro-
JOHNSON ET AL
1184
FIG 1 Survivin mRNA induction in rat brain after traumaticbrain injury Rats were subjected to craniotomy followed bycontrolled cortical impact brain injury Total RNA was isolatedfrom injured (ipsilateral) cortex (ic) and hippocampus (ih) atindicated post-injury times cDNA was synthesized and quan-titative PCR using survivin primers was performed Data aregiven as percent of survivin expression over craniotomy con-trols (C Cont) each time point represents mean SEM offour independent measurements in craniotomy control or TBIgroup p 001 versus craniotomy control (one-wayANOVA test with post hoc Bonferroni analysis)
1185
FIG 2 Expression of survivin protein after TBI in rats Brain tissue homogenate proteins (40 g) were separated using SDS-PAGE immunoblotted with survivin antibody and visualized (A) Representative Western blot of survivin (17-kDa protein) inipsilateral cortex (ic) and hippocampus (ih) contralateral cortex (cc) and hippocampus (ch) obtained from injured rats and fromcraniotomy control rats without cortical impact (C Cont) Densitometry analysis representation of survivin-positive bands in ip-silateral (ic) and contralateral (cc) cortex (B) and ipsilateral (ih) and contralateral (ch) hippocampus (C) after TBI is shown aspercent of craniotomy control values Each data point represents the mean SEM of four to six independent experiments p 005 p 0001 versus craniotomy control (one-way ANOVA test with post hoc Bonferroni analysis)
SURVIVIN UPREGULATION AFTER TBI
gen Total RNA was isolated from the samples using TRI-zol reagent (Invitrogen Carlsbad CA) according to themanufacturerrsquos instructions cDNA synthesis was per-formed using 1 g of total RNA with the SuperScripttradeFirst-Strand Synthesis System for RT-PCR kit (Invitro-genLife Technologies Carlsbad CA) according to the
manufacturerrsquos instructions Q-PCR was performed aspreviously described (Tolentino et al 2002) using theLightCycler-FastStart DNA Master SYBR Green I reac-tion mix (Roche Diagnostics Indianapolis IN) in com-bination with 05 M primers 25 mM MgCl2 in theLight Cycler rapid thermal cycler system (Roche Diag-
JOHNSON ET AL
1186
FIG 3 Expression of PCNA after TBI in rats PVDF membranes visualized for survivin were stripped and re-probed withPCNA antibody Representative western blots showing PCNA (36 kDa) (A) and densitometry analysis of PCNA-positive bands(BC) are presented Experimental conditions sample size and abbreviations are identical to those in Figure 2 p 005 p 001 versus craniotomy control (one-way ANOVA test with post hoc Bonferroni analysis)
SURVIVIN UPREGULATION AFTER TBI
1187
FIG 4 Immunohistochemistry of survivin and PCNA Double fluorescent immunostaining for survivin (red) and PCNA (green)was performed in the ipsilateral cortex (A) and hippocampus (B) at 5 day post-injury Survivin is expressed in the cytoplasm (Cred) while PCNA is expressed in the nucleus (D green) The white arrow indicates the typical focal co-expression of survivinand PCNA as shown in merged survivin and PCNA images (E) PCNA expression was co-incident with DAPI staining (F bluewhite arrow) Original magnification 200 bar 50 m (AB) original magnification 400 bar 20 m (CndashF)
nostics Indianapolis IN) Briefly the primers were ampli-fied then quantified by online monitoring and identificationof the crossing point value (CPV) which is the exact timepoint at which the logarithmic linear phase could be dis-tinguished from the background The survivin primer setswere subjected to serial dilution and linear regression analy-sis of the logarithm of the dilution factor versus the CPVgenerated a standard curve for each transcript-speci-fic tem-plate Results are presented as percentage of cranio-tomycontrol Data were analyzed by ANOVA with a post-hocBonferroni-test and are given as mean SEM Differenceswere considered significant at the level of p 005
Western Blot Analyses
Brain tissue was removed as described above rinsedwith cold PBS snap frozen in liquid nitrogen and ho-
mogenized in ice-cold triple detergent lysis buffer con-taining a Completetrade protease inhibitor cocktail (RocheBiochemicals Indianapolis IN) Protein concentrationwas determined by bicinchoninic acid (BCA) micro pro-tein assays (Pierce Inc Rockford IL) Forty microgramsof protein per well was loaded and separated by SDS-PAGE transferred to PVDF membranes and probed withgoat-anti-rabbit survivin antibody (Novus BiologicalsLittleton CO 11000) After incubation with goat anti-rabbit HRP-labeled secondary antibody (Biorad Her-cules CA) the membranes were developed using En-hanced Chemiluminescence Plus reagents (ECL PlusAmersham Arlington Heights IL) For further PCNAanalysis developed PVDF membranes were incubated instripping buffer rinsed twice in TBST and incubated withPCNA antibody (Santa Cruz Biotech Santa Cruz CA
11000) as described above Semi-quantitative densito-metric analysis was performed using the AlphaImagertrade2000 Digital Imaging System (San Leandro CA) Trans-formed data (experimental densitometry value crani-otomy control densitometry value 100) was evaluatedby ANOVA and a post-hoc Bonferroni-test Values areexpressed as percentage of craniotomy controls and aregiven as mean SEM Differences were considered sig-nificant at the level of p 005
Characterization of Survivin Antibody (R51)
First we compared the specificity of the survivin an-tibody developed within our group (R51 Dr G Shaw)and a commercially available survivin antibody (Chemi-con Temecula CA) Our antibody showed characteris-tic staining of the cleavage furrow between dividingHeLa cells consistent with other reports (Li et al 19981999 Uren et al 2000) In addition double labeling withboth survivin antibodies showed co-localization at thecleavage furrow The peptides used to develop our sur-vivin antibody are specific to survivin and do not recog-nize other IAP family proteins according to SDSC Biol-ogy Workbench BLASTP (222) (Altschul et al 1997)and CLUSTAL W (181) analysis (Higgins et al 1992Thompson et al 1994) resulting in the survivin anti-bodyrsquos specificity
Immunohistochemistry (IHC)
Animals were transcardially perfused with 2 Heparin(Elkins-Sinn Inc Cherry Hill NJ) in 09 saline solu-tion (pH 74) followed by 4 paraformaldehyde in 01M phosphate buffer (pH 74) The brains were post-fixedin 4 paraformaldehyde and stored in 01M PBS or cryo-buffer Forty micron sections were fluorescent immuno-labeled with two primary antibodies in the following experiments survivin (1500)GFAP for astrocytes(Sternberger Lutherville MD 11000) survivinNeuNfor mature neurons (Chemicon Temecula CA 11000)survivinPCNA (Santa Cruz Biotech Santa Cruz CA1200) PCNAGFAP and PCNANeuN The nuclear dyeDAPI (in Vectashield H-1200 Vector LaboratoriesBurlingame CA) was used to label the nuclei The firstprimary antibody was incubated at 4degC for 24ndash48 h in a2 goat serum2 horse serum02 Triton-X 100 in01 M PBS (block) solution followed by the second pri-mary antibody at 4degC for 1 h in block solution Fluores-cent-tagged secondary antibody (Molecular Probes Eu-gene OR) was used for visualization For double-labelingusing same species antibodies we used the tyramide sig-nal amplification (TSA) kit (PerkinElmer Life SciencesBoston MA) according to the manufacturerrsquos instruc-tions and as previously described (Stone et al 2002) The
sections were viewed and digitally captured with a ZeissAxioplan 2 microscope (Zeiss Thornwood NY)equipped with a SPOT Real Time Slider high-resolutioncolor CCD digital camera (Diagnostic Instruments IncSterling Heights MI) The number of animals used fordual-labeling IHC is as follows survivin PCNA 4survivin GFAP 6 survivin NeuN 4 PCNA GFAP 4 and PCNA NeuN 4 Cell counts wereobtained by comparing the number of double-labeledcells to total single-labeled cells in the following groupssurvivinNeuN positive cells to total NeuN positive cellssurvivinPCNA positive cells to total PCNA positivecells PCNANeuN positive cells to total NeuN positivecells and survivinGFAP positive cells to total GFAPpositive cells Percentages were calculated by dividingthe number of double-labeled cells with the total numberof single-labeled cells For each group representativephotomicrographs were selected and counted Cells werecounted in a total area of at least 188000 m2 for eachgroup with no distinction made between cortical and hip-pocampal regions
RESULTS
Induction of Survivin Expression after TBI
Q-PCR analysis revealed an initial increase in survivinmRNA at 2 days post injury in the ipsilateral cortex andhippocampus These transcripts remained elevated inboth regions reached maximum levels at day 5 post-in-jury and declined at 7 days in the cortex and at 14 daysin the hippocampus All experimental animals remainedalive and exhibited slightly impaired motor and cogni-tive impairments (data not shown) Cortical mRNA lev-els reached a maximum of 448 100 whereas hip-pocampal mRNAs attained 606 100 compared tocraniotomy control values (Fig 1) To determine if theinduction of survivin mRNA resulted in correspondingincreases in survivin protein western blot analysis wasperformed Survivin (17-kDa protein) was readily de-tectable in the ipsilateral cortex and hippocampus of TBIrats while it was negligible in contralateral cortex andhippocampus (Fig 2A) Survivin was expressed in atime-dependent manner with a maximum increase at 5days after injury followed by a gradual decline by 14days Specifically the levels of survivin in cortical tis-sue were 616 257 at 3 days and 839 339 at 5days compared to craniotomy controls (Fig 2B) Similarincreases of survivin protein in the ipsilateral hippocam-pus were detected at 3 d and 5 days post injury 464 196 and 545 102 compared to craniotomy controlrespectively (Fig 2C)
JOHNSON ET AL
1188
FIG 5 Co-localization of survivin and GFAP in brain tissue after TBI Fluorescent immunohistochemistry for survivin (green)and GFAP (red) was performed in the ipsilateral and contralateral cortex (AB) and in the CA1 and dentate gyrus regions of thehippocampus (GH) at 5 day post-injury The injury has completely destroyed the cortex in G leaving only the hippocampus inthis picture Survivin was expressed in the cytoplasm (DJ green) of GFAP-positive astrocytes (CI red) of the ipsilateral cor-tex and hippocampus and was found to co-localize to these cells as shown in merged CD and IJ images (EK respectively yel-low) White arrows indicate typical survivin-positive astrocytes Nuclei are shown using DAPI (FL blue) Original magnifica-tion 100 bar 50 m (ABGH) 400 bar 20 m (CndashF and IndashL)
FIG 6 Co-localization of PCNA and GFAP in brain tissue after TBI Double immunostaining for PCNA (green) and GFAP(red) was performed in the ipsilateral and contralateral cortex (AB) and the CA1 and dentate gyrus regions of the hippocampus(GH) at 5 day post-injury PCNA is present in GFAP-positive cells of ipsilateral cortex (CD) and to a lesser extent hippocampus(IJ) (EK) Merged CD and IJ respectively White arrows indicate typical PCNA-positive astrocytes PCNA expression wasco-incident with DAPI staining (FL blue) Original magnification 100 bar 50 m (ABGH) 400 bar 20 m (CndashFand IndashL)
PCNA Expression after TBI
For detection of proliferating cell nuclear antigen(PCNA) PVDF membranes immunostained for survivinwere stripped and re-probed using a PCNA-specific an-tibody PCNA (36-kDa protein) was significantly de-tectable in the ipsilateral cortex and hippocampus of TBIrats but only negligible amounts were observed in thecontralateral cortex and hippocampus (Fig 3A) Thetemporal patterns exhibited by PCNA protein were sim-ilar to that of survivin protein Namely PCNA expressedin a time-dependant fashion with a maximum increase at5 days after injury followed by a gradual decline by 14days The levels of PCNA in ipsilateral cortical tissuewere raised over craniotomy control by 919 459 at3 days 2263 333 at 5 days and 1035 356 at 7days post injury (Fig 3B) Similar increases of PCNAprotein in ipsilateral hippocampus were detected at 5 dayspost injury with a maximum of 1006 229 comparedto craniotomy controls (Fig 3C) No significant increasewas found in the contralateral regions when compared tocraniotomy controls (Fig 3A)
Co-Expression of Survivin and PCNA following TBI
To examine spatial co-localization of survivin andPCNA double-label immunohistochemistry of brain tis-sue sections was performed on day 5 post injury whenpeak expression of these proteins was observed
At this time point a remarkable survivin and PCNAimmunoreactivity was found in the ipsilateral cortex (Fig4A) and ipsilateral hippocampus (Fig 4B) consistent withdata obtained using Western blot analyses Within bothregions focal co-expression patterns of survivin andPCNA in single cells were detected which was demon-strated by both separate fluorescent visualization of in-dividual proteins and by merging the images of double-stained slides (Fig 4CndashE) However the dual expressionof survivin and PCNA occurred infrequently as survivinand PCNA immunoreactivity could readily be found sep-arately (Fig 4CndashE) Approximately 12 of the total num-ber of PCNA-positive cells also labeled with survivin Thenuclear morphology of dual survivin and PCNA-positivecells was ambiguous as indicated by DAPI staining (Fig4F) Therefore DAPI staining was simply used for cellidentification in all subsequent experiments
Survivin and PCNA Are Expressed in Astrocytes after TBI
To determine the cell types expressing survivin andPCNA double-label immunohistochemistry for theseproteins and GFAP a marker of astrocytes was per-formed on day 5 post injury
In accordance with Western blot data remarkable sur-vivin-positive immunoreactivity was observed in the ip-silateral cortex and hippocampus proximal to the injurycavity (Fig 5AG green) but not in the contralateral ar-eas (Fig 5BH) Survivin was co-localized with GFAPin the cells of injured cortex and hippocampus whichstrongly suggested primary accumulation of survivin incells of astrocytic lineage (Fig 5CndashEIndashL) Survivin wasuniformly distributed in the cytoplasm and processes ofastrocytes in both cortex and hippocampus (Fig 5DJ)DAPI staining is shown in Figure 5FL3 Approximately88 of the total number of GFAP-positive cells also la-beled with survivin
PCNA-positive immunoreactivity staining was ob-served in the ipsilateral cortex (Fig 6A green) and hip-pocampus (Fig 6G green) of injured brain while con-tralateral cortex and hippocampus exhibited negligiblePCNA immunoreactivity (Fig 6BH) PCNA (Fig 6CI)was partially co-localized with GFAP (Fig 6DJ red) inboth regions and was characteristically distributed in thenucleus of the cells in both cortex and hippocampus (Fig6EK) DAPI staining is shown in Figure 6FL
Taken together double-label immunohistochemistrydata provides evidence that both survivin and PCNA canbe detected in GFAP-positive astrocytes following trau-matic insult Since survivin and PCNA immunoreactiv-ity was not exclusively localized in GFAP-positive cellswe next addressed a question what other cell type mightexpress survivin after TBI We suggested that a certainpopulation of mature neurons might express survivin inresponse to injury
Survivin and PCNA Are Expressed in a Sub-Set of Neurons after TBI
As can be seen in Figure 7 survivin and PCNA wereeach co-expressed with NeuN a marker of mature neu-rons NeuN-positive cells were found to express survivinin the ipsilateral cortex distal to the injury cavity (Fig7AndashD) and in the contralateral hippocampus (Fig 7EndashH)It should be noted however that NeuN-positive cells thatalso expressed survivin occurred infrequently For ex-ample we estimated the number of dual survivinNeuNpositive cells as 01ndash15 of the total number of NeuN-positive cells in these regions Survivin immunoreactiv-ity was negligible in either hemisphere of craniotomycontrol brains (Fig 7QR) No co-localization of survivinand NeuN was observed in ipsilateral hippocampus (datanot shown) As can be seen in Figure 7BF survivin waspredominantly localized to the cytoplasm and axons ofNeuN-positive neurons DAPI staining is shown in Fig-ure 7DH
PCNA-positive neurons were found in the ipsilateralcortex (Fig 7IndashL) and hippocampus after TBI (Fig
JOHNSON ET AL
1190
1191
FIG 7 A sub-set of NeuN-positive neurons express survivin and PCNA after TBI Double fluorescent immunohistochemistryfor survivin (green) and NeuN (red) in the ipsilateral cortex (AB) and the CA1 pyramidal layer of the contralateral hippocam-pus (EF) was performed Survivin is expressed in the cytoplasm and to a limited extent in the processes of NeuN-positive neu-rons (merged images CG) Dual staining for PCNA (green) and NeuN (red) is shown in the ipsilateral cortex (IJ) and the CA1pyramidal layer of the ipsilateral hippocampus (MN) The nuclei are shown using DAPI staining (DH blue) PCNA is expressedin the nucleus of NeuN-positive neurons (merged images KO) PCNA expression was co-incident with DAPI staining in theseexamples (LP blue) White arrows indicate focal co-localization of survivinNeuN and PCNANeuN SurvivinNeuN co-local-ization of survivin (green) and NeuN (red) was seen only in TBI rats as opposed to either hemisphere of craniotomy control(QR) Original magnification 400 bar 20 m (AndashP) original magnification 50 bar 100000 m (QR)
SURVIVIN UPREGULATION AFTER TBI
7MndashP) whereas craniotomy control tissue exhibited onlytrace amounts of PCNA (data not shown) Similar to thesurvivinNeuN co-localization data dual PCNANeuNimmunostaining was a rare event accounting for approx-imately 4 of the total number of NeuN positive cellsPCNA was distributed in the nuclei of these neurons (Fig7KO) although the nuclear morphology of these cellswas not clearly resolved by DAPI staining (Fig 7LP)
DISCUSSION
Traumatic brain injury (TBI) sets into motion variousbiochemical cascades that induce neural tissue injury andcell death Conversely several proteins expressed inneural cells after TBI are directed to resist cell death andpromote recovery in the injured CNS (Ridet et al 1997Chen and Swanson 2003) Survivin is a multi-functionalprotein that inhibits apoptosis and is also required for theproper completion of mitosis Anti-apoptotic and pro-mi-togenic roles for survivin have been documented in pro-liferating cells of neural origin in vitro such as in neu-roblastoma and glioma cells (LaCasse et al 1998 Tammet al 1998 Deveraux and Reed 1999 Conway et al2000 Shin et al 2001 Sasaki et al 2002) Howeverno studies have investigated the potential role of survivinin the adult brain after TBI when a sub-population ofCNS cells may initiate a cell cycle-related process in re-sponse to injury
In the present paper we demonstrate the induction ofsurvivin expression in rat brain subjected to TBI The ex-pression of survivin was time-dependent and cell-spe-cific and was present in astrocytes and to a much lesserextent in neurons in brain cortex and hippocampus In-duction of survivin in these cells was accompanied byoccasional expression of PCNA a cell cycle protein in-volved in mitotic G1S progression Our present data arethe first to show that survivin mRNA and protein are sig-nificantly up-regulated after traumatic brain injury in ratsPCNA expression after TBI has been described previously(Miyake et al 1992 Chen et al 2003) suggesting itsrole in mechanisms of brain recovery after injury Theconcurrent up-regulation of survivin with a similar tem-poral profile as PCNA shown herein further suggests thatsurvivin may play a role in cellular proliferation after TBI
Brain injury evoked the expression of survivin andPCNA in a time-dependent manner (Figs 2 and 3) West-ern blot analysis revealed maximal co-expression of bothsurvivin and PCNA at five days post injury Immuno-histochemistry at this time point demonstrated co-local-ization of these proteins (Fig 4) although most cells werelabeled separately with PCNA and survivin In fact only12 of the total number of PCNA-positive cells were
also survivin positive It has been reported that PCNA isexpressed predominantly in G1S (Bravo et al 1987)while survivin is found at the G2M phase of the cell cy-cle (Bravo et al 1987 Otaki et al 2000) Hence a lackof strict co-localization of survivin and PCNA in ourstudy may be explained by their expression at differentpoints in the cell cycle
In our experimental model we observed survivin- aswell as PCNA-positive astrocytes in the proximal area ofthe injury and in the ipsilateral hippocampus Prolifera-tion of astrocytes is well documented after TBI as shownby cell labeling with BrdU as well as expression of PCNA(Latov et al 1979 Dunn-Meynell and Levin 1997 Car-bonell and Grady 1999 Norton 1999 Csuka et al 2000Kernie et al 2001 Chen et al 2003) Because survivinand PCNA were expressed in astrocytes following TBI(Figs 5 and 6) it is possible that survivin plays an im-portant role linking astrocyte survival and proliferationafter traumatic insult Astrocyte proliferation has beenimplicated in the formation of the glial scar observed af-ter injury (Latov et al 1979) and creates a non-permis-sive environment for repair (Sykova et al 1999) How-ever glial proliferation may also enhance neuronalsurvival (Smith et al 2001 Wei et al 2001)
Of particular interest is a sub-set of NeuN-positive neu-rons found to express survivin only after TBI (Fig 7)These cells were much less abundant than survivin-pos-itive astrocytes and their functional significance is cur-rently unknown However both neurons and astrocyteshave been documented previously to express cell cycleproteins after various insults such as exposure to -amy-loidndashactivated microglia (Wu et al 2000) TBI (Kaya etal 1999ab) chlorine toxicity (Magavi et al 2000) oras a consequence of Alzheimerrsquos disease (Yang et al2001) These papers underscore the significant contro-versy that exists regarding the function of cell cycle pro-teins such as PCNA in neurons after different types ofinjury We are currently conducting further studies whichwill elucidate the roles for PCNA and survivin in neu-rons after TBI
It should be noted that dual staining of survivin andPCNA could not be directly attributed to a specific celltype due to the technical difficulties of triple labeling an-tibody-based IHC Therefore we cannot rule out the pos-sibility that other cell types such as endothelial (Conwayet al 2003) or inflammatory cells (Hill-Felberg et al1999) may also contribute to survivin and PCNA ex-pression after TBI The appearance of survivin and PCNAseparately in neurons (NeuN-positive) and astrocytes(GFAP-positive) along with co-localization of survivinwith PCNA in the same cells provide correlative data tosuggest an activation of cell cycle-like program in astro-cytes and possibly in a small subtype of neurons after
JOHNSON ET AL
1192
TBI In our experiments survivin co-localization withPCNA does suggest that survivin may be associated witha pro-mitotic process In an attempt to clarify these pro-teinrsquos roles after TBI we analyzed the nuclear morphol-ogy of survivin-positive cells to define the apoptotic ormitotic architecture of nuclei DAPI staining proved tooambiguous in identifying apoptotic versus mitotic phe-notypes likely due to the thickness of the brain sections(40 m) Further studies using direct markers of mitosissuch as BrdU incorporation as well as simultaneous la-beling with cell death related proteins is required to de-lineate anti-apoptotic and pro-mitotic activities of sur-vivin and PCNA in these cells
In conclusion our data demonstrate the induction ofsurvivin in the rat brain following TBI Expression of sur-vivin occurred predominantly in astrocytes as comparedto neurons in a time-dependant fashion and was accom-panied by expression of PCNA Taken together these re-sults suggest that survivin plays a role in neural cell re-sponses following traumatic brain injury in rats Futurestudies will investigate the implications of these findingsto the pathophysiology of TBI
ACKNOWLEDGMENTS
We thank Barbara OrsquoSteen and Tao Fan MD fortechnical assistance This work was funded by grants NIHRO1 NS39091 NIH RO1 NS40182 DAMD 17-99-1-9565 and DAMD 17-01-1-0765
REFERENCES
ADIDA C CROTTY PL MCGRATH J BERREBI DDIEBOLD J and ALTIERI DC (1998) Developmentallyregulated expression of the novel cancer anti-apoptosis genesurvivin in human and mouse differentiation Am J Pathol152 43ndash49
ALTIERI DC MARCHISIO PC and MARCHISIO C(1999) Survivin apoptosis an interloper between cell deathand cell proliferation in cancer Lab Invest 79 1327ndash1333
ALTSCHUL SF MADDEN TL SCHAFFER AA et al(1997) Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 253389ndash3402
AMBROSINI G ADIDA C and ALTIERI DC (1997) Anovel anti-apoptosis gene survivin expressed in cancer andlymphoma Nat Med 3 917ndash921
BRAVO R FRANK R BLUNDELL PA and MAC-DONALD-BRAVO H (1987) CyclinPCNA is the auxil-iary protein of DNA polymerase-delta Nature 326 515ndash517
BUSH TG PUVANACHANDRA N HORNER CH et al
(1999) Leukocyte infiltration neuronal degeneration andneurite outgrowth after ablation of scar-forming reactive as-trocytes in adult transgenic mice Neuron 23 297ndash308
CAMERON HA and MCKAY RD (2001) Adult neuroge-nesis produces a large pool of new granule cells in the den-tate gyrus J Comp Neurol 435 406ndash417
CARBONELL WS and GRADY MS (1999) Regional andtemporal characterization of neuronal glial and axonal re-sponse after traumatic brain injury in the mouse Acta Neu-ropathol (Berl) 98 396ndash406
CHEN XH IWATA A NONAKA M BROWNE KDand SMITH DH (2003) Neurogenesis and glial prolifera-tion persist for at least one year in the subventricular zonefollowing brain trauma in rats J Neurotrauma 20 623ndash631
CHEN Y and SWANSON RA (2003) Astrocytes and braininjury J Cereb Blood Flow Metab 23 137ndash149
CONWAY EM POLLEFEYT S CORNELISSEN J et al(2000) Three differentially expressed survivin cDNA vari-ants encode proteins with distinct antiapoptotic functionsBlood 95 1435ndash1442
CSUKA E HANS VH AMMANN E TRENTZ OKOSSMANN T and MORGANTI-KOSSMANN MC(2000) Cell activation and inflammatory response followingtraumatic axonal injury in the rat Neuroreport 112587ndash2590
DEVERAUX QL and REED JC (1999) IAP family pro-teinsmdashsuppressors of apoptosis Genes Dev 13 239ndash252
DIXON CE CLIFTON GL LIGHTHALL JW YAGH-MAI AA and HAYES RL (1991) A controlled corticalimpact model of traumatic brain injury in the rat J NeurosciMethods 39 253ndash262
DOETSCH F CAILLE I LIM DA GARCIA-VERDUGOJM and ALVAREZ-BUYLLA A (1999) Subventricularzone astrocytes are neural stem cells in the adult mammalianbrain Cell 97 703ndash716
DUNN-MEYNELL AA and LEVIN BE (1997) Histolog-ical markers of neuronal axonal and astrocytic changes af-ter lateral rigid impact traumatic brain injury Brain Res 76125ndash41
GIULIAN D (1991) Microgliamdashneuron interactions after in-jury to the central nervous system in Peripheral Signalingof the Brain Role in Neural-Immune Interactions Learningand Memory R Frederickson J McGaugh and D Felton(eds) Lewiston NY Hogrefe and Huber pps 73ndash82
HIGGINS DG BLEASBY AJ and FUCHS R (1992)CLUSTAL V improved software for multiple sequencealignment Comput Appl Biosci 8 189ndash191
HILL-FELBERG SJ MCINTOSH TK OLIVER DLRAGHUPATHI R and BARBARESE E (1999) Concur-rent loss and proliferation of astrocytes following lateral fluidpercussion brain injury in the adult rat J Neurosci Res 57271ndash279
1193
SURVIVIN UPREGULATION AFTER TBI
JIANG X WILFORD C DUENSING S MUNGER KJONES G and JONES D (2001) Participation of Survivinin mitotic and apoptotic activities of normal and tumor-de-rived cells J Cell Biochem 83 342ndash354
KAYA SS MAHMOOD A LI Y YAVUZ E andCHOPP M (1999a) Expression of cell cycle proteins (cy-clin D1 and cdk4) after controlled cortical impact in rat brainJ Neurotrauma 16 1187ndash1196
KAYA SS MAHMOOD A LI Y YAVUZ E GOKSELM and CHOPP M (1999b) Apoptosis and expression ofp53 response proteins and cyclin D1 after cortical impact inrat brain Brain Res 818 23ndash33
KERNIE SG ERWIN TM and PARADA LF (2001)Brain remodeling due to neuronal and astrocytic prolifera-tion after controlled cortical injury in mice J Neurosci Res66 317ndash326
KOBAYASHI K HATANO M OTAKI M OGA-SAWARA T and TOKUHISA T (1999) Expression of amurine homologue of the inhibitor of apoptosis protein is re-lated to cell proliferation Proc Natl Acad Sci USA 961457ndash1462
LACASSE EC BAIRD S KORNELUK RG andMACKENZIE AE (1998) The inhibitors of apoptosis(IAPs) and their emerging role in cancer Oncogene 173247ndash3259
LATOV N NILAVER G ZIMMERMAN EA et al(1979) Fibrillary astrocytes proliferate in response to braininjury a study combining immunoperoxidase technique forglial fibrillary acidic protein and radioautography of tritiatedthymidine Dev Biol 72 381ndash384
LI F and ALTIERI DC (1999) The cancer antiapoptosismouse survivin gene characterization of locus and tran-scriptional requirements of basal and cell cycle-dependentexpression Cancer Res 59 3143ndash3151
LI F AMBROSINI G CHU EY et al (1998) Control ofapoptosis and mitotic spindle checkpoint by survivin Nature396 580ndash584
LI F ACKERMANN EJ BENNETT CF et al (1999)Pleiotropic cell-division defects and apoptosis induced by in-terference with survivin function Nat Cell Biol 1 461ndash466
MAGAVI SS LEAVITT BR and MACKLIS JD (2000)Induction of neurogenesis in the neocortex of adult mice Na-ture 405 951ndash955
MIYAKE T OKADA M and KITAMURA T (1992) Re-active proliferation of astrocytes studied by immunohisto-chemistry for proliferating cell nuclear antigen Brain Res590 300ndash302
MUCHMORE SW CHEN J JAKOB C et al (2000)Crystal structure and mutagenic analysis of the inhibitor-of-apoptosis protein survivin Mol Cell 6 173ndash182
NORTON WT (1999) Cell reactions following acute braininjury a review Neurochem Res 24 213ndash218
OTAKI M HATANO M KOBAYASHI K et al (2000)Cell cyclendashdependent regulation of TIAPm-survivin expres-sion Biochim Biophys Acta 1493 188ndash194
PETERSON DA (2002) Stem cells in brain plasticity and re-pair Curr Opin Pharmacol 2 34ndash42
PIKE BR ZHAO X NEWCOMB JK POSMANTURRM WANG KK and Hayes RL (1998) Regional cal-pain and caspase-3 proteolysis of alpha-spectrin after trau-matic brain injury Neuroreport 9 2437ndash2442
RAGHUPATHI R GRAHAM DI MCINTOSH TK(2000) Apoptosis after traumatic brain injury J Neuro-trauma 17 927ndash938
RIDET JL MALHOTRA SK PRIVAT A and GAGEFH (1997) Reactive astrocytes cellular and molecular cuesto biological function Trends Neurosci 20 570ndash577
SASAKI T LOPES MB HANKINS GR and HELMGA (2002) Expression of survivin an inhibitor of apopto-sis protein in tumors of the nervous system Acta Neu-ropathol (Berl) 104 105ndash109
SHANKAR SL MANI S OrsquoGUIN KN KANDIMALLAER AGRAWAL S and SHAFIT-ZAGARDO B (2001)Survivin inhibition induces human neural tumor cell deaththrough caspase-independent and -dependent pathways JNeurochem 79 426ndash436
SHIN S SUNG BJ CHO YS et al (2001) An anti-apop-totic protein human survivin is a direct inhibitor of caspase-3 and -7 Biochemistry 40 1117ndash1123
SMITH C BERRY M CLARKE WE and LOGAN A(2001) Differential expression of fibroblast growth factorndash2and fibroblast growth factor receptor 1 in a scarring and non-scarring model of CNS injury in the rat Eur J Neurosci 13443ndash456
STONE JR OKONKWO DO SINGLETON RHMUTLU LK HELM GA and POVLISHOCK JT(2002) Caspase-3ndashmediated cleavage of amyloid precursorprotein and formation of amyloid Beta peptide in traumaticaxonal injury J Neurotrauma 19 601ndash614
SYKOVA E VARGOVA L PROKOPOVA S and SI-MONOVA Z (1999) Glial swelling and astrogliosis pro-duce diffusion barriers in the rat spinal cord Glia 25 56ndash70
TAKAHASHI R DEVERAUX Q TAMM I et al (1998)A single BIR domain of XIAP sufficient for inhibiting cas-pases J Biol Chem 273 7787ndash7790
TAMM I WANG Y SAUSVILLE E et al (1998) IAP-family protein survivin inhibits caspase activity and apopto-sis induced by Fas (CD95) Bax caspases and anticancerdrugs Cancer Res 58 5315ndash5320
THOMPSON JD HIGGINS DG and GIBSON TJ (1994)CLUSTAL W improving the sensitivity of progressive mul-tiple sequence alignment through sequence weighting posi-tion-specific gap penalties and weight matrix choice NucleicAcids Res 22 4673ndash4680
JOHNSON ET AL
1194
TOLENTINO PJ DEFORD SM NOTTERPEK L et al(2002) Up-regulation of tissue-type transglutaminase aftertraumatic brain injury J Neurochem 80 579ndash588
UREN AG WONG L PAKUSCH M et al (2000) Sur-vivin and the inner centromere protein INCENP show simi-lar cell-cycle localization and gene knockout phenotypeCurr Biol 10 1319ndash1328
WEI LH HUANG CY CHENG SP CHEN CA andHSIEH CY (2001) Carcinosarcoma of ovary associatedwith previous radiotherapy Int J Gynecol Cancer 11 81ndash84
WU Q COMBS C CANNADY SB GELDMACHERDS and HERRUP K (2000) Beta-amyloid activated mi-croglia induce cell cycling and cell death in cultured corticalneurons Neurobiol Aging 21 797ndash806
YAGITA Y KITAGAWA K OHTSUKI T et al (2001)
Neurogenesis by progenitor cells in the ischemic adult rathippocampus Stroke 32 1890ndash1896
YANG Y GELDMACHER DS and HERRUP K (2001)DNA replication precedes neuronal cell death in Alzheimerrsquosdisease J Neurosci 21 2661ndash2668
Address reprint requests toJose A Pineda PhD
EF and WL McKnight Brain InstituteUniversity of Florida
100 S Newell Dr Bldg 59L1-118 Dept of Pediatrics
Gainesville FL 32610
E-mail pinedjapedsufledu
SURVIVIN UPREGULATION AFTER TBI
1195

1185
FIG 2 Expression of survivin protein after TBI in rats Brain tissue homogenate proteins (40 g) were separated using SDS-PAGE immunoblotted with survivin antibody and visualized (A) Representative Western blot of survivin (17-kDa protein) inipsilateral cortex (ic) and hippocampus (ih) contralateral cortex (cc) and hippocampus (ch) obtained from injured rats and fromcraniotomy control rats without cortical impact (C Cont) Densitometry analysis representation of survivin-positive bands in ip-silateral (ic) and contralateral (cc) cortex (B) and ipsilateral (ih) and contralateral (ch) hippocampus (C) after TBI is shown aspercent of craniotomy control values Each data point represents the mean SEM of four to six independent experiments p 005 p 0001 versus craniotomy control (one-way ANOVA test with post hoc Bonferroni analysis)
SURVIVIN UPREGULATION AFTER TBI
gen Total RNA was isolated from the samples using TRI-zol reagent (Invitrogen Carlsbad CA) according to themanufacturerrsquos instructions cDNA synthesis was per-formed using 1 g of total RNA with the SuperScripttradeFirst-Strand Synthesis System for RT-PCR kit (Invitro-genLife Technologies Carlsbad CA) according to the
manufacturerrsquos instructions Q-PCR was performed aspreviously described (Tolentino et al 2002) using theLightCycler-FastStart DNA Master SYBR Green I reac-tion mix (Roche Diagnostics Indianapolis IN) in com-bination with 05 M primers 25 mM MgCl2 in theLight Cycler rapid thermal cycler system (Roche Diag-
JOHNSON ET AL
1186
FIG 3 Expression of PCNA after TBI in rats PVDF membranes visualized for survivin were stripped and re-probed withPCNA antibody Representative western blots showing PCNA (36 kDa) (A) and densitometry analysis of PCNA-positive bands(BC) are presented Experimental conditions sample size and abbreviations are identical to those in Figure 2 p 005 p 001 versus craniotomy control (one-way ANOVA test with post hoc Bonferroni analysis)
SURVIVIN UPREGULATION AFTER TBI
1187
FIG 4 Immunohistochemistry of survivin and PCNA Double fluorescent immunostaining for survivin (red) and PCNA (green)was performed in the ipsilateral cortex (A) and hippocampus (B) at 5 day post-injury Survivin is expressed in the cytoplasm (Cred) while PCNA is expressed in the nucleus (D green) The white arrow indicates the typical focal co-expression of survivinand PCNA as shown in merged survivin and PCNA images (E) PCNA expression was co-incident with DAPI staining (F bluewhite arrow) Original magnification 200 bar 50 m (AB) original magnification 400 bar 20 m (CndashF)
nostics Indianapolis IN) Briefly the primers were ampli-fied then quantified by online monitoring and identificationof the crossing point value (CPV) which is the exact timepoint at which the logarithmic linear phase could be dis-tinguished from the background The survivin primer setswere subjected to serial dilution and linear regression analy-sis of the logarithm of the dilution factor versus the CPVgenerated a standard curve for each transcript-speci-fic tem-plate Results are presented as percentage of cranio-tomycontrol Data were analyzed by ANOVA with a post-hocBonferroni-test and are given as mean SEM Differenceswere considered significant at the level of p 005
Western Blot Analyses
Brain tissue was removed as described above rinsedwith cold PBS snap frozen in liquid nitrogen and ho-
mogenized in ice-cold triple detergent lysis buffer con-taining a Completetrade protease inhibitor cocktail (RocheBiochemicals Indianapolis IN) Protein concentrationwas determined by bicinchoninic acid (BCA) micro pro-tein assays (Pierce Inc Rockford IL) Forty microgramsof protein per well was loaded and separated by SDS-PAGE transferred to PVDF membranes and probed withgoat-anti-rabbit survivin antibody (Novus BiologicalsLittleton CO 11000) After incubation with goat anti-rabbit HRP-labeled secondary antibody (Biorad Her-cules CA) the membranes were developed using En-hanced Chemiluminescence Plus reagents (ECL PlusAmersham Arlington Heights IL) For further PCNAanalysis developed PVDF membranes were incubated instripping buffer rinsed twice in TBST and incubated withPCNA antibody (Santa Cruz Biotech Santa Cruz CA
11000) as described above Semi-quantitative densito-metric analysis was performed using the AlphaImagertrade2000 Digital Imaging System (San Leandro CA) Trans-formed data (experimental densitometry value crani-otomy control densitometry value 100) was evaluatedby ANOVA and a post-hoc Bonferroni-test Values areexpressed as percentage of craniotomy controls and aregiven as mean SEM Differences were considered sig-nificant at the level of p 005
Characterization of Survivin Antibody (R51)
First we compared the specificity of the survivin an-tibody developed within our group (R51 Dr G Shaw)and a commercially available survivin antibody (Chemi-con Temecula CA) Our antibody showed characteris-tic staining of the cleavage furrow between dividingHeLa cells consistent with other reports (Li et al 19981999 Uren et al 2000) In addition double labeling withboth survivin antibodies showed co-localization at thecleavage furrow The peptides used to develop our sur-vivin antibody are specific to survivin and do not recog-nize other IAP family proteins according to SDSC Biol-ogy Workbench BLASTP (222) (Altschul et al 1997)and CLUSTAL W (181) analysis (Higgins et al 1992Thompson et al 1994) resulting in the survivin anti-bodyrsquos specificity
Immunohistochemistry (IHC)
Animals were transcardially perfused with 2 Heparin(Elkins-Sinn Inc Cherry Hill NJ) in 09 saline solu-tion (pH 74) followed by 4 paraformaldehyde in 01M phosphate buffer (pH 74) The brains were post-fixedin 4 paraformaldehyde and stored in 01M PBS or cryo-buffer Forty micron sections were fluorescent immuno-labeled with two primary antibodies in the following experiments survivin (1500)GFAP for astrocytes(Sternberger Lutherville MD 11000) survivinNeuNfor mature neurons (Chemicon Temecula CA 11000)survivinPCNA (Santa Cruz Biotech Santa Cruz CA1200) PCNAGFAP and PCNANeuN The nuclear dyeDAPI (in Vectashield H-1200 Vector LaboratoriesBurlingame CA) was used to label the nuclei The firstprimary antibody was incubated at 4degC for 24ndash48 h in a2 goat serum2 horse serum02 Triton-X 100 in01 M PBS (block) solution followed by the second pri-mary antibody at 4degC for 1 h in block solution Fluores-cent-tagged secondary antibody (Molecular Probes Eu-gene OR) was used for visualization For double-labelingusing same species antibodies we used the tyramide sig-nal amplification (TSA) kit (PerkinElmer Life SciencesBoston MA) according to the manufacturerrsquos instruc-tions and as previously described (Stone et al 2002) The
sections were viewed and digitally captured with a ZeissAxioplan 2 microscope (Zeiss Thornwood NY)equipped with a SPOT Real Time Slider high-resolutioncolor CCD digital camera (Diagnostic Instruments IncSterling Heights MI) The number of animals used fordual-labeling IHC is as follows survivin PCNA 4survivin GFAP 6 survivin NeuN 4 PCNA GFAP 4 and PCNA NeuN 4 Cell counts wereobtained by comparing the number of double-labeledcells to total single-labeled cells in the following groupssurvivinNeuN positive cells to total NeuN positive cellssurvivinPCNA positive cells to total PCNA positivecells PCNANeuN positive cells to total NeuN positivecells and survivinGFAP positive cells to total GFAPpositive cells Percentages were calculated by dividingthe number of double-labeled cells with the total numberof single-labeled cells For each group representativephotomicrographs were selected and counted Cells werecounted in a total area of at least 188000 m2 for eachgroup with no distinction made between cortical and hip-pocampal regions
RESULTS
Induction of Survivin Expression after TBI
Q-PCR analysis revealed an initial increase in survivinmRNA at 2 days post injury in the ipsilateral cortex andhippocampus These transcripts remained elevated inboth regions reached maximum levels at day 5 post-in-jury and declined at 7 days in the cortex and at 14 daysin the hippocampus All experimental animals remainedalive and exhibited slightly impaired motor and cogni-tive impairments (data not shown) Cortical mRNA lev-els reached a maximum of 448 100 whereas hip-pocampal mRNAs attained 606 100 compared tocraniotomy control values (Fig 1) To determine if theinduction of survivin mRNA resulted in correspondingincreases in survivin protein western blot analysis wasperformed Survivin (17-kDa protein) was readily de-tectable in the ipsilateral cortex and hippocampus of TBIrats while it was negligible in contralateral cortex andhippocampus (Fig 2A) Survivin was expressed in atime-dependent manner with a maximum increase at 5days after injury followed by a gradual decline by 14days Specifically the levels of survivin in cortical tis-sue were 616 257 at 3 days and 839 339 at 5days compared to craniotomy controls (Fig 2B) Similarincreases of survivin protein in the ipsilateral hippocam-pus were detected at 3 d and 5 days post injury 464 196 and 545 102 compared to craniotomy controlrespectively (Fig 2C)
JOHNSON ET AL
1188
FIG 5 Co-localization of survivin and GFAP in brain tissue after TBI Fluorescent immunohistochemistry for survivin (green)and GFAP (red) was performed in the ipsilateral and contralateral cortex (AB) and in the CA1 and dentate gyrus regions of thehippocampus (GH) at 5 day post-injury The injury has completely destroyed the cortex in G leaving only the hippocampus inthis picture Survivin was expressed in the cytoplasm (DJ green) of GFAP-positive astrocytes (CI red) of the ipsilateral cor-tex and hippocampus and was found to co-localize to these cells as shown in merged CD and IJ images (EK respectively yel-low) White arrows indicate typical survivin-positive astrocytes Nuclei are shown using DAPI (FL blue) Original magnifica-tion 100 bar 50 m (ABGH) 400 bar 20 m (CndashF and IndashL)
FIG 6 Co-localization of PCNA and GFAP in brain tissue after TBI Double immunostaining for PCNA (green) and GFAP(red) was performed in the ipsilateral and contralateral cortex (AB) and the CA1 and dentate gyrus regions of the hippocampus(GH) at 5 day post-injury PCNA is present in GFAP-positive cells of ipsilateral cortex (CD) and to a lesser extent hippocampus(IJ) (EK) Merged CD and IJ respectively White arrows indicate typical PCNA-positive astrocytes PCNA expression wasco-incident with DAPI staining (FL blue) Original magnification 100 bar 50 m (ABGH) 400 bar 20 m (CndashFand IndashL)
PCNA Expression after TBI
For detection of proliferating cell nuclear antigen(PCNA) PVDF membranes immunostained for survivinwere stripped and re-probed using a PCNA-specific an-tibody PCNA (36-kDa protein) was significantly de-tectable in the ipsilateral cortex and hippocampus of TBIrats but only negligible amounts were observed in thecontralateral cortex and hippocampus (Fig 3A) Thetemporal patterns exhibited by PCNA protein were sim-ilar to that of survivin protein Namely PCNA expressedin a time-dependant fashion with a maximum increase at5 days after injury followed by a gradual decline by 14days The levels of PCNA in ipsilateral cortical tissuewere raised over craniotomy control by 919 459 at3 days 2263 333 at 5 days and 1035 356 at 7days post injury (Fig 3B) Similar increases of PCNAprotein in ipsilateral hippocampus were detected at 5 dayspost injury with a maximum of 1006 229 comparedto craniotomy controls (Fig 3C) No significant increasewas found in the contralateral regions when compared tocraniotomy controls (Fig 3A)
Co-Expression of Survivin and PCNA following TBI
To examine spatial co-localization of survivin andPCNA double-label immunohistochemistry of brain tis-sue sections was performed on day 5 post injury whenpeak expression of these proteins was observed
At this time point a remarkable survivin and PCNAimmunoreactivity was found in the ipsilateral cortex (Fig4A) and ipsilateral hippocampus (Fig 4B) consistent withdata obtained using Western blot analyses Within bothregions focal co-expression patterns of survivin andPCNA in single cells were detected which was demon-strated by both separate fluorescent visualization of in-dividual proteins and by merging the images of double-stained slides (Fig 4CndashE) However the dual expressionof survivin and PCNA occurred infrequently as survivinand PCNA immunoreactivity could readily be found sep-arately (Fig 4CndashE) Approximately 12 of the total num-ber of PCNA-positive cells also labeled with survivin Thenuclear morphology of dual survivin and PCNA-positivecells was ambiguous as indicated by DAPI staining (Fig4F) Therefore DAPI staining was simply used for cellidentification in all subsequent experiments
Survivin and PCNA Are Expressed in Astrocytes after TBI
To determine the cell types expressing survivin andPCNA double-label immunohistochemistry for theseproteins and GFAP a marker of astrocytes was per-formed on day 5 post injury
In accordance with Western blot data remarkable sur-vivin-positive immunoreactivity was observed in the ip-silateral cortex and hippocampus proximal to the injurycavity (Fig 5AG green) but not in the contralateral ar-eas (Fig 5BH) Survivin was co-localized with GFAPin the cells of injured cortex and hippocampus whichstrongly suggested primary accumulation of survivin incells of astrocytic lineage (Fig 5CndashEIndashL) Survivin wasuniformly distributed in the cytoplasm and processes ofastrocytes in both cortex and hippocampus (Fig 5DJ)DAPI staining is shown in Figure 5FL3 Approximately88 of the total number of GFAP-positive cells also la-beled with survivin
PCNA-positive immunoreactivity staining was ob-served in the ipsilateral cortex (Fig 6A green) and hip-pocampus (Fig 6G green) of injured brain while con-tralateral cortex and hippocampus exhibited negligiblePCNA immunoreactivity (Fig 6BH) PCNA (Fig 6CI)was partially co-localized with GFAP (Fig 6DJ red) inboth regions and was characteristically distributed in thenucleus of the cells in both cortex and hippocampus (Fig6EK) DAPI staining is shown in Figure 6FL
Taken together double-label immunohistochemistrydata provides evidence that both survivin and PCNA canbe detected in GFAP-positive astrocytes following trau-matic insult Since survivin and PCNA immunoreactiv-ity was not exclusively localized in GFAP-positive cellswe next addressed a question what other cell type mightexpress survivin after TBI We suggested that a certainpopulation of mature neurons might express survivin inresponse to injury
Survivin and PCNA Are Expressed in a Sub-Set of Neurons after TBI
As can be seen in Figure 7 survivin and PCNA wereeach co-expressed with NeuN a marker of mature neu-rons NeuN-positive cells were found to express survivinin the ipsilateral cortex distal to the injury cavity (Fig7AndashD) and in the contralateral hippocampus (Fig 7EndashH)It should be noted however that NeuN-positive cells thatalso expressed survivin occurred infrequently For ex-ample we estimated the number of dual survivinNeuNpositive cells as 01ndash15 of the total number of NeuN-positive cells in these regions Survivin immunoreactiv-ity was negligible in either hemisphere of craniotomycontrol brains (Fig 7QR) No co-localization of survivinand NeuN was observed in ipsilateral hippocampus (datanot shown) As can be seen in Figure 7BF survivin waspredominantly localized to the cytoplasm and axons ofNeuN-positive neurons DAPI staining is shown in Fig-ure 7DH
PCNA-positive neurons were found in the ipsilateralcortex (Fig 7IndashL) and hippocampus after TBI (Fig
JOHNSON ET AL
1190
1191
FIG 7 A sub-set of NeuN-positive neurons express survivin and PCNA after TBI Double fluorescent immunohistochemistryfor survivin (green) and NeuN (red) in the ipsilateral cortex (AB) and the CA1 pyramidal layer of the contralateral hippocam-pus (EF) was performed Survivin is expressed in the cytoplasm and to a limited extent in the processes of NeuN-positive neu-rons (merged images CG) Dual staining for PCNA (green) and NeuN (red) is shown in the ipsilateral cortex (IJ) and the CA1pyramidal layer of the ipsilateral hippocampus (MN) The nuclei are shown using DAPI staining (DH blue) PCNA is expressedin the nucleus of NeuN-positive neurons (merged images KO) PCNA expression was co-incident with DAPI staining in theseexamples (LP blue) White arrows indicate focal co-localization of survivinNeuN and PCNANeuN SurvivinNeuN co-local-ization of survivin (green) and NeuN (red) was seen only in TBI rats as opposed to either hemisphere of craniotomy control(QR) Original magnification 400 bar 20 m (AndashP) original magnification 50 bar 100000 m (QR)
SURVIVIN UPREGULATION AFTER TBI
7MndashP) whereas craniotomy control tissue exhibited onlytrace amounts of PCNA (data not shown) Similar to thesurvivinNeuN co-localization data dual PCNANeuNimmunostaining was a rare event accounting for approx-imately 4 of the total number of NeuN positive cellsPCNA was distributed in the nuclei of these neurons (Fig7KO) although the nuclear morphology of these cellswas not clearly resolved by DAPI staining (Fig 7LP)
DISCUSSION
Traumatic brain injury (TBI) sets into motion variousbiochemical cascades that induce neural tissue injury andcell death Conversely several proteins expressed inneural cells after TBI are directed to resist cell death andpromote recovery in the injured CNS (Ridet et al 1997Chen and Swanson 2003) Survivin is a multi-functionalprotein that inhibits apoptosis and is also required for theproper completion of mitosis Anti-apoptotic and pro-mi-togenic roles for survivin have been documented in pro-liferating cells of neural origin in vitro such as in neu-roblastoma and glioma cells (LaCasse et al 1998 Tammet al 1998 Deveraux and Reed 1999 Conway et al2000 Shin et al 2001 Sasaki et al 2002) Howeverno studies have investigated the potential role of survivinin the adult brain after TBI when a sub-population ofCNS cells may initiate a cell cycle-related process in re-sponse to injury
In the present paper we demonstrate the induction ofsurvivin expression in rat brain subjected to TBI The ex-pression of survivin was time-dependent and cell-spe-cific and was present in astrocytes and to a much lesserextent in neurons in brain cortex and hippocampus In-duction of survivin in these cells was accompanied byoccasional expression of PCNA a cell cycle protein in-volved in mitotic G1S progression Our present data arethe first to show that survivin mRNA and protein are sig-nificantly up-regulated after traumatic brain injury in ratsPCNA expression after TBI has been described previously(Miyake et al 1992 Chen et al 2003) suggesting itsrole in mechanisms of brain recovery after injury Theconcurrent up-regulation of survivin with a similar tem-poral profile as PCNA shown herein further suggests thatsurvivin may play a role in cellular proliferation after TBI
Brain injury evoked the expression of survivin andPCNA in a time-dependent manner (Figs 2 and 3) West-ern blot analysis revealed maximal co-expression of bothsurvivin and PCNA at five days post injury Immuno-histochemistry at this time point demonstrated co-local-ization of these proteins (Fig 4) although most cells werelabeled separately with PCNA and survivin In fact only12 of the total number of PCNA-positive cells were
also survivin positive It has been reported that PCNA isexpressed predominantly in G1S (Bravo et al 1987)while survivin is found at the G2M phase of the cell cy-cle (Bravo et al 1987 Otaki et al 2000) Hence a lackof strict co-localization of survivin and PCNA in ourstudy may be explained by their expression at differentpoints in the cell cycle
In our experimental model we observed survivin- aswell as PCNA-positive astrocytes in the proximal area ofthe injury and in the ipsilateral hippocampus Prolifera-tion of astrocytes is well documented after TBI as shownby cell labeling with BrdU as well as expression of PCNA(Latov et al 1979 Dunn-Meynell and Levin 1997 Car-bonell and Grady 1999 Norton 1999 Csuka et al 2000Kernie et al 2001 Chen et al 2003) Because survivinand PCNA were expressed in astrocytes following TBI(Figs 5 and 6) it is possible that survivin plays an im-portant role linking astrocyte survival and proliferationafter traumatic insult Astrocyte proliferation has beenimplicated in the formation of the glial scar observed af-ter injury (Latov et al 1979) and creates a non-permis-sive environment for repair (Sykova et al 1999) How-ever glial proliferation may also enhance neuronalsurvival (Smith et al 2001 Wei et al 2001)
Of particular interest is a sub-set of NeuN-positive neu-rons found to express survivin only after TBI (Fig 7)These cells were much less abundant than survivin-pos-itive astrocytes and their functional significance is cur-rently unknown However both neurons and astrocyteshave been documented previously to express cell cycleproteins after various insults such as exposure to -amy-loidndashactivated microglia (Wu et al 2000) TBI (Kaya etal 1999ab) chlorine toxicity (Magavi et al 2000) oras a consequence of Alzheimerrsquos disease (Yang et al2001) These papers underscore the significant contro-versy that exists regarding the function of cell cycle pro-teins such as PCNA in neurons after different types ofinjury We are currently conducting further studies whichwill elucidate the roles for PCNA and survivin in neu-rons after TBI
It should be noted that dual staining of survivin andPCNA could not be directly attributed to a specific celltype due to the technical difficulties of triple labeling an-tibody-based IHC Therefore we cannot rule out the pos-sibility that other cell types such as endothelial (Conwayet al 2003) or inflammatory cells (Hill-Felberg et al1999) may also contribute to survivin and PCNA ex-pression after TBI The appearance of survivin and PCNAseparately in neurons (NeuN-positive) and astrocytes(GFAP-positive) along with co-localization of survivinwith PCNA in the same cells provide correlative data tosuggest an activation of cell cycle-like program in astro-cytes and possibly in a small subtype of neurons after
JOHNSON ET AL
1192
TBI In our experiments survivin co-localization withPCNA does suggest that survivin may be associated witha pro-mitotic process In an attempt to clarify these pro-teinrsquos roles after TBI we analyzed the nuclear morphol-ogy of survivin-positive cells to define the apoptotic ormitotic architecture of nuclei DAPI staining proved tooambiguous in identifying apoptotic versus mitotic phe-notypes likely due to the thickness of the brain sections(40 m) Further studies using direct markers of mitosissuch as BrdU incorporation as well as simultaneous la-beling with cell death related proteins is required to de-lineate anti-apoptotic and pro-mitotic activities of sur-vivin and PCNA in these cells
In conclusion our data demonstrate the induction ofsurvivin in the rat brain following TBI Expression of sur-vivin occurred predominantly in astrocytes as comparedto neurons in a time-dependant fashion and was accom-panied by expression of PCNA Taken together these re-sults suggest that survivin plays a role in neural cell re-sponses following traumatic brain injury in rats Futurestudies will investigate the implications of these findingsto the pathophysiology of TBI
ACKNOWLEDGMENTS
We thank Barbara OrsquoSteen and Tao Fan MD fortechnical assistance This work was funded by grants NIHRO1 NS39091 NIH RO1 NS40182 DAMD 17-99-1-9565 and DAMD 17-01-1-0765
REFERENCES
ADIDA C CROTTY PL MCGRATH J BERREBI DDIEBOLD J and ALTIERI DC (1998) Developmentallyregulated expression of the novel cancer anti-apoptosis genesurvivin in human and mouse differentiation Am J Pathol152 43ndash49
ALTIERI DC MARCHISIO PC and MARCHISIO C(1999) Survivin apoptosis an interloper between cell deathand cell proliferation in cancer Lab Invest 79 1327ndash1333
ALTSCHUL SF MADDEN TL SCHAFFER AA et al(1997) Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 253389ndash3402
AMBROSINI G ADIDA C and ALTIERI DC (1997) Anovel anti-apoptosis gene survivin expressed in cancer andlymphoma Nat Med 3 917ndash921
BRAVO R FRANK R BLUNDELL PA and MAC-DONALD-BRAVO H (1987) CyclinPCNA is the auxil-iary protein of DNA polymerase-delta Nature 326 515ndash517
BUSH TG PUVANACHANDRA N HORNER CH et al
(1999) Leukocyte infiltration neuronal degeneration andneurite outgrowth after ablation of scar-forming reactive as-trocytes in adult transgenic mice Neuron 23 297ndash308
CAMERON HA and MCKAY RD (2001) Adult neuroge-nesis produces a large pool of new granule cells in the den-tate gyrus J Comp Neurol 435 406ndash417
CARBONELL WS and GRADY MS (1999) Regional andtemporal characterization of neuronal glial and axonal re-sponse after traumatic brain injury in the mouse Acta Neu-ropathol (Berl) 98 396ndash406
CHEN XH IWATA A NONAKA M BROWNE KDand SMITH DH (2003) Neurogenesis and glial prolifera-tion persist for at least one year in the subventricular zonefollowing brain trauma in rats J Neurotrauma 20 623ndash631
CHEN Y and SWANSON RA (2003) Astrocytes and braininjury J Cereb Blood Flow Metab 23 137ndash149
CONWAY EM POLLEFEYT S CORNELISSEN J et al(2000) Three differentially expressed survivin cDNA vari-ants encode proteins with distinct antiapoptotic functionsBlood 95 1435ndash1442
CSUKA E HANS VH AMMANN E TRENTZ OKOSSMANN T and MORGANTI-KOSSMANN MC(2000) Cell activation and inflammatory response followingtraumatic axonal injury in the rat Neuroreport 112587ndash2590
DEVERAUX QL and REED JC (1999) IAP family pro-teinsmdashsuppressors of apoptosis Genes Dev 13 239ndash252
DIXON CE CLIFTON GL LIGHTHALL JW YAGH-MAI AA and HAYES RL (1991) A controlled corticalimpact model of traumatic brain injury in the rat J NeurosciMethods 39 253ndash262
DOETSCH F CAILLE I LIM DA GARCIA-VERDUGOJM and ALVAREZ-BUYLLA A (1999) Subventricularzone astrocytes are neural stem cells in the adult mammalianbrain Cell 97 703ndash716
DUNN-MEYNELL AA and LEVIN BE (1997) Histolog-ical markers of neuronal axonal and astrocytic changes af-ter lateral rigid impact traumatic brain injury Brain Res 76125ndash41
GIULIAN D (1991) Microgliamdashneuron interactions after in-jury to the central nervous system in Peripheral Signalingof the Brain Role in Neural-Immune Interactions Learningand Memory R Frederickson J McGaugh and D Felton(eds) Lewiston NY Hogrefe and Huber pps 73ndash82
HIGGINS DG BLEASBY AJ and FUCHS R (1992)CLUSTAL V improved software for multiple sequencealignment Comput Appl Biosci 8 189ndash191
HILL-FELBERG SJ MCINTOSH TK OLIVER DLRAGHUPATHI R and BARBARESE E (1999) Concur-rent loss and proliferation of astrocytes following lateral fluidpercussion brain injury in the adult rat J Neurosci Res 57271ndash279
1193
SURVIVIN UPREGULATION AFTER TBI
JIANG X WILFORD C DUENSING S MUNGER KJONES G and JONES D (2001) Participation of Survivinin mitotic and apoptotic activities of normal and tumor-de-rived cells J Cell Biochem 83 342ndash354
KAYA SS MAHMOOD A LI Y YAVUZ E andCHOPP M (1999a) Expression of cell cycle proteins (cy-clin D1 and cdk4) after controlled cortical impact in rat brainJ Neurotrauma 16 1187ndash1196
KAYA SS MAHMOOD A LI Y YAVUZ E GOKSELM and CHOPP M (1999b) Apoptosis and expression ofp53 response proteins and cyclin D1 after cortical impact inrat brain Brain Res 818 23ndash33
KERNIE SG ERWIN TM and PARADA LF (2001)Brain remodeling due to neuronal and astrocytic prolifera-tion after controlled cortical injury in mice J Neurosci Res66 317ndash326
KOBAYASHI K HATANO M OTAKI M OGA-SAWARA T and TOKUHISA T (1999) Expression of amurine homologue of the inhibitor of apoptosis protein is re-lated to cell proliferation Proc Natl Acad Sci USA 961457ndash1462
LACASSE EC BAIRD S KORNELUK RG andMACKENZIE AE (1998) The inhibitors of apoptosis(IAPs) and their emerging role in cancer Oncogene 173247ndash3259
LATOV N NILAVER G ZIMMERMAN EA et al(1979) Fibrillary astrocytes proliferate in response to braininjury a study combining immunoperoxidase technique forglial fibrillary acidic protein and radioautography of tritiatedthymidine Dev Biol 72 381ndash384
LI F and ALTIERI DC (1999) The cancer antiapoptosismouse survivin gene characterization of locus and tran-scriptional requirements of basal and cell cycle-dependentexpression Cancer Res 59 3143ndash3151
LI F AMBROSINI G CHU EY et al (1998) Control ofapoptosis and mitotic spindle checkpoint by survivin Nature396 580ndash584
LI F ACKERMANN EJ BENNETT CF et al (1999)Pleiotropic cell-division defects and apoptosis induced by in-terference with survivin function Nat Cell Biol 1 461ndash466
MAGAVI SS LEAVITT BR and MACKLIS JD (2000)Induction of neurogenesis in the neocortex of adult mice Na-ture 405 951ndash955
MIYAKE T OKADA M and KITAMURA T (1992) Re-active proliferation of astrocytes studied by immunohisto-chemistry for proliferating cell nuclear antigen Brain Res590 300ndash302
MUCHMORE SW CHEN J JAKOB C et al (2000)Crystal structure and mutagenic analysis of the inhibitor-of-apoptosis protein survivin Mol Cell 6 173ndash182
NORTON WT (1999) Cell reactions following acute braininjury a review Neurochem Res 24 213ndash218
OTAKI M HATANO M KOBAYASHI K et al (2000)Cell cyclendashdependent regulation of TIAPm-survivin expres-sion Biochim Biophys Acta 1493 188ndash194
PETERSON DA (2002) Stem cells in brain plasticity and re-pair Curr Opin Pharmacol 2 34ndash42
PIKE BR ZHAO X NEWCOMB JK POSMANTURRM WANG KK and Hayes RL (1998) Regional cal-pain and caspase-3 proteolysis of alpha-spectrin after trau-matic brain injury Neuroreport 9 2437ndash2442
RAGHUPATHI R GRAHAM DI MCINTOSH TK(2000) Apoptosis after traumatic brain injury J Neuro-trauma 17 927ndash938
RIDET JL MALHOTRA SK PRIVAT A and GAGEFH (1997) Reactive astrocytes cellular and molecular cuesto biological function Trends Neurosci 20 570ndash577
SASAKI T LOPES MB HANKINS GR and HELMGA (2002) Expression of survivin an inhibitor of apopto-sis protein in tumors of the nervous system Acta Neu-ropathol (Berl) 104 105ndash109
SHANKAR SL MANI S OrsquoGUIN KN KANDIMALLAER AGRAWAL S and SHAFIT-ZAGARDO B (2001)Survivin inhibition induces human neural tumor cell deaththrough caspase-independent and -dependent pathways JNeurochem 79 426ndash436
SHIN S SUNG BJ CHO YS et al (2001) An anti-apop-totic protein human survivin is a direct inhibitor of caspase-3 and -7 Biochemistry 40 1117ndash1123
SMITH C BERRY M CLARKE WE and LOGAN A(2001) Differential expression of fibroblast growth factorndash2and fibroblast growth factor receptor 1 in a scarring and non-scarring model of CNS injury in the rat Eur J Neurosci 13443ndash456
STONE JR OKONKWO DO SINGLETON RHMUTLU LK HELM GA and POVLISHOCK JT(2002) Caspase-3ndashmediated cleavage of amyloid precursorprotein and formation of amyloid Beta peptide in traumaticaxonal injury J Neurotrauma 19 601ndash614
SYKOVA E VARGOVA L PROKOPOVA S and SI-MONOVA Z (1999) Glial swelling and astrogliosis pro-duce diffusion barriers in the rat spinal cord Glia 25 56ndash70
TAKAHASHI R DEVERAUX Q TAMM I et al (1998)A single BIR domain of XIAP sufficient for inhibiting cas-pases J Biol Chem 273 7787ndash7790
TAMM I WANG Y SAUSVILLE E et al (1998) IAP-family protein survivin inhibits caspase activity and apopto-sis induced by Fas (CD95) Bax caspases and anticancerdrugs Cancer Res 58 5315ndash5320
THOMPSON JD HIGGINS DG and GIBSON TJ (1994)CLUSTAL W improving the sensitivity of progressive mul-tiple sequence alignment through sequence weighting posi-tion-specific gap penalties and weight matrix choice NucleicAcids Res 22 4673ndash4680
JOHNSON ET AL
1194
TOLENTINO PJ DEFORD SM NOTTERPEK L et al(2002) Up-regulation of tissue-type transglutaminase aftertraumatic brain injury J Neurochem 80 579ndash588
UREN AG WONG L PAKUSCH M et al (2000) Sur-vivin and the inner centromere protein INCENP show simi-lar cell-cycle localization and gene knockout phenotypeCurr Biol 10 1319ndash1328
WEI LH HUANG CY CHENG SP CHEN CA andHSIEH CY (2001) Carcinosarcoma of ovary associatedwith previous radiotherapy Int J Gynecol Cancer 11 81ndash84
WU Q COMBS C CANNADY SB GELDMACHERDS and HERRUP K (2000) Beta-amyloid activated mi-croglia induce cell cycling and cell death in cultured corticalneurons Neurobiol Aging 21 797ndash806
YAGITA Y KITAGAWA K OHTSUKI T et al (2001)
Neurogenesis by progenitor cells in the ischemic adult rathippocampus Stroke 32 1890ndash1896
YANG Y GELDMACHER DS and HERRUP K (2001)DNA replication precedes neuronal cell death in Alzheimerrsquosdisease J Neurosci 21 2661ndash2668
Address reprint requests toJose A Pineda PhD
EF and WL McKnight Brain InstituteUniversity of Florida
100 S Newell Dr Bldg 59L1-118 Dept of Pediatrics
Gainesville FL 32610
E-mail pinedjapedsufledu
SURVIVIN UPREGULATION AFTER TBI
1195

gen Total RNA was isolated from the samples using TRI-zol reagent (Invitrogen Carlsbad CA) according to themanufacturerrsquos instructions cDNA synthesis was per-formed using 1 g of total RNA with the SuperScripttradeFirst-Strand Synthesis System for RT-PCR kit (Invitro-genLife Technologies Carlsbad CA) according to the
manufacturerrsquos instructions Q-PCR was performed aspreviously described (Tolentino et al 2002) using theLightCycler-FastStart DNA Master SYBR Green I reac-tion mix (Roche Diagnostics Indianapolis IN) in com-bination with 05 M primers 25 mM MgCl2 in theLight Cycler rapid thermal cycler system (Roche Diag-
JOHNSON ET AL
1186
FIG 3 Expression of PCNA after TBI in rats PVDF membranes visualized for survivin were stripped and re-probed withPCNA antibody Representative western blots showing PCNA (36 kDa) (A) and densitometry analysis of PCNA-positive bands(BC) are presented Experimental conditions sample size and abbreviations are identical to those in Figure 2 p 005 p 001 versus craniotomy control (one-way ANOVA test with post hoc Bonferroni analysis)
SURVIVIN UPREGULATION AFTER TBI
1187
FIG 4 Immunohistochemistry of survivin and PCNA Double fluorescent immunostaining for survivin (red) and PCNA (green)was performed in the ipsilateral cortex (A) and hippocampus (B) at 5 day post-injury Survivin is expressed in the cytoplasm (Cred) while PCNA is expressed in the nucleus (D green) The white arrow indicates the typical focal co-expression of survivinand PCNA as shown in merged survivin and PCNA images (E) PCNA expression was co-incident with DAPI staining (F bluewhite arrow) Original magnification 200 bar 50 m (AB) original magnification 400 bar 20 m (CndashF)
nostics Indianapolis IN) Briefly the primers were ampli-fied then quantified by online monitoring and identificationof the crossing point value (CPV) which is the exact timepoint at which the logarithmic linear phase could be dis-tinguished from the background The survivin primer setswere subjected to serial dilution and linear regression analy-sis of the logarithm of the dilution factor versus the CPVgenerated a standard curve for each transcript-speci-fic tem-plate Results are presented as percentage of cranio-tomycontrol Data were analyzed by ANOVA with a post-hocBonferroni-test and are given as mean SEM Differenceswere considered significant at the level of p 005
Western Blot Analyses
Brain tissue was removed as described above rinsedwith cold PBS snap frozen in liquid nitrogen and ho-
mogenized in ice-cold triple detergent lysis buffer con-taining a Completetrade protease inhibitor cocktail (RocheBiochemicals Indianapolis IN) Protein concentrationwas determined by bicinchoninic acid (BCA) micro pro-tein assays (Pierce Inc Rockford IL) Forty microgramsof protein per well was loaded and separated by SDS-PAGE transferred to PVDF membranes and probed withgoat-anti-rabbit survivin antibody (Novus BiologicalsLittleton CO 11000) After incubation with goat anti-rabbit HRP-labeled secondary antibody (Biorad Her-cules CA) the membranes were developed using En-hanced Chemiluminescence Plus reagents (ECL PlusAmersham Arlington Heights IL) For further PCNAanalysis developed PVDF membranes were incubated instripping buffer rinsed twice in TBST and incubated withPCNA antibody (Santa Cruz Biotech Santa Cruz CA
11000) as described above Semi-quantitative densito-metric analysis was performed using the AlphaImagertrade2000 Digital Imaging System (San Leandro CA) Trans-formed data (experimental densitometry value crani-otomy control densitometry value 100) was evaluatedby ANOVA and a post-hoc Bonferroni-test Values areexpressed as percentage of craniotomy controls and aregiven as mean SEM Differences were considered sig-nificant at the level of p 005
Characterization of Survivin Antibody (R51)
First we compared the specificity of the survivin an-tibody developed within our group (R51 Dr G Shaw)and a commercially available survivin antibody (Chemi-con Temecula CA) Our antibody showed characteris-tic staining of the cleavage furrow between dividingHeLa cells consistent with other reports (Li et al 19981999 Uren et al 2000) In addition double labeling withboth survivin antibodies showed co-localization at thecleavage furrow The peptides used to develop our sur-vivin antibody are specific to survivin and do not recog-nize other IAP family proteins according to SDSC Biol-ogy Workbench BLASTP (222) (Altschul et al 1997)and CLUSTAL W (181) analysis (Higgins et al 1992Thompson et al 1994) resulting in the survivin anti-bodyrsquos specificity
Immunohistochemistry (IHC)
Animals were transcardially perfused with 2 Heparin(Elkins-Sinn Inc Cherry Hill NJ) in 09 saline solu-tion (pH 74) followed by 4 paraformaldehyde in 01M phosphate buffer (pH 74) The brains were post-fixedin 4 paraformaldehyde and stored in 01M PBS or cryo-buffer Forty micron sections were fluorescent immuno-labeled with two primary antibodies in the following experiments survivin (1500)GFAP for astrocytes(Sternberger Lutherville MD 11000) survivinNeuNfor mature neurons (Chemicon Temecula CA 11000)survivinPCNA (Santa Cruz Biotech Santa Cruz CA1200) PCNAGFAP and PCNANeuN The nuclear dyeDAPI (in Vectashield H-1200 Vector LaboratoriesBurlingame CA) was used to label the nuclei The firstprimary antibody was incubated at 4degC for 24ndash48 h in a2 goat serum2 horse serum02 Triton-X 100 in01 M PBS (block) solution followed by the second pri-mary antibody at 4degC for 1 h in block solution Fluores-cent-tagged secondary antibody (Molecular Probes Eu-gene OR) was used for visualization For double-labelingusing same species antibodies we used the tyramide sig-nal amplification (TSA) kit (PerkinElmer Life SciencesBoston MA) according to the manufacturerrsquos instruc-tions and as previously described (Stone et al 2002) The
sections were viewed and digitally captured with a ZeissAxioplan 2 microscope (Zeiss Thornwood NY)equipped with a SPOT Real Time Slider high-resolutioncolor CCD digital camera (Diagnostic Instruments IncSterling Heights MI) The number of animals used fordual-labeling IHC is as follows survivin PCNA 4survivin GFAP 6 survivin NeuN 4 PCNA GFAP 4 and PCNA NeuN 4 Cell counts wereobtained by comparing the number of double-labeledcells to total single-labeled cells in the following groupssurvivinNeuN positive cells to total NeuN positive cellssurvivinPCNA positive cells to total PCNA positivecells PCNANeuN positive cells to total NeuN positivecells and survivinGFAP positive cells to total GFAPpositive cells Percentages were calculated by dividingthe number of double-labeled cells with the total numberof single-labeled cells For each group representativephotomicrographs were selected and counted Cells werecounted in a total area of at least 188000 m2 for eachgroup with no distinction made between cortical and hip-pocampal regions
RESULTS
Induction of Survivin Expression after TBI
Q-PCR analysis revealed an initial increase in survivinmRNA at 2 days post injury in the ipsilateral cortex andhippocampus These transcripts remained elevated inboth regions reached maximum levels at day 5 post-in-jury and declined at 7 days in the cortex and at 14 daysin the hippocampus All experimental animals remainedalive and exhibited slightly impaired motor and cogni-tive impairments (data not shown) Cortical mRNA lev-els reached a maximum of 448 100 whereas hip-pocampal mRNAs attained 606 100 compared tocraniotomy control values (Fig 1) To determine if theinduction of survivin mRNA resulted in correspondingincreases in survivin protein western blot analysis wasperformed Survivin (17-kDa protein) was readily de-tectable in the ipsilateral cortex and hippocampus of TBIrats while it was negligible in contralateral cortex andhippocampus (Fig 2A) Survivin was expressed in atime-dependent manner with a maximum increase at 5days after injury followed by a gradual decline by 14days Specifically the levels of survivin in cortical tis-sue were 616 257 at 3 days and 839 339 at 5days compared to craniotomy controls (Fig 2B) Similarincreases of survivin protein in the ipsilateral hippocam-pus were detected at 3 d and 5 days post injury 464 196 and 545 102 compared to craniotomy controlrespectively (Fig 2C)
JOHNSON ET AL
1188
FIG 5 Co-localization of survivin and GFAP in brain tissue after TBI Fluorescent immunohistochemistry for survivin (green)and GFAP (red) was performed in the ipsilateral and contralateral cortex (AB) and in the CA1 and dentate gyrus regions of thehippocampus (GH) at 5 day post-injury The injury has completely destroyed the cortex in G leaving only the hippocampus inthis picture Survivin was expressed in the cytoplasm (DJ green) of GFAP-positive astrocytes (CI red) of the ipsilateral cor-tex and hippocampus and was found to co-localize to these cells as shown in merged CD and IJ images (EK respectively yel-low) White arrows indicate typical survivin-positive astrocytes Nuclei are shown using DAPI (FL blue) Original magnifica-tion 100 bar 50 m (ABGH) 400 bar 20 m (CndashF and IndashL)
FIG 6 Co-localization of PCNA and GFAP in brain tissue after TBI Double immunostaining for PCNA (green) and GFAP(red) was performed in the ipsilateral and contralateral cortex (AB) and the CA1 and dentate gyrus regions of the hippocampus(GH) at 5 day post-injury PCNA is present in GFAP-positive cells of ipsilateral cortex (CD) and to a lesser extent hippocampus(IJ) (EK) Merged CD and IJ respectively White arrows indicate typical PCNA-positive astrocytes PCNA expression wasco-incident with DAPI staining (FL blue) Original magnification 100 bar 50 m (ABGH) 400 bar 20 m (CndashFand IndashL)
PCNA Expression after TBI
For detection of proliferating cell nuclear antigen(PCNA) PVDF membranes immunostained for survivinwere stripped and re-probed using a PCNA-specific an-tibody PCNA (36-kDa protein) was significantly de-tectable in the ipsilateral cortex and hippocampus of TBIrats but only negligible amounts were observed in thecontralateral cortex and hippocampus (Fig 3A) Thetemporal patterns exhibited by PCNA protein were sim-ilar to that of survivin protein Namely PCNA expressedin a time-dependant fashion with a maximum increase at5 days after injury followed by a gradual decline by 14days The levels of PCNA in ipsilateral cortical tissuewere raised over craniotomy control by 919 459 at3 days 2263 333 at 5 days and 1035 356 at 7days post injury (Fig 3B) Similar increases of PCNAprotein in ipsilateral hippocampus were detected at 5 dayspost injury with a maximum of 1006 229 comparedto craniotomy controls (Fig 3C) No significant increasewas found in the contralateral regions when compared tocraniotomy controls (Fig 3A)
Co-Expression of Survivin and PCNA following TBI
To examine spatial co-localization of survivin andPCNA double-label immunohistochemistry of brain tis-sue sections was performed on day 5 post injury whenpeak expression of these proteins was observed
At this time point a remarkable survivin and PCNAimmunoreactivity was found in the ipsilateral cortex (Fig4A) and ipsilateral hippocampus (Fig 4B) consistent withdata obtained using Western blot analyses Within bothregions focal co-expression patterns of survivin andPCNA in single cells were detected which was demon-strated by both separate fluorescent visualization of in-dividual proteins and by merging the images of double-stained slides (Fig 4CndashE) However the dual expressionof survivin and PCNA occurred infrequently as survivinand PCNA immunoreactivity could readily be found sep-arately (Fig 4CndashE) Approximately 12 of the total num-ber of PCNA-positive cells also labeled with survivin Thenuclear morphology of dual survivin and PCNA-positivecells was ambiguous as indicated by DAPI staining (Fig4F) Therefore DAPI staining was simply used for cellidentification in all subsequent experiments
Survivin and PCNA Are Expressed in Astrocytes after TBI
To determine the cell types expressing survivin andPCNA double-label immunohistochemistry for theseproteins and GFAP a marker of astrocytes was per-formed on day 5 post injury
In accordance with Western blot data remarkable sur-vivin-positive immunoreactivity was observed in the ip-silateral cortex and hippocampus proximal to the injurycavity (Fig 5AG green) but not in the contralateral ar-eas (Fig 5BH) Survivin was co-localized with GFAPin the cells of injured cortex and hippocampus whichstrongly suggested primary accumulation of survivin incells of astrocytic lineage (Fig 5CndashEIndashL) Survivin wasuniformly distributed in the cytoplasm and processes ofastrocytes in both cortex and hippocampus (Fig 5DJ)DAPI staining is shown in Figure 5FL3 Approximately88 of the total number of GFAP-positive cells also la-beled with survivin
PCNA-positive immunoreactivity staining was ob-served in the ipsilateral cortex (Fig 6A green) and hip-pocampus (Fig 6G green) of injured brain while con-tralateral cortex and hippocampus exhibited negligiblePCNA immunoreactivity (Fig 6BH) PCNA (Fig 6CI)was partially co-localized with GFAP (Fig 6DJ red) inboth regions and was characteristically distributed in thenucleus of the cells in both cortex and hippocampus (Fig6EK) DAPI staining is shown in Figure 6FL
Taken together double-label immunohistochemistrydata provides evidence that both survivin and PCNA canbe detected in GFAP-positive astrocytes following trau-matic insult Since survivin and PCNA immunoreactiv-ity was not exclusively localized in GFAP-positive cellswe next addressed a question what other cell type mightexpress survivin after TBI We suggested that a certainpopulation of mature neurons might express survivin inresponse to injury
Survivin and PCNA Are Expressed in a Sub-Set of Neurons after TBI
As can be seen in Figure 7 survivin and PCNA wereeach co-expressed with NeuN a marker of mature neu-rons NeuN-positive cells were found to express survivinin the ipsilateral cortex distal to the injury cavity (Fig7AndashD) and in the contralateral hippocampus (Fig 7EndashH)It should be noted however that NeuN-positive cells thatalso expressed survivin occurred infrequently For ex-ample we estimated the number of dual survivinNeuNpositive cells as 01ndash15 of the total number of NeuN-positive cells in these regions Survivin immunoreactiv-ity was negligible in either hemisphere of craniotomycontrol brains (Fig 7QR) No co-localization of survivinand NeuN was observed in ipsilateral hippocampus (datanot shown) As can be seen in Figure 7BF survivin waspredominantly localized to the cytoplasm and axons ofNeuN-positive neurons DAPI staining is shown in Fig-ure 7DH
PCNA-positive neurons were found in the ipsilateralcortex (Fig 7IndashL) and hippocampus after TBI (Fig
JOHNSON ET AL
1190
1191
FIG 7 A sub-set of NeuN-positive neurons express survivin and PCNA after TBI Double fluorescent immunohistochemistryfor survivin (green) and NeuN (red) in the ipsilateral cortex (AB) and the CA1 pyramidal layer of the contralateral hippocam-pus (EF) was performed Survivin is expressed in the cytoplasm and to a limited extent in the processes of NeuN-positive neu-rons (merged images CG) Dual staining for PCNA (green) and NeuN (red) is shown in the ipsilateral cortex (IJ) and the CA1pyramidal layer of the ipsilateral hippocampus (MN) The nuclei are shown using DAPI staining (DH blue) PCNA is expressedin the nucleus of NeuN-positive neurons (merged images KO) PCNA expression was co-incident with DAPI staining in theseexamples (LP blue) White arrows indicate focal co-localization of survivinNeuN and PCNANeuN SurvivinNeuN co-local-ization of survivin (green) and NeuN (red) was seen only in TBI rats as opposed to either hemisphere of craniotomy control(QR) Original magnification 400 bar 20 m (AndashP) original magnification 50 bar 100000 m (QR)
SURVIVIN UPREGULATION AFTER TBI
7MndashP) whereas craniotomy control tissue exhibited onlytrace amounts of PCNA (data not shown) Similar to thesurvivinNeuN co-localization data dual PCNANeuNimmunostaining was a rare event accounting for approx-imately 4 of the total number of NeuN positive cellsPCNA was distributed in the nuclei of these neurons (Fig7KO) although the nuclear morphology of these cellswas not clearly resolved by DAPI staining (Fig 7LP)
DISCUSSION
Traumatic brain injury (TBI) sets into motion variousbiochemical cascades that induce neural tissue injury andcell death Conversely several proteins expressed inneural cells after TBI are directed to resist cell death andpromote recovery in the injured CNS (Ridet et al 1997Chen and Swanson 2003) Survivin is a multi-functionalprotein that inhibits apoptosis and is also required for theproper completion of mitosis Anti-apoptotic and pro-mi-togenic roles for survivin have been documented in pro-liferating cells of neural origin in vitro such as in neu-roblastoma and glioma cells (LaCasse et al 1998 Tammet al 1998 Deveraux and Reed 1999 Conway et al2000 Shin et al 2001 Sasaki et al 2002) Howeverno studies have investigated the potential role of survivinin the adult brain after TBI when a sub-population ofCNS cells may initiate a cell cycle-related process in re-sponse to injury
In the present paper we demonstrate the induction ofsurvivin expression in rat brain subjected to TBI The ex-pression of survivin was time-dependent and cell-spe-cific and was present in astrocytes and to a much lesserextent in neurons in brain cortex and hippocampus In-duction of survivin in these cells was accompanied byoccasional expression of PCNA a cell cycle protein in-volved in mitotic G1S progression Our present data arethe first to show that survivin mRNA and protein are sig-nificantly up-regulated after traumatic brain injury in ratsPCNA expression after TBI has been described previously(Miyake et al 1992 Chen et al 2003) suggesting itsrole in mechanisms of brain recovery after injury Theconcurrent up-regulation of survivin with a similar tem-poral profile as PCNA shown herein further suggests thatsurvivin may play a role in cellular proliferation after TBI
Brain injury evoked the expression of survivin andPCNA in a time-dependent manner (Figs 2 and 3) West-ern blot analysis revealed maximal co-expression of bothsurvivin and PCNA at five days post injury Immuno-histochemistry at this time point demonstrated co-local-ization of these proteins (Fig 4) although most cells werelabeled separately with PCNA and survivin In fact only12 of the total number of PCNA-positive cells were
also survivin positive It has been reported that PCNA isexpressed predominantly in G1S (Bravo et al 1987)while survivin is found at the G2M phase of the cell cy-cle (Bravo et al 1987 Otaki et al 2000) Hence a lackof strict co-localization of survivin and PCNA in ourstudy may be explained by their expression at differentpoints in the cell cycle
In our experimental model we observed survivin- aswell as PCNA-positive astrocytes in the proximal area ofthe injury and in the ipsilateral hippocampus Prolifera-tion of astrocytes is well documented after TBI as shownby cell labeling with BrdU as well as expression of PCNA(Latov et al 1979 Dunn-Meynell and Levin 1997 Car-bonell and Grady 1999 Norton 1999 Csuka et al 2000Kernie et al 2001 Chen et al 2003) Because survivinand PCNA were expressed in astrocytes following TBI(Figs 5 and 6) it is possible that survivin plays an im-portant role linking astrocyte survival and proliferationafter traumatic insult Astrocyte proliferation has beenimplicated in the formation of the glial scar observed af-ter injury (Latov et al 1979) and creates a non-permis-sive environment for repair (Sykova et al 1999) How-ever glial proliferation may also enhance neuronalsurvival (Smith et al 2001 Wei et al 2001)
Of particular interest is a sub-set of NeuN-positive neu-rons found to express survivin only after TBI (Fig 7)These cells were much less abundant than survivin-pos-itive astrocytes and their functional significance is cur-rently unknown However both neurons and astrocyteshave been documented previously to express cell cycleproteins after various insults such as exposure to -amy-loidndashactivated microglia (Wu et al 2000) TBI (Kaya etal 1999ab) chlorine toxicity (Magavi et al 2000) oras a consequence of Alzheimerrsquos disease (Yang et al2001) These papers underscore the significant contro-versy that exists regarding the function of cell cycle pro-teins such as PCNA in neurons after different types ofinjury We are currently conducting further studies whichwill elucidate the roles for PCNA and survivin in neu-rons after TBI
It should be noted that dual staining of survivin andPCNA could not be directly attributed to a specific celltype due to the technical difficulties of triple labeling an-tibody-based IHC Therefore we cannot rule out the pos-sibility that other cell types such as endothelial (Conwayet al 2003) or inflammatory cells (Hill-Felberg et al1999) may also contribute to survivin and PCNA ex-pression after TBI The appearance of survivin and PCNAseparately in neurons (NeuN-positive) and astrocytes(GFAP-positive) along with co-localization of survivinwith PCNA in the same cells provide correlative data tosuggest an activation of cell cycle-like program in astro-cytes and possibly in a small subtype of neurons after
JOHNSON ET AL
1192
TBI In our experiments survivin co-localization withPCNA does suggest that survivin may be associated witha pro-mitotic process In an attempt to clarify these pro-teinrsquos roles after TBI we analyzed the nuclear morphol-ogy of survivin-positive cells to define the apoptotic ormitotic architecture of nuclei DAPI staining proved tooambiguous in identifying apoptotic versus mitotic phe-notypes likely due to the thickness of the brain sections(40 m) Further studies using direct markers of mitosissuch as BrdU incorporation as well as simultaneous la-beling with cell death related proteins is required to de-lineate anti-apoptotic and pro-mitotic activities of sur-vivin and PCNA in these cells
In conclusion our data demonstrate the induction ofsurvivin in the rat brain following TBI Expression of sur-vivin occurred predominantly in astrocytes as comparedto neurons in a time-dependant fashion and was accom-panied by expression of PCNA Taken together these re-sults suggest that survivin plays a role in neural cell re-sponses following traumatic brain injury in rats Futurestudies will investigate the implications of these findingsto the pathophysiology of TBI
ACKNOWLEDGMENTS
We thank Barbara OrsquoSteen and Tao Fan MD fortechnical assistance This work was funded by grants NIHRO1 NS39091 NIH RO1 NS40182 DAMD 17-99-1-9565 and DAMD 17-01-1-0765
REFERENCES
ADIDA C CROTTY PL MCGRATH J BERREBI DDIEBOLD J and ALTIERI DC (1998) Developmentallyregulated expression of the novel cancer anti-apoptosis genesurvivin in human and mouse differentiation Am J Pathol152 43ndash49
ALTIERI DC MARCHISIO PC and MARCHISIO C(1999) Survivin apoptosis an interloper between cell deathand cell proliferation in cancer Lab Invest 79 1327ndash1333
ALTSCHUL SF MADDEN TL SCHAFFER AA et al(1997) Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 253389ndash3402
AMBROSINI G ADIDA C and ALTIERI DC (1997) Anovel anti-apoptosis gene survivin expressed in cancer andlymphoma Nat Med 3 917ndash921
BRAVO R FRANK R BLUNDELL PA and MAC-DONALD-BRAVO H (1987) CyclinPCNA is the auxil-iary protein of DNA polymerase-delta Nature 326 515ndash517
BUSH TG PUVANACHANDRA N HORNER CH et al
(1999) Leukocyte infiltration neuronal degeneration andneurite outgrowth after ablation of scar-forming reactive as-trocytes in adult transgenic mice Neuron 23 297ndash308
CAMERON HA and MCKAY RD (2001) Adult neuroge-nesis produces a large pool of new granule cells in the den-tate gyrus J Comp Neurol 435 406ndash417
CARBONELL WS and GRADY MS (1999) Regional andtemporal characterization of neuronal glial and axonal re-sponse after traumatic brain injury in the mouse Acta Neu-ropathol (Berl) 98 396ndash406
CHEN XH IWATA A NONAKA M BROWNE KDand SMITH DH (2003) Neurogenesis and glial prolifera-tion persist for at least one year in the subventricular zonefollowing brain trauma in rats J Neurotrauma 20 623ndash631
CHEN Y and SWANSON RA (2003) Astrocytes and braininjury J Cereb Blood Flow Metab 23 137ndash149
CONWAY EM POLLEFEYT S CORNELISSEN J et al(2000) Three differentially expressed survivin cDNA vari-ants encode proteins with distinct antiapoptotic functionsBlood 95 1435ndash1442
CSUKA E HANS VH AMMANN E TRENTZ OKOSSMANN T and MORGANTI-KOSSMANN MC(2000) Cell activation and inflammatory response followingtraumatic axonal injury in the rat Neuroreport 112587ndash2590
DEVERAUX QL and REED JC (1999) IAP family pro-teinsmdashsuppressors of apoptosis Genes Dev 13 239ndash252
DIXON CE CLIFTON GL LIGHTHALL JW YAGH-MAI AA and HAYES RL (1991) A controlled corticalimpact model of traumatic brain injury in the rat J NeurosciMethods 39 253ndash262
DOETSCH F CAILLE I LIM DA GARCIA-VERDUGOJM and ALVAREZ-BUYLLA A (1999) Subventricularzone astrocytes are neural stem cells in the adult mammalianbrain Cell 97 703ndash716
DUNN-MEYNELL AA and LEVIN BE (1997) Histolog-ical markers of neuronal axonal and astrocytic changes af-ter lateral rigid impact traumatic brain injury Brain Res 76125ndash41
GIULIAN D (1991) Microgliamdashneuron interactions after in-jury to the central nervous system in Peripheral Signalingof the Brain Role in Neural-Immune Interactions Learningand Memory R Frederickson J McGaugh and D Felton(eds) Lewiston NY Hogrefe and Huber pps 73ndash82
HIGGINS DG BLEASBY AJ and FUCHS R (1992)CLUSTAL V improved software for multiple sequencealignment Comput Appl Biosci 8 189ndash191
HILL-FELBERG SJ MCINTOSH TK OLIVER DLRAGHUPATHI R and BARBARESE E (1999) Concur-rent loss and proliferation of astrocytes following lateral fluidpercussion brain injury in the adult rat J Neurosci Res 57271ndash279
1193
SURVIVIN UPREGULATION AFTER TBI
JIANG X WILFORD C DUENSING S MUNGER KJONES G and JONES D (2001) Participation of Survivinin mitotic and apoptotic activities of normal and tumor-de-rived cells J Cell Biochem 83 342ndash354
KAYA SS MAHMOOD A LI Y YAVUZ E andCHOPP M (1999a) Expression of cell cycle proteins (cy-clin D1 and cdk4) after controlled cortical impact in rat brainJ Neurotrauma 16 1187ndash1196
KAYA SS MAHMOOD A LI Y YAVUZ E GOKSELM and CHOPP M (1999b) Apoptosis and expression ofp53 response proteins and cyclin D1 after cortical impact inrat brain Brain Res 818 23ndash33
KERNIE SG ERWIN TM and PARADA LF (2001)Brain remodeling due to neuronal and astrocytic prolifera-tion after controlled cortical injury in mice J Neurosci Res66 317ndash326
KOBAYASHI K HATANO M OTAKI M OGA-SAWARA T and TOKUHISA T (1999) Expression of amurine homologue of the inhibitor of apoptosis protein is re-lated to cell proliferation Proc Natl Acad Sci USA 961457ndash1462
LACASSE EC BAIRD S KORNELUK RG andMACKENZIE AE (1998) The inhibitors of apoptosis(IAPs) and their emerging role in cancer Oncogene 173247ndash3259
LATOV N NILAVER G ZIMMERMAN EA et al(1979) Fibrillary astrocytes proliferate in response to braininjury a study combining immunoperoxidase technique forglial fibrillary acidic protein and radioautography of tritiatedthymidine Dev Biol 72 381ndash384
LI F and ALTIERI DC (1999) The cancer antiapoptosismouse survivin gene characterization of locus and tran-scriptional requirements of basal and cell cycle-dependentexpression Cancer Res 59 3143ndash3151
LI F AMBROSINI G CHU EY et al (1998) Control ofapoptosis and mitotic spindle checkpoint by survivin Nature396 580ndash584
LI F ACKERMANN EJ BENNETT CF et al (1999)Pleiotropic cell-division defects and apoptosis induced by in-terference with survivin function Nat Cell Biol 1 461ndash466
MAGAVI SS LEAVITT BR and MACKLIS JD (2000)Induction of neurogenesis in the neocortex of adult mice Na-ture 405 951ndash955
MIYAKE T OKADA M and KITAMURA T (1992) Re-active proliferation of astrocytes studied by immunohisto-chemistry for proliferating cell nuclear antigen Brain Res590 300ndash302
MUCHMORE SW CHEN J JAKOB C et al (2000)Crystal structure and mutagenic analysis of the inhibitor-of-apoptosis protein survivin Mol Cell 6 173ndash182
NORTON WT (1999) Cell reactions following acute braininjury a review Neurochem Res 24 213ndash218
OTAKI M HATANO M KOBAYASHI K et al (2000)Cell cyclendashdependent regulation of TIAPm-survivin expres-sion Biochim Biophys Acta 1493 188ndash194
PETERSON DA (2002) Stem cells in brain plasticity and re-pair Curr Opin Pharmacol 2 34ndash42
PIKE BR ZHAO X NEWCOMB JK POSMANTURRM WANG KK and Hayes RL (1998) Regional cal-pain and caspase-3 proteolysis of alpha-spectrin after trau-matic brain injury Neuroreport 9 2437ndash2442
RAGHUPATHI R GRAHAM DI MCINTOSH TK(2000) Apoptosis after traumatic brain injury J Neuro-trauma 17 927ndash938
RIDET JL MALHOTRA SK PRIVAT A and GAGEFH (1997) Reactive astrocytes cellular and molecular cuesto biological function Trends Neurosci 20 570ndash577
SASAKI T LOPES MB HANKINS GR and HELMGA (2002) Expression of survivin an inhibitor of apopto-sis protein in tumors of the nervous system Acta Neu-ropathol (Berl) 104 105ndash109
SHANKAR SL MANI S OrsquoGUIN KN KANDIMALLAER AGRAWAL S and SHAFIT-ZAGARDO B (2001)Survivin inhibition induces human neural tumor cell deaththrough caspase-independent and -dependent pathways JNeurochem 79 426ndash436
SHIN S SUNG BJ CHO YS et al (2001) An anti-apop-totic protein human survivin is a direct inhibitor of caspase-3 and -7 Biochemistry 40 1117ndash1123
SMITH C BERRY M CLARKE WE and LOGAN A(2001) Differential expression of fibroblast growth factorndash2and fibroblast growth factor receptor 1 in a scarring and non-scarring model of CNS injury in the rat Eur J Neurosci 13443ndash456
STONE JR OKONKWO DO SINGLETON RHMUTLU LK HELM GA and POVLISHOCK JT(2002) Caspase-3ndashmediated cleavage of amyloid precursorprotein and formation of amyloid Beta peptide in traumaticaxonal injury J Neurotrauma 19 601ndash614
SYKOVA E VARGOVA L PROKOPOVA S and SI-MONOVA Z (1999) Glial swelling and astrogliosis pro-duce diffusion barriers in the rat spinal cord Glia 25 56ndash70
TAKAHASHI R DEVERAUX Q TAMM I et al (1998)A single BIR domain of XIAP sufficient for inhibiting cas-pases J Biol Chem 273 7787ndash7790
TAMM I WANG Y SAUSVILLE E et al (1998) IAP-family protein survivin inhibits caspase activity and apopto-sis induced by Fas (CD95) Bax caspases and anticancerdrugs Cancer Res 58 5315ndash5320
THOMPSON JD HIGGINS DG and GIBSON TJ (1994)CLUSTAL W improving the sensitivity of progressive mul-tiple sequence alignment through sequence weighting posi-tion-specific gap penalties and weight matrix choice NucleicAcids Res 22 4673ndash4680
JOHNSON ET AL
1194
TOLENTINO PJ DEFORD SM NOTTERPEK L et al(2002) Up-regulation of tissue-type transglutaminase aftertraumatic brain injury J Neurochem 80 579ndash588
UREN AG WONG L PAKUSCH M et al (2000) Sur-vivin and the inner centromere protein INCENP show simi-lar cell-cycle localization and gene knockout phenotypeCurr Biol 10 1319ndash1328
WEI LH HUANG CY CHENG SP CHEN CA andHSIEH CY (2001) Carcinosarcoma of ovary associatedwith previous radiotherapy Int J Gynecol Cancer 11 81ndash84
WU Q COMBS C CANNADY SB GELDMACHERDS and HERRUP K (2000) Beta-amyloid activated mi-croglia induce cell cycling and cell death in cultured corticalneurons Neurobiol Aging 21 797ndash806
YAGITA Y KITAGAWA K OHTSUKI T et al (2001)
Neurogenesis by progenitor cells in the ischemic adult rathippocampus Stroke 32 1890ndash1896
YANG Y GELDMACHER DS and HERRUP K (2001)DNA replication precedes neuronal cell death in Alzheimerrsquosdisease J Neurosci 21 2661ndash2668
Address reprint requests toJose A Pineda PhD
EF and WL McKnight Brain InstituteUniversity of Florida
100 S Newell Dr Bldg 59L1-118 Dept of Pediatrics
Gainesville FL 32610
E-mail pinedjapedsufledu
SURVIVIN UPREGULATION AFTER TBI
1195
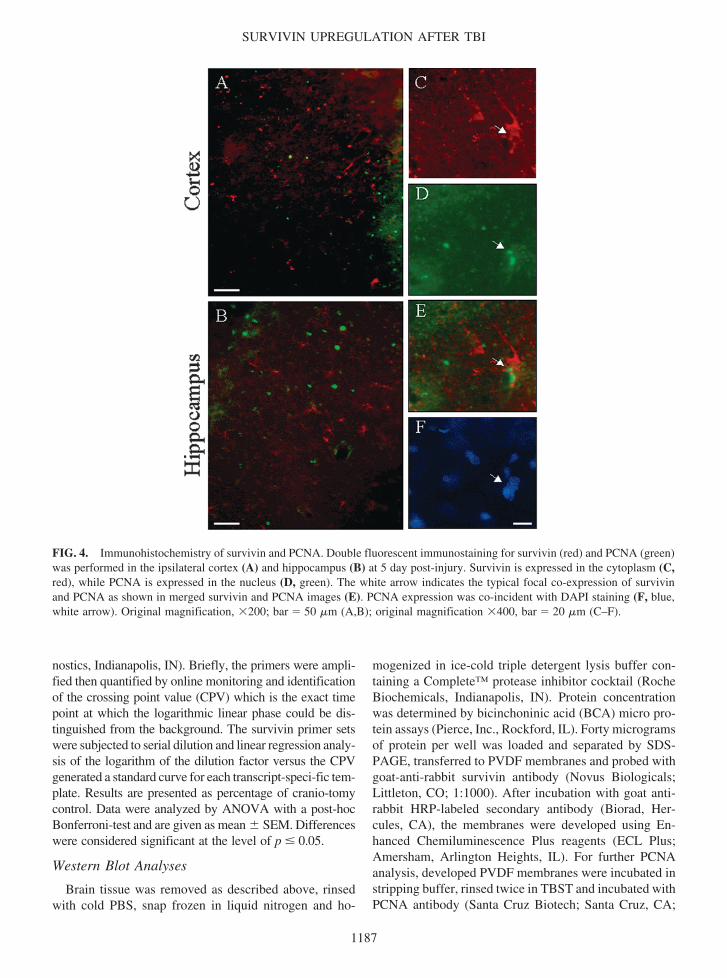
SURVIVIN UPREGULATION AFTER TBI
1187
FIG 4 Immunohistochemistry of survivin and PCNA Double fluorescent immunostaining for survivin (red) and PCNA (green)was performed in the ipsilateral cortex (A) and hippocampus (B) at 5 day post-injury Survivin is expressed in the cytoplasm (Cred) while PCNA is expressed in the nucleus (D green) The white arrow indicates the typical focal co-expression of survivinand PCNA as shown in merged survivin and PCNA images (E) PCNA expression was co-incident with DAPI staining (F bluewhite arrow) Original magnification 200 bar 50 m (AB) original magnification 400 bar 20 m (CndashF)
nostics Indianapolis IN) Briefly the primers were ampli-fied then quantified by online monitoring and identificationof the crossing point value (CPV) which is the exact timepoint at which the logarithmic linear phase could be dis-tinguished from the background The survivin primer setswere subjected to serial dilution and linear regression analy-sis of the logarithm of the dilution factor versus the CPVgenerated a standard curve for each transcript-speci-fic tem-plate Results are presented as percentage of cranio-tomycontrol Data were analyzed by ANOVA with a post-hocBonferroni-test and are given as mean SEM Differenceswere considered significant at the level of p 005
Western Blot Analyses
Brain tissue was removed as described above rinsedwith cold PBS snap frozen in liquid nitrogen and ho-
mogenized in ice-cold triple detergent lysis buffer con-taining a Completetrade protease inhibitor cocktail (RocheBiochemicals Indianapolis IN) Protein concentrationwas determined by bicinchoninic acid (BCA) micro pro-tein assays (Pierce Inc Rockford IL) Forty microgramsof protein per well was loaded and separated by SDS-PAGE transferred to PVDF membranes and probed withgoat-anti-rabbit survivin antibody (Novus BiologicalsLittleton CO 11000) After incubation with goat anti-rabbit HRP-labeled secondary antibody (Biorad Her-cules CA) the membranes were developed using En-hanced Chemiluminescence Plus reagents (ECL PlusAmersham Arlington Heights IL) For further PCNAanalysis developed PVDF membranes were incubated instripping buffer rinsed twice in TBST and incubated withPCNA antibody (Santa Cruz Biotech Santa Cruz CA
11000) as described above Semi-quantitative densito-metric analysis was performed using the AlphaImagertrade2000 Digital Imaging System (San Leandro CA) Trans-formed data (experimental densitometry value crani-otomy control densitometry value 100) was evaluatedby ANOVA and a post-hoc Bonferroni-test Values areexpressed as percentage of craniotomy controls and aregiven as mean SEM Differences were considered sig-nificant at the level of p 005
Characterization of Survivin Antibody (R51)
First we compared the specificity of the survivin an-tibody developed within our group (R51 Dr G Shaw)and a commercially available survivin antibody (Chemi-con Temecula CA) Our antibody showed characteris-tic staining of the cleavage furrow between dividingHeLa cells consistent with other reports (Li et al 19981999 Uren et al 2000) In addition double labeling withboth survivin antibodies showed co-localization at thecleavage furrow The peptides used to develop our sur-vivin antibody are specific to survivin and do not recog-nize other IAP family proteins according to SDSC Biol-ogy Workbench BLASTP (222) (Altschul et al 1997)and CLUSTAL W (181) analysis (Higgins et al 1992Thompson et al 1994) resulting in the survivin anti-bodyrsquos specificity
Immunohistochemistry (IHC)
Animals were transcardially perfused with 2 Heparin(Elkins-Sinn Inc Cherry Hill NJ) in 09 saline solu-tion (pH 74) followed by 4 paraformaldehyde in 01M phosphate buffer (pH 74) The brains were post-fixedin 4 paraformaldehyde and stored in 01M PBS or cryo-buffer Forty micron sections were fluorescent immuno-labeled with two primary antibodies in the following experiments survivin (1500)GFAP for astrocytes(Sternberger Lutherville MD 11000) survivinNeuNfor mature neurons (Chemicon Temecula CA 11000)survivinPCNA (Santa Cruz Biotech Santa Cruz CA1200) PCNAGFAP and PCNANeuN The nuclear dyeDAPI (in Vectashield H-1200 Vector LaboratoriesBurlingame CA) was used to label the nuclei The firstprimary antibody was incubated at 4degC for 24ndash48 h in a2 goat serum2 horse serum02 Triton-X 100 in01 M PBS (block) solution followed by the second pri-mary antibody at 4degC for 1 h in block solution Fluores-cent-tagged secondary antibody (Molecular Probes Eu-gene OR) was used for visualization For double-labelingusing same species antibodies we used the tyramide sig-nal amplification (TSA) kit (PerkinElmer Life SciencesBoston MA) according to the manufacturerrsquos instruc-tions and as previously described (Stone et al 2002) The
sections were viewed and digitally captured with a ZeissAxioplan 2 microscope (Zeiss Thornwood NY)equipped with a SPOT Real Time Slider high-resolutioncolor CCD digital camera (Diagnostic Instruments IncSterling Heights MI) The number of animals used fordual-labeling IHC is as follows survivin PCNA 4survivin GFAP 6 survivin NeuN 4 PCNA GFAP 4 and PCNA NeuN 4 Cell counts wereobtained by comparing the number of double-labeledcells to total single-labeled cells in the following groupssurvivinNeuN positive cells to total NeuN positive cellssurvivinPCNA positive cells to total PCNA positivecells PCNANeuN positive cells to total NeuN positivecells and survivinGFAP positive cells to total GFAPpositive cells Percentages were calculated by dividingthe number of double-labeled cells with the total numberof single-labeled cells For each group representativephotomicrographs were selected and counted Cells werecounted in a total area of at least 188000 m2 for eachgroup with no distinction made between cortical and hip-pocampal regions
RESULTS
Induction of Survivin Expression after TBI
Q-PCR analysis revealed an initial increase in survivinmRNA at 2 days post injury in the ipsilateral cortex andhippocampus These transcripts remained elevated inboth regions reached maximum levels at day 5 post-in-jury and declined at 7 days in the cortex and at 14 daysin the hippocampus All experimental animals remainedalive and exhibited slightly impaired motor and cogni-tive impairments (data not shown) Cortical mRNA lev-els reached a maximum of 448 100 whereas hip-pocampal mRNAs attained 606 100 compared tocraniotomy control values (Fig 1) To determine if theinduction of survivin mRNA resulted in correspondingincreases in survivin protein western blot analysis wasperformed Survivin (17-kDa protein) was readily de-tectable in the ipsilateral cortex and hippocampus of TBIrats while it was negligible in contralateral cortex andhippocampus (Fig 2A) Survivin was expressed in atime-dependent manner with a maximum increase at 5days after injury followed by a gradual decline by 14days Specifically the levels of survivin in cortical tis-sue were 616 257 at 3 days and 839 339 at 5days compared to craniotomy controls (Fig 2B) Similarincreases of survivin protein in the ipsilateral hippocam-pus were detected at 3 d and 5 days post injury 464 196 and 545 102 compared to craniotomy controlrespectively (Fig 2C)
JOHNSON ET AL
1188
FIG 5 Co-localization of survivin and GFAP in brain tissue after TBI Fluorescent immunohistochemistry for survivin (green)and GFAP (red) was performed in the ipsilateral and contralateral cortex (AB) and in the CA1 and dentate gyrus regions of thehippocampus (GH) at 5 day post-injury The injury has completely destroyed the cortex in G leaving only the hippocampus inthis picture Survivin was expressed in the cytoplasm (DJ green) of GFAP-positive astrocytes (CI red) of the ipsilateral cor-tex and hippocampus and was found to co-localize to these cells as shown in merged CD and IJ images (EK respectively yel-low) White arrows indicate typical survivin-positive astrocytes Nuclei are shown using DAPI (FL blue) Original magnifica-tion 100 bar 50 m (ABGH) 400 bar 20 m (CndashF and IndashL)
FIG 6 Co-localization of PCNA and GFAP in brain tissue after TBI Double immunostaining for PCNA (green) and GFAP(red) was performed in the ipsilateral and contralateral cortex (AB) and the CA1 and dentate gyrus regions of the hippocampus(GH) at 5 day post-injury PCNA is present in GFAP-positive cells of ipsilateral cortex (CD) and to a lesser extent hippocampus(IJ) (EK) Merged CD and IJ respectively White arrows indicate typical PCNA-positive astrocytes PCNA expression wasco-incident with DAPI staining (FL blue) Original magnification 100 bar 50 m (ABGH) 400 bar 20 m (CndashFand IndashL)
PCNA Expression after TBI
For detection of proliferating cell nuclear antigen(PCNA) PVDF membranes immunostained for survivinwere stripped and re-probed using a PCNA-specific an-tibody PCNA (36-kDa protein) was significantly de-tectable in the ipsilateral cortex and hippocampus of TBIrats but only negligible amounts were observed in thecontralateral cortex and hippocampus (Fig 3A) Thetemporal patterns exhibited by PCNA protein were sim-ilar to that of survivin protein Namely PCNA expressedin a time-dependant fashion with a maximum increase at5 days after injury followed by a gradual decline by 14days The levels of PCNA in ipsilateral cortical tissuewere raised over craniotomy control by 919 459 at3 days 2263 333 at 5 days and 1035 356 at 7days post injury (Fig 3B) Similar increases of PCNAprotein in ipsilateral hippocampus were detected at 5 dayspost injury with a maximum of 1006 229 comparedto craniotomy controls (Fig 3C) No significant increasewas found in the contralateral regions when compared tocraniotomy controls (Fig 3A)
Co-Expression of Survivin and PCNA following TBI
To examine spatial co-localization of survivin andPCNA double-label immunohistochemistry of brain tis-sue sections was performed on day 5 post injury whenpeak expression of these proteins was observed
At this time point a remarkable survivin and PCNAimmunoreactivity was found in the ipsilateral cortex (Fig4A) and ipsilateral hippocampus (Fig 4B) consistent withdata obtained using Western blot analyses Within bothregions focal co-expression patterns of survivin andPCNA in single cells were detected which was demon-strated by both separate fluorescent visualization of in-dividual proteins and by merging the images of double-stained slides (Fig 4CndashE) However the dual expressionof survivin and PCNA occurred infrequently as survivinand PCNA immunoreactivity could readily be found sep-arately (Fig 4CndashE) Approximately 12 of the total num-ber of PCNA-positive cells also labeled with survivin Thenuclear morphology of dual survivin and PCNA-positivecells was ambiguous as indicated by DAPI staining (Fig4F) Therefore DAPI staining was simply used for cellidentification in all subsequent experiments
Survivin and PCNA Are Expressed in Astrocytes after TBI
To determine the cell types expressing survivin andPCNA double-label immunohistochemistry for theseproteins and GFAP a marker of astrocytes was per-formed on day 5 post injury
In accordance with Western blot data remarkable sur-vivin-positive immunoreactivity was observed in the ip-silateral cortex and hippocampus proximal to the injurycavity (Fig 5AG green) but not in the contralateral ar-eas (Fig 5BH) Survivin was co-localized with GFAPin the cells of injured cortex and hippocampus whichstrongly suggested primary accumulation of survivin incells of astrocytic lineage (Fig 5CndashEIndashL) Survivin wasuniformly distributed in the cytoplasm and processes ofastrocytes in both cortex and hippocampus (Fig 5DJ)DAPI staining is shown in Figure 5FL3 Approximately88 of the total number of GFAP-positive cells also la-beled with survivin
PCNA-positive immunoreactivity staining was ob-served in the ipsilateral cortex (Fig 6A green) and hip-pocampus (Fig 6G green) of injured brain while con-tralateral cortex and hippocampus exhibited negligiblePCNA immunoreactivity (Fig 6BH) PCNA (Fig 6CI)was partially co-localized with GFAP (Fig 6DJ red) inboth regions and was characteristically distributed in thenucleus of the cells in both cortex and hippocampus (Fig6EK) DAPI staining is shown in Figure 6FL
Taken together double-label immunohistochemistrydata provides evidence that both survivin and PCNA canbe detected in GFAP-positive astrocytes following trau-matic insult Since survivin and PCNA immunoreactiv-ity was not exclusively localized in GFAP-positive cellswe next addressed a question what other cell type mightexpress survivin after TBI We suggested that a certainpopulation of mature neurons might express survivin inresponse to injury
Survivin and PCNA Are Expressed in a Sub-Set of Neurons after TBI
As can be seen in Figure 7 survivin and PCNA wereeach co-expressed with NeuN a marker of mature neu-rons NeuN-positive cells were found to express survivinin the ipsilateral cortex distal to the injury cavity (Fig7AndashD) and in the contralateral hippocampus (Fig 7EndashH)It should be noted however that NeuN-positive cells thatalso expressed survivin occurred infrequently For ex-ample we estimated the number of dual survivinNeuNpositive cells as 01ndash15 of the total number of NeuN-positive cells in these regions Survivin immunoreactiv-ity was negligible in either hemisphere of craniotomycontrol brains (Fig 7QR) No co-localization of survivinand NeuN was observed in ipsilateral hippocampus (datanot shown) As can be seen in Figure 7BF survivin waspredominantly localized to the cytoplasm and axons ofNeuN-positive neurons DAPI staining is shown in Fig-ure 7DH
PCNA-positive neurons were found in the ipsilateralcortex (Fig 7IndashL) and hippocampus after TBI (Fig
JOHNSON ET AL
1190
1191
FIG 7 A sub-set of NeuN-positive neurons express survivin and PCNA after TBI Double fluorescent immunohistochemistryfor survivin (green) and NeuN (red) in the ipsilateral cortex (AB) and the CA1 pyramidal layer of the contralateral hippocam-pus (EF) was performed Survivin is expressed in the cytoplasm and to a limited extent in the processes of NeuN-positive neu-rons (merged images CG) Dual staining for PCNA (green) and NeuN (red) is shown in the ipsilateral cortex (IJ) and the CA1pyramidal layer of the ipsilateral hippocampus (MN) The nuclei are shown using DAPI staining (DH blue) PCNA is expressedin the nucleus of NeuN-positive neurons (merged images KO) PCNA expression was co-incident with DAPI staining in theseexamples (LP blue) White arrows indicate focal co-localization of survivinNeuN and PCNANeuN SurvivinNeuN co-local-ization of survivin (green) and NeuN (red) was seen only in TBI rats as opposed to either hemisphere of craniotomy control(QR) Original magnification 400 bar 20 m (AndashP) original magnification 50 bar 100000 m (QR)
SURVIVIN UPREGULATION AFTER TBI
7MndashP) whereas craniotomy control tissue exhibited onlytrace amounts of PCNA (data not shown) Similar to thesurvivinNeuN co-localization data dual PCNANeuNimmunostaining was a rare event accounting for approx-imately 4 of the total number of NeuN positive cellsPCNA was distributed in the nuclei of these neurons (Fig7KO) although the nuclear morphology of these cellswas not clearly resolved by DAPI staining (Fig 7LP)
DISCUSSION
Traumatic brain injury (TBI) sets into motion variousbiochemical cascades that induce neural tissue injury andcell death Conversely several proteins expressed inneural cells after TBI are directed to resist cell death andpromote recovery in the injured CNS (Ridet et al 1997Chen and Swanson 2003) Survivin is a multi-functionalprotein that inhibits apoptosis and is also required for theproper completion of mitosis Anti-apoptotic and pro-mi-togenic roles for survivin have been documented in pro-liferating cells of neural origin in vitro such as in neu-roblastoma and glioma cells (LaCasse et al 1998 Tammet al 1998 Deveraux and Reed 1999 Conway et al2000 Shin et al 2001 Sasaki et al 2002) Howeverno studies have investigated the potential role of survivinin the adult brain after TBI when a sub-population ofCNS cells may initiate a cell cycle-related process in re-sponse to injury
In the present paper we demonstrate the induction ofsurvivin expression in rat brain subjected to TBI The ex-pression of survivin was time-dependent and cell-spe-cific and was present in astrocytes and to a much lesserextent in neurons in brain cortex and hippocampus In-duction of survivin in these cells was accompanied byoccasional expression of PCNA a cell cycle protein in-volved in mitotic G1S progression Our present data arethe first to show that survivin mRNA and protein are sig-nificantly up-regulated after traumatic brain injury in ratsPCNA expression after TBI has been described previously(Miyake et al 1992 Chen et al 2003) suggesting itsrole in mechanisms of brain recovery after injury Theconcurrent up-regulation of survivin with a similar tem-poral profile as PCNA shown herein further suggests thatsurvivin may play a role in cellular proliferation after TBI
Brain injury evoked the expression of survivin andPCNA in a time-dependent manner (Figs 2 and 3) West-ern blot analysis revealed maximal co-expression of bothsurvivin and PCNA at five days post injury Immuno-histochemistry at this time point demonstrated co-local-ization of these proteins (Fig 4) although most cells werelabeled separately with PCNA and survivin In fact only12 of the total number of PCNA-positive cells were
also survivin positive It has been reported that PCNA isexpressed predominantly in G1S (Bravo et al 1987)while survivin is found at the G2M phase of the cell cy-cle (Bravo et al 1987 Otaki et al 2000) Hence a lackof strict co-localization of survivin and PCNA in ourstudy may be explained by their expression at differentpoints in the cell cycle
In our experimental model we observed survivin- aswell as PCNA-positive astrocytes in the proximal area ofthe injury and in the ipsilateral hippocampus Prolifera-tion of astrocytes is well documented after TBI as shownby cell labeling with BrdU as well as expression of PCNA(Latov et al 1979 Dunn-Meynell and Levin 1997 Car-bonell and Grady 1999 Norton 1999 Csuka et al 2000Kernie et al 2001 Chen et al 2003) Because survivinand PCNA were expressed in astrocytes following TBI(Figs 5 and 6) it is possible that survivin plays an im-portant role linking astrocyte survival and proliferationafter traumatic insult Astrocyte proliferation has beenimplicated in the formation of the glial scar observed af-ter injury (Latov et al 1979) and creates a non-permis-sive environment for repair (Sykova et al 1999) How-ever glial proliferation may also enhance neuronalsurvival (Smith et al 2001 Wei et al 2001)
Of particular interest is a sub-set of NeuN-positive neu-rons found to express survivin only after TBI (Fig 7)These cells were much less abundant than survivin-pos-itive astrocytes and their functional significance is cur-rently unknown However both neurons and astrocyteshave been documented previously to express cell cycleproteins after various insults such as exposure to -amy-loidndashactivated microglia (Wu et al 2000) TBI (Kaya etal 1999ab) chlorine toxicity (Magavi et al 2000) oras a consequence of Alzheimerrsquos disease (Yang et al2001) These papers underscore the significant contro-versy that exists regarding the function of cell cycle pro-teins such as PCNA in neurons after different types ofinjury We are currently conducting further studies whichwill elucidate the roles for PCNA and survivin in neu-rons after TBI
It should be noted that dual staining of survivin andPCNA could not be directly attributed to a specific celltype due to the technical difficulties of triple labeling an-tibody-based IHC Therefore we cannot rule out the pos-sibility that other cell types such as endothelial (Conwayet al 2003) or inflammatory cells (Hill-Felberg et al1999) may also contribute to survivin and PCNA ex-pression after TBI The appearance of survivin and PCNAseparately in neurons (NeuN-positive) and astrocytes(GFAP-positive) along with co-localization of survivinwith PCNA in the same cells provide correlative data tosuggest an activation of cell cycle-like program in astro-cytes and possibly in a small subtype of neurons after
JOHNSON ET AL
1192
TBI In our experiments survivin co-localization withPCNA does suggest that survivin may be associated witha pro-mitotic process In an attempt to clarify these pro-teinrsquos roles after TBI we analyzed the nuclear morphol-ogy of survivin-positive cells to define the apoptotic ormitotic architecture of nuclei DAPI staining proved tooambiguous in identifying apoptotic versus mitotic phe-notypes likely due to the thickness of the brain sections(40 m) Further studies using direct markers of mitosissuch as BrdU incorporation as well as simultaneous la-beling with cell death related proteins is required to de-lineate anti-apoptotic and pro-mitotic activities of sur-vivin and PCNA in these cells
In conclusion our data demonstrate the induction ofsurvivin in the rat brain following TBI Expression of sur-vivin occurred predominantly in astrocytes as comparedto neurons in a time-dependant fashion and was accom-panied by expression of PCNA Taken together these re-sults suggest that survivin plays a role in neural cell re-sponses following traumatic brain injury in rats Futurestudies will investigate the implications of these findingsto the pathophysiology of TBI
ACKNOWLEDGMENTS
We thank Barbara OrsquoSteen and Tao Fan MD fortechnical assistance This work was funded by grants NIHRO1 NS39091 NIH RO1 NS40182 DAMD 17-99-1-9565 and DAMD 17-01-1-0765
REFERENCES
ADIDA C CROTTY PL MCGRATH J BERREBI DDIEBOLD J and ALTIERI DC (1998) Developmentallyregulated expression of the novel cancer anti-apoptosis genesurvivin in human and mouse differentiation Am J Pathol152 43ndash49
ALTIERI DC MARCHISIO PC and MARCHISIO C(1999) Survivin apoptosis an interloper between cell deathand cell proliferation in cancer Lab Invest 79 1327ndash1333
ALTSCHUL SF MADDEN TL SCHAFFER AA et al(1997) Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 253389ndash3402
AMBROSINI G ADIDA C and ALTIERI DC (1997) Anovel anti-apoptosis gene survivin expressed in cancer andlymphoma Nat Med 3 917ndash921
BRAVO R FRANK R BLUNDELL PA and MAC-DONALD-BRAVO H (1987) CyclinPCNA is the auxil-iary protein of DNA polymerase-delta Nature 326 515ndash517
BUSH TG PUVANACHANDRA N HORNER CH et al
(1999) Leukocyte infiltration neuronal degeneration andneurite outgrowth after ablation of scar-forming reactive as-trocytes in adult transgenic mice Neuron 23 297ndash308
CAMERON HA and MCKAY RD (2001) Adult neuroge-nesis produces a large pool of new granule cells in the den-tate gyrus J Comp Neurol 435 406ndash417
CARBONELL WS and GRADY MS (1999) Regional andtemporal characterization of neuronal glial and axonal re-sponse after traumatic brain injury in the mouse Acta Neu-ropathol (Berl) 98 396ndash406
CHEN XH IWATA A NONAKA M BROWNE KDand SMITH DH (2003) Neurogenesis and glial prolifera-tion persist for at least one year in the subventricular zonefollowing brain trauma in rats J Neurotrauma 20 623ndash631
CHEN Y and SWANSON RA (2003) Astrocytes and braininjury J Cereb Blood Flow Metab 23 137ndash149
CONWAY EM POLLEFEYT S CORNELISSEN J et al(2000) Three differentially expressed survivin cDNA vari-ants encode proteins with distinct antiapoptotic functionsBlood 95 1435ndash1442
CSUKA E HANS VH AMMANN E TRENTZ OKOSSMANN T and MORGANTI-KOSSMANN MC(2000) Cell activation and inflammatory response followingtraumatic axonal injury in the rat Neuroreport 112587ndash2590
DEVERAUX QL and REED JC (1999) IAP family pro-teinsmdashsuppressors of apoptosis Genes Dev 13 239ndash252
DIXON CE CLIFTON GL LIGHTHALL JW YAGH-MAI AA and HAYES RL (1991) A controlled corticalimpact model of traumatic brain injury in the rat J NeurosciMethods 39 253ndash262
DOETSCH F CAILLE I LIM DA GARCIA-VERDUGOJM and ALVAREZ-BUYLLA A (1999) Subventricularzone astrocytes are neural stem cells in the adult mammalianbrain Cell 97 703ndash716
DUNN-MEYNELL AA and LEVIN BE (1997) Histolog-ical markers of neuronal axonal and astrocytic changes af-ter lateral rigid impact traumatic brain injury Brain Res 76125ndash41
GIULIAN D (1991) Microgliamdashneuron interactions after in-jury to the central nervous system in Peripheral Signalingof the Brain Role in Neural-Immune Interactions Learningand Memory R Frederickson J McGaugh and D Felton(eds) Lewiston NY Hogrefe and Huber pps 73ndash82
HIGGINS DG BLEASBY AJ and FUCHS R (1992)CLUSTAL V improved software for multiple sequencealignment Comput Appl Biosci 8 189ndash191
HILL-FELBERG SJ MCINTOSH TK OLIVER DLRAGHUPATHI R and BARBARESE E (1999) Concur-rent loss and proliferation of astrocytes following lateral fluidpercussion brain injury in the adult rat J Neurosci Res 57271ndash279
1193
SURVIVIN UPREGULATION AFTER TBI
JIANG X WILFORD C DUENSING S MUNGER KJONES G and JONES D (2001) Participation of Survivinin mitotic and apoptotic activities of normal and tumor-de-rived cells J Cell Biochem 83 342ndash354
KAYA SS MAHMOOD A LI Y YAVUZ E andCHOPP M (1999a) Expression of cell cycle proteins (cy-clin D1 and cdk4) after controlled cortical impact in rat brainJ Neurotrauma 16 1187ndash1196
KAYA SS MAHMOOD A LI Y YAVUZ E GOKSELM and CHOPP M (1999b) Apoptosis and expression ofp53 response proteins and cyclin D1 after cortical impact inrat brain Brain Res 818 23ndash33
KERNIE SG ERWIN TM and PARADA LF (2001)Brain remodeling due to neuronal and astrocytic prolifera-tion after controlled cortical injury in mice J Neurosci Res66 317ndash326
KOBAYASHI K HATANO M OTAKI M OGA-SAWARA T and TOKUHISA T (1999) Expression of amurine homologue of the inhibitor of apoptosis protein is re-lated to cell proliferation Proc Natl Acad Sci USA 961457ndash1462
LACASSE EC BAIRD S KORNELUK RG andMACKENZIE AE (1998) The inhibitors of apoptosis(IAPs) and their emerging role in cancer Oncogene 173247ndash3259
LATOV N NILAVER G ZIMMERMAN EA et al(1979) Fibrillary astrocytes proliferate in response to braininjury a study combining immunoperoxidase technique forglial fibrillary acidic protein and radioautography of tritiatedthymidine Dev Biol 72 381ndash384
LI F and ALTIERI DC (1999) The cancer antiapoptosismouse survivin gene characterization of locus and tran-scriptional requirements of basal and cell cycle-dependentexpression Cancer Res 59 3143ndash3151
LI F AMBROSINI G CHU EY et al (1998) Control ofapoptosis and mitotic spindle checkpoint by survivin Nature396 580ndash584
LI F ACKERMANN EJ BENNETT CF et al (1999)Pleiotropic cell-division defects and apoptosis induced by in-terference with survivin function Nat Cell Biol 1 461ndash466
MAGAVI SS LEAVITT BR and MACKLIS JD (2000)Induction of neurogenesis in the neocortex of adult mice Na-ture 405 951ndash955
MIYAKE T OKADA M and KITAMURA T (1992) Re-active proliferation of astrocytes studied by immunohisto-chemistry for proliferating cell nuclear antigen Brain Res590 300ndash302
MUCHMORE SW CHEN J JAKOB C et al (2000)Crystal structure and mutagenic analysis of the inhibitor-of-apoptosis protein survivin Mol Cell 6 173ndash182
NORTON WT (1999) Cell reactions following acute braininjury a review Neurochem Res 24 213ndash218
OTAKI M HATANO M KOBAYASHI K et al (2000)Cell cyclendashdependent regulation of TIAPm-survivin expres-sion Biochim Biophys Acta 1493 188ndash194
PETERSON DA (2002) Stem cells in brain plasticity and re-pair Curr Opin Pharmacol 2 34ndash42
PIKE BR ZHAO X NEWCOMB JK POSMANTURRM WANG KK and Hayes RL (1998) Regional cal-pain and caspase-3 proteolysis of alpha-spectrin after trau-matic brain injury Neuroreport 9 2437ndash2442
RAGHUPATHI R GRAHAM DI MCINTOSH TK(2000) Apoptosis after traumatic brain injury J Neuro-trauma 17 927ndash938
RIDET JL MALHOTRA SK PRIVAT A and GAGEFH (1997) Reactive astrocytes cellular and molecular cuesto biological function Trends Neurosci 20 570ndash577
SASAKI T LOPES MB HANKINS GR and HELMGA (2002) Expression of survivin an inhibitor of apopto-sis protein in tumors of the nervous system Acta Neu-ropathol (Berl) 104 105ndash109
SHANKAR SL MANI S OrsquoGUIN KN KANDIMALLAER AGRAWAL S and SHAFIT-ZAGARDO B (2001)Survivin inhibition induces human neural tumor cell deaththrough caspase-independent and -dependent pathways JNeurochem 79 426ndash436
SHIN S SUNG BJ CHO YS et al (2001) An anti-apop-totic protein human survivin is a direct inhibitor of caspase-3 and -7 Biochemistry 40 1117ndash1123
SMITH C BERRY M CLARKE WE and LOGAN A(2001) Differential expression of fibroblast growth factorndash2and fibroblast growth factor receptor 1 in a scarring and non-scarring model of CNS injury in the rat Eur J Neurosci 13443ndash456
STONE JR OKONKWO DO SINGLETON RHMUTLU LK HELM GA and POVLISHOCK JT(2002) Caspase-3ndashmediated cleavage of amyloid precursorprotein and formation of amyloid Beta peptide in traumaticaxonal injury J Neurotrauma 19 601ndash614
SYKOVA E VARGOVA L PROKOPOVA S and SI-MONOVA Z (1999) Glial swelling and astrogliosis pro-duce diffusion barriers in the rat spinal cord Glia 25 56ndash70
TAKAHASHI R DEVERAUX Q TAMM I et al (1998)A single BIR domain of XIAP sufficient for inhibiting cas-pases J Biol Chem 273 7787ndash7790
TAMM I WANG Y SAUSVILLE E et al (1998) IAP-family protein survivin inhibits caspase activity and apopto-sis induced by Fas (CD95) Bax caspases and anticancerdrugs Cancer Res 58 5315ndash5320
THOMPSON JD HIGGINS DG and GIBSON TJ (1994)CLUSTAL W improving the sensitivity of progressive mul-tiple sequence alignment through sequence weighting posi-tion-specific gap penalties and weight matrix choice NucleicAcids Res 22 4673ndash4680
JOHNSON ET AL
1194
TOLENTINO PJ DEFORD SM NOTTERPEK L et al(2002) Up-regulation of tissue-type transglutaminase aftertraumatic brain injury J Neurochem 80 579ndash588
UREN AG WONG L PAKUSCH M et al (2000) Sur-vivin and the inner centromere protein INCENP show simi-lar cell-cycle localization and gene knockout phenotypeCurr Biol 10 1319ndash1328
WEI LH HUANG CY CHENG SP CHEN CA andHSIEH CY (2001) Carcinosarcoma of ovary associatedwith previous radiotherapy Int J Gynecol Cancer 11 81ndash84
WU Q COMBS C CANNADY SB GELDMACHERDS and HERRUP K (2000) Beta-amyloid activated mi-croglia induce cell cycling and cell death in cultured corticalneurons Neurobiol Aging 21 797ndash806
YAGITA Y KITAGAWA K OHTSUKI T et al (2001)
Neurogenesis by progenitor cells in the ischemic adult rathippocampus Stroke 32 1890ndash1896
YANG Y GELDMACHER DS and HERRUP K (2001)DNA replication precedes neuronal cell death in Alzheimerrsquosdisease J Neurosci 21 2661ndash2668
Address reprint requests toJose A Pineda PhD
EF and WL McKnight Brain InstituteUniversity of Florida
100 S Newell Dr Bldg 59L1-118 Dept of Pediatrics
Gainesville FL 32610
E-mail pinedjapedsufledu
SURVIVIN UPREGULATION AFTER TBI
1195

11000) as described above Semi-quantitative densito-metric analysis was performed using the AlphaImagertrade2000 Digital Imaging System (San Leandro CA) Trans-formed data (experimental densitometry value crani-otomy control densitometry value 100) was evaluatedby ANOVA and a post-hoc Bonferroni-test Values areexpressed as percentage of craniotomy controls and aregiven as mean SEM Differences were considered sig-nificant at the level of p 005
Characterization of Survivin Antibody (R51)
First we compared the specificity of the survivin an-tibody developed within our group (R51 Dr G Shaw)and a commercially available survivin antibody (Chemi-con Temecula CA) Our antibody showed characteris-tic staining of the cleavage furrow between dividingHeLa cells consistent with other reports (Li et al 19981999 Uren et al 2000) In addition double labeling withboth survivin antibodies showed co-localization at thecleavage furrow The peptides used to develop our sur-vivin antibody are specific to survivin and do not recog-nize other IAP family proteins according to SDSC Biol-ogy Workbench BLASTP (222) (Altschul et al 1997)and CLUSTAL W (181) analysis (Higgins et al 1992Thompson et al 1994) resulting in the survivin anti-bodyrsquos specificity
Immunohistochemistry (IHC)
Animals were transcardially perfused with 2 Heparin(Elkins-Sinn Inc Cherry Hill NJ) in 09 saline solu-tion (pH 74) followed by 4 paraformaldehyde in 01M phosphate buffer (pH 74) The brains were post-fixedin 4 paraformaldehyde and stored in 01M PBS or cryo-buffer Forty micron sections were fluorescent immuno-labeled with two primary antibodies in the following experiments survivin (1500)GFAP for astrocytes(Sternberger Lutherville MD 11000) survivinNeuNfor mature neurons (Chemicon Temecula CA 11000)survivinPCNA (Santa Cruz Biotech Santa Cruz CA1200) PCNAGFAP and PCNANeuN The nuclear dyeDAPI (in Vectashield H-1200 Vector LaboratoriesBurlingame CA) was used to label the nuclei The firstprimary antibody was incubated at 4degC for 24ndash48 h in a2 goat serum2 horse serum02 Triton-X 100 in01 M PBS (block) solution followed by the second pri-mary antibody at 4degC for 1 h in block solution Fluores-cent-tagged secondary antibody (Molecular Probes Eu-gene OR) was used for visualization For double-labelingusing same species antibodies we used the tyramide sig-nal amplification (TSA) kit (PerkinElmer Life SciencesBoston MA) according to the manufacturerrsquos instruc-tions and as previously described (Stone et al 2002) The
sections were viewed and digitally captured with a ZeissAxioplan 2 microscope (Zeiss Thornwood NY)equipped with a SPOT Real Time Slider high-resolutioncolor CCD digital camera (Diagnostic Instruments IncSterling Heights MI) The number of animals used fordual-labeling IHC is as follows survivin PCNA 4survivin GFAP 6 survivin NeuN 4 PCNA GFAP 4 and PCNA NeuN 4 Cell counts wereobtained by comparing the number of double-labeledcells to total single-labeled cells in the following groupssurvivinNeuN positive cells to total NeuN positive cellssurvivinPCNA positive cells to total PCNA positivecells PCNANeuN positive cells to total NeuN positivecells and survivinGFAP positive cells to total GFAPpositive cells Percentages were calculated by dividingthe number of double-labeled cells with the total numberof single-labeled cells For each group representativephotomicrographs were selected and counted Cells werecounted in a total area of at least 188000 m2 for eachgroup with no distinction made between cortical and hip-pocampal regions
RESULTS
Induction of Survivin Expression after TBI
Q-PCR analysis revealed an initial increase in survivinmRNA at 2 days post injury in the ipsilateral cortex andhippocampus These transcripts remained elevated inboth regions reached maximum levels at day 5 post-in-jury and declined at 7 days in the cortex and at 14 daysin the hippocampus All experimental animals remainedalive and exhibited slightly impaired motor and cogni-tive impairments (data not shown) Cortical mRNA lev-els reached a maximum of 448 100 whereas hip-pocampal mRNAs attained 606 100 compared tocraniotomy control values (Fig 1) To determine if theinduction of survivin mRNA resulted in correspondingincreases in survivin protein western blot analysis wasperformed Survivin (17-kDa protein) was readily de-tectable in the ipsilateral cortex and hippocampus of TBIrats while it was negligible in contralateral cortex andhippocampus (Fig 2A) Survivin was expressed in atime-dependent manner with a maximum increase at 5days after injury followed by a gradual decline by 14days Specifically the levels of survivin in cortical tis-sue were 616 257 at 3 days and 839 339 at 5days compared to craniotomy controls (Fig 2B) Similarincreases of survivin protein in the ipsilateral hippocam-pus were detected at 3 d and 5 days post injury 464 196 and 545 102 compared to craniotomy controlrespectively (Fig 2C)
JOHNSON ET AL
1188
FIG 5 Co-localization of survivin and GFAP in brain tissue after TBI Fluorescent immunohistochemistry for survivin (green)and GFAP (red) was performed in the ipsilateral and contralateral cortex (AB) and in the CA1 and dentate gyrus regions of thehippocampus (GH) at 5 day post-injury The injury has completely destroyed the cortex in G leaving only the hippocampus inthis picture Survivin was expressed in the cytoplasm (DJ green) of GFAP-positive astrocytes (CI red) of the ipsilateral cor-tex and hippocampus and was found to co-localize to these cells as shown in merged CD and IJ images (EK respectively yel-low) White arrows indicate typical survivin-positive astrocytes Nuclei are shown using DAPI (FL blue) Original magnifica-tion 100 bar 50 m (ABGH) 400 bar 20 m (CndashF and IndashL)
FIG 6 Co-localization of PCNA and GFAP in brain tissue after TBI Double immunostaining for PCNA (green) and GFAP(red) was performed in the ipsilateral and contralateral cortex (AB) and the CA1 and dentate gyrus regions of the hippocampus(GH) at 5 day post-injury PCNA is present in GFAP-positive cells of ipsilateral cortex (CD) and to a lesser extent hippocampus(IJ) (EK) Merged CD and IJ respectively White arrows indicate typical PCNA-positive astrocytes PCNA expression wasco-incident with DAPI staining (FL blue) Original magnification 100 bar 50 m (ABGH) 400 bar 20 m (CndashFand IndashL)
PCNA Expression after TBI
For detection of proliferating cell nuclear antigen(PCNA) PVDF membranes immunostained for survivinwere stripped and re-probed using a PCNA-specific an-tibody PCNA (36-kDa protein) was significantly de-tectable in the ipsilateral cortex and hippocampus of TBIrats but only negligible amounts were observed in thecontralateral cortex and hippocampus (Fig 3A) Thetemporal patterns exhibited by PCNA protein were sim-ilar to that of survivin protein Namely PCNA expressedin a time-dependant fashion with a maximum increase at5 days after injury followed by a gradual decline by 14days The levels of PCNA in ipsilateral cortical tissuewere raised over craniotomy control by 919 459 at3 days 2263 333 at 5 days and 1035 356 at 7days post injury (Fig 3B) Similar increases of PCNAprotein in ipsilateral hippocampus were detected at 5 dayspost injury with a maximum of 1006 229 comparedto craniotomy controls (Fig 3C) No significant increasewas found in the contralateral regions when compared tocraniotomy controls (Fig 3A)
Co-Expression of Survivin and PCNA following TBI
To examine spatial co-localization of survivin andPCNA double-label immunohistochemistry of brain tis-sue sections was performed on day 5 post injury whenpeak expression of these proteins was observed
At this time point a remarkable survivin and PCNAimmunoreactivity was found in the ipsilateral cortex (Fig4A) and ipsilateral hippocampus (Fig 4B) consistent withdata obtained using Western blot analyses Within bothregions focal co-expression patterns of survivin andPCNA in single cells were detected which was demon-strated by both separate fluorescent visualization of in-dividual proteins and by merging the images of double-stained slides (Fig 4CndashE) However the dual expressionof survivin and PCNA occurred infrequently as survivinand PCNA immunoreactivity could readily be found sep-arately (Fig 4CndashE) Approximately 12 of the total num-ber of PCNA-positive cells also labeled with survivin Thenuclear morphology of dual survivin and PCNA-positivecells was ambiguous as indicated by DAPI staining (Fig4F) Therefore DAPI staining was simply used for cellidentification in all subsequent experiments
Survivin and PCNA Are Expressed in Astrocytes after TBI
To determine the cell types expressing survivin andPCNA double-label immunohistochemistry for theseproteins and GFAP a marker of astrocytes was per-formed on day 5 post injury
In accordance with Western blot data remarkable sur-vivin-positive immunoreactivity was observed in the ip-silateral cortex and hippocampus proximal to the injurycavity (Fig 5AG green) but not in the contralateral ar-eas (Fig 5BH) Survivin was co-localized with GFAPin the cells of injured cortex and hippocampus whichstrongly suggested primary accumulation of survivin incells of astrocytic lineage (Fig 5CndashEIndashL) Survivin wasuniformly distributed in the cytoplasm and processes ofastrocytes in both cortex and hippocampus (Fig 5DJ)DAPI staining is shown in Figure 5FL3 Approximately88 of the total number of GFAP-positive cells also la-beled with survivin
PCNA-positive immunoreactivity staining was ob-served in the ipsilateral cortex (Fig 6A green) and hip-pocampus (Fig 6G green) of injured brain while con-tralateral cortex and hippocampus exhibited negligiblePCNA immunoreactivity (Fig 6BH) PCNA (Fig 6CI)was partially co-localized with GFAP (Fig 6DJ red) inboth regions and was characteristically distributed in thenucleus of the cells in both cortex and hippocampus (Fig6EK) DAPI staining is shown in Figure 6FL
Taken together double-label immunohistochemistrydata provides evidence that both survivin and PCNA canbe detected in GFAP-positive astrocytes following trau-matic insult Since survivin and PCNA immunoreactiv-ity was not exclusively localized in GFAP-positive cellswe next addressed a question what other cell type mightexpress survivin after TBI We suggested that a certainpopulation of mature neurons might express survivin inresponse to injury
Survivin and PCNA Are Expressed in a Sub-Set of Neurons after TBI
As can be seen in Figure 7 survivin and PCNA wereeach co-expressed with NeuN a marker of mature neu-rons NeuN-positive cells were found to express survivinin the ipsilateral cortex distal to the injury cavity (Fig7AndashD) and in the contralateral hippocampus (Fig 7EndashH)It should be noted however that NeuN-positive cells thatalso expressed survivin occurred infrequently For ex-ample we estimated the number of dual survivinNeuNpositive cells as 01ndash15 of the total number of NeuN-positive cells in these regions Survivin immunoreactiv-ity was negligible in either hemisphere of craniotomycontrol brains (Fig 7QR) No co-localization of survivinand NeuN was observed in ipsilateral hippocampus (datanot shown) As can be seen in Figure 7BF survivin waspredominantly localized to the cytoplasm and axons ofNeuN-positive neurons DAPI staining is shown in Fig-ure 7DH
PCNA-positive neurons were found in the ipsilateralcortex (Fig 7IndashL) and hippocampus after TBI (Fig
JOHNSON ET AL
1190
1191
FIG 7 A sub-set of NeuN-positive neurons express survivin and PCNA after TBI Double fluorescent immunohistochemistryfor survivin (green) and NeuN (red) in the ipsilateral cortex (AB) and the CA1 pyramidal layer of the contralateral hippocam-pus (EF) was performed Survivin is expressed in the cytoplasm and to a limited extent in the processes of NeuN-positive neu-rons (merged images CG) Dual staining for PCNA (green) and NeuN (red) is shown in the ipsilateral cortex (IJ) and the CA1pyramidal layer of the ipsilateral hippocampus (MN) The nuclei are shown using DAPI staining (DH blue) PCNA is expressedin the nucleus of NeuN-positive neurons (merged images KO) PCNA expression was co-incident with DAPI staining in theseexamples (LP blue) White arrows indicate focal co-localization of survivinNeuN and PCNANeuN SurvivinNeuN co-local-ization of survivin (green) and NeuN (red) was seen only in TBI rats as opposed to either hemisphere of craniotomy control(QR) Original magnification 400 bar 20 m (AndashP) original magnification 50 bar 100000 m (QR)
SURVIVIN UPREGULATION AFTER TBI
7MndashP) whereas craniotomy control tissue exhibited onlytrace amounts of PCNA (data not shown) Similar to thesurvivinNeuN co-localization data dual PCNANeuNimmunostaining was a rare event accounting for approx-imately 4 of the total number of NeuN positive cellsPCNA was distributed in the nuclei of these neurons (Fig7KO) although the nuclear morphology of these cellswas not clearly resolved by DAPI staining (Fig 7LP)
DISCUSSION
Traumatic brain injury (TBI) sets into motion variousbiochemical cascades that induce neural tissue injury andcell death Conversely several proteins expressed inneural cells after TBI are directed to resist cell death andpromote recovery in the injured CNS (Ridet et al 1997Chen and Swanson 2003) Survivin is a multi-functionalprotein that inhibits apoptosis and is also required for theproper completion of mitosis Anti-apoptotic and pro-mi-togenic roles for survivin have been documented in pro-liferating cells of neural origin in vitro such as in neu-roblastoma and glioma cells (LaCasse et al 1998 Tammet al 1998 Deveraux and Reed 1999 Conway et al2000 Shin et al 2001 Sasaki et al 2002) Howeverno studies have investigated the potential role of survivinin the adult brain after TBI when a sub-population ofCNS cells may initiate a cell cycle-related process in re-sponse to injury
In the present paper we demonstrate the induction ofsurvivin expression in rat brain subjected to TBI The ex-pression of survivin was time-dependent and cell-spe-cific and was present in astrocytes and to a much lesserextent in neurons in brain cortex and hippocampus In-duction of survivin in these cells was accompanied byoccasional expression of PCNA a cell cycle protein in-volved in mitotic G1S progression Our present data arethe first to show that survivin mRNA and protein are sig-nificantly up-regulated after traumatic brain injury in ratsPCNA expression after TBI has been described previously(Miyake et al 1992 Chen et al 2003) suggesting itsrole in mechanisms of brain recovery after injury Theconcurrent up-regulation of survivin with a similar tem-poral profile as PCNA shown herein further suggests thatsurvivin may play a role in cellular proliferation after TBI
Brain injury evoked the expression of survivin andPCNA in a time-dependent manner (Figs 2 and 3) West-ern blot analysis revealed maximal co-expression of bothsurvivin and PCNA at five days post injury Immuno-histochemistry at this time point demonstrated co-local-ization of these proteins (Fig 4) although most cells werelabeled separately with PCNA and survivin In fact only12 of the total number of PCNA-positive cells were
also survivin positive It has been reported that PCNA isexpressed predominantly in G1S (Bravo et al 1987)while survivin is found at the G2M phase of the cell cy-cle (Bravo et al 1987 Otaki et al 2000) Hence a lackof strict co-localization of survivin and PCNA in ourstudy may be explained by their expression at differentpoints in the cell cycle
In our experimental model we observed survivin- aswell as PCNA-positive astrocytes in the proximal area ofthe injury and in the ipsilateral hippocampus Prolifera-tion of astrocytes is well documented after TBI as shownby cell labeling with BrdU as well as expression of PCNA(Latov et al 1979 Dunn-Meynell and Levin 1997 Car-bonell and Grady 1999 Norton 1999 Csuka et al 2000Kernie et al 2001 Chen et al 2003) Because survivinand PCNA were expressed in astrocytes following TBI(Figs 5 and 6) it is possible that survivin plays an im-portant role linking astrocyte survival and proliferationafter traumatic insult Astrocyte proliferation has beenimplicated in the formation of the glial scar observed af-ter injury (Latov et al 1979) and creates a non-permis-sive environment for repair (Sykova et al 1999) How-ever glial proliferation may also enhance neuronalsurvival (Smith et al 2001 Wei et al 2001)
Of particular interest is a sub-set of NeuN-positive neu-rons found to express survivin only after TBI (Fig 7)These cells were much less abundant than survivin-pos-itive astrocytes and their functional significance is cur-rently unknown However both neurons and astrocyteshave been documented previously to express cell cycleproteins after various insults such as exposure to -amy-loidndashactivated microglia (Wu et al 2000) TBI (Kaya etal 1999ab) chlorine toxicity (Magavi et al 2000) oras a consequence of Alzheimerrsquos disease (Yang et al2001) These papers underscore the significant contro-versy that exists regarding the function of cell cycle pro-teins such as PCNA in neurons after different types ofinjury We are currently conducting further studies whichwill elucidate the roles for PCNA and survivin in neu-rons after TBI
It should be noted that dual staining of survivin andPCNA could not be directly attributed to a specific celltype due to the technical difficulties of triple labeling an-tibody-based IHC Therefore we cannot rule out the pos-sibility that other cell types such as endothelial (Conwayet al 2003) or inflammatory cells (Hill-Felberg et al1999) may also contribute to survivin and PCNA ex-pression after TBI The appearance of survivin and PCNAseparately in neurons (NeuN-positive) and astrocytes(GFAP-positive) along with co-localization of survivinwith PCNA in the same cells provide correlative data tosuggest an activation of cell cycle-like program in astro-cytes and possibly in a small subtype of neurons after
JOHNSON ET AL
1192
TBI In our experiments survivin co-localization withPCNA does suggest that survivin may be associated witha pro-mitotic process In an attempt to clarify these pro-teinrsquos roles after TBI we analyzed the nuclear morphol-ogy of survivin-positive cells to define the apoptotic ormitotic architecture of nuclei DAPI staining proved tooambiguous in identifying apoptotic versus mitotic phe-notypes likely due to the thickness of the brain sections(40 m) Further studies using direct markers of mitosissuch as BrdU incorporation as well as simultaneous la-beling with cell death related proteins is required to de-lineate anti-apoptotic and pro-mitotic activities of sur-vivin and PCNA in these cells
In conclusion our data demonstrate the induction ofsurvivin in the rat brain following TBI Expression of sur-vivin occurred predominantly in astrocytes as comparedto neurons in a time-dependant fashion and was accom-panied by expression of PCNA Taken together these re-sults suggest that survivin plays a role in neural cell re-sponses following traumatic brain injury in rats Futurestudies will investigate the implications of these findingsto the pathophysiology of TBI
ACKNOWLEDGMENTS
We thank Barbara OrsquoSteen and Tao Fan MD fortechnical assistance This work was funded by grants NIHRO1 NS39091 NIH RO1 NS40182 DAMD 17-99-1-9565 and DAMD 17-01-1-0765
REFERENCES
ADIDA C CROTTY PL MCGRATH J BERREBI DDIEBOLD J and ALTIERI DC (1998) Developmentallyregulated expression of the novel cancer anti-apoptosis genesurvivin in human and mouse differentiation Am J Pathol152 43ndash49
ALTIERI DC MARCHISIO PC and MARCHISIO C(1999) Survivin apoptosis an interloper between cell deathand cell proliferation in cancer Lab Invest 79 1327ndash1333
ALTSCHUL SF MADDEN TL SCHAFFER AA et al(1997) Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 253389ndash3402
AMBROSINI G ADIDA C and ALTIERI DC (1997) Anovel anti-apoptosis gene survivin expressed in cancer andlymphoma Nat Med 3 917ndash921
BRAVO R FRANK R BLUNDELL PA and MAC-DONALD-BRAVO H (1987) CyclinPCNA is the auxil-iary protein of DNA polymerase-delta Nature 326 515ndash517
BUSH TG PUVANACHANDRA N HORNER CH et al
(1999) Leukocyte infiltration neuronal degeneration andneurite outgrowth after ablation of scar-forming reactive as-trocytes in adult transgenic mice Neuron 23 297ndash308
CAMERON HA and MCKAY RD (2001) Adult neuroge-nesis produces a large pool of new granule cells in the den-tate gyrus J Comp Neurol 435 406ndash417
CARBONELL WS and GRADY MS (1999) Regional andtemporal characterization of neuronal glial and axonal re-sponse after traumatic brain injury in the mouse Acta Neu-ropathol (Berl) 98 396ndash406
CHEN XH IWATA A NONAKA M BROWNE KDand SMITH DH (2003) Neurogenesis and glial prolifera-tion persist for at least one year in the subventricular zonefollowing brain trauma in rats J Neurotrauma 20 623ndash631
CHEN Y and SWANSON RA (2003) Astrocytes and braininjury J Cereb Blood Flow Metab 23 137ndash149
CONWAY EM POLLEFEYT S CORNELISSEN J et al(2000) Three differentially expressed survivin cDNA vari-ants encode proteins with distinct antiapoptotic functionsBlood 95 1435ndash1442
CSUKA E HANS VH AMMANN E TRENTZ OKOSSMANN T and MORGANTI-KOSSMANN MC(2000) Cell activation and inflammatory response followingtraumatic axonal injury in the rat Neuroreport 112587ndash2590
DEVERAUX QL and REED JC (1999) IAP family pro-teinsmdashsuppressors of apoptosis Genes Dev 13 239ndash252
DIXON CE CLIFTON GL LIGHTHALL JW YAGH-MAI AA and HAYES RL (1991) A controlled corticalimpact model of traumatic brain injury in the rat J NeurosciMethods 39 253ndash262
DOETSCH F CAILLE I LIM DA GARCIA-VERDUGOJM and ALVAREZ-BUYLLA A (1999) Subventricularzone astrocytes are neural stem cells in the adult mammalianbrain Cell 97 703ndash716
DUNN-MEYNELL AA and LEVIN BE (1997) Histolog-ical markers of neuronal axonal and astrocytic changes af-ter lateral rigid impact traumatic brain injury Brain Res 76125ndash41
GIULIAN D (1991) Microgliamdashneuron interactions after in-jury to the central nervous system in Peripheral Signalingof the Brain Role in Neural-Immune Interactions Learningand Memory R Frederickson J McGaugh and D Felton(eds) Lewiston NY Hogrefe and Huber pps 73ndash82
HIGGINS DG BLEASBY AJ and FUCHS R (1992)CLUSTAL V improved software for multiple sequencealignment Comput Appl Biosci 8 189ndash191
HILL-FELBERG SJ MCINTOSH TK OLIVER DLRAGHUPATHI R and BARBARESE E (1999) Concur-rent loss and proliferation of astrocytes following lateral fluidpercussion brain injury in the adult rat J Neurosci Res 57271ndash279
1193
SURVIVIN UPREGULATION AFTER TBI
JIANG X WILFORD C DUENSING S MUNGER KJONES G and JONES D (2001) Participation of Survivinin mitotic and apoptotic activities of normal and tumor-de-rived cells J Cell Biochem 83 342ndash354
KAYA SS MAHMOOD A LI Y YAVUZ E andCHOPP M (1999a) Expression of cell cycle proteins (cy-clin D1 and cdk4) after controlled cortical impact in rat brainJ Neurotrauma 16 1187ndash1196
KAYA SS MAHMOOD A LI Y YAVUZ E GOKSELM and CHOPP M (1999b) Apoptosis and expression ofp53 response proteins and cyclin D1 after cortical impact inrat brain Brain Res 818 23ndash33
KERNIE SG ERWIN TM and PARADA LF (2001)Brain remodeling due to neuronal and astrocytic prolifera-tion after controlled cortical injury in mice J Neurosci Res66 317ndash326
KOBAYASHI K HATANO M OTAKI M OGA-SAWARA T and TOKUHISA T (1999) Expression of amurine homologue of the inhibitor of apoptosis protein is re-lated to cell proliferation Proc Natl Acad Sci USA 961457ndash1462
LACASSE EC BAIRD S KORNELUK RG andMACKENZIE AE (1998) The inhibitors of apoptosis(IAPs) and their emerging role in cancer Oncogene 173247ndash3259
LATOV N NILAVER G ZIMMERMAN EA et al(1979) Fibrillary astrocytes proliferate in response to braininjury a study combining immunoperoxidase technique forglial fibrillary acidic protein and radioautography of tritiatedthymidine Dev Biol 72 381ndash384
LI F and ALTIERI DC (1999) The cancer antiapoptosismouse survivin gene characterization of locus and tran-scriptional requirements of basal and cell cycle-dependentexpression Cancer Res 59 3143ndash3151
LI F AMBROSINI G CHU EY et al (1998) Control ofapoptosis and mitotic spindle checkpoint by survivin Nature396 580ndash584
LI F ACKERMANN EJ BENNETT CF et al (1999)Pleiotropic cell-division defects and apoptosis induced by in-terference with survivin function Nat Cell Biol 1 461ndash466
MAGAVI SS LEAVITT BR and MACKLIS JD (2000)Induction of neurogenesis in the neocortex of adult mice Na-ture 405 951ndash955
MIYAKE T OKADA M and KITAMURA T (1992) Re-active proliferation of astrocytes studied by immunohisto-chemistry for proliferating cell nuclear antigen Brain Res590 300ndash302
MUCHMORE SW CHEN J JAKOB C et al (2000)Crystal structure and mutagenic analysis of the inhibitor-of-apoptosis protein survivin Mol Cell 6 173ndash182
NORTON WT (1999) Cell reactions following acute braininjury a review Neurochem Res 24 213ndash218
OTAKI M HATANO M KOBAYASHI K et al (2000)Cell cyclendashdependent regulation of TIAPm-survivin expres-sion Biochim Biophys Acta 1493 188ndash194
PETERSON DA (2002) Stem cells in brain plasticity and re-pair Curr Opin Pharmacol 2 34ndash42
PIKE BR ZHAO X NEWCOMB JK POSMANTURRM WANG KK and Hayes RL (1998) Regional cal-pain and caspase-3 proteolysis of alpha-spectrin after trau-matic brain injury Neuroreport 9 2437ndash2442
RAGHUPATHI R GRAHAM DI MCINTOSH TK(2000) Apoptosis after traumatic brain injury J Neuro-trauma 17 927ndash938
RIDET JL MALHOTRA SK PRIVAT A and GAGEFH (1997) Reactive astrocytes cellular and molecular cuesto biological function Trends Neurosci 20 570ndash577
SASAKI T LOPES MB HANKINS GR and HELMGA (2002) Expression of survivin an inhibitor of apopto-sis protein in tumors of the nervous system Acta Neu-ropathol (Berl) 104 105ndash109
SHANKAR SL MANI S OrsquoGUIN KN KANDIMALLAER AGRAWAL S and SHAFIT-ZAGARDO B (2001)Survivin inhibition induces human neural tumor cell deaththrough caspase-independent and -dependent pathways JNeurochem 79 426ndash436
SHIN S SUNG BJ CHO YS et al (2001) An anti-apop-totic protein human survivin is a direct inhibitor of caspase-3 and -7 Biochemistry 40 1117ndash1123
SMITH C BERRY M CLARKE WE and LOGAN A(2001) Differential expression of fibroblast growth factorndash2and fibroblast growth factor receptor 1 in a scarring and non-scarring model of CNS injury in the rat Eur J Neurosci 13443ndash456
STONE JR OKONKWO DO SINGLETON RHMUTLU LK HELM GA and POVLISHOCK JT(2002) Caspase-3ndashmediated cleavage of amyloid precursorprotein and formation of amyloid Beta peptide in traumaticaxonal injury J Neurotrauma 19 601ndash614
SYKOVA E VARGOVA L PROKOPOVA S and SI-MONOVA Z (1999) Glial swelling and astrogliosis pro-duce diffusion barriers in the rat spinal cord Glia 25 56ndash70
TAKAHASHI R DEVERAUX Q TAMM I et al (1998)A single BIR domain of XIAP sufficient for inhibiting cas-pases J Biol Chem 273 7787ndash7790
TAMM I WANG Y SAUSVILLE E et al (1998) IAP-family protein survivin inhibits caspase activity and apopto-sis induced by Fas (CD95) Bax caspases and anticancerdrugs Cancer Res 58 5315ndash5320
THOMPSON JD HIGGINS DG and GIBSON TJ (1994)CLUSTAL W improving the sensitivity of progressive mul-tiple sequence alignment through sequence weighting posi-tion-specific gap penalties and weight matrix choice NucleicAcids Res 22 4673ndash4680
JOHNSON ET AL
1194
TOLENTINO PJ DEFORD SM NOTTERPEK L et al(2002) Up-regulation of tissue-type transglutaminase aftertraumatic brain injury J Neurochem 80 579ndash588
UREN AG WONG L PAKUSCH M et al (2000) Sur-vivin and the inner centromere protein INCENP show simi-lar cell-cycle localization and gene knockout phenotypeCurr Biol 10 1319ndash1328
WEI LH HUANG CY CHENG SP CHEN CA andHSIEH CY (2001) Carcinosarcoma of ovary associatedwith previous radiotherapy Int J Gynecol Cancer 11 81ndash84
WU Q COMBS C CANNADY SB GELDMACHERDS and HERRUP K (2000) Beta-amyloid activated mi-croglia induce cell cycling and cell death in cultured corticalneurons Neurobiol Aging 21 797ndash806
YAGITA Y KITAGAWA K OHTSUKI T et al (2001)
Neurogenesis by progenitor cells in the ischemic adult rathippocampus Stroke 32 1890ndash1896
YANG Y GELDMACHER DS and HERRUP K (2001)DNA replication precedes neuronal cell death in Alzheimerrsquosdisease J Neurosci 21 2661ndash2668
Address reprint requests toJose A Pineda PhD
EF and WL McKnight Brain InstituteUniversity of Florida
100 S Newell Dr Bldg 59L1-118 Dept of Pediatrics
Gainesville FL 32610
E-mail pinedjapedsufledu
SURVIVIN UPREGULATION AFTER TBI
1195
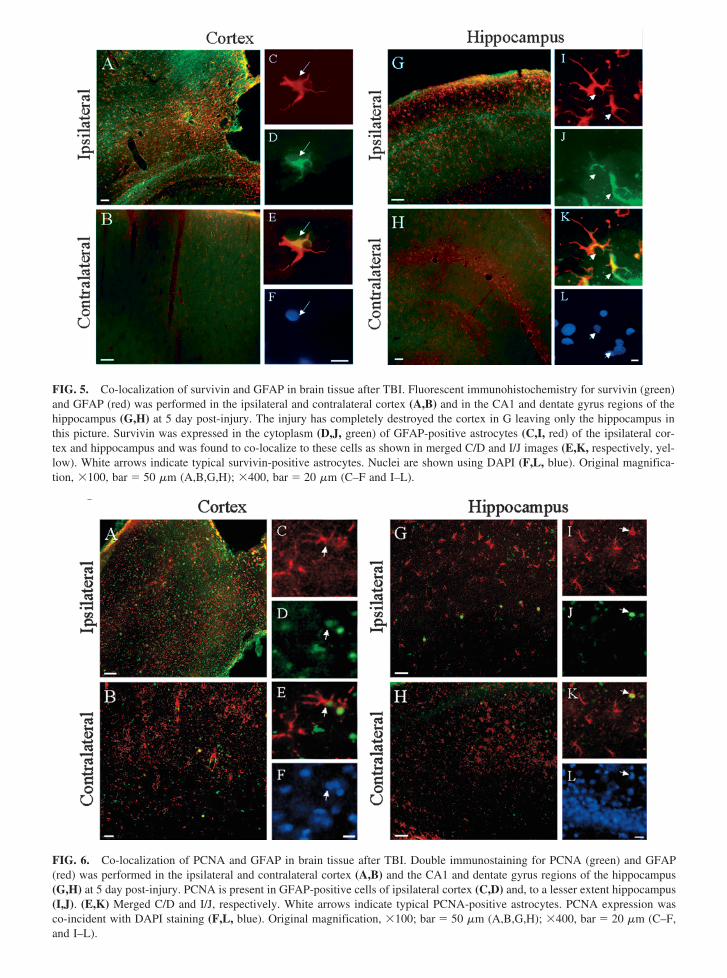
FIG 5 Co-localization of survivin and GFAP in brain tissue after TBI Fluorescent immunohistochemistry for survivin (green)and GFAP (red) was performed in the ipsilateral and contralateral cortex (AB) and in the CA1 and dentate gyrus regions of thehippocampus (GH) at 5 day post-injury The injury has completely destroyed the cortex in G leaving only the hippocampus inthis picture Survivin was expressed in the cytoplasm (DJ green) of GFAP-positive astrocytes (CI red) of the ipsilateral cor-tex and hippocampus and was found to co-localize to these cells as shown in merged CD and IJ images (EK respectively yel-low) White arrows indicate typical survivin-positive astrocytes Nuclei are shown using DAPI (FL blue) Original magnifica-tion 100 bar 50 m (ABGH) 400 bar 20 m (CndashF and IndashL)
FIG 6 Co-localization of PCNA and GFAP in brain tissue after TBI Double immunostaining for PCNA (green) and GFAP(red) was performed in the ipsilateral and contralateral cortex (AB) and the CA1 and dentate gyrus regions of the hippocampus(GH) at 5 day post-injury PCNA is present in GFAP-positive cells of ipsilateral cortex (CD) and to a lesser extent hippocampus(IJ) (EK) Merged CD and IJ respectively White arrows indicate typical PCNA-positive astrocytes PCNA expression wasco-incident with DAPI staining (FL blue) Original magnification 100 bar 50 m (ABGH) 400 bar 20 m (CndashFand IndashL)
PCNA Expression after TBI
For detection of proliferating cell nuclear antigen(PCNA) PVDF membranes immunostained for survivinwere stripped and re-probed using a PCNA-specific an-tibody PCNA (36-kDa protein) was significantly de-tectable in the ipsilateral cortex and hippocampus of TBIrats but only negligible amounts were observed in thecontralateral cortex and hippocampus (Fig 3A) Thetemporal patterns exhibited by PCNA protein were sim-ilar to that of survivin protein Namely PCNA expressedin a time-dependant fashion with a maximum increase at5 days after injury followed by a gradual decline by 14days The levels of PCNA in ipsilateral cortical tissuewere raised over craniotomy control by 919 459 at3 days 2263 333 at 5 days and 1035 356 at 7days post injury (Fig 3B) Similar increases of PCNAprotein in ipsilateral hippocampus were detected at 5 dayspost injury with a maximum of 1006 229 comparedto craniotomy controls (Fig 3C) No significant increasewas found in the contralateral regions when compared tocraniotomy controls (Fig 3A)
Co-Expression of Survivin and PCNA following TBI
To examine spatial co-localization of survivin andPCNA double-label immunohistochemistry of brain tis-sue sections was performed on day 5 post injury whenpeak expression of these proteins was observed
At this time point a remarkable survivin and PCNAimmunoreactivity was found in the ipsilateral cortex (Fig4A) and ipsilateral hippocampus (Fig 4B) consistent withdata obtained using Western blot analyses Within bothregions focal co-expression patterns of survivin andPCNA in single cells were detected which was demon-strated by both separate fluorescent visualization of in-dividual proteins and by merging the images of double-stained slides (Fig 4CndashE) However the dual expressionof survivin and PCNA occurred infrequently as survivinand PCNA immunoreactivity could readily be found sep-arately (Fig 4CndashE) Approximately 12 of the total num-ber of PCNA-positive cells also labeled with survivin Thenuclear morphology of dual survivin and PCNA-positivecells was ambiguous as indicated by DAPI staining (Fig4F) Therefore DAPI staining was simply used for cellidentification in all subsequent experiments
Survivin and PCNA Are Expressed in Astrocytes after TBI
To determine the cell types expressing survivin andPCNA double-label immunohistochemistry for theseproteins and GFAP a marker of astrocytes was per-formed on day 5 post injury
In accordance with Western blot data remarkable sur-vivin-positive immunoreactivity was observed in the ip-silateral cortex and hippocampus proximal to the injurycavity (Fig 5AG green) but not in the contralateral ar-eas (Fig 5BH) Survivin was co-localized with GFAPin the cells of injured cortex and hippocampus whichstrongly suggested primary accumulation of survivin incells of astrocytic lineage (Fig 5CndashEIndashL) Survivin wasuniformly distributed in the cytoplasm and processes ofastrocytes in both cortex and hippocampus (Fig 5DJ)DAPI staining is shown in Figure 5FL3 Approximately88 of the total number of GFAP-positive cells also la-beled with survivin
PCNA-positive immunoreactivity staining was ob-served in the ipsilateral cortex (Fig 6A green) and hip-pocampus (Fig 6G green) of injured brain while con-tralateral cortex and hippocampus exhibited negligiblePCNA immunoreactivity (Fig 6BH) PCNA (Fig 6CI)was partially co-localized with GFAP (Fig 6DJ red) inboth regions and was characteristically distributed in thenucleus of the cells in both cortex and hippocampus (Fig6EK) DAPI staining is shown in Figure 6FL
Taken together double-label immunohistochemistrydata provides evidence that both survivin and PCNA canbe detected in GFAP-positive astrocytes following trau-matic insult Since survivin and PCNA immunoreactiv-ity was not exclusively localized in GFAP-positive cellswe next addressed a question what other cell type mightexpress survivin after TBI We suggested that a certainpopulation of mature neurons might express survivin inresponse to injury
Survivin and PCNA Are Expressed in a Sub-Set of Neurons after TBI
As can be seen in Figure 7 survivin and PCNA wereeach co-expressed with NeuN a marker of mature neu-rons NeuN-positive cells were found to express survivinin the ipsilateral cortex distal to the injury cavity (Fig7AndashD) and in the contralateral hippocampus (Fig 7EndashH)It should be noted however that NeuN-positive cells thatalso expressed survivin occurred infrequently For ex-ample we estimated the number of dual survivinNeuNpositive cells as 01ndash15 of the total number of NeuN-positive cells in these regions Survivin immunoreactiv-ity was negligible in either hemisphere of craniotomycontrol brains (Fig 7QR) No co-localization of survivinand NeuN was observed in ipsilateral hippocampus (datanot shown) As can be seen in Figure 7BF survivin waspredominantly localized to the cytoplasm and axons ofNeuN-positive neurons DAPI staining is shown in Fig-ure 7DH
PCNA-positive neurons were found in the ipsilateralcortex (Fig 7IndashL) and hippocampus after TBI (Fig
JOHNSON ET AL
1190
1191
FIG 7 A sub-set of NeuN-positive neurons express survivin and PCNA after TBI Double fluorescent immunohistochemistryfor survivin (green) and NeuN (red) in the ipsilateral cortex (AB) and the CA1 pyramidal layer of the contralateral hippocam-pus (EF) was performed Survivin is expressed in the cytoplasm and to a limited extent in the processes of NeuN-positive neu-rons (merged images CG) Dual staining for PCNA (green) and NeuN (red) is shown in the ipsilateral cortex (IJ) and the CA1pyramidal layer of the ipsilateral hippocampus (MN) The nuclei are shown using DAPI staining (DH blue) PCNA is expressedin the nucleus of NeuN-positive neurons (merged images KO) PCNA expression was co-incident with DAPI staining in theseexamples (LP blue) White arrows indicate focal co-localization of survivinNeuN and PCNANeuN SurvivinNeuN co-local-ization of survivin (green) and NeuN (red) was seen only in TBI rats as opposed to either hemisphere of craniotomy control(QR) Original magnification 400 bar 20 m (AndashP) original magnification 50 bar 100000 m (QR)
SURVIVIN UPREGULATION AFTER TBI
7MndashP) whereas craniotomy control tissue exhibited onlytrace amounts of PCNA (data not shown) Similar to thesurvivinNeuN co-localization data dual PCNANeuNimmunostaining was a rare event accounting for approx-imately 4 of the total number of NeuN positive cellsPCNA was distributed in the nuclei of these neurons (Fig7KO) although the nuclear morphology of these cellswas not clearly resolved by DAPI staining (Fig 7LP)
DISCUSSION
Traumatic brain injury (TBI) sets into motion variousbiochemical cascades that induce neural tissue injury andcell death Conversely several proteins expressed inneural cells after TBI are directed to resist cell death andpromote recovery in the injured CNS (Ridet et al 1997Chen and Swanson 2003) Survivin is a multi-functionalprotein that inhibits apoptosis and is also required for theproper completion of mitosis Anti-apoptotic and pro-mi-togenic roles for survivin have been documented in pro-liferating cells of neural origin in vitro such as in neu-roblastoma and glioma cells (LaCasse et al 1998 Tammet al 1998 Deveraux and Reed 1999 Conway et al2000 Shin et al 2001 Sasaki et al 2002) Howeverno studies have investigated the potential role of survivinin the adult brain after TBI when a sub-population ofCNS cells may initiate a cell cycle-related process in re-sponse to injury
In the present paper we demonstrate the induction ofsurvivin expression in rat brain subjected to TBI The ex-pression of survivin was time-dependent and cell-spe-cific and was present in astrocytes and to a much lesserextent in neurons in brain cortex and hippocampus In-duction of survivin in these cells was accompanied byoccasional expression of PCNA a cell cycle protein in-volved in mitotic G1S progression Our present data arethe first to show that survivin mRNA and protein are sig-nificantly up-regulated after traumatic brain injury in ratsPCNA expression after TBI has been described previously(Miyake et al 1992 Chen et al 2003) suggesting itsrole in mechanisms of brain recovery after injury Theconcurrent up-regulation of survivin with a similar tem-poral profile as PCNA shown herein further suggests thatsurvivin may play a role in cellular proliferation after TBI
Brain injury evoked the expression of survivin andPCNA in a time-dependent manner (Figs 2 and 3) West-ern blot analysis revealed maximal co-expression of bothsurvivin and PCNA at five days post injury Immuno-histochemistry at this time point demonstrated co-local-ization of these proteins (Fig 4) although most cells werelabeled separately with PCNA and survivin In fact only12 of the total number of PCNA-positive cells were
also survivin positive It has been reported that PCNA isexpressed predominantly in G1S (Bravo et al 1987)while survivin is found at the G2M phase of the cell cy-cle (Bravo et al 1987 Otaki et al 2000) Hence a lackof strict co-localization of survivin and PCNA in ourstudy may be explained by their expression at differentpoints in the cell cycle
In our experimental model we observed survivin- aswell as PCNA-positive astrocytes in the proximal area ofthe injury and in the ipsilateral hippocampus Prolifera-tion of astrocytes is well documented after TBI as shownby cell labeling with BrdU as well as expression of PCNA(Latov et al 1979 Dunn-Meynell and Levin 1997 Car-bonell and Grady 1999 Norton 1999 Csuka et al 2000Kernie et al 2001 Chen et al 2003) Because survivinand PCNA were expressed in astrocytes following TBI(Figs 5 and 6) it is possible that survivin plays an im-portant role linking astrocyte survival and proliferationafter traumatic insult Astrocyte proliferation has beenimplicated in the formation of the glial scar observed af-ter injury (Latov et al 1979) and creates a non-permis-sive environment for repair (Sykova et al 1999) How-ever glial proliferation may also enhance neuronalsurvival (Smith et al 2001 Wei et al 2001)
Of particular interest is a sub-set of NeuN-positive neu-rons found to express survivin only after TBI (Fig 7)These cells were much less abundant than survivin-pos-itive astrocytes and their functional significance is cur-rently unknown However both neurons and astrocyteshave been documented previously to express cell cycleproteins after various insults such as exposure to -amy-loidndashactivated microglia (Wu et al 2000) TBI (Kaya etal 1999ab) chlorine toxicity (Magavi et al 2000) oras a consequence of Alzheimerrsquos disease (Yang et al2001) These papers underscore the significant contro-versy that exists regarding the function of cell cycle pro-teins such as PCNA in neurons after different types ofinjury We are currently conducting further studies whichwill elucidate the roles for PCNA and survivin in neu-rons after TBI
It should be noted that dual staining of survivin andPCNA could not be directly attributed to a specific celltype due to the technical difficulties of triple labeling an-tibody-based IHC Therefore we cannot rule out the pos-sibility that other cell types such as endothelial (Conwayet al 2003) or inflammatory cells (Hill-Felberg et al1999) may also contribute to survivin and PCNA ex-pression after TBI The appearance of survivin and PCNAseparately in neurons (NeuN-positive) and astrocytes(GFAP-positive) along with co-localization of survivinwith PCNA in the same cells provide correlative data tosuggest an activation of cell cycle-like program in astro-cytes and possibly in a small subtype of neurons after
JOHNSON ET AL
1192
TBI In our experiments survivin co-localization withPCNA does suggest that survivin may be associated witha pro-mitotic process In an attempt to clarify these pro-teinrsquos roles after TBI we analyzed the nuclear morphol-ogy of survivin-positive cells to define the apoptotic ormitotic architecture of nuclei DAPI staining proved tooambiguous in identifying apoptotic versus mitotic phe-notypes likely due to the thickness of the brain sections(40 m) Further studies using direct markers of mitosissuch as BrdU incorporation as well as simultaneous la-beling with cell death related proteins is required to de-lineate anti-apoptotic and pro-mitotic activities of sur-vivin and PCNA in these cells
In conclusion our data demonstrate the induction ofsurvivin in the rat brain following TBI Expression of sur-vivin occurred predominantly in astrocytes as comparedto neurons in a time-dependant fashion and was accom-panied by expression of PCNA Taken together these re-sults suggest that survivin plays a role in neural cell re-sponses following traumatic brain injury in rats Futurestudies will investigate the implications of these findingsto the pathophysiology of TBI
ACKNOWLEDGMENTS
We thank Barbara OrsquoSteen and Tao Fan MD fortechnical assistance This work was funded by grants NIHRO1 NS39091 NIH RO1 NS40182 DAMD 17-99-1-9565 and DAMD 17-01-1-0765
REFERENCES
ADIDA C CROTTY PL MCGRATH J BERREBI DDIEBOLD J and ALTIERI DC (1998) Developmentallyregulated expression of the novel cancer anti-apoptosis genesurvivin in human and mouse differentiation Am J Pathol152 43ndash49
ALTIERI DC MARCHISIO PC and MARCHISIO C(1999) Survivin apoptosis an interloper between cell deathand cell proliferation in cancer Lab Invest 79 1327ndash1333
ALTSCHUL SF MADDEN TL SCHAFFER AA et al(1997) Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 253389ndash3402
AMBROSINI G ADIDA C and ALTIERI DC (1997) Anovel anti-apoptosis gene survivin expressed in cancer andlymphoma Nat Med 3 917ndash921
BRAVO R FRANK R BLUNDELL PA and MAC-DONALD-BRAVO H (1987) CyclinPCNA is the auxil-iary protein of DNA polymerase-delta Nature 326 515ndash517
BUSH TG PUVANACHANDRA N HORNER CH et al
(1999) Leukocyte infiltration neuronal degeneration andneurite outgrowth after ablation of scar-forming reactive as-trocytes in adult transgenic mice Neuron 23 297ndash308
CAMERON HA and MCKAY RD (2001) Adult neuroge-nesis produces a large pool of new granule cells in the den-tate gyrus J Comp Neurol 435 406ndash417
CARBONELL WS and GRADY MS (1999) Regional andtemporal characterization of neuronal glial and axonal re-sponse after traumatic brain injury in the mouse Acta Neu-ropathol (Berl) 98 396ndash406
CHEN XH IWATA A NONAKA M BROWNE KDand SMITH DH (2003) Neurogenesis and glial prolifera-tion persist for at least one year in the subventricular zonefollowing brain trauma in rats J Neurotrauma 20 623ndash631
CHEN Y and SWANSON RA (2003) Astrocytes and braininjury J Cereb Blood Flow Metab 23 137ndash149
CONWAY EM POLLEFEYT S CORNELISSEN J et al(2000) Three differentially expressed survivin cDNA vari-ants encode proteins with distinct antiapoptotic functionsBlood 95 1435ndash1442
CSUKA E HANS VH AMMANN E TRENTZ OKOSSMANN T and MORGANTI-KOSSMANN MC(2000) Cell activation and inflammatory response followingtraumatic axonal injury in the rat Neuroreport 112587ndash2590
DEVERAUX QL and REED JC (1999) IAP family pro-teinsmdashsuppressors of apoptosis Genes Dev 13 239ndash252
DIXON CE CLIFTON GL LIGHTHALL JW YAGH-MAI AA and HAYES RL (1991) A controlled corticalimpact model of traumatic brain injury in the rat J NeurosciMethods 39 253ndash262
DOETSCH F CAILLE I LIM DA GARCIA-VERDUGOJM and ALVAREZ-BUYLLA A (1999) Subventricularzone astrocytes are neural stem cells in the adult mammalianbrain Cell 97 703ndash716
DUNN-MEYNELL AA and LEVIN BE (1997) Histolog-ical markers of neuronal axonal and astrocytic changes af-ter lateral rigid impact traumatic brain injury Brain Res 76125ndash41
GIULIAN D (1991) Microgliamdashneuron interactions after in-jury to the central nervous system in Peripheral Signalingof the Brain Role in Neural-Immune Interactions Learningand Memory R Frederickson J McGaugh and D Felton(eds) Lewiston NY Hogrefe and Huber pps 73ndash82
HIGGINS DG BLEASBY AJ and FUCHS R (1992)CLUSTAL V improved software for multiple sequencealignment Comput Appl Biosci 8 189ndash191
HILL-FELBERG SJ MCINTOSH TK OLIVER DLRAGHUPATHI R and BARBARESE E (1999) Concur-rent loss and proliferation of astrocytes following lateral fluidpercussion brain injury in the adult rat J Neurosci Res 57271ndash279
1193
SURVIVIN UPREGULATION AFTER TBI
JIANG X WILFORD C DUENSING S MUNGER KJONES G and JONES D (2001) Participation of Survivinin mitotic and apoptotic activities of normal and tumor-de-rived cells J Cell Biochem 83 342ndash354
KAYA SS MAHMOOD A LI Y YAVUZ E andCHOPP M (1999a) Expression of cell cycle proteins (cy-clin D1 and cdk4) after controlled cortical impact in rat brainJ Neurotrauma 16 1187ndash1196
KAYA SS MAHMOOD A LI Y YAVUZ E GOKSELM and CHOPP M (1999b) Apoptosis and expression ofp53 response proteins and cyclin D1 after cortical impact inrat brain Brain Res 818 23ndash33
KERNIE SG ERWIN TM and PARADA LF (2001)Brain remodeling due to neuronal and astrocytic prolifera-tion after controlled cortical injury in mice J Neurosci Res66 317ndash326
KOBAYASHI K HATANO M OTAKI M OGA-SAWARA T and TOKUHISA T (1999) Expression of amurine homologue of the inhibitor of apoptosis protein is re-lated to cell proliferation Proc Natl Acad Sci USA 961457ndash1462
LACASSE EC BAIRD S KORNELUK RG andMACKENZIE AE (1998) The inhibitors of apoptosis(IAPs) and their emerging role in cancer Oncogene 173247ndash3259
LATOV N NILAVER G ZIMMERMAN EA et al(1979) Fibrillary astrocytes proliferate in response to braininjury a study combining immunoperoxidase technique forglial fibrillary acidic protein and radioautography of tritiatedthymidine Dev Biol 72 381ndash384
LI F and ALTIERI DC (1999) The cancer antiapoptosismouse survivin gene characterization of locus and tran-scriptional requirements of basal and cell cycle-dependentexpression Cancer Res 59 3143ndash3151
LI F AMBROSINI G CHU EY et al (1998) Control ofapoptosis and mitotic spindle checkpoint by survivin Nature396 580ndash584
LI F ACKERMANN EJ BENNETT CF et al (1999)Pleiotropic cell-division defects and apoptosis induced by in-terference with survivin function Nat Cell Biol 1 461ndash466
MAGAVI SS LEAVITT BR and MACKLIS JD (2000)Induction of neurogenesis in the neocortex of adult mice Na-ture 405 951ndash955
MIYAKE T OKADA M and KITAMURA T (1992) Re-active proliferation of astrocytes studied by immunohisto-chemistry for proliferating cell nuclear antigen Brain Res590 300ndash302
MUCHMORE SW CHEN J JAKOB C et al (2000)Crystal structure and mutagenic analysis of the inhibitor-of-apoptosis protein survivin Mol Cell 6 173ndash182
NORTON WT (1999) Cell reactions following acute braininjury a review Neurochem Res 24 213ndash218
OTAKI M HATANO M KOBAYASHI K et al (2000)Cell cyclendashdependent regulation of TIAPm-survivin expres-sion Biochim Biophys Acta 1493 188ndash194
PETERSON DA (2002) Stem cells in brain plasticity and re-pair Curr Opin Pharmacol 2 34ndash42
PIKE BR ZHAO X NEWCOMB JK POSMANTURRM WANG KK and Hayes RL (1998) Regional cal-pain and caspase-3 proteolysis of alpha-spectrin after trau-matic brain injury Neuroreport 9 2437ndash2442
RAGHUPATHI R GRAHAM DI MCINTOSH TK(2000) Apoptosis after traumatic brain injury J Neuro-trauma 17 927ndash938
RIDET JL MALHOTRA SK PRIVAT A and GAGEFH (1997) Reactive astrocytes cellular and molecular cuesto biological function Trends Neurosci 20 570ndash577
SASAKI T LOPES MB HANKINS GR and HELMGA (2002) Expression of survivin an inhibitor of apopto-sis protein in tumors of the nervous system Acta Neu-ropathol (Berl) 104 105ndash109
SHANKAR SL MANI S OrsquoGUIN KN KANDIMALLAER AGRAWAL S and SHAFIT-ZAGARDO B (2001)Survivin inhibition induces human neural tumor cell deaththrough caspase-independent and -dependent pathways JNeurochem 79 426ndash436
SHIN S SUNG BJ CHO YS et al (2001) An anti-apop-totic protein human survivin is a direct inhibitor of caspase-3 and -7 Biochemistry 40 1117ndash1123
SMITH C BERRY M CLARKE WE and LOGAN A(2001) Differential expression of fibroblast growth factorndash2and fibroblast growth factor receptor 1 in a scarring and non-scarring model of CNS injury in the rat Eur J Neurosci 13443ndash456
STONE JR OKONKWO DO SINGLETON RHMUTLU LK HELM GA and POVLISHOCK JT(2002) Caspase-3ndashmediated cleavage of amyloid precursorprotein and formation of amyloid Beta peptide in traumaticaxonal injury J Neurotrauma 19 601ndash614
SYKOVA E VARGOVA L PROKOPOVA S and SI-MONOVA Z (1999) Glial swelling and astrogliosis pro-duce diffusion barriers in the rat spinal cord Glia 25 56ndash70
TAKAHASHI R DEVERAUX Q TAMM I et al (1998)A single BIR domain of XIAP sufficient for inhibiting cas-pases J Biol Chem 273 7787ndash7790
TAMM I WANG Y SAUSVILLE E et al (1998) IAP-family protein survivin inhibits caspase activity and apopto-sis induced by Fas (CD95) Bax caspases and anticancerdrugs Cancer Res 58 5315ndash5320
THOMPSON JD HIGGINS DG and GIBSON TJ (1994)CLUSTAL W improving the sensitivity of progressive mul-tiple sequence alignment through sequence weighting posi-tion-specific gap penalties and weight matrix choice NucleicAcids Res 22 4673ndash4680
JOHNSON ET AL
1194
TOLENTINO PJ DEFORD SM NOTTERPEK L et al(2002) Up-regulation of tissue-type transglutaminase aftertraumatic brain injury J Neurochem 80 579ndash588
UREN AG WONG L PAKUSCH M et al (2000) Sur-vivin and the inner centromere protein INCENP show simi-lar cell-cycle localization and gene knockout phenotypeCurr Biol 10 1319ndash1328
WEI LH HUANG CY CHENG SP CHEN CA andHSIEH CY (2001) Carcinosarcoma of ovary associatedwith previous radiotherapy Int J Gynecol Cancer 11 81ndash84
WU Q COMBS C CANNADY SB GELDMACHERDS and HERRUP K (2000) Beta-amyloid activated mi-croglia induce cell cycling and cell death in cultured corticalneurons Neurobiol Aging 21 797ndash806
YAGITA Y KITAGAWA K OHTSUKI T et al (2001)
Neurogenesis by progenitor cells in the ischemic adult rathippocampus Stroke 32 1890ndash1896
YANG Y GELDMACHER DS and HERRUP K (2001)DNA replication precedes neuronal cell death in Alzheimerrsquosdisease J Neurosci 21 2661ndash2668
Address reprint requests toJose A Pineda PhD
EF and WL McKnight Brain InstituteUniversity of Florida
100 S Newell Dr Bldg 59L1-118 Dept of Pediatrics
Gainesville FL 32610
E-mail pinedjapedsufledu
SURVIVIN UPREGULATION AFTER TBI
1195

PCNA Expression after TBI
For detection of proliferating cell nuclear antigen(PCNA) PVDF membranes immunostained for survivinwere stripped and re-probed using a PCNA-specific an-tibody PCNA (36-kDa protein) was significantly de-tectable in the ipsilateral cortex and hippocampus of TBIrats but only negligible amounts were observed in thecontralateral cortex and hippocampus (Fig 3A) Thetemporal patterns exhibited by PCNA protein were sim-ilar to that of survivin protein Namely PCNA expressedin a time-dependant fashion with a maximum increase at5 days after injury followed by a gradual decline by 14days The levels of PCNA in ipsilateral cortical tissuewere raised over craniotomy control by 919 459 at3 days 2263 333 at 5 days and 1035 356 at 7days post injury (Fig 3B) Similar increases of PCNAprotein in ipsilateral hippocampus were detected at 5 dayspost injury with a maximum of 1006 229 comparedto craniotomy controls (Fig 3C) No significant increasewas found in the contralateral regions when compared tocraniotomy controls (Fig 3A)
Co-Expression of Survivin and PCNA following TBI
To examine spatial co-localization of survivin andPCNA double-label immunohistochemistry of brain tis-sue sections was performed on day 5 post injury whenpeak expression of these proteins was observed
At this time point a remarkable survivin and PCNAimmunoreactivity was found in the ipsilateral cortex (Fig4A) and ipsilateral hippocampus (Fig 4B) consistent withdata obtained using Western blot analyses Within bothregions focal co-expression patterns of survivin andPCNA in single cells were detected which was demon-strated by both separate fluorescent visualization of in-dividual proteins and by merging the images of double-stained slides (Fig 4CndashE) However the dual expressionof survivin and PCNA occurred infrequently as survivinand PCNA immunoreactivity could readily be found sep-arately (Fig 4CndashE) Approximately 12 of the total num-ber of PCNA-positive cells also labeled with survivin Thenuclear morphology of dual survivin and PCNA-positivecells was ambiguous as indicated by DAPI staining (Fig4F) Therefore DAPI staining was simply used for cellidentification in all subsequent experiments
Survivin and PCNA Are Expressed in Astrocytes after TBI
To determine the cell types expressing survivin andPCNA double-label immunohistochemistry for theseproteins and GFAP a marker of astrocytes was per-formed on day 5 post injury
In accordance with Western blot data remarkable sur-vivin-positive immunoreactivity was observed in the ip-silateral cortex and hippocampus proximal to the injurycavity (Fig 5AG green) but not in the contralateral ar-eas (Fig 5BH) Survivin was co-localized with GFAPin the cells of injured cortex and hippocampus whichstrongly suggested primary accumulation of survivin incells of astrocytic lineage (Fig 5CndashEIndashL) Survivin wasuniformly distributed in the cytoplasm and processes ofastrocytes in both cortex and hippocampus (Fig 5DJ)DAPI staining is shown in Figure 5FL3 Approximately88 of the total number of GFAP-positive cells also la-beled with survivin
PCNA-positive immunoreactivity staining was ob-served in the ipsilateral cortex (Fig 6A green) and hip-pocampus (Fig 6G green) of injured brain while con-tralateral cortex and hippocampus exhibited negligiblePCNA immunoreactivity (Fig 6BH) PCNA (Fig 6CI)was partially co-localized with GFAP (Fig 6DJ red) inboth regions and was characteristically distributed in thenucleus of the cells in both cortex and hippocampus (Fig6EK) DAPI staining is shown in Figure 6FL
Taken together double-label immunohistochemistrydata provides evidence that both survivin and PCNA canbe detected in GFAP-positive astrocytes following trau-matic insult Since survivin and PCNA immunoreactiv-ity was not exclusively localized in GFAP-positive cellswe next addressed a question what other cell type mightexpress survivin after TBI We suggested that a certainpopulation of mature neurons might express survivin inresponse to injury
Survivin and PCNA Are Expressed in a Sub-Set of Neurons after TBI
As can be seen in Figure 7 survivin and PCNA wereeach co-expressed with NeuN a marker of mature neu-rons NeuN-positive cells were found to express survivinin the ipsilateral cortex distal to the injury cavity (Fig7AndashD) and in the contralateral hippocampus (Fig 7EndashH)It should be noted however that NeuN-positive cells thatalso expressed survivin occurred infrequently For ex-ample we estimated the number of dual survivinNeuNpositive cells as 01ndash15 of the total number of NeuN-positive cells in these regions Survivin immunoreactiv-ity was negligible in either hemisphere of craniotomycontrol brains (Fig 7QR) No co-localization of survivinand NeuN was observed in ipsilateral hippocampus (datanot shown) As can be seen in Figure 7BF survivin waspredominantly localized to the cytoplasm and axons ofNeuN-positive neurons DAPI staining is shown in Fig-ure 7DH
PCNA-positive neurons were found in the ipsilateralcortex (Fig 7IndashL) and hippocampus after TBI (Fig
JOHNSON ET AL
1190
1191
FIG 7 A sub-set of NeuN-positive neurons express survivin and PCNA after TBI Double fluorescent immunohistochemistryfor survivin (green) and NeuN (red) in the ipsilateral cortex (AB) and the CA1 pyramidal layer of the contralateral hippocam-pus (EF) was performed Survivin is expressed in the cytoplasm and to a limited extent in the processes of NeuN-positive neu-rons (merged images CG) Dual staining for PCNA (green) and NeuN (red) is shown in the ipsilateral cortex (IJ) and the CA1pyramidal layer of the ipsilateral hippocampus (MN) The nuclei are shown using DAPI staining (DH blue) PCNA is expressedin the nucleus of NeuN-positive neurons (merged images KO) PCNA expression was co-incident with DAPI staining in theseexamples (LP blue) White arrows indicate focal co-localization of survivinNeuN and PCNANeuN SurvivinNeuN co-local-ization of survivin (green) and NeuN (red) was seen only in TBI rats as opposed to either hemisphere of craniotomy control(QR) Original magnification 400 bar 20 m (AndashP) original magnification 50 bar 100000 m (QR)
SURVIVIN UPREGULATION AFTER TBI
7MndashP) whereas craniotomy control tissue exhibited onlytrace amounts of PCNA (data not shown) Similar to thesurvivinNeuN co-localization data dual PCNANeuNimmunostaining was a rare event accounting for approx-imately 4 of the total number of NeuN positive cellsPCNA was distributed in the nuclei of these neurons (Fig7KO) although the nuclear morphology of these cellswas not clearly resolved by DAPI staining (Fig 7LP)
DISCUSSION
Traumatic brain injury (TBI) sets into motion variousbiochemical cascades that induce neural tissue injury andcell death Conversely several proteins expressed inneural cells after TBI are directed to resist cell death andpromote recovery in the injured CNS (Ridet et al 1997Chen and Swanson 2003) Survivin is a multi-functionalprotein that inhibits apoptosis and is also required for theproper completion of mitosis Anti-apoptotic and pro-mi-togenic roles for survivin have been documented in pro-liferating cells of neural origin in vitro such as in neu-roblastoma and glioma cells (LaCasse et al 1998 Tammet al 1998 Deveraux and Reed 1999 Conway et al2000 Shin et al 2001 Sasaki et al 2002) Howeverno studies have investigated the potential role of survivinin the adult brain after TBI when a sub-population ofCNS cells may initiate a cell cycle-related process in re-sponse to injury
In the present paper we demonstrate the induction ofsurvivin expression in rat brain subjected to TBI The ex-pression of survivin was time-dependent and cell-spe-cific and was present in astrocytes and to a much lesserextent in neurons in brain cortex and hippocampus In-duction of survivin in these cells was accompanied byoccasional expression of PCNA a cell cycle protein in-volved in mitotic G1S progression Our present data arethe first to show that survivin mRNA and protein are sig-nificantly up-regulated after traumatic brain injury in ratsPCNA expression after TBI has been described previously(Miyake et al 1992 Chen et al 2003) suggesting itsrole in mechanisms of brain recovery after injury Theconcurrent up-regulation of survivin with a similar tem-poral profile as PCNA shown herein further suggests thatsurvivin may play a role in cellular proliferation after TBI
Brain injury evoked the expression of survivin andPCNA in a time-dependent manner (Figs 2 and 3) West-ern blot analysis revealed maximal co-expression of bothsurvivin and PCNA at five days post injury Immuno-histochemistry at this time point demonstrated co-local-ization of these proteins (Fig 4) although most cells werelabeled separately with PCNA and survivin In fact only12 of the total number of PCNA-positive cells were
also survivin positive It has been reported that PCNA isexpressed predominantly in G1S (Bravo et al 1987)while survivin is found at the G2M phase of the cell cy-cle (Bravo et al 1987 Otaki et al 2000) Hence a lackof strict co-localization of survivin and PCNA in ourstudy may be explained by their expression at differentpoints in the cell cycle
In our experimental model we observed survivin- aswell as PCNA-positive astrocytes in the proximal area ofthe injury and in the ipsilateral hippocampus Prolifera-tion of astrocytes is well documented after TBI as shownby cell labeling with BrdU as well as expression of PCNA(Latov et al 1979 Dunn-Meynell and Levin 1997 Car-bonell and Grady 1999 Norton 1999 Csuka et al 2000Kernie et al 2001 Chen et al 2003) Because survivinand PCNA were expressed in astrocytes following TBI(Figs 5 and 6) it is possible that survivin plays an im-portant role linking astrocyte survival and proliferationafter traumatic insult Astrocyte proliferation has beenimplicated in the formation of the glial scar observed af-ter injury (Latov et al 1979) and creates a non-permis-sive environment for repair (Sykova et al 1999) How-ever glial proliferation may also enhance neuronalsurvival (Smith et al 2001 Wei et al 2001)
Of particular interest is a sub-set of NeuN-positive neu-rons found to express survivin only after TBI (Fig 7)These cells were much less abundant than survivin-pos-itive astrocytes and their functional significance is cur-rently unknown However both neurons and astrocyteshave been documented previously to express cell cycleproteins after various insults such as exposure to -amy-loidndashactivated microglia (Wu et al 2000) TBI (Kaya etal 1999ab) chlorine toxicity (Magavi et al 2000) oras a consequence of Alzheimerrsquos disease (Yang et al2001) These papers underscore the significant contro-versy that exists regarding the function of cell cycle pro-teins such as PCNA in neurons after different types ofinjury We are currently conducting further studies whichwill elucidate the roles for PCNA and survivin in neu-rons after TBI
It should be noted that dual staining of survivin andPCNA could not be directly attributed to a specific celltype due to the technical difficulties of triple labeling an-tibody-based IHC Therefore we cannot rule out the pos-sibility that other cell types such as endothelial (Conwayet al 2003) or inflammatory cells (Hill-Felberg et al1999) may also contribute to survivin and PCNA ex-pression after TBI The appearance of survivin and PCNAseparately in neurons (NeuN-positive) and astrocytes(GFAP-positive) along with co-localization of survivinwith PCNA in the same cells provide correlative data tosuggest an activation of cell cycle-like program in astro-cytes and possibly in a small subtype of neurons after
JOHNSON ET AL
1192
TBI In our experiments survivin co-localization withPCNA does suggest that survivin may be associated witha pro-mitotic process In an attempt to clarify these pro-teinrsquos roles after TBI we analyzed the nuclear morphol-ogy of survivin-positive cells to define the apoptotic ormitotic architecture of nuclei DAPI staining proved tooambiguous in identifying apoptotic versus mitotic phe-notypes likely due to the thickness of the brain sections(40 m) Further studies using direct markers of mitosissuch as BrdU incorporation as well as simultaneous la-beling with cell death related proteins is required to de-lineate anti-apoptotic and pro-mitotic activities of sur-vivin and PCNA in these cells
In conclusion our data demonstrate the induction ofsurvivin in the rat brain following TBI Expression of sur-vivin occurred predominantly in astrocytes as comparedto neurons in a time-dependant fashion and was accom-panied by expression of PCNA Taken together these re-sults suggest that survivin plays a role in neural cell re-sponses following traumatic brain injury in rats Futurestudies will investigate the implications of these findingsto the pathophysiology of TBI
ACKNOWLEDGMENTS
We thank Barbara OrsquoSteen and Tao Fan MD fortechnical assistance This work was funded by grants NIHRO1 NS39091 NIH RO1 NS40182 DAMD 17-99-1-9565 and DAMD 17-01-1-0765
REFERENCES
ADIDA C CROTTY PL MCGRATH J BERREBI DDIEBOLD J and ALTIERI DC (1998) Developmentallyregulated expression of the novel cancer anti-apoptosis genesurvivin in human and mouse differentiation Am J Pathol152 43ndash49
ALTIERI DC MARCHISIO PC and MARCHISIO C(1999) Survivin apoptosis an interloper between cell deathand cell proliferation in cancer Lab Invest 79 1327ndash1333
ALTSCHUL SF MADDEN TL SCHAFFER AA et al(1997) Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 253389ndash3402
AMBROSINI G ADIDA C and ALTIERI DC (1997) Anovel anti-apoptosis gene survivin expressed in cancer andlymphoma Nat Med 3 917ndash921
BRAVO R FRANK R BLUNDELL PA and MAC-DONALD-BRAVO H (1987) CyclinPCNA is the auxil-iary protein of DNA polymerase-delta Nature 326 515ndash517
BUSH TG PUVANACHANDRA N HORNER CH et al
(1999) Leukocyte infiltration neuronal degeneration andneurite outgrowth after ablation of scar-forming reactive as-trocytes in adult transgenic mice Neuron 23 297ndash308
CAMERON HA and MCKAY RD (2001) Adult neuroge-nesis produces a large pool of new granule cells in the den-tate gyrus J Comp Neurol 435 406ndash417
CARBONELL WS and GRADY MS (1999) Regional andtemporal characterization of neuronal glial and axonal re-sponse after traumatic brain injury in the mouse Acta Neu-ropathol (Berl) 98 396ndash406
CHEN XH IWATA A NONAKA M BROWNE KDand SMITH DH (2003) Neurogenesis and glial prolifera-tion persist for at least one year in the subventricular zonefollowing brain trauma in rats J Neurotrauma 20 623ndash631
CHEN Y and SWANSON RA (2003) Astrocytes and braininjury J Cereb Blood Flow Metab 23 137ndash149
CONWAY EM POLLEFEYT S CORNELISSEN J et al(2000) Three differentially expressed survivin cDNA vari-ants encode proteins with distinct antiapoptotic functionsBlood 95 1435ndash1442
CSUKA E HANS VH AMMANN E TRENTZ OKOSSMANN T and MORGANTI-KOSSMANN MC(2000) Cell activation and inflammatory response followingtraumatic axonal injury in the rat Neuroreport 112587ndash2590
DEVERAUX QL and REED JC (1999) IAP family pro-teinsmdashsuppressors of apoptosis Genes Dev 13 239ndash252
DIXON CE CLIFTON GL LIGHTHALL JW YAGH-MAI AA and HAYES RL (1991) A controlled corticalimpact model of traumatic brain injury in the rat J NeurosciMethods 39 253ndash262
DOETSCH F CAILLE I LIM DA GARCIA-VERDUGOJM and ALVAREZ-BUYLLA A (1999) Subventricularzone astrocytes are neural stem cells in the adult mammalianbrain Cell 97 703ndash716
DUNN-MEYNELL AA and LEVIN BE (1997) Histolog-ical markers of neuronal axonal and astrocytic changes af-ter lateral rigid impact traumatic brain injury Brain Res 76125ndash41
GIULIAN D (1991) Microgliamdashneuron interactions after in-jury to the central nervous system in Peripheral Signalingof the Brain Role in Neural-Immune Interactions Learningand Memory R Frederickson J McGaugh and D Felton(eds) Lewiston NY Hogrefe and Huber pps 73ndash82
HIGGINS DG BLEASBY AJ and FUCHS R (1992)CLUSTAL V improved software for multiple sequencealignment Comput Appl Biosci 8 189ndash191
HILL-FELBERG SJ MCINTOSH TK OLIVER DLRAGHUPATHI R and BARBARESE E (1999) Concur-rent loss and proliferation of astrocytes following lateral fluidpercussion brain injury in the adult rat J Neurosci Res 57271ndash279
1193
SURVIVIN UPREGULATION AFTER TBI
JIANG X WILFORD C DUENSING S MUNGER KJONES G and JONES D (2001) Participation of Survivinin mitotic and apoptotic activities of normal and tumor-de-rived cells J Cell Biochem 83 342ndash354
KAYA SS MAHMOOD A LI Y YAVUZ E andCHOPP M (1999a) Expression of cell cycle proteins (cy-clin D1 and cdk4) after controlled cortical impact in rat brainJ Neurotrauma 16 1187ndash1196
KAYA SS MAHMOOD A LI Y YAVUZ E GOKSELM and CHOPP M (1999b) Apoptosis and expression ofp53 response proteins and cyclin D1 after cortical impact inrat brain Brain Res 818 23ndash33
KERNIE SG ERWIN TM and PARADA LF (2001)Brain remodeling due to neuronal and astrocytic prolifera-tion after controlled cortical injury in mice J Neurosci Res66 317ndash326
KOBAYASHI K HATANO M OTAKI M OGA-SAWARA T and TOKUHISA T (1999) Expression of amurine homologue of the inhibitor of apoptosis protein is re-lated to cell proliferation Proc Natl Acad Sci USA 961457ndash1462
LACASSE EC BAIRD S KORNELUK RG andMACKENZIE AE (1998) The inhibitors of apoptosis(IAPs) and their emerging role in cancer Oncogene 173247ndash3259
LATOV N NILAVER G ZIMMERMAN EA et al(1979) Fibrillary astrocytes proliferate in response to braininjury a study combining immunoperoxidase technique forglial fibrillary acidic protein and radioautography of tritiatedthymidine Dev Biol 72 381ndash384
LI F and ALTIERI DC (1999) The cancer antiapoptosismouse survivin gene characterization of locus and tran-scriptional requirements of basal and cell cycle-dependentexpression Cancer Res 59 3143ndash3151
LI F AMBROSINI G CHU EY et al (1998) Control ofapoptosis and mitotic spindle checkpoint by survivin Nature396 580ndash584
LI F ACKERMANN EJ BENNETT CF et al (1999)Pleiotropic cell-division defects and apoptosis induced by in-terference with survivin function Nat Cell Biol 1 461ndash466
MAGAVI SS LEAVITT BR and MACKLIS JD (2000)Induction of neurogenesis in the neocortex of adult mice Na-ture 405 951ndash955
MIYAKE T OKADA M and KITAMURA T (1992) Re-active proliferation of astrocytes studied by immunohisto-chemistry for proliferating cell nuclear antigen Brain Res590 300ndash302
MUCHMORE SW CHEN J JAKOB C et al (2000)Crystal structure and mutagenic analysis of the inhibitor-of-apoptosis protein survivin Mol Cell 6 173ndash182
NORTON WT (1999) Cell reactions following acute braininjury a review Neurochem Res 24 213ndash218
OTAKI M HATANO M KOBAYASHI K et al (2000)Cell cyclendashdependent regulation of TIAPm-survivin expres-sion Biochim Biophys Acta 1493 188ndash194
PETERSON DA (2002) Stem cells in brain plasticity and re-pair Curr Opin Pharmacol 2 34ndash42
PIKE BR ZHAO X NEWCOMB JK POSMANTURRM WANG KK and Hayes RL (1998) Regional cal-pain and caspase-3 proteolysis of alpha-spectrin after trau-matic brain injury Neuroreport 9 2437ndash2442
RAGHUPATHI R GRAHAM DI MCINTOSH TK(2000) Apoptosis after traumatic brain injury J Neuro-trauma 17 927ndash938
RIDET JL MALHOTRA SK PRIVAT A and GAGEFH (1997) Reactive astrocytes cellular and molecular cuesto biological function Trends Neurosci 20 570ndash577
SASAKI T LOPES MB HANKINS GR and HELMGA (2002) Expression of survivin an inhibitor of apopto-sis protein in tumors of the nervous system Acta Neu-ropathol (Berl) 104 105ndash109
SHANKAR SL MANI S OrsquoGUIN KN KANDIMALLAER AGRAWAL S and SHAFIT-ZAGARDO B (2001)Survivin inhibition induces human neural tumor cell deaththrough caspase-independent and -dependent pathways JNeurochem 79 426ndash436
SHIN S SUNG BJ CHO YS et al (2001) An anti-apop-totic protein human survivin is a direct inhibitor of caspase-3 and -7 Biochemistry 40 1117ndash1123
SMITH C BERRY M CLARKE WE and LOGAN A(2001) Differential expression of fibroblast growth factorndash2and fibroblast growth factor receptor 1 in a scarring and non-scarring model of CNS injury in the rat Eur J Neurosci 13443ndash456
STONE JR OKONKWO DO SINGLETON RHMUTLU LK HELM GA and POVLISHOCK JT(2002) Caspase-3ndashmediated cleavage of amyloid precursorprotein and formation of amyloid Beta peptide in traumaticaxonal injury J Neurotrauma 19 601ndash614
SYKOVA E VARGOVA L PROKOPOVA S and SI-MONOVA Z (1999) Glial swelling and astrogliosis pro-duce diffusion barriers in the rat spinal cord Glia 25 56ndash70
TAKAHASHI R DEVERAUX Q TAMM I et al (1998)A single BIR domain of XIAP sufficient for inhibiting cas-pases J Biol Chem 273 7787ndash7790
TAMM I WANG Y SAUSVILLE E et al (1998) IAP-family protein survivin inhibits caspase activity and apopto-sis induced by Fas (CD95) Bax caspases and anticancerdrugs Cancer Res 58 5315ndash5320
THOMPSON JD HIGGINS DG and GIBSON TJ (1994)CLUSTAL W improving the sensitivity of progressive mul-tiple sequence alignment through sequence weighting posi-tion-specific gap penalties and weight matrix choice NucleicAcids Res 22 4673ndash4680
JOHNSON ET AL
1194
TOLENTINO PJ DEFORD SM NOTTERPEK L et al(2002) Up-regulation of tissue-type transglutaminase aftertraumatic brain injury J Neurochem 80 579ndash588
UREN AG WONG L PAKUSCH M et al (2000) Sur-vivin and the inner centromere protein INCENP show simi-lar cell-cycle localization and gene knockout phenotypeCurr Biol 10 1319ndash1328
WEI LH HUANG CY CHENG SP CHEN CA andHSIEH CY (2001) Carcinosarcoma of ovary associatedwith previous radiotherapy Int J Gynecol Cancer 11 81ndash84
WU Q COMBS C CANNADY SB GELDMACHERDS and HERRUP K (2000) Beta-amyloid activated mi-croglia induce cell cycling and cell death in cultured corticalneurons Neurobiol Aging 21 797ndash806
YAGITA Y KITAGAWA K OHTSUKI T et al (2001)
Neurogenesis by progenitor cells in the ischemic adult rathippocampus Stroke 32 1890ndash1896
YANG Y GELDMACHER DS and HERRUP K (2001)DNA replication precedes neuronal cell death in Alzheimerrsquosdisease J Neurosci 21 2661ndash2668
Address reprint requests toJose A Pineda PhD
EF and WL McKnight Brain InstituteUniversity of Florida
100 S Newell Dr Bldg 59L1-118 Dept of Pediatrics
Gainesville FL 32610
E-mail pinedjapedsufledu
SURVIVIN UPREGULATION AFTER TBI
1195
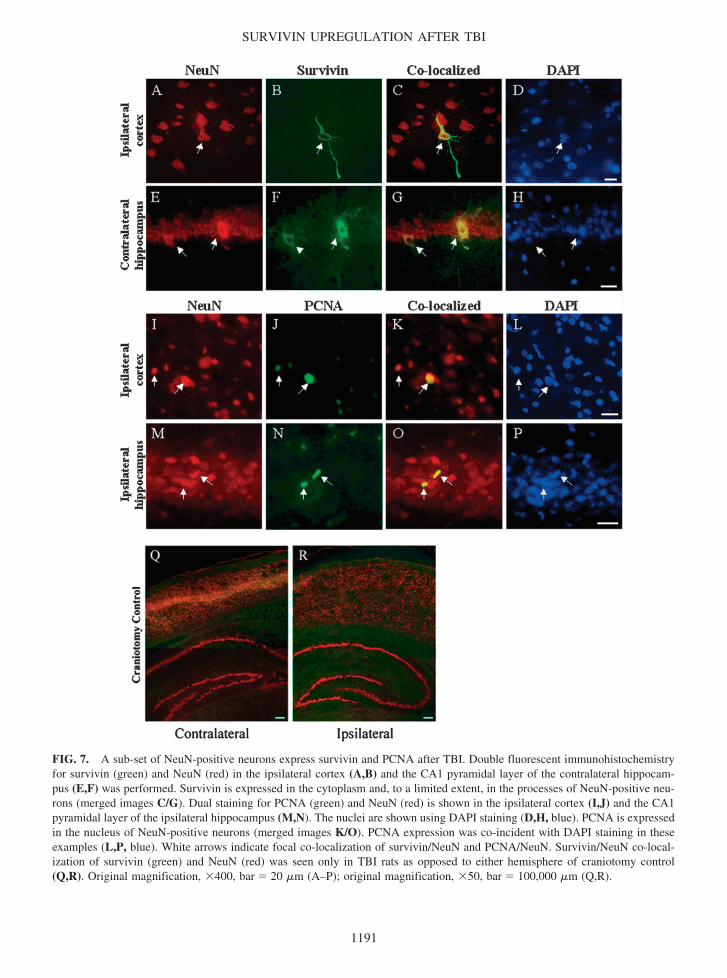
1191
FIG 7 A sub-set of NeuN-positive neurons express survivin and PCNA after TBI Double fluorescent immunohistochemistryfor survivin (green) and NeuN (red) in the ipsilateral cortex (AB) and the CA1 pyramidal layer of the contralateral hippocam-pus (EF) was performed Survivin is expressed in the cytoplasm and to a limited extent in the processes of NeuN-positive neu-rons (merged images CG) Dual staining for PCNA (green) and NeuN (red) is shown in the ipsilateral cortex (IJ) and the CA1pyramidal layer of the ipsilateral hippocampus (MN) The nuclei are shown using DAPI staining (DH blue) PCNA is expressedin the nucleus of NeuN-positive neurons (merged images KO) PCNA expression was co-incident with DAPI staining in theseexamples (LP blue) White arrows indicate focal co-localization of survivinNeuN and PCNANeuN SurvivinNeuN co-local-ization of survivin (green) and NeuN (red) was seen only in TBI rats as opposed to either hemisphere of craniotomy control(QR) Original magnification 400 bar 20 m (AndashP) original magnification 50 bar 100000 m (QR)
SURVIVIN UPREGULATION AFTER TBI
7MndashP) whereas craniotomy control tissue exhibited onlytrace amounts of PCNA (data not shown) Similar to thesurvivinNeuN co-localization data dual PCNANeuNimmunostaining was a rare event accounting for approx-imately 4 of the total number of NeuN positive cellsPCNA was distributed in the nuclei of these neurons (Fig7KO) although the nuclear morphology of these cellswas not clearly resolved by DAPI staining (Fig 7LP)
DISCUSSION
Traumatic brain injury (TBI) sets into motion variousbiochemical cascades that induce neural tissue injury andcell death Conversely several proteins expressed inneural cells after TBI are directed to resist cell death andpromote recovery in the injured CNS (Ridet et al 1997Chen and Swanson 2003) Survivin is a multi-functionalprotein that inhibits apoptosis and is also required for theproper completion of mitosis Anti-apoptotic and pro-mi-togenic roles for survivin have been documented in pro-liferating cells of neural origin in vitro such as in neu-roblastoma and glioma cells (LaCasse et al 1998 Tammet al 1998 Deveraux and Reed 1999 Conway et al2000 Shin et al 2001 Sasaki et al 2002) Howeverno studies have investigated the potential role of survivinin the adult brain after TBI when a sub-population ofCNS cells may initiate a cell cycle-related process in re-sponse to injury
In the present paper we demonstrate the induction ofsurvivin expression in rat brain subjected to TBI The ex-pression of survivin was time-dependent and cell-spe-cific and was present in astrocytes and to a much lesserextent in neurons in brain cortex and hippocampus In-duction of survivin in these cells was accompanied byoccasional expression of PCNA a cell cycle protein in-volved in mitotic G1S progression Our present data arethe first to show that survivin mRNA and protein are sig-nificantly up-regulated after traumatic brain injury in ratsPCNA expression after TBI has been described previously(Miyake et al 1992 Chen et al 2003) suggesting itsrole in mechanisms of brain recovery after injury Theconcurrent up-regulation of survivin with a similar tem-poral profile as PCNA shown herein further suggests thatsurvivin may play a role in cellular proliferation after TBI
Brain injury evoked the expression of survivin andPCNA in a time-dependent manner (Figs 2 and 3) West-ern blot analysis revealed maximal co-expression of bothsurvivin and PCNA at five days post injury Immuno-histochemistry at this time point demonstrated co-local-ization of these proteins (Fig 4) although most cells werelabeled separately with PCNA and survivin In fact only12 of the total number of PCNA-positive cells were
also survivin positive It has been reported that PCNA isexpressed predominantly in G1S (Bravo et al 1987)while survivin is found at the G2M phase of the cell cy-cle (Bravo et al 1987 Otaki et al 2000) Hence a lackof strict co-localization of survivin and PCNA in ourstudy may be explained by their expression at differentpoints in the cell cycle
In our experimental model we observed survivin- aswell as PCNA-positive astrocytes in the proximal area ofthe injury and in the ipsilateral hippocampus Prolifera-tion of astrocytes is well documented after TBI as shownby cell labeling with BrdU as well as expression of PCNA(Latov et al 1979 Dunn-Meynell and Levin 1997 Car-bonell and Grady 1999 Norton 1999 Csuka et al 2000Kernie et al 2001 Chen et al 2003) Because survivinand PCNA were expressed in astrocytes following TBI(Figs 5 and 6) it is possible that survivin plays an im-portant role linking astrocyte survival and proliferationafter traumatic insult Astrocyte proliferation has beenimplicated in the formation of the glial scar observed af-ter injury (Latov et al 1979) and creates a non-permis-sive environment for repair (Sykova et al 1999) How-ever glial proliferation may also enhance neuronalsurvival (Smith et al 2001 Wei et al 2001)
Of particular interest is a sub-set of NeuN-positive neu-rons found to express survivin only after TBI (Fig 7)These cells were much less abundant than survivin-pos-itive astrocytes and their functional significance is cur-rently unknown However both neurons and astrocyteshave been documented previously to express cell cycleproteins after various insults such as exposure to -amy-loidndashactivated microglia (Wu et al 2000) TBI (Kaya etal 1999ab) chlorine toxicity (Magavi et al 2000) oras a consequence of Alzheimerrsquos disease (Yang et al2001) These papers underscore the significant contro-versy that exists regarding the function of cell cycle pro-teins such as PCNA in neurons after different types ofinjury We are currently conducting further studies whichwill elucidate the roles for PCNA and survivin in neu-rons after TBI
It should be noted that dual staining of survivin andPCNA could not be directly attributed to a specific celltype due to the technical difficulties of triple labeling an-tibody-based IHC Therefore we cannot rule out the pos-sibility that other cell types such as endothelial (Conwayet al 2003) or inflammatory cells (Hill-Felberg et al1999) may also contribute to survivin and PCNA ex-pression after TBI The appearance of survivin and PCNAseparately in neurons (NeuN-positive) and astrocytes(GFAP-positive) along with co-localization of survivinwith PCNA in the same cells provide correlative data tosuggest an activation of cell cycle-like program in astro-cytes and possibly in a small subtype of neurons after
JOHNSON ET AL
1192
TBI In our experiments survivin co-localization withPCNA does suggest that survivin may be associated witha pro-mitotic process In an attempt to clarify these pro-teinrsquos roles after TBI we analyzed the nuclear morphol-ogy of survivin-positive cells to define the apoptotic ormitotic architecture of nuclei DAPI staining proved tooambiguous in identifying apoptotic versus mitotic phe-notypes likely due to the thickness of the brain sections(40 m) Further studies using direct markers of mitosissuch as BrdU incorporation as well as simultaneous la-beling with cell death related proteins is required to de-lineate anti-apoptotic and pro-mitotic activities of sur-vivin and PCNA in these cells
In conclusion our data demonstrate the induction ofsurvivin in the rat brain following TBI Expression of sur-vivin occurred predominantly in astrocytes as comparedto neurons in a time-dependant fashion and was accom-panied by expression of PCNA Taken together these re-sults suggest that survivin plays a role in neural cell re-sponses following traumatic brain injury in rats Futurestudies will investigate the implications of these findingsto the pathophysiology of TBI
ACKNOWLEDGMENTS
We thank Barbara OrsquoSteen and Tao Fan MD fortechnical assistance This work was funded by grants NIHRO1 NS39091 NIH RO1 NS40182 DAMD 17-99-1-9565 and DAMD 17-01-1-0765
REFERENCES
ADIDA C CROTTY PL MCGRATH J BERREBI DDIEBOLD J and ALTIERI DC (1998) Developmentallyregulated expression of the novel cancer anti-apoptosis genesurvivin in human and mouse differentiation Am J Pathol152 43ndash49
ALTIERI DC MARCHISIO PC and MARCHISIO C(1999) Survivin apoptosis an interloper between cell deathand cell proliferation in cancer Lab Invest 79 1327ndash1333
ALTSCHUL SF MADDEN TL SCHAFFER AA et al(1997) Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 253389ndash3402
AMBROSINI G ADIDA C and ALTIERI DC (1997) Anovel anti-apoptosis gene survivin expressed in cancer andlymphoma Nat Med 3 917ndash921
BRAVO R FRANK R BLUNDELL PA and MAC-DONALD-BRAVO H (1987) CyclinPCNA is the auxil-iary protein of DNA polymerase-delta Nature 326 515ndash517
BUSH TG PUVANACHANDRA N HORNER CH et al
(1999) Leukocyte infiltration neuronal degeneration andneurite outgrowth after ablation of scar-forming reactive as-trocytes in adult transgenic mice Neuron 23 297ndash308
CAMERON HA and MCKAY RD (2001) Adult neuroge-nesis produces a large pool of new granule cells in the den-tate gyrus J Comp Neurol 435 406ndash417
CARBONELL WS and GRADY MS (1999) Regional andtemporal characterization of neuronal glial and axonal re-sponse after traumatic brain injury in the mouse Acta Neu-ropathol (Berl) 98 396ndash406
CHEN XH IWATA A NONAKA M BROWNE KDand SMITH DH (2003) Neurogenesis and glial prolifera-tion persist for at least one year in the subventricular zonefollowing brain trauma in rats J Neurotrauma 20 623ndash631
CHEN Y and SWANSON RA (2003) Astrocytes and braininjury J Cereb Blood Flow Metab 23 137ndash149
CONWAY EM POLLEFEYT S CORNELISSEN J et al(2000) Three differentially expressed survivin cDNA vari-ants encode proteins with distinct antiapoptotic functionsBlood 95 1435ndash1442
CSUKA E HANS VH AMMANN E TRENTZ OKOSSMANN T and MORGANTI-KOSSMANN MC(2000) Cell activation and inflammatory response followingtraumatic axonal injury in the rat Neuroreport 112587ndash2590
DEVERAUX QL and REED JC (1999) IAP family pro-teinsmdashsuppressors of apoptosis Genes Dev 13 239ndash252
DIXON CE CLIFTON GL LIGHTHALL JW YAGH-MAI AA and HAYES RL (1991) A controlled corticalimpact model of traumatic brain injury in the rat J NeurosciMethods 39 253ndash262
DOETSCH F CAILLE I LIM DA GARCIA-VERDUGOJM and ALVAREZ-BUYLLA A (1999) Subventricularzone astrocytes are neural stem cells in the adult mammalianbrain Cell 97 703ndash716
DUNN-MEYNELL AA and LEVIN BE (1997) Histolog-ical markers of neuronal axonal and astrocytic changes af-ter lateral rigid impact traumatic brain injury Brain Res 76125ndash41
GIULIAN D (1991) Microgliamdashneuron interactions after in-jury to the central nervous system in Peripheral Signalingof the Brain Role in Neural-Immune Interactions Learningand Memory R Frederickson J McGaugh and D Felton(eds) Lewiston NY Hogrefe and Huber pps 73ndash82
HIGGINS DG BLEASBY AJ and FUCHS R (1992)CLUSTAL V improved software for multiple sequencealignment Comput Appl Biosci 8 189ndash191
HILL-FELBERG SJ MCINTOSH TK OLIVER DLRAGHUPATHI R and BARBARESE E (1999) Concur-rent loss and proliferation of astrocytes following lateral fluidpercussion brain injury in the adult rat J Neurosci Res 57271ndash279
1193
SURVIVIN UPREGULATION AFTER TBI
JIANG X WILFORD C DUENSING S MUNGER KJONES G and JONES D (2001) Participation of Survivinin mitotic and apoptotic activities of normal and tumor-de-rived cells J Cell Biochem 83 342ndash354
KAYA SS MAHMOOD A LI Y YAVUZ E andCHOPP M (1999a) Expression of cell cycle proteins (cy-clin D1 and cdk4) after controlled cortical impact in rat brainJ Neurotrauma 16 1187ndash1196
KAYA SS MAHMOOD A LI Y YAVUZ E GOKSELM and CHOPP M (1999b) Apoptosis and expression ofp53 response proteins and cyclin D1 after cortical impact inrat brain Brain Res 818 23ndash33
KERNIE SG ERWIN TM and PARADA LF (2001)Brain remodeling due to neuronal and astrocytic prolifera-tion after controlled cortical injury in mice J Neurosci Res66 317ndash326
KOBAYASHI K HATANO M OTAKI M OGA-SAWARA T and TOKUHISA T (1999) Expression of amurine homologue of the inhibitor of apoptosis protein is re-lated to cell proliferation Proc Natl Acad Sci USA 961457ndash1462
LACASSE EC BAIRD S KORNELUK RG andMACKENZIE AE (1998) The inhibitors of apoptosis(IAPs) and their emerging role in cancer Oncogene 173247ndash3259
LATOV N NILAVER G ZIMMERMAN EA et al(1979) Fibrillary astrocytes proliferate in response to braininjury a study combining immunoperoxidase technique forglial fibrillary acidic protein and radioautography of tritiatedthymidine Dev Biol 72 381ndash384
LI F and ALTIERI DC (1999) The cancer antiapoptosismouse survivin gene characterization of locus and tran-scriptional requirements of basal and cell cycle-dependentexpression Cancer Res 59 3143ndash3151
LI F AMBROSINI G CHU EY et al (1998) Control ofapoptosis and mitotic spindle checkpoint by survivin Nature396 580ndash584
LI F ACKERMANN EJ BENNETT CF et al (1999)Pleiotropic cell-division defects and apoptosis induced by in-terference with survivin function Nat Cell Biol 1 461ndash466
MAGAVI SS LEAVITT BR and MACKLIS JD (2000)Induction of neurogenesis in the neocortex of adult mice Na-ture 405 951ndash955
MIYAKE T OKADA M and KITAMURA T (1992) Re-active proliferation of astrocytes studied by immunohisto-chemistry for proliferating cell nuclear antigen Brain Res590 300ndash302
MUCHMORE SW CHEN J JAKOB C et al (2000)Crystal structure and mutagenic analysis of the inhibitor-of-apoptosis protein survivin Mol Cell 6 173ndash182
NORTON WT (1999) Cell reactions following acute braininjury a review Neurochem Res 24 213ndash218
OTAKI M HATANO M KOBAYASHI K et al (2000)Cell cyclendashdependent regulation of TIAPm-survivin expres-sion Biochim Biophys Acta 1493 188ndash194
PETERSON DA (2002) Stem cells in brain plasticity and re-pair Curr Opin Pharmacol 2 34ndash42
PIKE BR ZHAO X NEWCOMB JK POSMANTURRM WANG KK and Hayes RL (1998) Regional cal-pain and caspase-3 proteolysis of alpha-spectrin after trau-matic brain injury Neuroreport 9 2437ndash2442
RAGHUPATHI R GRAHAM DI MCINTOSH TK(2000) Apoptosis after traumatic brain injury J Neuro-trauma 17 927ndash938
RIDET JL MALHOTRA SK PRIVAT A and GAGEFH (1997) Reactive astrocytes cellular and molecular cuesto biological function Trends Neurosci 20 570ndash577
SASAKI T LOPES MB HANKINS GR and HELMGA (2002) Expression of survivin an inhibitor of apopto-sis protein in tumors of the nervous system Acta Neu-ropathol (Berl) 104 105ndash109
SHANKAR SL MANI S OrsquoGUIN KN KANDIMALLAER AGRAWAL S and SHAFIT-ZAGARDO B (2001)Survivin inhibition induces human neural tumor cell deaththrough caspase-independent and -dependent pathways JNeurochem 79 426ndash436
SHIN S SUNG BJ CHO YS et al (2001) An anti-apop-totic protein human survivin is a direct inhibitor of caspase-3 and -7 Biochemistry 40 1117ndash1123
SMITH C BERRY M CLARKE WE and LOGAN A(2001) Differential expression of fibroblast growth factorndash2and fibroblast growth factor receptor 1 in a scarring and non-scarring model of CNS injury in the rat Eur J Neurosci 13443ndash456
STONE JR OKONKWO DO SINGLETON RHMUTLU LK HELM GA and POVLISHOCK JT(2002) Caspase-3ndashmediated cleavage of amyloid precursorprotein and formation of amyloid Beta peptide in traumaticaxonal injury J Neurotrauma 19 601ndash614
SYKOVA E VARGOVA L PROKOPOVA S and SI-MONOVA Z (1999) Glial swelling and astrogliosis pro-duce diffusion barriers in the rat spinal cord Glia 25 56ndash70
TAKAHASHI R DEVERAUX Q TAMM I et al (1998)A single BIR domain of XIAP sufficient for inhibiting cas-pases J Biol Chem 273 7787ndash7790
TAMM I WANG Y SAUSVILLE E et al (1998) IAP-family protein survivin inhibits caspase activity and apopto-sis induced by Fas (CD95) Bax caspases and anticancerdrugs Cancer Res 58 5315ndash5320
THOMPSON JD HIGGINS DG and GIBSON TJ (1994)CLUSTAL W improving the sensitivity of progressive mul-tiple sequence alignment through sequence weighting posi-tion-specific gap penalties and weight matrix choice NucleicAcids Res 22 4673ndash4680
JOHNSON ET AL
1194
TOLENTINO PJ DEFORD SM NOTTERPEK L et al(2002) Up-regulation of tissue-type transglutaminase aftertraumatic brain injury J Neurochem 80 579ndash588
UREN AG WONG L PAKUSCH M et al (2000) Sur-vivin and the inner centromere protein INCENP show simi-lar cell-cycle localization and gene knockout phenotypeCurr Biol 10 1319ndash1328
WEI LH HUANG CY CHENG SP CHEN CA andHSIEH CY (2001) Carcinosarcoma of ovary associatedwith previous radiotherapy Int J Gynecol Cancer 11 81ndash84
WU Q COMBS C CANNADY SB GELDMACHERDS and HERRUP K (2000) Beta-amyloid activated mi-croglia induce cell cycling and cell death in cultured corticalneurons Neurobiol Aging 21 797ndash806
YAGITA Y KITAGAWA K OHTSUKI T et al (2001)
Neurogenesis by progenitor cells in the ischemic adult rathippocampus Stroke 32 1890ndash1896
YANG Y GELDMACHER DS and HERRUP K (2001)DNA replication precedes neuronal cell death in Alzheimerrsquosdisease J Neurosci 21 2661ndash2668
Address reprint requests toJose A Pineda PhD
EF and WL McKnight Brain InstituteUniversity of Florida
100 S Newell Dr Bldg 59L1-118 Dept of Pediatrics
Gainesville FL 32610
E-mail pinedjapedsufledu
SURVIVIN UPREGULATION AFTER TBI
1195

7MndashP) whereas craniotomy control tissue exhibited onlytrace amounts of PCNA (data not shown) Similar to thesurvivinNeuN co-localization data dual PCNANeuNimmunostaining was a rare event accounting for approx-imately 4 of the total number of NeuN positive cellsPCNA was distributed in the nuclei of these neurons (Fig7KO) although the nuclear morphology of these cellswas not clearly resolved by DAPI staining (Fig 7LP)
DISCUSSION
Traumatic brain injury (TBI) sets into motion variousbiochemical cascades that induce neural tissue injury andcell death Conversely several proteins expressed inneural cells after TBI are directed to resist cell death andpromote recovery in the injured CNS (Ridet et al 1997Chen and Swanson 2003) Survivin is a multi-functionalprotein that inhibits apoptosis and is also required for theproper completion of mitosis Anti-apoptotic and pro-mi-togenic roles for survivin have been documented in pro-liferating cells of neural origin in vitro such as in neu-roblastoma and glioma cells (LaCasse et al 1998 Tammet al 1998 Deveraux and Reed 1999 Conway et al2000 Shin et al 2001 Sasaki et al 2002) Howeverno studies have investigated the potential role of survivinin the adult brain after TBI when a sub-population ofCNS cells may initiate a cell cycle-related process in re-sponse to injury
In the present paper we demonstrate the induction ofsurvivin expression in rat brain subjected to TBI The ex-pression of survivin was time-dependent and cell-spe-cific and was present in astrocytes and to a much lesserextent in neurons in brain cortex and hippocampus In-duction of survivin in these cells was accompanied byoccasional expression of PCNA a cell cycle protein in-volved in mitotic G1S progression Our present data arethe first to show that survivin mRNA and protein are sig-nificantly up-regulated after traumatic brain injury in ratsPCNA expression after TBI has been described previously(Miyake et al 1992 Chen et al 2003) suggesting itsrole in mechanisms of brain recovery after injury Theconcurrent up-regulation of survivin with a similar tem-poral profile as PCNA shown herein further suggests thatsurvivin may play a role in cellular proliferation after TBI
Brain injury evoked the expression of survivin andPCNA in a time-dependent manner (Figs 2 and 3) West-ern blot analysis revealed maximal co-expression of bothsurvivin and PCNA at five days post injury Immuno-histochemistry at this time point demonstrated co-local-ization of these proteins (Fig 4) although most cells werelabeled separately with PCNA and survivin In fact only12 of the total number of PCNA-positive cells were
also survivin positive It has been reported that PCNA isexpressed predominantly in G1S (Bravo et al 1987)while survivin is found at the G2M phase of the cell cy-cle (Bravo et al 1987 Otaki et al 2000) Hence a lackof strict co-localization of survivin and PCNA in ourstudy may be explained by their expression at differentpoints in the cell cycle
In our experimental model we observed survivin- aswell as PCNA-positive astrocytes in the proximal area ofthe injury and in the ipsilateral hippocampus Prolifera-tion of astrocytes is well documented after TBI as shownby cell labeling with BrdU as well as expression of PCNA(Latov et al 1979 Dunn-Meynell and Levin 1997 Car-bonell and Grady 1999 Norton 1999 Csuka et al 2000Kernie et al 2001 Chen et al 2003) Because survivinand PCNA were expressed in astrocytes following TBI(Figs 5 and 6) it is possible that survivin plays an im-portant role linking astrocyte survival and proliferationafter traumatic insult Astrocyte proliferation has beenimplicated in the formation of the glial scar observed af-ter injury (Latov et al 1979) and creates a non-permis-sive environment for repair (Sykova et al 1999) How-ever glial proliferation may also enhance neuronalsurvival (Smith et al 2001 Wei et al 2001)
Of particular interest is a sub-set of NeuN-positive neu-rons found to express survivin only after TBI (Fig 7)These cells were much less abundant than survivin-pos-itive astrocytes and their functional significance is cur-rently unknown However both neurons and astrocyteshave been documented previously to express cell cycleproteins after various insults such as exposure to -amy-loidndashactivated microglia (Wu et al 2000) TBI (Kaya etal 1999ab) chlorine toxicity (Magavi et al 2000) oras a consequence of Alzheimerrsquos disease (Yang et al2001) These papers underscore the significant contro-versy that exists regarding the function of cell cycle pro-teins such as PCNA in neurons after different types ofinjury We are currently conducting further studies whichwill elucidate the roles for PCNA and survivin in neu-rons after TBI
It should be noted that dual staining of survivin andPCNA could not be directly attributed to a specific celltype due to the technical difficulties of triple labeling an-tibody-based IHC Therefore we cannot rule out the pos-sibility that other cell types such as endothelial (Conwayet al 2003) or inflammatory cells (Hill-Felberg et al1999) may also contribute to survivin and PCNA ex-pression after TBI The appearance of survivin and PCNAseparately in neurons (NeuN-positive) and astrocytes(GFAP-positive) along with co-localization of survivinwith PCNA in the same cells provide correlative data tosuggest an activation of cell cycle-like program in astro-cytes and possibly in a small subtype of neurons after
JOHNSON ET AL
1192
TBI In our experiments survivin co-localization withPCNA does suggest that survivin may be associated witha pro-mitotic process In an attempt to clarify these pro-teinrsquos roles after TBI we analyzed the nuclear morphol-ogy of survivin-positive cells to define the apoptotic ormitotic architecture of nuclei DAPI staining proved tooambiguous in identifying apoptotic versus mitotic phe-notypes likely due to the thickness of the brain sections(40 m) Further studies using direct markers of mitosissuch as BrdU incorporation as well as simultaneous la-beling with cell death related proteins is required to de-lineate anti-apoptotic and pro-mitotic activities of sur-vivin and PCNA in these cells
In conclusion our data demonstrate the induction ofsurvivin in the rat brain following TBI Expression of sur-vivin occurred predominantly in astrocytes as comparedto neurons in a time-dependant fashion and was accom-panied by expression of PCNA Taken together these re-sults suggest that survivin plays a role in neural cell re-sponses following traumatic brain injury in rats Futurestudies will investigate the implications of these findingsto the pathophysiology of TBI
ACKNOWLEDGMENTS
We thank Barbara OrsquoSteen and Tao Fan MD fortechnical assistance This work was funded by grants NIHRO1 NS39091 NIH RO1 NS40182 DAMD 17-99-1-9565 and DAMD 17-01-1-0765
REFERENCES
ADIDA C CROTTY PL MCGRATH J BERREBI DDIEBOLD J and ALTIERI DC (1998) Developmentallyregulated expression of the novel cancer anti-apoptosis genesurvivin in human and mouse differentiation Am J Pathol152 43ndash49
ALTIERI DC MARCHISIO PC and MARCHISIO C(1999) Survivin apoptosis an interloper between cell deathand cell proliferation in cancer Lab Invest 79 1327ndash1333
ALTSCHUL SF MADDEN TL SCHAFFER AA et al(1997) Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 253389ndash3402
AMBROSINI G ADIDA C and ALTIERI DC (1997) Anovel anti-apoptosis gene survivin expressed in cancer andlymphoma Nat Med 3 917ndash921
BRAVO R FRANK R BLUNDELL PA and MAC-DONALD-BRAVO H (1987) CyclinPCNA is the auxil-iary protein of DNA polymerase-delta Nature 326 515ndash517
BUSH TG PUVANACHANDRA N HORNER CH et al
(1999) Leukocyte infiltration neuronal degeneration andneurite outgrowth after ablation of scar-forming reactive as-trocytes in adult transgenic mice Neuron 23 297ndash308
CAMERON HA and MCKAY RD (2001) Adult neuroge-nesis produces a large pool of new granule cells in the den-tate gyrus J Comp Neurol 435 406ndash417
CARBONELL WS and GRADY MS (1999) Regional andtemporal characterization of neuronal glial and axonal re-sponse after traumatic brain injury in the mouse Acta Neu-ropathol (Berl) 98 396ndash406
CHEN XH IWATA A NONAKA M BROWNE KDand SMITH DH (2003) Neurogenesis and glial prolifera-tion persist for at least one year in the subventricular zonefollowing brain trauma in rats J Neurotrauma 20 623ndash631
CHEN Y and SWANSON RA (2003) Astrocytes and braininjury J Cereb Blood Flow Metab 23 137ndash149
CONWAY EM POLLEFEYT S CORNELISSEN J et al(2000) Three differentially expressed survivin cDNA vari-ants encode proteins with distinct antiapoptotic functionsBlood 95 1435ndash1442
CSUKA E HANS VH AMMANN E TRENTZ OKOSSMANN T and MORGANTI-KOSSMANN MC(2000) Cell activation and inflammatory response followingtraumatic axonal injury in the rat Neuroreport 112587ndash2590
DEVERAUX QL and REED JC (1999) IAP family pro-teinsmdashsuppressors of apoptosis Genes Dev 13 239ndash252
DIXON CE CLIFTON GL LIGHTHALL JW YAGH-MAI AA and HAYES RL (1991) A controlled corticalimpact model of traumatic brain injury in the rat J NeurosciMethods 39 253ndash262
DOETSCH F CAILLE I LIM DA GARCIA-VERDUGOJM and ALVAREZ-BUYLLA A (1999) Subventricularzone astrocytes are neural stem cells in the adult mammalianbrain Cell 97 703ndash716
DUNN-MEYNELL AA and LEVIN BE (1997) Histolog-ical markers of neuronal axonal and astrocytic changes af-ter lateral rigid impact traumatic brain injury Brain Res 76125ndash41
GIULIAN D (1991) Microgliamdashneuron interactions after in-jury to the central nervous system in Peripheral Signalingof the Brain Role in Neural-Immune Interactions Learningand Memory R Frederickson J McGaugh and D Felton(eds) Lewiston NY Hogrefe and Huber pps 73ndash82
HIGGINS DG BLEASBY AJ and FUCHS R (1992)CLUSTAL V improved software for multiple sequencealignment Comput Appl Biosci 8 189ndash191
HILL-FELBERG SJ MCINTOSH TK OLIVER DLRAGHUPATHI R and BARBARESE E (1999) Concur-rent loss and proliferation of astrocytes following lateral fluidpercussion brain injury in the adult rat J Neurosci Res 57271ndash279
1193
SURVIVIN UPREGULATION AFTER TBI
JIANG X WILFORD C DUENSING S MUNGER KJONES G and JONES D (2001) Participation of Survivinin mitotic and apoptotic activities of normal and tumor-de-rived cells J Cell Biochem 83 342ndash354
KAYA SS MAHMOOD A LI Y YAVUZ E andCHOPP M (1999a) Expression of cell cycle proteins (cy-clin D1 and cdk4) after controlled cortical impact in rat brainJ Neurotrauma 16 1187ndash1196
KAYA SS MAHMOOD A LI Y YAVUZ E GOKSELM and CHOPP M (1999b) Apoptosis and expression ofp53 response proteins and cyclin D1 after cortical impact inrat brain Brain Res 818 23ndash33
KERNIE SG ERWIN TM and PARADA LF (2001)Brain remodeling due to neuronal and astrocytic prolifera-tion after controlled cortical injury in mice J Neurosci Res66 317ndash326
KOBAYASHI K HATANO M OTAKI M OGA-SAWARA T and TOKUHISA T (1999) Expression of amurine homologue of the inhibitor of apoptosis protein is re-lated to cell proliferation Proc Natl Acad Sci USA 961457ndash1462
LACASSE EC BAIRD S KORNELUK RG andMACKENZIE AE (1998) The inhibitors of apoptosis(IAPs) and their emerging role in cancer Oncogene 173247ndash3259
LATOV N NILAVER G ZIMMERMAN EA et al(1979) Fibrillary astrocytes proliferate in response to braininjury a study combining immunoperoxidase technique forglial fibrillary acidic protein and radioautography of tritiatedthymidine Dev Biol 72 381ndash384
LI F and ALTIERI DC (1999) The cancer antiapoptosismouse survivin gene characterization of locus and tran-scriptional requirements of basal and cell cycle-dependentexpression Cancer Res 59 3143ndash3151
LI F AMBROSINI G CHU EY et al (1998) Control ofapoptosis and mitotic spindle checkpoint by survivin Nature396 580ndash584
LI F ACKERMANN EJ BENNETT CF et al (1999)Pleiotropic cell-division defects and apoptosis induced by in-terference with survivin function Nat Cell Biol 1 461ndash466
MAGAVI SS LEAVITT BR and MACKLIS JD (2000)Induction of neurogenesis in the neocortex of adult mice Na-ture 405 951ndash955
MIYAKE T OKADA M and KITAMURA T (1992) Re-active proliferation of astrocytes studied by immunohisto-chemistry for proliferating cell nuclear antigen Brain Res590 300ndash302
MUCHMORE SW CHEN J JAKOB C et al (2000)Crystal structure and mutagenic analysis of the inhibitor-of-apoptosis protein survivin Mol Cell 6 173ndash182
NORTON WT (1999) Cell reactions following acute braininjury a review Neurochem Res 24 213ndash218
OTAKI M HATANO M KOBAYASHI K et al (2000)Cell cyclendashdependent regulation of TIAPm-survivin expres-sion Biochim Biophys Acta 1493 188ndash194
PETERSON DA (2002) Stem cells in brain plasticity and re-pair Curr Opin Pharmacol 2 34ndash42
PIKE BR ZHAO X NEWCOMB JK POSMANTURRM WANG KK and Hayes RL (1998) Regional cal-pain and caspase-3 proteolysis of alpha-spectrin after trau-matic brain injury Neuroreport 9 2437ndash2442
RAGHUPATHI R GRAHAM DI MCINTOSH TK(2000) Apoptosis after traumatic brain injury J Neuro-trauma 17 927ndash938
RIDET JL MALHOTRA SK PRIVAT A and GAGEFH (1997) Reactive astrocytes cellular and molecular cuesto biological function Trends Neurosci 20 570ndash577
SASAKI T LOPES MB HANKINS GR and HELMGA (2002) Expression of survivin an inhibitor of apopto-sis protein in tumors of the nervous system Acta Neu-ropathol (Berl) 104 105ndash109
SHANKAR SL MANI S OrsquoGUIN KN KANDIMALLAER AGRAWAL S and SHAFIT-ZAGARDO B (2001)Survivin inhibition induces human neural tumor cell deaththrough caspase-independent and -dependent pathways JNeurochem 79 426ndash436
SHIN S SUNG BJ CHO YS et al (2001) An anti-apop-totic protein human survivin is a direct inhibitor of caspase-3 and -7 Biochemistry 40 1117ndash1123
SMITH C BERRY M CLARKE WE and LOGAN A(2001) Differential expression of fibroblast growth factorndash2and fibroblast growth factor receptor 1 in a scarring and non-scarring model of CNS injury in the rat Eur J Neurosci 13443ndash456
STONE JR OKONKWO DO SINGLETON RHMUTLU LK HELM GA and POVLISHOCK JT(2002) Caspase-3ndashmediated cleavage of amyloid precursorprotein and formation of amyloid Beta peptide in traumaticaxonal injury J Neurotrauma 19 601ndash614
SYKOVA E VARGOVA L PROKOPOVA S and SI-MONOVA Z (1999) Glial swelling and astrogliosis pro-duce diffusion barriers in the rat spinal cord Glia 25 56ndash70
TAKAHASHI R DEVERAUX Q TAMM I et al (1998)A single BIR domain of XIAP sufficient for inhibiting cas-pases J Biol Chem 273 7787ndash7790
TAMM I WANG Y SAUSVILLE E et al (1998) IAP-family protein survivin inhibits caspase activity and apopto-sis induced by Fas (CD95) Bax caspases and anticancerdrugs Cancer Res 58 5315ndash5320
THOMPSON JD HIGGINS DG and GIBSON TJ (1994)CLUSTAL W improving the sensitivity of progressive mul-tiple sequence alignment through sequence weighting posi-tion-specific gap penalties and weight matrix choice NucleicAcids Res 22 4673ndash4680
JOHNSON ET AL
1194
TOLENTINO PJ DEFORD SM NOTTERPEK L et al(2002) Up-regulation of tissue-type transglutaminase aftertraumatic brain injury J Neurochem 80 579ndash588
UREN AG WONG L PAKUSCH M et al (2000) Sur-vivin and the inner centromere protein INCENP show simi-lar cell-cycle localization and gene knockout phenotypeCurr Biol 10 1319ndash1328
WEI LH HUANG CY CHENG SP CHEN CA andHSIEH CY (2001) Carcinosarcoma of ovary associatedwith previous radiotherapy Int J Gynecol Cancer 11 81ndash84
WU Q COMBS C CANNADY SB GELDMACHERDS and HERRUP K (2000) Beta-amyloid activated mi-croglia induce cell cycling and cell death in cultured corticalneurons Neurobiol Aging 21 797ndash806
YAGITA Y KITAGAWA K OHTSUKI T et al (2001)
Neurogenesis by progenitor cells in the ischemic adult rathippocampus Stroke 32 1890ndash1896
YANG Y GELDMACHER DS and HERRUP K (2001)DNA replication precedes neuronal cell death in Alzheimerrsquosdisease J Neurosci 21 2661ndash2668
Address reprint requests toJose A Pineda PhD
EF and WL McKnight Brain InstituteUniversity of Florida
100 S Newell Dr Bldg 59L1-118 Dept of Pediatrics
Gainesville FL 32610
E-mail pinedjapedsufledu
SURVIVIN UPREGULATION AFTER TBI
1195

TBI In our experiments survivin co-localization withPCNA does suggest that survivin may be associated witha pro-mitotic process In an attempt to clarify these pro-teinrsquos roles after TBI we analyzed the nuclear morphol-ogy of survivin-positive cells to define the apoptotic ormitotic architecture of nuclei DAPI staining proved tooambiguous in identifying apoptotic versus mitotic phe-notypes likely due to the thickness of the brain sections(40 m) Further studies using direct markers of mitosissuch as BrdU incorporation as well as simultaneous la-beling with cell death related proteins is required to de-lineate anti-apoptotic and pro-mitotic activities of sur-vivin and PCNA in these cells
In conclusion our data demonstrate the induction ofsurvivin in the rat brain following TBI Expression of sur-vivin occurred predominantly in astrocytes as comparedto neurons in a time-dependant fashion and was accom-panied by expression of PCNA Taken together these re-sults suggest that survivin plays a role in neural cell re-sponses following traumatic brain injury in rats Futurestudies will investigate the implications of these findingsto the pathophysiology of TBI
ACKNOWLEDGMENTS
We thank Barbara OrsquoSteen and Tao Fan MD fortechnical assistance This work was funded by grants NIHRO1 NS39091 NIH RO1 NS40182 DAMD 17-99-1-9565 and DAMD 17-01-1-0765
REFERENCES
ADIDA C CROTTY PL MCGRATH J BERREBI DDIEBOLD J and ALTIERI DC (1998) Developmentallyregulated expression of the novel cancer anti-apoptosis genesurvivin in human and mouse differentiation Am J Pathol152 43ndash49
ALTIERI DC MARCHISIO PC and MARCHISIO C(1999) Survivin apoptosis an interloper between cell deathand cell proliferation in cancer Lab Invest 79 1327ndash1333
ALTSCHUL SF MADDEN TL SCHAFFER AA et al(1997) Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 253389ndash3402
AMBROSINI G ADIDA C and ALTIERI DC (1997) Anovel anti-apoptosis gene survivin expressed in cancer andlymphoma Nat Med 3 917ndash921
BRAVO R FRANK R BLUNDELL PA and MAC-DONALD-BRAVO H (1987) CyclinPCNA is the auxil-iary protein of DNA polymerase-delta Nature 326 515ndash517
BUSH TG PUVANACHANDRA N HORNER CH et al
(1999) Leukocyte infiltration neuronal degeneration andneurite outgrowth after ablation of scar-forming reactive as-trocytes in adult transgenic mice Neuron 23 297ndash308
CAMERON HA and MCKAY RD (2001) Adult neuroge-nesis produces a large pool of new granule cells in the den-tate gyrus J Comp Neurol 435 406ndash417
CARBONELL WS and GRADY MS (1999) Regional andtemporal characterization of neuronal glial and axonal re-sponse after traumatic brain injury in the mouse Acta Neu-ropathol (Berl) 98 396ndash406
CHEN XH IWATA A NONAKA M BROWNE KDand SMITH DH (2003) Neurogenesis and glial prolifera-tion persist for at least one year in the subventricular zonefollowing brain trauma in rats J Neurotrauma 20 623ndash631
CHEN Y and SWANSON RA (2003) Astrocytes and braininjury J Cereb Blood Flow Metab 23 137ndash149
CONWAY EM POLLEFEYT S CORNELISSEN J et al(2000) Three differentially expressed survivin cDNA vari-ants encode proteins with distinct antiapoptotic functionsBlood 95 1435ndash1442
CSUKA E HANS VH AMMANN E TRENTZ OKOSSMANN T and MORGANTI-KOSSMANN MC(2000) Cell activation and inflammatory response followingtraumatic axonal injury in the rat Neuroreport 112587ndash2590
DEVERAUX QL and REED JC (1999) IAP family pro-teinsmdashsuppressors of apoptosis Genes Dev 13 239ndash252
DIXON CE CLIFTON GL LIGHTHALL JW YAGH-MAI AA and HAYES RL (1991) A controlled corticalimpact model of traumatic brain injury in the rat J NeurosciMethods 39 253ndash262
DOETSCH F CAILLE I LIM DA GARCIA-VERDUGOJM and ALVAREZ-BUYLLA A (1999) Subventricularzone astrocytes are neural stem cells in the adult mammalianbrain Cell 97 703ndash716
DUNN-MEYNELL AA and LEVIN BE (1997) Histolog-ical markers of neuronal axonal and astrocytic changes af-ter lateral rigid impact traumatic brain injury Brain Res 76125ndash41
GIULIAN D (1991) Microgliamdashneuron interactions after in-jury to the central nervous system in Peripheral Signalingof the Brain Role in Neural-Immune Interactions Learningand Memory R Frederickson J McGaugh and D Felton(eds) Lewiston NY Hogrefe and Huber pps 73ndash82
HIGGINS DG BLEASBY AJ and FUCHS R (1992)CLUSTAL V improved software for multiple sequencealignment Comput Appl Biosci 8 189ndash191
HILL-FELBERG SJ MCINTOSH TK OLIVER DLRAGHUPATHI R and BARBARESE E (1999) Concur-rent loss and proliferation of astrocytes following lateral fluidpercussion brain injury in the adult rat J Neurosci Res 57271ndash279
1193
SURVIVIN UPREGULATION AFTER TBI
JIANG X WILFORD C DUENSING S MUNGER KJONES G and JONES D (2001) Participation of Survivinin mitotic and apoptotic activities of normal and tumor-de-rived cells J Cell Biochem 83 342ndash354
KAYA SS MAHMOOD A LI Y YAVUZ E andCHOPP M (1999a) Expression of cell cycle proteins (cy-clin D1 and cdk4) after controlled cortical impact in rat brainJ Neurotrauma 16 1187ndash1196
KAYA SS MAHMOOD A LI Y YAVUZ E GOKSELM and CHOPP M (1999b) Apoptosis and expression ofp53 response proteins and cyclin D1 after cortical impact inrat brain Brain Res 818 23ndash33
KERNIE SG ERWIN TM and PARADA LF (2001)Brain remodeling due to neuronal and astrocytic prolifera-tion after controlled cortical injury in mice J Neurosci Res66 317ndash326
KOBAYASHI K HATANO M OTAKI M OGA-SAWARA T and TOKUHISA T (1999) Expression of amurine homologue of the inhibitor of apoptosis protein is re-lated to cell proliferation Proc Natl Acad Sci USA 961457ndash1462
LACASSE EC BAIRD S KORNELUK RG andMACKENZIE AE (1998) The inhibitors of apoptosis(IAPs) and their emerging role in cancer Oncogene 173247ndash3259
LATOV N NILAVER G ZIMMERMAN EA et al(1979) Fibrillary astrocytes proliferate in response to braininjury a study combining immunoperoxidase technique forglial fibrillary acidic protein and radioautography of tritiatedthymidine Dev Biol 72 381ndash384
LI F and ALTIERI DC (1999) The cancer antiapoptosismouse survivin gene characterization of locus and tran-scriptional requirements of basal and cell cycle-dependentexpression Cancer Res 59 3143ndash3151
LI F AMBROSINI G CHU EY et al (1998) Control ofapoptosis and mitotic spindle checkpoint by survivin Nature396 580ndash584
LI F ACKERMANN EJ BENNETT CF et al (1999)Pleiotropic cell-division defects and apoptosis induced by in-terference with survivin function Nat Cell Biol 1 461ndash466
MAGAVI SS LEAVITT BR and MACKLIS JD (2000)Induction of neurogenesis in the neocortex of adult mice Na-ture 405 951ndash955
MIYAKE T OKADA M and KITAMURA T (1992) Re-active proliferation of astrocytes studied by immunohisto-chemistry for proliferating cell nuclear antigen Brain Res590 300ndash302
MUCHMORE SW CHEN J JAKOB C et al (2000)Crystal structure and mutagenic analysis of the inhibitor-of-apoptosis protein survivin Mol Cell 6 173ndash182
NORTON WT (1999) Cell reactions following acute braininjury a review Neurochem Res 24 213ndash218
OTAKI M HATANO M KOBAYASHI K et al (2000)Cell cyclendashdependent regulation of TIAPm-survivin expres-sion Biochim Biophys Acta 1493 188ndash194
PETERSON DA (2002) Stem cells in brain plasticity and re-pair Curr Opin Pharmacol 2 34ndash42
PIKE BR ZHAO X NEWCOMB JK POSMANTURRM WANG KK and Hayes RL (1998) Regional cal-pain and caspase-3 proteolysis of alpha-spectrin after trau-matic brain injury Neuroreport 9 2437ndash2442
RAGHUPATHI R GRAHAM DI MCINTOSH TK(2000) Apoptosis after traumatic brain injury J Neuro-trauma 17 927ndash938
RIDET JL MALHOTRA SK PRIVAT A and GAGEFH (1997) Reactive astrocytes cellular and molecular cuesto biological function Trends Neurosci 20 570ndash577
SASAKI T LOPES MB HANKINS GR and HELMGA (2002) Expression of survivin an inhibitor of apopto-sis protein in tumors of the nervous system Acta Neu-ropathol (Berl) 104 105ndash109
SHANKAR SL MANI S OrsquoGUIN KN KANDIMALLAER AGRAWAL S and SHAFIT-ZAGARDO B (2001)Survivin inhibition induces human neural tumor cell deaththrough caspase-independent and -dependent pathways JNeurochem 79 426ndash436
SHIN S SUNG BJ CHO YS et al (2001) An anti-apop-totic protein human survivin is a direct inhibitor of caspase-3 and -7 Biochemistry 40 1117ndash1123
SMITH C BERRY M CLARKE WE and LOGAN A(2001) Differential expression of fibroblast growth factorndash2and fibroblast growth factor receptor 1 in a scarring and non-scarring model of CNS injury in the rat Eur J Neurosci 13443ndash456
STONE JR OKONKWO DO SINGLETON RHMUTLU LK HELM GA and POVLISHOCK JT(2002) Caspase-3ndashmediated cleavage of amyloid precursorprotein and formation of amyloid Beta peptide in traumaticaxonal injury J Neurotrauma 19 601ndash614
SYKOVA E VARGOVA L PROKOPOVA S and SI-MONOVA Z (1999) Glial swelling and astrogliosis pro-duce diffusion barriers in the rat spinal cord Glia 25 56ndash70
TAKAHASHI R DEVERAUX Q TAMM I et al (1998)A single BIR domain of XIAP sufficient for inhibiting cas-pases J Biol Chem 273 7787ndash7790
TAMM I WANG Y SAUSVILLE E et al (1998) IAP-family protein survivin inhibits caspase activity and apopto-sis induced by Fas (CD95) Bax caspases and anticancerdrugs Cancer Res 58 5315ndash5320
THOMPSON JD HIGGINS DG and GIBSON TJ (1994)CLUSTAL W improving the sensitivity of progressive mul-tiple sequence alignment through sequence weighting posi-tion-specific gap penalties and weight matrix choice NucleicAcids Res 22 4673ndash4680
JOHNSON ET AL
1194
TOLENTINO PJ DEFORD SM NOTTERPEK L et al(2002) Up-regulation of tissue-type transglutaminase aftertraumatic brain injury J Neurochem 80 579ndash588
UREN AG WONG L PAKUSCH M et al (2000) Sur-vivin and the inner centromere protein INCENP show simi-lar cell-cycle localization and gene knockout phenotypeCurr Biol 10 1319ndash1328
WEI LH HUANG CY CHENG SP CHEN CA andHSIEH CY (2001) Carcinosarcoma of ovary associatedwith previous radiotherapy Int J Gynecol Cancer 11 81ndash84
WU Q COMBS C CANNADY SB GELDMACHERDS and HERRUP K (2000) Beta-amyloid activated mi-croglia induce cell cycling and cell death in cultured corticalneurons Neurobiol Aging 21 797ndash806
YAGITA Y KITAGAWA K OHTSUKI T et al (2001)
Neurogenesis by progenitor cells in the ischemic adult rathippocampus Stroke 32 1890ndash1896
YANG Y GELDMACHER DS and HERRUP K (2001)DNA replication precedes neuronal cell death in Alzheimerrsquosdisease J Neurosci 21 2661ndash2668
Address reprint requests toJose A Pineda PhD
EF and WL McKnight Brain InstituteUniversity of Florida
100 S Newell Dr Bldg 59L1-118 Dept of Pediatrics
Gainesville FL 32610
E-mail pinedjapedsufledu
SURVIVIN UPREGULATION AFTER TBI
1195

JIANG X WILFORD C DUENSING S MUNGER KJONES G and JONES D (2001) Participation of Survivinin mitotic and apoptotic activities of normal and tumor-de-rived cells J Cell Biochem 83 342ndash354
KAYA SS MAHMOOD A LI Y YAVUZ E andCHOPP M (1999a) Expression of cell cycle proteins (cy-clin D1 and cdk4) after controlled cortical impact in rat brainJ Neurotrauma 16 1187ndash1196
KAYA SS MAHMOOD A LI Y YAVUZ E GOKSELM and CHOPP M (1999b) Apoptosis and expression ofp53 response proteins and cyclin D1 after cortical impact inrat brain Brain Res 818 23ndash33
KERNIE SG ERWIN TM and PARADA LF (2001)Brain remodeling due to neuronal and astrocytic prolifera-tion after controlled cortical injury in mice J Neurosci Res66 317ndash326
KOBAYASHI K HATANO M OTAKI M OGA-SAWARA T and TOKUHISA T (1999) Expression of amurine homologue of the inhibitor of apoptosis protein is re-lated to cell proliferation Proc Natl Acad Sci USA 961457ndash1462
LACASSE EC BAIRD S KORNELUK RG andMACKENZIE AE (1998) The inhibitors of apoptosis(IAPs) and their emerging role in cancer Oncogene 173247ndash3259
LATOV N NILAVER G ZIMMERMAN EA et al(1979) Fibrillary astrocytes proliferate in response to braininjury a study combining immunoperoxidase technique forglial fibrillary acidic protein and radioautography of tritiatedthymidine Dev Biol 72 381ndash384
LI F and ALTIERI DC (1999) The cancer antiapoptosismouse survivin gene characterization of locus and tran-scriptional requirements of basal and cell cycle-dependentexpression Cancer Res 59 3143ndash3151
LI F AMBROSINI G CHU EY et al (1998) Control ofapoptosis and mitotic spindle checkpoint by survivin Nature396 580ndash584
LI F ACKERMANN EJ BENNETT CF et al (1999)Pleiotropic cell-division defects and apoptosis induced by in-terference with survivin function Nat Cell Biol 1 461ndash466
MAGAVI SS LEAVITT BR and MACKLIS JD (2000)Induction of neurogenesis in the neocortex of adult mice Na-ture 405 951ndash955
MIYAKE T OKADA M and KITAMURA T (1992) Re-active proliferation of astrocytes studied by immunohisto-chemistry for proliferating cell nuclear antigen Brain Res590 300ndash302
MUCHMORE SW CHEN J JAKOB C et al (2000)Crystal structure and mutagenic analysis of the inhibitor-of-apoptosis protein survivin Mol Cell 6 173ndash182
NORTON WT (1999) Cell reactions following acute braininjury a review Neurochem Res 24 213ndash218
OTAKI M HATANO M KOBAYASHI K et al (2000)Cell cyclendashdependent regulation of TIAPm-survivin expres-sion Biochim Biophys Acta 1493 188ndash194
PETERSON DA (2002) Stem cells in brain plasticity and re-pair Curr Opin Pharmacol 2 34ndash42
PIKE BR ZHAO X NEWCOMB JK POSMANTURRM WANG KK and Hayes RL (1998) Regional cal-pain and caspase-3 proteolysis of alpha-spectrin after trau-matic brain injury Neuroreport 9 2437ndash2442
RAGHUPATHI R GRAHAM DI MCINTOSH TK(2000) Apoptosis after traumatic brain injury J Neuro-trauma 17 927ndash938
RIDET JL MALHOTRA SK PRIVAT A and GAGEFH (1997) Reactive astrocytes cellular and molecular cuesto biological function Trends Neurosci 20 570ndash577
SASAKI T LOPES MB HANKINS GR and HELMGA (2002) Expression of survivin an inhibitor of apopto-sis protein in tumors of the nervous system Acta Neu-ropathol (Berl) 104 105ndash109
SHANKAR SL MANI S OrsquoGUIN KN KANDIMALLAER AGRAWAL S and SHAFIT-ZAGARDO B (2001)Survivin inhibition induces human neural tumor cell deaththrough caspase-independent and -dependent pathways JNeurochem 79 426ndash436
SHIN S SUNG BJ CHO YS et al (2001) An anti-apop-totic protein human survivin is a direct inhibitor of caspase-3 and -7 Biochemistry 40 1117ndash1123
SMITH C BERRY M CLARKE WE and LOGAN A(2001) Differential expression of fibroblast growth factorndash2and fibroblast growth factor receptor 1 in a scarring and non-scarring model of CNS injury in the rat Eur J Neurosci 13443ndash456
STONE JR OKONKWO DO SINGLETON RHMUTLU LK HELM GA and POVLISHOCK JT(2002) Caspase-3ndashmediated cleavage of amyloid precursorprotein and formation of amyloid Beta peptide in traumaticaxonal injury J Neurotrauma 19 601ndash614
SYKOVA E VARGOVA L PROKOPOVA S and SI-MONOVA Z (1999) Glial swelling and astrogliosis pro-duce diffusion barriers in the rat spinal cord Glia 25 56ndash70
TAKAHASHI R DEVERAUX Q TAMM I et al (1998)A single BIR domain of XIAP sufficient for inhibiting cas-pases J Biol Chem 273 7787ndash7790
TAMM I WANG Y SAUSVILLE E et al (1998) IAP-family protein survivin inhibits caspase activity and apopto-sis induced by Fas (CD95) Bax caspases and anticancerdrugs Cancer Res 58 5315ndash5320
THOMPSON JD HIGGINS DG and GIBSON TJ (1994)CLUSTAL W improving the sensitivity of progressive mul-tiple sequence alignment through sequence weighting posi-tion-specific gap penalties and weight matrix choice NucleicAcids Res 22 4673ndash4680
JOHNSON ET AL
1194
TOLENTINO PJ DEFORD SM NOTTERPEK L et al(2002) Up-regulation of tissue-type transglutaminase aftertraumatic brain injury J Neurochem 80 579ndash588
UREN AG WONG L PAKUSCH M et al (2000) Sur-vivin and the inner centromere protein INCENP show simi-lar cell-cycle localization and gene knockout phenotypeCurr Biol 10 1319ndash1328
WEI LH HUANG CY CHENG SP CHEN CA andHSIEH CY (2001) Carcinosarcoma of ovary associatedwith previous radiotherapy Int J Gynecol Cancer 11 81ndash84
WU Q COMBS C CANNADY SB GELDMACHERDS and HERRUP K (2000) Beta-amyloid activated mi-croglia induce cell cycling and cell death in cultured corticalneurons Neurobiol Aging 21 797ndash806
YAGITA Y KITAGAWA K OHTSUKI T et al (2001)
Neurogenesis by progenitor cells in the ischemic adult rathippocampus Stroke 32 1890ndash1896
YANG Y GELDMACHER DS and HERRUP K (2001)DNA replication precedes neuronal cell death in Alzheimerrsquosdisease J Neurosci 21 2661ndash2668
Address reprint requests toJose A Pineda PhD
EF and WL McKnight Brain InstituteUniversity of Florida
100 S Newell Dr Bldg 59L1-118 Dept of Pediatrics
Gainesville FL 32610
E-mail pinedjapedsufledu
SURVIVIN UPREGULATION AFTER TBI
1195

TOLENTINO PJ DEFORD SM NOTTERPEK L et al(2002) Up-regulation of tissue-type transglutaminase aftertraumatic brain injury J Neurochem 80 579ndash588
UREN AG WONG L PAKUSCH M et al (2000) Sur-vivin and the inner centromere protein INCENP show simi-lar cell-cycle localization and gene knockout phenotypeCurr Biol 10 1319ndash1328
WEI LH HUANG CY CHENG SP CHEN CA andHSIEH CY (2001) Carcinosarcoma of ovary associatedwith previous radiotherapy Int J Gynecol Cancer 11 81ndash84
WU Q COMBS C CANNADY SB GELDMACHERDS and HERRUP K (2000) Beta-amyloid activated mi-croglia induce cell cycling and cell death in cultured corticalneurons Neurobiol Aging 21 797ndash806
YAGITA Y KITAGAWA K OHTSUKI T et al (2001)
Neurogenesis by progenitor cells in the ischemic adult rathippocampus Stroke 32 1890ndash1896
YANG Y GELDMACHER DS and HERRUP K (2001)DNA replication precedes neuronal cell death in Alzheimerrsquosdisease J Neurosci 21 2661ndash2668
Address reprint requests toJose A Pineda PhD
EF and WL McKnight Brain InstituteUniversity of Florida
100 S Newell Dr Bldg 59L1-118 Dept of Pediatrics
Gainesville FL 32610
E-mail pinedjapedsufledu
SURVIVIN UPREGULATION AFTER TBI
1195




















