
Non-traumatic adhesive arachnoiditis as a cause of spinal ...
Upload
independentCategory
view
1download
0
*Spinal Cord and Brain Injury Research Center and Department of Anatomy and Neurobiology, University of Kentucky College of
Medicine, Lexington, Kentucky, USA
�Department of Orthodontics and Neuroscience, College of Dentistry and the Comprehensive Center for Pain Research, University of
Florida, Gainesville, Florida, USA
Traumatic spinal cord injury (SCI) is a leading cause ofpermanent locomotor disability among young adults in theUnited States, with 11 000 new injures each year. At present,effective treatments for locomotor disability associated withSCI are inadequate (French et al. 2007). Understanding thecomplex biochemical mechanisms contributing to locomotordeficits after SCI and developing novel therapeutic strategiesis of great clinical importance.
Extracellular signal-regulated kinase 1 and 2 (ERK1, a44 kDa protein and ERK2, a 42 kDa protein) are mitogen-activated protein kinase family members (Chen et al. 2001).They are encoded by distinct genes, but share 83% homologyat the amino acid level and are expressed in all tissues. Inresting cells, the proteins are largely cytosolic and associatedwith the cytoskeleton. Upon activation, ERK1/2 can trans-locate to the nucleus or to membrane specializations.Activation of ERK1/2 by phosphorylation of threonine andtyrosine residues was originally identified as an effect of thegrowth factor signaling cascades that involve activation ofRas, Raf, mitogen-activated protein kinase kinase (MEK),
and ERK1/2 and are associated with neuronal survival andneuroprotection (Hetman and Gozdz 2004).
However, recent reports have challenged this view andsuggest a neurodegenerative role of ERK1/2 (Yu and Yezier-ski 2005; Zhuang and Schnellmann 2006; Genovese et al.2008). For instance, blockade of ERK1/2 by MEK inhibitors(U0126 or PD98059) protects against neurodegeneration
Received July 22, 2009; revised manuscript received December 19,2009; accepted January 4, 2010.Address correspondence and reprint requests to Chen-Guang Yu,
M.D., Ph.D., Research Assistant Professor, Spinal Cord and Brain InjuryResearch Center, B465, Biomedical & Biological Sciences ResearchBuilding (BBSRB), 741 S. Limestone Street, Lexington, KY 40536-0509, USA. E-mail: [email protected] used: ATCC, American type culture collection; BBB,
Basso,Beattie,Bresnahan;CMV, cytomegalovirus;CY-3, cyanine 3; eGFP,enhanced green fluorescent protein; ERK1/2, extracellular signal-regulatedprotein kinase 1/2l; GADPH, glyceraldehyde-3-phosphate dehydrogenase;IH, infinite horizons;LV, lentivirus;MEK,mitogen-activatedproteinkinasekinase; QUIS, quisqualic acid; RNAi, RNA interference; SCI, spinal cordinjury.siRNA, small interfering RNA.
Abstract
Activation of extracellular signal-regulated protein kinase 1/2
(ERK1/2) are implicated in the pathophysiology of spinal cord
injury (SCI). However, the specific functions of individual ERK
isoforms in neurodegeneration are largely unknown. We
investigated the hypothesis that ERK2 activation may con-
tribute to pathological and functional deficits following SCI and
that ERK2 knockdown using RNA interference may provide a
novel therapeutic strategy for SCI. Lentiviral ERK2 shRNA
and siRNA were utilized to knockdown ERK2 expression in
the spinal cord following SCI. Pre-injury intrathecal adminis-
tration of ERK2 siRNA significantly reduced excitotoxic injury-
induced activation of ERK2 (p < 0.001) and caspase 3
(p < 0.01) in spinal cord. Intraspinal administration of lentiviral
ERK2 shRNA significantly reduced ERK2 expression in the
spinal cord (p < 0.05), but did not alter ERK1 expression.
Administration of the lentiviral ERK2 shRNA vector 1 week
prior to severe spinal cord contusion injury resulted in a sig-
nificant improvement in locomotor function (p < 0.05), total
tissue sparing (p < 0.05), white matter sparing (p < 0.05), and
gray matter sparing (p < 0.05) 6 weeks following severe
contusive SCI. Our results suggest that ERK2 signaling is a
novel target associated with the deleterious consequences of
spinal injury.
Keywords: extracellular signal-regulated protein kinase 2,
locomotor function, RNA interference, spinal cord injury, tis-
sue damage.
J. Neurochem. (2010) 113, 131–142.
JOURNAL OF NEUROCHEMISTRY | 2010 | 113 | 131–142 doi: 10.1111/j.1471-4159.2010.06579.x
� 2010 The AuthorsJournal Compilation � 2010 International Society for Neurochemistry, J. Neurochem. (2010) 113, 131–142 131

induced by excitotoxic SCI and spinal cord compressioninjury (Yu and Yezierski 2005; Genovese et al. 2008); ERK2knockdown with RNA interference (RNAi) inhibits neuroki-nin-1 receptor or protein kinase-C a-mediated cell death(Matsumoto et al. 2005) and complete Freund’s adjuvant-induced inflammatory pain (Xu et al. 2008). However, thespecific functions of the individual ERK isoforms in neurode-generation after SCI are unknown. Isoform-specific inhibitorsof ERK 1 or ERK2 are not available. As a potent and specificinhibitor of gene expression,RNAi has emerged as a promisingtool to analyze individual gene function, using local delivery ofsequence-specific small interfering RNA (siRNA) (Breckpotet al. 2007; Lu and Woodle 2008). To test the hypothesis thatERK2 activation is involved in the injury-induced neurode-generation of spinal tissue and locomotor dysfunction, weutilizedmicroinjection of lentiviral-ERK2 (LV-ERK2) shRNAor intrathecal administration of ERK2 siRNA to knockdownspinal ERK2 prior to contusive or excitotoxic SCI in rats.
Materials and methods
AnimalsFor this study, 38 female and 12male Long-Evans rats (Charles River,
Indianapolis, IN, USA), weighing 200–250 g, were used. Animals
were kept under standard housing conditions for at least 1 week
following arrival. All experimental procedures were approved and
carried out in accordance with the Guidelines of the US National
Institutes of Health and Institutional Animal Care and Use Committee
of the University of Kentucky and University of Florida.
ERK2 siRNA sequenceERK2 siRNA sequences (Genebank accession no. M64300) were
selected and chemically modified based on accepted criteria for the
rational design of siRNAwith off-target control using two web-based
programs (Dharmacon, Inc., Lafayette, CO, USA and Invitrogen,
Carlsbad, CA, USA) (Reynolds et al. 2004). To minimize the
potential off-target effects of siRNA, the ERK2 siRNA sequences
(Sense, start 522, 5¢-pGCACCUCAGCAAUGAUCAUdTdT-3¢ andantisense 5¢-pAUGAUCAUUGCUGAGGUGCdTdT-3¢) were sub-
jected to BLAST analysis and synthesized in the ‘ready-to-use invivo’ and modification option with off-target control by Thermo
Scientific-Dharmacon Products (Lafayette, CO, USA). ON-TAR-
GET plus Non-Targeting siRNA was used as the negative control
siRNA (Dharmacon).
Cell culture and transfectionTransfection was performed as described in the Invitrogen siRNA
Transfection Manual using Lipofectamine 2000. To determine the
efficacy of ERK2 siRNA sequences, PC-12 cells (3 · 105/well in
six-well plates) were transfected with 100 nM ERK2 siRNAs
complexed with 6 lL Lipofectamine 2000, based on the manufac-
turer’s instructions (Invitrogen). ERK2 mRNA and protein levels in
the cells were examined 72 h following transfection. Real-time PCR
was employed to determine ERK2 mRNA levels. Western blot
analysis was used to estimate protein levels. A 70% reduction in
both mRNA and protein levels was considered an effective gene
silencing effect. PC-12 cells were purchased from American Type
Culture Collection (ATCC, Manassas, VA) and maintained in
Dulbecco’s Modified Eagle Medium supplemented with 5% fetal
bovine serum, 10% heat-inactivated donor horse serum, 50 lg/ml
streptomycin, and 50 IU/ml penicillin in a humidified atmosphere at
37�C and 5% CO2 as described in the protocol provided by ATCC.
Cells were maintained at a medium density (3 · 105/cm2).
Construction and production of lentiviral vectorsERK2 shRNA was designed and synthesized based on the active
ERK2 siRNA sequence using Invitrogen online software and
cloned into BLOCK-iT U6 RNAi Entry Vector using U6 Entry
Vector Kit (Invitrogen). The U6-ERK2 shRNA expression cassette
was inserted between BamHI and XbaI sites in a lentiviral-
cytomegalovirus (CMV)-RNAi vector (third generation lentiviral
vector was kindly provided by Dr George Smith, University of
Kentucky). Expression of the shRNA is under control of a U6
promoter, while the enhanced green fluorescent protein (eGFP)
reporter is linked to a CMV promoter. The lentiviral vector
containing eGFP and ERK2 shRNA or the empty vector
expressing only eGFP (LV-control) was co-transfected with
packaging plasmid PAX2 and envelope plasmid pMD2G (obtained
from Dr Didier Trono, University of Geneva, Switzerland) into
293FT cells (Invitrogen) using calcium-phosphate precipitation.
Viral supernatants were harvested and concentrated after 72 h. The
titer of the lentiviral stocks was assessed using 10-fold serial
dilutions to HT-1080 human fibrosarcoma cells (ATCC) using the
eGFP reporter of the lenti-CMV-eGFP-RNAi vector to identify
infected cells. The viral titer was 4 · 108 TU/mL for LV-ERK2
shRNA and 5 · 107 TU/mL for LV-eGFP control.
Intrathecal siRNAThe siRNA-transfectamine 2000 complexes were prepared by
diluting ERK2 or control siRNA (2.5 lg) in opti-MEM (7.5 lL)and diluting lipofectamine 2000 (5 lg) in opti-MEM (7.5 lL) usinglipofectamine 2000 kit (Invitrogen). The two mixtures were incu-
bated for 5 min at 25�C, combined, and incubated for an additional
20 min. Complexes of ERK2 siRNA-lipofectamine 2000 or control
siRNA-lipofectamine 2000 (one dose) were intrathecally delivered
onto the surface of dorsal horn (QUIS injection site or L1 level) for a
period of 72 h prior to QUIS injections. The dose of ERK2 siRNA
was chosen based on previous studies (Cai et al. 2009).
Uptake of CY-3-labeled ERK2 siRNA in spinal cordCyanine 3 (CY-3)-ERK2 siRNA (2.5 lg) complexed with lipofec-
tamine 2000 (5 lg) were intrathecally delivered onto the surface of
dorsal horn in uninjured rats. 24 h after delivery, the rat was
perfused and spinal cords dissected and processed for epifluorescent
microscopy. The uptake of ERK2 siRNAwas verified in spinal cord
of rats by direct CY-3 visualization with fluorescent microscopy.
Intraspinal microinjectionLentiviral vectors encoding ERK2 shRNA or lentiviral control
vectors were bilaterally injected into the gray and white matter of the
spinal cord at T10 (four injections) in rats. Injections were made into
the lateral funiculus 1.1 mm lateral to the midline at a depth of
1 mm. For gray matter, bilateral injections were made at 0.5 mm
lateral to the midline at a depth of 1.3 mm to target motor neurons in
the ventral horn. Injections were made using a beveled glass
Journal Compilation � 2010 International Society for Neurochemistry, J. Neurochem. (2010) 113, 131–142� 2010 The Authors
132 | C.-G. Yu et al.

micropipette (60 lm tip diameter) and nano-injector (Stoelting Co.,
Wood Dale, IL, USA). For each injection site, a volume of up to
1.0 lL was slowly infused over 4 min (100 nL/min). The injection
protocol is similar to that described in detail by Ruitenberg et al.(2002) for delivery of adeno-associated viral vectors to the spinal
cord. Intraspinal injection of LV-ERK2 shRNA or LV-control was
verified using western blot, immunofluorescence staining, and
fluorescent microscopy.
Excitotoxic spinal cord injuryExcitotoxic injury was initiated by intraspinal injection of the
glutamatergic alpha-amino-3-hydroxy-5-methy-4-isoxazole propio-
nate (AMPA)/metabotropic receptor agonist quisqualic acid (QUIS)
as previously described (Yu and Yezierski 2005). The intraspinal
QUIS injection has been used for study of glutamatergic excitotox-
icity (Yu and Yezierski 2005). Briefly, male Long-Evan rats
weighing 200–250 g were anesthetized with a mixture of ketamine,
acepromazine, and rompun (0.65 mL/kg, subcutaneously). Animals
were placed in a stereotaxic unit and the spinal column immobilized
with a vertebral clamp. One injection window for intraspinalinjection was made by laminectomy between spinal segments T12-
L2. QUIS (125 mmol/L) was injected unilaterally at three levels of
the cord. Injections were made at a depth of 1000 lm below the
dura in segments between the dorsal root entry zone and the dorsal
vein. These coordinates place injections in the center of the gray
matter between spinal laminae IV–VI. The total volume of QUIS
injected was 1.2 lL (0.4 lL each site, delivered over 60 s).
Contusive SCIContusion SCI was produced following a T10 laminectomy using an
Infinite Horizons (IH) SCI device (Precision Systems & Instrumen-
tation, Lexington, KY, USA) as described previously (Scheff et al.2003; Yu and Geddes 2007). Briefly, adult female Long-Evans rats,
weighing 200–250 g, were anesthetized with ketamine (80 mg/kg,
ip) and xylazine (10 mg/kg, ip), and a laminectomy was made to
expose spinal segment T10. The exposed vertebral column was
stabilized by clamping the rostral T9 and caudal T11 vertebral bodies
with two spinal forceps. SCI was then applied with the IH device
using a 230 kdyn force setting, which resulted in severe contusion
injury. Impact analyses, including actual force applied to the spinal
cord, displacement of spinal cord, and velocity, were recorded. The
impact tip was automatically retracted immediately, the wound
irrigated with saline, and the muscle and skin openings were closed
with sutures. The surgical procedure and post-operative care were
similar to that described previously (Yu and Geddes 2007).
Real-time PCRTotal RNA was isolated from PC-12 cell lyses sample using a
modified guanidium thiocyanate-phenol-chloroform extraction
method as described previously (Yu and Yezierski 2005). First-
strand cDNA was synthesized using 1.0 lg of total RNA with the
SuperScript First-strand Synthesis System (Invitrogen). The first-
strand cDNA serves as a template for initial PCR and real-time PCR
analysis. Initial PCR using Taq DNA Polymerase PCR system
(QIAGEN, Valencia, CA, USA) was performed with Glyceralde-
hyde-3-phosphate dehydrogenase (GAPDH) and ERK2 specific
primers (see below) that resulted in 369-bp and 297-bp products,
respectively, according to the protocol provided by the manufac-
turer. Agarose gels were used to evaluate the amplification product
size. The use of DNase and no-RT control ensured lack of false-
positive/negative data and genomic DNA contamination. The
quantitative real-time PCR analysis was carried out using a Roche
Light Cycler (Roche Diagnostics, Mannheim, Germany) as
described previously (Yu and Yezierski 2005). The quantitative
assessment of PCR amplification was determined using the
fluorescent dye SYBR Green I, which binds to the minor grove of
the DNA double helix. Melting curve analysis of PCR products and
gel electrophoresis confirmed the specificity of real-time PCR
products. Two relative standard curves were obtained with 5- to 10-
fold dilutions from a PC-12 cell sample template: one for the target
gene and another for the endogenous control GAPDH gene. The
Light-Cycler program calculated a concentration value for each
reaction. Calculated concentrations of RNA target and endogenous
control (GAPDH) were determined by extrapolating from the
relative standard curves. The target value was expressed relative to
GAPDH value and normalized. All genes were amplified with high
efficiency during real-time PCR.
ERK2 primers: Sense 5¢-GCGCTACACTAATCTCTCGT-3¢;Size: 297 bp
Anti-sense 5¢-CTGAGGTGCTGTGTCTTCAA-3¢GAPDH primers: Sense 5¢-GGTGAAGGTCGGTGTGAACGG-
3¢; Size: 369-bpAnti-sense 5¢-GGCAGAAGGGGCGGAGATG-3¢
Western blottingPC-12 cells were washed with 1· phosphate-buffered saline, lysed
by adding buffer, and sonicated for 15 s to shear DNA and reduce
sample viscosity. For spinal cord samples, animals were euthana-
tized by pentobarbital (100 mg/kg, ip injection) and decapitated
2 weeks post-injury with treatment of LV-ERK2 shRNA/control
vectors or 2 days post-injury with treatment of ERK2 siRNA/control
siRNA. A 5-mm block of spinal cord centered on lesion site was
removed and snap-frozen on dry ice, then stored at )80�C. Thespinal cord samples were homogenized in a lysis buffer, sonicated,
and centrifuged at 20 426 ·g for 10 min. Protein content of the
supernatant was determined using the bicinchoninic acid method.
Western blotting was performed as previously described (Yu and
Yezierski 2005) except that infrared-labeled secondary antibodies
(IRDye Conjugates; Rockland Immunochemicals, Gilbertsville, PA,
USA) were used, designed for visualization with the LI-COR
Odyssey infrared imaging system (Lincoln, NE, USA). Briefly, PC-
12 cell or spinal cord protein samples were loaded on SDS–PAGE
gels and following electrophoresis were electrotransfered to nitro-
cellulose membranes. Blots were probed with a polyclonal antibody
against ERK2 (1 : 1000; Cell Signaling Technology, Beverly, MA,
USA) and reprobed with a monoclonal antibody against GAPDH
(1 : 1000; Chemicon, Temecula, CA, USA), ERK1, pERK1, or
pERK2 (1 : 1000; Cell Signaling). Blots were then incubated with
infrared-labeled anti-rabbit or anti-mouse secondary antibodies
(1 : 5000). All blots were visualized and analyzed on the LI-COR
Odyssey infrared imaging system.
ImmunofluorescenceFollowing anesthesia, the injured or naı̈ve rats at 4 h post-QUIS
injection or 2 weeks post-injection with LV-ERK2 shRNA or
� 2010 The AuthorsJournal Compilation � 2010 International Society for Neurochemistry, J. Neurochem. (2010) 113, 131–142
ERK2 following spinal injury | 133

control vectors were transcardially perfused with ice-cold saline
followed by phosphate-buffered 4% paraformaldehyde. The spinal
cord was removed, cut into blocks according to segments, and post-
fixed for 6 h. The fixed spinal cords were serially cryosectioned at
20 lm. Spinal expression of eGFP in the lentiviral vector-infected
sections and uptake of CY-3-labeled ERK2 siRNA in spinal sections
were directly viewed using an epifluorescent microscopy system.
Immunofluorescence staining was performed using Cell Signaling
Technology protocols. Briefly, sections were immunolabeled with
polyclonal antibodies against ERK2, pERK2 (1 : 200; Cell Signal-
ing), cleaved caspase 3 (1 : 200, Cell Signaling), or ED-1 (1 : 200;
Chemicon) followed by incubation with Alexa Fluor 594 or FITC-
conjugated secondary antibodies. The fluorescent ERK2 or ED-1
signal in the lentiviral vector injection epicenter were imaged/
analyzed and the pERK2-positive or cleaved caspase 3-positive
cells in the QUIS lesion epicenter were counted using an Olympus
spinning disk fluorescent microscopy image system (Olympus,
Center Valley, PA, USA) and Adobe Photoshop CS2 software
(Adobe Systems Incorporated, New York, NY, USA) (n = 3 sections
separated 60 lm apart per animal, n = 3–4 animals per group). For
each animal, the calculated fluorescent density for expression of
ERK2 and ED-1 or cell counts for activation of ERK2 and caspase 3
of three sections was averaged. Subsequently, the average density or
cell counts per section of all animals in each group was averaged to
obtain a mean value per section.
Assessment of locomotor functionAt 7 days post-injection with lentiviral-ERK2 shRNA or control
vectors, the rats received severe contusion SCI (230 kdyn). Open-
field locomotor function was assessed pre-injury and 0, 3, 7, 14, 21,
28, 35, and 42 days post-injury using the Basso, Beattie, Bresnahan
(BBB) locomotor rating scale as described previously (Basso et al.1996; Yu and Geddes 2007). The two examiners participating in the
BBB evaluation were blinded to the experimental treatment received
by each animal.
Assessment of tissue sparingAt 6 weeks post-injury, animals were euthanatized and transcar-
dially perfused with ice-cold saline followed by phosphate-
buffered 4% paraformaldehyde. The spinal cord was removed
and prepared for histological assessment as previously described
(Rabchevsky et al. 2002; Yu and Geddes 2007). Spinal cords were
serially cryosectioned at a thickness of 20 lm. Every fifth section
was mounted onto gelatin-coated slides and stored at )20�C.A modified eriochrome cyanine staining protocol for myelin that
differentiates both white matter and cell bodies was used to
visualize spared spinal tissue. Area measurements in lesion, gray
matter, white matter, and total spinal tissue and calculation of
lesion volume, total tissue sparing, white matter sparing, and gray
matter sparing in transverse sections of the injured cords were
performed as previously described (Rabchevsky et al. 2002; Yu
and Geddes 2007).
Experimental groupsFor siRNA treatment in PC-12 cells: PC-12 cells were assigned to
the following groups: for real-time PCR analysis, (i) ERK2 siRNA
treatment and (ii) control siRNA treatment; for western blot
analysis, (i) ERK2 siRNA treatment and (ii) control siRNA
treatment. Real-time PCR analysis was used to measure mRNA
levels of ERK2 gene expression 72 h post-transfections (n = 4 per
group). Western blot analysis was used to evaluate the protein levels
of ERK2 at 72 h post-transfections (n = 4 per group).
For injury-induced ERK1/2 activation: Female Long-Evans rats
were randomly assigned to the following groups: (i) sham control
(laminectomy without injury), and (ii) contusive SCI (n = 4/group).
Western blot analysis was used to determine whether injury causes
ERK1/2 activation 2 days after contusion SCI.
For lentiviral-shRNA pre-treatment for contusion SCI: Female
Long-Evans rats were randomly assigned to the following groups:
(i) LV-ERK2 shRNA plus sham control (laminectomy without
injury), (ii) LV-Control plus sham control (laminectomy without
injury), (iii) LV-ERK2 shRNA plus SCI, and (iv) LV-Control plus
SCI. Fluorescent microscopy and immunofluorescent staining
analysis were used to observe whether LV-ERK2 shRNA intraspinal
administration results in eGFP expression or ERK2 knockdown in
spinal cord at 2 weeks post-delivery of lentiviral vectors in
uninjured rats (n = 4). Western blot analysis was used to confirm
whether LV-ERK2 shRNA intraspinal administration results in
ERK2 knockdown in spinal cord 2 weeks post-injury (n = 4).
Fourteen virus-infected animals in the SCI group were used to
assess pathological and behavioral outcome 6 weeks following
injury using histological and behavioral assessments (n = 7).
General evaluation observed in the present in vivo study includes
daily observation of respiration, weight, hematuria, locomotor
deficits, other abnormal behaviors, skin/spinal tissue damage, and
inflammatory response (ED-1 marker for microglial activation) in
virus-infected spinal tissue (sham animals).
For siRNA pre-treatment for excitotoxic insult: Male Long-
Evans rats were randomly assigned to the following groups: (i)
CY-3-labeled ERK2 siRNA/lipofectamine 2000 with sham control
(laminectomy without injury), (ii) control siRNA/lipofectamine
2000 with sham control (laminectomy without injury), (iii) ERK2
siRNA/lipofectamine 2000 wtih excitotoxic insult, and (iv) control
siRNA/lipofectamine 2000 complexes with excitotoxic insult(n =
3/group). Fluorescent microscopy was used to observe whether
intrathecal delivery with CY-3-labeled ERK2 siRNA results in
CY-3 uptake in the spinal cord cells in uninjured rats (n = 3).
Immunofluorescent staining and cell counts were used to
determine whether intrathecal delivery with ERK2 siRNA inhibits
activation of ERK2 and caspase 3 in the spinal cord 4 h
following excitotoxic insult using lipofectamine 2000 delivery
system (three sections separated 60 lm apart per animal, three
rats per groups).
Statistical analysis was performed using StatView (SAS Institute,
Cary, NC, USA). Data are presented as mean ± SEM. Group
differences were evaluated by t-test or repeated measures ANOVA and
Bonferroni post hoc test (McTigtue et al. 2007). Null hypotheseswererejected at the p < 0.05 level. Although the BBB scale is an ordinal
scale, differences between the treatments were compared using
parametric statistical methods recommended by Scheff et al. (2002).
Results
ERK2 siRNA in vitroThe silencing efficacy of the ERK2 siRNA sequencewas validated on expression of the targeted mRNA and
Journal Compilation � 2010 International Society for Neurochemistry, J. Neurochem. (2010) 113, 131–142� 2010 The Authors
134 | C.-G. Yu et al.

protein levels in PC-12 cells using real-time PCRand western blots. Transfection of PC-12 cells withERK2 siRNA or control siRNA showed that ERK2siRNA (100 nm) significantly decreased levels of ERK2mRNA (Fig. 1a, p < 0.001, t-test) and ERK2 protein(Fig. 1b and c, p < 0.001, t-test) by 70% at 72 h aftertransfection in PC-12 cells compared with control siRNAtransfection.
ERK1/2 activation after contusion SCIWestern blot analysis showed that contusion injury signifi-cantly induced activation of ERK1/2 at 2 days after contu-sive SCI (Fig. 2a–c, p < 0.01, t-test).
LV-ERK2 shRNATo evaluate effects of spinal ERK2 knockdown followingSCI, we constructed ERK2 shRNA expression cassettes
(a)
(b)
(c)
Fig. 1 Silencing of extracellular signal-regulated protein kinase 2
(ERK2) with siRNA in vitro. (a) ERK2 siRNA decreased ERK2 mRNA
levels in PC-12 cells. PC-12 cells (3 · 105/well in 6-well plates) were
transfected with 100 nM ERK2 siRNA complexed with 6 ll lipofecta-
mine 2000. The mRNA levels of ERK2 in the cells were measured at
72 h after transfection using real-time PCR. ***p < 0.001, t-test, n = 4
per group. Dark bar = control siRNA, Light bar = ERK2siRNA (ERK2
siRNA: Sense (start 522) 5¢-pGCACCUCAGCAAUGAUCAUdTdT-3¢).(b and c) ERK2 siRNA decreased ERK2 protein levels in vitro. PC-12
cells (3 · 105/well in six-well plates) were transfected with ERK2
siRNA complexed with lipofectamine 2000 at 100 nm of siRNA and
liposome (6 lL). The protein levels of ERK2 in the cells were mea-
sured at 72 h after transfection using western blot analysis
(***p < 0.001, t-test, n = 4 per group).
44 KDa42 KDa
44 KDa42 KDa
pERK1pERK2
ERK1ERK2
0
0.5
1
1.5
2
2.5
3
ER
K2
phos
phor
ylat
ion
leve
l(p
ER
K2/
Tota
l ER
K2)
**
0
0.2
0.4
0.6
0.8
1
1.2E
RK
1 ph
osph
oryl
atio
n le
vel
(pE
RK
1/To
tal E
RK
1)
**
SCI Sham
Sham
Sham
SCI
SCI
(a)
(b)
(c)
Fig. 2 Contusive spinal cord injury (SCI) induces up-regulation of
extracellular signal-regulated protein kinase 1/2 (ERK1/2) phosphor-
ylation in the spinal cord 2 days after contusive SCI. Western blot
analysis showed injury-induced ERK1/2 phosphorylation of serine
residues of both ERK1 and ERK2 compared with sham-controls. The
bottom panels in the Fig. 2a show levels of total ERK1 and ERK2
protein expression, as loading controls. Antibodies on the top panels
were specific for the phosphorylated form of the ERK proteins.
**p < 0.001, t-test.
� 2010 The AuthorsJournal Compilation � 2010 International Society for Neurochemistry, J. Neurochem. (2010) 113, 131–142
ERK2 following spinal injury | 135

driven by the U6 promoter, cloned these into lentiviral-CMV-RNAi vectors and produced high-titer lentiviral ERK2shRNA or control vectors as described in protocols providedby Invitrogen. As both vectors contain an eGFP reporter,expression of LV-ERK2 shRNA or LV-control vectors wasverified in spinal cord of rats (Fig. 3a and b) by direct eGFPvisualization with fluorescent microscopy. Intraspinal micro-injection of LV-ERK2 shRNA resulted in eGFP expressionthroughout the gray matter and to a lesser extent in the whitematter of spinal cord at injection epicenter (�4 mm rostro-caudally, Fig. 3b). Intraspinal injection of LV-ERK2 shRNAreduced ERK2 protein levels, as compared to LV-control, asdemonstrated by immunohistochemistry (Fig. 3c–e) andwestern blots (Fig. 4a and b). LV-ERK2 shRNA constructsdid not decrease ERK1 levels (Fig. 4a top panel and c).These data indicate that intraspinal administration ofLV-ERK2 shRNA selective reduces ERK2 expression inthe rat spinal cord.
Following administration of the lentiviral particles insham control rats, we did not observe pathological orbehavioral alterations. Parameters examined included weightloss, hematuria, locomotor deficits, skin or spinal tissuedamage, and inflammatory response in spinal tissue. Immu-nofluorescent staining revealed that no obvious ED-1 signalwas seen in lentiviral vector-infected spinal tissue (data notshown).
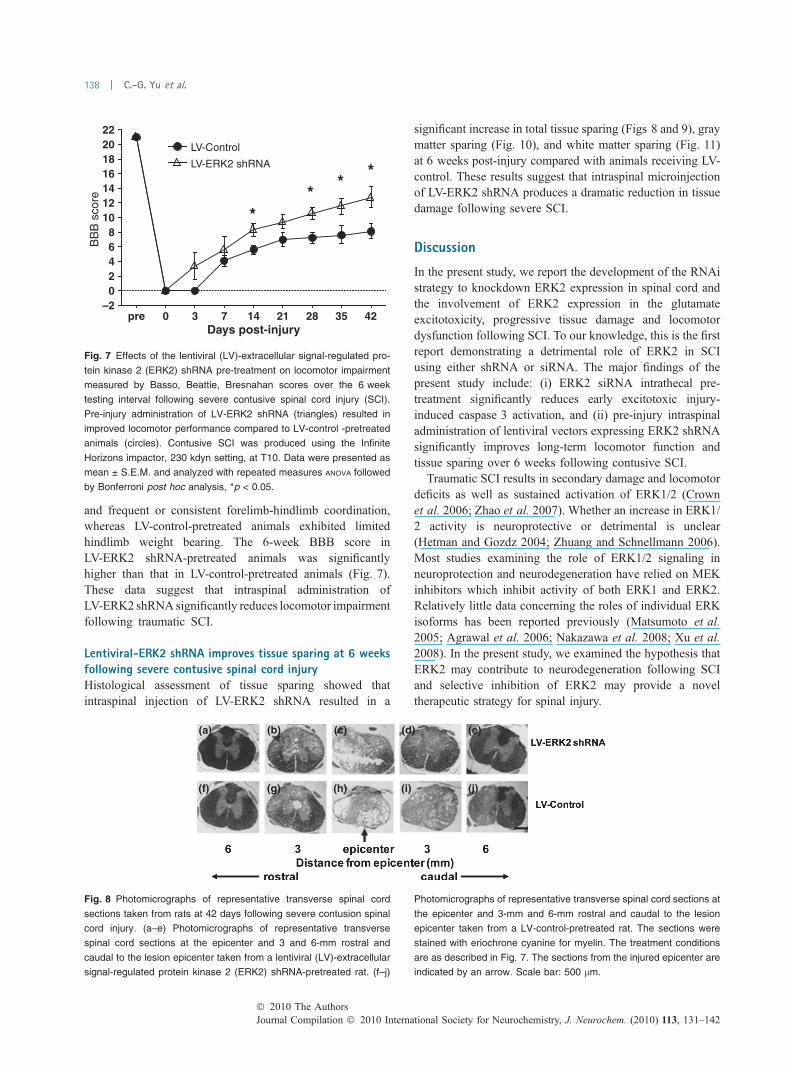
ERK2 siRNA reduces excitotoxic injuryDirect epifluorescent visualization demonstrated uptake ofCY-3-ERK2 siRNA throughout the spinal tissue 24 hfollowing intrathecal delivery (Fig. 5). Intrathecal pre-treat-ment for 72 h with ERK2 siRNA significantly reduced levelsof pERK2 (Fig. 6a–c) and activated caspase 3 (Fig. 6d–f) at4 h following excitotoxic insult.
(a) (b)
(c) (d)
(e)
Fig. 3 Lentiviral (LV)-extracellular signal-regulated protein kinase 2
(ERK2) shRNA reduces ERK2 expression in rat spinal cord. (a and b):
Intraspinal microinjection of LV-ERK2 shRNA (four injections in bilat-
eral gray matter and gray matter at T10) resulted in expression
throughout the spinal tissue (b), compared to naive control (a), as
visualized by the eGFP reporter. (c–e): Knockdown of ERK2 protein in
the spinal cord with LV-ERK2 shRNA treatment was evident using
Immunofluorescent staining (c, d, and e) with antibody specific for
ERK2. Scale bar: 400 lm. ***p < 0.001, t-test.
(a)
(b)
(c)
Fig. 4 Lentiviral (LV)-extracellular signal-regulated protein kinase 2
(ERK2) shRNA reduces ERK2 expression in rat spinal cord. Intraspi-
nal microinjection of LV-ERK2 shRNA (four injections in bilateral gray
matter and gray matter at T10) resulted in knockdown of ERK2 protein
in the spinal cord 2 weeks post-injury using western blotting (a and b)
with antibody specific for ERK2. These shRNA constructs do not alter
ERK1 levels using western blot with antibody against ERK1 (a, top
panel and c). *p < 0.05, t-test.
Journal Compilation � 2010 International Society for Neurochemistry, J. Neurochem. (2010) 113, 131–142� 2010 The Authors
136 | C.-G. Yu et al.
Lentiviral-ERK2 shRNA improves locomotor function6 weeks following contusive spinal cord injuryTo investigate the functional effects of LV-ERK2 shRNAon long-term locomotor deficits following traumatic SCI,fourteen lentiviral vector-infected rats received contusiveinjury using the IH impactor at the 230 kdyn force setting.No significant differences in actual force, displacement, orvelocity were found between LV-ERK2 shRNA andLV-control-treated groups, indicating similar injuries toall animals (Table 1). Immediately following SCI, allanimals exhibited complete bilateral hindlimb paralysis.Behavioral assessment demonstrated that administration ofLV-ERK2 shRNA at 7 days pre-injury resulted in signif-icant improvement in locomotor function measured byBBB scores over 6 weeks after severe contusive SCIcompared with the administration of LV-control (Fig. 7,
p < 0.05 at 2, 4, 5, and 6 weeks post-injury, repeatedmeasures ANOVA and Bonferroni post hoc test). At 6-weekspost-injury, most LV-ERK2 shRNA-pretreated animalsexhibited consistent weight-supported hindlimb movements
Sham control CY-3 ERK2 siRNA
(a) (b) (c)
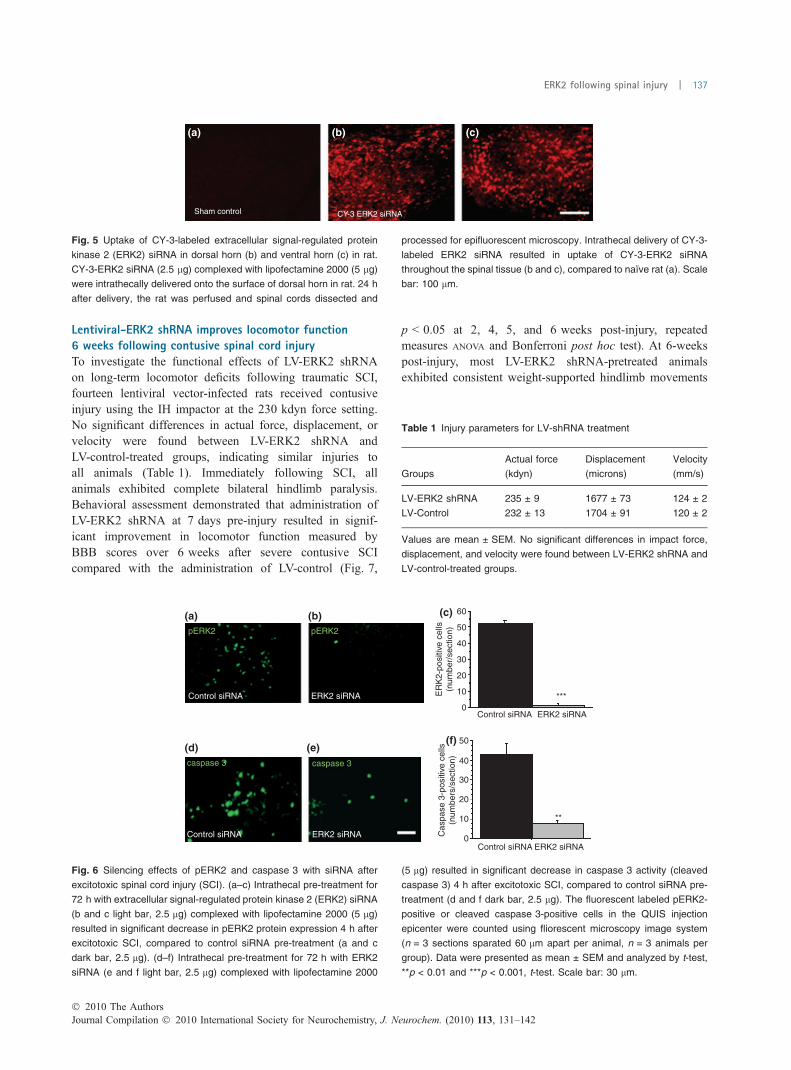
Fig. 5 Uptake of CY-3-labeled extracellular signal-regulated protein
kinase 2 (ERK2) siRNA in dorsal horn (b) and ventral horn (c) in rat.
CY-3-ERK2 siRNA (2.5 lg) complexed with lipofectamine 2000 (5 lg)
were intrathecally delivered onto the surface of dorsal horn in rat. 24 h
after delivery, the rat was perfused and spinal cords dissected and
processed for epifluorescent microscopy. Intrathecal delivery of CY-3-
labeled ERK2 siRNA resulted in uptake of CY-3-ERK2 siRNA
throughout the spinal tissue (b and c), compared to naı̈ve rat (a). Scale
bar: 100 lm.
(a) (b) (c)
(d) (e)(f)
0Control siRNA ERK2 siRNA
Control siRNA
Control siRNA
ERK2 siRNA
ERK2 siRNA
Control siRNA
pERK2 pERK2
caspase 3 caspase 3
ERK2 siRNA
***
**
ER
K2-
posi
tive
cells
(num
ber/
sect
ion)
Cas
pase
3-p
ositi
ve c
ells
(num
bers
/sec
tion)
10
20
30
40
50
0
10
20
30
40
50
60
Fig. 6 Silencing effects of pERK2 and caspase 3 with siRNA after
excitotoxic spinal cord injury (SCI). (a–c) Intrathecal pre-treatment for
72 h with extracellular signal-regulated protein kinase 2 (ERK2) siRNA
(b and c light bar, 2.5 lg) complexed with lipofectamine 2000 (5 lg)
resulted in significant decrease in pERK2 protein expression 4 h after
excitotoxic SCI, compared to control siRNA pre-treatment (a and c
dark bar, 2.5 lg). (d–f) Intrathecal pre-treatment for 72 h with ERK2
siRNA (e and f light bar, 2.5 lg) complexed with lipofectamine 2000
(5 lg) resulted in significant decrease in caspase 3 activity (cleaved
caspase 3) 4 h after excitotoxic SCI, compared to control siRNA pre-
treatment (d and f dark bar, 2.5 lg). The fluorescent labeled pERK2-
positive or cleaved caspase 3-positive cells in the QUIS injection
epicenter were counted using fliorescent microscopy image system
(n = 3 sections sparated 60 lm apart per animal, n = 3 animals per
group). Data were presented as mean ± SEM and analyzed by t-test,
**p < 0.01 and ***p < 0.001, t-test. Scale bar: 30 lm.
Table 1 Injury parameters for LV-shRNA treatment
Groups
Actual force
(kdyn)
Displacement
(microns)
Velocity
(mm/s)
LV-ERK2 shRNA 235 ± 9 1677 ± 73 124 ± 2
LV-Control 232 ± 13 1704 ± 91 120 ± 2
Values are mean ± SEM. No significant differences in impact force,
displacement, and velocity were found between LV-ERK2 shRNA and
LV-control-treated groups.
� 2010 The AuthorsJournal Compilation � 2010 International Society for Neurochemistry, J. Neurochem. (2010) 113, 131–142
ERK2 following spinal injury | 137
and frequent or consistent forelimb-hindlimb coordination,whereas LV-control-pretreated animals exhibited limitedhindlimb weight bearing. The 6-week BBB score inLV-ERK2 shRNA-pretreated animals was significantlyhigher than that in LV-control-pretreated animals (Fig. 7).These data suggest that intraspinal administration ofLV-ERK2 shRNA significantly reduces locomotor impairmentfollowing traumatic SCI.
Lentiviral-ERK2 shRNA improves tissue sparing at 6 weeksfollowing severe contusive spinal cord injuryHistological assessment of tissue sparing showed thatintraspinal injection of LV-ERK2 shRNA resulted in a
significant increase in total tissue sparing (Figs 8 and 9), graymatter sparing (Fig. 10), and white matter sparing (Fig. 11)at 6 weeks post-injury compared with animals receiving LV-control. These results suggest that intraspinal microinjectionof LV-ERK2 shRNA produces a dramatic reduction in tissuedamage following severe SCI.
Discussion
In the present study, we report the development of the RNAistrategy to knockdown ERK2 expression in spinal cord andthe involvement of ERK2 expression in the glutamateexcitotoxicity, progressive tissue damage and locomotordysfunction following SCI. To our knowledge, this is the firstreport demonstrating a detrimental role of ERK2 in SCIusing either shRNA or siRNA. The major findings of thepresent study include: (i) ERK2 siRNA intrathecal pre-treatment significantly reduces early excitotoxic injury-induced caspase 3 activation, and (ii) pre-injury intraspinaladministration of lentiviral vectors expressing ERK2 shRNAsignificantly improves long-term locomotor function andtissue sparing over 6 weeks following contusive SCI.
Traumatic SCI results in secondary damage and locomotordeficits as well as sustained activation of ERK1/2 (Crownet al. 2006; Zhao et al. 2007). Whether an increase in ERK1/2 activity is neuroprotective or detrimental is unclear(Hetman and Gozdz 2004; Zhuang and Schnellmann 2006).Most studies examining the role of ERK1/2 signaling inneuroprotection and neurodegeneration have relied on MEKinhibitors which inhibit activity of both ERK1 and ERK2.Relatively little data concerning the roles of individual ERKisoforms has been reported previously (Matsumoto et al.2005; Agrawal et al. 2006; Nakazawa et al. 2008; Xu et al.2008). In the present study, we examined the hypothesis thatERK2 may contribute to neurodegeneration following SCIand selective inhibition of ERK2 may provide a noveltherapeutic strategy for spinal injury.
–202468
10121416182022
BB
B s
core
pre 0 3 7 14 21 28 35 42
LV-ERK2 shRNA
LV-Control
**
**
Days post-injury
Fig. 7 Effects of the lentiviral (LV)-extracellular signal-regulated pro-
tein kinase 2 (ERK2) shRNA pre-treatment on locomotor impairment
measured by Basso, Beattie, Bresnahan scores over the 6 week
testing interval following severe contusive spinal cord injury (SCI).
Pre-injury administration of LV-ERK2 shRNA (triangles) resulted in
improved locomotor performance compared to LV-control -pretreated
animals (circles). Contusive SCI was produced using the Infinite
Horizons impactor, 230 kdyn setting, at T10. Data were presented as
mean ± S.E.M. and analyzed with repeated measures ANOVA followed
by Bonferroni post hoc analysis, *p < 0.05.
(a) (b) (c) (d) (e)
(f) (g) (h) (i) (j)
Fig. 8 Photomicrographs of representative transverse spinal cord
sections taken from rats at 42 days following severe contusion spinal
cord injury. (a–e) Photomicrographs of representative transverse
spinal cord sections at the epicenter and 3 and 6-mm rostral and
caudal to the lesion epicenter taken from a lentiviral (LV)-extracellular
signal-regulated protein kinase 2 (ERK2) shRNA-pretreated rat. (f–j)
Photomicrographs of representative transverse spinal cord sections at
the epicenter and 3-mm and 6-mm rostral and caudal to the lesion
epicenter taken from a LV-control-pretreated rat. The sections were
stained with eriochrone cyanine for myelin. The treatment conditions
are as described in Fig. 7. The sections from the injured epicenter are
indicated by an arrow. Scale bar: 500 lm.
Journal Compilation � 2010 International Society for Neurochemistry, J. Neurochem. (2010) 113, 131–142� 2010 The Authors
138 | C.-G. Yu et al.

–0.2
0
0.2
0.4
0.6
0.8
1
1.2
Gra
y m
atte
r sp
arin
g (m
m2 )
–7 –6 –5 –4 –3 –2 –1 0 1 2 3 4 5 6 7
LV-ERK2 shRNA
LV-Control
Rostral Epicenter CaudalDistance from epicenter (mm)
*
**
*****
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
Tota
l gra
y m
atte
r sp
arin
g (m
m3 )
**
LV-Control LV-ERK2 shRNA
(a) (b)
Fig. 10 Effects of the LV-extracellular signal-regulated protein kinase
2 (ERK2) shRNA pre-treatment on gray matter sparing 6 weeks fol-
lowing severe contusive spinal cord injury. Pre-injury administration of
LV-ERK2 shRNA resulted in a significant increase in total gray matter
sparing (a) and gray matter sparing at 3–6 mm caudal to the injury
epicenter (b) following contusion injury to the spinal cord. Injury con-
ditions and treatment groups are as described in Fig. 7. Data were
presented as mean ± SEM and analyzed by t-test (a) or repeated
measures ANOVA followed by Bonferroni post hoc analysis (b).
*p < 0.05, **p < 0.01, ***p < 0.001.
0
0.5
1
1.5
2
2.5
3
3.5
4
Tota
l tis
sue
spar
ing
(mm
3 )
*
LV-Control LV-ERK2 shRNA
–0.5
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
Tis
sue
spar
ing
(mm
2 )
–7 –6 –5 –4 –3 –2 –1 0 1 2 3 4 5 6 7
LV-ERK2 shRNA
LV-Control
Rostral Epicenter CaudalDistance from epicenter (mm)
*
* **
*(a) (b)
Fig. 9 Effects of the lentiviral (LV)-extracellular signal-regulated
protein kinase 2 (ERK2) shRNA pre-treatment on tissue sparing
6 weeks following severe contusive spinal cord injury. Pre-injury
administration of LV-ERK2 shRNA resulted in a significant increase
in total tissue sparing (a) and tissue sparing at 3–6 mm caudal to the
injury epicenter (b) following contusion injury to the spinal cord. Injury
conditions and treatment groups are as described in Fig. 7. Data
were presented as mean ± SEM and analyzed by t-test (a) or re-
peated measures ANOVA followed by Bonferroni post hoc analysis (b).
*p < 0.05, **p < 0.01.
–0.5
0
0.5
1
1.5
2
2.5
3
3.5
Whi
te m
atte
r sp
arin
g (m
m2 )
–7 –6 –5 –4 –3 –2 –1 0 1 2 3 4 5 6 7
LV-ERK2 shRNALV-Control
Rostral Epicenter CaudalDistance from epicenter (mm)
*
*
* * *
0
0.5
1
1.5
2
2.5
3
3.5
Tota
l whi
te m
atte
r sp
arin
g (m
m3 )
LV-Control LV-ERK2 shRNA
(a) (b)
Fig. 11 Effects of the LV-extracellular signal-regulated protein kinase
2 (ERK2) shRNA pre-treatment on white matter sparing 6 weeks fol-
lowing severe contusive spinal cord injury. Pre-injury administration of
LV-ERK2 shRNA resulted in a significant increase in white matter
tissue sparing at 3–6 mm caudal to the injury epicenter (b, 300% in-
crease at 3 mm, 100% increase at 4 mm, 89% increase at 5 mm, and
64% increase at 6 mm caudal to the epicenter) and non-significant
increase in total white matter sparing (a) following contusion injury to
the spinal cord. Injury conditions and treatment groups are as de-
scribed in Fig. 7. Data were presented as mean ± SEM and analyzed
by t-test (a) or repeated measures ANOVA and Bonferroni post hoc
analysis (b). *p < 0.05.
� 2010 The AuthorsJournal Compilation � 2010 International Society for Neurochemistry, J. Neurochem. (2010) 113, 131–142
ERK2 following spinal injury | 139

Selective inhibitor for ERK2 is not available. To test thehypothesis that ERK2 expression is involved in the patho-genesis of SCI, we utilized intraspinal microinjection oflentiviral-ERK2 shRNA vectors to knockdown ERK2expression in the spinal cord prior to contusive SCI in rats.We initially evaluated ERK2 siRNA efficacy in PC-12 cells,then examined intrathecal delivery of ERK2 siRNA into thespinal cord in vivo using CY-3 labeled ERK2 siRNA.Although intravenous administration of lipid-based transfec-tion reagents is problematic because of rapid liver metabo-lism, such agents have been used successfully for intrathecalsiRNA administration (Luo et al. 2005; Aigner 2006).Intrathecal ERK2 siRNA reduced activation of ERK2 andcaspase 3 at 4 h after excitotoxic insult. These resultsprovided support for the hypothesis that ERK2 inhibitionmay protect against neurodegeneration following SCI.
For subsequent studies, we utilized Lentiviral-shRNAvectors which have the ability to infect both dividing andnon-dividing cells and are an effective method for RNAi inpost-mitotic cells, with a longer time course of gene knock-down and less inflammatory/immune response (Rubinsonet al. 2003; Breckpot et al. 2007; Meunier et al. 2008).Lentiviral vector-based RNAi has been approved in clinicaltrials for gene therapy in HIV, human beta-thalassemia, andsickle-cell anemia with no obvious side effects (Chen et al.2007). However, the use of lentiviral vector expressingshRNA for analysis of individual gene function and genetherapy in SCI has not been reported. This study developedlentiviral-mediated delivery of ERK2 shRNA in vivo andverified the efficacy of this approach in locally reducing ERK2expression in the spinal cord. In the present study, no sideeffects including spinal tissue damage, weight loss, hematuria,locomotor deficits, other abnormal behaviors, or inflammatoryresponse in spinal tissue were observed after lentiviral-ERK2shRNA intraspinal administration in uninjured controls. Theimportant finding of the present study is that selective ERK2knockdown by intraspinal injection of lentiviral-ERK2shRNA (pre-injury) resulted in significant improvement inlocomotor function and tissue sparing over 6 weeks followingsevere contusive SCI using IH SCI impact device.
The IH impact device-induced contusive SCI at T10 is aclinically relevant model of thoracic traumatic SCI (Scheffet al. 2003). The 230 kdyn force setting used in the presentstudy results in severe secondary damage and locomotordysfunction (Scheff et al. 2003). The final outcome of SCIdirectly depends on the degree of the initial traumatic insultsand the secondary damage (Scheff et al. 2003). In the severecontusion injury model, the BBB score assessment andhistological assessment demonstrated that LV-ERK2 shRNApre-treatment resulted in significant improvement in loco-motor function and tissue sparing over 6 weeks post-injury.These data suggest that ERK2 knockdown with LV-ERK2shRNA intraspinal administration is a highly potent approach
for long-term recovery of locomotor dysfunction and tissuedamage after SCI.
Secondary tissue damage after SCI spreads both rostraland caudal to the lesion epicenter with increasing functionaldeficits (Wells et al. 2003). Tissue loss, including loss ofneurons (gray matter) and/or axons (white matter), is theunderlying cause of permanent disability following cervicaland/or thoracic SCI (el-Bohy et al. 1998; Magnuson et al.1999; Reier et al. 2002). In the T10 contusive injury model,it is likely that only white matter sparing contributes to thefunctional improvement because the locomotor deficits atthis level of injury are largely due to demyelination andloss of white matter axons (Blight 1983; Teng and Wrathall1997; Magnuson et al. 1999; Reier et al. 2002). Gray matterdamage at the thoracic level has relatively minor conse-quences, including loss of sensory input from one or twodermatomes and deficits in motor output to some axialmusculature (Magnuson et al. 1999). Preservation of as fewas 5–10% of myelinated axons in white mater can conferlocomotor recovery (Azari et al. 2006). Thus, strategiesaimed at improving white matter sparing by ERK2 knock-down are likely to be of therapeutic significance. On theother hand, very small sparing of tissue at the lesionepicenter or rostral/caudal to the lesion site has profoundeffects on locomotor recovery (Basso et al. 1996; Wrathallet al. 1996, 1997; Lankhorst et al. 1999; Rabchevsky et al.2000; Hillard et al. 2004). In the present study, although totalwhite matter sparing was not statistically different betweenthe groups, 300–64% increase in white matter sparing at3–6 mm caudal to the lesion epicenter was observed withintraspinal administration of LV-ERK2 shRNA. Theimprovement in locomotor function obtained with intraspinalLV-ERK2 shRNA is presumably associated with whitematter sparing caudal to the lesion epicenter. These findingsare similar to those observed with other pharmacologicalinterventions in studies of spinal cord contusion injury byother investigators and may be responsible for the functionalrecovery (Wrathall et al. 1996, 1997; Teng and Wrathall1997; Madsen et al. 1998; Lankhorst et al. 1999; Rabchev-sky et al. 2000).
The tissue sparing achieved with intraspinal administrationof LV-ERK2 shRNA was primarily caudal to the injury.Although the reason for this marked rostral and caudaldifferences is uncertain, it may be related to the followingfactors: (i) an unequal local gene knockdown in differentcells (Meunier et al. 2008), (ii) an unequal local expression/activation of ERK2 downstream targeted genes (Zhao et al.2007), and/or, (iii) an unequal local gain in blood perfusion(Oudega et al. 1999). Further studies in the future arenecessary to examine the local pattern in eGFP expression,cell type-selective ERK2 knockdown, and downstreamtargeted gene expression/activation with LV-ERK2 shRNAvectors in the spinal cord after SCI.
Journal Compilation � 2010 International Society for Neurochemistry, J. Neurochem. (2010) 113, 131–142� 2010 The Authors
140 | C.-G. Yu et al.

Although the mechanisms of pathological and behavioraldysfunction after SCI are not fully understood, it is knownthat SCI activates ERK1/2 that targets multiple secondaryinjury events including glutamate excitotoxicity, inflamma-tion, apoptosis, pain hypersensitivity, and mitochondrialdysfunction (Klussmann and Martin-Villalba 2005; Yu andYezierski 2005; Barrett et al. 2006; Genovese et al. 2008).Glutamatergic excitotoxicity is believed to be a majormechanism in neurotrauma (Park et al. 2004). Strategiesaimed at blocking the deleterious effects of ERK signalingleading to inhibition of glutamatergic cascades may be oftherapeutic significance. In the present study, we showed thatselective ERK2 knockdown with ERK2 siRNA significantlyreduced glutamatergic signaling-mediated caspase 3 activa-tion after excitotoxic insult (Yu et al. 2009). The presentfindings suggest that ERK2 activation after SCI contributesto activation of glutamatergic cascades, resulting in second-ary damage and locomotor deficits.
The major objective of the present study was to investigatethe pathological contributions of ERK2 in the secondarydamage following contusive SCI. Although pre-treatment withlentiviral vectors before SCI is not a viable therapeutic strategy,the strategy enabled examination of the role of ERK2 isoform,which is not possible using current pharmacologic inhibitors.The lentiviral vectors were administered pre-injury to allowsufficient time for genomic integration of the retroviral cDNAand the decreased gene and protein expression.
Treating SCI remains a medical challenge and developingnew therapeutic strategies is clinically important. Our studyprovides a proof-of-concept that ERK2 knockdown canreduce tissue damage and locomotor impairment, therebyrevealing a detrimental role for ERK2 pathway in secondaryinjury and paving the way for further investigation with post-injury administration of lentiviral shRNA or siRNA targetingERK2 in the pathophysiology and treatment of SCI.
Acknowledgements
This research was supported by the Paralysis Project of America, the
Kentucky Spinal Cord and Head Injury Research Trust, and a
University of Florida Seed grant. The authors wish to thank Dr
George Smith for lentiviral-CMV-RNAi vectors, and Paula Thoma-
son for manuscript editing.
References
Agrawal A., Dillon S., Denning T. L. and Pulendran B. (2006) ERK1-/-mice exhibit Th1 cell polarization and increased susceptibility toexperimental autoimmune encephalomyelitis. J. Immunol. 176,5788–5796.
Aigner A. (2006) Gene silencing through RNA interference (RNAi)in vivo: strategies based on the direct application of siRNAs.J. Biotechnol.. 124, p. 12–25.
Azari M. F., Prolyri S. C., Karnezis T., Bernard C. C., Small D. H.,Cheema S. S., Ozturk E., Hatzinisirious I. and Petratos S. (2006)Leukemia inhibitory factor arrests oligodendrocyte death and
demyelination in spinal cord injury. J. Neuropathol. Exp. Neurol.65, 914–929.
Barrett L. E., Van Bockstaele E. J., Sul J. Y., Takano H., Haydon P. G.and Eberwine J. H. (2006) Elk-1 associates with the mitochondrialpermeability transition pore complex in neurons. Proc. Natl Acad.Sci. USA 103, 5155–5160.
Basso D. M., Beattie M. S. and Bresnahan J. C. (1996) Graded histo-logical and locomotor outcomes after spinal cord contusion usingthe NYU weight-drop device versus transection. Exp. Neurol. 139,244–256.
Blight A. R. (1983) Cellular morphology of chronic spinal cord injury inthe cat: analysis of myelinated axons by line-sampling. Neurosci-ence 10, 521–543.
el-Bohy A. A., Schrimsher G. W., Reier J. P. and Goshgarian H. G. (1998)Quantitative assessment of respiratory function following contusioninjury of the cervical spinal cord. Exp. Neurol. 150, 143–152.
Breckpot K., Aerts J. L. and Thielemans K. (2007) Lentiviral vectors forcancer immunotherapy: transforming infectious particles intotherapeutics. Gene Ther. 14, 847–862.
Cai Y., Chen S., Han H., Sood A. K., Lopex-Berestein G. and Pan H.(2009) Role of M2, M3, and M4 muscarinic receptor subtypes inthe spinal cholonergic control of nociception revealed using siRNAin rats. J. Neurochem. 111, 1000–1010.
Chen Z., Gibson T. B., Robinson F., Silvestro L., Pearson G., Xu B.,Wright A., Vanderbilt C. and Cobb M. H. (2001) MAP kinases.Chem Rev. 101, 2449–2476.
Chen Y., Lin M. C., Yao H. et al. (2007) Lentivirus-mediated RNAinterference targeting enhancer of zeste homolog 2 inhibits hepa-tocellular carcinoma growth through down-regulation of stathmin.Hepatology 46, 200–208.
Crown E. D., Ye Z., Johnson K. M., Xu G. Y., McAdoo D. J. andHulsebosch C. E. (2006) Increases in the activated forms of ERK1/2, p38 MAPK, and CREB are correlated with the expression ofat-level mechanical allodynia following spinal cord injury. Exp.Neurol. 199, 397–407.
French D. D., Campbell R. R., Sabharwal S., Nelson A. L., PalaciosP. A. and Gavin-Dreschnack D. (2007) Health care costs forpatients with chronic spinal cord injury in the Veterans HealthAdministration. J. Spinal Cord Med. 30, 477–481.
Genovese T., Esposito E., Mazzon E., Muia C., Di Paola R., Meli R.,Bramanti P. and Cuzzocrea S. (2008) Evidence for the role ofmitogen-activated protein kinase signaling pathways in thedevelopment of spinal cord injury. J. Pharmacol. Exp. Ther. 325,100–114.
Hetman M. and Gozdz A. (2004) Role of extracellular signal regulatedkinases 1 and 2 in neuronal survival. Eur. J. Biochem. 271, 2050–2055.
Hillard V. H., Peng H., Zhang Y., Das K., Murali R., Etlinger J. D. andZeman R. J. (2004) Tempol, a nitroxide antioxidant, improveslocomotor and histological outcomes after spinal cord contusion inrats. J. Neurotrauma 21, 1405–1414.
Klussmann S. and Martin-Villalba A. (2005) Molecular targets in spinalcord injury. J. Mol. Med. 83, 657–671.
Lankhorst A. J., Duis S. E., Ter Laak M. P., Joosten E. A., Hamers F. P.and Gispen W. H. (1999) Functional recovery after central infusionof alpha-melanocyte-stimulating hormone in rats with spinal cordcontusion injury. J. Neurotrauma 16, 323–331.
Lu P. Y. and Woodle M. C. (2008) Delivering small interfering RNA fornovel therapeutics. Methods Mol. Biol. 437, 93–107.
Luo M. C., Zhang D. Q., Ma S. W., Huang Y. Y., Shuster S. J., Porreca F.and Lai J. (2005)An efficient intrathecal delivery of small interferingRNA to the spinal cord and peripheral neurons.Mol. Pain. 1, 29.
Madsen J. R., MacDonald P., Irwin N., Goldberg D. E., Yao G. L., MeiriK. F., Rimm I. J., Stieg P. E. and Benowitz L. I. (1998) Tacrolimus
� 2010 The AuthorsJournal Compilation � 2010 International Society for Neurochemistry, J. Neurochem. (2010) 113, 131–142
ERK2 following spinal injury | 141

(FK506) increases neuronal expression of GAP-43 and improvesfunctional recovery after spinal cord injury in rats. Exp. Neurol.154, 673–683.
Magnuson D. S. K., Trinder T. C., Zhang Y. P., Burke D., Morassutti D.J. and Shields C. B. (1999) Comparing deficits following excito-toxic and contusion injuries in the thoracic and lumber spinal cordof the adult rat. Exp. Neurol. 156, 191–204.
Matsumoto S., Miyagishi M., Akashi H., Nagai R. and Taira K. (2005)Analysis of double-stranded RNA-induced apoptosis pathwaysusing interferon-response noninducible small interfering RNAexpression vector library. J. Biol. Chem. 280, 25687–25696.
McTigtue D. M., Tripathi R., Wei P. and Lash A. T. (2007) The PPARgamma agonist pioglitazone improves anatomical and locomotorrecovery after rodent spinal cord injury. Exp. Neurol. 205,396–406.
Meunier A., Mauborgne A., Masson J., Mallet J. and Pohl M. (2008)Lentiviral-mediated targeted transgene expression in dorsal spinalcord glia: tool for the study of glial cell implication in mechanismsunderlying chronic pain development. J. Neurosci. Methods 167,148–159.
Nakazawa T., Shimura M., Ryu M., Nishida K., Pages G., Pouyssegur J.and Endo S. (2008) ERK1 plays a critical protective role against N-methyl-D-aspartate-induced retinal injury. J. Neurosci. Res. 86,136–144.
Oudega M., Vargas C. G., Weber A. B., Kleitman N. and Bunge M. B.(1999) Long-term effects of methylprednisolone followingtransfection of adult rat spinal cord. Eur. J. Neurosci. 11, 2453–2464.
Park E., Velumian A. A. and Fehlings M. G. (2004) The role ofexcitotoxicity in secondary mechanisms of spinal cord injury: areview with an emphasis on the implications for white matterdegeneration. J. Neurotrauma 21, 754–774.
Rabchevsky A. G., Fugaccia I., Turner A. F., Blades D. A., Mattson M.P. and Scheff S. W. (2000) Basic fibroblast growth factor (bFGF)enhances functional recovery following severe spinal cord injury tothe rat. Exp. Neurol. 164, 280–291.
Rabchevsky A. G., Fugaccia I., Sullivan P. G., Blades D. A. and ScheffS. W. (2002) Efficacy of methylprednisolone therapy for theinjured rat spinal cord. J. Neurosci. Res. 68, 7–18.
Reier P. J., Golder F. J., Bolser D. C., Hubscher C., Johnson R.,Schrimsher G. W. and Velardo M. J. (2002) Gray matter repair inthe cervical spinal cord. Prog. Brain Res. 137, 49–70.
Reynolds A., Leake D., Boese Q., Scaringe S., Marshall W. S. andKhvorova A. (2004) Rational siRNA design for RNA interference.Nat Biotechnol. 22, 326–330.
Rubinson D. A., Dillon C. P., Kwiatkowski A. V. et al. (2003) A len-tivirus-based system to functionally silence genes in primarymammalian cells, stem cells and transgenic mice by RNA inter-ference. Nat. Genet. 33, 401–406.
Ruitenberg M. J., Eggers R., Boer G. J. and Verhaagen J. (2002) Adeno-associated viral vectors as agents for gene delivery: application indisorders and trauma of the central nervous system. Methods 28,182–194.
Scheff S. W., Saucier D. A. and Cain M. E. (2002) A statistical methodfor analyzing rating scale data: the BBB locomotor score.J. Neurotrauma 19, 1251–1260.
Scheff S.W., RabchevskyA.G., Fugaccia I.,Main J. A. and Lumpp J. E. Jr(2003) Experimental modeling of spinal cord injury: characteriza-tion of a force-defined injury device. J. Neurotrauma 20, 179–193.
Teng Y. D. and Wrathall J. R. (1997) Local blockade of sodium channelsby tetrodotoxin ameliorates tissue loss and long-term functionaldeficits resulting from experimental spinal cord injury. J. Neurosci.17, 4359–4366.
Wells J. E., Hurlbert R. J., Fehlings M. G. and Yong V. W. (2003)Neuroprotection by minocycline facilitates significant recoveryfrom spinal cord injury in mice. Brain 126, 1628–1637.
Wrathall J. R., Teng Y. D. and Choiniere D. (1996) Amelioration offunctional deficits from spinal cord trauma with systemicallyadministered NBQX, an antagonist of non-N-methyl-D-aspartatereceptors. Exp. Neurol. 137, 119–126.
Wrathall J. R., Teng Y. D. and Marriott R. (1997) Delayed antago-nism of AMPA/kainate receptors reduces long-term functionaldeficits resulting from spinal cord trauma. Exp. Neurol. 145,565–573.
Xu Q., Garraway S. M., Weyerbacher A. R., Shin S. J. and Inturrisi C. E.(2008) Activation of the neuronal extracellular signal-regulatedkinase 2 in the spinal cord dorsal horn is required for completeFreund’s adjuvant-induced pain hypersensitivity. J. Neurosci. 28,14087–14096.
Yu C. G. and Geddes J. W. (2007) Sustained calpain inhibition improveslocomotor function and tissue sparing following contusive spinalcord injury. Neurochem. Res. 32, 2046–2053.
Yu C. G. and Yezierski R. P. (2005) Activation of the ERK1/2 signalingcascade by excitotoxic spinal cord injury. Brain Res. Mol. BrainRes. 138, 244–255.
Yu C. G., Yezierski R. P., Joshi A., Raza K., Li Y. and Geddes J. W..(2009). RNAi approach reveals a specific role of ERK2 in thedeleterious consequences of spinal cord injury. J. Neurotrauma26(8): A-73. Online (Abstract from The Second Joint Symposiumof The National and International Neurotrauma Societies,September 7–11, 2009, Santa Barbara, CA).
Zhao P., Waxman S. G. and Hains B. C. (2007) Extracellular signal-regulated kinase-regulated microglia-neuron signaling byprostaglandin E2 contributes to pain after spinal cord injury.J. Neurosci. 27, 2357–2368.
Zhuang S. and Schnellmann R. G. (2006) A death-promoting role forextracellular signal-regulated kinase. J. Pharmacol. Exp. Ther. 319,991–997.
Journal Compilation � 2010 International Society for Neurochemistry, J. Neurochem. (2010) 113, 131–142� 2010 The Authors
142 | C.-G. Yu et al.
Copyright © 2022 FDOKUMEN




















