
Blocking EphA4 upregulation after spinal cord injury results in enhanced chronic pain
Upload
independentCategory
view
9download
0
GRS
GARa
gb
ic
i
Arpgcdodt8m(rd(nibgtSflvnbGSayTtir
*TfEAagpti
Neuroscience 130 (2005) 639–649
0d
LUTATHIONE MONOETHYL ESTER IMPROVES FUNCTIONALECOVERY, ENHANCES NEURON SURVIVAL, AND STABILIZES
PINAL CORD BLOOD FLOW AFTER SPINAL CORD INJURY IN RATSKp
Tnomsintar
Hihawordp2
mtse(ciht12
tIct12imuW
. GUÍZAR-SAHAGÚN,a,b* A. IBARRA,a,b A. ESPITIA,b
. MARTÍNEZ,b I. MADRAZOb AND
. E. FRANCO-BOURLANDb,c
Research Unit for Neurological Diseases, Instituto Mexicano del Se-uro Social, Mexico City, Mexico
IMSS-Proyecto Camina A.C., Tlalpan 4430, Mexico City 14050, Mex-co
Department of Biochemistry, Centro Nacional de Rehabilitación, Mex-co City, Mexico
bstract—Secondary damage after spinal cord (SC) injuryemains without a clinically effective drug treatment. To ex-lore the neuroprotective effects of cell-permeable reducedlutathione monoethyl ester (GSHE), rats subjected to SContusion using the New York University impactor were ran-omly assigned to receive intraperitoneally GSHE (total dosef 12 mg/kg), methylprednisolone sodium succinate (totalose of 120 mg/kg), or saline solution as vehicle. Motor func-ion, assessed using the Basso–Beattie–Bresnahan scale for
weeks, was significantly better in GSHE (11.2�0.6,ean�S.E.M., n�8, at 8 weeks) than methylprednisolone
9.3�0.6) and vehicle (9.4�0.7) groups. The number of neu-ons in the red nuclei labeled with FluoroRuby placed cau-ally to the injury site was significantly higher in GSHE158�9.3 mean�S.E.M., n�4) compared with methylpred-isolone (53�14.7) and vehicle (46�16.4) groups. Differences
n the amount of spared SC tissue at the epicenter and neigh-oring areas were not significant among experimentalroups. In a second series of experiments, using similarreatment groups (n�6), regional changes in microvascularC blood flow were evaluated for 100 min by laser-Dopplerowmetry after clip compression injury. SC blood flow fell inehicle-treated rats 20% below baseline and increased sig-ificantly with methylprednisolone approximately 12% aboveaseline; changes were not greater than 5% in rats givenSHE. In conclusion, GSHE given to rats early after moderateC contusion/compression improves functional outcomend red nuclei neuron survival significantly better than meth-lprednisolone and vehicle, and stabilizes SC blood flow.hese results support further investigation of reduced gluta-hione supplementation after acute SC injury for future clin-cal application. © 2005 IBRO. Published by Elsevier Ltd. Allights reserved.
Correspondence to: G. Guı́zar-Sahagún, IMSS-Proyecto Camina,lalpan 4430, Mexico City 14050, Mexico. Tel: �52-55-5573-0029;
ax: �52-55-5573-5545.-mail address: [email protected] (G. Guı́zar-Sahagún).bbreviations: BBB, Basso–Beattie–Bresnahan; GFAP, glial fibrillarycidic protein; GSH, glutathione in its reduced form; GSHE, reducedlutathione monoethyl ester; LPO, lipid peroxidation; MPSS, methyl-rednisolone sodium succinate; PBS, phosphate buffer saline solu-
mion; SC, spinal cord; SCBF, spinal cord blood flow; SCI, spinal cordnjury.
306-4522/05$30.00�0.00 © 2005 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2004.09.056
639
ey words: antioxidant, ischemia, microcirculation, neuro-rotective, oxidative stress, secondary damage.
rauma to spinal cord (SC) causes primary damage toeural and vascular tissues, and is followed by a cascadef secondary autodestructive events that worsen the pri-ary lesion increasing the likelihood of irreversible motor,
ensory and autonomic dysfunctions. Oxidative stress andschemia are considered, among others, as major mecha-isms for secondary injury (Anderson and Hall, 1993; Ta-or, 1995; Kaynar et al., 1998; Dumont et al., 2001; Sekhonnd Fehlings, 2001) for which current clinical treatmentemains ineffective.
Oxidative damage occurs soon after SC injury (SCI).ighly reactive oxygen species, including superoxide an-
on (Liu et al., 1998), hydrogen peroxide (Liu et al., 1999),ydroxyl radical (Liu et al., 2003), and peroxynitrite (Liu etl., 2000), are excessively generated early after injury,hile natural mechanisms to inactivate them (i.e. glutathi-ne system, superoxide dismutase, and catalase) do notespond opportunely or efficiently (Azbill et al., 1997). Un-er these circumstances, oxidative degradation of lipids,roteins, and DNA, contribute to cell death (Liu et al., 1998,000, 2003; Leski et al., 2001; Aksenova et al., 2002).
Vascular events associated with initial trauma and withechanisms of secondary damage including vascular rup-
ure, thromboses, loss of vascular autoregulation, vaso-pasm, disruption of the blood–SC barrier, hypotension,tc., result in a marked fall in microvascular SC blood flowSCBF) which leads to ischemia in traumatized and adja-ent segments of the SC. Due to the pathophysiological
mportance of this hypo-perfusion state, multiple studiesave been performed to evaluate and try to counteracthese phenomena (Tator and Fehlings 1991; Anthes et al.,996; Carlson et al., 1997; Guha et al., 1989; Mautes et al.,000).
Glutathione in its reduced form (GSH) is quantitativelyhe most important endogenous rechargeable antioxidant.t has also been shown to act as a vasodilator underonditions of oxidative stress that alter endothelial func-ion, thereby improving ischemia (Cheung and Schulz,997; Lopez et al., 1998; Prasad et al., 1999; Weiss et al.,001; Arosio et al., 2002). In addition, it has a number of
mportant functions related to amino acid transport acrossembranes, protein synthesis and degradation, gene reg-lation, and cellular redox regulation (Anderson, 1998;ernerman and Hammarqvist, 1999). Glutathione ho-
eostasis is seriously affected after SCI, contributing toved.
tS
hiSsh1cbvi1m
itlr
E
TfSts(Sbpa2Cwitm
Fc
Srs
(tacsAe2c(vibtimhkl
Lfm8emselspwk
wtifrno
wstso
T
H
G
�
�
�
�
�
�
�
*
G. Guı́zar-Sahagún et al. / Neuroscience 130 (2005) 639–649640
he state of oxidative stress that ensues after trauma to theC (Taskiran et al., 2000; Lucas et al., 2002).
In accordance with the abovementioned data, we haveypothesized that increasing cellular levels of GSH should
mprove the general outcome of rats subjected to SCI.ince by itself GSH is not effectively transported into cells,everal alternatives to increase intracellular levels of GSHave been developed (Wernerman and Hammarqvist,999). Reduced glutathione monoethyl ester (GSHE) isell-permeable and is effectively transported into the cere-rospinal fluid (Anderson et al., 1989), and therefore pro-ides the most direct and convenient means available toncrease intracellular GSH concentration (Anderson et al.,985, 1989; Robinson et al., 1992; Wernerman and Ham-arqvist, 1999).
Recently we reported on the effect of GSHE to signif-cantly diminish SC lipid peroxidation (LPO) after SCI (San-oscoy et al., 2002). Here we have evaluated its effect onocomotor performance, neural degeneration, and SCBF inats early after moderate SCI.
EXPERIMENTAL PROCEDURES
xperimental design
o evaluate the effect of GSHE on long-term morphological andunctional evolution, we studied three groups of rats (n�8) withCI by contusion. The experimental group was given GSHE and
he other two control groups received methylprednisolone sodiumuccinate (MPSS; standard treatment for acute SCI), or vehiclesaline solution). To evaluate the effect of GSHE on microvascularCBF immediately after injury, three groups of rats (n�6) with SCIy compression received similar drug treatments to the first ex-erimental series; we included a group of sham-injured rats as andditional control. Adult female Long-Evans rats weighing 210–40 g were used for both experiments. The Animal Care and Useommittee of our Institution approved this study, and animalsere handled in compliance with the ethical guidelines of our
nstitution in accord with the National Institutes of Health guide forhe Care and Use of Laboratory Animals. Efforts were made toinimize the number of animals used and their suffering.
unctional and morphological studies after SContusion
pinal surgery and postoperative cares. Prior to surgery, rats wereandomly assigned to one of the treatment groups. For spinal
able 1. Percentage of error in height and velocity of impact for each
eight, % error
SHE MPSS SALINE
0.9 �0.8 �0.70.9 �1.1 0.21.1 0.1 �1.30.3 �0.8 �1.00.9 �1.6 �1.51.4 �1.0 �0.40.8 �0.5 �1.01.1 �0.6 �0.9
Difference between expected and actual parameter values.
urgery, animals were anesthetized with a mixture of ketamine n
80 mg/kg) and xylazine (8 mg/kg) given i.m. A bilateral laminec-omy was performed aseptically at T9 level to expose the dorsalspect of the SC, maintaining integrity of the meninges. The spinalolumn was then stabilized with angled clamps on T8 and T10pinous processes which were in turn held by a stereotaxic frame.
moderate SC contusion was inflicted by dropping onto thexposed dura, a stainless steel rod weighing 10 g with a flat tip of
mm in diameter, from a height of 25 mm using the well-haracterized New York University weight-drop impactor deviceBasso et al., 1996). No errors greater than 3% in height orelocity of impact were tolerated; errors for each animal are givenn Table 1. Post-surgical care included manual expression ofladders twice a day until bladder function returned, usually be-ween 2 and 3 weeks. Body weight changes were used as anndicator of the general condition of the animals; body weight was
easured daily for the first 7 day, then weekly. Animals wereoused individually in a temperature-regulated environment andept on a 12-h light/dark cycle. Food and water were provided adibitum.
Drug administration. GSHE (G-1404, Sigma Chem. Co., St.ouis, MO, USA) was given at a total dose of 12 mg/kg divided intoour injections (i.p.) at 20 min, 4 h, 10 h, and 20 h post-injury. Fourg/kg GSHE were given in the first injection, and the remainingmg/kg were divided equally for the additional three injections;
ach dose was dissolved in 1 ml of saline solution. MPSS (Phar-acia and Upjohn, Mexico City, Mexico) was also given at the
ame time points after injury in four i.p. injections of 30 mg/kgach. As prophylactic for infections, 8 mg/kg of ciprofloxacine
actate (Bayer, Mexico City, Mexico) were given SC every 12 h,tarting at the end of surgery and for 7 consecutive days. Torevent autophagia, acetaminophen (Cilag, Mexico City, Mexico)as given in the drinking water at an approximate dose of 64 mg/g/day for 3 weeks.
Assessment of motor function. Normal mobility of animalsas assessed before surgery. Starting at day 1 after surgery, and
hen once a week for 8 weeks, contused rats were assessedndividually for 4 min in an open field for hindlimb locomotorunction assessment using the Basso–Beattie–Bresnahan (BBB)ating scale, where 0 represents no hindlimb movement and 21ormal mobility (Basso et al., 1996). Rats were scored by twobservers unaware of the treatment received by the animals.
Histological studies. After the last functional assessment (8eeks after injury), rats of each group were divided into twoubgroups, matched according to their functional outcome. SCissue from one subgroup was used to assess the amount ofpared tissue at the injury site (n�4) and brain tissue from thether subgroup was used to quantify the number of surviving
e three experimental groups*
Velocity, % error
GSHE MPSS SALINE
�1.6 �1.1 �1.7�1.8 �1.9 �0.4�1.5 �0.2 �1.9�1.1 �1.1 �2.0�1.2 �1.5 �1.3�2.2 �1.8 �1.4�1.4 �2.0 �0.3�0.8 �1.5 �1.5
rat of th
eurons in the red nuclei (n�4).

ai4pcprronp
mTwEdmitmsata
asSoomtaomsA
rrOiodwrtpTo
M
t(Tefwtpmsct
riia5C3k
fapptcacb
gU(smc(topS
wMbuiEcc1p1e
S
MaSrcsalfw
M
A1twG
G. Guı́zar-Sahagún et al. / Neuroscience 130 (2005) 639–649 641
Assessment of spared SC tissue. Each rat was deeplynesthetized, and then given 1000 U of heparin i.p. and perfused
ntracardially with 100 ml of saline solution followed by 400 ml of% paraformaldehyde in 0.1 M phosphate buffer, pH 7.4, using aeristaltic pump (Masterflex, Chicago, IL, USA) at 30 ml/min. Onem of SC, with the injured area in the center, was dissected andlaced in the same fixative for 24 h at 4 °C; the tissue was thenoutinely processed and embedded in paraffin. Starting from theostral edge in the coronal plane, 8 �m thick sections, werebtained sequentially every 1 mm. Sections mounted on gelati-ized slides were stained with several techniques for further mor-hological and morphometric analysis.
Sections were stained with Luxol Fast Blue and Cresyl Violet foryelin and cell bodies respectively, using standardized procedures.o identify astrocytes, sections from all groups were immunostainedith a monoclonal antibody to glial fibrillary acidic protein (GFAP).ndogenous peroxidase was blocked with 3% hydrogen peroxide ineionized water for 10 min, and then sections were incubated for 10in with serum free blocker. The blocker was removed and the tissue
ncubated with the GFAP antibody (1:800) for 1 h at room tempera-ure. Afterward, tissues were incubated with biotinylated goat anti-ouse IgG for 10 min. Visualization was accomplished using the
treptavidin–horseradish peroxidase complex and diaminobenzidines a chromogen; counterstaining was performed with Harris’ hema-oxylin. Tissues were rinsed in phosphate buffer saline solution (PBS)fter each incubation.
GFAP-immunostained sections were used to quantify themount of spared tissue through the lesion site (in our hands, thistaining defines spared tissue better than Luxol Fast Blue).pared tissue was identified on the basis of intense GFAP stainingf tissue located predominately at the ventral and lateral aspectsf the SC. Morphometric analysis of spared SC parenchyma wereeasured at the epicenter, as well as 1–3 mm rostral and caudal
o it. Epicenter was defined as the section with the smallestmount of spared tissue. The transverse section area at T9 levelf non-injured SC tissue was measured as control. Area measure-ents were performed using the Image-Pro Plus image analysis
oftware from Media Cybernetics (V 4.5, Silver Spring, MD, USA).ll slides were assessed blindly with respect to the treatment.
Retrograde labeling of rubrospinal neurons. Surviving neu-ons in the red nuclei were stained with 5 �l of 10% tetramethyl-hodamine dextran dye (FluoroRuby; Molecular Probes, Eugene,R, USA) in PBS, applied to both rubrospinal areas of the prox-
mal stump after a complete transection of the SC below the sitef contusion at T12, in anesthetized rats. After 5 days, rats wereecapitated and their brains were excised; the areas of interestere cryosectioned. Every other 20 �m thick section through the
ed nuclei (an average of 44 sections) was qualitatively and quan-itatively examined by confocal microscopy. Only large and com-letely labeled cell bodies were counted in every collected section.he number of labeled neurons for each rat brain is the averagef cells counted in both red nuclei.
icrovascular SCBF after SC compression
Animal preparation and spinal surgery. Rats were anesthe-ized as previously described. Prior to spinal surgery, a catheterPE 50) was inserted into the femoral vein for fluid infusion.rachea was cannulated (by tracheotomy) with a 14-gauge cath-ter. Then a two segment laminectomy (T8 and T9) was per-ormed. With the aid of a surgical microscope dura and arachnoidsere sectioned longitudinally 1 mm in the most rostral portion of
he laminectomy (T8). This procedure was performed to directlylace the probe on the SC surface and hence avoid recordingixed flows of meninges and SC. Rats were then placed in a
tereotaxic frame. The laser-Doppler probe was affixed to a mi-romanipulator and placed perpendicularly to the SC, barely
ouching the dorsal surface of the pia mater. To avoid artifacts eelated to body movements (hyperextension) that usually occurmmediately after SCI, full muscular relaxation was produced by.v. administration of 3.2 mg/kg vecuronium for 5 min. Rats werertificially ventilated through the trachea, with room air at a rate of5–60 breaths/min using a small-animal respirator (Harvard type,.F. Palmer, London, UK). Rectal temperature was kept at6.0�0.2 °C with a thermostatically controlled homothermic blan-et (Harvard Apparatus, Edenbridge, UK).
SC compression was performed with an extradural 135-gorce clip for 5 s at the caudal part of the laminectomy (T9), 2 mmway from the probe. This is the shortest distance between therobe and the clip that ensures a continuous contact between therobe and the pia mater, without protrusion of SC tissue throughhe open meninges during the procedure. This type of injuryauses SCBF changes at the site of compression and peri-injuryreas (Guha et al., 1989; Westergren et al., 2001). We usedompression instead of contusion in order to measure SCBFefore, during and after SCI.
Fluids and drug administration. Fluids and drugs wereiven using a syringe pump (Sage Instruments, Cambridge, MA,SA): first 0.3 ml GSHE (4 mg/kg in saline solution), MPSS
30 mg/kg in saline solution), or saline solution for the injured andham-injured control groups, were administered as a bolus for 1in, 15 min after injury. All rats were given an infusion of 1 ml/h
ontaining vecuronium (53 �g/kg/min) and ketamine/xylazine2.13/0.21 mg/kg/min) to maintain muscular relaxation and anes-hesia until the end of the study. All rats received the same volumef fluids. At the end of the study, animals were killed by an i.v.otassium chloride overdose; immediately after heart arrest, theCBF fell to near 0.
SCBF measurements. Measurements of regional SCBFere performed with a MoorLab laser-Doppler flowmeter and aP3 probe with afferent and efferent fibers separated by 0.5 mm,oth from Moor Instruments, Devon, England. SCBF was contin-ously monitored for 110 min. Recordings were made before
njury (basal), 10 min after injury, and every 10 min thereafter.ach flux value was taken as the mean from about 200 individualonsecutive spikes in the region of interest. The signal was re-orded and analyzed using the Moorsoft software for Windows V.1. Arbitrary units obtained from the flowmeter were converted toercentage, considering the baseline value for each animal as00%. Differences between basal recordings and those obtainedvery 10 min for 100 min were plotted and statistically analyzed.
tatistical analysis
ean and S.E.M. values for motor function, body weight changes,nd regional SCBF were plotted as a function of time post-injury.tatistical analysis for these variables was assessed using the
epeated measures ANOVA test followed by Tukey’s multipleomparison test (Scheff et al., 2002). To compare final functionalcores of individual rats from each experimental group, themount of spared tissue at the injured site, and the number of
abeled cells in the red nuclei, we used the one-way ANOVAollowed by Tukey’s test. Differences were considered significanthen P�0.05.
RESULTS
otor function outcome
ll rats showed complete bilateral hindlimb paralysis at dayfollowing SC contusion, and gradually improved thereaf-
er. Functional recovery during the first 2 weeks post-injuryas similar among all groups, but from the 3rd week on,SHE-treated rats showed the greatest degree of recov-
ry, while MPSS- and vehicle-treated rats showed very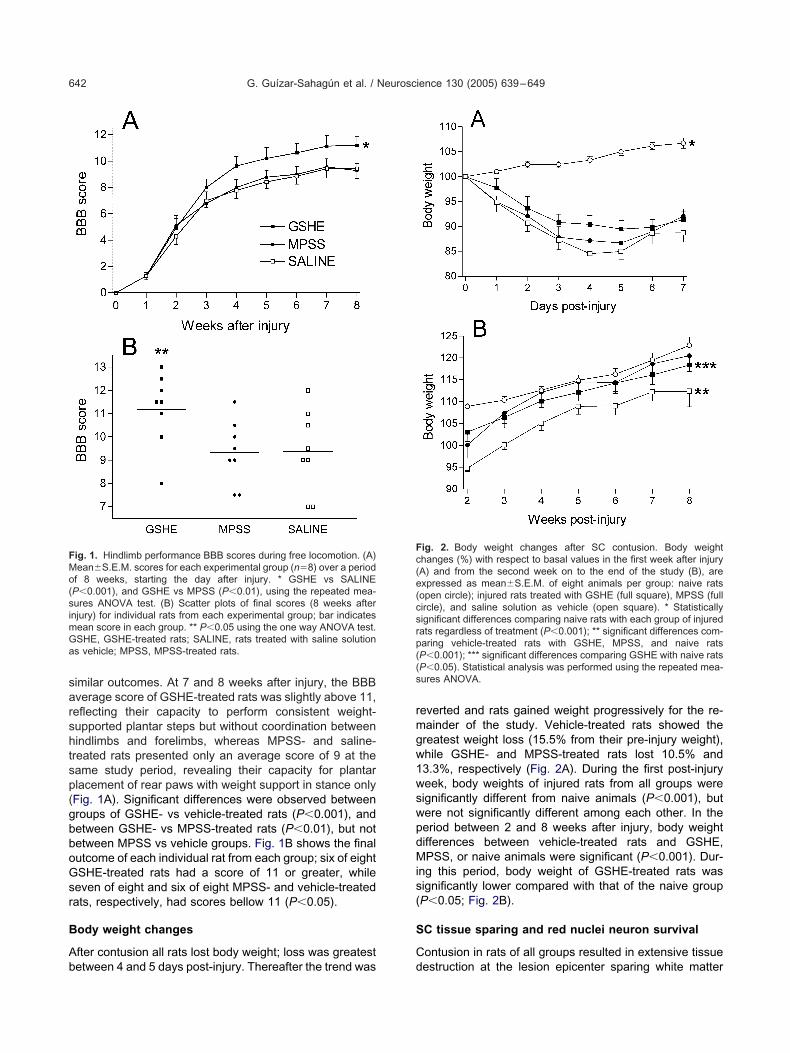
sarshtsp(gbboGsr
B
Ab
rmgw1wswpdMis(
S
C
FMo(simGa
Fc(e(csrp((s
G. Guı́zar-Sahagún et al. / Neuroscience 130 (2005) 639–649642
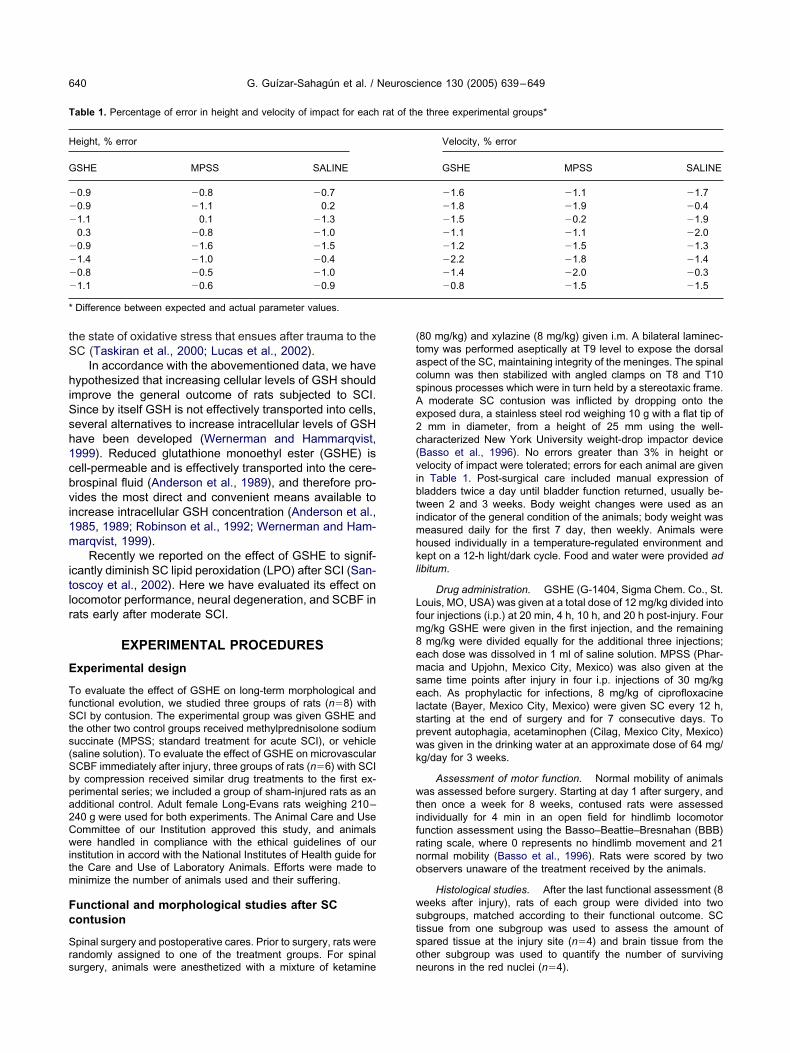
imilar outcomes. At 7 and 8 weeks after injury, the BBBverage score of GSHE-treated rats was slightly above 11,eflecting their capacity to perform consistent weight-upported plantar steps but without coordination betweenindlimbs and forelimbs, whereas MPSS- and saline-reated rats presented only an average score of 9 at theame study period, revealing their capacity for plantarlacement of rear paws with weight support in stance onlyFig. 1A). Significant differences were observed betweenroups of GSHE- vs vehicle-treated rats (P�0.001), andetween GSHE- vs MPSS-treated rats (P�0.01), but notetween MPSS vs vehicle groups. Fig. 1B shows the finalutcome of each individual rat from each group; six of eightSHE-treated rats had a score of 11 or greater, whileeven of eight and six of eight MPSS- and vehicle-treatedats, respectively, had scores bellow 11 (P�0.05).
ody weight changes
fter contusion all rats lost body weight; loss was greatest
ig. 1. Hindlimb performance BBB scores during free locomotion. (A)ean�S.E.M. scores for each experimental group (n�8) over a periodf 8 weeks, starting the day after injury. * GSHE vs SALINEP�0.001), and GSHE vs MPSS (P�0.01), using the repeated mea-ures ANOVA test. (B) Scatter plots of final scores (8 weeks afternjury) for individual rats from each experimental group; bar indicates
ean score in each group. ** P�0.05 using the one way ANOVA test.SHE, GSHE-treated rats; SALINE, rats treated with saline solutions vehicle; MPSS, MPSS-treated rats.
etween 4 and 5 days post-injury. Thereafter the trend was d
everted and rats gained weight progressively for the re-ainder of the study. Vehicle-treated rats showed thereatest weight loss (15.5% from their pre-injury weight),hile GSHE- and MPSS-treated rats lost 10.5% and3.3%, respectively (Fig. 2A). During the first post-injuryeek, body weights of injured rats from all groups wereignificantly different from naive animals (P�0.001), butere not significantly different among each other. In theeriod between 2 and 8 weeks after injury, body weightifferences between vehicle-treated rats and GSHE,PSS, or naive animals were significant (P�0.001). Dur-
ng this period, body weight of GSHE-treated rats wasignificantly lower compared with that of the naive groupP�0.05; Fig. 2B).
C tissue sparing and red nuclei neuron survival
ontusion in rats of all groups resulted in extensive tissue
ig. 2. Body weight changes after SC contusion. Body weighthanges (%) with respect to basal values in the first week after injuryA) and from the second week on to the end of the study (B), arexpressed as mean�S.E.M. of eight animals per group: naive ratsopen circle); injured rats treated with GSHE (full square), MPSS (fullircle), and saline solution as vehicle (open square). * Statisticallyignificant differences comparing naive rats with each group of injuredats regardless of treatment (P�0.001); ** significant differences com-aring vehicle-treated rats with GSHE, MPSS, and naive ratsP�0.001); *** significant differences comparing GSHE with naive ratsP�0.05). Statistical analysis was performed using the repeated mea-ures ANOVA.
estruction at the lesion epicenter sparing white matter

mwvna
mGco
FeismA tted for re
G. Guı́zar-Sahagún et al. / Neuroscience 130 (2005) 639–649 643
ainly at ventral and lateral margins. Newly formed tissueas found at central and dorsal portions, and cavitations ofariable extension were located between spared andewly formed tissues. No apparent sparing of gray matter
ig. 3. Spared SC tissue. G FAP immunostained specimens are shopicenter. (A, D) GSHE treated-rats; B, E, vehicle-treated rats; C,
mmunoreactivity in all groups. * Area of newly formed tissue; � areaspecimens per injured group which were analyzed 1–3 mm rostral toean�S.E.M. of spared area (mm2). No statistically significant differenNOVA). Non-injured cross-sectional SC area at T-9 (SHAM-I) is plo
t the epicenter was observed in any group. Spared white i
atter at this site was intensely and evenly stained forFAP in all groups (Fig. 3A–F) showing a persistent pro-
ess of gliosis. In most cases a gradient stain pattern wasbserved using the Luxol Fast Blue stain, which was more
ighest (A–C) and lowest (D–F) amount of spared white matter at the-treated rats. Spared white matter (outlined) shows intense GFAP
ion; v, ventral portion of the SC. Scale bar�550 �m. Histogram of fourenter, at the epicenter, and 1–3 mm caudal to it (G). Bars representfound among the injured groups for any of the sites studied (one wayference.
wn with hF, MPSSof cavitatthe epicces were
ntense in the most peripheral zone and weaker in the

dsra
sgTwi
tS((
S
TmbcrmSeowD
wsiao
F* d MPSSt nd (D) S
Fr(ci*(
G. Guı́zar-Sahagún et al. / Neuroscience 130 (2005) 639–649644
eepest portion, indicating variable myelination in thepared tissues (data not shown). Most sections at 1 mmostral and caudal to the epicenter showed preserved graynd white matter, as well as areas of cavitation.
The amount of spared SC tissue at each of the seventudied sites was not significantly different amongroups. Cross sectional SC area of sham-injured rats at9, and areas of spared SC tissues at the epicenter, asell as 1–3 mm cephalic and caudal to it are illustrated
n Fig. 3G.The number of surviving red nuclei cells in GSHE-
reated rats was significantly higher (158.8�9.3, mean�.E.M., n�4) compared with vehicle-treated animals
46.2�16.4) and MPSS-treated rats (53.5�14.7), P�0.01Fig. 4).
CBF
he mean SCBF in SC compressed GSHE-treated rats wasaintained with minor variations (not greater than 5% ofasal) during the study. No significant differences were foundomparing GSHE vs sham-injured groups. SCBF in injuredats treated with saline solution remained stable for up to 20in post-injury, and then fell progressively to 80% of basalCBF at the end of the study. MPSS-treated rats showed anarly and sustained increase of SCBF of approximately 12%ver baseline, which was significantly different comparedith SCBF in GSHE-treated and sham-injured rats (P�0.01).
ig. 4. Red nuclei neuron survival. (A) Number of retrograde labeleStatistically significant differences comparing GSHE with SALINE an
he greatest number of stained neurons from (B) GSHE, (C) MPSS, a
ifferences were also significant comparing the vehicle grouppr
ith MPSS-treated rats (P�0.001), with GSHE-treated andham-injured rats (P�0.05). By 30 min post-surgery, sham-
njured rats showed a 10% decrease in SCBF, which gradu-lly rose to values slightly above basal recordings by the endf the study (Fig. 5).
dies. Data represent the mean�S.E.M. of four animals per group.(P�0.01, one way ANOVA). Photomicrographs of the red nuclei withALINE groups, respectively. Scale bar�100 �m.
ig. 5. Temporal course of microvascular SCBF changes. Plots rep-esent mean�S.E.M. of six animals per group: sham-injured ratsopen circle); injured rats treated with GSHE (full square), MPSS (fullircle), and saline solution as vehicle (open square). Solid arrowndicates the time of injury; dashed arrow, the start of drug infusion.
Significant differences comparing MPSS with GSHE and SHAM-IP�0.01) and with SALINE (P�0.001); ** significant differences com-
d cell bo
aring SALINE with SHAM-I and GSHE (P�0.05). Statistical analysis:epeated measures ANOVA.

TgmSSace
R
Gtu21rtear1amd(
topaHaimai1gpivcrS
M
Fwsoomc
ut
iG
msTsu1dmitrdaia
faimntciScdbfomsgi
imgarpt1twsvoesiamRr
G. Guı́zar-Sahagún et al. / Neuroscience 130 (2005) 639–649 645
DISCUSSION
his study was undertaken to determine whether GSHEiven i.p. to rats subjected to SCI of moderate intensityodifies functional outcome, post-operative body weight,C tissue sparing, survival of red nuclei neurons, andCBF, compared with MPSS (the standard treatment forcute SCI) or vehicle. Rats given GSHE showed: signifi-ant motor function recovery and red nuclei neuron pres-rvation; SCBF was stabilized.
ationale for testing GSHE
SH plays a major antioxidant role as scavenger of reac-ive oxygen and nitrogen species, and as a dynamic reg-lator of the intracellular redox state (Klatt and Lamas,000), which are critical for cell function and survival (Hall,999). In addition, GSH could also play an anti-excitotoxicole by inhibiting the binding of specific ligands to iono-ropic glutamate receptors under redox modulation (Vargat al., 1997), which have been involved in excitotoxicityfter SCI (Park et al., 2004). Trauma to the SC seriouslyeduces GSH which results in SC neuron loss (Lucas et al.,998), possibly due to increased oxidative stress (Azbill etl., 1997; Taskiran et al., 2000; Lucas et al., 2002), anditochondrial dysfunction (Azbill et al., 1997). In vitro, GSHepletion enhances apoptosis of cultured motor neuronsRizzardini et al., 2003).
Recent experimental SCI studies have reported func-ional improvements, tissue preservation, and decreasedxidative stress by enhancing GSH levels, using the GSHrecursors L-oxothiazolidine-4-carboxylate (Kamencic etl., 2001) and �-glutamylcysteine (Lucas et al., 2002).ere we have chosen GSHE to enhance intracellular GSHfter SCI, because of its membrane permeability, and its
ntracellular conversion to GSH by the action of cytoplas-ic esterases (Anderson et al., 1985, 1989; Robinson etl., 1992; Wernerman and Hammarqvist, 1999); GSH itself
s not effectively transported into cells (Anderson et al.,985). In a model of excitotoxic damage to cerebellarranule neurons (Dabbeni-Sala et al., 2001) GSHE sup-lementation increased mitochondrial GSH pool preserv-
ng mitochondrial activity. GSHE has been shown to pro-ide neuroprotection following experimental transient focalerebral ischemia (Anderson et al., 2004a,b), and we haveecently reported its effectiveness to significantly diminishC LPO after SCI (Santoscoy et al., 2002).
orphological and functional outcome
unctional outcome was significantly better in rats treatedith GSHE vs controls (vehicle- and MPSS-treated rats). Ithould be stressed that although the difference in motorutcome between GSHE-treated rats and controls is onlyf two points on the BBB scale, its clinical significance isuch greater: it is the difference between an animal that
an walk vs an animal that can only stand.Similarity in percentage error of impact parameter val-
es among rats of the three experimental groups rules out
he possibility of differences in the mechanics of contusion tnjury as an explanation for the improved behavior inSHE-treated rats.
Despite the improved locomotor activity using GSHE,orphometric analysis revealed no significant SC tissue
paring at the epicenter of injury and neighboring areas.he functional significance of tissue sparing is controver-ial. Studies on models of SC contusion or compressionsing graded injury (Basso et al., 1996; Kuhn and Wrathall,998; Joshi and Fehlings, 2002; Scheff et al., 2003) haveemonstrated an expected positive correlation betweenotor function outcome and spared SC tissue at the site of
njury. This correlation has also been observed in studiesesting diverse neuroprotective strategies: following inju-ies of the same intensity, animals receiving treatmentemonstrated significantly enhanced functional recoveryttributed to the presence of significant neural tissue spar-
ng at the site of injury (Popovich et al., 1999; Rosenberg etl., 1999; Gaviria et al., 2000; Wells et al., 2003).
On the other hand, diverse treatments like ours haveound no relation between functional outcome and themount of spared tissue at the site of injury. Functional
mprovement of SC-injured rats treated with BDNF (Jake-an et al., 1998) or bFGF (Rabchevsky et al., 2000) wasot accompanied by an increased amount of spared SCissue. Using several anti-excitotoxic strategies, signifi-antly improved trunk musculature, as measured by rear-
ng events, was observed in the absence of differences inC tissue sparing in contused treated rats compared withontrols (Mills et al., 2002a). In another study, significantifferences in the amount of spared SC tissue have noteen associated with significant improvements in motorunction in later stages of SCI (Mills et al., 2002b). Thesebservations as well as our results suggest that improvedotor function can or cannot be related to the amount of
pared SC tissue. Taken together, these observations sug-est that other mechanisms, yet to be defined, underlie the
mprovements observed.In the absence of significant differences in tissue spar-
ng, improvement in weight-supported hindlimb move-ents in GSHE-treated rats may result in part from thereater preservation of red nuclei neurons that projectxons below the site of injury (the population of neuronsetrogradely stained here). It has been shown that su-raspinal axons, including non-pyramidal tracts, are impor-ant in modulating gross motor performance (Houk et al.,993; Fehlings and Tator, 1995). Although not analyzed inhe present study, preservation of other descending path-ays including serotoninergic and monoaminergic brain-tem projections, as well as propriospinal neurons in theicinity and further away from the lesion, could also haveccurred and contribute to the improved functional recov-ry observed in GSHE-treated animals. Experimental datauggest that both monoamines and serotonin are involved
n various aspects of locomotor control including initiationnd modulation of the central pattern-generator for loco-otion located in the lumbar SC (Jacobs and Fornal, 1997;ossignol et al., 1998). Moreover, supplementation of se-
otonin by agonists or sublesional transplantation of sero-
oninergic neurons (Ribotta et al., 2000; Antri et al., 2002;
Gtsi
ioim
etatdssiohoagmas
osttoo2sda(2t
B
L2bbliltbsasg
M
Lc
temi0oatSmaVc
creissic(1(ntirga
1poptaGi(Aipnc1
Gsvntgtt
G. Guı́zar-Sahagún et al. / Neuroscience 130 (2005) 639–649646
imenez y Ribotta et al., 2002) improves locomotor func-ion in SC-injured animals. Therefore, it could be that be-ides quantity, quality of spared tissue at the site of injurys a factor for functional improvement.
Preserved white matter at the site of injury showedntense GFAP staining, revealing an enhanced metabolismf persistent gliosis. Regardless of its significance, the
ntense GFAP immunoreactivity was used here as aeans to visualize and measure spared SC tissue.
Our GSHE-treated rats showed a better motor recov-ry than controls starting 3 weeks after injury. This fact,
ogether with the lack of a significant difference in themount of spared tissue among the groups, suggests that
he beneficial effects of acute GSHE treatment could beue to the influence of this compound on processes ofelf-repair, such as enhanced segmental plasticity (Gold-tein et al., 1997; Fouad et al., 2001; Weidner et al., 2001),nstead of a direct action to block secondary mechanismsf injury. This hypothesis is supported by the fact that GSHolds multiple important functions besides its major anti-xidant role. GSH participates in amino acid transportcross membranes, protein synthesis and degradation,ene regulation (Anderson, 1998; Wernerman and Ham-arqvist, 1999), and immune function regulation (Utsugi etl., 2002; Villa et al., 2002), all of which are mechanisms ofelf-repair after SCI (Hulsebosch, 2002).
Functional outcome and survival of red nuclei neuronsf rats treated with MPSS were very similar to those ob-erved in vehicle-treated rats, but significantly lower thanhose seen in the GSHE-treated animals. This suggestshat treatment of acute SCI using GSHE could be a betterption than standard MPSS treatment. In agreement withur results, several human (Hurlbert, 2001; Pointillart et al.,000) and experimental (Lankhorst et al., 2000; Rabchev-ky et al., 2002; Takami et al., 2002) studies have failed toemonstrate MPSS-induced improvement in SC functionfter injury, although there are also reports to the contraryBraughler et al., 1987; Behrmann et al., 1994; Bracken,001). A discussion regarding this controversy is beyondhe scope of the present work.
ody weight changes
ike in previous studies of SCI in rats (Lankhorst et al.,000 and 2001; Van Meeteren et al., 2003), monitoringody weight was used as a measure of the general well-eing of the animals. Although differences in body weight
oss during the first week post-injury among groups werensignificant, GSHE-treated rats showed a trend to loseess weight than MPSS- and saline-treated groups. In con-rast, from the second week on there were differences inody weight gain among groups. Weight increase wasignificantly greater in both drug-treated groups (MPSSnd GSHE) as compared with the saline-treated animals,uggesting a beneficial effect of both drugs on the animals’eneral condition.
icrocirculation
aser-Doppler flowmetry used here permits non-invasive,
ontinuous, on-line assessment of SC microcirculation in ahe rat (Young et al., 1994; Hitchon et al., 1996; Kristensent al., 1998). To sample SCBF in a volume of approxi-ately 0.4 mm3 of SC tissue (at T9 the diameter of the SC
s approximately 1.6 mm thick) we used a probe with.5 mm fiber separation (Larsson et al., 2002). This volumef SC tissue includes dorsal white and gray matters. Tovoid measurement of dura and arachnoid microcircula-ion, we placed the probe directly on the pial surface of theC. Fixation of both animal and probe, and the use of auscular relaxant, prevented movement artifacts, whichre commonly a source of interference (Obeid et al., 1990).entilation and body temperature were also variables weontrolled.
In this report, soon after SC compression, SCBF de-reased progressively at the site of lesion in saline-treatedats. Of the various events that have been described toxplain diminished SCBF after SCI, arteriolar vasospasm
s an important contributor (Anthes et al., 1996). Vaso-pasms could result from an imbalance of vasoactive sub-tances. Early after traumatic SCI there is a significant
ncrease of endothelins in parenchymal tissue and extra-ellular fluid, as well as in cerebrospinal fluid and plasmaMcKenzie et al., 1995; Salzman et al., 1996; Uesugi et al.,996), leading to vasoconstriction of SC blood vesselsWestmark et al., 1995), and ultimately stress and death ofeurons (Sato and Noble, 1998). Further contributing tohe imbalance of vasoactive substances after SCI is thenactivation of the major vasodilator nitric oxide, by itseaction with superoxide anion to form peroxynitrite (Gry-lewski et al., 1986; Loscalzo, 1996), both reactive speciesbundantly produced after SCI (Liu et al., 1998, 2000).
In agreement with earlier studies (Young and Flamm,982; Hall et al., 1984; Holtz et al., 1990) MPSS treatmentroduced a significant increase of SCBF compared with allther studied groups, while treatment with GSHE blocked theost-traumatic hypoperfusion seen after injury in vehicle-
reated rats. These observations suggest that both MPSSnd GSHE cause local vasodilation of SC blood vessels. LikeSH, its precursor GSHE could exert its effect on SCBF by
mproving endothelial function altered by oxidative stressLopez et al., 1998; Prasad et al., 1999; Weiss et al., 2001;rosio et al., 2002). GSH causes vasodilation and therefore
mprovement of macro- and microcirculation, by reacting witheroxynitrite to form S-nitrosoglutathione, thereby increasingitric oxide levels which in turn activate nitric oxide-guanylateyclase-dependent mechanisms (Cheung and Schulz,997).
CONCLUSIONS
SHE given early following moderate SC contusion in ratsignificantly enhances motor function recovery and sur-ival of suprasegmentary neurons in the absence of sig-ificant tissue sparing at the injury site, and is associatedo SCBF stabilization. Beneficial actions of GSHE werereater than those obtained with MPSS which is currentlyhe treatment of choice for SC-injured patients. Evenhough we cannot rule out the possibility of neuroprotection
fter GSHE treatment due to the significant survival of
nfsigpwaeu
A3t
A
A
A
A
A
A
A
A
A
A
A
B
B
B
B
C
C
D
D
F
F
G
G
G
G
G
H
H
H
H
H
H
H
J
J
J
K
G. Guı́zar-Sahagún et al. / Neuroscience 130 (2005) 639–649 647
eurons measured in the red nuclei, the time-course ofunctional improvement, given the lack of gross tissueparing at the site of injury, suggests that mechanisms ofntrinsic plasticity and self-repair may be relevant to thelobal effect of GSHE on motor function. These interestingossibilities warrant further analysis. The managementith GSHE of SC lesions of varying severity and the ther-peutic window of its application are some of the param-ters that still must be evaluated before recommending itsse in the clinical setting.
cknowledgments—This work was supported by CONACYT grant0658-M. We thank Francisco Márquez for his invaluable assis-ance with histology techniques.
REFERENCES
ksenova M, Butterfield DA, Zhang SX, Underwood M, Geddes JW(2002) Increased protein oxidation and decreased creatine kinaseBB expression and activity after spinal cord contusion injury.J Neurotrauma 19:491–502.
nderson DK, Hall ED (1993) Pathophysiology of spinal cord trauma.Ann Emerg Med 22:987–992.
nderson ME, Powrie F, Puri RN, Meister A (1985) Glutathione mo-noethyl ester: preparation, uptake by tissues, and conversion toglutathione. Arch Biochem Biophys 239:538–548.
nderson ME, Underwood M, Bridges RJ, Meister A (1989) Glutathi-one metabolism at the blood-cerebrospinal fluid barrier. FASEB J3:2527–2531.
nderson ME (1998) Glutathione: an overview of biosynthesis andmodulation. Chem Biol Interact 111–112:1–14
nderson MF, Nilsson M, Eriksson PS, Sims NR (2004a) Glutathionemonoethyl ester provides neuroprotection in a rat model of stroke.Neurosci Lett 354:163–165.
nderson MF, Nilsson M, Sims NR (2004b) Glutathione monoethyl-ester prevents mitochondrial glutathione depletion during focalcerebral ischemia. Neurochem Int 44:153–159.
nthes DL, Theriault E, Tator CH (1996) Ultrastructural evidence forarteriolar vasospasm after spinal cord trauma. Neurosurgery39:804–814.
ntri M, Orsal D, Barthe JY (2002) Locomotor recovery in the chronicspinal rat: effects of long-term treatment with a 5-HT2 agonist. EurJ Neurosci 16:467–476.
rosio E, De Marchi S, Zannoni M, Prior M, Lechi A (2002) Effect ofglutathione infusion on leg arterial circulation, cutaneous microcir-culation, and pain-free walking distance in patients with peripheralobstructive arterial disease: a randomized, double-blind, placebo-controlled trial. Mayo Clin Proc 77:754–759.
zbill RD, Mu X, Bruce-Keller AJ, Mattson MP, Springer JE (1997)Impaired mitochondrial function, oxidative stress and altered anti-oxidant enzyme activities following traumatic spinal cord injury.Brain Res 765:283–290.
asso DM, Beattie MS, Bresnahan JC (1996) Graded histological andlocomotor outcomes after spinal cord contusion using the NYUweight-drop device versus transection. Exp Neurol 139:244–256.
ehrmann DL, Bresnahan JC, Beattie MS (1994) Modeling of acutespinal cord injury in the rat: neuroprotection and enhanced recov-ery with methylprednisolone, U-74006F and YM-14673. Exp Neu-rol 126:61–75.
racken MB (2001) Methylprednisolone and acute spinal cord injury:an update of the randomized evidence. Spine 26:S47–S54.
raughler JM, Hall ED, Means ED, Waters TR, Anderson DK (1987)Evaluation of an intensive methylprednisolone sodium succinatedosing regimen in experimental spinal cord injury. J Neurosurg67:102–105.
arlson GD, Minato Y, Okada A, Gorden CD, Warden KE, Barbeau
JM, Biro CL, Bahnuik E, Bohlman HH, Lamanna JC (1997) Earlytime-dependent decompression for spinal cord injury: vascularmechanisms of recovery. J Neurotrauma 14:951–962.
heung PY, Schulz R (1997) Glutathione causes coronary vasodila-tion via a nitric oxide- and soluble guanylate cyclase-dependentmechanism. Am J Physiol 273:H1231–H1238.
abbeni-Sala F, Floreani M, Franceschini D, Skaper SD, Giusti P(2001) Kainic acid induces selective mitochondrial oxidative phos-phorylation enzyme dysfunction in cerebellar granule neurons:protective effects of melatonin and GSH ethyl ester. FASEB J15:1786–1788.
umont RJ, Okonkwo DO, Verma S, Hurlbert RJ, Boulos PT, EllegalaDB, Dumont AS (2001) Acute spinal cord injury, part I: pathophys-iologic mechanisms. Clin Neuropharmacol 24:254–264.
ehlings MG, Tator CH (1995) The relationships among the severity ofspinal cord injury, residual neurological function, axon counts, andcounts of retrogradely labeled neurons after experimental spinalcord injury. Exp Neurol 132:220–228.
ouad K, Pedersen V, Schwab ME, Brosamle C (2001) Cervicalsprouting of corticospinal fibers after thoracic spinal cord injuryaccompanies shifts in evoked motor responses. Curr Biol11:1766–1770.
aviria M, Privat A, d’Arbigny P, Kamenka J, Haton H, Ohanna F(2000) Neuroprotective effects of a novel NMDA antagonist, gacy-clidine, after experimental contusive spinal cord injury in adult rats.Brain Res 874:200–209.
imenez y Ribotta M, Gaviria M, Menet V, Privat A (2002) Strategiesfor regeneration and repair in spinal cord traumatic injury. ProgBrain Res 137:191–212.
oldstein B, Little JW, Harris RM (1997) Axonal sprouting followingincomplete spinal cord injury: an experimental model. J SpinalCord Med 20:200–206.
ryglewski RJ, Palmer RM, Moncada S (1986) Superoxide anion isinvolved in the breakdown of endothelium-derived vascular relax-ing factor. Nature 320:454–456.
uha A, Tator CH, Smith CR, Piper I (1989) Improvement in post-traumatic spinal cord blood flow with a combination of a calciumchannel blocker and a vasopressor. J Trauma 29:1440–1447.
all AG (1999) Review: the role of glutathione in the regulation ofapoptosis. Eur J Clin Invest 29:238–245.
all ED, Wolf DL, Braughler JM (1984) Effects of a single large doseof methylprednisolone sodium succinate on experimental posttrau-matic spinal cord ischemia: dose-response and time-action analy-sis. J Neurosurg 61:124–130.
itchon PW, Mouw LJ, Rogge TN, Torner JC, Miller AK (1996) Re-sponse of spinal cord blood flow to the nitric oxide inhibitor ni-troarginine. Neurosurgery 39:795–803.
oltz A, Nystrom B, Gerdin B (1990) Effect of methylprednisolone onmotor function and spinal cord blood flow after spinal cord com-pression in rats. Acta Neurol Scand 82:68–73.
ouk JC, Keifer J, Barto AG (1993) Distributed motor commands in thelimb premotor network. Trends Neurosci 16:27–33.
ulsebosch CE (2002) Recent advances in pathophysiology and treat-ment of spinal cord injury. Adv Physiol Educ 26:238–255.
urlbert RJ (2001) The role of steroids in acute spinal cord injury: anevidence-based analysis. Spine 26:S39–S46.
acobs BL, Fornal CA (1997) Serotonin and motor activity. Curr OpinNeurobiol 7:820–825.
akeman LB, Wei P, Guan Z, Stokes BT (1998) Brain-derived neuro-trophic factor stimulates hindlimb stepping and sprouting of cho-linergic fibers after spinal cord injury. Exp Neurol 154:170–184.
oshi M, Fehlings MG (2002) Development and characterization of anovel, graded model of clip compressive spinal cord injury in themouse: part 2. Quantitative neuroanatomical assessment andanalysis of the relationships between axonal tracts, residual tissue,and locomotor recovery. J Neurotrauma 19:191–203.
amencic H, Griebel RW, Lyon AW, Paterson PG, Juurlink BH (2001)
Promoting glutathione synthesis after spinal cord trauma de-
K
K
K
K
L
L
L
L
L
L
L
L
L
L
L
L
M
M
M
M
O
P
P
P
P
R
R
R
R
R
R
R
S
S
S
S
S
S
T
T
G. Guı́zar-Sahagún et al. / Neuroscience 130 (2005) 639–649648
creases secondary damage and promotes retention of function.FASEB J 15:243–250.
aynar MY, Hanci M, Kafadar A, Gumustas K, Belce A, Ciplak N(1998) The effect of duration of compression on lipid peroxidationafter experimental spinal cord injury. Neurosurg Rev 21:117–120.
latt P, Lamas S (2000) Regulation of protein function by S-glutathiolation in response to oxidative and nitrosative stress. EurJ Biochem 267:4928–4944.
ristensen JD, Karlsten R, Gordh T (1998) Spinal cord blood flow afterintrathecal injection of ropivacaine and bupivacaine with or withoutepinephrine in rats. Acta Anaesthesiol Scand 42:685–690.
uhn PL, Wrathall JR (1998) A mouse model of graded contusivespinal cord injury. J Neurotrauma 15:125–140.
ankhorst AJ, ter Laak MP, Hamers FP, Gispen WH (2000) Combinedtreatment with alphaMSH and methylprednisolone fails to improvefunctional recovery after spinal injury in the rat. Brain Res859:334–340.
ankhorst AJ, ter Laak MP, van Laar TJ, van Meeteren NL, de GrootJC, Schrama LH, Hamers FP, Gispen WH (2001) Effects of en-riched housing on functional recovery after spinal cord contusiveinjury in the adult rat. J Neurotrauma 8:203–215.
arsson M, Steenbergen W, Stromberg T (2002) Influence of opticalproperties and fiber separation on laser Doppler flowmetry.J Biomed Opt 7:236–243.
eski ML, Bao F, Wu L, Qian H, Sun D, Liu D (2001) Protein and DNAoxidation in spinal injury: neurofilaments: an oxidation target. FreeRadic Biol Med 30:613–624.
iu D, Ling X, Wen J, Liu J (2000) The role of reactive nitrogen speciesin secondary spinal cord injury: formation of nitric oxide, peroxyni-trite, and nitrated protein. J Neurochem 75:2144–2154.
iu D, Liu J, Sun D, Alcock NW, Wen J (2003) Spinal cord injuryincreases iron levels: catalytic production of hydroxyl radicals. FreeRadic Biol Med 34:64–71.
iu D, Liu J, Wen J (1999) Elevation of hydrogen peroxide after spinalcord injury detected by using the Fenton reaction. Free Radic BiolMed 27:478–482.
iu D, Sybert TE, Qian H, Liu J (1998) Superoxide production afterspinal injury detected by microperfusion of cytochrome c. FreeRadic Biol Med 25:298–304.
opez BL, Snyder JW, Birenbaum DS, Ma XI (1998) N-acetylcysteineenhances endothelium-dependent vasorelaxation in the isolatedrat mesenteric artery. Ann Emerg Med 32:405–410.
oscalzo J (1996) The oxidant stress of hyperhomocyst(e)inemia.J Clin Invest 98:5–7.
ucas JH, Wheeler DG, Emery DG, Mallery SR (1998) The endoge-nous antioxidant glutathione as a factor in the survival of physicallyinjured mammalian spinal cord neurons. J Neuropathol Exp Neurol57:937–954.
ucas JH, Wheeler DG, Guan Z, Suntres Z, Stokes BT (2002) Effect ofglutathione augmentation on lipid peroxidation after spinal cordinjury. J Neurotrauma 19:763–775.
autes AE, Weinzierl MR, Donovan F, Noble LJ (2000) Vascularevents after spinal cord injury: contribution to secondary pathogen-esis. Phys Ther 80:673–687.
cKenzie AL, Hall JJ, Aihara N, Fukuda K, Noble LJ (1995) Immuno-localization of endothelin in the traumatized spinal cord: relation-ship to blood-spinal cord barrier breakdown. J Neurotrauma12:257–268.
ills CD, Johnson KM, Hulsebosch CE (2002a) Role of group II andgroup III metabotropic glutamate receptors in spinal cord injury.Exp Neurol 173:153–167.
ills CD, Johnson KM, Hulsebosch CE (2002b) Group I metabotropicglutamate receptors in spinal cord injury: roles in neuroprotectionand the development of chronic central pain. J Neurotrauma19:23–42.
beid AN, Barnett NJ, Dougherty G, Ward G (1990) A critical review
of laser Doppler flowmetry. J Med Eng Technol 14:178–181.ark E, Velumian AA, Fehlings MG (2004) The role of excitotoxicity insecondary mechanisms of spinal cord injury: a review with anemphasis on the implications for white matter degeneration. J Neu-rotrauma 21:754–774.
ointillart V, Petitjean ME, Wiart L, Vital JM, Lassie P, Thicoipe M,Dabadie P (2000) Pharmacological therapy of spinal cord injuryduring the acute phase. Spinal Cord 38:71–76.
opovich PG, Guan Z, Wei P, Huitinga I, van Rooijen N, Stokes BT(1999) Depletion of hematogenous macrophages promotes partialhindlimb recovery and neuroanatomical repair after experimentalspinal cord injury. Exp Neurol 158:351–365.
rasad A, Andrews NP, Padder FA, Husain M, Quyyumi AA (1999)Glutathione reverses endothelial dysfunction and improves nitricoxide bioavailability. J Am Coll Cardiol 34:507–514.
abchevsky AG, Fugaccia I, Turner AF, Blades DA, Mattson MP,Scheff SW (2000) Basic fibroblast growth factor (bFGF) enhancesfunctional recovery following severe spinal cord injury to the rat.Exp Neurol 164:280–291.
abchevsky AG, Fugaccia I, Sullivan PG, Blades DA, Scheff SW(2002) Efficacy of methylprednisolone therapy for the injured ratspinal cord. J Neurosci Res 68:7–18.
ibotta MG, Provencher J, Feraboli-Lohnherr D, Rossignol S, Privat A,Orsal D (2000) Activation of locomotion in adult chronic spinal ratsis achieved by transplantation of embryonic raphe cells reinnervat-ing a precise lumbar level. J Neurosci 20:5144–5152.
izzardini M, Lupi M, Bernasconi S, Mangolini A, Cantoni L (2003)Mitochondrial dysfunction and death in motor neurons exposed tothe glutathione-depleting agent ethacrynic acid. J Neurol Sci207:51–58.
obinson MK, Ahn MS, Rounds JD, Cook JA, Jacobs DO, WilmoreDW (1992) Parenteral glutathione monoester enhances tissue an-tioxidant stores. JPEN J Parenter Enteral Nutr 16:413–418.
osenberg LJ, Teng YD, Wrathall JR (1999) Effects of the sodiumchannel blocker tetrodotoxin on acute white matter pathology afterexperimental contusive spinal cord injury. J Neurosci 19:6122–6133.
ossignol S, Chau C, Brustein E, Giroux N, Bouyer L, Barbeau H,Reader TA (1998) Pharmacological activation and modulation ofthe central pattern generator for locomotion in the cat. Ann NYAcad Sci 860:346–359.
alzman SK, Acosta R, Beck G, Madden J, Boxer B, Ohlstein EH(1996) Spinal endothelin content is elevated after moderate localtrauma in the rat to levels associated with locomotor dysfunctionafter intrathecal injection. J Neurotrauma 13:93–101.
antoscoy C, Rios C, Franco-Bourland RE, Hong E, Bravo G, RojasG, Guizar-Sahagun G (2002) Lipid peroxidation by nitric oxidesupplements after spinal cord injury: effect of antioxidants in rats.Neurosci Lett 330:94–98.
ato M, Noble LJ (1998) Involvement of the endothelin receptor sub-type A in neuronal pathogenesis after traumatic brain injury. BrainRes 809:39–49.
cheff SW, Saucier DA, Cain ME (2002) A statistical method foranalyzing rating scale data: the BBB locomotor score. J Neuro-trauma 19:1251–1260.
cheff SW, Rabchevsky AG, Fugaccia I, Main JA, Lumpp JE Jr (2003)Experimental modeling of spinal cord injury: characterization of aforce-defined injury device. J Neurotrauma 20:179–193.
ekhon LH, Fehlings MG (2001) Epidemiology, demographics, andpathophysiology of acute spinal cord injury. Spine 26:S2–S12.
akami T, Oudega M, Bethea JR, Wood PM, Kleitman N, Bunge MB(2002) Methylprednisolone and interleukin-10 reduce gray matterdamage in the contused Fischer rat thoracic spinal cord but do notimprove functional outcome. J Neurotrauma 19:653–666.
askiran D, Tanyalcin T, Sozmen EY, Peker GO, Gulmen V, Cagli S,Kanit L, Tekeli G, Barcin E, Zileli M, Kutay FZ (2000) The effects ofmelatonin on the antioxidant systems in experimental spinal injury.
Int J Neurosci 104:63–73.
T
T
U
U
V
V
V
W
W
W
W
W
W
Y
Y
G. Guı́zar-Sahagún et al. / Neuroscience 130 (2005) 639–649 649
ator CH (1995) Update on the pathophysiology and pathology ofacute spinal cord injury. Brain Pathol 5:407–413.
ator CH, Fehlings MG (1991) Review of the secondary injury theoryof acute spinal cord trauma with emphasis on vascular mecha-nisms. J Neurosurg 75:15–26.
esugi M, Kasuya Y, Hama H, Yamamoto M, Hayashi K, Masaki T,Goto K (1996) Endogenous endothelin-1 initiates astrocytic growthafter spinal cord injury. Brain Res 728:255–259.
tsugi M, Dobashi K, Koga Y, Shimizu Y, Ishizuka T, Iizuka K, HamuroJ, Nakazawa T, Mori M (2002) Glutathione redox regulates lipopo-lysaccharide-induced IL-12 production through p38 mitogen-activated protein kinase activation in human monocytes: role ofglutathione redox in IFN-gamma priming of IL-12 production.J Leukoc Biol 71:339–347.
an Meeteren NL, Eggers R, Lankhorst AJ, Gispen WH, Hamers FP(2003) Locomotor recovery after spinal cord contusion injury in rats isimproved by spontaneous exercise. J Neurotrauma 20:1029–1037.
arga V, Jenei Z, Janaky R, Saransaari P, Oja SS (1997) Glutathioneis an endogenous ligand of rat brain N-methyl-D-aspartate (NMDA)and 2-amino-3-hydroxy-5-methyl-4-isoxazolepropionate (AMPA)receptors. Neurochem Res 22:1165–1171.
illa P, Saccani A, Sica A, Ghezzi P (2002) Glutathione protects micefrom lethal sepsis by limiting inflammation and potentiating hostdefense. J Infect Dis 185:1115–1120.
eidner N, Ner A, Salimi N, Tuszynski MH (2001) Spontaneous
corticospinal axonal plasticity and functional recovery after adultcentral nervous system injury. Proc Natl Acad Sci USA98:3513–3518.
eiss N, Zhang YY, Heydrick S, Bierl C, Loscalzo J (2001) Overexpres-sion of cellular glutathione peroxidase rescues homocyst(e)ine-induced endothelial dysfunction. Proc Natl Acad Sci USA 98:12503–12508.
ells JE, Hurlbert RJ, Fehlings MG, Yong VW (2003) Neuroprotectionby minocycline facilitates significant recovery from spinal cordinjury in mice. Brain 126:1628–1637.
ernerman J, Hammarqvist F (1999) Modulation of endogenous glu-tathione availability. Curr Opin Clin Nutr Metab Care 2:487–492.
estergren H, Farooque M, Olsson Y, Holtz A (2001) Spinal cordblood flow changes following systemic hypothermia and spinalcord compression injury: an experimental study in the rat usingLaser-Doppler flowmetry. Spinal Cord 39:74–84.
estmark R, Noble LJ, Fukuda K, Aihara N, McKenzie AL (1995)Intrathecal administration of endothelin-1 in the rat: impact onspinal cord blood flow and the blood-spinal cord barrier. NeurosciLett 192:173–176.
oung WF, Rosenwasser RH, Vasthare US, Tuma RF (1994) Preser-vation of post-compression spinal cord function by infusion ofhypertonic saline. J Neurosurg Anesthesiol 6:122–127.
oung W, Flamm ES (1982) Effect of high-dose corticosteroid therapyon blood flow, evoked potentials, and extracellular calcium in ex-
perimental spinal injury. J Neurosurg 57:667–673.(Accepted 25 September 2004)(Available online 23 November 2004)
Copyright © 2022 FDOKUMEN




















