
Influence of uremia and hemodialysis on circulating interleukin-1 and tumor necrosis factor
10
Kidney International, Vol. 37 (1990), pp. 116—125 Influence of uremia and hemodialysis on circulating interleukin-1 and tumor necrosis factor a ANDRE HERBELIN, ANH THU NGUYEN, JOHANNA ZINGRAFF, PABLO UREA, and BEATRICE DESCAMPS-LATSCHA INSERM U 25 and CNRS UA 122, INSERM U 90 Hôpital Necker, Paris, France Influence of uremia and hemodlalysis on circulating interleukin-1 and tumor necrosis factor a. Interleukin-1 (IL-I) and tumor necrosis factor a (TNFa) were determined in the plasma of long-term hemodialysis (HD) patients and uremic (UR) patients undergoing their first dialysis session using either cellulosic (CUP) or synthetic (PAN-AN 69) mem- brane-equipped dialyzers. In long-term HD patients, plasma IL-I and TNFa levels were significantly increased compared to their levels in normal subjects. During a single dialysis session, a significant increase in IL-I but not in TNFa was observed. In not yet dialyzed UR patients, LL-l plasma levels did not differ from those observed in normal subjects. By contrast, TNFa was found significantly increased although less than in long-term HD patients. During the first dialysis session, no significant increase was observed in the levels of either monokine, Lastly, regardless of the group of patients, no significant influence of the dialysis membrane could be detected, suggesting that the observed changes are not exclusively secondary to the activation of complement. Altogether, these results suggest that the passage of the blood through the extracorporeal dialysis circuit triggers the secretion of IL-i and further exacerbates that of TNFa by monocytes. The presence of increased TNFa in the plasma of first-dialysis UR patients suggests that factors unrelated to dialysis contribute to the activation of monocytes in these patients. Lastly, the concomitant presence of IL-I and TNFa in the plasma of long-term HD patients could be responsible for some of the clinical features observed in these patients, and provides strong evidence favoring the concept that HD can be assimilated to a recurrent acute-phase inflammatory response. Growing interest has focused in recent years on the role of cytokines in relation to the acute phase response of the inflam- matory process [1—3]. Among these cytokines, interleukin-1 (IL-i), which is mainly derived from monocyte-macrophages, has been shown to exert a broad range of activities associated with both acute and chronic inflammatory changes [4, 5], and to possess distinct and multiple biological properties recently confirmed by molecular cloning [6]. As early as 1983, Henderson et al [7] postulated that IL-l may be involved in some of the acute and chronic secondary effects of long-term hemodialysis (HD) treatment. However, the actual immunopathological role of IL-i in HD patients is just begin- ning to be investigated [8—10]. Another monokine that may well be relevant in HD-induced Received for publication October 25, 1988 and in revised form May 11, 1989 Accepted for publication July 24, 1989 © 1990 by the International Society of Nephrology pathology symptoms is the tumor necrosis factor a (TNFa) [3, 11]. Although TNFa and IL-i are biochemically and immuno- logically distinct, they share remarkable similarities in their biological properties [12, 13]. The fact that both monokines exert pleiotropic activities affecting distinct organs [14] along with the observation that they can circulate in the blood, could suggest that they be considered as hormones of inflammation [3, 4]. Lastly, TNFa is able to induce the production of IL-i in vivo and in vitro [15], and a synergy between the two monokines has also been shown in vivo during inflammation [16] and septic shock [171. To our knowledge, TNFa has not been extensively investigated in long-term HD patients [18], whereas more consistent studies have been reported in acute clinical situa- tions such as severe septicemia [19] or renal allograft rejection [201. It has also long been documented that uremia can itself impair the immune response capacity, as has been shown both in patients with end-stage renal failure [21] and in experimental uremia models in the rabbit [22, 23]. Most of these studies have focused on T cell-mediated reactions [241, or on neutrophil and monocytic functions such as chemotactic, adherence, phago- cytic or bactericidal activities in order to explain the high susceptibility of uremic patients to infections [25—27]. How- ever, the possibility that terminal renal failure could also result in monocyte activation leading to monokine production has not yet been investigated. Among the dialysis-related factors which can result in mono- cyte alteration and/or activation, the cellulosic nature of the dialysis membrane has been incriminated [28—30], mainly via its capacity to activate complement system. In this regard, we previously showed both in vivo and in vitro that cellulosic cuprophane (CUP) but not synthetic polyacrilonitrile (PAN-AN 69) dialysis membrane leads to a strong activation of phagocyte oxidative metabolism which is detectable within whole blood and is, at least partly, induced by activated complement com- ponents generated in the plasma following blood interaction with the membrane [31]. In 1986, we first reported a preliminary study showing that plasma IL- 1 is significantly increased in long-term HD patients and that this elevation occurred regardless of the nature of the dialysis membrane [8]. Furthermore, the capacity of monocytes to release IL-i on contact with dialysis membranes has also been observed in vitro and even in the absence of complement [32]. 116
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Influence of uremia and hemodialysis on circulating interleukin-1 and tumor necrosis factor

Kidney International, Vol. 37 (1990), pp. 116—125
Influence of uremia and hemodialysis on circulatinginterleukin-1 and tumor necrosis factor a
ANDRE HERBELIN, ANH THU NGUYEN, JOHANNA ZINGRAFF, PABLO UREA,and BEATRICE DESCAMPS-LATSCHA
INSERM U 25 and CNRS UA 122, INSERM U 90 Hôpital Necker, Paris, France
Influence of uremia and hemodlalysis on circulating interleukin-1 andtumor necrosis factor a. Interleukin-1 (IL-I) and tumor necrosis factora (TNFa) were determined in the plasma of long-term hemodialysis(HD) patients and uremic (UR) patients undergoing their first dialysissession using either cellulosic (CUP) or synthetic (PAN-AN 69) mem-brane-equipped dialyzers. In long-term HD patients, plasma IL-I andTNFa levels were significantly increased compared to their levels innormal subjects. During a single dialysis session, a significant increasein IL-I but not in TNFa was observed. In not yet dialyzed UR patients,LL-l plasma levels did not differ from those observed in normalsubjects. By contrast, TNFa was found significantly increased althoughless than in long-term HD patients. During the first dialysis session, nosignificant increase was observed in the levels of either monokine,Lastly, regardless of the group of patients, no significant influence ofthe dialysis membrane could be detected, suggesting that the observedchanges are not exclusively secondary to the activation of complement.Altogether, these results suggest that the passage of the blood throughthe extracorporeal dialysis circuit triggers the secretion of IL-i andfurther exacerbates that of TNFa by monocytes. The presence ofincreased TNFa in the plasma of first-dialysis UR patients suggests thatfactors unrelated to dialysis contribute to the activation of monocytes inthese patients. Lastly, the concomitant presence of IL-I and TNFa inthe plasma of long-term HD patients could be responsible for some ofthe clinical features observed in these patients, and provides strongevidence favoring the concept that HD can be assimilated to a recurrentacute-phase inflammatory response.
Growing interest has focused in recent years on the role ofcytokines in relation to the acute phase response of the inflam-matory process [1—3]. Among these cytokines, interleukin-1(IL-i), which is mainly derived from monocyte-macrophages,has been shown to exert a broad range of activities associatedwith both acute and chronic inflammatory changes [4, 5], and topossess distinct and multiple biological properties recentlyconfirmed by molecular cloning [6].
As early as 1983, Henderson et al [7] postulated that IL-l maybe involved in some of the acute and chronic secondary effectsof long-term hemodialysis (HD) treatment. However, the actualimmunopathological role of IL-i in HD patients is just begin-ning to be investigated [8—10].
Another monokine that may well be relevant in HD-induced
Received for publication October 25, 1988and in revised form May 11, 1989Accepted for publication July 24, 1989
© 1990 by the International Society of Nephrology
pathology symptoms is the tumor necrosis factor a (TNFa) [3,11]. Although TNFa and IL-i are biochemically and immuno-logically distinct, they share remarkable similarities in theirbiological properties [12, 13]. The fact that both monokinesexert pleiotropic activities affecting distinct organs [14] alongwith the observation that they can circulate in the blood, couldsuggest that they be considered as hormones of inflammation [3,4]. Lastly, TNFa is able to induce the production of IL-i in vivoand in vitro [15], and a synergy between the two monokines hasalso been shown in vivo during inflammation [16] and septicshock [171. To our knowledge, TNFa has not been extensivelyinvestigated in long-term HD patients [18], whereas moreconsistent studies have been reported in acute clinical situa-tions such as severe septicemia [19] or renal allograft rejection[201.
It has also long been documented that uremia can itself impairthe immune response capacity, as has been shown both inpatients with end-stage renal failure [21] and in experimentaluremia models in the rabbit [22, 23]. Most of these studies havefocused on T cell-mediated reactions [241, or on neutrophil andmonocytic functions such as chemotactic, adherence, phago-cytic or bactericidal activities in order to explain the highsusceptibility of uremic patients to infections [25—27]. How-ever, the possibility that terminal renal failure could also resultin monocyte activation leading to monokine production has notyet been investigated.
Among the dialysis-related factors which can result in mono-cyte alteration and/or activation, the cellulosic nature of thedialysis membrane has been incriminated [28—30], mainly via itscapacity to activate complement system. In this regard, wepreviously showed both in vivo and in vitro that cellulosiccuprophane (CUP) but not synthetic polyacrilonitrile (PAN-AN69) dialysis membrane leads to a strong activation of phagocyteoxidative metabolism which is detectable within whole bloodand is, at least partly, induced by activated complement com-ponents generated in the plasma following blood interactionwith the membrane [31].
In 1986, we first reported a preliminary study showing thatplasma IL- 1 is significantly increased in long-term HD patientsand that this elevation occurred regardless of the nature of thedialysis membrane [8]. Furthermore, the capacity of monocytesto release IL-i on contact with dialysis membranes has alsobeen observed in vitro and even in the absence of complement[32].
116

Herbelin et a!: Cytokines in ure,nic and hemodialyzed patients 117
The present study attempts to determine the respectiveinfluence of hemodialysis and uremia on circulating IL-I andTNFa and to evaluate the influence of the nature of the dialysismembrane on the presence of these monokines. For this pur-pose, plasma from both first-dialysis UR and long-term HDpatients have been investigated at the start and at the end of thedialysis session, using either CUP or PAN-AN 69 membranes.
Methods
Patients and control subjects
Forty-four patients with end-stage renal failure, including 18males and 22 females ranging in age from 21 to 69 years (mean50.1 years), participated in the study upon informed consent.Twenty-six of them were long-term hemodialyzed (HD) pa-tients (46 9 months, mean suM), undergoing from four tofive hours of maintenance thrice a week. Eighteen uremicpatients (UR) not yet dialyzed were tested just before andduring their first dialysis session. The etiology of renal failuredid not differ greatly from that of the general dialysis population(glomerulonephritis, 15; polycystic kidney disease, 8; renalvascular disease, 6; hereditary nephropathy, 5; and unknownetiology, 10). Inclusion criteria were the absence of intercurrentinfection and/or immunosuppressive therapy.
Two types of dialyzers of comparable surface (1 to 1.4 m2)were used: a disposable plate kidney equipped with syntheticpolyacrilonitrile (PAN-AN 69 membrane, Hospal, Meysieu,France) and a kidney equipped with cellulosic cuprophanecapillary membrane (CUP, Hospal). All long-term HD patientswere dialyzed with the same type of membrane for at least threemonths. Acetate baths were used in all patients except two whowere dialyzed with bicarbonate baths.
Twenty-five healthy subjects recruited among volunteerblood donors served as controls.
Plasma preparation
Patient blood samples of 15 ml were collected in heparinizedtubes (Liquemine Roche, Paris, France, 10 U/mI of blood) fromthe arterial site of the arteriovenous fistula at the start and at theend of dialysis sessions. In normal subjects, blood sampleswere obtained by cubital venous puncture.
Blood samples were centrifuged (600 X g, 10 mm). Theplasma was separated into small aliquots and frozen at —70°Cand all assays were carried out on samples thawed only once. Inorder to remove inhibitory factors, plasma were gel filtratedbefore testing for IL-i according to the methodology firstproposed by Cannon and Dinareilo [331 and also used by othergroups [8, 9]. Briefly, 0.3 ml of plasma was applied to a I x 30cm sterile column with packed autoclaved Sephadex 0-50 fine(Pharmacia Fine Chemicals, Uppsala, Sweden). The columnwas equilibrated with RPM! 1640 medium (Gibco Laboratories,Grand Island, New York, USA) supplemented with 25 mMHepes (Gibco), 2 mri L-glutamine (Gibco), 100 lU/mi penicillin(Gibco), 100 pg/ml streptomycin (Gibco), and 5 X iO M2-mercaptoethanol (2-ME, Merck, Darmstadt, FRG). Thirty-two 0.7 ml fractions were collected in sterile, pyrogen-freepolypropylene tubes at a flow rate of 1 mI/mm.
Cytokines
Human IL-i (hu IL-i), purified from monocyte supernatant,was purchased from Genzyme (Suffolk, England, UK). Recom-binant human TNFa (hu rTNF), recombinant human IL-1/3 (hurIL-1.3) and recombinant human IL-la (hu rIL-la) were fromDr. F. Floc'h, (Rhône Poulenc, Vitry, France). Based onhalf-maximal PHA-induced thymocyte stimulation, 1 U/mI ofhu rIL-13 preparation corresponds to 100 pg/mi as defined byimmunodetection (ELISA, Cistron Biotechnology, Pine Brook,New Jersey, USA). The activity of recombinant human inter-leukin-2 (hu rIL-2), provided by the Sandoz Forschunginstitut,Vienna, Austria, was calculated based on the NIH standard.Recombinant munne interleukin-4 (mu rIL-4), of Dr. F.Melchers (Basel Institute for Immunology, Basel, Switzerland),was provided by Dr. E. Ruuth [34], and recombinant humanIL-4 (hu rIL-4) was provided by Dr. J. Banchereau (Unicet,Dardilly, France). The effects of mu rIL-4 and hu rIL-4 in theLAF assay were tested by adding twofold serial dilutions ofIL-4 to the thymocytes starting at concentrations far above theoptimal concentration determined in their respective assay.Recombinant human interieukin-6 (hu rIL-6, Janssen Biochem-ica, Beerse, Belgium) was from the Institut Henri Beaufour(Les Ulis, France).
Bioassays for cytokines
The lymphocyte activating factor (LAF) assay. IL-i activitywas determined by measuring the augmentation of phytohemag-glutinin (PHA)-induced murine thymocyte proliferation, as de-scribed [35]. Briefly, io thymocytes obtained from lipopolysac-charide-low responder C3H/HeJ mice, at four to six weeks ofage, (C.S.E.A.L. Orleans, France) were incubated in RPM!1640 medium supplemented with 5% heat-inactivated fetal calfserum (FCS, Flow Laboratories Limited, Irvine, Scotland,UK), 25 mi Hepes, 2 mri L-glutamine, 100 lU/mi penicillin(Gibco), 100 sg/mi streptomycin (Gibco), 5 X i0 M 2-ME, 1pg/ml of PHA (Weilcome Research Laboratories, Beckenham,England, UK), and plasma samples. Each plasma sample wastested in triplicate. DNA synthesis was assayed for the lasteight hours of a 72 hour incubation at 37°C under 5% CO2atmosphere by pulsing the cultures with 1 pCi/well of [3H1-thymidine ([3H]-TdR, 5 Ci/mmol, Centre d'Energie Atomique,Saclay, France). The standard deviation of triplicate sampleswas less than 15% of the mean. Since responsiveness to bothPHA and IL-i varied among thymocyte preparations, pre- andpost-dialysis plasma fractions were assayed on the same thy-mocyte population. IL-l activity in plasma fractions was ex-pressed as a costimulation index (CI) defined by the mean oftriplicate determination of [3H]-TdR incorporation in eacheluted fraction, divided by the mean background incorporationin the presence of fractions eluting before the void volume.
To discriminate the optimal fractions corresponding to IL-ielution, normal plasma supplemented with various concentra-tions of hu rIL-1/3 was subjected to gel filtration. A single peakof IL-i activity eluting approximately 12,000 to 18,000 daltonswas detected. Based on these findings, all plasma from patientsand controls were fractionated and the 12,000 to 18,000 daltonrange fractions were tested for their LAF(IL- 1) activity. PlasmaIL-i level was expressed as the mean activity of these fractions.

118 Herbelin et al: Cytokines in uremic and hemodialyzed patients
The detection limit of hu rIL-1f3 added to plasma and sub-jected to gel filtration was 15 pg/mi, corresponding to a CI of1.8. Fifty pg/mi of hu rIL- 113 gave a significant index of 4, and1 pg of hu rIL-1/3 corresponds to approximately 0.12 CI.
In addition to IL- 1, the possible participation of IL-2, IL-4 orIL-6, known to mediate thymocyte proliferation 135—37] in theLAF assay, was investigated. In the presence of 1 tg/ml ofPHA (that is, the concentration used for detecting IL-i activi-ty), no significant increase in thymocyte proliferation could bedetected with hu rIL-6 up to 12,500 U/mi. By contrast, and asalready described by other groups, significant increase in LAFactivity was observed upon addition of hu rIL-6 together with10 jig/mi of PHA. As reported by Ziotnik et al [36], we alsofound that murine rIL-4 induced significant LAF activity.However, in our test conditions this LAP activity could not beinduced with human rIL-4, regardless of the concentrationused, thus confirming its species-specificity [38]. Lastly, plasmafractions were tested for their IL-2 activity by the CTLL assay.
In some experiments, plasma IL-i activity was confirmed byneutralization with an antibody against human IL-i (rabbitpolyclonal antibody, Cistron Biotechnology). In these experi-ments, plasma fractions with significant IL-i activity wereincubated for six hours at 37°C with 2% to 0.25% (vollvol) rabbitantibody or normal rabbit serum (as control) and then tested inthe LAF assay. In these conditions, we found that this anti-body, when used at 2% (voL1vol), inhibited 90% of the biologicalactivity mediated by 80 pg of hu rIL- 1/3. This neutralization wasobserved as well with chromatographed plasma supplementedwith hu rIL-la, hu rIL-i/3 or hu IL-i. In addition, this poly-clonal anti-IL-i was tested for its effect on hu rIL-6-mediatedthymocyte proliferation and no significant inhibition could bedetected.
The TNFx activity assay. TNFa was measured by its cy-tolytic activity on the L929 tumor cell line [39]. L929 cells weremaintained in RPM! 1640 medium supplemented with antibiot-ics and 10% FCS. For the assay, cells were treated with trypsin(0.125%) (Gibco), washed twice with RPMI and adjusted to adensity of 2.5 x l05/ml. The cells were seeded in 96-wellflat-bottomed culture plates (Falcon 3072, Becton-Dickinsonand C°, New Jersey, USA) at a density of 2.5 x 104/well andincubated 24 hours at 37°C under 5% CO2 atmosphere in 100 jilmedium consisting of RPM! 1640 supplemented with antibioticsand 10% FCS. The supernatant was eliminated and replaced by75 ji! ofdiluted plasma supplemented with 75 jil of actinomycinD (1 jig/mi; Behring Diagnostics, La Jolla, California, USA),and the plate was further incubated for 18 hours.
Cytotoxicity was measured by the MTT (dimethyl-thiazol-2-yl diphenyltetrazolium bromide, Sigma Chemical Co., St.Louis, Missouri, USA) colorimetric assay [401 at 570 nm usinga microplate reader (MR 600, Dynatech, Alexandria, Virginia,USA) with a reference filter at 630 nm. Plasma samples weretested in duplicate and results of optical density of the samplewell as a percentage of that of the control well was taken as ameasure of cell survival. RPM! 1640 supplemented with 10%FCS was used in the control wells. The amount of TNFa ofeach plasma sample was calculated on the basis of dilutioncurves plotted against cell survival. One TNFa unit was definedas the amount of TNFa giving 50% cell survival. Before testing,plasma samples were heated at 56°C for 30 minutes. Thedetection limit of TNFa in plasma was about 10 U/mI cone-
sponding to a concentration of hu rTNFa of 28 pg/ml. It wascontrolled that addition of up to 15% plasma had no effect onthe TNF cytolytic assay.
The bioassay for IL-2. The plasma fractionated samples weretested for their ability to maintain the growth of an IL-2-dependent murine T-cell line (CTLL), as described [35].Briefly, cells were adjusted to a density of io cells/mi in RPM!1640 medium containing antibiotics and 5% FCS. A total of i0cells/well of a 96-well plate (Falcon) were incubated at 37°C in5% CO2 for 48 hours with various sample concentrations. Cellswere pulsed for the final eight hours with [3H]-TdR and har-vested as already described above. Recombinant human IL-2,titrated along with the unknown samples, was used as positivecontrol.
Immunoassays for cytokines
Enzyme-linked immunosorbent assay (ELISA) for IL-I /3. Acommercial, enzyme-linked immunosorbent assay (ELISA,Cistron Biotechnology) was used according to the manufactur-er's suggestions for measuring IL-i/3 levels in diluted plasmasamples. The antiserum in this assay was shown to be specificfor IL-l /3, and the lowest concentration of hu rIL- 1/3 detectablefrom the zero level was 15 to 20 pg/mI. Samples below thedetection level of 15 pg/mi were assumed to have a value of 7.5pg/mi, that is, the midpoint of the interval between 0 and 15.
Radioimmunoassays (RIA). A commercial radioimmunoas-say (RIA, Cistron Biotechnology), which was standardized withhu rIL-1f3, was used for measuring IL-l/3 levels in dilutedplasma samples. Previous experiments to validate the immu-noassay showed a detection limit of 250 pg/mI for hu rIL- 1/3 inplasma, less than 10% interassay variation and more than 80%recovery in the detection of recombinant IL-1/3 added to plasmaregardless of plasma origin (control subjects, HD or UR pa-tients).
TNFa, IL-2 and gamma interferon (IFNy) were assessed inthe plasma of patients and controls by means of soluble phaseRIA (Prof. P. Franchimont, University of Liege, Belgium) andIRE-MEDGENIX (Fleurus, Belgium), using specific rabbitpolyclonal antisera and iodinated human recombinant cytokinemolecules as described in [41, 42]. The sensitivities of the RIAused, defined as the smallest amount of unlabelled antigen ableto significantly reduce the binding of the corresponding cyto-kine to each given specific antibody, were as follows: 50 pg/mifor TNFa, 0.4 U/mI for IFNy and 0.5 U/mi for IL-2. As alreadydetailed elsewhere [41, 42] the antiserum used in each RIA didnot show cross reactivities that could affect the specificity of theassays in the experimental conditions used.
Presentation of results and statistical analysis
In long-term HD patients, each post-hemodialysis cytokineplasma level was corrected according to hemoconcentrationmeasurement using the method described by Bergstrom andWehle [43]. Such correction was not made in nondialyzed URpatients undergoing their first dialysis since no ultrafiltrationwas performed.
All data are given as mean SEM and compared either by theStudent's 1-test or by the paired 1-test depending on the samplegroups (unpaired or paired) to be compared. Differences wereconsidered to be significant when P was less than 0.05.

Herbelin et al: Cytokines in uremic and hemodialyzed patients 119
0C UR HD
Fig. 1. Plasma IL-I activity in 25 healthy control subjects (C), 13first-dialysis uremic patients (UR) and 25 long-term heinodialysispatients (HD) before a dialysis session. Each point represents the meanof IL-i activity (CI) in plasma fractions ranging from 12 to 18 kDamolecular mass, obtained in a given patient. Plasma IL-I activity issignificantly increased in HD patients, compared both to healthycontrol subjects and UR patients (P < 0.001).
Results
Plasma LAF(IL-I) activityComparison between first dialysis and long-term dialysis
patients. Figure 1 compares the LAF(IL-l) activity observed inplasma fractions from 13 UR patients undergoing their firstdialysis session, 25 long-term HD patients and 25 normalsubjects. All patients were tested at the start of the dialysissession. Compared to normal subjects (CI: 1.05 0.08, mean5EM), no significant LAF(IL-l) activity was detected in plasmafractions from UR patients (CI: 1.27 0.07, mean SEM). Bycontrast, and although great variations existed among individ-ual cases, the mean LAF(IL-l) activity was significantly higherin HD patients (CI: 2.91 0.30, mean SEM; P <0.001). Theresults obtained in control normal subjects show that a signifi-cant IL-i activity gives a CI superior to 1.8 (mean + 2 SD ofcontrols). According to data obtained with normal plasmasupplemented with hu rIL-1/3, this CI corresponded to approx-imately 15 pg/mi of IL-i. On this basis, significant LAF(IL- 1)activity was found in 1 out of 25 control subjects and I out of 13UR first-dialysis patients against 20 out of 25 long-term HDpatients (P < 0.001; test). Among the latter, four patientsshowed a CI of 5 to 6 corresponding to that obtained withfractions from plasma supplemented with 60 pg/ml of hu rIL-lJ3(data not shown).
In these patients with the most pronounced IL-i activity, wealso verified that this activity was totally neutralized by anti-IL-i, and in a dose-dependent manner (Table 1). Interestingly,
Table 1. Inhibition of plasma ILagainst h
-1 activity by a polyclonal antibodyuman IL-l
PatientPlasma ILia
(CI)
% Inhibitionb with anti IL-Idilution of the antibody
1/25 1/50 1/100 1/200
1
234
4.24.55.96.2
90 82 89 7586 73 79 7079 63 76 3585 86 45 21
hurlL-1/3200 pg/ml
18.2 92 85 60 24
a Plasma LAF (IL-i) activity expressed as costimulation index (CI).b % inhibition: CI of plasma sample in the presence of the antibody
xIOO.CI of plasma sample in the presence of control serum
UR HD
Fig. 2. Plasma IL-i activity in 12 first-dialysis uremic (UR) and 24long-term dialyzed (HD) patients at the start (T0) and the end (Tf) of asingle dialysis session. Plasma IL-I activity (CI) is measured asdescribed in legend of Figure 1. Each point represents the plasma IL-iactivity in a given patient. The differences observed between the s iiiand the end of the dialysis session in HD patients are significant (P <0.001).
a similar neutralization pattern was obtained from fractionatednormal plasma supplemented with hu rIL-l/3.
Effect of the dialysis session. Figure 2 compares the plasmaLAF(IL- 1) activity at the start and at the end of a single dialysissession in both first-dialyzed UR and long-term HD patients. In
8
6
4
2
0>U
S..S
S
I:
0..4458
6
0>->
2.
0. I I
To Tf To Tf

120 Herbelin et at: Cytokines in uremic and hemodialyzed patients
8
6
(-)>-� 4
2
0
CliP PANFig. 3. Comparative effect of cellulosic-CUP membrane and synthetic-PAN-AN 69 ,nembrane equipped dialyzer on plasma IL-I activityduring a single dialysis session in long-term dialyzed patients. PlasmaIL-i activity (CI) is measured as described in legend of Figure 1. PlasmaIL-i activity is significantly elevated in both CUP and PAN-AN 69treated, long-term dialyzed patients compared to healthy control sub-jects (P < 0.001) and in both groups further increased between the start(Io) and the end (Tf) of the dialysis session (respectively P < 0.001 andP < 0.005).
first-dialysis UR patients, the mean LAF(IL-l) activity re-mained at its initial level, that is, similar to that observed innormal subjects (Fig. 1). By contrast, in long-term HD patients,the already high LAF(IL-l) activity observed at the start of thedialysis session further increased in 18 out of 24 patients (P <0.00 1).
As shown on Figure 3, which illustrates the influence of themembrane equipping the dialyzer, HD patients routinelytreated with the cellulosic (CUP) dialysis membrane showedhigher predialysis levels of LAF(IL-1) plasma activity thanthose with the synthetic (PAN-AN 69) membrane, but theobserved differences in the mean CI values (respectively 3.310.52 vs. 2.51 0.21, mean sEM) were not statisticallysignificant. At the end of the dialysis session, LAF(IL-l)activity significantly increased in both CUP- (P < 0.001) andPAN-AN 69- (P < 0.005) treated patients.
Plasma IL-I /3 immunoreactivitvTable 2 shows results obtained when plasma IL-lJ3 level was
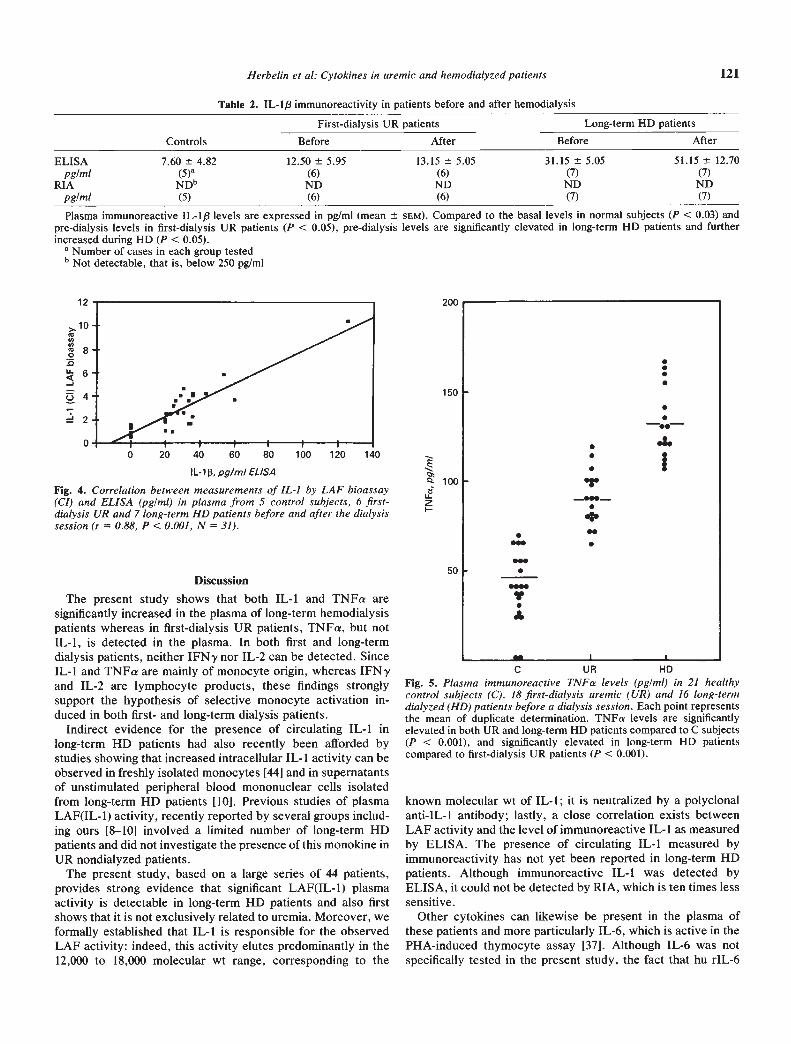
determined by its immunoreactivity in ELISA and RIA. Usingthe ELISA, a significant increase in plasmatic IL-113 concentra-tion was found in long-term HD patients as compared to itsalmost undetectable level (<15 pglml according to the sensitiv-
ity of the assay) in first-dialysis UR patients (P < 0.05) andcontrol subjects (P < 0.03). In addition, this immunoreactiveIL- 1 activity increased during the dialysis session (P < 0.05),thus corroborating the LAF assay determination. By contrast,regardless of plasma origin and time of dialysis, no significantIL-l/3 could be detected by RIA in any of the plasma tested.This result confirms our previous observation that the LAFassay is approximately ten times more sensitive than the RIAspecific for human IL- 1/3 (data not shown).
A strong linear correlation was observed (r =0.88,P <0.001,N 31) when IL-I measured in the above experiments byELISA was compared to that obtained by the LAF assay in thesame plasma (Fig. 4).
Plasma TNFs immunoreactivity
Comparison between first dialysis and long-term dialysispatients. Figure 5 illustrates the results obtained in the deter-mination of plasma TNFa by radioimmunoassay in long-termHD and first-dialysis UR patients before the dialysis session. Itis observed that: 1) compared to its level in normal subjects(46.7 4.2 pg/mi, mean SuM), plasma TNFa immunoreac-tivity was significantly higher in both long-term HD and first-dialysis UR patients (P < 0.001); and 2) among the two groupsof patients, long-term HD had significantly higher TNFa levels(131.8 5 pg/mI, mean SEM) than first-dialysis UR patients(89.7 3.8 pg/ml, mean SEM, P < 0.00 1).
Effect of the dialysis session. As shown on Figure 6, RIAdetermination of plasma TNFa levels before and at the end ofthe dialysis session showed that TNFa remained constant, thatis, mean level comprised between 125 and 135 pg/mI in bothgroups of patients. Moreover, regardless of the time of sam-pling, plasma TNFa levels did not significantly differ betweenthe patients treated with CUP and those treated with PAN-AN69 dialysis membranes (Fig. 7).
Plasma TNFa biological activity
To determine whether TNFa immunoreactivity in plasmafrom dialyzed patients could exert biological activity, we testedthe capacity of some of these plasma to induce cytolytic activityon the TNFa-sensitive L929 cell line.
Table 3 compares the TNFa activity measured in dilutedplasma from five long-term HD patients, four first-dialysis URpatients and six normal subjects. No significant TNFa activitycould be detected in the plasma of normal subjects, except inone case which showed TNFa level close to the detection limitof the assay (10 TNFa U/mi). Compared to normal subjects,significant TNFa activity was detected before the dialysissession in three out of four first-dialysis UR patients and fourout of five long-term HD patients. Plasma TNFa activity wasalso significantly higher in long-term HD patients than infirst-dialysis UR patients, although in both groups it remainedconstant between the start and the end of the dialysis session.
IFNy and IL-2 determinations
Neither immunoreactive IFNy nor IL-2, determined either bythe IL-2 CTLL-dependent assay or by RIA, was detected in anyof the plasma from UR (N = 14) or long-term HD (N = 16)
patients.
r
— I ITo If To Tf
Herbeiin et a!: Cytokines in uremic and hemodialyzed patients 121
Table 2. IL-l/3 immunoreactivity in patients before and after hemodialysis
ControlsFirst-dialysis UR patients Long-term HD patients
Before After Before After
ELISA 7.60 4,82 12.50 5.95 13.15 5.05 31.15 5.05 51.15 12.70
pg/mi (5) (6) (6) (7) (7)RIA NDb ND ND ND ND
pg/mi (5) (6) (6) (7) (7)
Plasma immunoreactive IL-l/3 levels are expressed in pg/mi (mean sEM). Compared to the basal levels in normal subjects (P < 0.03) andpre-dialysis levels in first-dialysis UR patients (P < 0.05), pre-dialysis levels are significantly elevated in long-term HD patients and furtherincreased during HD (P < 0.05).a Number of cases in each group tested
b Not detectable, that is, below 250 pg/mi
Discussion
The present study shows that both IL-i and TNFa aresignificantly increased in the plasma of long-term hemodialysispatients whereas in first-dialysis UR patients, TNFa, but notIL-l, is detected in the plasma. In both first and long-termdialysis patients, neither IFNy nor IL-2 can be detected. SinceIL-i and TNFa are mainly of monocyte origin, whereas IFNyand IL-2 are lymphocyte products, these findings stronglysupport the hypothesis of selective monocyte activation in-duced in both first- and long-term dialysis patients.
Indirect evidence for the presence of circulating IL- 1 inlong-term HD patients had also recently been afforded bystudies showing that increased intracellular IL- 1 activity can beobserved in freshly isolated monocytes [44] and in supernatantsof unstimulated peripheral blood mononuclear cells isolatedfrom long-term HD patients [10]. Previous studies of plasmaLAF(IL-l) activity, recently reported by several groups includ-ing ours [8—10] involved a limited number of long-term HDpatients and did not investigate the presence of this monokine inUR nondialyzed patients.
The present study, based on a large series of 44 patients,provides strong evidence that significant LAF(IL- 1) plasmaactivity is detectable in long-term HD patients and also firstshows that it is not exclusively related to uremia. Moreover, weformally established that IL-i is responsible for the observedLAF activity: indeed, this activity elutes predominantly in the12,000 to 18,000 molecular wt range, corresponding to the
C UR HO
Fig. 5. Plasma immunoreactive TNFa levels (pg/mi) in 21 healthycontrol subjects (C), 18 first-dialysis uremic (UR) and 16 long-termdialyzed (HD) patients before a dialysis session. Each point representsthe mean of duplicate determination. TNFa levels are significantlyelevated in both UR and long-term HD patients compared to C subjects(P < 0.001), and significantly elevated in long-term HD patientscompared to first-dialysis UR patients (P < 0.001).
known molecular wt of IL- 1; it is neutralized by a polyclonalanti-IL-l antibody; lastly, a close correlation exists betweenLAF activity and the level of immunoreactive IL- 1 as measuredby ELISA. The presence of circulating IL-i measured byimmunoreactivity has not yet been reported in long-term HDpatients. Although immunoreactive IL-i was detected byELISA, it could not be detected by RIA, which is ten times lesssensitive.
Other cytokines can likewise be present in the plasma ofthese patients and more particularly IL-6, which is active in thePHA-induced thymocyte assay [37]. Although IL-6 was notspecifically tested in the present study, the fact that hu rIL-6
20 40 60 80
200 -
150
100
50
IL-i 3, pg/mi EL ISA
Fig. 4. Correlation between measurements of IL-I by LAF bioassay(CI) and ELISA (pg/mi) in plasma from 5 control subjects, 6 first-dialysis UR and 7 long-term HD patients before and after the dialysissession (r 0.88, P < 0.001, N = 31).
U-z
..
S—..—
• $
-.••-.+. S.
S.. ••5•S
S...S£

122 Herbelin et a!: Cytokines in uremic and hemodialyzed patients
UR HDFig. 6. Plasma immunoreactive TNFa levels (pg/mi) in 14 first-dialysisuremic (UR) and 14 long-term dialyzed (HD) patients at the start (T0)and the end (1') of dialysis session. Each point represents the mean ofduplicate determination, TNFa levels are not significantly different atthe end of dialysis compared to its level at the start in both UR and HDpatients.
induced no significant thymocyte proliferation at the PHAconcentration used in our LAF assay argues against the possi-bility that it could interfere in the IL-i bioassay. Moreover, wecould exclude the presence of IL-2, which is also known tointerfere in the LAF assay, and the possibility that human IL-4could induce significant proliferation of murine thymocytes.
The presence of plasma TNFa, a cytokine intimately associ-ated with the acute phase response and sharing almost all of itsproperties with IL-i, has recently been reported in one long-term HD patient by Lonnemann et al [18]. The present studyshows that TNFa is considerably increased in plasma of long-term HD patients compared to normal subjects. Unlike IL-l,TNFa is also significantly increased in UR patients before theirfirst dialysis, although not as much as in long-term HD patients.This apparent discrepancy between IL-i and TNFa plasmalevels observed in uremic patients suggests that uremia-depen-dent factors may selectively influence the secretion of TNFawithout interfering with that of IL-i. Alternatively, one maypropose that these factors induce the production of bothmonokines but that while the former is secreted, the latterremains intracellular and/or membrane-associated. Previousstudies have reported the presence of such a form of IL-i,especially in long-term HD patients [44]. By contrast, no
I L
To Tf To Tf
CUP PANFig. 7. Comparative effect of celiulosic-CUP and synthetic-PAN-AN69 membrane-equipped dialyzers on plasma immunoreactive TNFalevels (pg/mi) during a single dialysis session in long-term dialyzed(HD) patients. Each point represents the mean of duplicate determina-tions. TNFa levels are not significantly different in CUP compared toPAN-AN 69 treated HD patients. No significant change in TNFa levelsis observed at the end (Tf) compared to the start (T0) of dialysis sessionregardless of the type of membrane equipping the dialyzer.
Table 3. TNFa activity in patients before and after hemodialysis
First-dialysis UR Long-term HD
Controlspatients patients
Before After Before After
7.05 3.58 39 14.8 35 12.6 60 28.3 76 24(6) (4) (4) (5) (5)
Plasma TNFa activity is expressed in units/mi (mean 5EM).Compared to the basal levels in normal subjects, pre-dialysis levels aresignificantly elevated both in first-dialysis UR (P < 0.02) and long-termHD (P < 0.05) patients, but not further increased during HD in bothgroups of patients.a Number of cases in each group tested.
evidence for cell-associated TNFcr has been reported; in addi-tion, we failed to detect its presence in LPS stimulated humanmonocytes (unpublished observations).
Increased TNFa levels in the plasma of not yet dialyzed URpatients could be induced by damaged cells. Baud et al [45]recently showed that rat mesangial cells produce cytokinesrelated to TNFa. Similar observations were also reported byother groups with rat [46] or rabbit [47] glomeruli. Thesefindings suggest that mesangial cells could be responsible for
200
150
100aU-zI-
50
0
200
150
S.-
' 100a
U-zI—
50
0
To Tf To Tf

Herbelin et a!: Cytokines in uremic and hemodialyzed patients 123
the increased TNFa production observed in patients withend-stage renal failure. Alternatively, a possible role of in-creased TNFa levels in the development of renal impairmentbefore dialysis should be considered. In fact, some authorshave recently shown that TNFa infused in rats can inducemetabolic acidosis and acute renal failure [3]. Moreover, Rhee,McGoldnck and Meuwissen [271 reported the presence of aneutrophil stimulatory reduced oxygen species (ROS) factor inserum from nondialyzed UR patients and suggested that it isspecifically associated with renal dysfunction. Although thisserum factor was found in the range of 1,000 dalton molecularmass, the role of TNFa, known to induce ROS production byphagocytic cells [48], remains to be investigated in nondialyzedUR patients. Progressive renal damage may indeed result fromTNFa-induced production of ROS by neutrophils in the vascu-lar membrane of the kidney.
Interestingly, during the first dialysis session, IL- 1 remainedundetectable and TNFa maintained its initial level. Thesefindings may suggest that repeated dialysis sessions and/oraccumulation of dialysis-related factors are necessary to induceor to amplify the secretion of these cytokines. Comparativeinvestigation of TNFa and IL-i plasma levels at the beginningand the end of a single dialysis session in long-term HD patientsshowed a significant increase in IL-l but not in TNFa. Thisdiscrepancy between the evolution of IL-i and TNFa levelscould be explained by differences in the kinetics of secretion ofthe two monokines. In this regard, Michie et a! [49] havereported that TNFa response following injection of endotoxinin human volunteers occurs between the 90th and 120th minuteand is no longer detected at the 4th hour.
In contrast to our previous report showing that CUP but notPAN-AN 69 membrane induces a significant activation ofphagocyte oxidative metabolism [311, we did not detect a majorinfluence of CUP membrane on IL-l or TNFa secretion. SinceCUP but not PAN-AN 69 membrane has the potential ofactivating the complement system, our observations suggestthat IL-i and TNFa production in long-term HD patients is notexclusively caused by the activation complement fragmentswhich are well known to be potent inducers of both IL-i andTNFa secretion [50—52]. Interestingly, Binge! et al [531 recentlyreported that long-term HD patients treated with CUP mem-branes display high plasma levels of IL-l activity but that thiswas markedly decreased upon a subsequent dialysis with PANmembrane, thus suggesting that this membrane might favor thebinding or the clearance of IL- 1. The discrepancy between thatstudy and ours remains to be explained. However, the fact thatour patients were treated for at least three months with CUP orPAN membrane, allows a more accurate appreciation of theproper effect of each of the two membranes. Lastly, Haeffner-Cavaillon et a! [44], measuring intramonocytic IL-! activity,also found that both CUP- and PAN-treated patients displayincreased IL-l activity and that no significant difference couldbe detected between the two groups of patients.
In vitro, Betz et al [32] have shown that a complementindependent stimulation of IL-i release by human monocytesmay occur following a brief exposure to cuprammonium dialy-sis membranes. Concerning the synthetic (PAN) dialysis mem-brane, Lonnemann et al [54] have reported its intrinsic propertyto stimulate IL-i production. Since circulating IL-i and/orTNFa are present in first-dialysis UR and long-term HD pa-
tients regardless of the type of the membrane equipping thedialyzer, and given the above-mentioned in vitro studies, weconclude that the type of membrane does not exert a pivotalrole on the presence of these circulating cytokines.
Among other factors which may contribute to triggering thesecretion of these monokines in hemodialysis patients andconsequently to masking complement-dependent monokineproduction, the microbial contamination of the dialysate shouldbe considered. Although all the evidence is not yet in hand, it ishighly probable that repeated exposure of blood to minimaldoses of endotoxin may lead to chronic monocyte activation. Inaddition, according to a recent report from Nisbeth et a! [55],significant amounts of circulating endotoxins can be present innot yet dialyzed UR patients, thus suggesting a possible role ofLPS in the increased levels of TNFa observed at the beginningof the first dialysis. The fact that IL-i was not detectable inthese patients could be related to a recent report [49] showingincreased plasma TNFa but not IL-i in normal subjects treatedwith endotoxins.
Even though we show that IL- 1 and TNFa can be detected inplasma of long-term HD patients, it remains to be verifiedwhether these two monokines are present in sufficient amountsto be responsible for the clinical manifestations seen in long-term dialysis patients. Concerning IL-i, this study providesevidence for the presence of circulating immunoreactive IL-i inlong-term HD patients, thus corroborating previous [8] andpresent findings obtained by the LAF assay. However, the IL-ilevels observed, ranging between 30 and 60 pg/mI (except in onecase in which it rose over 100 pg/mi), are much lower than thosereported by Maury and Teppo [56] in renal allograft recipientsundergoing acute rejection. Concerning TNFa, recent studieshave demonstrated elevated levels of plasma TNFa in infec-tious diseases. Scuderi et al [57], using an enzyme immunoas-say, found a mean level of TNFa reaching 119 pg/ml in patientswith malaria, while Waage, Halstensen and Espevik [19], usinga bioassay, found raised TNFa levels (440 U/mI correspondingto 100 pg/mI) in septicemic patients. The fact that in our study,on the one hand, similar immunoreactive TNFc levels arefound in the plasma from long-term HD patients and, on theother hand, that this TNFs is biologically active, favors thehypothesis that TNFa could be at least partly implicated in theclinical manifestations observed in these patients.
Moreover, particular attention must be paid to synergisticproperties of TNFa and IL-l monokines. Recent studies sup-port the concept that IL-! and TNFa act synergistically in avariety of biological processes such as fibroblast prostaglandinE2 production [58] and induction of polymorphonuclear leuko-cyte migration [161. Furthermore, the possibility that plasmaTNFa when present at sufficient levels could itself induce IL-lsecretion by monocytes [15], must also be considered in long-term HD patients. In this regard, recent results in the study ofthe hypotensive effect of IL-l and TNFa in rabbits are relevantto the synergy resulting from the combination of TNFa itselfplus IL-i induced by TNFa in these rabbits [12]. Similarly, theparticipation of IL-6 which has been shown to be involved inthe acute phase response [59] and to be produced by monocytesin response to TNFa or IL-I [60, 61], requires further study.Furthermore, endotoxin contaminants entering the blood com-partment from the dialysate may not only induce monokineproduction but also act as a potentiator of several monokine

124 Herbelin et a!: Cytokines in uremic and hemodialyzed patients
functions. Likewise, strong synergy between TNFa and bacte-ria and their products, causing hemorrhagic necrosis and lethalshock in normal mice, has been recently described [62].
In conclusion, the present study supports the hypothesis thatboth dialysis and uremia related factors contribute to monocyteactivation and consequent IL-i and/or TNFa production. Themultifaceted biological effects of monokines, which we showedto be produced in large amounts during HD treatment, mayexplain some of the pathological findings observed in thesepatients. In addition, our findings demonstrate that the releaseof IL-i and TNFa into the circulation in man is not confined tosevere bacterial infections, but also occurs in the immunologi-cal tissue injury caused by the inflammatory response. Lastly, itis tempting to propose that long-term production of these twomonokines, which are known for their synergistic effects, mayaccount for the occurrence of acute phase changes in HDpatients.
Acknowledgments
The authors gratefully thank: Dr. B. Zins for her help in patientmanagement; Prof. P. Franchimont and Dr. M. Baudrihaye for the giftof TNFa, IL-2 and INFyRIA; Dr. L. Chatenoud for her contribution inthese RIA determinations; Dr. B. Dugas for the gift of hu rIL-6; F.Tresset for her technical assistance; Dr. F. Praz for her contribution inpreparing the manuscript; Dr. F. Zavala, and Dr. C. Michel-Herbelinfor valuable discussions and suggestions; and D. Broneer for revisingthe manuscript. Recombinant human TNFa and recombinant humanIL-l/3 and IL-la were a gift from Dr. F. Floc'h, Rhóne Poulenc, Vitry,France. Recombinant human interleukin-2 used in this study wasprovided by the Sandoz Forschunginstitut, Vienna, Austria. Recombi-nant murine interleukin-4 used in this study was a gift from Dr. F.Melchers, Basel Institute for Immunology, Basel, Switzerland andrecombinant human IL-4 was provided by Dr. J. Banchereau, Unicet,Dardilly, France. Recombinant human interleukin-6 used in this studywas donated by Institut Henri Beaufour, Les Ulis, France.
Reprint requests to Dr. Beatrice Descamps-Lwscha, INSERM U25,Hópita! Necker, 161 rue de Sèvres, 75743 Paris Cedex 15, France.
References
I. DINARELLO CA, MIER JW: Current concepts: lymphokines. N Eng!JMed 317:940—945, 1987
2. RUDDLE NH: Tumor necrosis factor and related cytotoxins. Im-munol Today 8:129—130, 1987
3. BEUTLER B, CERAMI A: Cachectin: more than a tumor necrosisfactor. N Engi J Med 316:379—385, 1987
4. OPPENHEIM JJ, KOVACS EJ, MATSUSHIMA K, DURUM SK: There ismore than one interleukin 1. Immunol Today 7:45—56, 1986
5. DINARELLO CA: Interleukin-1 and the pathogenesis of the acutephase response. NEnglJMed3ll:1413—1418, 1984
6. DINARELLO CA, CANNON JG, MIER JW, BERNHEIM HA, Lo-PRESTE G, LYNN DL, LOVE RN, WEBB AC, AURON PE, REUBENRC, RICH A, WOLFF SM, PUTNEY SD: Multiple biological activi-ties of human recombinant interleukin I. J Cliii Invest 77:1734—1739, 1986
7. HENDERSON LW, KOCH KM, DINARELLO CA, SHALDON S: He-modialysis hypotension: The interleukin hypothesis. Blood Purif
1:3—8, 19838. DESCAMPS-LATSCHA B, HERBELIN A, NGUYEN AT, UZAN M,
ZINGRAFF J: Haemodialysis-membrane-induced phagocyte oxida-tive metabolism activation and interleukin-1 production. Lfe Sup-port Syst 4:349—353, 1986
9. LONNEMANN G, BINGEL M, KOCH KM, SHALDON S, DINARELLOCA: Plasma interleukin- 1 activity in humans undergoing hemodial-ysis with regenerated cellulosic membranes. Lymphokine Res 6:
63—70, 198710. LUGER A, KOVARIK J, STUMMVOLL HK, URBANSKA A, LUGER
TA: Blood-membrane interaction in hemodialysis leads to in-creased cytokine production. Kidney mt 32:84—88, 1988
11. ZIEGLER EJ: Tumor necrosis factor in humans. N Engi J Med318:1533—1535, 1988
12. DINARELLO CA: The biology of interleukin 1 and comparison totumor necrosis factor. Immunol Lett 16:227—232, 1987
13. LE J, VILCEK J: Tumor necrosis factor and interleukin 1: Cytokineswith multiple overlapping biological activities. Lab Invest 56:
234—248, 198714. OPPENHEIM JJ, MATSUSHIMA K, Y0sHIMuRA T, LEONARD EJ: The
activities of cytokines are pleiotropic and interdependent. ImmunolLett 16:179—184, 1987
15. DINARELLO CA, CANNON JG, WOLFF SM, BERNHEIM HA, BEUT-LER B, CEIM1 A, FIGARI IS, PALLADINO MA, O'CONNOR JVTumor necrosis factor (cachectin) is an endogenous pyrogen andinduces production of interleukin 1. J Exp Med 163:1433-1450, 1986
16. WANKOWICZ Z, MEGYERI P, I5SEKUTZ A: Synergy between tumournecrosis factor a and interleukin-1 in the induction of polymorpho-nuclear leucocyte migration during inflammation. JLeukocyte Biol43:349—356, 1988
17. OKUSAWAS,GELFAND JA, IKEJIMA T, CONNOLLY RJ, DINARELLOCA: Interleukin 1 induces a shock-like state in rabbits. J C/in Invest81:1162—1172, 1988
18. LONNEMANN G, VAN DER MEER JWM, CANNON JG, DINARELLOCA, KOCH KM, GRANOLLERAS C, DESCHODT G, SHALDON 5:Induction of tumor necrosis factor during extracorporeal bloodpurification. NEngi J Med 317:963—964, 1987
19. WAAGE A, HALSTENSEN A, ESPEVIK T: Association betweentumour necrosis factor in serum and fatal outcome in patients withmeningococcal disease. Lancet 1:355—357, 1987
20. MAURY CPJ, TEPPO AM: Raised serum levels of cachectin/tumornecrosis factor a in renal allograft rejection. J Exp Med 166:1132—1137, 1987
21. HANICKI Z, CICHOCKI T, KoMoRowsRA Z, SULOWICZ W, SM0-LENSKI 0: Some aspects of cellular immunity in untreated andmaintenance hemodialysis patients. Nephron 23:273—275, 1979
22. SILVERBLATT FJ, BEATY HN, PETERSDORF RG: The effect ofuremia on the pyrogenic response of rabbits. Proc Soc Exp Biol
Med 132:492—496, 196923. TURNBULL AJ, KLUGER Mi, CRANSTON WI, TOWNSEND Y: Are
the kidneys involved in the removal of endogenous pyrogens in therabbit?. J Therm Biol 7:95—98, 1982
24. BEAURAIN G, NARET C, MARCON L, GRATEAU G, DRUEKE T,URENA F, NELSON DL, BACH iF, CHATENOUD L: In vivo T cellpreactivation in chronic uremic hemodialyzed and non-hemodia-lyzed patients. Kidney mt (in press)
25. MONTGOMERIE JZ, KALMANSON GM, GUZE LB: Leukocytephagocytosis and serum bactericidal activity in chronic renal fail-ure. Am J Med Sd 264:385—393, 1972
26. LESPIER-DEXTER LE, GUERRA C, OJEDA W, MARTINEZ-MAL-DONADO M: Granulocyte adherence in uremia and hemodialysis.Nephron 24:64—68, 1979
27. RHEE MS, MCGOLDRICK MD, MEUWISSEN Hi: Serum factor frompatients with chronic renal failure enhances polymorphonuclearleukocyte oxidative metabolism. Nephron 42:6—13, 1986
28. SHALDON S: Future trends in biocompatibility aspects of hemodi-alysis and related therapies. Cliii Nephrol 26:513—516, 1986
29. HENDERSON LW, CHENOWETH D: Biocompatibility of artificialorgans: An overview. Blood Purif 5:100—Ill, 1987
30. CHENOWETH D, CHEUNG AK, HENDERSON LW: Anaphylatoxinformation during hemodialysis: Effects of different dialyzer mem-branes. Kidney Int 24:764—769, 1983
31. NGUYEN AT, LETHIAS C, ZINGRAFF J, HERBELIN A, NARETC,DESCAMPS-LATSCHA B: Hemodialysis membrane-induced activa-tion of phagocyte oxidative metabolism detected in vivo and invitro within microamounts of whole blood. Kidney mt 28:158—167,1985
32. BETZ M, HAENSCH GM, RAUTERBERG EW, BOMMER J, RITZ E:Cuprammonium membranes stimulate interleukin 1 release andarachidonic acid metabolism in monocytes in the absence ofcomplement. Kidney Int 34:67—73, 1988
33. CANNON JG, DINARELLO CA: Increased plasma interleukin-1 ac-tivity in women after ovulation. Science 227:1247—1249, 1985

Herbelin et al: Cytokines in urernic and hernodialyzed patients 125
34. RUUTH E, COUILLIN I, HERBELIN A, Piz F: Functional bioassaysfor B cell growth factors using polyclonally activated murine spleenB cells. Lymphokine Res 8: 147—158, 1989
35. GEARING AJH, JOHNSTONE AP, THORPE R: Production and assayof the interleukins. J Immuno! Meth 83:1—27, 1985
36. ZLOTNIK A, RANSOM J, FRANK G, FISCHER M, HOWARD M:Interleukin 4 is a growth factor for activated thymocytes: Possiblerole in T-cell ontogeny. Proc Nat! Acad Sci USA 84:3856—3860,1987
37. LE J, FREDRICKSON G, REIS LFL, DIAMANTSTEIN T, HIRAN0 T,KISHIMOTO T, VILCEK J: Interleukin 2-dependent and interleukin2-independent pathways of regulation of thymocyte function byinterleukin 6. Proc Nat! Acad Sci USA 85:8643—8647, 1988
38. MOSMANN TR, YOKOTA T, KASTELEIN R, ZURAWSKI SM, ARA! N,TAKEBE Y: Species-specificity of T cell stimulating activities of IL2 and BSF-1 (IL 4): Comparison of normal and recombinant, mouseand human IL 2 and BSF-l (IL 4). Jlmmunol 138:1813—1816, 1987
39. RUFF M, GIFFORD GE: Tumor necrosis factor, in Lymphokines,edited by PICK E, Academic Press, New York, 1981, 2, p. 235
40. MOSMANN T: Rapid colorimetric assay for cellular growth andsurvival: Application to proliferation and cytotoxicity assays. JImmunol Meth 65:55-63, 1983
41. REUTER A, BERNIER J, VRINDTS-GEVAERT Y, MEULEMAN-GATHYR, MALAISE M, FIERS W, FRANCHIMONT P: Production of inter-feron gamma by peripheral blood mononuclear cells from normalsubjects and from patients with rheumatoid arthritis. C!in ExpRheumato! (in press)
42. REUTER A, BERNIER J, GYSEN P, GEVAERT Y, GATHY R, LOPEZM, DUPONT G, DAMAS P, FRANCI-IIMONT P: A RIA for tumornecrosis factor (TNFa) and interleukin 1(3 (IL-1p) and their directdetermination in serum, in Monokines and Other Non-!ymphocyticCytokines, edited by POWANDA M, New York, Alan Liss Inc, 1988,pp. 377—381
43. BERGSTROM J, WEHLE B: No change in corrected /3-microglobulinconcentration alter cuprophane haemodialysis. Lancet 1:628—629,1987
44. HAEFFNER-CAVAILON N, FISCHER E, BACLE F, CARRENO MP,MAILLET F, CAVAILLON JM, KAZATCHKINE MD: Complementactivation and induction of interleukin-l production during hemo-dialysis. Contrib Nephrol 62:86—98, 1988
45. BAUD L, OUDINET JP, BENS M, NOE L, ETIENNE J, ARDAILLOUR: Production of tumor necrosis factor by rat mesangial cells inresponse to bacterial lipopolysaccharide. Kidney mt (in press)
46. LOWRY RP, RUBY ZH: Glomeruli of nephritic rats release tumornecrosis factor a and serine protease with cytotoxic activity.(abstract) Xe" Internationa! Congress of Nephrology, July 26-31,London, p. 339, 1987
47. WIGGINS RC, ELDREDGE C, KUNKEL S: Monokine production byglomeruli at different stages of crescent formation in the rabbit.(abstract). Xth International Congress of Nephro!ogy, July 26-31,London, p. 368, 1987
48. TSUJIMOTO M, YOKOTA S, VILCEK J, WEISSMANN G: Tumor
necrosis factor provokes superoxide anion generation from neutro-phils. Bioch Biophys Res Corn 137:1094—1100, 1986
49. MICHIE HR, MANOGUE KR, SPRIGGS DR. REVHAUG A, O'DWYERS, DINARELLO CA, CERAMI A, WOLFF SM, WILMORE DW: Detec-tion of circulating necrosis factor alter endotoxin administration. NEng! J Med 318:1481—1486, 1988
50. OKUSAWA 5, DINARELLO CA, YANCEY KB, ENDRES 5, LAWLEYTJ, FRANK MM, BURKE JF, GELFAND JA: C5a induction of humaninterleukin 1. Synergistic effect with endotoxin or interferon-y. J.Immunol 139:2635—2640, 1987
51. OKUSAWA S, YANCEY KB, VAN DER MEER JWM, ENDRES 5,LONNEMANN 0, HEFTER K, FRANK MM, BURKE JF, DINARELLOCA, GELFAND JA: C5a stimulates secretion of tumor necrosisfactor from human mononuclear cells in vitro. Comparison withsecretion of interleukin 1/3 and interleukin la. J Exp Med 168:443—448, 1988
52. HAEFFNER-CAVAILLON N, CAVAILLON JM, LAUDE M, KAZATCH-KINE MD: C3a (C3adesArg) induces production and release ofinterleukin I by cultured human monocytes. J Immuno! 139:794—799, 1987
53. BINGEL M, LONNEMANN 0, KOCH KM. DINARELLO CA, SHAL-DON S: Plasma interleukin-l activity during hemodialysis: Theinfluence of dialysis membranes. Nephron 50:273—276, 1988
54. LONNEMANN G, KOCH KM, SHALDON 5, DINARELLO CA: Studieson the ability of hemodialysis membranes to induce, bind, and clearhuman interleukin- I. J Lab C!in Med 112:76—86, 1988
55. NISBETH U, HALLGREN R, ERIKSSON 0, DANIELSON BG: Endo-toxemia in chronic renal failure. Nephron 45:93—97, 1987
56. MAURY CPJ, TEPPO AM: Serum immunoreactive interleukin 1 inrenal transplant recipients. Transp!ant 45:143—147, 1988
57. SCUDERI P, STERLING KE, LAM KS, FINLEY PR, RYAN KJ, RAYCG, PETERSEN E, SLYMEN Di, SALMON SE: Raised serum levels oftumour necrosis factor in parasitic infections. Lancet 2:1364—1365,1986
58. ELIAS JA, GUSTILO K, BAEDER W, FREUNDLICH B: Synergisticstimulation of fibroblast prostaglandin production by recombinantinterleukin 1 and tumor necrosis factor. J Immuno! 138:3812—3816,1987
59. NLJSTEN MW, DE GROOT ER, TEN DUIS HJ, KLASEN HJ, HACKCE, AARDEN LA: Serum levels of interleukin-6 and acute phaseresponses. Lancet 2:921, 1987
60. KOHASE M, HENRIKSEN-DE STEPHANO D, MAY LT, VILCEK i,SEHGAL PB: Induction of /32-interferon by tumor necrosis factor: Ahomeostatic mechanism in the control of cell proliferation. Ce!!45:659—666, 1986
61. CONTENT J, DE WITT L, POUPART P, OPDENAKKER G, VANDAMME J, BILLIAU A: Induction of a 26-kDa-protein mRNA inhuman cells treated with an interleukin-l-related, leukocyte-de-rived factor. Eur J Biochem 152:253—257, 1985
62. ROTHSTEIN JL, SCHREIBER H: Synergy between tumor necrosisfactor and bacterial products causes hemorrhagic necrosis andlethal shock in normal mice. Proc Nat! Acad Sci USA 85:607—611,1988




















